

ABSTRACT
Infectious bronchitis virus (IBV) represents a major threat to poultry production worldwide particularly when complicated with bacterial infection. In the present study samples were collected from forty broiler farms with respiratory manifestations to characterize IBV and E. coli. Bacteriophages were isolated and enriched from sampled farms to study its efficacy to control single and mixed infections with E. coli and IBV in vivo. Twelve out of forty farms were positive for IBV. Phylogenetic analysis of partial spike protein revealed that all positive cases clustered within the GI-23 genotype. Eight out of forty farms were positive for E. coli serogroups O26, O78, O86, O114, O119, with O125 found on three farms. Bacteriophage treatment delayed the onset and reduced the severity of clinical signs, and prevented the mortality associated with single and mixed infection with IBV and E. coli. Furthermore, in mixed infections, bacteriophage treatment significantly reduced E. coli as well as IBV shedding. Groups treated with bacteriophages showed a significant reduction of E. coli shedding that gradually decreased over time, in contrast to higher and gradually increasing shedding without bacteriophage treatment. In conclusion, bacteriophage treatment significantly reduced the pathogenicity and shedding of IBVand E. coli in mixed infections.
KEYWORDS: Bacteriophage, E. coli, IBV, mixed infection
Introduction
Avian respiratory tract infections are associated with massive economic losses particularly under poor intensive rearing conditions in winter seasons. Several pathogens, adverse environmental conditions and poor managemental factors are involved [1–3]. Pathogens include avian influenza virus (AIV), Newcastle disease virus (NDV), infectious bronchitis virus (IBV), avian pneumovirus (APV), Mycoplasma gallisepticum (MG), and avian pathogenic E. coli (APEC). Avian respiratory tract infections can be caused by single pathogens or in combination with each other in form of preceding, concurrent or secondary infections [3–5]. Change in the ecosystem of the respiratory microbiome [6], mechanical damage of the epithelium, loss of ciliated cells and the impairment of innate immune responses are a consequence of primary respiratory infections and associated with enhanced disease and economical losses during superinfections [7].
In Egypt, respiratory diseases outbreaks caused significant economic losses in commercial poultry production during recent years [8]. Laboratory investigations revealed that IBV represents the most common pathogen detected during these outbreaks [8–10]. IB is a highly contagious disease affecting the respiratory, renal and reproductive tract of chickens. The disease caused by single-stranded, enveloped RNA virus belongs to the genus of Gamma coronavirus [11]. The spike (S) glycoprotein represents the major structural and immunogenic domain, and plays a pivotal role in tissue tropism and carry the receptor-binding site [12]. The S glycoprotein is cleaved post transitionary into S1 (globular head) and S2 (stalk) domain [13]. The S1 domain possesses three hypervariable regions (HVR 1, 2 and 3) responsible for the high genetic and antigenic variability of IBV [14]. Despite massive vaccination strategies used in Egypt, IBV extensively circulates in all over Egypt in vaccinated and non-vaccinated chickens with continuous viral evolution [9,15,16]. Complications between APEC and IBV infections are widely observed under field conditions and are reproduced under experimental conditions resulting in enhanced severe colibacillosis [7] and enhanced pathogenicity post IBV infection.
Under field conditions, commercial broiler farms with inadequate biosecurity measures and with the presence of colibacillosis in early age bird are associated with more severe IBV infections in late age birds (>20 days old). Despite the wide spread use of antibiotics and vaccination against IBV, complications as a consequence of mixed infections of avian bacterial pathogens including APEC and IBV became a general phenomenon associated with significant economic losses.
In the present study samples were collected from broiler farms with birds with respiratory manifestations. All samples were tested for the presence of IBV and APEC. Recovered IBV isolates were genetically characterized by partial spike protein sequencing. Recovered APEC were characterized by serotyping and antibiotic profiling. Bacteriophages were isolated from water samples collected from the same farms and enriched against APEC O78. In vivo study was conducted to evaluate the efficacy of bacteriophages in reducing the pathogenicity of single and mixed infections with APEC and IBV.
Material and methods
Sampling and sample preparation
One hundred recently succumbed chickens suspected of being IBV infected were collected from forty broiler farms. The study was conducted between late 2017 and early 2018 and samples collected in northern Egypt (Dakahila and Damietta governorates). Lung, trachea, kidney, and liver samples were aseptically collected. On the same day of sampling, half of the organs were cultured for bacteriological investigation in buffered peptone water at 37ºC for 24 h. For IBV screening, lung and trachea samples were homogenized with an equal mass of phosphate buffer saline and then centrifuged at 7,000 × g for 5 min. Supernatant was collected and stored at −80ºC until further investigation.
IBV and E.coli detection and characterization
Viral RNA was extracted using the QIAamp viral RNA mini kit (QIAGEN) in accordance with the manufacturer instructions. One-step RT-PCR was performed for IBV screening as previously described [17]. Genotyping with forward and reverse primers for amplification of HVR1, amplicon purification and sequencing was performed as previously described [17]. Obtained sequences in this study aligned with sequences represent all infectious bronchitis virus genotypes proposed by [18]. All samples used in the phylogenetic analysis were downloaded from GenBank (http://www.ncbi.nlm.nih.gov). The sequences were aligned using multiple alignment MAFFT version 7 (https://mafft.cbrc.jp/alignment/server/). The tree was constructed with the MEGA 6 software [19] using the nucleotide substitution of the Hasegawa-Kishino-Yano model with the gamma-distributed rate (with four rate categories) with bootstrap value based on 1000 replicates [20]. Then, the tree was viewed and edited using the FigTree v1.4.2 software (http://tree.bio.ed.ac.uk/software/figtree/).
For the bacteriological investigation, E.coli isolation and confirmation was performed in accordance with [21]. Confirmed, positive E.coli strains were subjected to serotyping using all available O (O1 to O181) antisera in accordance with [22], cross-reacting antigens were used to ensure the removal of cross-reactivity. All confirmed strains were tested against antibiotics commonly used in Egyptian farms by disc diffusion, with testing procedures and interpretations of the results performed in accordance with reference laboratory protocols. One of the field isolates E.coli (serotype O78) was tested for purity using API 20E (bioMérieux, Inc.) and counted in colony-forming units (CFU).
Bacteriophage isolation, enrichment, and titration
Over the surveillance period, sewage water samples were collected from poultry farms and used for the isolation of bacteriophages specific to APEC O78. Samples were centrifuged at 10,000 rpm for 10 min and then filtered through a 0.22 μm filter (Millipore). Bacteriophage purification and enrichment was performed in accordance with [23]. Briefly, filtered samples were mixed with Luria Bertani (LB) broth (Sigma), early-log grown APEC O78 added and samples incubated overnight at 37ºC with shaking set to 120 rpm/min. Then, samples were centrifuged at 10,000 rpm for 10 min and filtered through a 0.22 μm filter. The presence of bacteriophages was initially tested by the spot test method based on the double layer plaque technique in accordance with [24]. For this, 100 µl of APEC O78 were cultured on LB agar for 8 h, then 10 µl of the prepared bacteriophage suspension spotted onto it, and plates incubated at 37ºC overnight. The appearance of a clear zone in the plate indicated the presence of the lytic phage. Plaque assay was used for the titration of bacteriophages as previously described [25].
In vivo evaluation of bacteriophage efficacy in the reduction of E.coli and/or IB pathogenicity
Ethics statement
Animal experimental designs and procedures were reviewed and approved by RLQP in accordance with guidelines of the ministry of agriculture and land reclamation and ministry of environment of Egypt.
Virus and bacteria
Pure E.coli O78 isolate recovered in the present study were titrated by colony-forming units (CFU). Birds were challenged with 108 CFU diluted in 100 µl sterile PBS. The IBV strain IBV-EG/1212B-2012 (accession number: JQ839287) belonging to Egy/Var II [15] was used for the intratracheal challenge with 106 EID50 in 100 µl per bird. Previously described primers and probes were used to create a standard curve for absolute quantification of IBV shedding [26].
Experimental design
A total of 70 healthy one-day old chicks were purchased from a local poultry hatchery and housed in isolators with ad libitum access to feed and water. One day later, chicks were monitored for the presence of IBV or APEC O78 as previously described and then randomly divided into seven groups. Groups 1, 2, and 3 were treated with bacteriophages by intratracheal inoculation with 108 PFU at 1, 5, 8 and 13 days; whilst groups 4, 5, 6 and 7 were not treated with bacteriophages. Groups 1 and 4 were challenged intratracheally at day 2 with 108 CFU APEC O78. Groups 2 and 5 were challenged intratracheally with 106 EID50 of IBV. Groups 3 and 6 were infected with both pathogens in a 3 days interval. Group 7 was kept as a negative control and received PBS. The experimental design is shown in Figure 1. Three birds per time point were humanely culled at 4, 9 and 15 days post-infection (dpi). The carcasses were weighed and dissected and trachea and lung samples collected aseptically for the detection and titration of APEC O78, bacteriophages and IBV as previously described.
Figure 1.
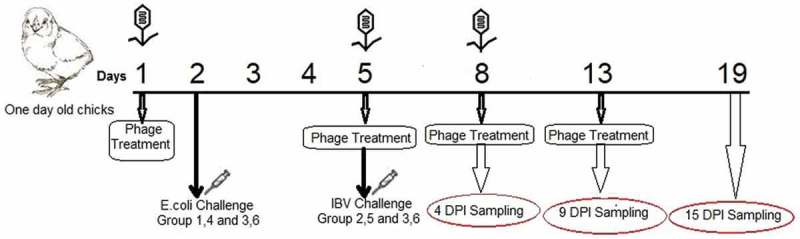
Schematic outline of the experimental design
Clinical signs scoring and sampling
All birds were observed twice daily and clinical signs recorded in accordance with [27]. Briefly, no signs were recorded as 0, mild signs included mild gasping, coughing or depression and were recorded as 1, and severe gasping, coughing or depression with ruffled feathers was considered as severe signs and recorded as 2. Birds with severe signs unable to move were recorded as 3 and humanely culled and samples collected. Birds which unexpectedly succumbed to disease were also recorded as 3 and samples collected.
Statistical analysis
Statistical significances of viral and bacterial shedding between different groups were evaluated using Student’s t-test in Microsoft Excel based on the mean of shedding quantities in three tested birds sampled from each group at each time point.
Results
History, clinical findings, and prevalence of APEC and IBV in the examined flocks
Forty broiler flocks, aged between 30 and 40 days and suffering from respiratory affections, from the Dakahlia and Damietta governorate were examined. Unwell birds showed respiratory manifestations visible as nasal discharge, sneezing, gasping and swollen head. In all farms examined unwell birds were isolated and subjected to intense antibiotic treatments. However, clear differences in activity and feed intake remained. Screening of collected samples focused on the detection of IBV and E.coli.
Eight farms were positive for E.coli (20% of all farms examined), five farms from the Dakahlia and three farms from the Damietta governate. Serotyping of the isolated strains revealed the presence of the O26, O78, O86, O114, O119 serogroups, with the O125 serogroup detected on three farms. Disc diffusion tests showed the following resistance rates: 100% to penicillin, 75% to gentamycin and streptomycin, 62.5% to ciprofloxacin, 50% to cefotaxime and 25% to doxycycline. 75% of all isolated strains were multidrug-resistant (MDR).
Twelve farms were positive for IBV (30% of all farms examined) determined by real-time PCR, with ten farms from the Dakahlia and two farms from the Damietta governate. Postmortem examination of the recently succumbed birds revealed the presence of caseous plug at the tracheal bifurcation, caseous trachitis, and fibrinous pericarditis, perihepatitis and air sacculitis. For virus isolation, specific pathogen free embryonated chicken eggs were inoculated. Total RNA was extracted from the allantoic fluid and the partial S1 gene of each isolate sequenced. Results obtained revealed that the tested samples clustered within the Egyptian variant group of GI-23 genotype subgroups (Figure 2). Nucleotide sequences tested in this study were designated as IBV_Egypt_Chicken_2017_partial, IBV_Egypt_Chicken _2018_ IBV_Egypt_Chicken_2017_partial_2, IBV_Egypt_Chicken_2017_partial_3, IBV_ Egypt_Chicken_2017_partial_4, IBV_Egypt_Chicken_2017_partial_5, IBV_ Egypt _Chicken_2018_partial_2, IBV_Egypt_Chicken_2018_partial_3, IBV_Egypt_ Chicken_2018_partial_4, IBV_Egypt_Chicken_2018_partial_5,IBV_Egypt_Chicken _2018_partial_6 and IBV_Egypt_Chicken_2018_partial_7, and were submitted to GenBank under accession number MK415385 and MK415386, MK882512, MK882513, MK882514, MK882515, MK882516, MK882517, MK882518, MK882519, MK882520 and MK882521 respectively.
Figure 2.
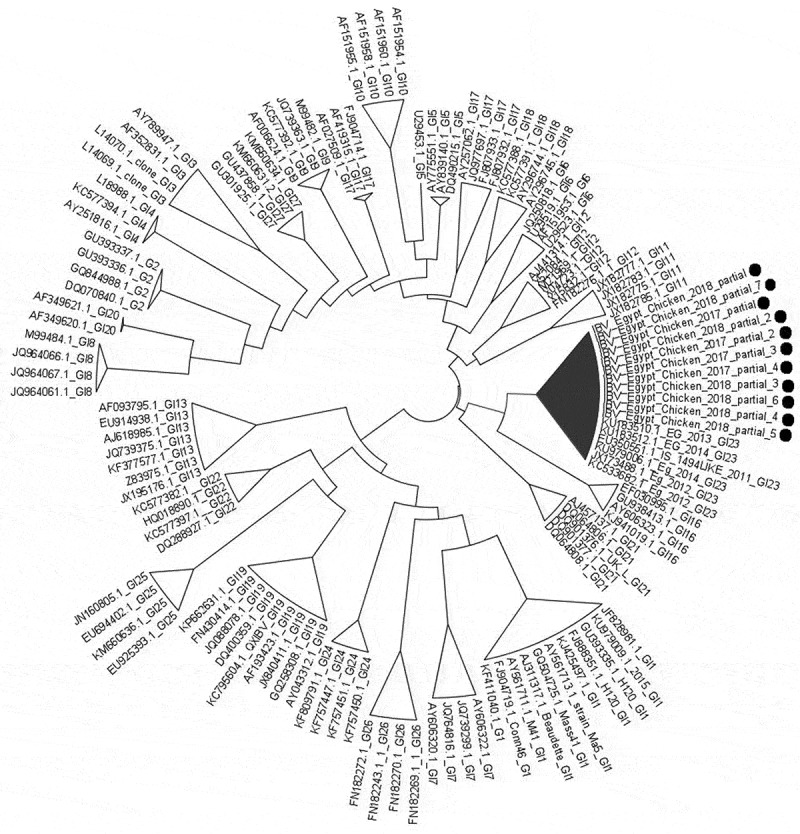
Phylogenetic analysis of IBV samples based on partial HVR1 sequences. Shown sequences represent all infectious bronchitis virus genotypes (GI) proposed by [18] with samples from the present study indicated by black circles. The tree was constructed using the nucleotide substitution of the Hasegawa-Kishino-Yano model with the gammadistributed rate (with four rate categories) with bootstrap value based on 1000 replicates
In vivo evaluation of bacteriophage treatment
Clinical signs observed including gasping, coughing or depression started to appear from three days post-challenge with APEC or a mixed APEC and IBV infection. Bacteriophage treatment delayed the onset of the clinical signs to 6 days post-challenge (dpc) and in addition markedly reduced their severity in both groups (Figure 3). Regarding IBV infection, clinical signs were observed from four-days post-challenge, with bacteriophage treatment leading to a reduction of their severity, but not delaying their onset (Figure 3).
Figure 3.
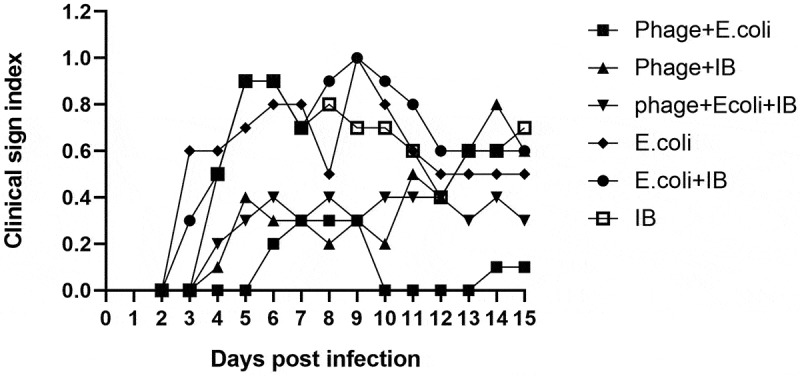
Clinical signs scoring in different groups. No signs were recorded as 0, mild signs included mild gasping, coughing or depression and were recorded as 1, and severe gasping, coughing or depression with ruffled feathers was considered as severe signs and recorded as 2. Birds with severe signs unable to move were recorded as 3 and humanely culled and samples collected. Birds which unexpectedly succumbed to disease were also recorded as 3 and samples collected. Each dot represents the mean of the clinical signs scoring of all birds in the group at each time point.
Bacteriophage treatment was not associated with mortality in single APEC or mixed APEC and IBV infected groups. In contrast, birds challenged with APEC alone and mixed APEC and IBV infection without bacteriophage treatment showed a 16% and 29% mortality rate at 8 and 7 days post-infection respectively (Figure 4). Bacteriophage treatment in combination with single IBV infection did not reduce the mortality of 26% (Figure 4).
Figure 4.
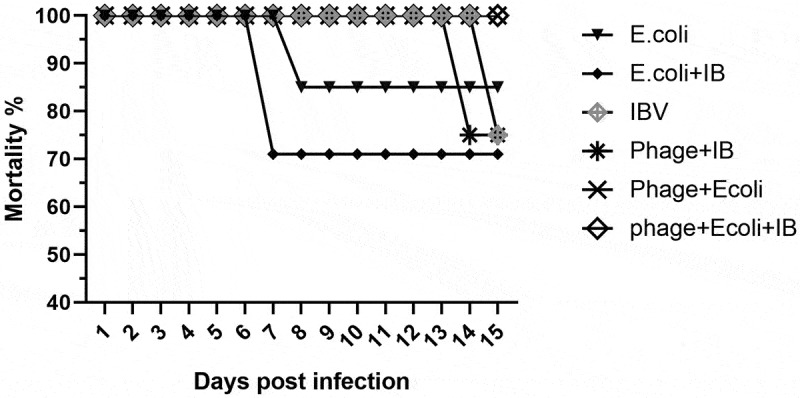
Average percentage of mortality in different groups (birds per group at each time point). The highest and earliest mortality was associated with mixed infections without bacteriophage treatment. In contrast, no deaths occurred in bacteriophage treated groups with mixed infection.
Bacteriophage treatment significantly reduced APEC shedding after single APEC or mixed APEC and IBV challenge, with a gradual decrease of bacterial loads in lung tissues over time. In contrast, a non-treated and challenged group showed a significantly higher APEC load with a gradual increase over time especially at 9 and 15 dpc (Figure 5). Interestingly, bacteriophage treatment significantly reduced IBV shedding in the mixed infected group but not in the IBV alone infected group comparing to the mixed infected group without bacteriophage treatment. The bacteriophage treated group infected with IBV showed relatively comparable results to the infected non-treated group. Groups with single IBV infection and mixed APEC and IBV infection with bacteriophage treatment showed a reduction, but not statistically significant, of IBV comparing to single IBV infection without bacteriophage treatment, with the reduction only becoming statistically significant at 15 dpc (Figure 6).
Figure 5.
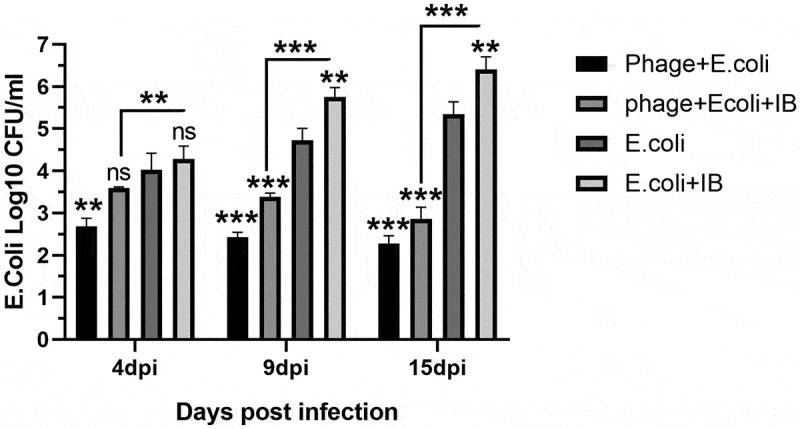
the E.coli bacterial counts. Samples were collected at 4, 9 and 15 dpc and are represented by the mean log10 CFU/ml of samples collected and tested from three birds. T-tests were performed. * p < 0.05, * * p < 0.01, ***p < 0.001 or ns for non-significant. The asterisks over columns represent a significant difference comparing to APEC the infected group. Asterisks over lines represent a significant difference between the mixed infection groups with and without bacteriophage treatments. Significantly higher counts with gradually increased bacterial shedding are observed in non-treated groups.
Figure 6.
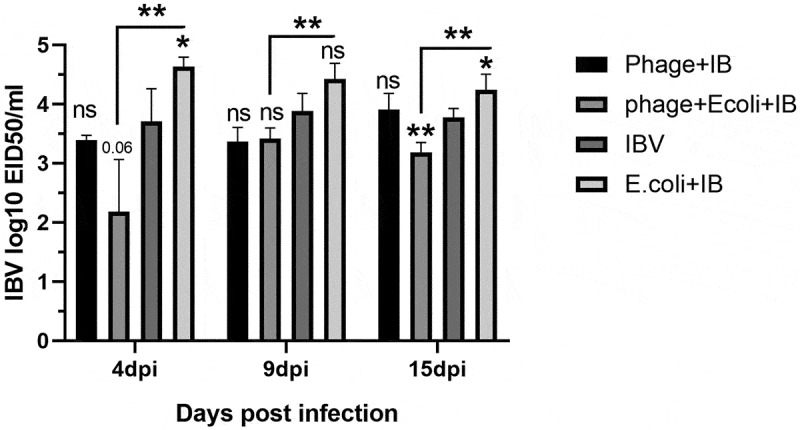
IBV titers. The IBV titers of samples collected at 4, 9 and 15 dpc are represented by the mean log10 EID50/ml of samples collected and tested from three birds. T-tests were performed. * p < 0.05, ** p < 0.01, and ns for non-significant. The asterisks over columns represent a significant difference compared to the IBV infected group. Asterisks over lines represent a significant difference between mixed infection groups with and without bacteriophage treatments. Significantly lower viral titers were observed in the mixed infection group treated with bacteriophages.
Discussion
Infectious bronchitis in particular complicated by mixed viral and/or bacterial infections is associated with catastrophic economic losses for the global poultry industry, and a significant threat for food security. Under field conditions complications caused by mixed IBV and APEC infections are linked with with accentuating respiratory signs in addition to airsacculitis, pericarditis and possible perihepatitis [28]. Under experimentally controlled conditions, mixed IBV and APEC infections can induce more pronounced clinical lesions in the air sacs that persist for a longer time [29]. In the present study samples were collected from birds with respiratory manifestations from a total of forty farms, and subjected to characterization based on currently circulating IBV and APEC strains. In addition, the efficacy of bacteriophage treatment in reducing APEC replication in the avian respiratory tract was studied in vivo, as well as their effect in reducing IBV pathogenicity after mixed APEC and IBV infections.
Out of forty farms, IBV was detected on 12 farms (30% of all farms examined). The high incidence of IBV in Egyptian poultry flocks suffering from respiratory manifestations (39–75.6%) has been recorded in several previous studies [8,10,30,31]. The range in the incidence rate is apparently due to the differences in the age of birds, with a higher incidence in 20–30 days old birds [8]. Based on partial spike protein gene sequencing, all positive IBV samples were clustered within the Egyptian variant IBV that belongs to the GI-23 genotype [18]. All recent molecular characterization of IBV circulating in Egypt revealed the circulation of two distinct genotypes, the GI-1 vicinal strains genotype and the GI-23 local variant genotype [32–34]. Despite wide spread vaccination with classical and/or variant strain based vaccines in Egypt, IBV outbreaks were frequently reported in vaccinated and non-vaccinated flocks of farms [16], indicating poor protection against the prevalent local variant strain [35].
In chickens, diseases associated with APEC cause significant economic losses due to mortality, decreased feed conversion rates, carcass contaminations and the high cost of its control and prevention. These losses in addition to the high prevalence of E.coli in poultry farms’ environment necessitate the use of antibiotics. However, the constant unwise use of antimicrobials in Egypt led to the high prevalence of multidrug-resistant strains that threaten human health and are associated with economic losses in the commercial poultry sector [36,37]. In the present study, 20% of the collected samples were positive for E.coli. 75% of them showed resistance to three or more antibiotics which were defined as multidrug resistant. This highlights the importance for the development of alternative therapeutic approaches preventing colibacillosis.
Bacteriophage therapy has been proven as an effective tool in the treatment of colibacillosis which initiated in the respiratory tract of chicken [38–40]. The ability to generate bacteriophages against selected bacterial species, serotypes or strains, that do not affect the commensal bacterial flora [41], make them safe in comparison to antibiotics and provide an alternative approach for eliminating pathogens.
In the present study, the efficacy of bacteriophage therapy in single and mixed infections with Egyptian IBV variant and/or APEC O78 has been evaluated. Results obtained revealed a significant higher persistence of APEC and IBV shedding together with higher morbidity and mortality in the mixed infection group comparing to single infected groups. This finding agrees with several previous studies [29,42,43]. Mechanisms behind the escalation of pathogenicity after mixed IBV and APEC infections are not yet fully understood. However, alteration of the immune response is most likely the cause not mechanical altering of the mucociliary barrier [29]. Bacteriophage treatment significantly reduced the severity of single APEC O78 challenge as well as a mixed infection with APEC and IBV. In brief, a high dose of bacteriophages administered intratracheally reduced the morbidity, prevented mortalities and significantly decreased APEC shedding compared to non-treated APEC challenge. Local application of bacteriophages at the site of APEC infection that is relatively inaccessible via the circulatory system, i.e. intra air sac administration, enables bacteriophages to eliminate the disease [44]. This explains the success of intrathoracic air sac injections in the reduction or prevention of mortality in several studies [38,44]. In the present study with a higher bacteriophage dose and intra-tracheal inoculation, APEC was significantly reduced in the bacteriophage treated lung tissue. In the case of mixed infections, bacteriophage treatment did not only reduce bacterial counts but also significantly reduced IBV detected in the lung, and associated with reduction in morbidity and mortality in the mixed infection group but not the single IBV infected group. These results show the efficient inhibition of APEC replication associated with a reduction of IBV severity during mixed infections.
In conclusion, the intra-tracheal administration of a high dose of bacteriophages limited APEC replication in the respiratory tract, and subsequently diminished the consequences of single and mixed infections with APEC and IBV. Bacteriophage treatment represents a promising tool for the prophylactic control of colibacillosis, and minimizes the consequences of mixed APEC and IBV infections.
Biographies
Maram Tawakol: Graduated 2004 from Mansoura university with PhD in bacteriology, immunology and mycology in parallel with work as microbiologist and head of biotechnology unit in the reference laboratory for veterinary quality control on poultry production animal health research institute (RLQP), Gamasa, Dakahlia, Egypt.
Nehal Nabil: Graduated 2004 from Mansoura university with PhD in bacteriology, immunology and mycology in parallel with work as microbiologist and quality manager in the RLQP, animal health research institute, Gamasa, Dakahlia, Egypt.
Ahmed Samy: Graduated from faculty of veterinary medicine zagazig university 2006. I got my PhD in veterinary microbiology and immunology 2013 from Cairo university. 2013 to 2015 work as a researcher in RLQP, Cairo, Egypt. currently I work as postdoctoral scientist, Pirbright Institute, Uk.
Acknowledgments
The authors would like to thank collogues in Reference Laboratory for Veterinary Quality Control on Poultry Production for their technical help. Also, we would like to thank the broilers farm’s owners for their cooperation and patience during samples collection.
Disclosure statement
All Authors declare that they have no conflict of interest.
References
- [1].Gross W. Factors affecting the development of respiratory disease complex in chickens. Avian Dis. 1990 Jul-Sep; 34(3):607–610.. [PubMed] [Google Scholar]
- [2].Ahmad SHK, Fawad N, Siddique B, et al. Current respiratory disease problem and the probes in chicken. Pak Vet J. 2002;22:17–8. [Google Scholar]
- [3].Roussan D, Haddad R, Khawaldeh G. Molecular survey of avian respiratory pathogens in commercial broiler chicken flocks with respiratory diseases in Jordan. Poult Sci. 2008;87(3):444–448. [DOI] [PubMed] [Google Scholar]
- [4].Malik YS, Patnayak DP, Goyal SM. Detection of three avian respiratory viruses by single-tube multiplex reverse transcription–polymerase chain reaction assay. J Vet Diagn Invest. 2004;16(3):244–248. [DOI] [PubMed] [Google Scholar]
- [5].Samy A, Naguib M. Avian respiratory coinfection and impact on avian influenza pathogenicity in domestic poultry: field and experimental findings. Vet Sci. 2018;5(1):23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Shabbir MZ, Malys T, Ivanov YV, et al. Microbial communities present in the lower respiratory tract of clinically healthy birds in Pakistan. Poult Sci. 2015;94(4):612–620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Ariaans MP, Matthijs MGR, van Haarlem D, et al. The role of phagocytic cells in enhanced susceptibility of broilers to colibacillosis after infectious bronchitis virus infection. Vet Immunol Immunopathol. 2008;123(3–4):240–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Hassan KE, Shany SAS, Ali A, et al. Prevalence of avian respiratory viruses in broiler flocks in Egypt. Poult Sci. 2016;95(6):1271–1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Magouz A, Abdo W, Abdelsabour A, et al. Molecular and histopathological investigation of avian infectious bronchitis virus in the Delta of Egypt between 2016 and 2017. Pak Vet J. 2018;38(3):243–248. [Google Scholar]
- [10].Setta A, S H, Elhady M, et al. Molecular and genetic characterization of infectious bronchitis viruses isolated from commercial chicken flocks in Egypt between 2014 and 2016. J World Poult Res. 2018;8(1):01–07. [Google Scholar]
- [11].Cavanagh D. A nomenclature for avian coronavirus isolates and the question of species status. Avian Pathol. 2001;30(2):109–115. [DOI] [PubMed] [Google Scholar]
- [12].Belouzard S, Millet JK, Licitra BN, et al. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses. 2012;4(6):1011–1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Cavanagh D. Coronavirus avian infectious bronchitis virus. Vet Res. 2007;38(2):281–297. [DOI] [PubMed] [Google Scholar]
- [14].Cavanagh D, Davis PJ, Mockett AA. Amino acids within hypervariable region 1 of avian coronavirus IBV (Massachusetts serotype) spike glycoprotein are associated with neutralization epitopes. Virus Res. 1988;11(2):141–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Zanaty A, Naguib MM, El-Husseiny MH, et al. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch Virol. 2016;161(12):3583–3587. [DOI] [PubMed] [Google Scholar]
- [16].Abd El Rahman S, Hoffmann M, Lueschow D, et al. Isolation and characterization of new variant strains of infectious bronchitis virus in Northern Egypt. Adv Anim Vet Sci. 2015;3:362–371. [Google Scholar]
- [17].Naguib MM, El-Kady MF, Lüschow D, et al. New real time and conventional RT-PCRs for updated molecular diagnosis of infectious bronchitis virus infection (IBV) in chickens in Egypt associated with frequent co-infections with avian influenza and Newcastle disease viruses. J Virol Methods. 2017;245:19–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Viviana Valastro ECH, Britton P, Fusaro A, et al. S1 gene-based phylogeny of infectious bronchitis virus: an attempt to harmonize virus classification. Infect Genet Evol. 2016;39(p):349–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Jackwood MW, Lee D-H, Cao Y. Different evolutionary trajectories of vaccine-controlled and non-controlled avian infectious bronchitis viruses in commercial poultry. PloS One. 2017;12(5):e0176709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Quinn PJ, Markey BK, Leonard FC, et al. Veterinary microbiology and microbial disease. UK: John Wiley & Sons; 2011. [Google Scholar]
- [22].Kok T, Worswich D, Gowans E. Some serological techniques for microbial and viral infections In: Collee J, Fraser A, Marmion B, et al, editors. Practical medical microbiology. 14th ed. Edinburgh, UK: Churchill Livingstone; 1996:179–204. [Google Scholar]
- [23].Oliveira A, Sillankorva S, Quinta R, et al. Isolation and characterization of bacteriophages for avian pathogenic E. coli strains. J Appl Microbiol. 2009;106(6):1919–1927. [DOI] [PubMed] [Google Scholar]
- [24].Carey-Smith GV, Billington C, Cornelius AJ, et al. Isolation and characterization of bacteriophages infecting Salmonella spp. FEMS Microbiol Lett. 2006;258(2):182–186. [DOI] [PubMed] [Google Scholar]
- [25].Shousha A, Awaiwanont N, Sofka D, et al. Bacteriophages isolated from chicken meat and the horizontal transfer of antimicrobial resistance genes. Appl Environ Microbiol. 2015;81(14):4600–4606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Callison SA, Hilt DA, Boynton TO, et al. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J Virol Methods. 2006;138(1–2):60–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Grgiæ H, Hunter DB, Hunton P, et al. Pathogenicity of infectious bronchitis virus isolates from Ontario chickens. Can J Vet Res. 2008;72(5):403. [PMC free article] [PubMed] [Google Scholar]
- [28].Ignjatovic J, Sapats S. Avian infectious bronchitis virus. Revue Scientifique Et Technique-Office International Des Epizooties. 2000;19(2):493–501. [DOI] [PubMed] [Google Scholar]
- [29].Matthijs MG, Ariaans MP, Dwars RM, et al. Course of infection and immune responses in the respiratory tract of IBV infected broilers after superinfection with E. coli. Vet Immunol Immunopathol. 2009;127(1–2):77–84. [DOI] [PubMed] [Google Scholar]
- [30].El-Hamid A, Hatem S, Ellakany HF, et al. Incidence of avian nephritis and infectious bronchitis viruses in broilers in Egypt. Alexandria J Vet Sci. 2016;51(2):33–45. [Google Scholar]
- [31].Selim K, Arafa AS, Hussein HA, et al. Molecular characterization of infectious bronchitis viruses isolated from broiler and layer chicken farms in Egypt during 2012. Int J Vet Sci Med. 2013;1(2):102–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Abdel-Moneim A, Madbouly HM, Gelb J, et al. Isolation and identification of Egypt/Beni-Seuf/01 a novel genotype of infectious bronchitis virus. Vet Med J-GIZA-. 2002;50(4):1065–1078. [Google Scholar]
- [33].Abdel-Moneim AS, Afifi MA, El-Kady MF. Emergence of a novel genotype of avian infectious bronchitis virus in Egypt. Arch Virol. 2012;157(12):2453–2457. [DOI] [PubMed] [Google Scholar]
- [34].Zanaty A, Arafa A-S, Hagag N, et al. Genotyping and pathotyping of diversified strains of infectious bronchitis viruses circulating in Egypt. World J Virol. 2016;5(3):125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Abozeid HH, Paldurai A, Varghese BP, et al. Development of a recombinant Newcastle disease virus-vectored vaccine for infectious bronchitis virus variant strains circulating in Egypt. Vet Res. 2019;50(1):12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Hussein AH, Ghanem IAI, Eid AAM, et al. Molecular and phenotypic characterization of Escherichia coli isolated from broiler chicken flocks in Egypt. Avian Dis. 2013;57(3):602–611. [DOI] [PubMed] [Google Scholar]
- [37].Ramadan H, Awad A, Ateya A. Detection of phenotypes, virulence genes and phylotypes of avian pathogenic and human diarrheagenic Escherichia coli in Egypt. J Infect Developing Countries. 2016;10(06):584–591. [DOI] [PubMed] [Google Scholar]
- [38].Oliveira A, Sereno R, Azeredo J. In vivo efficiency evaluation of a phage cocktail in controlling severe colibacillosis in confined conditions and experimental poultry houses. Vet Microbiol. 2010;146(3–4):303–308. [DOI] [PubMed] [Google Scholar]
- [39].Huff W, Huff GR, Rath NC, et al. Prevention of Escherichia coli respiratory infection in broiler chickens with bacteriophage (SPR02). Poult Sci. 2002;81(4):437–441. [DOI] [PubMed] [Google Scholar]
- [40].Huff W, Huff G, Rath N, et al. Evaluation of aerosol spray and intramuscular injection of bacteriophage to treat an Escherichia coli respiratory infection. Poult Sci. 2003;82(7):1108–1112. [DOI] [PubMed] [Google Scholar]
- [41].Wernicki A, Nowaczek A, Urban-Chmiel R. Bacteriophage therapy to combat bacterial infections in poultry. Virol J. 2017;14(1):179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Nakamura K, Cook JKA, Frazier JA, et al. Escherichia coli multiplication and lesions in the respiratory tract of chickens inoculated with infectious bronchitis virus and/or E. coli. Avian Dis. 1992;36(4):881–890. [PubMed] [Google Scholar]
- [43].Smith HW, Cook JK, Parsell ZE. The experimental infection of chickens with mixtures of infectious bronchitis virus and Escherichia coli. J Gen Virol. 1985;66(Pt 4):777–786. [DOI] [PubMed] [Google Scholar]
- [44].Huff WE, Huff GR, Rath NC, et al. Evaluation of the influence of bacteriophage titer on the treatment of colibacillosis in broiler chickens1. Poult Sci. 2006;85(8):1373–1377. [DOI] [PubMed] [Google Scholar]