

Abstract
In chronic kidney disease (CKD), hyperphosphatemia-induced inflammation aggravates vascular calcification (VC) by increasing vascular smooth muscle cell (VSMC) osteogenic differentiation, ADAM17-induced renal and vascular injury, and TNFα-induction of neutral-sphingomyelinase2 (nSMase2) to release pro-calcifying exosomes. This study examined anti-inflammatory β-glucans efficacy at attenuating systemic inflammation in health, and renal and vascular injury favoring VC in hyperphosphatemic CKD. In healthy adults, dietary barley β-glucans (Bβglucans) reduced leukocyte superoxide production, inflammatory ADAM17, TNFα, nSMase2, and pro-aging/pro-inflammatory STING (Stimulator of interferon genes) gene expression without decreasing circulating inflammatory cytokines, except for γ-interferon. In hyperphosphatemic rat CKD, dietary Bβglucans reduced renal and aortic ADAM17-driven inflammation attenuating CKD-progression (higher GFR and lower serum creatinine, proteinuria, kidney inflammatory infiltration and nSMase2), and TNFα-driven increases in aortic nSMase2 and calcium deposition without improving mineral homeostasis. In VSMC, Bβglucans prevented LPS- or uremic serum-induced rapid increases in ADAM17, TNFα and nSMase2, and reduced the 13-fold higher calcium deposition induced by prolonged calcifying conditions by inhibiting osteogenic differentiation and increases in nSMase2 through Dectin1-independent actions involving Bβglucans internalization. Thus, dietary Bβglucans inhibit leukocyte superoxide production and leukocyte, renal and aortic ADAM17- and nSMase2 gene expression attenuating systemic inflammation in health, and renal injury and aortic calcification despite hyperphosphatemia in CKD.
Subject terms: Nephrology, Phosphorus metabolism disorders
Introduction
In Chronic Kidney Disease (CKD), the development of hyperphosphatemia increases the risk of vascular calcification (VC) and cardiovascular mortality1. Elevations in serum phosphate (P), even to levels below the upper normal limit, increase the propensity for VC indirectly by worsening secondary hyperparathyroidism (SHPT) and bone de-mineralization2 and also directly, by inducing the osteogenic differentiation of vascular smooth muscle cells (VSMC)3.
Hyperphosphatemia also increases systemic inflammation4,5 and consequently, oxidative-stress-driven multi-organ injury6,7 worsening CKD-induced renal and vascular damage predisposing to VC and high mortality rates1,3. Part of the renal and vascular deleterious effects of oxidative stress involves the induction of two critical enzymes: ADAM17 (A Disintegrin And Metalloproteinase, also called TACE for Tumor necrosis factor Alpha Converting Enzyme)8,9 and neutral sphyngomyelinase 2 (nSMase2)10. Specifically, renal ADAM17 expression increases in CKD of all etiologies11 aggravating renal damage12 and systemic inflammation, the latter by releasing soluble TNFα to the circulation9. In turn, TNFα induces its own gene expression13, and also the ADAM17 gene14, generating a vicious ADAM17/TNFα feed-forward inflammatory loop that worsens multi-organ injury9. In fact, in the severe inflammation of LPS-induced endotoxic shock in mice, exclusive ablation of the ADAM17 gene in mouse myeloid cells is sufficient to markedly reduce mortality rates15.
In the vasculature, TNFα increases local inflammation aggravating VSMC osteogenic differentiation7, and also, nSMase2 gene expression and activity16, an essential determinant of aging-induced inflammation, atherosclerosis and VC10,16,17. In fact, nSMase2 gene ablation or pharmacological inhibition of nSMase2 activity markedly reduces age-enhanced inflammation in health10, the higher propensity for atherosclerotic lesions in the ApoE−/− mouse17 and, more significantly for CKD, the release of exosomes initiating medial calcification16.
Based upon current therapeutic limitations to lower hyperphosphatemia in CKD18, and considering that orally administered yeast ß-glucans efficaciously reduce multi-organ injury and mortality rates in LPS-challenged rats,19 this study was designed to examine whether an anti-inflammatory strategy with β-glucans could effectively attenuate systemic inflammation in health, including the mediator of early pro-senescent/pro-inflammatory interferon like responses, activated by the cGAS/STING pathway of cytosolic dsDNA recognition20 and also, renal and vascular injury predisposing to VC in a rat model of hyperphosphatemic rat CKD.
To test this hypothesis, among the multiple natural sources for β-glucans, we chose barley β-glucans (Bβglucans), based upon the safety of the FDA recommended daily intake of 3 to 5 g for their cholesterol21,22 lowering capacity. Specifically, we evaluated the efficacy of dietary Bβglucans to reduce: (a) Systemic inflammation in healthy adults with normal renal function; (b) Renal and aortic inflammation and the propensity for CKD progression and aortic calcium deposition in a rat model of hyperphosphatemic CKD, and (c) The contribution of Bβglucans regulation of ADAM17 and nSMase2 expression to their anti-inflammatory/anti-calcifying actions in vivo and in vitro in VSMC.
Results
Human study. Systemic anti-inflammatory actions by dietary BßGlucans
The daily intake of 3 g of BßGlucans, as barley bread by 10 individuals with normal renal function during 4 weeks, decreased circulating leukocyte mitochondrial superoxide production by 50% in one week (Supplementary Fig. 1).
In the 5 volunteers that completed the 3 g daily intake of BßGlucans for one month, the 50% inhibition of leukocyte superoxide production in the first week remained up to the end (week 4) (Fig. 1a). Furthermore, dietary BßGlucans also decreased leukocyte median mRNA levels of three recognized inflammatory markers: ADAM17 (80% at week 1 and 94% at week 4; p < 0.05) (Fig. 1b), TNFα (64% (not significant) at week 1 and 80% at week 4, p < 0.05) (Fig. 1c) and nSMase2 (58% at week 1 and 77% at week 4; p < 0.05) (Fig. 1d). These BßGlucans actions in circulating leukocytes after 1 or 4 weeks of a daily intake occurred despite no increases in serum levels either of β-glucans or in markers of bone and mineral homeostasis (calcium, phosphate, PTH, soluble klotho) or of systemic inflammation (C reactive protein and pro-inflammatory cytokines, except for a significant reduction of γ-interferon by week 4) (Table 1).
Figure 1.

Dietary Bßglucans inhibit mitochondrial superoxide production, ADAM17, TNFα, nSMase2 and STING gene expression in circulating leukocytes from healthy adults. Box plot analyses of changes in baseline superoxide production (a), ADAM17 (b), TNFα (c), nSMase2 (d) and STING (e) mRNA levels (each shape represents one individual) in peripheral blood monocytes from 5 healthy adults ingesting 3 g of Bßglucans daily, as slices of barley bread, during 4 weeks. *p < 0.05 and **p < 0.01 vs. Baseline, #p < 0.05 vs. 1week. Correlation between leukocyte STING mRNA and serum γ-interferon levels (f) and between leukocyte STING and nSMase2 mRNA levels (g). Each shape represent baseline (●), one week (■) and 4 weeks (▲) values.
Table 1.
Serum biochemistries in healthy volunteers.
| Baseline | 1 week BßGlucans | 4 weeks BßGlucans | |
|---|---|---|---|
| (n = 5) | (n = 5) | (n = 5) | |
| Serum ßglucans | |||
| (1,3)-β-D-glucan (pg/mL) | Undetectable | Undetectable | Undetectable |
| Renal function | |||
| Creatinine (mg/dL) | 0.86 ± 0.10 | 0.89 ± 0.12 | 0.90 ± 0.16 |
| Urea (mg/dL) | 23.9 [21.9–52.3] | 26.3 [25.6–39.1] | 26.8 [24.2–29.9] |
| Albumin (g/L) | 46.02 ± 2.57 | 45.3 ± 1.48 | 44.46 ± 2.15 |
| Mineral homeostasis | |||
| Calcium (mmol/L) | 2.37 ± 0.08 | 2.32 ± 0.07 | 2.32 ± 0.05 |
| Phosphorus (mmol/L) | 1.29 ± 0.18 | 1.18 ± 0.16 | 1.14 ± 0.23 |
| PTH (pg/dL) | 56.00 [36.40–58.40] | 51.70 [33.00–55.90] | 31.9 [30.9–43.2] |
| Klotho (pg/mL) | 925.33 [809.83–1,003.17] | 873.17[749.83–1,001.50] | 868.00 [819.83–1,010.83] |
| Inflammation | |||
| C Reactive Protein (mg/dL) | 0.11 [0.04–0.22] | 0.05 [0.04–0.10] | 0.10 [0.04–0.13] |
| IL-2 (pg/mL) | 1.37 ± 0.21 | 1.54 ± 0.26 | 1.47 ± 0.48 |
| IL-6 (pg/mL) | 4.31 ± 0.50 | 2.91 ± 1.52 | 3.55 ± 1.05 |
| TNFα (pg/mL) | 5.18 ± 3.15 | 5.20 ± 4.00 | 3.86 ± 1.47 |
| γ-INFγ (pg/mL) | 7.05 ± 1.28 | 6.31 ± 2.28 | 3.71 ± 1.73* |
| VCAM-1 (ng/mL) | 2,180.68 ± 117.97 | 2,130.27 ± 34.16 | 2,246.75 ± 139.80 |
Values indicate Mean ± SD or Median (interquartile range). *p < 0.05 vs. Baseline.
Dietary BßGlucans-driven reductions in serum γ-interferon levels were paralleled by significant decreases in leukocyte STING mRNA (r = 0.54; p = 0.05; Fig. 1f). In fact, leukocyte STING mRNA levels decreased by 90% (p < 0.01) by week 4 (Fig. 1e) of dietary BßGlucans intake. Furthermore, this reduction strongly correlated with that of leukocyte nSMase2 gene expression (r = 0.80; p = 0.001; Fig. 1g).
Animal study. Renal anti-inflammatory actions by dietary BßGlucans
The administration of the high P diet containing BßGlucans to uremic rats had no effect on daily food consumption (20 g/day) or body weights (CKD: 234 ± 13 g to 248 ± 23 g; n = 13; CKD + Bßglucans: 237 ± 9 g to 255 ± 18 g; n = 13). Furthermore, despite undetectable serum (1,3)-ß-D-glucan levels, the CKD + Bßglucans group showed better renal function compared to the CKD group (lower serum creatinine, BUN, proteinuria and higher renal klotho) (Table 2), as well as reduced inflammatory markers (lower renal inflammatory cell infiltration (Table 2), ADAM17 protein and mRNA (Fig. 2a,b) and nSMase2 mRNA) (Fig. 2c). Serum and urinary TNFα levels were undetectable in both dietary groups.
Table 2.
Biomarkers of renal dysfunction in rat CKD.
| CKD | CKD + BßGlucans | p value | |
|---|---|---|---|
| (n = 13) | (n = 13) | ||
| Serum Creatinine (mg/dL) | 1.0 ± 0.2 | 0.8 ± 0.2 | 0.03 |
| BUN (mg/dL) | 52.9 ± 9.6 | 44.3 ± 7.2 | 0.02 |
| Phosphaturia (nmol/24 hours) | 1.9 ± 0.5 | 2.3 ± 0.6 | 0.15 |
| Proteinuria (mg/24 hours) | 101.1 ± 80.0 | 21.4 ± 32.1 | 0.04 |
| α-Klotho (IOD/Area) | 1,477.2 ± 459.7 | 3,197.0 ± 1839.4 | 0.02 |
| Lymphocyte infiltration (% Area) | 2.2 ± 2.4 | 1.0 ± 0.7 | 0.04 |
Values indicate Mean ± SD.
Figure 2.

Renoprotection by dietary Bßglucans in hyperphosphatemic rat CKD. (a) Representative immunohistochemical images of renal ADAM17 from 5/6NX rats fed a high P diet with none (CKD; n = 13) or 2 mg of Bßglucans/g diet (CKD + Bßglucans; n = 13) during 4 weeks (Inset bars indicate relative scale); (b) Quantification of immunostaining. Bars and error bars represent mean ± SD of Histoscores values for all rats in both dietary groups; (c) Box plot analysis of nSMase2 gene expression in kidneys from rats described in (a). **p < 0.01 and ***p < 0.001 vs. CKD.
Regarding bone and mineral parameters, there were no significant differences in serum calcium, phosphate, PTH, 25-hydroxyvitamin D, FGF23 and bone alkaline phosphatase between groups (Table 3). The higher renal klotho found in the CKD + Bßglucans group (Table 2) did not result in increases in 24 hours phosphaturia (Table 3).
Table 3.
Serum biomarkers of bone and mineral metabolism in rat CKD.
| CKD | CKD + BßGlucans | p value | |
|---|---|---|---|
| (n = 13) | (n = 13) | ||
| Calcium (mg/dL) | 9.8 ± 0.5 | 9.2 ± 0.8 | 0.19 |
| Phosphate (mg/dL) | 8.1 ± 2.6 | 7.7 ± 2.2 | 0.66 |
| PTH (pg/mL) | 3107 ± 2152 | 2044 ± 1127 | 0.14 |
| 25-hydroxyvitamin D (ng/mL) | 29.4 ± 8.7 | 27.9 ± 6.5 | 0.63 |
| FGF23 (ng/mL) | 10.9 ± 5.6 | 12.5 ± 6.5 | 0.58 |
| Bone alkaline phosphatase (U/mL) | 42.1 (33.3–70.2) | 28.1 (16.9–74.0) | 0.28 |
| (1,3)-β-D-glucan (pg/mL) | Undetectable | Undetetectable | — |
Values indicate Mean ± SD or Median (interquartile range).
Animal study. Vascular anti-inflammatory actions by BßGlucans
The uremic rats fed the high P diet with Bßglucans had significantly lower median aortic ADAM17 protein (47%) (Fig. 3a,b), ADAM17 and TNFα mRNA levels (80%) (Fig. 3c) and nSMase2 mRNA levels (65%) and activity (55%) compared with the CKD group (Fig. 3d). Furthermore, in the aortas of these uremic rats, TNFα mRNA levels correlated directly with ADAM17 mRNA (r = 0.81; p < 0.001) and also with nSMase2 mRNA levels and activity (r = 0.89; p < 0.001 and r = 0.81; p < 0.01, respectively).
Figure 3.

Anti-inflammatory vascular protection by dietary BßGlucans. (a) Representative immunostaining for ADAM17 (Inset bars indicate relative scale) in thoracic aortas from 5/6NX rats fed a high P diet with none (CKD; n = 13) or 2 mg of Bßglucans/g diet (CKD + Bßglucans; n = 13) during 4 weeks. (b) Quantification of aortic ADAM17 immunostaining. Box plots represent median (interquartile range) of OD/Area values for all rats in both dietary groups. (c) Aortic ADAM17 and TNFα gene expression; (d) Aortic nSMase2 mRNA (white bars; CKD n = 9; CKD + Bßglucans n = 7) and activity (black bars; CKD n = 13; CKD + Barley n = 11). R.U.: Relative units. Bars and error bars represent mean ± SD; *p < 0.05 vs. CKD.
Animal study. Vascular anti-calcifying actions by dietary BßGlucans
In the CKD group, 36% of the aortas were Von Kossa positive while all aortas from the CKD + Bßglucans group stained negatively (Fig. 4a). Furthermore, median aortic total calcium was 8 times lower in the CKD + Bßglucans group (7.5 µg Ca/mg protein) compared to uremic controls (CKD group: 60 µg Ca/mg protein) (Fig. 4b). Despite the higher Ca deposition found in the CKD group, there were minor changes in median aortic mRNA levels of osteogenic differentiation markers (reductions in α-actin and increases in Runx2 and Osterix) compared to the CKD + Bßglucans group, which did not reach statistical significance (Table 4). Instead, total aortic calcium content strongly correlated with increases in nSMase2 activity (r = 0.60; p < 0.01).
Figure 4.

Anti-calcifying protection by dietary BßGlucans. (a) Representative calcium deposition measured by Von Kossa (black) staining (Inset bars indicate relative scale) in thoracic aortas from 5/6NX rats fed a high P diet with none (CKD; n = 13) or 2 mg of Bßglucans/g diet (CKD + Bßglucans; n = 13) during 4 weeks. (b) Quantification of calcium deposition in thoracic aortas described in (a); boxplot analysis of changes in calcium content, *p < 0.05 vs. CKD.
Table 4.
Biomarkers of vascular osteogenic differentiation in rat CKD.
| CKD | CKD + BßGlucans | p value | |
|---|---|---|---|
| α-actin mRNA (R.U.) | 0.97 (0.69–1.45) | 1.20 (0.37–1.41) | 0.696 |
| N = 12 | N = 9 | ||
| Runx2 mRNA (R.U.) | 1.67 (1.11–2.58) | 1.51 (0.9–1.91) | 0.481 |
| N = 10 | N = 11 | ||
| Osterix mRNA (R.U.) | 0.72 (0.47–1.13) | 0.53 (0.47–0.73) | 0.374 |
| N = 12 | N = 9 |
Values indicate Median (interquartile range).
In vitro studies. Direct anti-inflammatory/anti-calcifying actions by BßGlucans in VSMC
Protocol 1 searched for an effective anti-inflammatory dose of BßGlucans extracts in the murine monocyte cell line Raw 264.7, exposed to 5 µg/mL of E. Coli LPS for 16 hours to reproduce severe systemic inflammatory stimuli.
Exposure to LPS increased mitochondrial superoxide production by 2-fold (Supplementary Fig. 1). A dose of 100 µg/mL of commercial BßGlucans extracts was necessary to fully prevent LPS-induced superoxide production. In unstimulated monocytes, the 100 µg/mL dose of BßGlucans extracts elicited anti-inflammatory actions as it reduced baseline monocyte superoxide production by 50% (Supplementary Fig. 1).
Protocol 2 examined the impact of LPS-driven inflammation on VSMC phenotype. In A7r5 cells, the exposure to 100 µg of BßGlucans/mL for 16 hours had no effect on basal nSMase2, TNFα or ADAM17 gene expression (Fig. 5a–c). A7r5 exposure to LPS for 16 hours sufficed to significantly increase nSMase2, TNFα and ADAM17 gene expression (Fig. 5a–c) above the levels in unstimulated controls. Furthermore, the combination of the same dose of LPS with 100 µg/mL Bßglucans fully prevented LPS-driven increases in nSMase2 gene expression maintaining nSMase2 mRNA at the levels of the control group, failed to counteract LPS-induction of TNFα and decreased LPS-induced ADAM17 gene expression only marginally (Fig. 5a–c).
Figure 5.

Bßglucans attenuate the vascular smooth muscle cell inflammation and osteogenic differentiation induced by LPS. Gene expression of nSMase2 (a), TNFα (b), ADAM17 (c), α-actin (d), Runx2 (e) and Osterix (f) in A7r5 cells exposed to control medium (CONTROL), 100 μg/mL of Bßglucans (β-GL), 5 µg/mL of LPS (LPS) or the combination of 100 μg/mL of Bßglucans and 5 µg/mL of LPS (LPS + ß-GL) for 16 hours. Bars and error bars represent mean ± SD from three independent experiments, each performed in triplicate per experimental condition. R.U.: relative units. **p < 0.01 and ***p < 0.001 vs. Control, #p < 0.05 and ###p < 0.001 vs. LPS.
Regarding osteogenic differentiation, A7r5 exposure to 100 µg of BßGlucans/mL for 16 hours had no effect on basal mRNA levels of α-actin, Runx2 or osterix. Instead, while LPS exposure significantly decreased A7r5 levels of α-actin and increased Runx2 and Osterix gene expression compared to those in the control group (Fig. 5d–f), the combination of LPS with 100 µg/mL Bßglucans attenuated the stimulatory effect of LPS maintaining the osteogenic differentiation markers at levels similar to those in the control group (Fig. 5d–f).
Protocol 3 examined the impact of uremia-driven inflammation on VSMC phenotype. The exposure of A7r5 to uremic serum from hyperphosphatemic rats for 16 hours significantly increased TNFα and ADAM17 gene expression. However, the almost 2-fold increase in nSMase2 gene expression was not statistically significant (Fig. 6a–c). By contrast, exposure of A7r5 cells to uremic serum + Bßglucans maintained the mRNA of the three markers of inflammation at levels similar to those in the cells exposed to control serum, which was not affected by exposure to Bßglucans (Fig. 6a–c).
Figure 6.

Bßglucans attenuate vascular smooth muscle cell inflammation and osteogenic differentiation induced by uremic conditions. Gene expression of nSMase2 (a), TNFα (b), ADAM17 (c), α-actin (d), Runx2 (e) and Osterix (f) in A7r5 cells exposed to serum from rats with normal renal function (Control), with or without 100 μg/mL of Bßglucans (β-GL), or exposed to serum from rats with 14 weeks of uremia fed a high phosphorus diet (Uremic) with or without 100 μg/mL of Bßglucans for 16 hours. Bars and error bars represent mean ± SD from three independent experiments, each performed in triplicate per experimental condition. R.U.: relative units. **p < 0.01 and ***p < 0.001 vs. Control, ##p < 0.01, ###p < 0.001 and ####p < 0.0001 vs. Uremic.
In A7r5 cells exposed to normal serum, Bßglucans had no effect on basal levels of the osteogenic markers. Instead, the uremic serum only increased Runx2 gene expression with no significant changes in either α-actin or Osterix (Fig. 6d–f). However, the addition of 100 µg/mL of Bßglucans to the uremic serum prevented the increases in Runx2 gene expression maintaining its levels at basal values.
In vitro studies. Direct anti-calcifying and anti-nSMase2 actions by BßGlucans
In Protocol 4, A7r5 cells were exposed exclusively to calcifying media (CM: 2 mM Ca; 3 mM P) during 4 days, which increased calcium deposition by 13-fold. The addition of BßGlucans to the CM markedly attenuated (by 72%) the increases in calcium deposition induced by the CM (Fig. 7a), prevented the decrease in α-actin and the increases in osteogenic Osterix and Runx2 mRNA levels (Fig. 7b).
Figure 7.
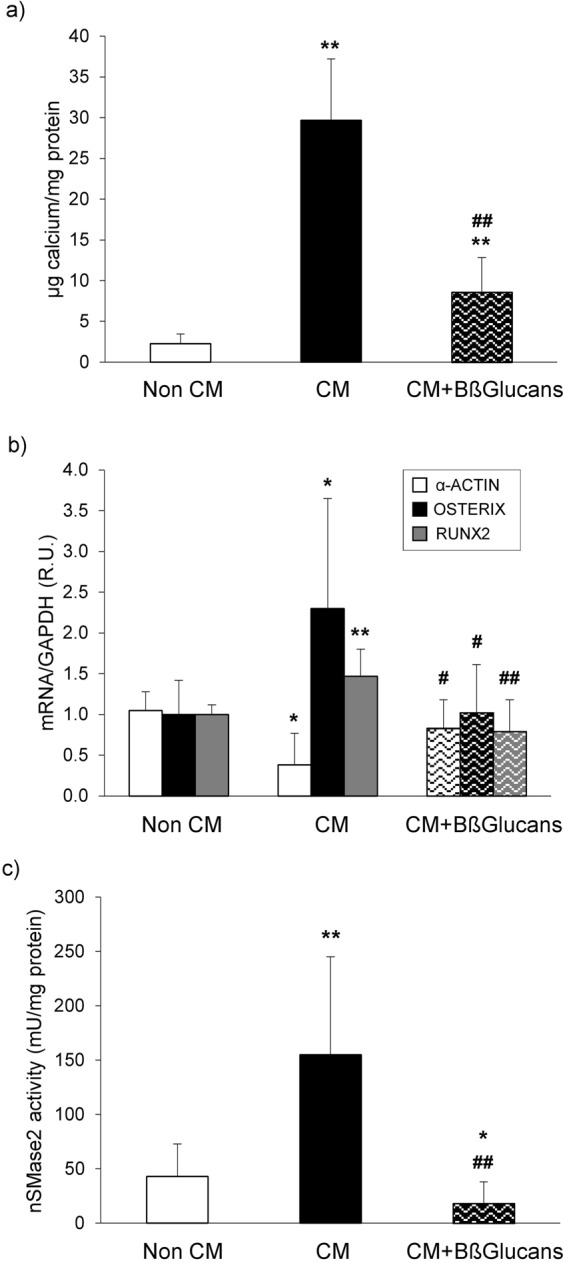
Bßglucans attenuate calcium deposition by inhibiting vascular smooth muscle cell osteogenic differentiation and nSMase2 activity. (a) Calcium deposition in A7r5 cells exposed to non-calcifying media (Non CM: 1 mM Ca; 1 mM P) or calcifying media (CM: 2 mM Ca; 3 mM P) with 0 or 100 μg/mL of Bßglucans (CM + β-GL) during 4 days. (b) Gene expression of α–actin (white bars), osterix (black bars) and Runx2 (gray bars) in A7r5 cells treated as described. (c) nSMase2 activity in A7r5 cells treated as described. Bars and error bars represent mean ± SD from three independent experiments performed in triplicate per experimental condition. R.U.: relative units. *p < 0.05 and **p < 0.01 vs. Non CM, #p < 0.05 and ##p < 0.01 vs. CM.
Similar results were found in ex vivo experiments culturing aortic rings from normal rats under the same calcifying conditions used in A7r5 (data not shown).
In addition, in A7r5 cells exposed to CM, the activity of nSMase2 increased by 3-fold, an induction totally prevented by adding 100 µg/mL of BßGlucans to the CM (Fig. 7c).
These direct anti-calcifying actions of BßGlucans extracts occurred in cells with undetectable mRNA levels of the β-glucan receptor Dectin 1 (not shown) and involved β-glucan internalization into A7r5 cells, as demonstrated by significant increases in intracellular BßGlucans levels from undetectable in cells exposed to the Non CM or CM alone to 2.3 ng/well (p < 0.01) in cells exposed to the CM + Bßglucans.
Discussion
This work presents unprecedented properties of dietary Bβglucans of high translational relevance to attenuate systemic inflammation in health, as well as the progression of renal and vascular damage predisposing to VC in hyperphosphatemic rat CKD. These Bβglucans actions extend beyond the inhibition of leukocyte superoxide production and of STING/interferon-like pro-aging/proinflammatory signals that precede any elevation of systemic inflammatory markers above normal levels, and involve a marked suppression of leukocyte, renal and vascular ADAM17 and nSMase2.
Bßglucans control of systemic inflammation in health
In healthy adults, the daily intake of 3 g of Bβglucans, recommended by the FDA to lower serum cholesterol21,22 could rapidly and efficaciously suppress systemic inflammation, a recognized inducer of VC. Indeed, dietary Bβglucans markedly reduced circulating leukocyte superoxide production (50%) as well as leukocyte ADAM17 and nSMase2 gene expression (60%) after a week intake, despite unchanged serum levels of CRP, TNFα and several other inflammatory cytokines, all within the normal range, and undetectable serum β-glucans concentrations. Despite the small size of this population of adults with normal renal function, Bβglucans rapid inhibition of these two pro-inflammatory genes in circulating leukocytes supports their potential to prevent/attenuate ADAM17-driven multi-organ injury and mortality rates9, and also nSMase2-driven age-enhanced inflammation regardless of serum TNFα10 and the propensity for atheromatous lesions17, as conclusively demonstrated in experimental models of ADAM17 or nSMase2 gene ablation.
Significantly, by week 4, dietary Bβglucans exerted a stronger (80 to 90%) inhibition of both ADAM17 and nSMase2 gene expression, and also markedly inhibited leukocyte TNFα mRNA levels, despite no further reductions of either of their inducers: superoxide production and serum TNFα. Thus, dietary Bβglucans inhibition of leukocyte ADAM17, nSMase2 and TNFα gene expression involved mechanisms other than suppressing oxidative stress induction of ADAM178,14 and nSMase2 gene expression10,23, or TNFα-self up-regulation13. Significantly, among the inflammatory cytokines measured, dietary BßGlucans markedly reduced only circulating γ-interferon levels by week 4. This finding led us to evaluate whether dietary BßGlucans could also attenuate interferon-likepro-senescence/pro-inflammatory responses initiated by activation of the cGAS/STING pathway20,24,25. We found that serum γ-interferon not only strongly correlated with leukocyte STING mRNA but, more significantly, dietary BßGlucans caused a 90% reduction of leukocyte STING mRNA by week 4. Thus, leukocyte STING emerged as a novel sensitive biomarker of leukocyte pro-aging/pro-inflammatory features, as its reductions by dietary BßGlucans precede any elevations above normal in circulating levels of inflammatory cytokines, including γ-interferon. Furthermore, the strong correlation between leukocyte STING and nSMase2 mRNA levels suggests that dietary BßGlucans´progressive and simultaneous suppression of leukocyte STING and nSMase2 gene expression could contribute to attenuate the age-enhanced inflammation attributed to increases in nSMase2 in health. Indeed, enhanced nSMase2 release of exosomes from leukocytes carrying a cGAS/STING-driven senescence/inflammation-associated secretory phenotype would propagate these signals to healthy neighboring cells26. Even though the accuracy of these early biomarkers of the anti-aging, anti-inflammatory actions of BßGlucans was only identified in the 5 normal individuals who completed the 4 weeks of BßGlucans intake, the strength of the leukocyte STING/nSMase2 association (r2 = 0.65) provides a solid base for early antinflammatory actions that need to be further corroborated in larger cohorts and long-term studies.
Bßglucans renal and vascular protection in hyperphosphatemic CKD
In hyperphosphatemic rat CKD, a daily intake of 40 mg of Bβglucans, as barley flour, effectively reduced inflammation-driven renal injury by targeting renal elevations in ADAM17, an enzyme induced by CKD of all etiologies12 and also by TNFα14 and oxidative stress8, despite undetectable serum β-glucan levels. The higher renal ADAM17 in uremic controls is sufficient to explain their increased inflammatory cell infiltration and 24 h proteinuria, as demonstrated by Lautrette and co-workers12. In turn, the higher proteinuria of uremic controls could mediate the faster decline in GFR, as recently reported27, and corroborated herein by the higher serum creatinine and BUN in the CKD group.
Dietary Bβglucans also reduced renal nSMase2 gene expression, which could extend the renoprotective benefits of this nutritional strategy to hypertensive individuals28, by counteracting angiotensin II-driven-increases in renal nSMase2 or to diabetic nephropathy, by reducing the release to the urine of megalin loaded exosomes29.
Bβglucans simultaneous attenuation of CKD progression and renal inflammatory cell infiltration, the two most potent down-regulators of renal α-klotho content5,30, resulted in a higher renal α-klotho in the CKD + Bβglucans group. However, phosphaturia was similar between dietary groups, which suggested that the higher renal klotho was insufficient to ameliorate kidney resistance to FGF23 and, consequently, the deleterious renal impact of phosphate retention.
In the inflamed aortas from these uremic rats, TNFα mRNA levels correlated directly with ADAM17 mRNA, partly reflecting the expected TNFα induction of ADAM179 and of its own gene13,14, and also with nSMase2 mRNA levels and activity. Dietary Bβglucans suppressed by 80% the self-perpetuating aortic ADAM17/TNFα-inflammatory loop9 and prevented the 3-fold elevations in nSMase2 reported to prompt pro-calcifying exosome release16, thereby reducing aortic calcium deposition in the CKD + Bβglucans group by 8-fold compared to that in uremic controls. Interestingly, the higher calcium in uremic controls was unrelated to changes in the expression of markers of osteogenic differentiation and strongly correlated only with increased nSMase2. Thus, our aortic findings support the evidence that exosomes released by nSMase2 are the earliest calcified particles preceding overt calcification31, and also, that there are differential increases in vascular nSMase2 release of calcified and noncalcified exosomes causing heterogeneous calcium deposition at sites of early calcification31,32. Furthermore, Von Kossa staining was positive only in 36% of the aortas from uremic controls supporting that overt calcification is a late event in the course of CKD. Significantly, dietary Bßglucans fully prevented overt calcification, as none of the aortas in the barley group was Von Kossa positive despite no amelioration of systemic calcium and phosphate homeostasis.
Importantly, a Bβglucans intake effective at reducing aortic nSMase2 gene expression and activity had no adverse impact on bone mineralization, as estimated by serum bone specific alkaline phosphatase levels. This is an important translational consideration to design therapeutic anti-nSMase2 strategies in CKD because the nSMase2 null mouse presents severe bone and dental mineralization defects33,34.
Mechanisms for Bßglucans´ actions
An intriguing aspect of Bβglucans potent anti-inflammatory, anti-ADAM17, anti-nSMase2, renal and vascular protective actions in vivo, is that they occurred with undetectable serum βglucans levels. Indeed, part of these actions may not require intestinal Bβglucans absorption because oral Bβglucans induce a saccharolytic shift in the microbiota that augments serum levels of anti-inflammatory short chain fatty acids, and also reduces circulating levels of p-cresyl-sulfate35, a uremic toxin promoting renal and cardiovascular injury36,37.
However, our studies in vitro in murine monocytes and VSMC exposed to commercially available Bβglucans extracts (95% purity) demonstrated direct anti-inflammatory, anti-calcifying actions that support the pharmacokinetics of dietary Bβglucans absorbed in the gut38–40. Specifically, in murine monocytes, we first identified that a dose of 100 µg/mL of Bβglucans extracts was necessary to prevent the 2-fold increases in mitochondrial superoxide production induced by a 16 h exposure to an LPS challenge (5 µg/mL). Significantly, this anti-inflammatory dose of Bβglucans also reduced by 50% the superoxide production in unstimulated monocytes, thus reproducing the 50% inhibition exerted in normal subjects after a week, or a month, of a daily intake of 3 g of Bβglucans.
Furthermore, the exposure of A7r5 cells to LPS (5 µg/mL for 16 h) corroborated that severe inflammation rapidly induced a switch to an osteogenic phenotype, as the increases in the mRNA levels for ADAM17, nSMase2 and TNFα were associated to decreases in α-actin and increases in the osteogenic markers Runx2 and osterix. Importantly, the simultaneous exposure of A7r5 cells to LPS and the effective anti-inflammatory dose of Bβglucans in immune cells, fully prevented LPS-induced inflammatory and osteogenic phenotypic changes without affecting VSMC baseline phenotype.
A7r5 cells also responded rapidly to uremic stimuli (serum from hyperphosphatemic rat CKD for 16 h) with significant elevations in ADAM17, TNFα and Runx2, but without significant increases in nSMase2 and osterix or decreases in α-actin. Nonetheless, Bβglucans fully prevented all uremia-induced changes, thus maintaining baseline levels of all of these inflammatory/osteogenic markers. These findings support that a threshold for inflammatory or uremic stimuli is necessary to increase nSMase2 and osteogenic markers to levels resulting in overt calcification.
In VSMC exposed exclusively to prolonged (4 days) high Ca/high P conditions, Bβglucans extracts decreased by 60% nSMase2 gene expression in the absence of any inflammatory or uremic stimuli, through a process involving β1,3-glucan internalization into VSMC that is Dectin1-independent, as A7r5 cells showed undetectable Dectin1 mRNA levels. Several β-glucans receptors expressed in VSMC (Complement Receptor 3 or TLR2/6)41,42 could mediate Bβglucans actions in A7r5 cells.
The internalization of (1,3)β-D-glucans into A7r5 cells exposed to Bβglucans extracts may also occur in other Bβglucans target cells, which could explain in part the systemic, renal and vascular anti-inflammatory/anti-nSMase2 actions of dietary Bβglucans in healthy adults and in hyperphosphatemic rat CKD despite undetectable serum levels. Indeed, there is evidence that soluble β1,3-glucose polymers, generated from oat and barley grains by the highly predominant β1,3-β1,4-glucanase in human gut bacteria43, are internalized and further processed by intestinal epithelial and immune cells to shorter β1,3-glucans. In turn, the shorter β1,3-glucans released to the circulation39 can be re-internalized into immune cells reducing serum levels while enhancing leukocyte systemic anti-oxidant/anti-inflammatory capacity41,44, as demonstrated herein in healthy adults.
Barley is the least expensive cereal grain. Therefore, dietary Bβglucans health benefits could reach the poorest populations of CKD patients and of healthy adults with systemic pro-aging or inflammatory problems and, therefore, at a higher risk for renal and vascular injury.
Prospective clinical trials are mandatory to evaluate the sensitivity and accuracy of leukocyte ADAM17, nSMase2 or TNFα mRNA and STING levels to personalize dietary Bβglucans interventions for efficacious systemic, renal and vascular protection in the general population and to attenuate CKD progression and the initiation of medial calcium deposition in human CKD.
Material and Methods
Ethics statements
Approval for the human study was obtained from the Institutional Review Board (Comité de Ética en Investigación Clínica del Hospital Universitario Central de Asturias), in compliance with the Declaration of Helsinki. All participants gave a written informed consent prior to their inclusion in the study.
Approval for the animal study was obtained from the Ethics Committee for Animal Experimentation at Lleida University in compliance with current international legislation for animal research.
All methods in the experimental protocols described below were carried out following the regulations for biomedical research of the University of Lleida and the University of Oviedo.
Human study
Ten volunteers (2 men, 8 women, age 20 to 60 y.o.), with normal renal function ingested daily during 4 weeks barley bread, manufactured with flour from barley grains selected for their high (1-3)(1-4)-β-D-glucans content (8%), to provide 3 g of BßGlucans (produced at Food Science and Technology Department, Lleida University). Blood (5 mL) was drawn at baseline and at week 1 and 4 to obtain plasma and circulating leukocytes (after red cell lysis). As systemic inflammation biomarkers, we measured leukocyte mitochondrial superoxide production by flow cytometry in freshly isolated leukocytes. In the five volunteers (1 man, 4 women) that complied with the daily intake for one-month, we also quantified the mRNA levels of TNFα, ADAM17, nSMase2 and STING, as described in the in vitro studies. Serum levels of (1, 3)β-Dglucans, cholesterol, glycemia, markers of renal function, calcium and phosphate homeostasis, and systemic inflammation were measured as specified in blood chemistries. The reason to abandon the study was the omission of the daily intake more than twice per week.
Animal study
Five-sixth nephrectomized (NX) female Sprague-Dawley rats (200–225 g) were fed during 4 weeks a high phosphate diet (HPD: 0.9% P; 0.6% calcium, Ca; Altromin) containing either 0 (CKD: n = 13) or 2 mg of BßGlucans/g diet (CKD + BßGlucans) from the barley flour described above, with no changes in dietary protein, P, carbohydrate or lipid content. The modified Megazyme method quantified final β-D-glucan content in the diet45.
In vitro studies
-
I.
BßGlucans anti-inflammatory actions were examined in the murine monocyte cell line Raw 264.7 and in the rat aortic VSMC line A7r5.
Protocol 1: Rested Raw264.7, synchronized at G0 by exposure to FBS-free DMEM medium for 6 h, were treated for 16 hours with 5 μg/mL LPS from Escherichia coli 0111:B4 (L4391, Sigma-Aldrich), 100 μg/mL of barley (1–3)(1–4)-β-D-glucans (G6513, Sigma-Aldrich; 95% purity) or both.
Protocol 2: A7r5 cells were exposed to 5 μg/mL LPS, 100 μg/mL of barley (1–3)(1–4)-β-D-glucans or both in DMEM + 1% foetal bovine serum (FBS) for 16 hours.
Protocol 3: To test uremia-driven inflammatory stimuli, A7r5 cells were exposed to either control serum (a pool from rats with normal renal function fed a normal P diet; DMEM + 10% control serum), or to uremic serum (a pool from rats with uremia of 14 weeks fed a high phosphorus diet; DMEM + 10% uremic serum), with or without 100 μg/mL of Bßglucans for 16 hours.
-
II.
BβGlucans anti-calcifying actions were examined in A7r5 cells and in aortic rings (1–2 mm) from normal rats.
Protocol 4: Aortic rings were washed in cold PBS containing P/S and then placed in fibronectin pre-coated (100 µg/mL) 6-well plates (8 rings/well) with growing media. A7r5 and aortic rings were exposed during 4 days to either non-calcifying medium (Non CM: DMEM-F12 + 0.1%BSA, 1 mM Ca, 1 mM P) or to calcifying medium (CM: DMEM-F12 + 0.1% BSA, 2 mM Ca, 3 mM P) with 0 or 100 μg/mL of BßGlucans. A7r5 levels of the β-glucan receptor Dectin1 were examined by qPCR and intracellular levels of (1–3) β-D-glucans as indicated in blood chemistries.
Blood chemistries and proteinuria
Human biochemical parameters were measured at the Medicine Laboratory of the Hospital Universitario Central de Asturias using Cobas 8000 (Roche Diagnostics) Module c702, for most parameters, and Module e801 for PTH immunoassay.
For animal studies spectrophotometry and immunoassay (Cobas 8000, Roche Diagnostics) were used to measure serum levels of Ca, P, creatinine and 25-hydroxyvitaminD (25(OH)D). QuantiChromTM Urea Assay Kit measured Blood Urea Nitrogen (BUN, BioAssay System). ELISA Kits were used to measure rat intact PTH (Immutopics), rat fibroblast growth factor 23 (FGF23; EMD Millipore), rat bone alkaline phosphatase (Biosource), blood and urinary rat TNFα (Abcam plc, Cambridge, UK). Serum human TNFα was measured by bead-based multiplex assay (BiolegendPlex, Biolegend, Germany) analyzed in a FACS Canto II flow cytometer (BD Biosciences) equipped with a FACS Diva 6.5 software, following manufacturers´ protocols. Test strips measured urinary protein (SIEMENS MULTISTICK 10SG, Analyticon Biochemistry). The FungitelTM kit (Associates of Cape Code, Inc) measured serum, plasma and intracellular (1,3)β-D-glucans.
Histological analyses
Rat renal and aortic 5 µm-paraffin sections were deparaffinized and hydrated. Renal ADAM17 and CD45 (leukocyte infiltration) immunostainings were performed as in46 and renal α-klotho with kit-CTS008 (R&D System). For aortic ADAM17 immunofluorescence nuclei were counterstained with Hoechst. For renal ADAM17, sections were counterstained with hematoxylin-eosin. Each slide had its negative control (no primary antibody). Supplementary Table 1 lists primary antibodies and dilutions. Quantifications used ImageJ or Histoscores47.
Mitochondrial superoxide production
Circulating human leukocytes and cultured Raw 264.7 cells were washed with PBS, resuspended in 5 µM superoxide sensitive probe MitoSOXTM (Thermo-Fisher Scientific) in PBS with either vehicle or the corresponding treatment for 10 min at 37 °C, protected from light and washed twice with PBS. A FACS CantoTMII flow cytometer (Becton Dickinson Bioscience) measured superoxide anion levels.
Von kossa staining
Deparaffinized, hydrated rat aorta sections were incubated in 5% silver nitrate before the revealing solution. Slides placed in 2% sodium thiosulfate were counterstained with nuclear fast red.
Total calcium measurements
A 20 mm segment of the abdominal rat aorta proximal to the iliac bifurcation was first grinded in liquid nitrogen and then decalcified with 0.6 N HCl. A7r5 cells, washed thrice with PBS, were decalcified with 0.6 N HCl.
Samples were shaken gently at 4 °C for 24 h. Upon centrifugation, O-cresolphthalein-complexone measured total calcium in supernatants. Pellets were re-suspended in lysis buffer (0.1 N NaOH, 0.1% SDS) for protein extraction and quantification (Lowry method, Bio-Rad).
For rat aortic rings, total calcium was measured as above in cell pellets from RNA extraction (below) and expressed as µg calcium/µg RNA.
Sphingomyelinase2 activity
The Amplex Red Sphingomyelinase assay kit (Molecular Probes, Invitrogen) was used in aorta sections and in A7r5 whole cell extracts.
Quantitative PCR
Total tissue or cell RNA, extracted with TRI reagent (Sigma-Aldrich), was reversed transcribed with a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems). Quantitative-real time PCR (qPCR) reactions used the Stratagene Mx3005P QPCR System (Agilent Technologies), Fast Start Universal Probe Master (Roche), pre-developed assays (Thermo-Fisher Scientific) and ΔΔCt quantification48.
Statistical analysis
T tests, ANOVA or Kruskal-Wallis with Bonferroni post-hoc test examined statistical differences between groups. Results are expressed as mean ± SD unless otherwise stated. Statistical analyses used GraphPad Prism, SPSS 17.0 for Windows or R.
Supplementary information
Acknowledgements
Authors thank Dr. Roxana Savin (Agrotechnio Research Center, Lleida University) for generating barley grain with high (8%) β-glucan content by multiplying lines selected for this trait (provided by Semillas Batlle, Spain) and for valuable suggestions for project design; Ana Martínez (IRB Lleida) for diet-chow preparation and Xenia Borràs (Food Technology Dept., Agrotecnio Research Center, Lleida University) for Barley bread preparation. A grant to A.S.D. and M.J.M. from IRBLleida and Agrotecnio Research collaborative projects from the Consell Social at Lleida University supported initial work, Instituto de Salud Carlos III and co-funded by European Union (ERDF/FEDER) (FIS PI11/00259, PI14/01452, PI17/02181), Plan de Ciencia, Tecnología e Innovación 2013–2017 y 2018–2022 del Principado de Asturias (GRUPIN14-028, IDI-2018-000152), RedInRen from ISCIII (ISCIII-RETIC REDINREN RD16/0009). Investigator support included: NC-L by GRUPIN14-028 and IDI-2018-000152, LM-A by GRUPIN14-028, SP by FICYT; MVA and PV by Educational Grant 2 A/2015 from ERA-EDTA CKD-MBD Working Group; PV and AC by ERA-EDTA fellowships 2011 and 2012; JR-C by MINECO (“Juan de la Cierva” program, FJCI-2015-23849); A.S.D. by Asociación Investigación de Fisiología Aplicada. A.S.D. and M.J.M. are members of the Campus Iberus (Ebro Valley Campus of International Excellence).
Author contributions
M.J.M., C.R., J.B.C.-A. and A.S.D. designed the study; M.V.A., N.C.-L., S.P., A.L.C.-G., P.V., C.U., A.C., C.Q.-C., L.M.-A., J.R.-C. and AS.D. performed experiments and analyzed results; M.V.A., N.C.-L. made figures; M.V.A., N.C.-L., S.P., C.M.-S., M.J.M., C.R., J.B.C.-A. and A.S.D. drafted and reviewed the manuscript. All authors approved the final version.
Data availability
No datasets were generated or analyzed during the current study.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Maria Vittoria Arcidiacono, Natalia Carrillo-López and Sara Panizo.
These authors jointly supervised this work: Jorge B. Cannata-Andía and Adriana S. Dusso.
Supplementary information
is available for this paper at 10.1038/s41598-019-54306-8.
References
- 1.Vervloet M, Cozzolino M. Vascular calcification in chronic kidney disease: different bricks in the wall? Kidney Int. 2017;91:808–817. doi: 10.1016/j.kint.2016.09.024. [DOI] [PubMed] [Google Scholar]
- 2.Slatopolsky E, Brown A, Dusso A. Role of phosphorus in the pathogenesis of secondary hyperparathyroidism. Am J Kidney Dis. 2001;37:S54–57. doi: 10.1053/ajkd.2001.20740. [DOI] [PubMed] [Google Scholar]
- 3.Paloian NJ, Giachelli CM. A current understanding of vascular calcification in CKD. Am J Physiol Renal Physiol. 2014;307:F891–900. doi: 10.1152/ajprenal.00163.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Navarro-Gonzalez JF, Mora-Fernandez C, Muros M, Herrera H, Garcia J. Mineral metabolism and inflammation in chronic kidney disease patients: a cross-sectional study. Clin J Am Soc Nephrol. 2009;4:1646–1654. doi: 10.2215/CJN.02420409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Izquierdo MC, et al. Klotho, phosphate and inflammation/ageing in chronic kidney disease. Nephrol Dial Transplant. 2012;27(Suppl 4):iv6–10. doi: 10.1093/ndt/gfs426. [DOI] [PubMed] [Google Scholar]
- 6.Massy ZA, Stenvinkel P, Drueke TB. The role of oxidative stress in chronic kidney disease. Semin Dial. 2009;22:405–408. doi: 10.1111/j.1525-139X.2009.00590.x. [DOI] [PubMed] [Google Scholar]
- 7.Zhao MM, et al. Mitochondrial reactive oxygen species promote p65 nuclear translocation mediating high-phosphate-induced vascular calcification in vitro and in vivo. Kidney Int. 2011;79:1071–1079. doi: 10.1038/ki.2011.18. [DOI] [PubMed] [Google Scholar]
- 8.Brill A, et al. Oxidative stress activates ADAM17/TACE and induces its target receptor shedding in platelets in a p38-dependent fashion. Cardiovasc Res. 2009;84:137–144. doi: 10.1093/cvr/cvp176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gooz M. ADAM-17: the enzyme that does it all. Crit Rev Biochem Mol Biol. 2010;45:146–169. doi: 10.3109/10409231003628015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rutkute K, Asmis RH, Nikolova-Karakashian MN. Regulation of neutral sphingomyelinase-2 by GSH: a new insight to the role of oxidative stress in aging-associated inflammation. Journal of lipid research. 2007;48:2443–2452. doi: 10.1194/jlr.M700227-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Melenhorst WB, et al. ADAM17 upregulation in human renal disease: a role in modulating TGF-alpha availability? Am J Physiol Renal Physiol. 2009;297:F781–790. doi: 10.1152/ajprenal.90610.2008. [DOI] [PubMed] [Google Scholar]
- 12.Lautrette A, et al. Angiotensin II and EGF receptor cross-talk in chronic kidney diseases: a new therapeutic approach. Nat Med. 2005;11:867–874. doi: 10.1038/nm1275. [DOI] [PubMed] [Google Scholar]
- 13.Tabibzadeh S, Satyaswaroop PG, von Wolff M, Strowitzki T. Regulation of TNF-alpha mRNA expression in endometrial cells by TNF-alpha and by oestrogen withdrawal. Mol Hum Reprod. 1999;5:1141–1149. doi: 10.1093/molehr/5.12.1141. [DOI] [PubMed] [Google Scholar]
- 14.Charbonneau M, et al. Hypoxia-inducible factor mediates hypoxic and tumor necrosis factor alpha-induced increases in tumor necrosis factor-alpha converting enzyme/ADAM17 expression by synovial cells. J Biol Chem. 2007;282:33714–33724. doi: 10.1074/jbc.M704041200. [DOI] [PubMed] [Google Scholar]
- 15.Horiuchi K, et al. Cutting edge: TNF-alpha-converting enzyme (TACE/ADAM17) inactivation in mouse myeloid cells prevents lethality from endotoxin shock. J Immunol. 2007;179:2686–2689. doi: 10.4049/jimmunol.179.5.2686. [DOI] [PubMed] [Google Scholar]
- 16.Kapustin AN, et al. Vascular smooth muscle cell calcification is mediated by regulated exosome secretion. Circ Res. 2015;116:1312–1323. doi: 10.1161/CIRCRESAHA.116.305012. [DOI] [PubMed] [Google Scholar]
- 17.Lallemand T, et al. nSMase2 (Type 2-Neutral Sphingomyelinase) Deficiency or Inhibition by GW4869 Reduces Inflammation and Atherosclerosis in Apoe(−/−) Mice. Arterioscler Thromb Vasc Biol. 2018;38:1479–1492. doi: 10.1161/ATVBAHA.118.311208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Habbous S, et al. The efficacy and safety of sevelamer and lanthanum versus calcium-containing and iron-based binders in treating hyperphosphatemia in patients with chronic kidney disease: a systematic review and meta-analysis. Nephrol Dial Transplant. 2017;32:111–125. doi: 10.1093/ndt/gfw312. [DOI] [PubMed] [Google Scholar]
- 19.Sandvik A, et al. Oral and systemic administration of beta-glucan protects against lipopolysaccharide-induced shock and organ injury in rats. Clin Exp Immunol. 2007;148:168–177. doi: 10.1111/j.1365-2249.2006.03320.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kreienkamp R, et al. A Cell-Intrinsic Interferon-like Response Links Replication Stress to Cellular Aging Caused by Progerin. Cell Rep. 2018;22:2006–2015. doi: 10.1016/j.celrep.2018.01.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Liu S, et al. Whole-grain consumption and risk of coronary heart disease: results from the Nurses’ Health Study. Am J Clin Nutr. 1999;70:412–419. doi: 10.1093/ajcn/70.3.412. [DOI] [PubMed] [Google Scholar]
- 22.Karmally W, et al. Cholesterol-lowering benefits of oat-containing cereal in Hispanic americans. J Am Diet Assoc. 2005;105:967–970. doi: 10.1016/j.jada.2005.03.006. [DOI] [PubMed] [Google Scholar]
- 23.Liu B, Hannun YA. Inhibition of the neutral magnesium-dependent sphingomyelinase by glutathione. J Biol Chem. 1997;272:16281–16287. doi: 10.1074/jbc.272.26.16281. [DOI] [PubMed] [Google Scholar]
- 24.Cai X, Chiu YH, Chen ZJ. The cGAS-cGAMP-STING pathway of cytosolic DNA sensing and signaling. Molecular cell. 2014;54:289–296. doi: 10.1016/j.molcel.2014.03.040. [DOI] [PubMed] [Google Scholar]
- 25.Yang H, Wang H, Ren J, Chen Q, Chen ZJ. cGAS is essential for cellular senescence. Proc Natl Acad Sci USA. 2017;114:E4612–e4620. doi: 10.1073/pnas.1705499114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Torralba D, et al. Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat Commun. 2018;9:2658. doi: 10.1038/s41467-018-05077-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Melsom T, et al. Mild Albuminuria Is a Risk Factor for Faster GFR Decline in the Nondiabetic Population. Kidney Int Rep. 2018;3:817–824. doi: 10.1016/j.ekir.2018.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bautista-Perez R, et al. Involvement of neutral sphingomyelinase in the angiotensin II signaling pathway. Am J Physiol Renal Physiol. 2015;308:F1178–1187. doi: 10.1152/ajprenal.00079.2014. [DOI] [PubMed] [Google Scholar]
- 29.De S, et al. Exocytosis-Mediated Urinary Full-Length Megalin Excretion Is Linked With the Pathogenesis of Diabetic Nephropathy. Diabetes. 2017;66:1391–1404. doi: 10.2337/db16-1031. [DOI] [PubMed] [Google Scholar]
- 30.Hu MC, et al. Renal Production, Uptake, and Handling of Circulating alphaKlotho. J Am Soc Nephrol. 2016;27:79–90. doi: 10.1681/ASN.2014101030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kapustin AN, et al. Calcium regulates key components of vascular smooth muscle cell-derived matrix vesicles to enhance mineralization. Circ Res. 2011;109:e1–12. doi: 10.1161/CIRCRESAHA.110.238808. [DOI] [PubMed] [Google Scholar]
- 32.Shroff RC, et al. Dialysis accelerates medial vascular calcification in part by triggering smooth muscle cell apoptosis. Circulation. 2008;118:1748–1757. doi: 10.1161/CIRCULATIONAHA.108.783738. [DOI] [PubMed] [Google Scholar]
- 33.Khavandgar Z, et al. A cell-autonomous requirement for neutral sphingomyelinase 2 in bone mineralization. J Cell Biol. 2011;194:277–289. doi: 10.1083/jcb.201102051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Aubin I, et al. A deletion in the gene encoding sphingomyelin phosphodiesterase 3 (Smpd3) results in osteogenesis and dentinogenesis imperfecta in the mouse. Nat Genet. 2005;37:803–805. doi: 10.1038/ng1603. [DOI] [PubMed] [Google Scholar]
- 35.Cosola C, et al. Beta-Glucans Supplementation Associates with Reduction in P-Cresyl Sulfate Levels and Improved Endothelial Vascular Reactivity in Healthy Individuals. PLoS One. 2017;12:e0169635. doi: 10.1371/journal.pone.0169635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ito S, Yoshida M. Protein-bound uremic toxins: new culprits of cardiovascular events in chronic kidney disease patients. Toxins (Basel) 2014;6:665–678. doi: 10.3390/toxins6020665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lin CJ, Wu V, Wu PC, Wu CJ. Meta-Analysis of the Associations of p-Cresyl Sulfate (PCS) and Indoxyl Sulfate (IS) with Cardiovascular Events and All-Cause Mortality in Patients with Chronic Renal Failure. PLoS One. 2015;10:e0132589. doi: 10.1371/journal.pone.0132589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Iraz M, Bilgic S, Samdanci E, Ozerol E, Tanbek K. Preventive and early therapeutic effects of beta-glucan on the bleomycin-induced lung fibrosis in rats. Eur Rev Med Pharmacol Sci. 2015;19:1505–1516. [PubMed] [Google Scholar]
- 39.Hong F, et al. Mechanism by which orally administered beta-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models. J Immunol. 2004;173:797–806. doi: 10.4049/jimmunol.173.2.797. [DOI] [PubMed] [Google Scholar]
- 40.Rice PJ, et al. Oral delivery and gastrointestinal absorption of soluble glucans stimulate increased resistance to infectious challenge. J Pharmacol Exp Ther. 2005;314:1079–1086. doi: 10.1124/jpet.105.085415. [DOI] [PubMed] [Google Scholar]
- 41.Chan GC, Chan WK, Sze DM. The effects of beta-glucan on human immune and cancer cells. J Hematol Oncol. 2009;2:25. doi: 10.1186/1756-8722-2-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Han Y, et al. Role of complement 3a in the synthetic phenotype and angiotensin II-production in vascular smooth muscle cells from spontaneously hypertensive rats. American journal of hypertension. 2012;25:284–289. doi: 10.1038/ajh.2011.214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tamura K, et al. Molecular Mechanism by which Prominent Human Gut Bacteroidetes Utilize Mixed-Linkage Beta-Glucans, Major Health-Promoting Cereal Polysaccharides. Cell Rep. 2017;21:417–430. doi: 10.1016/j.celrep.2017.09.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Novak M, Vetvicka V. Beta-glucans, history, and the present: immunomodulatory aspects and mechanisms of action. J Immunotoxicol. 2008;5:47–57. doi: 10.1080/15476910802019045. [DOI] [PubMed] [Google Scholar]
- 45.Motilva MJ, et al. Adaptation of the standard enzymatic protocol (Megazyme method) to microplaque format for β-(1,3)(1,4)-d-glucan determination in cereal based samples with a wide range of β-glucan content. Journal of Cereal Science. 2014;59:224–227. doi: 10.1016/j.jcs.2014.01.007. [DOI] [Google Scholar]
- 46.Arcidiacono MV, Yang J, Fernandez E, Dusso A. The induction of C/EBPbeta contributes to vitamin D inhibition of ADAM17 expression and parathyroid hyperplasia in kidney disease. Nephrol Dial Transplant. 2015;30:423–433. doi: 10.1093/ndt/gfu311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Arcidiacono MV, Yang J, Fernandez E, Dusso A. Parathyroid-specific epidermal growth factor-receptor inactivation prevents uremia-induced parathyroid hyperplasia in mice. Nephrol Dial Transplant. 2015;30:434–440. doi: 10.1093/ndt/gfu318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
No datasets were generated or analyzed during the current study.