

Abstract
Background
Chronic Helicobacter pylori infection is the cause of peptic ulcers in a subpopulation of individuals and a risk factor for the development of gastric cancer. A vaccine against H pylori infection can prevent the acquisition of the infection and protect against reinfections. Clinical trials to date evaluating the efficacy of H pylori vaccines in human challenge models have shown moderate to poor protection with difficulties in predicting efficacy. Thus, while further studies are needed to design an effective vaccine, we also need to find relevant correlates for vaccine efficacy.
Objective
To find immune correlates to vaccine efficacy, the frequencies of neutrophils, eosinophils and inflammatory monocytes and CD4+ T‐cell memory and mucosa homing integrin α4β7+ cells were assessed by flow cytometry in the blood of mice after vaccination.
Materials and Methods
H pylori antigens and cholera toxin or the multiple mutant CT (mmCT) were administered via the sublingual (SL) and intragastric route (IG). The vaccinated mice were infected with H pylori strain SS1 bacteria, and colonization in the stomach and immune responses were evaluated.
Results
The H pylori vaccine was effective in reducing bacterial load in the stomach of mice and enhancing immune responses compared to unvaccinated infection controls. In the blood of mice after SL or IG route of vaccination, we observed changes in frequencies of innate and adaptive immune cell subsets compared to infection controls. Remarkably, the frequency of circulating mucosal homing α4β7+CD4+ T cells after vaccination correlated with low bacterial load in the stomach of individual mice irrespective of the immunization route.
Conclusions
Our study shows that the innate and adaptive immune cell subsets can be measured in the blood after vaccination and that increased frequency of α4β7+CD4+ in the blood after immunization could be used as a predictive marker for the efficacy of vaccine against H pylori infection.
Keywords: α4β7, CD4+T cells, Helicobacter pylori, IL‐17A and IFNγ, inflammatory monocytes, neutrophils
1. INTRODUCTION
Helicobacter pylori are spiral‐shaped gram‐negative bacteria that persistently colonize the gastric mucosa of over 50% of the world's population. Infection is associated with the development of gastrointestinal disorders such as peptic ulcer disease, gastric adenocarcinoma, and gastric mucosa‐associated lymphoid tissue (MALT) lymphoma in sub‐populations of individuals.1 The role of chronic H pylori infection and inflammation in the etiology of gastric cancer classifies H pylori a class I carcinogen, according to the World Health Organization.2 Natural infection does not induce protective immunity in a majority of infected individuals, and the infection can persist for decades with a mucosal infiltration of immune cells such as dendritic cells (DCs), macrophages, granulocytes and lymphocytes mediated by secreted cytokines and chemokines.3, 4
Protective immunity against H pylori infection can instead be induced by mucosal vaccination resulting in the reduction of bacterial load compared to sham‐vaccinated mice.5, 6 Vaccine‐induced reduction in bacterial load has been shown to largely depend on migration of CD4+ Th1 and Th17 cells, mast cells and neutrophils to the stomach mucosa but not CD8+ T cells or B cells.6, 7, 8, 9 Since H pylori infects via the mucosal route, it would be desirable that the vaccine is also administered via the mucosal route to generate local immune responses that could protect against reinfection. In this regard, the sublingual (under the tongue; SL) and intragastric (IG) routes of vaccination have been reported as the most relevant routes for mucosal vaccination against H pylori infection due to their potential to induce mucosal CD4+ T‐cell immune responses and cytokine secretion in the stomach which correlated with protection.6
The induction of mucosal immune responses against H pylori antigens requires a mucosal adjuvant, and cholera toxin (CT) has been included as an adjuvant of choice in the preclinical evaluation of many vaccine candidates in animal models. Although enterotoxic in humans, CT can promote strong mucosal CD4+ T‐cell responses to co‐administered antigens, that are protective against H pylori infection in mice.10 Extensive research over the last few years has focused on the generation of non‐toxic derivatives of CT while retaining significant adjuvant activity.11 The multiple mutated cholera toxin (mmCT) is a product of such research and has been developed by introducing multiple mutations in the enzymatically active cholera toxin A subunit (CTA), making it resistant to proteolytic cleavage which is required for the enterotoxicity.12 A detailed mechanism of adjuvant activity of mmCT has also been recently demonstrated in human in vitro activated co‐cultures of T cells and antigen‐presenting cells.13 Furthermore, preclinical studies have shown that mmCT is as effective as CT in promoting mucosal and systemic immune responses to the co‐administered model antigen ovalbumin and a H pylori whole cell vaccine and thus seems to be a promising mucosal adjuvant.14
During vaccination, dendritic cells can take up the vaccine antigens from the delivery site and migrate to the draining lymph node for presentation and activation of CD4+ T cells. Activated and differentiated CD4+ T cells can then migrate back to the mucosal sites including stomach and intestine via the blood guided by the expression of the mucosal tissue‐specific integrin molecule α4β7 which binds to the endothelial cell adhesion molecule, mucosal vascular addressin cell adhesion molecule 1 (MAdCAM‐1). At the mucosal site, the antigen‐specific CD4+α4β7+ T cells can then orchestrate anti‐bacterial effector functions. Most studies in mice have focused on evaluation of H pylori vaccine‐induced CD4+ T‐cell activation and correlates to protection locally in the stomach. However, for vaccine trials in humans, it would be useful to identify immune cell populations in circulation that are related to effective protection against H pylori infection in the stomach. Identification of such correlates of protection would greatly streamline the design and analysis of future vaccine trials.
In the current study, we have analyzed the innate and adaptive immune cell populations in the blood shortly after immunization of mice and correlated the immune cell populations to the levels of bacterial colonization. We report that, while the SL route of immunization with lysate antigens and adjuvant led to an increase in circulating neutrophils and CD62L− memory CD4+ T cells, the IG immunization with H pylori whole cell vaccine (WCV) and adjuvant led to an increase in circulation of inflammatory monocytes, memory CD62L− and CD4+α4β7+ cells. Protection in immunized mice only correlated with high frequency of CD4+α4β7+ cells in the circulation but not with the frequency of neutrophils, inflammatory monocytes or memory CD4+CD62L− T cells (both SL and IG). Furthermore, in accordance with our previous studies, we also confirmed significantly lower bacterial colonization rates in sublingually compared to intragastrically immunized mice.15 Altogether, the data presented in the current study will be useful for the design and evaluation of a H pylori vaccine in clinical trials in high‐risk H pylori‐infected individuals/patients.
2. MATERIALS AND METHODS
2.1. Animals
Female C57BL/6 mice (8‐week‐old) were purchased from Taconic and housed in microisolators at the Laboratory for Experimental Biomedicine, University of Gothenburg, Sweden. All experiments were approved by the animal ethics committee of the Gothenburg region.
2.2. Preparation of inactivated H pylori whole cell vaccine and antigens for immunizations
2.2.1. Inactivated H pylori bacteria strain Hel 305
Bacteria were grown in Brucella broth as previously described.16 The bacteria were harvested and washed in PBS and resuspended to an optical density of 1.5. Formaldehyde was added to a final concentration of 0.01 mol/L to the suspension and incubated for 2 hours at 37°C on a slow shaker, and then overnight shaking at room temperature. After washing to remove all formaldehyde, the inactivated bacteria were resuspended in PBS to an optical density of 1.5 and stored at 4°C. Complete inactivation of bacteria was confirmed by plating the bacteria on horse blood agar plates, and no colonies were found growing.
2.2.2. Lysates
Helicobacter pylori lysate antigens from strain Hel 305 (CagA+VacA+ Lex) were prepared and protein content measured as previously described.17 Briefly, the H pylori Hel 305 strain of bacteria were was grown on blood agar plates for 48 hours and the bacteria were harvested and resuspended in sterile PBS. The bacterial suspension was then sonicated for 30 minutes (30 seconds pulse at 50% capacity). The bacterial suspension was placed on ice during the entire sonication time. The protein content in the H pylori Hel 305 lysate was measured using the noninterfering protein assay kit (Calbiochem). To reduce the volume for sublingual immunization, the H pylori Hel 305 lysate antigens were aliquoted into sterile glass vials and freeze‐dried to a concentration of 50 mg/mL. The lyophilized H pylori Hel 305 lysate antigens were stored at 4°C until further use. In‐house monoclonal antibodies were used to carry out semi‐quantitative Western blot analysis of the H pylori Hel 305 lysate antigens and inactivated H pylori whole cell vaccine to confirm that both the vaccine preparations expressed H pylori antigens, urease, flagellin, CagA, and HpaA.
2.3. Immunizations
2.3.1. Intragastric immunizations
Groups of mice were immunized intragastrically on two consecutive days at bi‐weekly intervals with OD 0.5 of formalin‐killed whole cell bacteria corresponding to 1 × 109 bacteria/mL and 7.5 μg of CT (List Biological Laboratories) or 15 μg of mmCT12 with bicarbonate buffer on each occasion (hereafter referred to as IG + CT or IG + mmCT) (Figure 3A).
Figure 3.
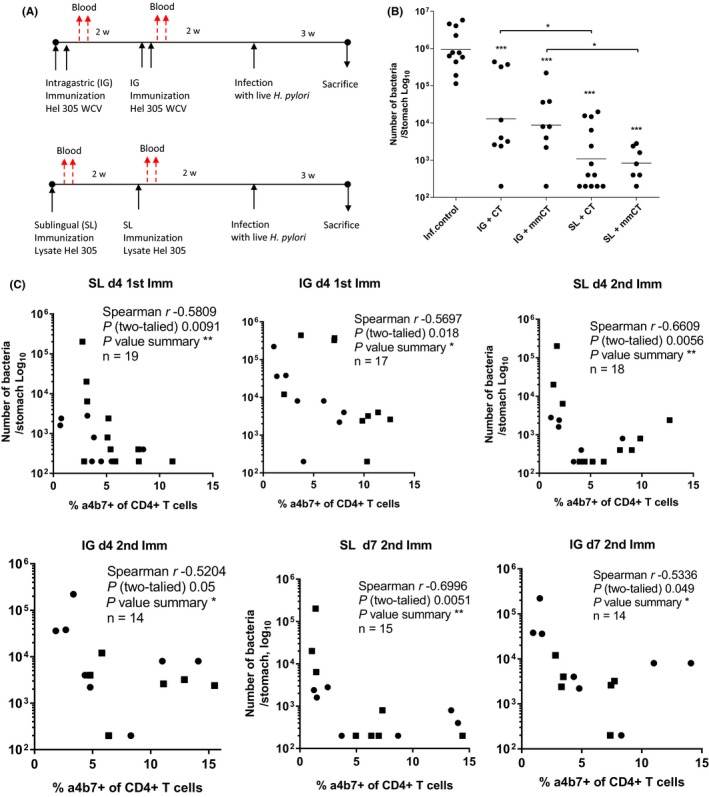
Protection against H pylori infection induced by intragastric or sublingual immunization with H pylori antigens and CT or mmCT. A, Groups of mice were immunized via the SL or IG route as indicated. Immunized and unimmunized mice were infected with a high dose of live H pylori SS1 bacteria, and the stomachs were collected to assess the protection against H pylori infection. B, Number of bacteria enumerated from the stomach of mice. Each dot represents one mouse, and the line indicates geometric mean. Combined data shown from three independent experiments giving similar results with n = 3‐5 mice per group per experiment. Statistical analysis of the reduction in gastric bacterial loads compared to inf. control was assessed by one‐way ANOVA with Dunnett's posttest, as denoted by ***P < .0001. C, Spearman correlation analysis between the frequency of α4β7 + CD4+ T cells in circulation days 4 and 7 after the 1st and 2nd immunization via the SL (left panel) or IG (right panel) route and the number of bacteria in the stomach in the same mice
2.3.2. Sublingual immunizations
Groups of mice were immunized sublingually twice at a bi‐weekly interval with 200 μg of H pylori lysate antigens reconstituted in 7.5 μg of CT or 15 μg of mmCT on each occasion (hereafter referred to as SL + CT or SL + mmCT) (Figure 3A). H pylori vaccination regimen (route and vaccine) was carefully chosen based on our previous studies.14, 18
2.3.3. Blood collection and Flow cytometry analysis
Blood was collected from the tail vein pre‐immunization (day 0) and on day 4, day 7 after the 1st and day 4, day 7 after the 2nd immunization for phenotypic analysis of leukocytes by multi‐parameter flow cytometry. Surface staining of the peripheral immune cells was performed using anti‐α4β7 PE (DATK‐32), anti‐Siglec‐F PE (E50‐2440), anti‐CD19 APC (eBio 1D3), anti‐CD62L BV605 (Mel‐14), anti‐Ly6C BV605 (HK 1.4), anti‐CD25 PerCP Cy5.5 (PC‐61), anti‐Ly6G PerCP Cy5.5 (1A8), anti‐TcRβ PE Cy7 (H57‐597), anti‐CD11b PE Cy7 (M1/70), anti‐CD11c V450 (N418), anti‐CD4 Alexa 700 (RM4‐5), and anti‐MHC II Alexa 700 (M5/114.15.2) followed by lysis of red blood cells and fixation in formaldehyde. Stained cells were immediately acquired on BD LSRII Flow Cytometer, and results were analyzed with FlowJo software (FlowJo, Treestar).
2.3.4. Cultivation of H pylori SS1 for infection
The mouse‐adapted H pylori Sydney strain 1 (SS1) (CagA+VacA+Ley) was cultured for infection as described in Ref. 17. A one‐time infectious dose consisting of approximately 3 × 108 viable bacteria/mouse was administered intragastrically to each mouse.19
2.3.5. Quantitation of bacteria in the stomach of mice
Immunized and unimmunized control mice were sacrificed 3 weeks after the infection to assess the protection induced by the prophylactic immunizations. The number of bacteria in the stomach was determined by quantitative culture as described previously.19
2.4. H pylori antigen‐specific cellular immune responses
2.4.1. Cell culture
Spleens from mice were collected after the sacrifice, and single‐cell suspensions were prepared and cultured.20 In short, cells were plated (2 × 105 cells per well) in the presence or absence of boiled H pylori strain Hel 305 lysate antigens (0.1 and 1 μg/mL) in complete Iscoves modified Dulbecco medium supplemented with supplemented with 10% heat‐inactivated fetal calf serum (Sigma), 50 µmol/L 2‐mercaptoethanol (Sigma), 1 mmol/L l‐glutamine (Biochrome), and 50 µg/mL gentamicin (Sigma) and incubated for 72 hours at 37°C in a 5% CO2 atmosphere.
2.4.2. Proliferation assay
The proliferation of the cells was assessed by incorporation of radioactive thymidine which was added in the last 7 hours of culture as previously described in detail in Ref. (18).
2.4.3. Detection of cytokines
Supernatants were collected before addition of thymidine and stored at −70°C for subsequent cytokine analysis. The cell culture supernatants were analyzed for IL‐2, IL‐17A, GM‐CSF, and IFNγ using the Luminex assay (Bio‐Plex Pro Assay; Bio‐Rad Laboratories) and analyzed according to the manufacturer's instructions.
2.5. Statistical and multivariate data analysis
Analysis of variance (ANOVA) with Dunnett's posttest was used to compare groups of mice with observations at various time points using GraphPad Prism software (GraphPad Software v6.0). For all tests, a P value of <0.05 was considered to be statistically significant. Spearman correlation analysis was carried out between the frequency of α4β7 + CD4+ T cells in circulation after immunization and the number of bacteria in the stomach in the same mice. To examine the relation between route of immunization (SL or IG) and the frequency of neutrophils, α4β7, in vitro proliferation and cytokines IL‐17A, IFNγ, and GM‐CSF in supernatant (X‐variables), multivariate factor analysis was used (SIMCA‐P+ software; Sartorius Stedim Data Analytics AB). Orthogonal partial least squares discriminant analyses (OPLS‐DA) was implemented to correlate selected Y‐variables and X‐variables with each other in linear multivariate models. The quality of the OPLS‐DA was based on the parameters R2Y, that is, the goodness of the fit of the model (values of ≥0.5 defines good discrimination, best possible fit, R2Y = 1) and Q2, that is, the goodness of prediction of the model. A Q2 value >0.4 is considered satisfactory with biological variables; furthermore, the difference between the Q2 and R2Y values should not exceed 0.2‐0.3. Variable influence on projection (VIP) was used as a variable selection based on discriminatory power. While variables with a VIP >1 are most influential for the model and are most relevant for explaining the Y observations, a VIP of 0.7‐0.8 is commonly used. In this study, a VIP cutoff of 0.7 was implemented. Finally, the statistical significance of the OPLS‐DA was also calculated and found to be significant (P < .001).
3. RESULTS
3.1. Increased frequency of innate immune cells in circulation after mucosal immunization against H pylori infection in mice
Our aim was to characterize the phenotype of circulating leukocytes in response to mucosal immunization with H pylori antigens and CT or mmCT as an adjuvant administered via sublingual (SL) or intragastric (IG) routes. We first analyzed the frequency of neutrophils (CD11b+Ly6G+), eosinophils (SSChighSiglec‐F+), and CD11b+Ly6G‐inflammatory monocytes (Ly6ChiMHCIIneg) in the blood of mice by multi‐parameter flow cytometry at day 0 (pre‐immunization), and then day 4, and day 7 after both 1st and 2nd immunization (Figure 1A).
Figure 1.
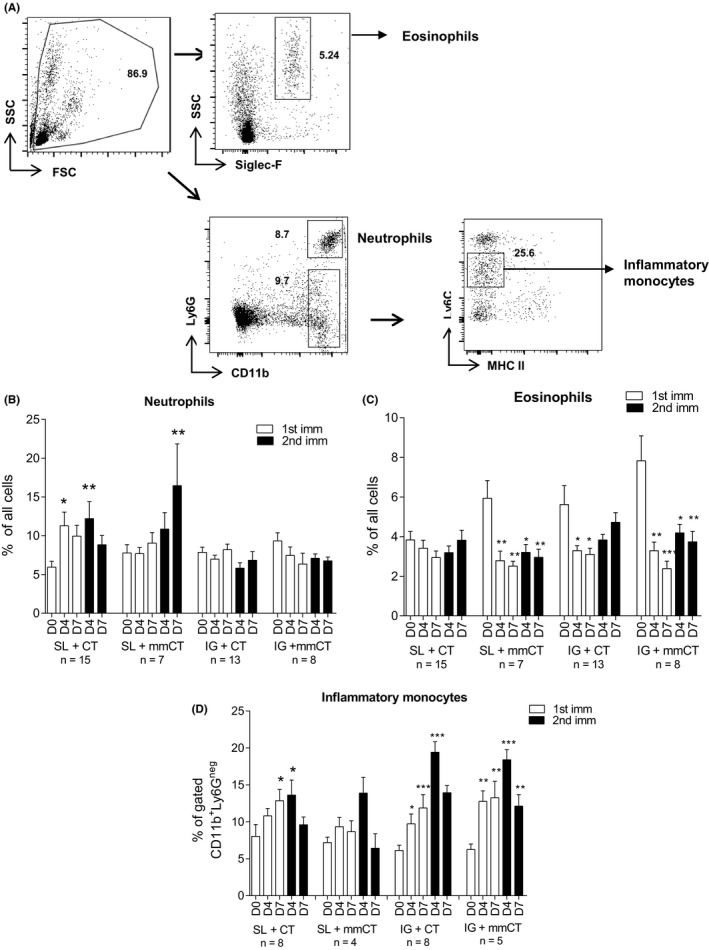
Sublingual immunization induced strong upregulation of neutrophils and inflammatory monocytes in circulation. A, Representative flow cytometry plots of leukocytes, Siglec‐F+ (eosinophils) CD11b+Ly6G+ (neutrophils) and CD11b+Ly6G−Ly6CintMHCII− (inflammatory monocytes among CD11b+Ly6G− cells). The numbers denote the frequency of cells within the gate. Blood was collected from mice at day 0 (D0), day (D4), and day 7 (D7) after both 1st and 2nd immunizations, and analyzed by flow cytometry the frequency of circulating (B) CD11b+Ly6G+ neutrophils (C) SSChighSiglec‐F+ eosinophils and (D) CD11b+Ly6GnegLy6Cint.MHCIIneg inflammatory monocytes. Data combined are shown from three independent experiments having similar results with n = 3‐5 mice per group per experiment. Bars indicate mean frequencies, and error bars represent standard errors of the mean (SEM) values. Significant upregulation in the frequency of cells at post‐immunizations compared to pre‐immunization was assessed by one‐way ANOVA with Dunnett's posttest, as denoted by *P < .05; **P < .001; ***P < .0001
In response to the SL + CT immunization, the frequency of circulating CD11b+Ly6G+ neutrophils was significantly upregulated at day 4 after 1st immunization (P < .05) and at day 4 (P < .05) after the 2nd immunization, compared to day 0 (Figure 1B). SL + mmCT immunization also induced an increase in circulating neutrophils, and this was significant only day 7 after the 2nd immunization (P < .05). The neutrophil frequency in blood was not affected by IG immunizations. In summary, neutrophils were enhanced in circulation shortly after SL but not IG immunizations, using CT and mmCT as adjuvants in the H pylori vaccine.
Although we found rather small population of Siglec‐F+ eosinophils in the circulation before vaccination, the frequency of Siglec‐F+ eosinophils was clearly downregulated from day 4 after the 1st immunization to day 7 after the 2nd immunization compared to day 0 in all groups of mice, except in the SL + CT immunized mice. Statistically significant downregulation in frequency of eosinophils in circulation was found in SL + mmCT, IG + CT, and IG + mmCT immunized mice (Figure 1C), suggesting a possible migration of these cells into the gastric mucosal tissue.7
We next analyzed the frequency of circulating CD11b+Ly6G− inflammatory monocytes (Ly6CintMHCIIneg) using a gating strategy used by Moyat et al21 and found them to be upregulated after SL + CT immunization day 7 after the 1st immunization (P < .05) and day 4 after the 2nd immunization (P < .05). A similar trend was seen in the SL + mmCT immunized mice, although the data were not statistically significant (Figure 1D). The increase in circulating CD11b+Ly6G− inflammatory monocytes was most striking after IG + CT immunization both at day 4 (P < .05) and day 7 (P < .001) after the 1st immunization, and even higher upregulation was observed at day 4 (P < .0001) after the 2nd immunization, compared to day 0. The patterns were similar in IG + mmCT immunized mice with significant upregulation of circulating CD11b+Ly6G− inflammatory monocytes at day 4 (P < .001) and day 7 (P < .001) after 1st immunization, and at day 4 (P < .0001) and day 7 (P < .001) after 2nd immunization, compared to day 0. All four groups of immunized mice had the highest peak circulation of CD11b+Ly6G− inflammatory monocytes day 4 after 2nd immunization.
No significant change in the frequencies of neutrophils, eosinophils, or CD11b+Ly6G‐inflammatory monocytes was found in the blood of naïve mice at any of the indicated time points (data not shown).
3.2. Increase in the frequency of either circulating memory and migrating CD4+T cells after immunization via mucosal routes
The crucial role of CD4+T cells in vaccine‐induced protection against H pylori infection has been reported in previous studies.9, 22 Therefore, we aimed to characterize the phenotype of circulating memory, mucosal and gut homing α4β7 +CD4+T cells in response to SL and IG immunizations in an attempt to find correlates of protection (Figure 2A). The total frequency of circulating CD4+T cells was unchanged at all post‐immunization time points compared to the pre‐immunization levels, (data not shown). The kinetics of the upregulation in frequency of circulating memory CD62L−CD4+T cells in response to immunization was found to be slightly different depending on whether CT or mmCT was the choice of adjuvant in the H pylori vaccine. Thus, immunization with H pylori antigens and CT via the SL or IG route led to significant upregulation in the frequency of memory CD4+CD62L− T cells both at day 4 (P < .05) and at day 7 (P < .01) after 1st immunization, and also at day 4 (P < .0001) and day 7 (P < .001) after 2nd immunization, compared to day 0. In contrast, in the mice immunized with H pylori antigens and mmCT as an adjuvant, a delayed response was observed compared to when using CT as an adjuvant, with an upregulation in the frequency of circulating CD62L‐CD4+T cells at day 7 after 1st immunization (P < .001) and day 7 after 2nd immunization (P < .001), compared to day 0 (Figure 2B).
Figure 2.
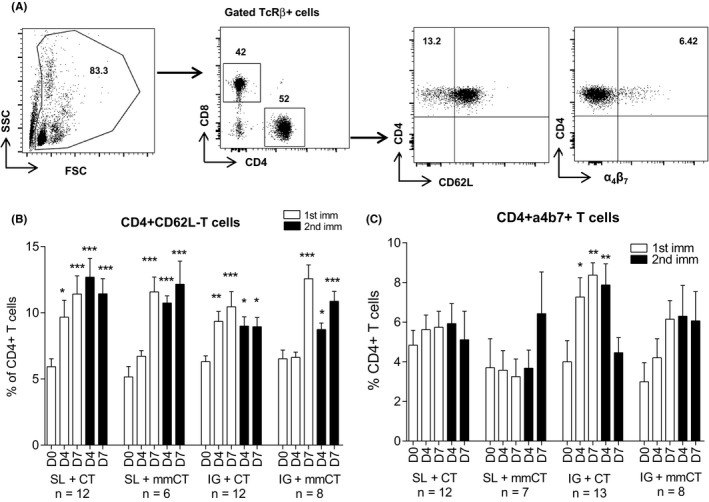
Upregulation in the frequency of CD4+TcRβ+ memory CD62L‐CD4+T and gut homing α4β7 + CD4+ T cells in the peripheral blood after intragastric immunization. A, Flow cytometry plots of CD4+T‐cell populations (TcRβ+) and CD62L and α4β7 +. In peripheral blood of mice at day 0 (D0), day (D4), and day 7 (D7) after both 1st and 2nd immunizations and frequency of the immunization induced (B) effector memory CD62L−CD4+T cells and (C) gut homing α4β7 + CD4+ T cells. Combined data are shown from three independent experiments giving similar results with n = 3‐5 mice per group per experiment. Bars indicate mean frequencies, and error bars represent standard errors of the mean (SEM) values. Significant upregulation in frequencies of cells at post‐immunizations compared to pre‐immunization was assessed by one‐way ANOVA with Dunnett's posttest, as denoted by *P < .05; **P < .001; ***P < .0001
We next chose to analyze the frequency of gut homing α4β7 +CD4+T cells among circulating CD4+T cells both after SL and IG immunizations8, 23 (Figure 2C). No clear trends in upregulation of α4β7 +CD4+T cells could be observed in the mice administered the H pylori vaccine and adjuvant CT or mmCT via the SL route. In contrast, immunization with H pylori antigens and CT via the IG route resulted in significant upregulation of the frequency of circulating α4β7 +CD4+T cells at day 4 (P < .05) and day 7 (P < .001) after the 1st immunization, and day 4 (P < .001) after the 2nd immunization compared to day 0. While IG + mmCT immunized groups of mice also showed tendencies for increased frequencies of α4β7 +CD4+ T cells both after 1st and 2nd immunizations compared to day 0, although the differences were not statistically significant (Figure 2C).
No particular trend for up‐ or downregulation of CD4+α4β7 +, CD44+CD62L− CD4+T cells could be found at any of the indicated time points in the blood of naïve unimmunized mice (data not shown).
In summary, the frequency of CD62L‐CD4+ memory T cells was upregulated after both SL and IG immunizations, while the frequency of gut homing α4β7 + CD4+T cells in circulation was upregulated after IG immunizations and to a lesser extent after SL immunizations.
3.3. Mucosal immunization with H pylori antigens and CT or mmCT promotes protection against H pylori infection
In order to evaluate the protection induced by the prophylactic SL and IG immunizations, the same groups of immunized and unimmunized mice analyzed for the frequencies of immune cells in circulation were also infected by gavage with live H pylori SS1 bacteria two weeks after the last immunization. Three weeks post‐infection, mice were sacrificed and the stomachs were collected to determine the numbers of H pylori bacteria (CFU) (Figure 3A). When compared to the unimmunized infection control group, the highest level of protection was found after SL + mmCT (P < .0001; mean, 1132‐fold reduction) as well as SL + CT immunizations (P < .0001, mean, 868‐fold reduction), while significant but not as large reduction in the gastric bacterial loads was found after IG + mmCT (P < .0001; mean, 107‐fold reduction) as well as IG + CT immunization (P < .0001; mean, 72‐fold reduction). The potent adjuvant effect of mmCT in the H pylori vaccine for reducing H pylori bacterial burden in the stomach of mice was completely comparable to when using CT as adjuvant (Figure 3B).
We further carried out correlation analysis between the frequency of CD4+α4β7 + cells in the blood day 4 or day 7 after 1st or 2nd immunization and the number of bacteria after challenge in the same mouse. We found a significant inverse correlation between the frequency of CD4+α4β7 + cells in the blood after SL or IG immunization, day 4 after 1st or day 4 and 7 after 2nd immunization and the number of bacteria in the stomach. Thus, mice with a higher frequency of CD4+α4β7 + cells in the blood early after SL or IG immunization were protected against H pylori infection (Figure 3C).
3.4. H pylori antigen‐specific proliferation and cytokine responses after sublingual or intragastric immunization
We next investigated the H pylori antigen‐specific cellular immune responses in immunized mice following H pylori infection. The proliferative response of splenocytes isolated from sublingually or intragastrically immunized and unimmunized infected mice (infection control) was examined by stimulating in vitro with or without H pylori Hel 305 lysate antigens. Compared to infection control, SL immunization showed significantly higher spleen proliferative responses to both 0.1 μg/mL (P < .0001) and 1 μg/mL (P < .0001) of Hel 305 lysate antigens. In mice immunized intragastrically with H pylori antigens and CT or mmCT as adjuvant, a significant increase in spleen proliferative responses was also seen to both 0.1 μg/mL (P < .05) and 1 μg/mL (P < .001) of lysate antigens, compared to infection control (Figure 4A). Overall, we found significantly higher spleen proliferative response and IL‐2 secretion specific to 0.1 and 1 μg/mL of lysate antigens and after both mucosal routes of immunizations with CT or mmCT adjuvant. We also determined the levels of the cytokines IL‐17A, GM‐CSF, and IFNγ (Figure 4B‐C). The concentrations of all of these cytokines were significantly elevated in response to SL or IG immunization irrespective of the type of adjuvant used. Splenocytes isolated from SL + CT immunized groups secreted the highest mean concentration for IFNγ in vitro in response H pylori antigens (mean, 170‐fold increase) and GM‐CSF (mean, 31‐fold increase), while splenocytes isolated from SL + mmCT immunized mice secreted high levels of IL‐17A (mean, 412‐fold increase) and IL‐2 (mean, 52‐fold increase), compared to splenocytes isolated from unimmunized but infected control group.
Figure 4.
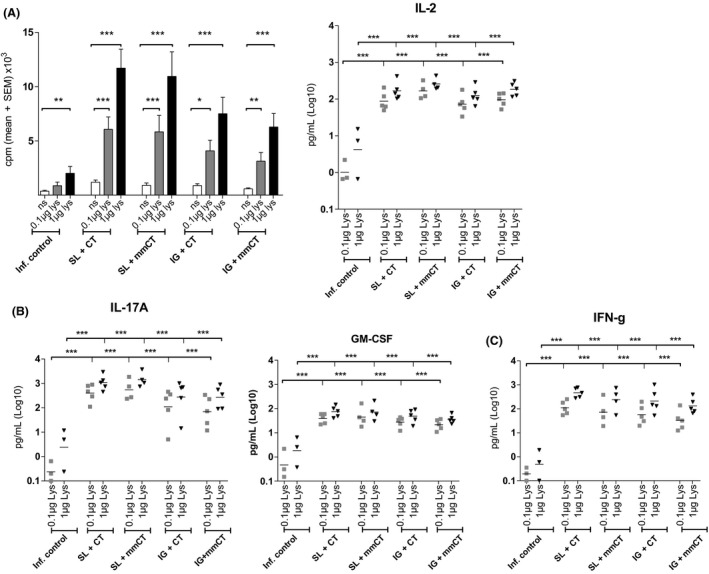
Cellular and cytokine responses specific to H pylori antigens in response to sublingual (SL) and intragastric (IG) immunization following infection. Splenocyte proliferative responses to H pylori antigens (0.1 or 1 μg/mL) by immunized infected and unimmunized infected mice, determined in vitro by the incorporation of radioactive thymidine indicated as mean counts per minute (cpm) + SEM and IL‐2 levels secreted by the splenocytes in cell culture supernatants (A). Combined data from two independent experiments giving similar results with n = 3‐5 mice per group. Supernatants from stimulation cultures analyzed using the Luminex assay for IL‐17A and GM‐CSF (B) and IFNγ (C). Data are shown from one experiment with n = 3‐5 mice per group. Each dot represents one mouse, and line indicates geometric mean. Statistical significance calculated using the one‐way ANOVA with Dunnett's multiple comparisons test, as denoted by *P < .05; **P < .001; ***P < .0001
3.5. Multivariate analysis reveals clustering of individual mice into groups depending on the route of immunization
We utilized a multivariate discriminant analysis (OPLS‐DA) to investigate the relationship between the route of immunization (SL or IG) and immune parameters studied in each mouse (X‐variables). The analysis showed that the two immunization groups SL and IG indeed clustered separately from each other based on the differences in bacterial colonization and adaptive immune response parameters (frequency of α4β7+CD4+ T cells, CD4+CD62L− day 4 after 1st immunization and proliferation counts (cpm) and cytokine secretion IL‐2, IL‐17A, GM‐CSF, and IFNγ measured at the time of sacrifice (R2 0.522, Q2 0.447) (Figure 5)). The clustering of groups of SL and IG immunized was found to be statistically significant (P < .001). A similar analysis, comparing the influence of the adjuvant included in the H pylori vaccine on the bacterial colonization and adaptive immune response (as above), showed that the mice immunized with vaccine and CT or mmCT (square and circle symbols) did not cluster to form distinct groups (R2 = 0.3 and Q2 0.1). In summary, our data show that protection against H pylori infection and the pattern of immune responses was unique to the route of immunization and not the adjuvant CT or mmCT used in the vaccine. Further, the selected immune parameters measured in this study can be used to predict the response to vaccination.
Figure 5.
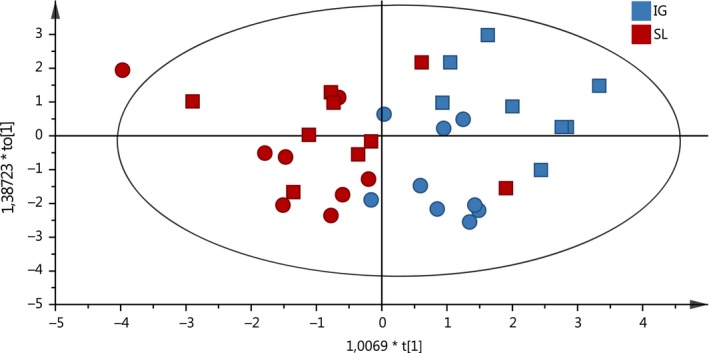
Multivariate analysis to elucidate the relationship between SL and IG immunized mice based on analysis of multiple immune parameters in the same mice. OPLS discriminant analysis (OPLS‐DA) displaying the separation between groups mice immunized via the SL (red) or IG (blue) route and with adjuvant CT (square symbol) or mmCT (circle). The X‐variables in the OPLS‐DA analysis were the frequency of α4β7 + CD4+ T cells, counts per minute of proliferation of spleen cells and cytokines IL‐17A, IFNγ, GM‐CSF, and IL‐2 in supernatant. R2 denotes the goodness of the fit of the model (values of ≥0.5 defines good discrimination), and Q2 denotes the goodness of prediction of the model (values of no more than 0.2‐0.3 less than the R2 value defines predictive ability)
4. DISCUSSION
In the current study, we have evaluated immune correlates in the blood to vaccine‐induced protection against H pylori infection. We were able to confirm previous findings that the non‐toxic adjuvant mmCT was as effective as an adjuvant as native CT in a H pylori vaccine for enhancing antigen‐specific T‐cell responses and cytokine secretion14 and report several important new findings, namely that (a) the populations of innate and memory cells are increased in the circulation after vaccination and that (b) the frequency of α4β7+CD4+ T cells emerging in the blood after 1st and 2nd immunization, irrespective of route or adjuvant correlated inversely with the bacterial load in the stomach of mice. We believe that the results from this study will be useful for the design of a H pylori vaccine and the evaluation of vaccine efficacy in clinical trials.
One of our first observations in vaccinated mice was the increase in the frequency of neutrophils in the blood particularly after SL immunization. This spike in the frequency neutrophils in the blood after SL vaccination was transient and returned to baseline level at the time of challenge with H pylori (data not shown). We have shown in a previous study a selective upregulation in the frequency of IL‐17A secreting CD4+ T cells in the draining lymph nodes of sublingually immunized but not intragastrically immunized mice. Thus, we believe that the mobilization of neutrophils in blood particularly after SL vaccination might play a role in the polarization of CD4+ T‐cell responses to Th17 responses in the draining lymph nodes. Indeed, we have observed a significantly higher frequency of IL‐17A+ cells in the stomach after infection with H pylori in sublingually immunized mice compared with intragastrically immunized mice.15 We also observed an increase in frequency of CD11b+Ly6G− inflammatory monocytes in the circulation after vaccination particularly after IG route of immunization in accordance with the study by Moyat et al Furthermore, we have previously reported the dependence of IFNγ on the protective immune response after IG immunization,20 and thus, the CD11b+Ly6G‐ inflammatory monocytes activated after IG route of vaccination might play a role in the polarization of the CD4+ T‐cell response to a Th1 phenotype.15, 20 Finally, we saw a decrease in frequency of eosinophils in circulation after vaccination possibly due to their migration to the effector tissues. We have previously reported an increase in frequency of eosinophils in the stomach of sublingually vaccinated mice after H pylori infection compared to unvaccinated mice.7 Eosinophils recruited to the stomach of H pylori‐infected mice may not contribute to the anti‐bacterial response but can promote the recruitment of immune cells to the stomach tissue through production of inflammatory mediators and chemokines. Thus, the pattern innate immune cell migration in the blood can be indicative of the route of immunization and polarization of the CD4+T‐cell response to a H pylori vaccine. We next focused our studies on the evaluation of the frequencies of circulating adaptive immune cells and particularly CD4+ T cells since they have been shown to be essential for protection against H pylori infection.
Previous studies have reported MAdCAM‐1 expression in the stomach of H pylori‐infected individuals24 and the presence of α4β7+ cells in the stomach of H pylori SS1‐infected mice.25 The function of α4β7+ cells in protection against Helicobacter infection was shown by blocking migration of α4β7+ cells to the gastric mucosa with antibodies which led to an abrogation of the protection in mice vaccinated and then infected with H felis. Having knowledge of these studies, we set out to investigate whether expression of the integrin α4β7 on CD4+ T cells in circulation after vaccination could be used as a biomarker for vaccine efficacy. We found that the frequencies of α4β7 + CD4+ T cells were elevated in the blood after vaccination and that the effect could be detected as early as 4 days after intragastric immunization with H pylori antigens and CT or mmCT as adjuvant. Remarkably, the frequency of α4β7 + CD4+ T cells in the circulation postvaccination (SL or IG) inversely correlated with bacterial numbers in the stomach of mice. A single clinical study has evaluated the frequency of α4β7 + CD4+ T cells in circulation after vaccination with recombinant H pylori urease and heat‐labile toxin from enterotoxigenic bacteria (LT) as an adjuvant.26 An increase in the frequency of α4β7 + CD4+ T cells in the circulation of urease and LT‐vaccinated compared to sham‐vaccinated individuals was documented. However, the frequency of α4β7 + CD4+ T cells in circulation could not be correlated with bacterial numbers in the stomach as the volunteers were not infected. Two human H pylori infection studies have been performed so far, using a challenge model that utilized a defined strain of H pylori 27, 28 but the expression of α4β7 + on CD4+ T cells in circulation was not studied. Based on our results, further studies are warranted both in mice and humans to further understand the migratory properties, antigen specificity, and phenotype of the α4β7 + CD4+ T cells if they will be utilized as correlates for protection. Ideally, the design of clinical trials would be enhanced if several biomarkers, including the frequency of α4β7 + CD4+ T cells could be studied in the blood of volunteers early after vaccination.
Our data show that mmCT is a promising candidate for a safe and effective mucosal adjuvant in a H pylori vaccine due to its non‐toxic properties and the ability to induce antigen‐specific T‐cell activation and cytokines IFNγ, IL‐17A, and IL‐2 secretion. The effects of mmCT that we observed on protection have also previously been reported for another non‐toxic mucosal adjuvant, the double mutant heat‐labile toxin from Escherichia coli (dmLT).18 The mutations in proteolytic cleavage site of the A subunit in both mmCT and dmLT lead to markedly reduced activity of adenylate cyclase, reduced cAMP levels and fluid secretion when compared to the native toxin.12 The advantage of mmCT over dmLT is that the mmCT protein can be produced in large quantities from V cholerae with high purity since it is secreted into the extracellular medium compared to dmLT that is found in inclusion bodies and requires additional purification steps which compromises the yield.12 If mmCT can be produced on an industrial GMP scale and tested for its safety in humans, it could be considered for inclusion in a low cost H pylori vaccine.
In summary, H pylori vaccines containing mmCT administered via mucosal routes lead to increased frequency of several immune cell populations during the prime‐boost stage of the vaccination protocol. In particular, the CD4+α4β7+ T‐cell population in the blood correlated with vaccine‐ induced protection and could potentially serve as a predictor for vaccine efficacy. These results will aid in the rational design and testing of a H pylori vaccine for use in humans.
ACKNOWLEDGEMENTS
This work was supported by the Sahlgrenska Academy, University of Gothenburg, Swedish Cancer Foundation (grant: 13 0411), Swedish Research Council (grant 348‐2014‐3071), and Swedish Research Links (grant 348‐2014‐4228). ICDDR,B is thankful to the donors for their support to its research efforts. ICDDR,B also gratefully acknowledges the following donors who provided unrestricted support: Government of the Peoplés Republic of Bangladesh, Global Affairs Canada (GAC), Swedish International Development Cooperation Agency (Sida), and the Department for International Development, UK (UKAid). The funding bodies did not play any role in the study design, collection, and interpretation of the data or writing of the manuscript.
Akter S, Jeverstam F, Lundgren A, et al. The frequency of circulating integrin α4β7+ cells correlates with protection against Helicobacter pylori infection in immunized mice. Helicobacter. 2019;24:e12658 10.1111/hel.12658
Akter and Jeverstam contributed equally to this study.
REFERENCES
- 1. Ernst PB, Gold BD. The disease spectrum of Helicobacter pylori: the immunopathogenesis of gastroduodenal ulcer and gastric cancer. Annu Rev Microbiol. 2000;54:615‐640. [DOI] [PubMed] [Google Scholar]
- 2. Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Cancer Incidence and Mortality Worldwide: IARC Cancer Base No. 10. GLOBCAN. 2008. Lyon: International Agency for Research on Cancer; 2010. [Google Scholar]
- 3. Raghavan S, Holmgren J, Svennerholm A‐M. Chapter 51 – Helicobacter pylori infection of the gastric mucosa In: Strober W, Russell MW, Kelsall BL, Cheroutre H, Lambrecht BN, eds. Mucosal Immunology (4th ed.). Boston, MA: Academic Press; 2015:985‐1001. [Google Scholar]
- 4. Lindholm C, Quiding‐Jarbrink M, Lonroth H, Hamlet A, Svennerholm AM. Local cytokine response in Helicobacter pylori‐infected subjects. Infect Immun. 1998;66(12):5964‐5971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Czinn SJ, Blanchard T. Vaccinating against Helicobacter pylori infection. Nat Rev Gastroenterol Hepatol. 2011;8(3):133‐140. [DOI] [PubMed] [Google Scholar]
- 6. Raghavan S, Quiding‐Järbrink M.Vaccination against Helicobacter pylori infection In: Backert S, Yamaoka Y. eds. Helicobacter pylori Research: From Bench to Bedside. Berlin: Springer; 2016:575‐603. [Google Scholar]
- 7. Flach CF, Mozer M, Sundquist M, Holmgren J, Raghavan S. Mucosal vaccination increases local chemokine production attracting immune cells to the stomach mucosa of Helicobacter pylori infected mice. Vaccine. 2012;30(9):1636‐1643. [DOI] [PubMed] [Google Scholar]
- 8. Michetti M, Kelly CP, Kraehenbuhl JP, Bouzourene H, Michetti P. Gastric mucosal alpha(4)beta(7)‐integrin‐positive CD4 T lymphocytes and immune protection against helicobacter infection in mice. Gastroenterology. 2000;119(1):109‐118. [DOI] [PubMed] [Google Scholar]
- 9. Pappo J, Torrey D, Castriotta L, Savinainen A, Kabok Z, Ibraghimov A. Helicobacter pylori infection in immunized mice lacking major histocompatibility complex class I and class II functions. Infect Immun. 1999;67(1):337‐341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lycke N. Recent progress in mucosal vaccine development: potential and limitations. Nat Rev Immunol. 2012;12(8):592‐605. [DOI] [PubMed] [Google Scholar]
- 11. Pizza M, Giuliani MM, Fontana MR, et al. Mucosal vaccines: non toxic derivatives of LT and CT as mucosal adjuvants. Vaccine. 2001;19(17–19):2534‐2541. [DOI] [PubMed] [Google Scholar]
- 12. Lebens M, Terrinoni M, Karlsson SL, et al. Construction and preclinical evaluation of mmCT, a novel mutant cholera toxin adjuvant that can be efficiently produced in genetically manipulated Vibrio cholerae . Vaccine. 2016;34(18):2121‐2128. [DOI] [PubMed] [Google Scholar]
- 13. Larena M, Holmgren J, Lebens M, Terrinoni M, Lundgren A. Cholera toxin, and the related nontoxic adjuvants mmCT and dmLT, promote human Th17 responses via cyclic AMP‐protein kinase A and inflammasome‐dependent IL‐1 signaling. J Immunol. 2015;194(8):3829‐3839. [DOI] [PubMed] [Google Scholar]
- 14. Holmgren J, Nordqvist S, Blomquist M, Jeverstam F, Lebens M, Raghavan S. Preclinical immunogenicity and protective efficacy of an oral Helicobacter pylori inactivated whole cell vaccine and multiple mutant cholera toxin: a novel and non‐toxic mucosal adjuvant. Vaccine. 2018;36(41):6223‐6230. [DOI] [PubMed] [Google Scholar]
- 15. Sjokvist Ottsjo L, Jeverstam F, Yrlid L, Wenzel AU, Walduck AK, Raghavan S. Induction of mucosal immune responses against Helicobacter pylori infection after sublingual and intragastric route of immunization. Immunology. 2017;150(2):172‐183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Flach CF, Ostberg AK, Nilsson AT, Malefyt Rde W, Raghavan S. Proinflammatory cytokine gene expression in the stomach correlates with vaccine‐induced protection against Helicobacter pylori infection in mice: an important role for interleukin‐17 during the effector phase. Infect Immun. 2011;79(2):879‐886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Raghavan S, Ostberg AK, Flach C‐F, et al. Sublingual immunization protects against Helicobacter pylori infection and induces T and B cell responses in the stomach. Infect Immun. 2010;78(10):4251‐4260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Sjokvist Ottsjo L, Flach CF, Clements J, Holmgren J, Raghavan S. A double mutant heat‐labile toxin from Escherichia coli, LT(R192G/L211A), is an effective mucosal adjuvant for vaccination against Helicobacter pylori infection. Infect Immun. 2013;81(5):1532‐1540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Raghavan S, Svennerholm AM, Holmgren J. Effects of oral vaccination and immunomodulation by cholera toxin on experimental Helicobacter pylori infection, reinfection, and gastritis. Infect Immun. 2002;70(8):4621‐4627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Sjokvist Ottsjo L, Flach CF, Nilsson S, Malefyt Rde W, Walduck AK, Raghavan S. Defining the Roles of IFN‐gamma and IL‐17A in inflammation and protection against Helicobacter pylori Infection. PLoS ONE. 2015;10(7):e0131444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Moyat M, Mack M, Bouzourene H, Velin D. Role of Inflammatory Monocytes in vaccine‐induced reduction of Helicobacter felis infection. Infect Immun. 2015;83(11):4217‐4228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ermak TH, Giannasca PJ, Nichols R, et al. Immunization of mice with urease vaccine affords protection against Helicobacter pylori infection in the absence of antibodies and is mediated by MHC class II‐restricted responses. J Exp Med. 1998;188(12):2277‐2288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Quiding‐Jarbrink M, Ahlstedt I, Lindholm C, Johansson EL, Lonroth H. Homing commitment of lymphocytes activated in the human gastric and intestinal mucosa. Gut. 2001;49(4):519‐525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Quiding‐Jarbrink M, Lonroth H, Ahlstedt I, Holmgren J, Svennerholm AM. Human gastric B cell responses can be induced by intestinal immunisation. Gut. 2001;49(4):512‐518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Hatanaka K, Hokari R, Matsuzaki K, et al. Increased expression of mucosal addressin cell adhesion molecule‐1 (MAdCAM‐1) and lymphocyte recruitment in murine gastritis induced by Helicobacter pylori . Clin Exp Immunol. 2002;130(2):183‐189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Michetti P, Kreiss C, Kotloff KL, et al. Oral immunization with urease and Escherichia coli heat‐labile enterotoxin is safe and immunogenic in Helicobacter pylori‐infected adults. Gastroenterology. 1999;116(4):804‐812. [DOI] [PubMed] [Google Scholar]
- 27. Graham DY, Opekun AR, Osato MS, et al. Challenge model for Helicobacter pylori infection in human volunteers. Gut. 2004;53(9):1235‐1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Aebischer T, Bumann D, Epple HJ, et al. Correlation of T‐cell response and bacterial clearance in human volunteers challenged with Helicobacter pylori revealed by randomised controlled vaccination with Ty21a‐based Salmonella vaccines. Gut. 2008;57(8):1065‐1072. [DOI] [PMC free article] [PubMed] [Google Scholar]