

Abstract
Sex differences exist in the neurochemical mechanisms underlying tobacco smoking and smoking-related behaviors. Men tend to smoke for the reinforcing effects of nicotine, whereas women tend to smoke for stress and mood regulation, and have a harder time maintaining long-term abstinence. The mesolimbic dopamine (DA) system drives the reinforcing effects of tobacco smoking, whereas the mesocortical DA system—including the dorsolateral prefrontal cortex (dlPFC)—is critical for stress-related cognitive functioning and inhibitory control. This study is the first to investigate dlPFC D2/3-type receptor (D2R) availability and amphetamine-induced cortical DA release in smokers and nonsmokers. Forty-nine subjects (24 tobacco smokers (12 females) and 25 sex- and age-matched nonsmokers) participated in two same-day [11C]FLB457 positron emission tomography (PET) scans before and 3-hours after amphetamine administration (0.4–0.5 mg/kg, PO). D2R availability (non-displaceable binding potential; BPND) was measured pre- and post-amphetamine. The percent fractional change in BPND (%ΔBPND) between pre- and post-amphetamine, an index of DA release, was compared between male and female smokers and nonsmokers. Smokers showed significantly lower dlPFC D2R availability (BPND = 0.77 ± 0.05) than nonsmokers (BPND = 0.92 ± 0.04), p = 0.016, driven by males. Female smokers showed significantly less amphetamine-induced DA release in dlPFC (%ΔBPND = 1.9 ± 3.0%) than male smokers (%ΔBPND = 14.0 ± 4.3%), p < 0.005, and female nonsmokers (%ΔBPND = 9.3 ± 3.3%), p < 0.005. This study shows that in the prefrontal cortex, smokers have lower D2R availability than nonsmokers and that female vs. male smokers have a blunted amphetamine-induced DA release. These findings demonstrate that tobacco smoking differentially affects the mesocortical DA system in men vs. women, suggesting a potential target for gender-specific treatments.
Subject terms: Addiction, Addiction
Introduction
Tobacco smoking is the world’s leading cause of preventable death [1]. Deaths related to the use of combustible tobacco products greatly exceed those from alcohol, firearms, AIDS, and all other drugs including opioids, combined [2–6]. Tobacco smoking is largely driven by the reinforcing effects of nicotine—the primary addictive chemical in tobacco cigarettes. Sex differences have been documented both in the reinforcing effects of nicotine and in tobacco smoking treatment. Men experience greater nicotine-induced reinforcement than women [7, 8]. Men also respond better than women to nicotine-replacement therapies (NRTs) [9]—the first line of treatment for smoking cessation. Women are more reinforced by smoking cues [10], report greater psychological withdrawal [11], tend to relapse to smoking in response to stress [12, 13], and have a harder time maintaining long-term abstinence [14]. To treat female tobacco smokers more effectively, it is important to understand the molecular mechanisms underlying sex/gender-based behavioral differences.
Nicotine binds to and activates nicotinic acetylcholine receptors, which, in turn, facilitates DA release in striatal and cortical brain regions [15, 16] via the mesolimbic and mesocortical pathways, respectively. The mesolimbic (“reward”) DA pathway drives the reinforcing effects of tobacco smoking, while the mesocortical (“goal-directed”) DA pathway—including the dlPFC—is critical for inhibitory control [17], which is compromised by stress [18, 19]. In both non-human primates and humans, stress impairs prefrontal cortex (PFC) functioning [18, 20]. Systematic nicotine exposure alters medial PFC–ventral tegmental area (mPFC–VTA) coupling, particularly connections from the dlPFC [18]. Thus, stress-induced impairment of PFC function could contribute to sex differences in smoking-related behaviors.
It is well known from PET brain imaging studies that individuals with drug and alcohol use disorders, including tobacco smoking, have significantly lower D2R availability in various subregions of the striatum compared to healthy controls [21–27]. Importantly, lower striatal D2R availability has only been shown in male [21, 28], but not female tobacco smokers [28]. Sex differences in D2R availability have also been found in the midbrain [29], where DA neurons originate, and in the ventral striatum specifically, in response to smoking a cigarette [30].
Although previous PET studies have investigated the mesolimbic DA pathway, the mesocortical DA system remains largely unexplored in tobacco smokers. This is due, in part, to the limitations of widely available radiotracers, such as [11C]raclopride, which are best for measuring striatal D2R availability. The development of the radiotracer [11C]FLB457, a high-affinity D2R radiotracer [31, 32], provides a tool to measure extrastriatal (i.e. PFC) D2R availability. Due to its high-affinity and long equilibration time, [11C]FLB457 is not suitable for analysis of D2R availability in striatum. D2R radiotracers can also be used with drug challenges, i.e., amphetamine, to measure changes in synaptic DA [33] levels and infer DA system function.
To examine sex differences in D2R availability and DA function in the mesocortical pathway, we used [11C]FLB457 and PET imaging. The goals of this study were to measure cortical D2R availability and amphetamine-induced cortical DA release in male and female tobacco smokers and nonsmokers. The dlPFC was our a priori region-of-interest (ROI) because of its primary role in the mesocortical pathway, executive control, and stress-related cognitive function and impairment, in animal and human studies [18, 34, 35]. We hypothesized that smokers compared to nonsmokers would have lower dlPFC D2R availability, consistent with findings in the striatum [21, 28, 30]. Further, since stress compromises PFC functioning [20] and female smokers are more likely to relapse to smoking in response to stress [12, 36], we hypothesized that female smokers would show a smaller magnitude amphetamine-induced dlPFC DA response compared to male smokers and to female nonsmokers.
Materials and methods
Subjects
Twenty-five tobacco smokers (12 female) and twenty-five age- and sex-matched healthy controls/nonsmokers (12 female) participated in two same-day [11C]FLB457 PET scans, one scan before (‘baseline’), and the second scan 3-h after amphetamine administration (0.4–0.5 mg/kg, PO). One female smoker had her amphetamine administration and subsequent scan 5 days after the baseline scan. One male smoker was excluded due to a cerebellar abnormality. All subjects also underwent structural magnetic resonance imaging (MRI) required to delineate anatomical information [37]. A subset (n = 16, 10 males) of the smokers were included in a previous study testing different hypotheses [38]. Written informed consent for all study procedures, approved by the Yale Human Investigation Committee and the Yale-New Haven Hospital Radiation Safety Committee, was obtained from all subjects prior to participation. The study adhered to the Protection of Human Subjects of Research and Ethical Principles and Guidelines.
Subject screening procedures included a physical exam, electrocardiogram, blood tests, and urine toxicologies. Subjects had no history of significant major medical disorders and did not meet DSM-IV criteria for current or past psychiatric or substance abuse diagnosis (except nicotine dependence for smokers). Smokers were required to have been smoking cigarettes daily for at least one year (Table 1). Current tobacco smoking status was confirmed by spirometer carbon monoxide (CO) levels >10 parts per million (ppm) and urine cotinine—the primary metabolite of nicotine—levels >150 ng/ml on intake day. On scan day, smokers were required to be overnight abstinent, verified by CO levels <11 ppm or ≤50% of their intake level. All female subjects were required to have a negative pregnancy test on intake day and PET scan day prior to radiotracer administration. Menstrual cycle phase was not controlled and use of hormonal contraception was not exclusionary. Plasma samples were collected in all subjects on the day of the PET scan prior to the first scan for analysis of sex steroid hormone levels: follicle-stimulating hormone (FSH), estradiol, progesterone and testosterone.
Table 1.
Group demographics
| NS (N = 25) | S (N = 24) | NS v. S | FS (N = 12) | MS (N = 12) | FS v. MS | FNS (N = 12) | FNS v. FS | MNS (N = 13) | MNS v. MS | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | P-value | Mean ± SE | Mean ± SE | P-value | Mean ± SE | P-value | Mean ± SE | P-value | |
| Demographic | ||||||||||
| Age (years) | 29.3 ± 1.8 | 34.0 ± 1.9 | 0.08 | 32.8 ± 2.8 | 35.3 ± 2.8 | 0.52 | 30.3 ± 2.9 | 0.55 | 28.4 ± 2.4 | 0.07 |
| Weight (kg) | 74.4 ± 2.7 | 77.5 ± 2.8 | 0.44 | 70.3 ± 2.4 | 84.7 ± 4.1 | 0.01* | 66.6 ± 3.1 | 0.36 | 81.7 ± 3.2 | 0.57 |
| Smoking measures | ||||||||||
| Cigarettes/day | 12.8 ± 1.5 | 13.6 ± 1.5 | 0.70 | |||||||
| Years smoked | 13.5 ± 2.0 | 16.1 ± 1.9 | 0.36 | |||||||
| FTND | 4.6 ± 0.53 | 6.1 ± 0.9 | 0.15 | |||||||
| CO level | 10.0 ± 1.4 | 8.8 ± 1.3 | 0.51 | |||||||
| Cotinine level | 5.4 ± 0.5 | 6.0 ± 0.0 | 0.27 | |||||||
| MNWQ | 9.75 ± 1.90 | 5.58 ± 1.38 | 0.09 | |||||||
| QSU | 43.3 ± 3.37 | 36.8 ± 5.06 | 0.29 | |||||||
| Self-Report Mood Scales | ||||||||||
| STAI state | 26.2 ± 1.4 | 32.9 ± 2.2 | 0.01* | 35.8 ± 2.8 | 30.0 ± 3.3 | 0.19 | 24.6 ± 1.6 | 0.002* | 27.6 ± 2.3 | 0.55 |
| STAI trait | 27.7 ± 1.38 | 28.9 ± 2.09 | 0.64 | 31.9 ± 2.31 | 25.8 ± 3.37 | 0.15 | 28.1 ± 1.66 | 0.20 | 27.4 ± 2.19 | 0.70 |
| BDI | 0.9 ± 0.5 | 3.0 ± 0.8 | 0.03* | 3.1 ± 0.9 | 2.8 ± 1.3 | 0.88 | 0.8 ± 0.6 | 0.049* | 0.9 ± 0.8 | 0.22 |
| BIS-11 | 53.8 ± 1.96 | 57.1 ± 2.17 | 0.28 | 59.2 ± 2.98 | 54.9 ± 3.15 | 0.33 | 54.0 ± 2.47 | 0.19 | 53.7 ± 3.10 | 0.79 |
Mean ± SE shown
BDI Beck’s Depression Inventory, BIS-11 Barratt Impulsiveness Scale 11, CO carbon monoxide, FNS female nonsmokers, FS female smokers, FTND Fagerström’s Test for Nicotine Dependence, MNS male nonsmokers, MNWQ Minnesota Nicotine Withdrawal Scale, MS male smokers, NS nonsmokers, QSU Questionnaire of Smoking Urge, STAI State/Trait Anxiety Inventory, S smokers, SE standard error of the mean
*Significant at p < 0.05
On PET scan day, all subjects completed mood and other relevant questionnaires including the Beck’s Depression Inventory (BDI) [39], the State/Trait Anxiety Inventory (STAI) [40], and Barratt Impulsiveness Scale Version11 (BIS-11) [41]. Tobacco smokers completed measures of tobacco smoking behavior including the Fagerström Test for Nicotine Dependence (FTND) [42], Minnesota Nicotine Withdrawal Scale (MNWQ) [43], and Questionnaire of Smoking Urge (QSU) [44]. CO and urine cotinine were collected on PET scan day.
Amphetamine administration
Amphetamine (0.4–0.5 mg/kg PO) was administered 150–180-minutes prior to the second scan. Amphetamine levels in plasma have been shown to peak during this timeframe following oral amphetamine administration [32]. Peak amphetamine levels relate to peak extracellular DA [45] and provide maximum sensitivity for us to detect differences in BPND between pre- and post-amphetamine conditions. Blood samples to measure plasma amphetamine concentrations were collected at t = 0, 60, 120, 180, 225, and 270-minutes relative to time of amphetamine administration. Out of 49 subjects, two (one male smoker and one female smoker) received a dose of 0.5 mg/kg. All others received 0.4 mg/kg as previously discussed [38, 46]. Mean amphetamine dose did not differ between smokers and nonsmokers (Table 1). Male and female smokers did not differ in mean amphetamine dose per bodyweight.
Imaging data acquisition
The high-affinity D2/3 radioligand [11C]FLB457 was synthesized as previously described [37, 38]. [11C]FLB457 was injected intravenously as a bolus over 1-min by a computer-controlled pump (Harvard Apparatus, Holliston, MA, USA), and emission data were collected for 90 min. PET data were acquired using an ECAT EXACT HR + (Siemens/CTI, Knoxville, TN, USA). A 6-min transmission scan was acquired prior to the emission scan for attenuation correction. Subjects participated in an MRI on a 3T whole-body scanner (Trio, Siemens Medical Systems, Erlangen, Germany) on average ±13 days relative to the PET scan day. Structural T1 MRI was acquired for anatomical localization of the PET ROIs.
Imaging data processing and analysis
Sinograms were reconstructed with filter-back projection (FBP) with all corrections (attenuation, normalization, scatter, randoms, and deadtime) into a sequence of 27 frames: 6 × 30-s; 3 × 1-min; 2 × 2-min; 16 × 5-min. Motion correction was performed on dynamic image data by registering each frame to a summed early frame (e.g. the first 10-min of data) using a six-parameter mutual information algorithm [47] (FMRIB’s Linear Image Registration Tool, FMRIB Software library, Version3.2). PET summed images were smoothed at 3 × 3 × 3 voxel FWHM Gaussian filter. Image dimensions and voxel size were 128 × 128 × 63 and 2.06 × 2.06 × 2.43 mm3, respectively. The final reconstructed image resolution was ~6 mm full-width at half maximum.
Each MR image was normalized to Montreal Neurological Institute (MNI) space [48] using an affine linear plus nonlinear registration (Bioimage Suite 2.5, http://www.bioimagesuite.org/index.html), to extract ROIs from the automated anatomic labeling (AAL) template, as previously described [37]. The ROIs were then mapped from AAL space to PET space via the two transformations (e.g., PET-MR and MR-AAL template) to compute time activity curves in ROIs. All ROIs were bilateral summations and were defined using the AAL template including the dlPFC which was defined by combining the frontal superior, frontal mid, and frontal inferior triangularis corresponding to Brodmann’s areas 9 and 46 [49]. Primary analyses focused on the a priori dlPFC ROI (Fig. S1). Exploratory analyses were conducted on additional extrastriatal ROIs with a measurable [11C]FLB457 specific signal (defined as BPND > 0.5) [50], which included amygdala, anterior cingulate cortex, hippocampus, midbrain, occipital cortex, parietal cortex, temporal cortex, and thalamus.
PET data were fitted with the simplified reference tissue model (SRTM) [51, 52] using the cerebellum as a reference region to estimate BPND (an index of D2R availability that is proportional to the number of available binding sites) as previously validated and described [37, 38]. The cerebellum reference region was gray matter masked and excluded the cerebellar vermis because of its low levels of D2/3R expression. This reference region approach is sensitive to detecting amphetamine-induced DA release with [11C]FLB457 in extrastriatal regions, as validated in our prior study [37], which included 6 (4M, 2F) smokers from the current study. The %ΔBPND between pre- and post-amphetamine was calculated as:
Outcome measurements were estimated in defined ROIs with a measurable [11C]FLB457 specific signal (listed above) for primary and exploratory analyses.
Statistics
Student’s t-tests were used to evaluate basic demographics (i.e., age, weight), mood questionnaires, and smoking characteristics between groups. Linear regressions were performed to examine potential associations between [11C]FLB457 BPND and %ΔBPND values with clinical correlates of tobacco smoking including cigarettes smoked per day, years smoked and, FTND, MNWQ, QSU, and BIS-11 scores. These were corrected for multiple comparisons. Exploratory analyses (t-tests and linear regressions) were conducted on the hormone data to examine group differences and associations with BPND and %ΔBPND values (Table S1). Hormone analyses were not corrected for multiple comparisons because it was an exploratory aim.
To compare group differences in baseline dlPFC D2R availability, [11C]FLB457 BPND values were statistically analyzed using univariate analysis of variance (ANOVA) with sex and smoking status as between-subjects factors. The main effect of smoking status was examined to test the a priori hypothesis that smokers have lower dlPFC D2R availability compared to nonsmokers. To address our a priori hypothesis, we conducted pairwise F-tests within the ANOVA model to compare male smokers to male nonsmokers and female smokers to female nonsmokers. All main effects and interactions were tested with appropriate post-hoc contrasts within the model.
To compare differences in amphetamine-induced DA release, a repeated-measures ANOVA featuring time (pre- vs. post-amphetamine) as a within-subjects factor with sex and smoking status as between-subjects factors was performed on dlPFC BPND values. To address our a priori hypothesis, we conducted pairwise F-tests within the ANOVA model to compare pre- vs. post-amphetamine BPND measures between female smokers and male smokers, and between female smokers and female nonsmokers. All main effects and interactions were tested with appropriate post-hoc comparisons, within the model. Each exploratory ROI was examined with a separate but identical repeated-measures ANOVA statistical model post-hoc and were not corrected for multiple comparisons.
Results
Smoking and mood characteristics
Twenty-four smokers and 25 nonsmokers were included in the final analysis (Table 1). On average, smokers smoked 13.2 ± 1.0 cigarettes per day for 14.8 ± 1.4 years and had FTND scores of 5.3 ± 0.5, indicating moderate dependence levels. Male and female smokers were matched for age, cigarettes smoked per day, years of smoking, and FTND scores. Overnight abstinent smokers had significantly higher STAI state (p = 0.01) and BDI scores (p = 0.03) compared to nonsmokers. Female smokers had significantly higher STAI state (p = 0.002) and BDI scores (p = 0.049) compared to female nonsmokers, while male subgroups were comparable (p > 0.22). All subgroup comparisons in Table 1 showed no differences in STAI trait and BIS-11 scores. Male and female smoker subgroups did not differ on MNWQ and QSU smoking-related measures. There were no significant relationships between questionnaire scores and D2R availability, or amphetamine-induced DA release.
PET scan parameters
On average, pre- and post-amphetamine scans did not differ in injected activity, injected mass, or injected mass per bodyweight. Nonsmokers received lower injected activity, but higher injected mass, than smokers for baseline [11C]FLB457 scans (p ≤ 0.02). Male nonsmokers compared to male smokers had a lower injected activity and higher injected mass for baseline [11C]FLB457 scans (p ≤ 0.04). These mass differences might have mitigated as opposed to accentuated our group differences (see Supplemental Information). There were no significant differences in injected activity or injected mass between male and female smokers, for baseline or post-amphetamine [11C]FLB457 scans (Table S1).
Amphetamine administration and plasma levels
Amphetamine levels in plasma were not different at any time points between smokers and nonsmokers, or between male smokers and female smokers. Amphetamine levels increased following administration [38] and peaked at the start of the PET scan (150–225-min post-amphetamine; Figure S2).
Hormone levels
We found no significant differences in hormone levels (FSH, estradiol, progesterone, testosterone) between groups—females vs. males, smokers vs. nonsmokers or female vs. male smokers (Table S2), except, male smokers had higher testosterone levels than female smokers (17.1 ± 6.70 and 1.97 ± 0.30, respectively p = 0.047, uncorrected). There were no significant relationships between hormone levels and D2R availability or amphetamine-induced DA release in females, female smokers, or female nonsmokers.
D2R availability
D2R availability was measured in the baseline scan, prior to amphetamine administration. A priori planned contrast testing revealed a main effect of smoking status; dlPFC D2R availability was significantly lower in smokers (BPND = 0.77 ± 0.05) than nonsmokers (BPND = 0.92 ± 0.04), p = 0.016 (Fig. 1). A priori planned contrast testing also revealed that male smokers had a significantly lower dlPFC D2R availability (BPND = 0.69 ± 0.06) than male nonsmokers (BPND = 0.90 ± 0.05), p = 0.009, while female smokers had lower, but not significantly different dlPFC D2R availability (BPND = 0.85 ± 0.06) compared to female nonsmokers (BPND = 0.95 ± 0.08), p = 0.29. The main effect of sex (p = 0.10) and the interaction between sex and smoking status (p = 0.40) were not significant. There were no baseline differences in D2R availability between groups in exploratory ROIs (Table S3).
Fig. 1.
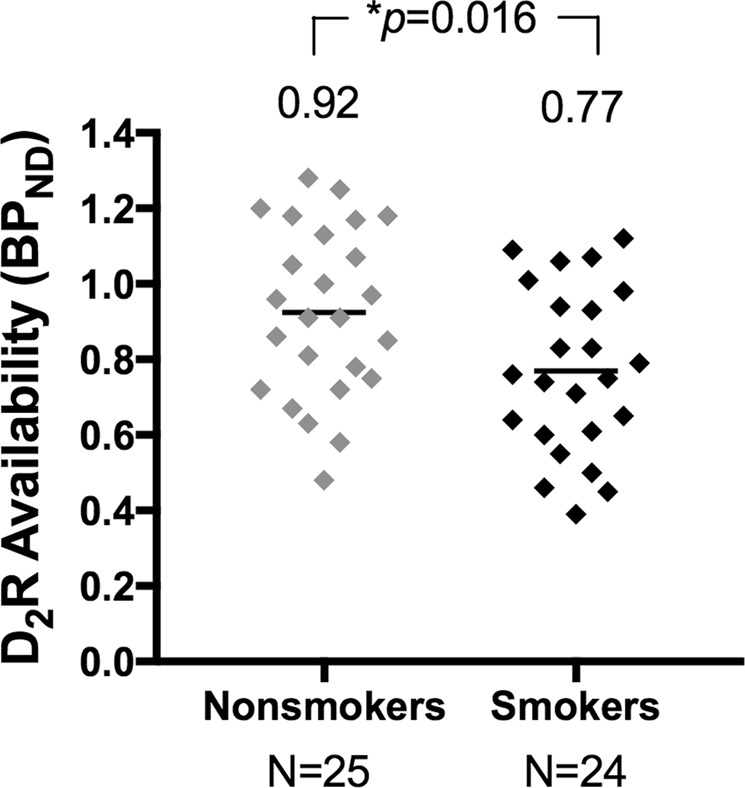
Baseline dlPFC D2R availability comparison of smokers and nonsmokers. Smokers (black) showed significantly lower dlPFC D2R availability than nonsmokers (gray), p = 0.016. Group means are shown above data points. BPND = binding potential
Amphetamine-induced DA release
Planned contrast testing of our a priori hypotheses demonstrated that female smokers had significantly less amphetamine-induced DA release in the dlPFC (%ΔBPND = 1.9 ± 3.0%) compared to male smokers (%ΔBPND = 14.0 ± 4.3%), p < 0.005, and compared to female nonsmokers (%ΔBPND = 9.3 ± 3.3%), p < 0.005; Bonferroni corrected (Fig. 2). Repeated-measures ANOVA also demonstrated a significant effect of time (amphetamine administration; p < 0.0005). Pre- vs. post-amphetamine dlPFC D2R availability was significantly different within the group of male smokers (p = 0.007), male nonsmokers (p = 0.0001), and female nonsmokers (p = 0.01), but not in the group of female smokers (p = 0.37), suggesting that there was no significant amphetamine-induced DA release in the female smokers. There were also significant main effects of sex (p = 0.04) and smoking status (p = 0.02). The interaction between time, sex, and smoking status was not significant (p = 0.10). In all exploratory ROIs, there was significant amphetamine-induced DA release (pre- vs. post-amphetamine; p < 0.02, uncorrected; except the midbrain) and no main effects of sex or smoking status (Table S3).
Fig. 2.
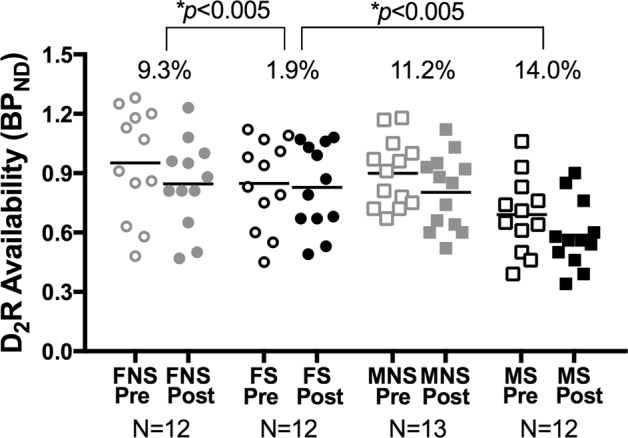
Baseline and post-amphetamine dlPFC D2R availability for all subgroups. Female smokers (black circles) showed significantly less amphetamine-induced DA release in the dlPFC than both male smokers (black squares) and female nonsmokers (gray circles). Male nonsmokers are shown in gray squares. Open shapes represent pre-amphetamine data and closed shapes represent post-amphetamine data. Group mean %ΔBPND is shown above data points. BPND = binding potential, FNS = female nonsmokers, FS = female smokers, MNS = male nonsmoker, MS = male smoker, Pre = pre-amphetamine, Post = post-amphetamine
Discussion
There are two primary findings from this neuroimaging study. First, tobacco smokers had significantly lower D2R availability in the dlPFC compared to nonsmokers. Although it is well known that tobacco smokers and individuals with other addictive disorders have lower D2R availability in the striatum [21–26] compared to matched control groups, this is the first study to extend these findings to the PFC. We also found differences by sex in smokers vs. nonsmokers; male smokers had lower dlPFC D2R availability (DA receptor levels) than male nonsmokers, but female smokers did not differ from female nonsmokers. Second, female smokers had significantly lower amphetamine-induced DA release (DA neurotransmission) in the dlPFC compared to male smokers, suggesting that tobacco smoking has no impact on stimulated DA release in males but blunts stimulated DA release in females. In addition, female smokers had significantly less amphetamine-induced DA release compared to female nonsmokers. Taken together, these findings suggest that tobacco smoking differentially affects the mesocortical DA system in males vs. females.
The main finding of lower D2R availability in tobacco smokers vs. controls extends our current understanding of the effect of tobacco smoking on the DA system. Numerous PET studies have demonstrated that individuals with addictive disorders (including tobacco smokers [21, 28, 53], and methamphetamine [24], alcohol [23], cocaine [22, 25], and heroin [26] use disorders) have significantly lower striatal D2R availability than comparison groups. Here, we show that dlPFC D2R availability is compromised, and potentially downregulated, by tobacco smoking, but only in males.
Lower dlPFC D2R availability in smokers vs. nonsmokers was driven by the difference between male smokers and male nonsmokers. We did not find a significant difference in dlPFC D2R availability between female smokers and female nonsmokers. Two studies previously reported lower striatal D2R availability in tobacco smokers vs. nonsmokers [21, 28]. Only one included females [28] and similarly reported differences in males (smokers < nonsmokers), but not females (smokers≈nonsmokers). Lower D2R availability in chronic smokers may be specific to males only, underscoring the importance of including females in these studies.
We found that tobacco smoking differentially alters DA function in males vs. females as indexed by amphetamine-induced DA release. Drug challenge paradigms has been used in many studies of addictive [23, 54] and other psychiatric [55–57] disorders to probe function of the DA system [58]. In this study, amphetamine-induced DA release in the dlPFC was negligible in female smokers compared to both male smokers and to female nonsmokers. Several studies have shown that amphetamine-induced DA release is blunted in individuals with addictive disorders compared to control groups [59], and in individuals with cocaine addiction, the more blunted the DA response, the worse the treatment outcome [22]. These data alone do not address whether this deficit is pre-existing or a consequence of drug use. However, preclinical longitudinal imaging studies, performed pre- and post- chronic cocaine administration, have shown that lower D2R availability in individuals with substance use disorders may be a consequence of use [60, 61]. While the neurobiological processes underlying a blunted DA response are unclear, these data suggest that blunted DA response is a biomarker of addiction development and treatment resistance.
Although our a priori ROI was the dlPFC because of its primary role in the mesocortical pathway, including executive control and stress-related cognitive function and impairment [18, 34, 35], we also examined other ROIs. For all exploratory ROIs, we observed a pattern in D2R availability between subgroups similar to the dlPFC; however, these differences did not reach statistical significance. These results suggest that the disruption in DA neurotransmission is specific to the dlPFC. For all ROIs (except the midbrain) we also observed a statistically significant amphetamine-induced DA response between pre- and post-amphetamine scans, suggesting that DA neurotransmission is intact in these brain regions in tobacco smokers. For the midbrain ROI, we did not observe any main effects of sex or smoking status at baseline, contrary to one prior study [29], nor did we observe an amphetamine-induced DA response. This study found a sex by smoking status interaction in baseline D2R availability using a different radiotracer and with a smaller sample size [29]. More research is required to disentangle these conflicting findings.
The mechanisms underlying the differential sex effects of tobacco smoking on the DA system are not established. Both the mesolimbic and mesocortical pathways originate in the VTA of the midbrain. In one prior study, female smokers have higher D2R availability in the midbrain than female nonsmokers, whereas male smokers and nonsmokers are not different [29]. Midbrain D2Rs are predominantly inhibitory, and it has been postulated that higher midbrain D2R availability in female smokers may lead to a suppression of ventral striatal smoking-induced DA release (compared to male smokers) [30]. In males, chronic tobacco smoking and the associated smoking-induced DA release in the ventral striatum likely downregulates D2R availability in that region. This is supported by the finding that male smokers have less D2R availability in striatum than male nonsmokers. Our sex-specific finding of lower D2R availability in the dlPFC of male smokers (but not female smokers) is consistent with striatal findings.
The neurochemical differences reported in the present study may underlie some of the behavioral sex differences reported in the literature [62–65]. For example, female smokers report smoking under stress or to reduce negative mood and tend to relapse in response to stress and negative mood [12, 13]. Under acute stress conditions, DA levels increase in the dlPFC and impair executive functions [20, 66], i.e., working memory and inhibitory control. This stress-induced disruption of the dlPFC may underlie the ability to resist smoking during stress and negative mood, which may lead to stress-induced smoking relapse [67] to manage these symptoms. Previous PET studies in humans have shown that dysregulated DA signaling in striatum is associated with maladaptive behavior such as drug seeking [54] and poorer treatment outcomes [22]. DA signaling in dlPFC is critical for stress management and behavioral disinhibition. We hypothesized that a dysfunctional mesocortical DA system in female smokers could facilitate stress-induced relapse and impede quit attempts. Thus, it is possible that a blunted amphetamine-induced DA response in the dlPFC may explain why women persistently smoke in response to stress. Although the neural mechanism of this blunted DA transmission in female smokers is unknown, it can be mechanistically explained by (1) decreased pre-synaptic DA release [54, 68], (2) reduction in pre-synaptic neuronal stores of dopamine [69], and/or (3) reduced baseline levels of endogenous dopamine [70] as observed in cocaine-dependent individuals. Another example of smoking-related sex difference is that male tobacco smokers are more sensitive to smoking-related rewards and more responsive to NRT than females [63, 71]. Our previous study showed that male smokers have a higher DA response in the striatum (the reinforcement center) in response to a cigarette [30] and the current study showed that female smokers have a blunted DA response to amphetamine (a robust DA probe). This heightened DA response to nicotine in male smokers and lack of a DA response in female smokers may explain why NRTs are more effective in men [22] since NRTs are thought to dampen the nicotine-induced dopaminergic response to smoking and extinguish drug-seeking behavior [72].
This study has several strengths including well-matched groups, a well-validated amphetamine challenge paradigm, and use of a novel high-affinity D2R radioligand that allowed for measurement of extrastriatal D2R availability and amphetamine-induced DA release. This study also has some limitations that can be addressed in future studies. Although the sample size allowed for systematic examination of sex differences, the size and makeup of the sample may have made it difficult to detect an influence of sex steroid hormones. The relationship between menstrual phase-related fluctuations in estrogen and/or progesterone on DA has produced mixed results [73–75]. Future studies should examine the relationship between sex steroid hormones and DA, controlling for menstrual cycle phase [76, 77]. It is important to note that the average amphetamine dose (0.38 mg/kg) in the current study is slightly lower than previous studies (0.4–0.5 mg/kg). Future studies should also include objective measurements of stress reactivity and nicotine-related behavior to determine if dlPFC D2R availability and amphetamine-induced DA release are differentially associated with stress-related cognitive impairment and behavioral disinhibition, respectively, in male and female smokers.
Much of our understanding of disease and treatment has come from clinical and preclinical studies in males [78]. Literature has shown that these findings do not necessarily translate in women [79]. Studying sex-related differences is essential for disentangling sex-related biological differences and promoting the health of both males and females. The current study identified a biological sexual dimorphism—tobacco smoking impairs mesocortical DA release in female smokers but not male smokers—thus highlighting the importance of including females in research studies. The dlPFC is a critical component of the stress pathway, therefore this sex difference may mediate the effects of stress impeding abstinence from tobacco smoking to a greater degree in women than men. In conjunction with the current findings, the limited efficacy of current dopaminergic and nicotine-replacement treatments—particularly in women—we propose that there is a critical need for the development of gender-specific treatment that target the stress pathway.
Funding and disclosure
The study was supported by the Office of Research on Women’s Health and the National Institutes of Health (P50DA033945, R01DA038832, R01DA14241, K02DA03175, and K01 AA024788). YZ was supported by the Interdepartmental Neuroscience Training Program at Yale University (T32NS4122813), Gruber Science Foundation Fellowship, the National Science Foundation Graduate Research Fellowship Program (NSF-GRFP), and the Neuroimaging Sciences Training Program at Yale University (T32DA022975). The authors declare no competing interests.
Supplementary information
Acknowledgements
We would like to thank Jon-Michael Anderson, Erin McGovern, Evgenia Perkins, Patrick Worhunsky, and the staff at the Yale PET Center.
Footnotes
Sixteen of the participants in this study took part in the following clinical trial: imaging extrastriatal dopamine release in tobacco smokers and nonsmokers; NCT02348385.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information accompanies this paper at (10.1038/s41386-019-0456-y).
References
- 1.Center for Disease Control and Prevention. Morbidity and Mortality Weekly Report (MMWR). US Department of Health & Human Services. 2015.
- 2.Center for Disease Control and Prevention. Drug Overdose Deaths. US Department of Health & Human Services. 2018.
- 3.Center for Disease Control and Prevention. Tobacco-Related Mortality. US Department of Health & Human Services. 2018.
- 4.Center for Disease Control and Prevention. All Injuries. US Department of Health & Human Services. 2017.
- 5.Center for Disease Control and Prevention. AIDS and HIV. US Department of Health & Human Services. 2018.
- 6.Center for Disease Control and Prevention. Alcohol and Public Health. US Department of Health & Human Services. 2018.
- 7.Perkins KA. Sex differences in nicotine versus nonnicotine reinforcement as determinants of tobacco smoking. Exp Clin Psychopharmacol. 1996;4:166–77. [Google Scholar]
- 8.Perkins KA, Donny E, Caggiula AR. Sex differences in nicotine effects and self-administration: review of human and animal evidence. Nicotine Tob Res. 1999;1:301–15. doi: 10.1080/14622299050011431. [DOI] [PubMed] [Google Scholar]
- 9.McKee SA, Smith PH, Kaufman M, Mazure CM, Weinberger AH. Sex differences in varenicline efficacy for smoking cessation: a meta-analysis. Nicotine Tob Res. 2016;18:1002–11. doi: 10.1093/ntr/ntv207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McBride D, Barrett SP, Kelly JT, Aw A, Dagher A. Effects of expectancy and abstinence on the neural response to smoking cues in cigarette smokers: an fMRI study. Neuropsychopharmacology. 2006;31:2728. doi: 10.1038/sj.npp.1301075. [DOI] [PubMed] [Google Scholar]
- 11.Faulkner P, Petersen N, Ghahremani DG, Cox CM, Tyndale RF, Hellemann GS, et al. Sex differences in tobacco withdrawal and responses to smoking reduced-nicotine cigarettes in young smokers. Psychopharmacology. 2018;235:193–202. doi: 10.1007/s00213-017-4755-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Xu J, Azizian A, Monterosso J, Domier CP, Brody AL, London ED, et al. Gender effects on mood and cigarette craving during early abstinence and resumption of smoking. Nicotine Tob Res. 2008;10:1653–61. doi: 10.1080/14622200802412929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cofta-Woerpel L, McClure JB, Li Y, Urbauer D, Cinciripini P, Wetter DW. Early cessation success or failure among women attempting to quit smoking: trajectories and volatility of urge and negative mood during the first post-cessation week. J Abnorm Psychol. 2011;120:596–606. doi: 10.1037/a0023755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Smith PH, Bessette AJ, Weinberger AH, Sheffer CE, McKee SA. Sex/gender differences in smoking cessation: a review. Prev Med. 2016;92:135–40. doi: 10.1016/j.ypmed.2016.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cosgrove Kelly P., Esterlis Irina, Sandiego Christine, Petrulli Ryan, Morris Evan D. The Neuropharmacology of Nicotine Dependence. Cham: Springer International Publishing; 2015. Imaging Tobacco Smoking with PET and SPECT; pp. 1–17. [DOI] [PubMed] [Google Scholar]
- 16.Benowitz NL. Nicotine addiction. New Engl J Med. 2010;362:2295–303. doi: 10.1056/NEJMra0809890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wu J, Gao M, Shen J-X, Shi W-X, Oster AM, Gutkin BS. Cortical control of VTA function and influence on nicotine reward. Biochem Pharmacol. 2013;86:1173–80. doi: 10.1016/j.bcp.2013.07.013. [DOI] [PubMed] [Google Scholar]
- 18.Arnsten AFT. Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci. 2009;10:410–22. doi: 10.1038/nrn2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Goldstein RZ, Volkow ND. Dysfunction of the prefrontal cortex in addiction: neuroimaging findings and clinical implications. Nat Rev Neurosci. 2011;12:652–69. doi: 10.1038/nrn3119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Arnsten AF. Stress weakens prefrontal networks: molecular insults to higher cognition. Nat Neurosci. 2015;18:1376–85. doi: 10.1038/nn.4087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Christoph Fehr MD, Igor Yakushev MD, Nina Hohmann DP, Hans-Georg Buchholz MS, Christian Landvogt MD, Hanna D, et al. Association of low striatal dopamine D 2 receptor availability with nicotine dependence similar to that seen with other drugs of abuse. Am J Psychiatry. 2008;165:507–14. doi: 10.1176/appi.ajp.2007.07020352. [DOI] [PubMed] [Google Scholar]
- 22.Martinez D, Carpenter KM, Liu F, Slifstein M, Broft A, Friedman AC, et al. Imaging dopamine transmission in cocaine dependence: link between neurochemistry and response to treatment. Am J Psychiatry. 2011;168:634–41. doi: 10.1176/appi.ajp.2010.10050748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Martinez D, Gil R, Slifstein M, Hwang D-R, Huang Y, Perez A, et al. Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biol Psychiatry. 2005;58:779–86. doi: 10.1016/j.biopsych.2005.04.044. [DOI] [PubMed] [Google Scholar]
- 24.Volkow ND, Chang L, Wang G-J, Fowler JS, Ding Y-S, Sedler M, et al. Low level of brain dopamine d2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am J Psychiatry. 2001;158:2015–21. doi: 10.1176/appi.ajp.158.12.2015. [DOI] [PubMed] [Google Scholar]
- 25.Martinez D, Broft A, Foltin RW, Slifstein M, Hwang D-R, Huang Y, et al. Cocaine dependence and D2 receptor availability in the functional subdivisions of the striatum: relationship with cocaine-seeking behavior. Neuropsychopharmacology. 2004;29:1190. doi: 10.1038/sj.npp.1300420. [DOI] [PubMed] [Google Scholar]
- 26.Martinez D, Saccone PA, Liu F, Slifstein M, Orlowska D, Grassetti A, et al. Deficits in dopamine D2 receptors and presynaptic dopamine in heroin dependence: commonalities and differences with other types of addiction. Biol Psychiatry. 2012;71:192–8. doi: 10.1016/j.biopsych.2011.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Worhunsky PD, Matuskey D, Gallezot J-D, Gaiser EC, Nabulsi N, Angarita GA, et al. Regional and source-based patterns of [(11)C]-(+)-PHNO binding potential reveal concurrent alterations in dopamine D(2) and D(3) receptor availability in cocaine-use disorder. Neuroimage. 2017;148:343–51. doi: 10.1016/j.neuroimage.2017.01.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brown AK, Mandelkern MA, Farahi J, Robertson C, Ghahremani DG, Sumerel B, et al. Sex differences in striatal dopamine D2/D3 receptor availability in smokers and nonsmokers. Int J Neuropsychopharmacol. 2012;15:989–94. doi: 10.1017/S1461145711001957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Okita K, Petersen N, Robertson CL, Dean AC, Mandelkern MA, London ED. Sex differences in midbrain dopamine d2-type receptor availability and association with nicotine dependence. Neuropsychopharmacology. 2016;41:2913–9. doi: 10.1038/npp.2016.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cosgrove KP, Wang S, Kim SJ, McGovern E, Nabulsi N, Gao H, et al. Sex differences in the brain’s dopamine signature of cigarette smoking. J Neurosci. 2014;34:16851–5. doi: 10.1523/JNEUROSCI.3661-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.SUDO Y., SUHARA T., INOUE M., ITO H., SUZUKI K., SAIJO T., HALLDIN C., FARDE L. Reproducibility of [11C]FLB 457 binding in extrastriatal regions. Nuclear Medicine Communications. 2001;22(11):1215–1221. doi: 10.1097/00006231-200111000-00008. [DOI] [PubMed] [Google Scholar]
- 32.Narendran R, Himes M, Mason NS. Reproducibility of Post-Amphetamine [11C]FLB 457 Binding to Cortical D2/3 Receptors. PLoS One. 2013;8:e76905. doi: 10.1371/journal.pone.0076905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Morris ED, Lucas MV, Cosgrove KP. How to study smoking and drinking with pet, positron emission tomography. In: Sandro Misciagna I, editor. London: IntechOpen Limited, 2013.
- 34.Arnsten AFT, Pliszka SR. Catecholamine influences on prefrontal cortical function: relevance to treatment of attention deficit/hyperactivity disorder and related disorders. Pharmacol Biochem Behav. 2011;99:211–6. doi: 10.1016/j.pbb.2011.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Arnsten AF, Wang M, Paspalas CD. Dopamine’s actions in primate prefrontal cortex: challenges for treating cognitive disorders. Pharm Rev. 2015;67:681–96. doi: 10.1124/pr.115.010512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Perkins KA, Karelitz JL, Giedgowd GE, Conklin CA. Negative mood effects on craving to smoke in women versus men. Addict Behav. 2013;38:1527–31. doi: 10.1016/j.addbeh.2012.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sandiego CM, Gallezot J-D, Lim K, Ropchan J, Lin S-f, Gao H, et al. Reference region modeling approaches for amphetamine challenge studies with [11C]FLB 457 and PET. J Cereb Blood Flow Metab. 2015;35:623–9. doi: 10.1038/jcbfm.2014.237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sandiego CM, Matuskey D, Lavery M, McGovern E, Huang Y, Nabulsi N, et al. The effect of treatment with guanfacine, an alpha2 adrenergic agonist, on dopaminergic tone in tobacco smokers: An [11C]FLB457 PET study. Neuropsychopharmacology. 2017;43:1–7. doi: 10.1038/npp.2017.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Beck AT. An inventory for measuring depression. Arch Gen Psychiatry. 1961;4:561. doi: 10.1001/archpsyc.1961.01710120031004. [DOI] [PubMed] [Google Scholar]
- 40.Spielberger CD, Gorsuch RL, Lushene RE. Manual for the State‐Trait Anxiety Inventory. Palo Alto: Consulting Psychologists Press; 1970. [Google Scholar]
- 41.Patton JH, Stanford MS, Barratt ES. Factor structure of the barratt impulsiveness scale. J Clin Psychol. 1995;51:768–74. doi: 10.1002/1097-4679(199511)51:6<768::aid-jclp2270510607>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- 42.Etter J-F, Duc TV, Perneger TV. Validity of the Fagerstrom test for nicotine dependence and of the Heaviness of Smoking Index among relatively light smokers. Addiction. 1999;94:269–81. doi: 10.1046/j.1360-0443.1999.94226910.x. [DOI] [PubMed] [Google Scholar]
- 43.Hughes JR, Hatsukami D. Signs and symptoms of tobacco withdrawal. Arch Gen Psychiatry. 1986;43:289–94. doi: 10.1001/archpsyc.1986.01800030107013. [DOI] [PubMed] [Google Scholar]
- 44.Tiffany ST, Drobes DJ. The development and initial validation of a questionnaire on smoking urges. Br J Addict. 1991;86:1467–76. doi: 10.1111/j.1360-0443.1991.tb01732.x. [DOI] [PubMed] [Google Scholar]
- 45.Narendran R, Jedema HP, Lopresti BJ, Mason NS, Gurnsey K, Ruszkiewicz J, et al. Imaging dopamine transmission in the frontal cortex: a simultaneous microdialysis and [11C]FLB 457 PET study. Mol Psychiatry. 2014;19:302–10. doi: 10.1038/mp.2013.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gaiser EC, Matuskey D, Perkins E, D’Amico C, Abdelghany O, McKee SA, et al. A case series on the heightened autonomic response due to guanfacine and amphetamine interaction. J Clin Psychopharmacol. 2015;35:197–9. doi: 10.1097/JCP.0000000000000273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Viola P, Wells Iii WM. Alignment by maximization of mutual information. Int J Comput Vis. 1997;24:137–54. [Google Scholar]
- 48.Holmes CJ, Hoge R, Collins L, Woods R, Toga AW, Evans AC. Enhancement of MR images using registration for signal averaging. J Comput Assist Tomogr. 1998;22:324–33. doi: 10.1097/00004728-199803000-00032. [DOI] [PubMed] [Google Scholar]
- 49.Petrides M, Pandya DN. Dorsolateral prefrontal cortex: comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. Eur J Neurosci. 1999;11:1011–36. doi: 10.1046/j.1460-9568.1999.00518.x. [DOI] [PubMed] [Google Scholar]
- 50.Narendran R, Frankle WG, Mason NS, Rabiner EA, Gunn RN, Searle GE, et al. Positron emission tomography imaging of amphetamine-induced dopamine release in the human cortex: a comparative evaluation of the high affinity dopamine D2/3 radiotracers [11C]FLB 457 and [11C]fallypride. Synapse. 2009;63:447–61. doi: 10.1002/syn.20628. [DOI] [PubMed] [Google Scholar]
- 51.Lammertsma AA, Hume SP. Simplified reference tissue model for PET receptor studies. Neuroimage. 1996;4:153–8. doi: 10.1006/nimg.1996.0066. [DOI] [PubMed] [Google Scholar]
- 52.Gunn RN, Lammertsma AA, Hume SP, Cunningham VJ. Parametric imaging of ligand-receptor binding in PET using a simplified reference region model. Neuroimage. 1997;6:279–87. doi: 10.1006/nimg.1997.0303. [DOI] [PubMed] [Google Scholar]
- 53.Wiers CE, Cabrera EA, Tomasi D, Wong CT, Demiral ŞB, Kim SW, et al. Striatal dopamine D2/D3 receptor availability varies across smoking status. Neuropsychopharmacology. 2017;42:2325. doi: 10.1038/npp.2017.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Martinez D, Narendran R, Foltin R, Slifstein M, Hwang D, Broft A, et al. Amphetamine-induced dopamine release: markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am J Psychiatry. 2007;164:622–9. doi: 10.1176/ajp.2007.164.4.622. [DOI] [PubMed] [Google Scholar]
- 55.Busto UE, Redden L, Mayberg H, Kapur S, Houle S, Zawertailo LA. Dopaminergic activity in depressed smokers: A positron emission tomography study. Synapse. 2009;63:681–9. doi: 10.1002/syn.20646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bertolino A, Breier A, Callicott JH, Adler C, Mattay VS, Shapiro M, et al. The Relationship between dorsolateral prefrontal neuronal N-acetylaspartate and evoked release of striatal dopamine in schizophrenia. Neuropsychopharmacology. 2000;22:125. doi: 10.1016/S0893-133X(99)00096-2. [DOI] [PubMed] [Google Scholar]
- 57.Abi-Dargham A, van de Giessen E, Slifstein M, Kegeles LS, Laruelle M. Baseline and amphetamine-stimulated dopamine activity are related in drug-naïve schizophrenic subjects. Biol Psychiatry. 2009;65:1091–3. doi: 10.1016/j.biopsych.2008.12.007. [DOI] [PubMed] [Google Scholar]
- 58.Laruelle M. Imaging synaptic neurotransmission with in vivo binding competition techniques: a critical review. J Cereb Blood Flow Metab. 2000;20:423–51. doi: 10.1097/00004647-200003000-00001. [DOI] [PubMed] [Google Scholar]
- 59.Trifilieff P, Ducrocq F, van der Veldt S, Martinez D. Blunted dopamine transmission in addiction: potential mechanisms and implications for behavior. Semin Nucl Med. 2017;47:64–74. doi: 10.1053/j.semnuclmed.2016.09.003. [DOI] [PubMed] [Google Scholar]
- 60.Nader MA, Morgan D, Gage HD, Nader SH, Calhoun TL, Buchheimer N, et al. PET imaging of dopamine D2 receptors during chronic cocaine self-administration in monkeys. Nat Neurosci. 2006;9:1050. doi: 10.1038/nn1737. [DOI] [PubMed] [Google Scholar]
- 61.Nader MA, Daunais JB, Moore T, Nader SH, Moore RJ, Smith HR, et al. Effects of cocaine self-administration on striatal dopamine systems in rhesus monkeys: initial and chronic exposure. Neuropsychopharmacology. 2002;27:35. doi: 10.1016/S0893-133X(01)00427-4. [DOI] [PubMed] [Google Scholar]
- 62.Perkins KA, Gerlach D, Vender J, Meeker J, Hutchison S, Grobe J. Sex differences in the subjective and reinforcing effects of visual and olfactory cigarette smoke stimuli. Nicotine Tob Res. 2001;3:141–50. doi: 10.1080/14622200110043059. [DOI] [PubMed] [Google Scholar]
- 63.Perkins KA, Jacobs L, Sanders M, Caggiula AR. Sex differences in the subjective and reinforcing effects of cigarette nicotine dose. Psychopharmacology. 2002;163:194–201. doi: 10.1007/s00213-002-1168-1. [DOI] [PubMed] [Google Scholar]
- 64.Perkins KA, Karelitz JL. Sex differences in acute relief of abstinence-induced withdrawal and negative affect due to nicotine content in cigarettes. Nicotine Tob Res. 2015;17:443–8. doi: 10.1093/ntr/ntu150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Perkins KA, Scott J. Sex differences in long-term smoking cessation rates due to nicotine patch. Nicotine Tob Res. 2008;10:1245–50. doi: 10.1080/14622200802097506. [DOI] [PubMed] [Google Scholar]
- 66.Arnsten AFT. Catecholamine influences on dorsolateral prefrontal cortical networks. Biol Psychiatry. 2011;69:e89–e99. doi: 10.1016/j.biopsych.2011.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sinha R. Chronic stress, drug use, and vulnerability to addiction. Ann N Y Acad Sci. 2008;1141:105–30. doi: 10.1196/annals.1441.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R, et al. Decreased striatal dopaminergic responsiveness in detoxified cocaine-dependent subjects. Nature. 1997;386:830–3. doi: 10.1038/386830a0. [DOI] [PubMed] [Google Scholar]
- 69.Wu JC, Bell K, Nijafi A, Widmark C, Keator D, Tang C, et al. Decreasing striatal 6-FDOPA uptake with increasing duration of cocaine withdrawal. Neuropsychopharmacology. 1997;17:402–9. doi: 10.1016/S0893-133X(97)00089-4. [DOI] [PubMed] [Google Scholar]
- 70.Martinez D, Greene K, Broft A, Kumar D, Liu F, Narendran R, et al. Lower level of endogenous dopamine in patients with cocaine dependence: findings from PET imaging of D 2 /D 3 receptors following acute dopamine depletion. Am J Psychiatry. 2009;166:1170–7. doi: 10.1176/appi.ajp.2009.08121801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Perkins KA, Grobe JE, Weiss D, Fonte C, Caggiula A. Nicotine preference in smokers as a function of smoking abstinence. Pharm Biochem Behav. 1996;55:257–63. doi: 10.1016/s0091-3057(96)00079-2. [DOI] [PubMed] [Google Scholar]
- 72.McFarland K, Kalivas PW. The circuitry mediating cocaine-induced reinstatement of drug-seeking behavior. J Neurosci. 2001;21:8655. doi: 10.1523/JNEUROSCI.21-21-08655.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Nordström A-L, Olsson H, Halldin CA. PET study of D2 dopamine receptor density at different phases of the menstrual cycle. Psychiatry Res. 1998;83:1–6. doi: 10.1016/s0925-4927(98)00021-3. [DOI] [PubMed] [Google Scholar]
- 74.Wong DF, Broussolle EP, Wand G, Villemagne V, Dannals RF, Links JM, et al. In vivo measurement of dopamine receptors in human brain by positron emission tomography age and sex differences. Ann N Y Acad Sci. 1988;515:203–14. doi: 10.1111/j.1749-6632.1988.tb32986.x. [DOI] [PubMed] [Google Scholar]
- 75.Munro CA, McCaul ME, Wong DF, Oswald LM, Zhou Y, Brasic J, et al. Sex differences in striatal dopamine release in healthy adults. Biol Psychiatry. 2006;59:966–74. doi: 10.1016/j.biopsych.2006.01.008. [DOI] [PubMed] [Google Scholar]
- 76.Mazure CM, Toll B, McKee SA, Wu R, O’Malley SS. Menstrual cycle phase at quit date and smoking abstinence at 6 weeks in an open label trial of bupropion. Drug Alcohol Depend. 2011;114:68–72. doi: 10.1016/j.drugalcdep.2010.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Allen SS, Bade T, Center B, Finstad D, Hatsukami D. Menstrual phase effects on smoking relapse. Addiction. 2008;103:809–21. doi: 10.1111/j.1360-0443.2008.02146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Beery AK, Zucker I. Sex bias in neuroscience and biomedical research. Neurosci Biobehav Rev. 2011;35:565–72. doi: 10.1016/j.neubiorev.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Heinrich J. GAO-01-286R. Washington, DC: ed Office USGA; 2001.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.