

Abstract
This study was conducted to assess the effects of dietary Clostridium butyricum on the growth, immunity, intestinal microbiota and disease resistance of tilapia (Oreochromis niloticus). Three hundreds of tilapia (56.21 ± 0.81 g) were divided into 5 groups and fed a diet supplemented with C. butyricum at 0, 1 x 104, 1 x 105, 1 x 106 or 1 x 107 CFU g-1 diet (denoted as CG, CB1, CB2, CB3 and CB4, respectively) for 56 days. Then 45 fish from each group were intraperitoneally injected with Streptococcus agalactiae, and the mortality was recorded for 14 days. The results showed that dietary C. butyricum significantly improved the specific growth rate (SGR) and feed intake in the CB2 group and decreased the cumulative mortality post-challenge with S. agalactiae in the CB2, CB3 and CB4 groups. The serum total antioxidant capacity and intestinal interleukin receptor-associated kinase-4 gene expression were significantly increased, and serum malondialdehyde content and diamine oxidase activity were significantly decreased in the CB1, CB2, CB3 and CB4 groups. Serum complement 3 and complement 4 concentrations and intestinal gene expression of tumour necrosis factor α, interleukin 8, and myeloid differentiation factor 88 were significantly higher in the CB2, CB3 and CB4 groups. Intestinal toll-like receptor 2 gene expression was significantly upregulated in the CB3 and CB4 groups. Dietary C. butyricum increased the diversity of the intestinal microbiota and the relative abundance of beneficial bacteria (such as Bacillus), and decreased the relative abundance of opportunistic pathogenic bacteria (such as Aeromonas) in the CB2 group. These results revealed that dietary C. butyricum at a suitable dose enhanced growth performance, elevated humoral and intestinal immunity, regulated the intestinal microbial components, and improved disease resistance in tilapia. The optimal dose was 1 x 105 CFU g-1 diet.
Introduction
In recent years, fish has increased in importance as a food source for humans, and aquaculture has rapidly developed [1]. With the expansion of the fish aquaculture industry, fish diseases have increased, resulting in serious economic losses. Antibiotics have been extensively used to prevent and control bacterial diseases [2]. However, the extensive usage of antibiotics may cause some negative effects, such as the emergence of antibiotic-resistant bacteria and antibiotic residues, which may affect the sustainable development of aquaculture and human health [3]. As an alternative, probiotics have received considerable attention in recent years [4]. Studies have shown that probiotics can improve growth [5–10], enhance the immune reponse [11–13], and improve disease resistance in fish [5,6,8–10,13,14]. Clostridium butyricum, a typical butyric acid bacterium, is a probiotic that occurs in the intestine of healthy humans [15] and other animals [16] and has been used in a wide range of human and veterinary intestinal diseases [17]. In aquatic animals, studies showed that survival post-challenge with pathogenic bacteria was significantly increased in Chinese drum (Miichthys miiuy) [18] and Pacific white shrimp (Litopenaeus vannamai) [19,20] fed a diet supplemented with C. butyricum for 30 and 42 days, respectively. Short-term supplementation with C. butyricum also improved the disease resistance of rainbow trout [21] and gibel carp [22]. Tilapia is the second most farmed fish worldwide [23]. To our knowledge, there are few studies about the effect of dietary C. butyricum on tilapia.
Fish disease resistance has been found to be associated with immunity [24,25]. The intestine is an important immune organ [25]. The intestinal immune response is related to inflammation mediated by cytokines [26]. Studies have shown that probiotics upregulate the gene expression of interleukin 8 (IL-8) in the intestine of rainbow trout (Oncorhynchus mykiss) [27] and grouper (Epinephelus coioides) [28] and that of tumour necrosis factor α (TNF-a) in the of intestine hybrid tilapia (Oreochromis niloticus x Oreochromis aureus) [29]. However, no studies have examined the effect of C. butyricum on cytokines in fish intestine. In HT-29 human colonic epithelial cells, C. butyricum improved the gene expression of the pro-inflammatory cytokines TNF-α and IL-8 [17,30] and the anti-inflammatory cytokine -IL-10 [17,31]. C. butyricum improved IL-6 and IL-8 gene expression in interstitial cells of Cajal (ICCs) [32]. Toll-like receptor (TLR) signalling pathways play an important role in the recognition of probiotics and activation of the intestinal immune system in mammals [28]. C. butyricum induced macrophages in inflamed mucosa producing IL-10 to prevent acute experimental colitis via the TLR2/Myeloid differentiation factor 88 (MyD88) signalling pathway [33]. TLR2 silencing alleviated C. butyricum-induced IL-6 and IL-8 expression and significantly inhibited C. butyricum-induced cell viability in ICCs [32]. C. butyricum upregulated TLR2 gene expression in HT-29 human colonic epithelial cells [17] and upregulated the gene expression of TLR2 and MyD88 in the intestine of weaning rex rabbits [34]. However, the roles of fish TLRs in the C. butyricum-induced intestinal immune response have not been reported. In grouper (Epinephelus coioides), the probiotic Psychrobacter sp. SE6 upregulated the gene expression of TLR2 and MyD88 in the intestine [28], which suggests that TLR2 signalling may play a key role in the modulation of intestinal immunity by C. butyricum.
The intestinal structural integrity is the foundation of intestinal immunity in fish [35]. The integrity of intestinal mucosa cells is reflected in serum diamine oxidase (DAO) activity [36]. Tight junction proteins are critical for maintaining the intercellular structural integrity of the intestine in mice [37,38]. A previous study found that dietary supplementation with C. butyricum decreased serum DAO in broiler chickens [39] and piglets [40]. Dietary C. butyricum upregulated the protein expression levels of ZO-1, claudin-3, and occludin in the intestines of weaned piglets [40] and the gene expression levels of ZO-1 and occludin in the ileum and colon of weaning rex rabbits [34]. To date, no reports have addressed the effect of C. butyricum on DAO gactivity and tight junction proteins in fish. Intestinal tissue showed close intestinal epithelium connections and healthy morphology in kuruma shrimp (Marsupenaeus japonicas) [41] and Pacific white shrimp (Litopenaeus vannamei) [42] fed diets supplemented with C. butyricum, suggesting that C. butyricum might affect intestinal structural integrity in fish, which needs to be further studied.
The intestinal microbiome is part of the host and interacts with the host immune system [43]. Studies revealed some disease association with dysbiosis or abnormal composition of the intestinal microbiome [44]. Studies have shown that C. butyricum can modify the intestinal microbiota. In vitro, C. butyricum inhibited the growth and adherence of potential pathogens to fish intestinal epithelial cells [45,46]. The potential intestinal pathogens were decreased, and the beneficial bacteria were increased in response to dietary C. butyricum in Litopenaeus vannamei [47], broiler chickens [48,49] and laying hens [50], and mice [16]. These data suggest that C. butyricum might affect the intestinal microbiome in fish, which needs to be further studied.
Therefore, the aim of the present study was to evaluate the effects of dietary supplementation with C. butyricum on growth performance and disease resistance in tilapia and the underlying mechanisms involved.
Materials and methods
Bacterial strains
C. butyricum (China Center for Type Culture Collection accession NO. M2014537) used in this experiment was obtained from Sun HY Bio Co., Ltd., Wuhan, China. Streptococcus agalactiae was kindly provided by Guangdong Ocean University, Zhanjiang, China.
Diets and feeding management
C. butyricum was cultured with reinforced Clostridium medium (RCM) (tryptone 10 g, beef extract 10 g, yeast extract 3 g, glucose 5 g, soluble starch 1 g, sodium chloride 5 g, sodium acetate 3 g, L-cysteine hydrochloride 0.5 g, distilled water 1000 mL) at 37°C for 24 h under anaerobic conditions. After cultivation, the bacteria were collected by centrifugation and resuspended in sterile saline at a concentration of 1x109CFU g-1. The bacterial cell quantity was examined using the spread RCM plate count method. The formulation of the basal diet is presented in Table 1. The bacterial suspension was gently sprayed on the basal diet, followed by thorough mixing, at five concentrations: 0 (control group), 1 x 104, 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet (denoted as CG, CB1, CB2, CB3 and CB4, respectively). The experimental diets were pelletized and stored at -20°C until use [51,52].
Table 1. Composition and nutrient content of the basal diet.
| Ingredients | g kg-1 diet | Nutrients contenta | g kg-1 diet |
|---|---|---|---|
| Fish meal | 50 | Crude protein | 323.4 |
| Soy bean meal | 300 | Crude fat | 60.6 |
| Cottonseed meal | 130 | Crude fibre | 48.3 |
| Rapeseed meal | 160 | Crude ash | 47.6 |
| Wheat flour | 200 | ||
| Rice bran | 54 | ||
| Cassava starch | 40 | ||
| Soybean oil | 37 | ||
| Ca(H2PO4)2 | 20 | ||
| Choline chloride (50%) | 3 | ||
| Vitamin premixb | 1 | ||
| Mineral premixb | 5 |
a The crude protein, crude fat, crude fibre and crude ash content are measured values.
b Kindly provided by New Hope Liuhe Co., Ltd, Chengdu, China.
The experimental protocol was approved by the Animal Care Advisory Committee of Huazhong Agricultural University. Juvenile tilapias were obtained from the Far East of Guangxi Agriculture and Animal Husbandry Fishery Development Co., Ltd. (Guangxi, China). Prior to the experiment, tilapia were cultured in experimental conditions (50 L/tank and 30 fish/tank) for 14 d and fed with the basal diet. After adaptation to the farming system, a total of 300 healthy tilapias (initial average weight 56.16 ± 0.82 g) were randomly divided into 15 tanks (50 L) with 20 fish/tank and fed one of five experimental diets to apparent satiety four times daily for 56 days. During the experiment, the water temperature and pH were 25–28°C and 7.0–7.5, respectively, and the dissolved oxygen content remained above 5 mg L-1.
Sample collection
At the end of the feeding trial, 5 fish per tank were anesthetized using tricaine methane sulfonate (MS222) according to Zhou et al. [53]. Blood samples were collected into non-heparinized tubes from the caudal vein and allowed to clot for 12 h (at 4°C). Then, serum was isolated by centrifugation (5000 g, 5 min) and stored at -20°C until further analysis as described by Hoseinifar et al. [54]. After blood sampling, these fishes were slaughtered and the hindguts were taken out. The luminal contents and hindgut tissues were collected and deposited at −80°C for later analysis. The remaining fishes were used for the challenge test.
Challenge test
The challenge test was approved by the Animal Care Advisory Committee of Huazhong Agricultural University and was conducted after the feeding trial. The remaining 45 fish in each group (triplicate) were anesthetized with MS222, and injected intraperitoneally with 0.3 mL of PBS containing 1.5 x 106 CFU bacteria mL-1. Then, these infected fish were returned to the original tank. All water quality parameters were the same as those in the feeding trial. The tanks were monitored every 2 h by a trained aquaculture technician. According to the previous description [55], any dead fish were removed, and any severely morbid fish were immediately anaesthetized and euthanized once noticed. All dead fish were examined for typical symptoms of streptococcosis. The challenge test lasted for 14 days.
The analysis of serum and hindgut tissue samples
According to Li et al. [56], serum total antioxidant capacity (T-AOC) and malondialdehyde (MDA) were analysed via kits provided by Nanjing Jiancheng Biological Engineering Research Institute (Nanjing, China). DAO activity was analysed via kits provided by Hangzhou Nuoyang Biological Technology Co. Ltd. (Hangzhou, China) according to Lei et al. [57]. Kits provided by Zhejiang Elikan Biological Technology Co. Ltd. (Wenzhou, China) were used for the determination of C3 and C4 according to Sun et al. [58].
Thirty hindgut tissue samples, six samples from each group, were used for RNA extraction. Total RNA was extracted using TRIzol according to the manufacturer's instructions (Aidlab Biotechnologies, Beijing, China). The quantity and quality of RNA were assessed using an ultraviolet spectrophotometer (Shanghai Sunny Hengping, Shanghai, China); the ratios of absorbance at 260 and 280 nm (A260/A280) were used to assess RNA purity and were between 1.8 and 2.0. Then, cDNA was synthesized using the Prime Script™ RT Reagent Kit according to the manufacturer's protocol (TaKaRa, Tokyo, Japan). Real-time PCR was applied to evaluate gene expression levels using gene-specific primers as shown in Table 2. β-actin was used as the house-keeping gene to standardize the data. Real-time PCR assays were conducted in an ABI 7900 real-time PCR detection system (Applied Biosystems, California, USA) with 20 μL reaction volumes containing: 4 μL of cDNA (10 times dilution), 0.4 μL of forward primer (100 μM), 0.4 μL of reverse primer (100 μM), 10 μL of 2× SYBR Premix Ex TaqTM (TaKaRa, Tokyo, Japan) and 5.2 μL of deionized H2O. The cycling conditions were as follows: 50°C for 2 min, 95°C for 10 min and then 40 cycles at 95°C for 30 s and 60°C for 30 s. The relative gene expression levels were analysed using the 2-ΔΔCT method according to Livak and Schmittgen [59].
Table 2. The gene specific primers sequences.
| Gene | Primer | Sequence | NCBI Gene ID |
|---|---|---|---|
| β-actin | Forward | 5’- TCCACGAAACCACCTACAACA -3’ | 100534414 |
| Reverse | 5’- CCAGACGGAGTATTTACGCTCA -3’ | ||
| claudin 1 | Forward | 5‘- CTTCACTCTGGTCGCCGTGTC -3’ | 100705074 |
| Reverse | 5‘- GCAGCAAAGCATAGATCCTCCC -3 | ||
| occludin | Forward | 5‘- AATCGGGATAATCTCCTACA -3’ | 100695261 |
| Reverse | 5‘- TTGGTCCTCTTTGCTATTTG -3’ | ||
| IL10 | Forward | 5‘- AGATGTCACCCAGTGTAGGAA -3’ | 100694754 |
| Reverse | 5‘- AAGCCAGGTACGTCTCAAAGT -3 | ||
| IL-8 | Forward | 5‘- ACCTGTGAAGGCATGGGTGT -3’ | 100534479 |
| Reverse | 5‘- TCGCAGTGGGAGTTGGGAAG -3 | ||
| TNF-a | Forward | 5‘- TCGTCGTCGTGGCTCTTTGT -3’ | 100534578 |
| Reverse | 5‘- GCCTTGGCTTTGCTGCTGAT -3 | ||
| TLR2 | Forward | 5‘- CATTCTGCTATCTGTGGTGCTGT -3’ | 100694547 |
| Reverse | 5‘- GCTGCTTTCGCTTGGCTCCTCTA -3 | ||
| MyD88 | Forward | 5‘- ATGCCTTCATCTGCTACTGC -3’ | 100700534 |
| Reverse | 5‘- ATCCGTTTACACCTCTTCTCG -3 | ||
| IRAK-4 | Forward | 5‘- ATACAAAGGTCTCCTGGATGA -3’ | 100707440 |
| Reverse | 5‘- AGCAAGCCAGTCGGTCTAAT -3 |
The analysis of the intestinal microbiota
According to the results of the feeding trial and challenge test, the most effective treatment in the present study was considered to be the CB2 group. Therefore, a comparison of bacterial structure and composition of the intestine between the CG and CB2 groups was performed. Twelve hindgut content samples, six each from the CG and CB2 groups, were used for analysis of the intestinal microbiota. Total microbial DNA was extracted using a TIANamp Stool DNA Kit (TIANGEN, Beijing, China) according to the manufacturer's guidelines, and the quality of the DNA samples was determined by agarose electrophoresis and a Nanodrop 8000 spectrophotometer (Thermo Fisher Scientific, Brisbane, Australia). The V3-V4 hypervariable region of the bacterial 16S rRNA gene was amplified by PCR (95°C for 2 min, followed by 25 cycles at 95°C for 30s, 55°C for 30 s, and 72°C for 30 s and a final extension at 72°C for 5 min) using forward primer 338F (5’- ACTCCTACGGGAGGCAGCAG-3’) and reverse primer 806R (5’–GGACTACHVGGGTWTCAAT-3’). PCR reactions were conducted in triplicate using a 20 μL reaction mixture consisting of template DNA (10 ng), FastPfu Polymerase (0.4 μL), each primer at 5 μM (0.8 μL), 2.5 mM dNTPs (2 μL) and 5 ×FastPfu Buffer (4 μL). Agarose gels (2%) were used to extract the amplicons, which were purified using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, U.S.) according to the manufacturer’s instructions and quantified using QuantiFluor™ -ST (Promega, U.S.). The amplicons from each sample were pooled in equimolar amounts and sent to Majorbio Bio-Pharm Technology Co., Ltd (Shanghai, China). The Illumina MiSeq platform was used to perform paired-end sequencing (2 × 250/300 bp) of these amplicons according to standard protocols.
In the present study, the Illumina sequencing reads were mainly analysed using QIIME pipeline software (version 1.9.1). The poor/low quality sequences, including those with uncertain nucleotides and three continuous nucleotides with an average quality of less than 20 over a 50bp sliding window, were removed. Operational Taxonomic Units (OTUs) were clustered with 97% similarity cut off using UPARSE (version 7.1 http://drive5.com/uparse/), and chimeric sequences were identified and removed using UCHIME. The taxonomy of each 16S rRNA gene sequence was analysed by RDP Classifier (http://rdp.cme.msu.edu/) against the Silva (SSU123) 16S rRNA database using a confidence threshold of 0.7. Mothur was also used to calculate the alpha diversity (Shannon index, Chao1, ACE and Sobs) and rarefaction curves. R programming language was used to perform the Venn diagram analysis. Afterwards, all sequence data were deposited to the SRA of the NCBI database under BioProject PRJNA559959.
Calculations and statistical analysis
Specific growth rate (SGR), feed conversion ratio (FCR) and cumulative mortality were calculated for each treatment according to [60,61].
SGR = {[ln (mean final weight) − ln (mean initial weight)] /number of days} x 100
FCR = feed intake (g)/wet weight gain (g)
Cumulative mortality = 100 x the number of dead fish/15
A nonparametric test of two samples was performed to assess the differences in bacterial relative abundance and diversity indices between the CB2 and CG groups. One-way ANOVA and Duncan's multiple comparison test were performed to assess the differences in growth performance, cumulative mortality, serum complement concentration, antioxidant capacity, gene expression, and DAO activity among the five groups. The significance level was set at P < 0.05. Statistical analyses were conducted using SPSS Statistics software v. 19.0 (IBM, Armonk, NY, USA). Results are presented as means ± standard deviation (SD).
Results
Growth performance
The effects of dietary C. butyricum on SGR, feed intake and FCR of tilapia are presented in Table 3. No fish died during the feeding experiment. SGR and feed intake were significantly improved in tilapia that were fed a diet with C. butyricum at a level of 1 x 105 CFU g-1 diet (P < 0.05), but FCR was not affected by dietary C. butyricum supplementation (P > 0.05).
Table 3. Growth Performance of tilapia fed diets supplemented with C.butyricum.
| CG | CB1 | CB2 | CB3 | CB4 | |
|---|---|---|---|---|---|
| Initial body weight (g) | 55.93±1.25a | 56.53±0.81a | 56.00±0.57a | 56.40±0.52a | 55.90±1.10a |
| Final body weight (g) | 178.77±4.70a | 187.20±8.55ab | 197.25±7.42b | 189.33±9.04ab | 184.53±7.22ab |
| SGR | 2.00±0.01a | 2.06±0.10ab | 2.17±0.08b | 2.09±0.10ab | 2.06±0.04ab |
| Feed intake (g fish-1) | 145.68±8.27a | 156.91±6.65ab | 163.24±6.39b | 156.94±7.47ab | 156.09±3.74ab |
| FCR | 1.19±0.04a | 1.20±0.04a | 1.16±0.02a | 1.18±0.03a | 1.22±0.03a |
SGR: Specific growth rate. FCR: feed conversion ratio.
Values are means ± standard deviation of three replicate groups. Mean values with the different superscripts in the same row are significantly different (P < 0.05).
Cumulative mortality
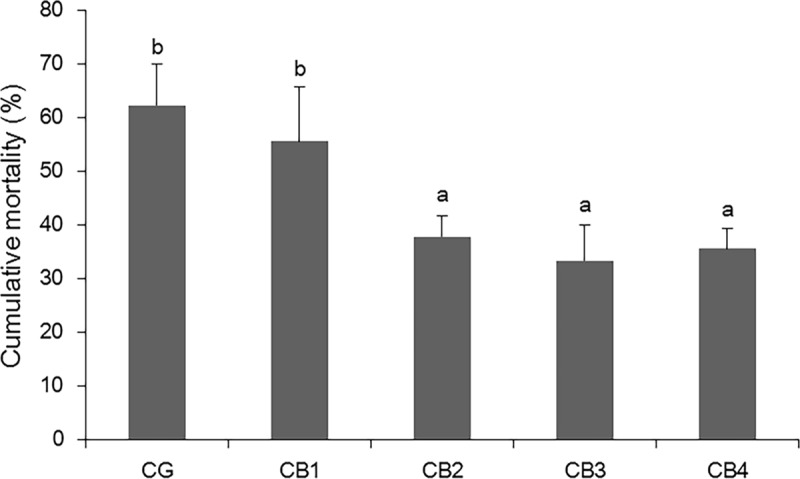
The cumulative mortality of fish at 14 days post-challenge with S. agalactiae is presented in Fig 1. C. butyricum significantly decreased the cumulative mortality of tilapia at levels of 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet (P < 0.05). Dead fish showed the typical symptoms of streptococcosis such as darkness of the body surface, exophthalmos, and basal fin haemorrhage.
Fig 1. The Effects of dietary C. butyricum on the cumulative mortality of tilapia post- challenge with S. agalactiae.
Serum complement concentration
The effects of dietary C. butyricum on serum C3 and C4 concentrations of tilapia are presented in Fig 2. Serum C3 and C4 concentrations increased significantly with dietary C. butyricum at levels of 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet (P < 0.05), with the highest levels observed in the CB3 group receiving the diet supplemented with C. butyricum at 1 x 106 CFU g-1 diet (P < 0.05).
Fig 2. The effects of dietary C. butyricum on serum complement 3 (C3) and 4 (C4) concentrations.

A: The effects on serum C3; B: The effects on serum C4.
Antioxidant capacity
The effects of dietary C. butyricum on the serum T-AOC and MDA contents of tilapia are presented in Fig 3. Serum T-AOC increased significantly with dietary C. butyricum, supplementation, with the highest levels observed in the CB4 group receiving the diet supplemented with C. butyricum at 1 x 107 CFU g-1 diet (P < 0.05). Serum MDA content significantly decreased with dietary C. butyricum supplementation, with the lowest content observed in the CB4 group receiving C. butyricum at 1 x 107 CFU g-1 diet (P < 0.05).
Fig 3. The effects of dietary C. butyricum on the serum T-AOC, MDA contents.

A: The effects on serum T-AOC; B: The effects on serum MDA.
Gene expression of cytokines, TLR2, MyD88, and IRAK-4
The effects of dietary C. butyricum on the gene expression of cytokines, TLR2, MyD88 and IRAK-4 in the hindgut of tilapia are presented in Fig 4. Compared with the unsupplemented group, gene expression of TNF-α, IL-8 and MyD88 was significantly upregulated by C. butyricum at levels of 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet (P < 0.05). TLR2 gene expression was significantly upregulated by C. butyricum supplementation, and significant upregulation was observed at levels of 1 x 106 and 1 x 107 CFU g-1 diet relative to the control group (P < 0.05). C. butyricum significantly upregulated IRAK-4 gene expression (P < 0.05) but did not affect IL-10 gene expression (P > 0.05).
Fig 4. The gene expression of TNF-α, IL-8, IL-10, TLR2, MyD88, IRAK-4 in the Hindgut of tilapia.

A: The effects on the gene expression levels of TNF-α; B: The effects on the gene expression levels of IL-8; C: The effects on the gene expression levels of IL-10; D: The effects on the gene expression levels of TLR2; E: The effects on the gene expression levels of MyD88; F: The effects on the gene expression levels of IRAK-4.
The intestinal physical barrier
The effects of dietary C. butyricum on serum DAO activity and the relative gene expression of claudin-1 and occludin in the hindgut of tilapia are presented in Fig 5. Serum DAO activity significantly decreased with dietary C. butyricum supplementation and was lowest in the CB4 group receiving C. butyricum at 1 x 107 CFU g-1 diet (P < 0.05). The relative gene expression of claudin-1 and occludin was not significantly affected by C. butyricum supplementation (P > 0.05).
Fig 5. The serum DAO activity and gene expression of Claudin -1 and Occludin in the hindgut.

A: The effects on serum DAO activity; B: The effects on the gene expression levels of claudin -1; C: The effects on the gene expression levels of occudin.
Intestinal microbiota analyses
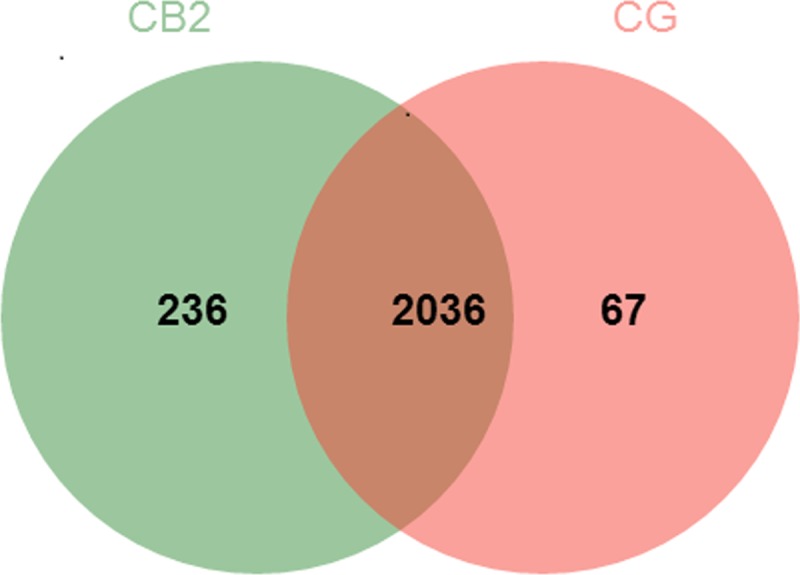
A total of 436184 valid sequences were obtained through Illumina sequencing analysis in the present study, with an average of 36348 (SD = 3585) per sample. A total of 2339 OTUs were identified at the 97% similarity level. There were 2036 OTUs shared between CG and CB2, 236 unique OTUS for the CB2 group and 67 unique OTUS for the CG (Fig 6).
Fig 6. Venn plot showing the shared and unique OTUs.
A rarefaction test was performed at the OTU level, and the results are presented in Fig 7. The rarefaction curves tended to approach the saturation plateau, indicating that the majority of microorganisms were revealed in the present study. Alpha diversity indices are presented in Table 4. Compared with the CG group, Sobs, Shannon, Ace and Chao1 indices were significantly increased by dietary supplementation with C. butyricum (1 x 105 CFU g-1 diet) (P < 0.05).
Fig 7. Rarefaction curves of samples.
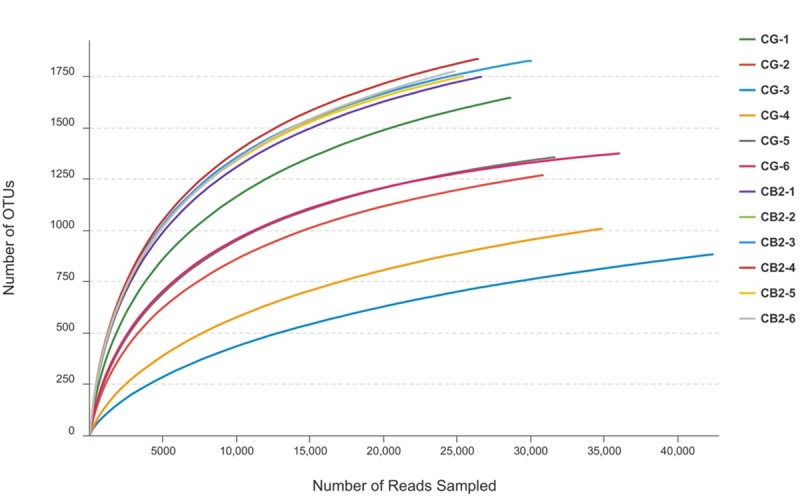
Table 4. Comparison of the diversity indices of bacteria communities between the CG and CB2 groups.
| Group | Sobs | Ace | Chao1 | Shannon |
|---|---|---|---|---|
| CG | 446.67±50.27a | 489.78±47.28a | 490.30±44.39a | 3.32±0.84a |
| CB2 | 514.00±7.90b | 547.30±5.30b | 552.24±5.73b | 4.99±0.05b |
Sobs: the observed richness; Ace: abundance-based coverage estimator; Vertical bars represented the means ± standard deviation (N = 6). Data indicated with different letters were significantly different (P < 0.05) between CG and CB2 group.
A total of 49 phyla, 108 classes, 210 orders, 394 families and 704 genera were identified in the present study. The bacterial compositions at the phylum and genus levels are presented in Fig 8. There were 42 phyla whose relative abundances were significantly different between the CG and CB2 groups, and the top 15 most abundance phyla are presented in Fig 9A. Compared to the control group, the relative abundances of Bacteroidetes, Firmicutes, Candidate-division-SR1, Chloroflexi, Chlorobi, Acidobacteria, Spirochaetae, Nitrospirae, Parcubacteria, Planctomycetes and WCHB1-60 were significantly enriched (P < 0.05), and those of Fusobacteria and CKC4 were significantly weakened in the CB2 group (P < 0.05). Proteobacteria was statistically the same between the two groups (P > 0.05). There were 354 genera whose relative abundances were significantly different between the CG and CG2 groups (P < 0.05), and the top15 most abundant genera are presented in Fig 9B. Compared with the CG group, the relative abundances of Cetobacterium, CKC4, Aeromonas and Gammaproteobacteria were significantly lower (P < 0.05), and those of Candidate_division_SR1_norank, Bacteroidetes_vadinHA17_norank, Dechloromonas, Zoogloea, Oligoflexales_noran, Comamonadaceae_unclassified, Draconibacteriaceae_uncultured, Saprospiraceae_uncultured, Nitrospira, Bacillus, and WCHB1-69_norank were significantly higher in the CB2 group (P < 0.05).
Fig 8. Bacterial composition at the phylum and genus levels of the CB2 and CG groups.

A: The bacterial compositions at the phylum level. B: The bacterial compositions at the genus level. Only the phylum and genus whose relative abundance was more than 1% were presented.
Fig 9. The differences at the phylum and genus level between the CB2 and CG groups.

A: at the phylum level. B: at the genus level. The vertical axis represented the species name at the phylum or genus level. Each column corresponding to the species represented the average relative abundance of the species in the CG and CB2 groups. Different colors represented different groups. Only the top15 most abundant genera were presented. 0.01< P < = 0.05 *, 0.001< P < = 0.01 **, P < = 0.001 ***.
Discussion
Probiotics have been extensively researched, and their effects on the growth and health of aquatic animals have been verified in recent years [3]. Compared with other probiotics, such as Bacillus and Lactobacillus, C. butyricum has a stronger tolerance to many antibiotics, higher temperature, and lower pH environment and has a wide range of application prospects [40,62]. Previous studies showed that a suitable dose of dietary C. butyricum increased growth performance in Pacific white shrimp (Litopenaeus vannamei) [19,20,42], black tiger shrimp (Penaeus monodon) [63], Macrobrachium rosenbergii [64], Chinese drum (Miichthys miiuy) [65] and silver pomfret (Pampus argenteus) [66]. The present study also found that dietary C. butyricum at a level of 1 x 105 CFU g-1 diet significantly improved the growth performance of tilapia. Fish growth performance is related to disease resistance [67]. Mortality post-challenge is an indicator of disease resistance [68,69]. In the present study, cumulative mortality post-challenge with S. agalactiae was decreased in tilapia fed a diet supplemented with C. butyricum at levels of 1 x 105, 1 x 106 and 1 x 107 CFU g-1diet. Similar results have been obtained in Chinese drum and rainbow trout. Pan et al. [18] reported that survival post-challenge with V. anguillarum or A. hydrophila was enhanced in Chinese drum fed diets supplemented with C. butyricum at the dose of 1 x 108 cells g-1 diet. After challenge with Vibrio parahaemolyticus, the cumulative mortality of Pacific white shrimp (Litopenaeus vannamei) significantly decreased with dietary C. butyricum at the dose of 1 x 108 CFU g-1diet [19] and 1 x 105, 1 x 107, 1 x 108, and 1 x 109 CFU g-1 diet [20]. These results revealed that C. butyricum could improve the disease resistance of aquatic animals.
Disease resistance is often related to humoural immunity, and the complement system is an important part of humoral immunity [70]. In fish, C3 and C4 play important roles in combatting bacterial pathogens [58]. Studies have shown that the probiotics Pediococcus acidilactici [54,71], Lactobacillus rhamnosus [72], Enterococcus faecium [73,74] and L. plantarum [5] can improve serum complement component levels in fish. However, no studies have examined the effect of C. butyricum on serum complement in fish. In this study, serum C3 and C4 concentrations were improved in tilapias fed a diet supplemented with C. butyricum at levels of 1 x 105, 1 x 106, 1 x 107CFU g-1 diet. Similar results were reported in which dietary C. butyricum improved serum C3 and C4 concentrations in broiler chickens [48,49]. The improvement of serum complement content induced by C. butyricum might be related to lipoteichoic acid (LTA). LTA is a major cell wall component of gram-positive bacteria [75,76]. In human, the classical complement pathway is activated by LTA in vitro [77]. In addition, immunity is related to antioxidative status in fish [78]. T-AOC is an anti-oxidative biomarker [79], and MDA is a typical parameter used to reflect oxidative injury [80]. In the present study, serum T-AOC was significantly increased and MDA content was significantly decreased in all four C. butyricum supplementary groups. Similar results were reported showing that dietary C. butyricum improved the antioxidant capacity in shrimp [20,41,42] and fish [66,81]. The effect of C. butyricum on antioxidant capacity of aquatic animals might be attributed to butyrate and H2. C. butyricum can produce butyric acid [82,83] and H2 [84]. Butyrate has been shown to reduce H2O2-induced DNA damage in human colon tumor cell [85] and to decrease MDA concentrations in the Caco-2 human colon carcinoma cell line [86]. Zhou et al. [87] reported that H2 reduced oxidative stress in acute pancreatitis both in vitro and in vivo.
The intestine is an important immune organ in fish [35]. Disease resistance in fish is related to intestinal immunity [25]. Cytokines play important roles in intestinal immunity [88]. Studies have shown that the gene expression of TNF-α and IL-8 in the fish intestine is upregulated by the dietary probiotics P. acidilactici [12] and L. plantarum [27]. Our results showed that the gene expression of TNF-α and IL-8 in the intestine was upregulated in tilapias fed a diet supplemented with C.butyricum at doses of 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet. Similar trends were reported in HT-29 human colonic epithelial cells [17,30] and the jejunal mucosa of broiler chickens [39]. The effect of C. butyricum on cytokines might be related to butyric acid. Sodium butyrate was found to upregulate the gene expression levels of TNF-α in the intestine of juvenile common carp [89] and in bovine mammary epithelial cells [90]. IL-10 is an anti-inflammatory cytokine that suppresses excessive immune responses [33]. In the present study, the gene expression of IL-10 was not influenced by dietary C. butyricum. This result differs from previous reports that C. butyricum upregulated the gene expression of IL-10 in HT-29 human colonic epithelial cells [17,31]. This difference might be related to immune status. In mouse intestinal macrophages, C. butyricum failed to increase the production of IL-10 in the steady state, whereas it induced IL-10 production under inflamed conditions [33]. The TLR2/MyD88/IRAK-4 signalling pathway plays an important role in intestinal immune responses, inducing the production of cytokines such as TNF-α and IL-8, in mammals [91]. Study has shown that the probiotic Psychrobacter sp. SE6 upregulated gene expression of TLR2 and MyD88 in the intestine of grouper (Epinephelus coioides) [28]. There has been no investigation of the effect of TLR signalling pathways in immune responses induced by C. butyricum in aquatic animals. In the present study, TLR2 gene expression was upregulated by dietary C. butyricum at levels of 1 x 106 and 1 x 107 CFU g-1 diet. MyD88 gene expression was significantly upregulated by dietary C. butyricum at levels of 1 x 105, 1 x 106 and 1 x 107 CFU g-1 diet. The gene expression of IRAK-4 was significantly upregulated in all four C. butyricum supplementary groups. Similar results were obtained showing that C. butyricum upregulate TLR2 gene expression in HT-29 human colonic epithelial cells [17], and upregulated the gene expression of TLR2 and MyD88 in the intestines of weaning rex rabbits [34]. The correlation analysis showed that TNF-α gene expression was significantly and positively related to gene expression of TLR2 (r = +0.932, P < 0.05), MyD88 (r = +0.969, P < 0.05) and IRAK-4 (r = +0.960, P < 0.05) and that IL-8 gene expression was significantly and positively related to gene expression of TLR2 (r = +0.957, P < 0.05), MyD88 (r = +0.936, P < 0.05) and IRAK-4 (r = +0.936, P < 0.05). These results indicated that the upregulation of TNF-α and IL-8 in response to dietary C. butyricum supplementation might be related to the upregulation of TLR2/MyD88/IRAK-4 signalling molecules in tilapia. To the best of our knowledge, this is the first study of the effects of C. butyricum on TLR2/MyD88/IRAK-4 signalling molecules in fish. The regulation of TLR2/MyD88/IRAK-4 signalling molecules by C. butyricum might be related to LTA. Rashidi et al. [92] reported that the gene expression of TLR2 and MyD88 was upregulated by LTA in human whole endometrial cells.
On the other hand, the effect of C. butyricum on intestinal immunity may be related to the modulation of intestinal microbial components. Studies have shown that the diversity and composition of the intestinal microbiota affect immune responses, including the regulation of immune cells, cytokines and TLR signalling molecule [93–95]. It has been reported that probiotics can regulate intestinal communities in tilapia [29,96–98]. In the present study, we investigated intestinal microbial composition in the CB2 and CG groups. Our results showed that a total of 49 phyla were identified, the relative abundances of 42 phyla were significantly different between the CG and CB2 groups, and the phylum with the greatest change was Fusobacteria, followed by CKC4, Bacteroidetes and Firmicutes. Cetobacterium is present in high numbers in many fish species [99–102] and produces large quantities of vitamin B12 [103]. In the present study, Cetobacterium was the dominant bacteria of Fusobacteria in the two groups, and its relative abundance was significantly decreased in the CB2 group. Similar to our results, a previous study showed that Cetobacterium abundance was significantly decreased by a multi-species probiotic (from 13.8% to 0.02%) in tilapia [96]. The reason that Cetobacterium was inhibited by probiotics was unclear and needs further research. CKC4, a phylum in the SILVA database, was detected in the intestines of zebrafish [104], white shrimp (Penaeus vannamei) [105] and Chinese mitten crab (Eriocheir sinensis) [106]. In the present study, dietary C. butyricum significantly decreased the relative abundance of CKC4. However, there is a lack of detailed information about the function of CKC4 in the fish intestine. The specific mechanism needs further research. Firmicutes is an abundant phylum in the tilapia intestine [96] and provides a good index of intestinal health [47]. In the present study, Firmicutes was enriched in the CB2 group relative to the CG group. Meanwhile, the relative abundance of Bacillus was significantly improved in the CB2 group. These results suggested that dietary C. butyricum could stimulate the growth of beneficial Firmicutes bacteria, such as Bacillus, in the tilapia intestine. A similar result was reported showing that Firmicutes, such as Bacillus and Clostridium, were abundant in Pacific white shrimp (Litopenaeus vannamei) fed a diet supplemented with C. butyricum [47]. Various opportunistic pathogens cause disease in stressed fish and fish affected by concurrent infection [107]. Aeromonas hydrophila is a conditional pathogen [108]. In the present study, the relative abundance of Aeromonas was significantly decreased in the CB2 group. A similar result was reported in Pacific white shrimp (Litopenaeus vannamei), showing that dietary C. butyricum inhibited some intestinal opportunistic pathogen genera [47]. In vitro, C. butyricum exhibited significant inhibitory activity on the growth and adherence of pathogenic bacteria to fish intestinal epithelial cells [45,46]. These results revealed that C. butyricum might decrease the risk of opportunistic pathogens to invade the host. Compared with disease states, there was a higher microbial diversity index in healthy humans [109], mice [110], pigs [111] and fish [110]. Stress caused a reduction in intestinal microbial diversity in fish [97]. In this study, alpha-diversity indices, including Sobs, Shannon, ACE and Chao1, were significantly increased by dietary C. butyricum. Similarly, C. butyricum increased the alpha-diversity indices, including ACE and Chao1, in Pacific white shrimp (Litopenaeus vannamei) [47]. These results indicated that C. butyricum improved the homeostasis of the intestinal microbial community.
In fish, the intestinal immunity is correlated with intestinal structural integrity [35]. DAO is an index of intestinal mucosal cell integrity [39]. In this study, serum DAO activity was decreased in tilapia that were fed diets supplemented with C. butyricum at all four levels. Similar results were reported in broiler chickens [39] and weaned piglets [40]. These results indicate that C.butyricum can improve intestinal mucosal cell integrity in fish. The observed decrease of DAO activity in the serum of fish fed C. butyricum might be associated with butyric acid. Butyric acid is a major respiratory fuel, is trophic to the colon and exerts proliferative effects on colonocytes [112]. Fang et al. [113] reported that serum DAO was significantly decreased by sodium butyrate in weanling piglets. In previous work, the intestinal intercellular structural integrity was found to be related to tight junction proteins, such as occludin and claudin, in the Caco-2 human colon carcinoma cell line [114]. Dietary C. Butyricum upregulated the protein expression levels of ZO-1, claudin-3, occludin in intestine of weaned piglets [40] and the gene expression levels of ZO-1 and occludin in ileum and colon of weaning rex rabbits [34]. In the present study, C. butyricum had no effect on the gene expression of claudin-1 and occludin in tilapia intestine. Nébot-Vivinus et al. [115] reported that in the T84 human colon epithelial cell line, multispecies probiotic LT did not affect the expression of occludin under basal conditions, but the expression of occludin was increased upon lipopolysaccharide stimulation. The effects of probiotics on tight junction protein might be related to health.
Conclusions
Dietary C. butyricum at a suitable dose significantly improved the growth performance, humoural and intestinal immunity, and integrity of the intestinal structure, elevated the diversity of the intestinal microbiota and the relative abundance of beneficial bacteria, decreased the relative abundance of opportunistic pathogenic bacteria, and enhanced the disease resistance in tilapia. According to the growth performance, immunity and disease resistance, the optimal dose of dietary C. butyricum was 1 x 105 CFU g-1 diet.
Acknowledgments
We are grateful to Lizhi Wang, Pei Wu, Weidan Jiang, Juan Zhao for their technical help.
Data Availability
All relevant data are within the manuscript and figures.
Funding Statement
Funded by New Hope Liuhe Co., Ltd. and Sun HY Bio Co., Ltd. The two funders provided support only in the form of salaries for authors [Hong-qin Li, Ying Zhou, Hua-yun Ling], and did not have any additional role in the study design, data collection and analysis, interpretation of data, writing of the paper, and decision to submit for publication. They don't serve on the editorial board of PLOS ONE, don't act as an expert witness in relevant legal proceedings, and don't sit on a committee for an organization that may benefit from publication of the paper.
References
- 1.Hai NV. The use of probiotics in aquaculture. J Appl Microbiol. 2015;119: 917–935. 10.1111/jam.12886 [DOI] [PubMed] [Google Scholar]
- 2.Wang A, Ran C, Wang Y, Zhang Z, Ding Q, Yang Y, et al. Use of probiotics in aquaculture of China-a review of the past decade. Fish Shellfish Immunol. 2019;86: 734–755. 10.1016/j.fsi.2018.12.026 [DOI] [PubMed] [Google Scholar]
- 3.Chauhan A, Singh R. Probiotics in aquaculture: a promising emerging alternative approach. Symbiosis. 2019;77: 99–113. [Google Scholar]
- 4.Lazado CC, Caipang CM, Estante EG. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions. Fish Shellfish Immunol. 2015;45: 2–12. 10.1016/j.fsi.2015.02.023 [DOI] [PubMed] [Google Scholar]
- 5.Son VM, Chang CC, Wu MC, Guu YK, Chiu CH, Cheng W. Dietary administration of the probiotic, Lactobacillus plantarum, enhanced the growth, innate immune responses, and disease resistance of the grouper Epinephelus coioides. Fish Shellfish Immunol. 2009;26: 691–698. 10.1016/j.fsi.2009.02.018 [DOI] [PubMed] [Google Scholar]
- 6.Gupta A, Gupta P, Dhawan A. Dietary supplementation of probiotics affects growth, immune response and disease resistance of Cyprinus carpio fry. Fish Shellfish Immunol. 2014;41: 113–119. 10.1016/j.fsi.2014.08.023 [DOI] [PubMed] [Google Scholar]
- 7.Ramos MA, Goncalves JF, Batista S, Costas B, Pires MA, Rema P, et al. Growth, immune responses and intestinal morphology of rainbow trout (Oncorhynchus mykiss) supplemented with commercial probiotics. Fish Shellfish Immunol. 2015;45: 19–26. 10.1016/j.fsi.2015.04.001 [DOI] [PubMed] [Google Scholar]
- 8.Aly SM, Ahmed YA-G, Ghareeb AA-A, Mohamed MF. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008;25: 128–136. 10.1016/j.fsi.2008.03.013 [DOI] [PubMed] [Google Scholar]
- 9.Aly SM, Mohamed MF, John G. Effect of probiotics on the survival, growth and challenge infection in tilapia nilotica (Oreochromis niloticus). Aquac Res. 2008;39: 647–656. [Google Scholar]
- 10.Geng X, Dong XH, Tan BP, Yang QH, Chi SY, Liu HY, et al. Effects of dietary chitosan and Bacillus subtilis on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Fish Shellfish Immunol. 2011;31: 400–406. 10.1016/j.fsi.2011.06.006 [DOI] [PubMed] [Google Scholar]
- 11.Panigrahi A, Kiron V, Satoh S, Hirono I, Kobayashi T, Sugita H, et al. Immune modulation and expression of cytokine genes in rainbow trout Oncorhynchus mykiss upon probiotic feeding. Dev Comp Immunol. 2007;31: 372–382. 10.1016/j.dci.2006.07.004 [DOI] [PubMed] [Google Scholar]
- 12.Standen BT, Rawling MD, Davies SJ, Castex M, Foey A, Gioacchini G, et al. Probiotic Pediococcus acidilactici modulates both localised intestinal- and peripheral-immunity in tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2013;35: 1097–1104. 10.1016/j.fsi.2013.07.018 [DOI] [PubMed] [Google Scholar]
- 13.Selim KM, Reda RM. Improvement of immunity and disease resistance in the nile tilapia, Oreochromis niloticus, by dietary supplementation with Bacillus amyloliquefaciens. Fish Shellfish Immunol. 2015;44: 496–503. 10.1016/j.fsi.2015.03.004 [DOI] [PubMed] [Google Scholar]
- 14.Iwashita MK, Nakandakare IB, Terhune JS, Wood T, Ranzani-Paiva MJ. Dietary supplementation with Bacillus subtilis, Saccharomyces cerevisiae and Aspergillus oryzae enhance immunity and disease resistance against Aeromonas hydrophila and Streptococcus iniae infection in juvenile tilapia Oreochromis niloticus. Fish Shellfish Immunol. 2015;43: 60–66. 10.1016/j.fsi.2014.12.008 [DOI] [PubMed] [Google Scholar]
- 15.Zhao X, Guo Y, Liu H, Gao J, Nie W. Clostridium butyricum reduce lipogenesis through bacterial wall components and butyrate. Appl Microbiol Biotechnol. 2014;98: 7549–7557. 10.1007/s00253-014-5829-x [DOI] [PubMed] [Google Scholar]
- 16.Kong Q, He GQ, Jia JL, Zhu QL, Ruan H. Oral administration of Clostridium butyricum for modulating gastrointestinal microflora in mice. Curr Microbiol. 2011;62: 512–517. 10.1007/s00284-010-9737-8 [DOI] [PubMed] [Google Scholar]
- 17.Gao Q, Qi L, Wu T, Xia T, Wang J. Immunomodulatory effects of Clostridium butyricum on human enterocyte-like HT-29 cells. Anim Cells Syst. 2013;17: 121–126. [Google Scholar]
- 18.Pan X, Wu T, Song Z, Tang H, Zhao Z. Immune responses and enhanced disease resistance in Chinese drum, Miichthys miiuy (Basilewsky), after oral administration of live or dead cells of Clostridium butyrium CB2. J Fish Dis. 2008;31: 679–686. 10.1111/j.1365-2761.2008.00955.x [DOI] [PubMed] [Google Scholar]
- 19.Li H, Tian X, Zhao K, Jiang W, Dong S. Effect of Clostridium butyricum in different forms on growth performance, disease resistance, expression of genes involved in immune responses and mTOR signaling pathway of Litopenaeus vannamai. Fish Shellfish Immunol. 2019;87: 13–21. 10.1016/j.fsi.2018.12.069 [DOI] [PubMed] [Google Scholar]
- 20.Li H-D, Tian X-L, Dong S-L. Growth performance, non-specific immunity, intestinal histology and disease resistance of Litopenaeus vannamei fed on a diet supplemented with live cells of Clostridium butyricum. Aquaculture. 2019;498: 470–481. [Google Scholar]
- 21.Sakai M, Yoshida T, Atsuta S, Kobayashi M. Enhancement of resistance to vibriosis in rainbow trout, Oncorhynchus mykiss (Walbaum), by oral administration of Clostridium butyricum bacterin. J Fish Dis. 1995;18: 187–190. [Google Scholar]
- 22.Li T, Ke F, Gui J-F, Zhou L, Zhang X-J, Zhang Q-Y. Protective effect of Clostridium butyricum against Carassius auratus herpesvirus in gibel carp. Aquac Int. 2019;27: 905–914. [Google Scholar]
- 23.Hai NV. Research findings from the use of probiotics in tilapia aquaculture: a review. Fish Shellfish Immunol. 2015;45: 592–597. 10.1016/j.fsi.2015.05.026 [DOI] [PubMed] [Google Scholar]
- 24.Wu P, Jiang J, Liu Y, Hu K, Jiang WD, Li SH, et al. Dietary choline modulates immune responses, and gene expressions of TOR and eIF4E-binding protein2 in immune organs of juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2013;35: 697–706. 10.1016/j.fsi.2013.05.030 [DOI] [PubMed] [Google Scholar]
- 25.Wen LM, Jiang WD, Liu Y, Wu P, Zhao J, Jiang J, et al. Evaluation the effect of thiamin deficiency on intestinal immunity of young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015;46: 501–515. 10.1016/j.fsi.2015.07.001 [DOI] [PubMed] [Google Scholar]
- 26.Wang B, Feng L, Chen GF, Jiang WD, Liu Y, Kuang SY, et al. Jian carp (Cyprinus carpio var. Jian) intestinal immune responses, antioxidant status and tight junction protein mRNA expression are modulated via Nrf2 and PKC in response to dietary arginine deficiency. Fish Shellfish Immunol. 2016;51: 116–124. 10.1016/j.fsi.2015.10.032 [DOI] [PubMed] [Google Scholar]
- 27.Perez-Sanchez T, Balcazar JL, Merrifield DL, Carnevali O, Gioacchini G, de Blas I, et al. Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish Immunol. 2011;31: 196–201. 10.1016/j.fsi.2011.05.005 [DOI] [PubMed] [Google Scholar]
- 28.Sun Y-Z, Xia H-Q, Yang H-L, Wang Y-L, Zou W-C. TLR2 signaling may play a key role in the probiotic modulation of intestinal microbiota in grouper Epinephelus coioides. Aquaculture. 2014;430: 50–56. [Google Scholar]
- 29.He S, Zhang Y, Xu L, Yang Y, Marubashi T, Zhou Z, et al. Effects of dietary Bacillus subtilis C-3102 on the production, intestinal cytokine expression and autochthonous bacteria of hybrid tilapia Oreochromis niloticus ♀×Oreochromis aureus ♂. Aquaculture. 2013;412–413: 125–130. [Google Scholar]
- 30.Gao Q, Qi L, Wu T, Wang J. Clostridium butyricum activates TLR2-mediated MyD88-independent signaling pathway in HT-29 cells. Mol Cell Biochem. 2012;361: 31–37. 10.1007/s11010-011-1084-y [DOI] [PubMed] [Google Scholar]
- 31.Gao Q, Qi L, Wu T, Wang J. An important role of interleukin-10 in counteracting excessive immune response in HT-29 cells exposed to Clostridium butyricum. BMC Microbiol. 2012;12: 100 10.1186/1471-2180-12-100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sui SJ, Tian ZB, Wang QC, Chen R, Nie J, Li JS, et al. Clostridium butyricum promotes intestinal motility by regulation of TLR2 in interstitial cells of Cajal. Eur Rev Med Pharmacol Sci. 2018;22: 4730–4738. 10.26355/eurrev_201807_15533 [DOI] [PubMed] [Google Scholar]
- 33.Kanai T, Mikami Y, Hayashi A. A breakthrough in probiotics: Clostridium butyricum regulates gut homeostasis and anti-inflammatory response in inflammatory bowel disease. J Gastroenterol. 2015;50: 928–939. 10.1007/s00535-015-1084-x [DOI] [PubMed] [Google Scholar]
- 34.Liu L, Zeng D, Yang M, Wen B, Lai J, Zhou Y, et al. Probiotic Clostridium butyricum improves the growth performance, immune function, and gut microbiota of weaning rex rabbits. Probiotics Antimicrob Proteins. 2018. 10.1007/s12602-018-9476-x [DOI] [PubMed] [Google Scholar]
- 35.Li L, Feng L, Jiang WD, Jiang J, Wu P, Kuang SY, et al. Dietary pantothenic acid deficiency and excess depress the growth, intestinal mucosal immune and physical functions by regulating NF-kappaB, TOR, Nrf2 and MLCK signaling pathways in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015;45: 399–413. 10.1016/j.fsi.2015.04.030 [DOI] [PubMed] [Google Scholar]
- 36.Ruan Z, Liu S, Zhou Y, Mi S, Liu G, Wu X, et al. Chlorogenic acid decreases intestinal permeability and increases expression of intestinal tight junction proteins in weaned rats challenged with LPS. PLoS One. 2014;9: e97815 10.1371/journal.pone.0097815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gong Y, Li H, Li Y. Effects of Bacillus subtilis on epithelial tight junctions of mice with inflammatory bowel disease. J Interferon Cytokine Res. 2016;36: 75–85. 10.1089/jir.2015.0030 [DOI] [PubMed] [Google Scholar]
- 38.Anderson RC, Cookson AL, McNabb WC, Park Z, McCann MJ, Kelly WJ, et al. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010;10: 316 10.1186/1471-2180-10-316 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zhang L, Zhang L, Zhan X, Zeng X, Zhou L, Cao G, et al. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function, and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. J Anim Sci Biotechnol. 2016;7: 3 10.1186/s40104-016-0061-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Li HH, Li YP, Zhu Q, Qiao JY, Wang WJ. Dietary supplementation with Clostridium butyricum helps to improve the intestinal barrier function of weaned piglets challenged with enterotoxigenic Escherichia coli K88. J Appl Microbiol. 2018;125: 964–975. 10.1111/jam.13936 [DOI] [PubMed] [Google Scholar]
- 41.Duan Y, Zhang Y, Dong H, Wang Y, Zhang J. Effect of the dietary probiotic Clostridium butyricum on growth, intestine antioxidant capacity and resistance to high temperature stress in kuruma shrimp Marsupenaeus japonicus. J Therm Biol. 2017;66: 93–100. 10.1016/j.jtherbio.2017.04.004 [DOI] [PubMed] [Google Scholar]
- 42.Duan Y, Zhang Y, Dong H, Wang Y, Zheng X, Zhang J. Effect of dietary Clostridium butyricum on growth, intestine health status and resistance to ammonia stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017;65: 25–33. 10.1016/j.fsi.2017.03.048 [DOI] [PubMed] [Google Scholar]
- 43.Li H, Gong Y, Xie Y, Sun Q, Li Y. Clostridium butyricum protects the epithelial barrier by maintaining tight junction protein expression and regulating microflora in a murine model of dextran sodium sulfate-induced colitis. Scand J Gastroenterol. 2018;53: 1031–1042. 10.1080/00365521.2016.1192678 [DOI] [PubMed] [Google Scholar]
- 44.Hagihara M, Yamashita R, Matsumoto A, Mori T, Inagaki T, Nonogaki T, et al. The impact of probiotic Clostridium butyricum MIYAIRI 588 on murine gut metabolic alterations. J Infect Chemother. 2019;25: 571–577. 10.1016/j.jiac.2019.02.008 [DOI] [PubMed] [Google Scholar]
- 45.Gao Q, Xiao Y, Sun P, Peng S, Yin F, Ma X, et al. In vitro protective efficacy of Clostridium butyricum against fish pathogen infections. Indian J Microbiol. 2013;53: 453–459. 10.1007/s12088-013-0394-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pan X, Wu T, Zhang L, Song Z, Tang H, Zhao Z. In vitro evaluation on adherence and antimicrobial properties of a candidate probiotic Clostridium butyricum CB2 for farmed fish. J Appl Microbiol. 2008;105: 1623–1629. 10.1111/j.1365-2672.2008.03885.x [DOI] [PubMed] [Google Scholar]
- 47.Duan Y, Wang Y, Dong H, Ding X, Liu Q, Li H, et al. Changes in the intestine microbial, digestive, and immune-related genes of litopenaeus vannamei in response to dietary probiotic Clostridium butyricum supplementation. Front Microbiol. 2018;9: 2191 10.3389/fmicb.2018.02191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhang L, Cao GT, Zeng XF, Zhou L, Ferket PR, Xiao YP, et al. Effects of Clostridium butyricum on growth performance, immune function, and cecal microflora in broiler chickens challenged with Escherichia coli K88. Poult Sci. 2014;93: 46–53. 10.3382/ps.2013-03412 [DOI] [PubMed] [Google Scholar]
- 49.Yang CM, Cao GT, Ferket PR, Liu TT, Zhou L, Zhang L, et al. Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens. Poult Sci. 2012;91: 2121–2129. 10.3382/ps.2011-02131 [DOI] [PubMed] [Google Scholar]
- 50.Zhan HQ, Dong XY, Li LL, Zheng YX, Gong YJ, Zou XT. Effects of dietary supplementation with Clostridium butyricum on laying performance, egg quality, serum parameters, and cecal microflora of laying hens in the late phase of production. Poult Sci. 2019;98: 896–903. 10.3382/ps/pey436 [DOI] [PubMed] [Google Scholar]
- 51.Lin Y-H, Ku C-Y, Shiau S-Y. Estimation of dietary magnesium requirements of juvenile tilapia, Oreochromis niloticus×Oreochromis aureus, reared in freshwater and seawater. Aquaculture. 2013;380–383: 47–51. [Google Scholar]
- 52.Shiau SY, Lo PS. Dietary choline requirements of juvenile hybrid tilapia, Oreochromis niloticus x O. aureus. J Nutr. 2000;130: 100–103. 10.1093/jn/130.1.100 [DOI] [PubMed] [Google Scholar]
- 53.Zhou P, Wang M, Xie F, Deng D-F, Zhou Q. Effects of dietary carbohydrate to lipid ratios on growth performance, digestive enzyme and hepatic carbohydrate metabolic enzyme activities of large yellow croaker (Larmichthys crocea). Aquaculture. 2016;452: 45–51. [Google Scholar]
- 54.Hoseinifar SH, Mirvaghefi A, Amoozegar MA, Sharifian M, Esteban MA. Modulation of innate immune response, mucosal parameters and disease resistance in rainbow trout (Oncorhynchus mykiss) upon synbiotic feeding. Fish Shellfish Immunol. 2015;45: 27–32. 10.1016/j.fsi.2015.03.029 [DOI] [PubMed] [Google Scholar]
- 55.Bayha KM, Ortell N, Ryan CN, Griffitt KJ, Krasnec M, Sena J, et al. Crude oil impairs immune function and increases susceptibility to pathogenic bacteria in southern flounder. PLoS One. 2017;12: e0176559 10.1371/journal.pone.0176559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Li W, Zhang X, Song W, Deng B, Liang Q, Fu L, et al. Effects of Bacillus preparations on immunity and antioxidant activities in grass carp (Ctenopharyngodon idellus). Fish Physiol Biochem. 2012;38: 1585–1592. 10.1007/s10695-012-9652-y [DOI] [PubMed] [Google Scholar]
- 57.Lei K, Li YL, Yu DY, Rajput IR, Li WF. Influence of dietary inclusion of Bacillus licheniformis on laying performance, egg quality, antioxidant enzyme activities, and intestinal barrier function of laying hens. Poult Sci. 2013;92: 2389–2395. 10.3382/ps.2012-02686 [DOI] [PubMed] [Google Scholar]
- 58.Sun YZ, Yang HL, Ma RL, Lin WY. Probiotic applications of two dominant gut Bacillus strains with antagonistic activity improved the growth performance and immune responses of grouper Epinephelus coioides. Fish Shellfish Immunol. 2010;29: 803–809. 10.1016/j.fsi.2010.07.018 [DOI] [PubMed] [Google Scholar]
- 59.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 2001;25: 402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- 60.Zhao J, Liu Y, Jiang J, Wu P, Chen G, Jiang W, et al. Effects of dietary isoleucine on growth, the digestion and absorption capacity and gene expression in hepatopancreas and intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture. 2012;368–369: 117–128. [Google Scholar]
- 61.Wang J, Xu H, Zuo R, Mai K, Xu W, Ai Q. Effects of oxidised dietary fish oil and high-dose vitamin E supplementation on growth performance, feed utilisation and antioxidant defence enzyme activities of juvenile large yellow croaker (Larmichthys crocea). Br J Nutr. 2016;115: 1531–1538. 10.1017/S0007114516000398 [DOI] [PubMed] [Google Scholar]
- 62.Liao XD, Ma G, Cai J, Fu Y, Yan XY, Wei XB, et al. Effects of Clostridium butyricum on growth performance, antioxidation, and immune function of broilers. Poult Sci. 2015;94: 662–667. 10.3382/ps/pev038 [DOI] [PubMed] [Google Scholar]
- 63.Duan Y, Zhang J, Huang J, Jiang S. Effects of dietary Clostridium butyricum on the growth, digestive enzyme activity, antioxidant capacity, and resistance to nitrite stress of Penaeus monodon. Probiotics Antimicrob Proteins. 2019;11: 938–945. 10.1007/s12602-018-9421-z [DOI] [PubMed] [Google Scholar]
- 64.Sumon MS, Ahmmed F, Khushi SS, Ahmmed MK, Rouf MA, Chisty MAH, et al. Growth performance, digestive enzyme activity and immune response of Macrobrachium rosenbergii fed with probiotic Clostridium butyricum incorporated diets. J King Saud Univ Sci. 2018;30: 21–28. [Google Scholar]
- 65.Song ZF, Wu TX, Cai LS, Zhang LJ, Zheng XD. Effects of dietary supplementation with Clostridium butyricum on the growth performance and humoral immune response in Miichthys miiuy. J Zhejiang Univ Sci B. 2006;7: 596–602. 10.1631/jzus.2006.B0596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gao Q, Xiao C, Min M, Zhang C, Peng S, Shi Z. Effects of probiotics dietary supplementation on growth performance, innate immunity and digestive enzymes of silver pomfret, Pampus argenteus. Indian J Anim Res. 2016;50: 936–941. [Google Scholar]
- 67.Xu HJ, Jiang WD, Feng L, Liu Y, Wu P, Jiang J, et al. Dietary vitamin C deficiency depresses the growth, head kidney and spleen immunity and structural integrity by regulating NF-kappaB, TOR, Nrf2, apoptosis and MLCK signaling in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2016;52: 111–138. 10.1016/j.fsi.2016.02.033 [DOI] [PubMed] [Google Scholar]
- 68.Wei Z, Yi L, Xu W, Zhou H, Zhang Y, Zhang W, et al. Effects of dietary nucleotides on growth, non-specific immune response and disease resistance of sea cucumber Apostichopus japonicas. Fish Shellfish Immunol. 2015;47: 1–6. 10.1016/j.fsi.2015.08.017 [DOI] [PubMed] [Google Scholar]
- 69.Zhou Q, Wang L, Wang H, Xie F, Wang T. Effect of dietary vitamin C on the growth performance and innate immunity of juvenile cobia (Rachycentron canadum). Fish Shellfish Immunol. 2012;32: 969–975. 10.1016/j.fsi.2012.01.024 [DOI] [PubMed] [Google Scholar]
- 70.Zhao J, Liu Y, Jiang J, Wu P, Jiang W, Li S, et al. Effects of dietary isoleucine on the immune response, antioxidant status and gene expression in the head kidney of juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2013;35: 572–580. 10.1016/j.fsi.2013.05.027 [DOI] [PubMed] [Google Scholar]
- 71.Neissi A, Rafiee G, Nematollahi M, Razavi SH, Maniei F. Influence of supplemented diet with Pediococcus acidilactici on non-specific immunity and stress indicators in green terror (Aequidens rivulatus) during hypoxia. Fish Shellfish Immunol. 2015;45: 13–18. 10.1016/j.fsi.2015.04.008 [DOI] [PubMed] [Google Scholar]
- 72.Pirarat N, Pinpimai K, Endo M, Katagiri T, Ponpornpisit A, Chansue N, et al. Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res Vet Sci. 2011;91: e92–e97. 10.1016/j.rvsc.2011.02.014 [DOI] [PubMed] [Google Scholar]
- 73.Wang Y-B, Tian Z-Q, Yao J-T, Li W-F. Effect of probiotics, Enteroccus faecium, on tilapia (Oreochromis niloticus) growth performance and immune response. Aquaculture. 2008;277: 203–207. [Google Scholar]
- 74.Sun YZ, Yang HL, Ma RL, Song K, Li JS. Effect of Lactococcus lactis and Enterococcus faecium on growth performance, digestive enzymes and immune response of grouper Epinephelus coioides. Aquac Nutr. 2012;18: 281–289. [Google Scholar]
- 75.Wang J, Qi L, Mei L, Wu Z, Wang H. C. butyricum lipoteichoic acid inhibits the inflammatory response and apoptosis in HT-29 cells induced by S. aureus lipoteichoic acid. Int J Biol Macromol. 2016;88: 81–87. 10.1016/j.ijbiomac.2016.03.054 [DOI] [PubMed] [Google Scholar]
- 76.Villeger R, Saad N, Grenier K, Falourd X, Foucat L, Urdaci MC, et al. Characterization of lipoteichoic acid structures from three probiotic Bacillus strains: involvement of D-alanine in their biological activity. Antonie Van Leeuwenhoek. 2014;106: 693–706. 10.1007/s10482-014-0239-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Monefeldt K, Helgeland K, Tollefsen T. In vitro activation of the classical pathway of complement by a streptococcal lipoteichoic acid. Oral Microbiol Immunol. 1994;9: 70–76. [DOI] [PubMed] [Google Scholar]
- 78.Biller-Takahashi JD, Takahashi LS, Mingatto FE, Urbinati EC. The immune system is limited by oxidative stress: dietary selenium promotes optimal antioxidative status and greatest immune defense in pacu Piaractus mesopotamicus. Fish Shellfish Immunol. 2015;47: 360–367. 10.1016/j.fsi.2015.09.022 [DOI] [PubMed] [Google Scholar]
- 79.Duan GL, Wang CN, Liu YJ, Yu Q, Tang XL, Ni X, et al. Resveratrol alleviates endotoxemia-associated adrenal insufficiency by suppressing oxidative/nitrative stress. Endocr J. 2016;63: 569–580. 10.1507/endocrj.EJ15-0610 [DOI] [PubMed] [Google Scholar]
- 80.Wang FY, Liu JM, Luo HH, Liu AH, Jiang Y. Potential protective effects of Clostridium butyricum on experimental gastric ulcers in mice. World J Gastroenterol. 2015;21: 8340–8351. 10.3748/wjg.v21.i27.8340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.He R-P, Feng J, Tian X-L, Dong S-L, Wen B. Effects of dietary supplementation of probiotics on the growth, activities of digestive and non-specific immune enzymes in hybrid grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀). Aquac Res. 2017;48: 5782–5790. [Google Scholar]
- 82.Zhang B, Yang X, Guo Y, Long F. Effects of dietary lipids and Clostridium butyricum on the performance and the digestive tract of broiler chickens. Arch Anim Nutr. 2011;65: 329–339. [DOI] [PubMed] [Google Scholar]
- 83.Nakanishi S, Kataoka K, Kuwahara T, Ohnishi Y. Effects of high amylose maize starch and Clostridium butyricum on metabolism in colonic microbiota and formation of azoxymethane-induced aberrant crypt foci in the rat colon. Microbiol Immunol. 2003;47: 951–958. 10.1111/j.1348-0421.2003.tb03469.x [DOI] [PubMed] [Google Scholar]
- 84.Dan J, Fang Z, Chin SX, Tian XF, Su TC. Biohydrogen production from hydrolysates of selected tropical biomass wastes with Clostridium butyricum. Sci Rep. 2016;6: 27205 10.1038/srep27205 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Rosignoli P, Fabiani R, De Bartolomeo A, Spinozzi F, Agea E, Pelli MA, et al. Protective activity of butyrate on hydrogen peroxide-induced DNA damage in isolated human colonocytes and HT29 tumour cells. Carcinogenesis. 2001;22: 1675–1680. 10.1093/carcin/22.10.1675 [DOI] [PubMed] [Google Scholar]
- 86.Courtois F, Seidman EG, Delvin E, Asselin C, Bernotti S, Ledoux M, et al. Membrane peroxidation by lipopolysaccharide and iron-ascorbate adversely affects Caco-2 cell function: beneficial role of butyric acid. Am J Clin Nutr. 2003;77: 744–750. 10.1093/ajcn/77.3.744 [DOI] [PubMed] [Google Scholar]
- 87.Zhou HX, Han B, Hou LM, An TT, Jia G, Cheng ZX, et al. Protective effects of hydrogen gas on experimental acute pancreatitis. PLoS One. 2016;11: e0154483 10.1371/journal.pone.0154483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhao J, Feng L, Liu Y, Jiang W, Wu P, Jiang J, et al. Effect of dietary isoleucine on the immunity, antioxidant status, tight junctions and microflora in the intestine of juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2014;41: 663–673. 10.1016/j.fsi.2014.10.002 [DOI] [PubMed] [Google Scholar]
- 89.Liu W, Yang Y, Zhang J, Gatlin DM, Ringo E, Zhou Z. Effects of dietary microencapsulated sodium butyrate on growth, intestinal mucosal morphology, immune response and adhesive bacteria in juvenile common carp (Cyprinus carpio) pre-fed with or without oxidised oil. Br J Nutr. 2014;112: 15–29. 10.1017/S0007114514000610 [DOI] [PubMed] [Google Scholar]
- 90.Alva-Murillo N, Medina-Estrada I, Baez-Magana M, Ochoa-Zarzosa A, Lopez-Meza JE. The activation of the TLR2/p38 pathway by sodium butyrate in bovine mammary epithelial cells is involved in the reduction of Staphylococcus aureus internalization. Mol Immunol. 2015;68: 445–455. 10.1016/j.molimm.2015.09.025 [DOI] [PubMed] [Google Scholar]
- 91.Kim TW, Staschke K, Bulek K, Yao J, Peters K, Oh KH, et al. A critical role for IRAK4 kinase activity in toll-like receptor-mediated innate immunity. J Exp Med. 2007;204: 1025–1036. 10.1084/jem.20061825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Rashidi N, Mirahmadian M, Jeddi-Tehrani M, Rezania S, Ghasemi J, Kazemnejad S, et al. Lipopolysaccharide- and lipoteichoic acid-mediated pro-inflammatory cytokine production and modulation of TLR2, TLR4 and MyD88 expression in human endometrial cells. J Reprod Infertil. 2015;16: 72–81. [PMC free article] [PubMed] [Google Scholar]
- 93.Kamada N, Seo SU, Chen GY, Nunez G. Role of the gut microbiota in immunity and inflammatory disease. Nat Rev Immunol. 2013;13: 321–335. 10.1038/nri3430 [DOI] [PubMed] [Google Scholar]
- 94.Molloy MJ, Bouladoux N, Belkaid Y. Intestinal microbiota: shaping local and systemic immune responses. Semin Immunol. 2012;24: 58–66. 10.1016/j.smim.2011.11.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Nowak P, Troseid M, Avershina E, Barqasho B, Neogi U, Holm K, et al. Gut microbiota diversity predicts immune status in HIV-1 infection. AIDS. 2015;29: 2409–2418. 10.1097/QAD.0000000000000869 [DOI] [PubMed] [Google Scholar]
- 96.Standen BT, Rodiles A, Peggs DL, Davies SJ, Santos GA, Merrifield DL. Modulation of the intestinal microbiota and morphology of tilapia, Oreochromis niloticus, following the application of a multi-species probiotic. Appl Microbiol Biotechnol. 2015;99: 8403–8417. 10.1007/s00253-015-6702-2 [DOI] [PubMed] [Google Scholar]
- 97.Zhai Q, Yu L, Li T, Zhu J, Zhang C, Zhao J, et al. Effect of dietary probiotic supplementation on intestinal microbiota and physiological conditions of nile tilapia (Oreochromis niloticus) under waterborne cadmium exposure. Antonie Van Leeuwenhoek. 2017;110: 501–513. 10.1007/s10482-016-0819-x [DOI] [PubMed] [Google Scholar]
- 98.Ferguson RM, Merrifield DL, Harper GM, Rawling MD, Mustafa S, Picchietti S, et al. The effect of Pediococcus acidilactici on the gut microbiota and immune status of on-growing red tilapia (Oreochromis niloticus). J Appl Microbiol. 2010;109: 851–862. 10.1111/j.1365-2672.2010.04713.x [DOI] [PubMed] [Google Scholar]
- 99.Zhang Z, Li D, Refaey MM, Xu W. High spatial and temporal variations of microbial community along the southern Catfish gastrointestinal tract: insights into dynamic food digestion. Front Microbiol. 2017;8: 1531 10.3389/fmicb.2017.01531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.She R, Li TT, Luo D, Li JB, Yin LY, Li H, et al. Changes in the intestinal microbiota of gibel carp (Carassius gibelio) associated with Cyprinid herpesvirus 2 (CyHV-2) infection. Curr Microbiol. 2017;74: 1130–1136. 10.1007/s00284-017-1294-y [DOI] [PubMed] [Google Scholar]
- 101.Larsen AM, Mohammed HH, Arias CR. Characterization of the gut microbiota of three commercially valuable warmwater fish species. J Appl Microbiol. 2014;116: 1396–1404. 10.1111/jam.12475 [DOI] [PubMed] [Google Scholar]
- 102.Li T, Long M, Gatesoupe FJ, Zhang Q, Li A, Gong X. Comparative analysis of the intestinal bacterial communities in different species of carp by pyrosequencing. Microb Ecol. 2015;69: 25–36. 10.1007/s00248-014-0480-8 [DOI] [PubMed] [Google Scholar]
- 103.Tsuchiya C, Sakata T, Sugita H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett Appl Microbiol. 2008;46: 43–48. 10.1111/j.1472-765X.2007.02258.x [DOI] [PubMed] [Google Scholar]
- 104.Liu Y, Yao Y, Li H, Qiao F, Wu J, Du ZY, et al. Influence of endogenous and exogenous estrogenic endocrine on intestinal microbiota in Zebrafish. PLoS One. 2016;11: e0163895 10.1371/journal.pone.0163895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Gainza O, Ramirez C, Ramos AS, Romero J. Intestinal microbiota of white shrimp Penaeus vannamei under intensive cultivation conditions in ecuador. Microb Ecol. 2018;75: 562–568. 10.1007/s00248-017-1066-z [DOI] [PubMed] [Google Scholar]
- 106.Zhang M, Sun Y, Chen L, Cai C, Qiao F, Du Z, et al. Symbiotic bacteria in gills and guts of Chinese mitten crab (Eriocheir sinensis) differ from the free-living bacteria in water. PLoS One. 2016;11: e0148135 10.1371/journal.pone.0148135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.van der Marel M, Schroers V, Neuhaus H, Steinhagen D. Chemotaxis towards, adhesion to, and growth in carp gut mucus of two Aeromonas hydrophila strains with different pathogenicity for common carp, Cyprinus carpio L. J Fish Dis. 2008;31: 321–330. 10.1111/j.1365-2761.2008.00902.x [DOI] [PubMed] [Google Scholar]
- 108.Harikrishnan R, Balasundaram C. Modern trends in Aeromonas hydrophila disease management with fish. Reviews in Fisheries Science. 2005;13: 281–320. [Google Scholar]
- 109.Ott SJ, Musfeldt M, Wenderoth DF, Hampe J, Brant O, Folsch UR, et al. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut. 2004;53: 685–693. 10.1136/gut.2003.025403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Sun J, Wang F, Ling Z, Yu X, Chen W, Li H, et al. Clostridium butyricum attenuates cerebral ischemia/reperfusion injury in diabetic mice via modulation of gut microbiota. Brain Res. 2016;1642: 180–188. 10.1016/j.brainres.2016.03.042 [DOI] [PubMed] [Google Scholar]
- 111.Konstantinov SR, Favier CF, Zhu WY, Williams BA, Klüß J, Souffrant W, et al. Microbial diversity studies of the porcine gastrointestinal ecosystem during weaning transition. Anim Res. 2004;53: 317–324. [Google Scholar]
- 112.Topping DL, Clifton PM. Short-chain fatty acids and human colonic function: roles of resistant starch and nonstarch polysaccharides. Physiol Rev. 2001;81: 1031–1064. 10.1152/physrev.2001.81.3.1031 [DOI] [PubMed] [Google Scholar]
- 113.Fang CL, Sun H, Wu J, Niu HH, Feng J. Effects of sodium butyrate on growth performance, haematological and immunological characteristics of weanling piglets. J Anim Physiol Anim Nutr (Berl). 2014;98: 680–685. [DOI] [PubMed] [Google Scholar]
- 114.Jiang WD, Tang RJ, Liu Y, Kuang SY, Jiang J, Wu P, et al. Manganese deficiency or excess caused the depression of intestinal immunity, induction of inflammation and dysfunction of the intestinal physical barrier, as regulated by NF-kappaB, TOR and Nrf2 signalling, in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015;46: 406–416. 10.1016/j.fsi.2015.06.007 [DOI] [PubMed] [Google Scholar]
- 115.Nebot-Vivinus M, Harkat C, Bzioueche H, Cartier C, Plichon-Dainese R, Moussa L, et al. Multispecies probiotic protects gut barrier function in experimental models. World J Gastroenterol. 2014;20: 6832–6843. 10.3748/wjg.v20.i22.6832 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the manuscript and figures.