

Abstract
Disturbed metabolism of copper ions can cause diseases such as Wilson’s disease (WD). In this work, we investigated the inhibitory effect of Cu(II) ion in vitro on the AlkB family DNA repair enzymes, which are members of the Fe(II)/alpha-ketoglutarate-dependent dioxygenase and include human ALKBH2, ALKBH3, and E. coli AlkB proteins. None of the three proteins was significantly inhibited under normal cellular copper concentrations. However, under WD related condition, we observed that the activities of all three enzymes were strongly suppressed (from 95.2 to 100.0%). We also noted the repair efficiency under ds-DNA condition was less susceptible than ss-DNA to the inhibition.
Graphical Abstract
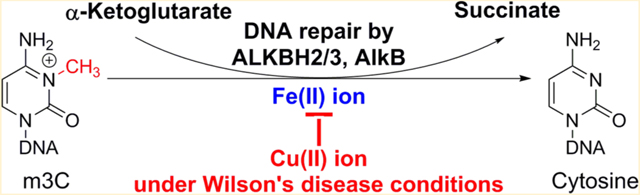
Copper is an essential yet toxic metal for human physiological functions, and the cellular concentrations of its ions, such as Cu(II) and Cu(I) ions, are tightly regulated. If the concentrations of these ions are dysregulated, certain diseases, such as Wilson’s disease (WD), may occur.1,2 For patients under WD condition, the level of copper in human liver samples raises as much as 249.9 ng/mg (dry weight), which is significantly higher (more than 10-fold) than the level in normal cells (21.5 ng/mg).1 These elevated copper levels create an inflammatory cellular environment in liver tissue cells and damage their lipid bilayers, thus forming a variety of DNA adducts. Nair et. al have reported that ethenoadenine (εA) and ethenocytosine (εC) DNA lesions are significantly increased under WD condition.1,3 These etheno DNA lesions are substrates of the AlkB family DNA repair enzymes, such as the human homologues ALKBH2 and 3, which use an Fe(II)/alpha-ketoglutarate (αKG)-dependent mechanism to oxidize the abnormal alkyl groups, ultimately restoring the undamaged DNA bases.4 Previously, the copper(II) ion has been reported to replace the Fe(II) ion in AlkB in a spectroscopic study and to inhibit several Fe(II)/αKG family enzymes such as JMJD2A, JMJD2E, and PHD2.5–7 Several divalent ions, such as Ni2+, Co2+, and Mn2+, have been demonstrated to inhibit other enzymes in this family such as C–P4H, PLOD1, FIH, TauD, JMJD1A, and ALKBH2.5,8 A recent study by Yu et al. demonstrated that Cu(II) ion inhibits the oxidation of 5-methylcytosine in DNA by the Fe(II)/αKG-dependent Tet family enzymes in vitro, and this inhibition was also reflected by decreased levels of Tet oxidation products in genomic DNA isolated from tissues of Long-Evans Agouti rat, a model of human Wilson’s disease.9 It is reasonable to predict, then, that the high level of copper ions accumulated under WD condition inhibits the DNA repair capacity of both ALKBH2 and 3. Therefore, the elevated level of copper ions found in WD condition induces damages and increases the number of etheno adducts within the cell in a “two-fold” manner; it not only induces alkyl DNA damage through lipid peroxidation, but also suppresses the repair of those adducts by the AlkB family DNA repair enzymes.
The AlkB protein in E. coli was discovered to be an Fe(II)/αKG- dependent dioxygenase that repairs various alkyl DNA lesions (Figure 1).10,11 Nine human homologues of AlkB have been identified as ALKBH1–8 and FTO. Among these homologues, ALKBH2 and ALKBH3 have been characterized as DNA repair proteins.12,13 A variety of alkyl DNA lesions have been proven as substrates for AlkB, ALKBH2, and ALKBH3, both in vitro and in vivo. The lesions include methyl adducts, like 3-methylcytosine (m3C) and 1-methyladenine (m1A), etheno adducts, such as εA and εC, and other lesions with complex alkyl modifications to the DNA bases.13 Among those adducts, m3C and m1A have been reported as the best substrates for the AlkB family enzymes.12,13 Previous in vitro studies on the inhibition of other Fe(II)/αKG-dependent enzymes by Cu(II) ion have only provided the IC50 values, and the results have not been extrapolated to physiologically and pathologically relevant concentrations.6 In this work, for the first time we not only measured the IC50 values of Cu(II) ion on the ABH2, ABH3, and AlkB proteins, but also tested the inhibitory effects under normal cellular and Wilson’s disease conditions.
Figure 1.

Repair mechanism of the AlkB family enzymes on alkyl DNA lesions and inhibition of repair by Cu(II) ion. Adduct m3C is used here as an example to show the enzymatic catalysis.
To test the inhibitory effect of Cu(II) ion on the AlkB family enzymes, we first site-specifically synthesized a 16mer oligonucleotide containing m3C, the major substrate of the AlkB family enzymes.13,14 We expressed and purified recombinant human ALKBH2 and ALKBH3 proteins and the E. coli AlkB protein.15 Then we evaluated the repair efficiency of the enzymes (Figure S1) and the inhibitory effect of Cu(II) ion under different conditions. It has been reported that the cellular concentrations of “free” copper and iron ions are very limited. Under normal cellular conditions, the concentration of free copper ions is 1.5 μM, which is 1.5-fold greater than that of free iron ions (1.0 μM).16,17 For the WD patients, the level of free copper ion is increased to 24.4 μM.18 In this work, we tested different Fe(II) ion concentrations and selected 5.0 μM to ensure the maximal efficiency of repair by the AlkB family enzymes (Figure S2); this concentration was similar to the Fe(II) ion concentrations used in other inhibition studies.6 The inhibitory effects of Cu(II) ion were tested from 0.0 to 100.0 μM, among which 7.5 μM of Cu(II) was used to mimic normal cellular condition. Though the cellular levels of copper ion under WD condition vary considerably from different reports, all of the levels are more than 10-fold higher than the normal cellular concentration.1,2,18 To make this work rigorous and relevant to pathological condition, we used 75.0 μM (10-fold) of copper ion to represent the WD condition.
For each enzymatic reaction, the m3C-containing oligonucleotides were incubated with the necessary cofactors for the AlkB reaction: Fe(II) ion, αKG, and ascorbic acid (see Experimental Section in Supporting Information) in either ss-or ds-DNA In a typical inhibition test (e.g., ALKBH2 repairing m3C), 5.0 μM oligonucleotide substrate and 2.0 μM ALKBH2 enzyme were mixed with a fixed concentration of Fe(II) ion (5.0 μM) plus different concentrations of Cu(II) ion, and the extent of the repair reaction was quantified (see Experimental Section for details). Because the AlkB family enzymes are able to repair DNA lesions in both ss- and ds-DNA, the repair reactions of m3C were carried out under both conditions.
For the Cu(II) ion inhibition of the ALKBH2 repair reaction on m3C in ss- and ds-DNA (Figure 2 and Table S1), the IC50 values were 14.8 μM and 54.0 μM, respectively (Table 1). The IC50 values for ALKBH3 and AlkB were also measured (Table 1). From the IC50 tests, we found the IC50 values are higher in ds-DNA condition than ss-DNA condition for all three enzymes, despite the preference of ALKBH3 and AlkB to repair m3C in ss-DNA and ALKBH2 preferring repair in ds-DNA.13 These results show the repair under ds condition is less susceptible than ss condition to the inhibition from Cu(II) ion. One possibility is that copper ion is harder to access to the active site of an enzyme and replace its iron ion due to the more crowded environment under ds-DNA binding condition.
Figure 2.

IC50 measurements of Cu(II) ion on the repair of m3C by ALKBH2, ALKBH3, and AlkB under ss- and ds-DNA conditions.
Table 1.
IC50 Values of Cu(II) Ion Inhibiting the AlkB Family Enzymesa
| protein | substrate | IC50 (μM) |
|---|---|---|
| ALKBH2 | ss-DNA | 14.8 |
| ds-DNA | 54.0 | |
| ALKBH3 | ss-DNA | 23.8 |
| ds-DNA | 28.4 | |
| AlkB | ss-DNA | 10.3 |
| ds-DNA | 29.3 |
Repair reactions of m3C were carried out under both ss- and ds-DNA conditions.
We then tested the inhibition of Cu(II) ion under normal physiological condition with the concentration at 7.5 μM. We found that none of the three proteins was significantly inhibited under this concentration, with the highest inhibition at 7.0% for ALKBH3 repairing m3C in ds-DNA (Table S2). These results indicated that the repair efficiency of the AlkB family enzymes was barely affected by Cu(II) ions under normal cellular conditions. Next, we tested the inhibition of Cu(II) ions under WD condition with the concentration of Cu(II) ion at 75.0 μM. We found that all three enzymes were strongly inhibited under WD condition (inhibition from 95.2 to 100.0%, Table S2).
In this work, we used m3C, the best substrate of AlkB, to demonstrate that the AlkB family enzymes are strongly inhibited by high concentration of Cu(II) ion under WD condition. Other substrates, such as the etheno DNA lesions, will be tested both in vitro and in vivo in the future, which may provide a new explanation to the previously observed elevation of etheno adducts in Wilson’s disease.1 Considering that there are about 80 Fe(II)/αKG-dependent enzymes in human body,12 it is highly possible that some of those enzymes could be inhibited in a similar mechanism by the high concentrations of the copper ion under Wilson’s disease condition.
Supplementary Material
ACKNOWLEDGMENTS
The authors want to thank the RI-INBRE program, its directors Prof. Zahir Shaikh and Prof. Bongsup Cho, and staff Dr. Al Bach, Kim Andrews, and Patricia Murray for their kind help. We also want to thank Michael Vittori and Kerri Bradshaw for their support and helpful discussions.
Funding
This work was supported by an Institutional Development Award from the National Institute of General Medical Sciences of the National Institutes of Health under Grant No. 2 P20 GM103430, 1R15CA213042-01 (to D.L.) and a Medical Research Funds grant from the Rhode Island foundation (to D.L.).
ABBREVIATIONS
- m3C
3-methylcytosine
- ss
single stranded
- ds
double stranded
- WD
Wilson’s disease
- αKG
alpha-ketoglutarate
- εA
ethenoadenine
- εC
ethenocytosine
Footnotes
ASSOCIATED CONTENT
Supporting Information
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acs.chemres-tox.7b00230.
Supplementary tables, figures, and experimental procedures (PDF)
The authors declare no competing financial interest.
REFERENCES
- (1).Nair J, Carmichael PL, Fernando RC, Phillips DH, Strain AJ, and Bartsch H (1998) Lipid peroxidation-induced etheno-DNA adducts in the liver of patients with the genetic metal storage disorders Wilson’s disease and primary hemochromatosis. Cancer Epidemiol. Biomark. Prev. 7, 435–440. [PubMed] [Google Scholar]
- (2).Wu F, Wang J, Pu C, Qiao L, and Jiang C (2015) Wilson’s disease: a comprehensive review of the molecular mechanisms. Int. J. Mol. Sci 16, 6419–6431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Nair J, Sone H, Nagao M, Barbin A, and Bartsch H (1996) Copper-dependent formation of miscoding etheno-DNA adducts in the liver of Long Evans cinnamon (LEC) rats developing hereditary hepatitis and hepatocellular carcinoma. Cancer Res. 56, 1267–1271. [PubMed] [Google Scholar]
- (4).Delaney JC, Smeester L, Wong C, Frick LE, Taghizadeh K, Wishnok JS, Drennan CL, Samson LD, and Essigmann JM (2005) AlkB reverses etheno DNA lesions caused by lipid oxidation in vitro and in vivo. Nat. Struct. Mol. Biol. 12, 855–860. [DOI] [PubMed] [Google Scholar]
- (5).Rose NR, McDonough MA, King ONF, Kawamura A, and Schofield CJ (2011) Inhibition of 2-oxoglutarate dependent oxygenases. Chem. Soc. Rev. 40, 4364–4397. [DOI] [PubMed] [Google Scholar]
- (6).Sekirnik R, Rose NR, Mecinović J, and Schofield CJ (2010) 2-Oxoglutarate oxygenases are inhibited by a range of transition metals. Met. Integr. Biometal Sci. 2, 397–399. [DOI] [PubMed] [Google Scholar]
- (7).Bleijlevens B, Shivarattan T, Sedgwick B, Rigby SEJ, and Matthews SJ (2007) Replacement of non-heme Fe(II) with Cu(II) in the alpha-ketoglutarate dependent DNA repair enzyme AlkB: spectroscopic characterization of the active site. J. Inorg. Biochem. 101, 1043–1048. [DOI] [PubMed] [Google Scholar]
- (8).Chen H, Giri NC, Zhang R, Yamane K, Zhang Y, Maroney M, and Costa M (2010) Nickel ions inhibit histone demethylase JMJD1A and DNA repair enzyme ABH2 by replacing the ferrous iron in the catalytic centers. J. Biol. Chem. 285, 7374–7383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Yu Y, Guerrero CR, Liu S, Amato NJ, Sharma Y, Gupta S, and Wang Y (2016) Comprehensive Assessment of Oxidatively Induced Modifications of DNA in a Rat Model of Human Wilson’s Disease. Mol. Cell. Proteomics 15, 810–817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Trewick SC, Henshaw TF, Hausinger RP, Lindahl T, and Sedgwick B (2002) Oxidative demethylation by Escherichia coli AlkB directly reverts DNA base damage. Nature 419, 174–178. [DOI] [PubMed] [Google Scholar]
- (11).Falnes PØ, Johansen RF, and Seeberg E (2002) AlkB-mediated oxidative demethylation reverses DNA damage in Escherichia coli. Nature 419, 178–182. [DOI] [PubMed] [Google Scholar]
- (12).(2015) 2-Oxoglutarate-Dependent Oxygenases (Hausinger R, and Schofield CJ, Eds.) Royal Society of Chemistry, Cambridge, UK. [Google Scholar]
- (13).Fedeles BI, Singh V, Delaney JC, Li D, and Essigmann JM (2015) The AlkB family of Fe(II)/α-ketoglutarate-dependent dioxygenases: repairing nucleic acid alkylation damage and beyond. J. Biol. Chem. 290, 20734–20742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Delaney JC, and Essigmann JM (2004) Mutagenesis, genotoxicity, and repair of 1-methyladenine, 3-alkylcytosines, 1-methylguanine, and 3-methylthymine in alkB Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 101, 14051–14056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Chen F, Tang Q, Bian K, Humulock ZT, Yang X, Jost M, Drennan CL, Essigmann JM, and Li D (2016) Adaptive response enzyme AlkB preferentially repairs 1-methylguanine and 3-methyl-thymine adducts in double-stranded DNA. Chem. Res. Toxicol. 29, 687–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Epsztejn S, Kakhlon O, Glickstein H, Breuer W, and Cabantchik I (1997) Fluorescence analysis of the labile iron pool of mammalian cells. Anal. Biochem. 248, 31–40. [DOI] [PubMed] [Google Scholar]
- (17).Ma J, Haldar S, Khan MA, Sharma SD, Merrick WC, Theil EC, and Goss DJ (2012) Fe2+ binds iron responsive element-RNA, selectively changing protein-binding affinities and regulating mRNA repression and activation. Proc. Natl. Acad. Sci. U. S. A. 109, 8417–8422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (18).McMillin GA, Travis JJ, and Hunt JW (2009) Direct measurement of free copper in serum or plasma ultrafiltrate. Am. J. Clin. Pathol. 131, 160–165. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.