

Abstract
Hybridization is a biological phenomenon increasingly recognized as an important evolutionary process in both plants and animals, as it is linked to speciation, radiation, extinction, range expansion and invasion, and allows for increased trait diversity in agricultural and horticultural systems. Estimates of hybridization frequency vary across taxonomic groups, but causes of this variation are unknown. Here, we ask on a global scale whether hybridization is linked to any of 11 traits related to plant life history, reproduction, genetic predisposition, and environment or opportunity. Given that hybridization is not evenly distributed across the plant tree of life, we use phylogenetic generalized least squares regression models and phylogenetic path analysis to detect statistical associations between hybridization and plant traits at both the family and genus levels. We find that perenniality and woodiness are each weakly associated with an increased frequency of hybridization in univariate analyses, but path analysis suggests that the direct linkage is between perenniality and increased hybridization (with woodiness having only an indirect relationship with hybridization via perenniality). Weak associations between higher rates of hybridization and higher outcrossing rates, abiotic pollination syndromes, vegetative reproductive modes, larger genomes, and less variable genome sizes are detectable in some cases but not others. We argue that correlational evidence at the global scale, such as that presented here, provides a robust framework for forming hypotheses to examine and test drivers of hybridization at a more mechanistic level.
Keywords: Hybridization, introgression, phylogenetically independent contrasts, vascular plant hybrids
Impact Summary.
Although historically thought of as rare, interspecific mating is increasingly recognized as an important evolutionary process. Hybridization can generate increased genetic and morphological variation and has been tied to increased diversification and other biological phenomena such as geographic range expansion and the success of invasive species. Here, we examine hybridization of plants on a global scale. Previous work has demonstrated that some plant groups hybridize more than others, but the reasons for this pattern remain unclear. We combine data from eight regional floras with trait data to test for associations between hybridization and different aspects of plant biology, such as life history, growth form, reproduction, and opportunity, all while accounting for the fact that plant lineages are related to each other.
We find that plant groups that are dominated by perennial species and species with woody growth forms tend to hybridize more than those dominated by annual or herbaceous species. We also find weak evidence that frequent hybridization is found in plant families that are predominantly pollinated abiotically (such as by wind or water) or have higher rates of outcrossing, plant genera that have less variable genome sizes, and plant groups (both genera and families) that can reproduce asexually and have larger genome sizes. This study provides the first analysis of the global correlates of hybridization in plants. Although this correlational evidence does not provide any mechanistic explanations for these patterns, the trends we find are novel in terms of both geographic and taxonomic scale. The correlations detected provide robust hypotheses for understanding the conditions for hybridization and its contributions to evolution.
Hybridization is increasingly recognized as an important evolutionary phenomenon in plants (Mallet 2005; Arnold 2006; Whitney et al. 2010), animals (Mallet 2005; Schwenk et al. 2008), and fungi (reviewed in Albertin and Marullo 2012). Hybridization has been linked to important processes such as evolution and diversification (Anderson and Stebbins 1954; Seehausen 2004), adaptive radiation (Anderson and Stebbins 1954; Stebbins 1959; Barton 2001; Seehausen 2004; Yakimowski and Rieseberg 2014; Marques et al. 2019), and speciation (Rieseberg 2003; Mallet 2007; Rieseberg et al. 2007; Soltis and Soltis 2009; Abbott et al. 2013). Hybridization has enabled plant breeders to transfer desirable traits among species for both agricultural and horticultural purposes (Allard 1999). In contrast, hybridization has also been linked to numerous conservation concerns such as biological invasion (Ellstrand and Schierenbeck 2000; Schierenbeck and Ellstrand 2009; Whitney et al. 2010; Hovick et al. 2012; Hovick and Whitney 2014), escape of novel traits via crop‐wild hybridization (Ellstrand and Hoffman 1990; Zapiola et al. 2008), and even extinction via hybridization (Rhymer and Simberloff 1996; Wolf et al. 2010; Todesco et al. 2016, Campbell et al. In press). A deep understanding of hybridization is thus necessary to understand evolutionary principles, to provide for agricultural needs, and to inform conservation management decisions.
There is evidence for hybridization in unexpected situations, for instance between distantly related species (Rothfels et al. 2015) or in cases of cryptic hybridization with molecular but little morphological evidence (Cronn and Wendel 2004; Soltis et al. 2007; McIntosh et al. 2014; Mitchell and Holsinger 2018). Focke (1881, in Stebbins 1959 and Grant 1981) first made the observation that rates of hybridization differ across plant taxa. More modern analyses based on floras or surveys of the literature have found different rates of hybridization in different taxonomic groups, with evidence for phylogenetic signal (Ellstrand et al. 1996; Whitney et al. 2010; Abbott 2017; Beddows and Rose 2018). Ferns and their allies and specific flowering plant families (such as Orchidaceae, Lamiaceae, Asparagaceae, and Asteraceae) contain high numbers of hybridizing species, while other families appear to contain few hybrids (such as Caryophyllaceae, Cyperaceae, and Apiaceae) (Whitney et al. 2010).
Hypotheses as to why some groups hybridize more than others center on traits related to life history, reproduction, genetics, and opportunity or environment. Researchers have either advanced theoretical reasons for a connection between a trait and increased hybridization, or have identified correlational evidence to support a connection without a theoretical justification (summarized in Table 1 and expanded on in Table S1). These traits may be associated with the formation of hybrids, i.e. allowing for interspecific mating and production of offspring, or may be associated with the persistence of hybrids, i.e. allowing for the continued propagation of a hybrid lineage after formation. Briefly, we expected that plant groups dominated by perennial species (Grant, 1958, 1981; Stace 1975; Ellstrand et al. 1996; Beddows and Rose 2018) or woody species (Stebbins 1959; Beddows and Rose 2018) will contain more hybrids than those dominated by annual or herbaceous life histories, because longer lifespans associated with perenniality and woodiness may allow hybrid individuals to produce offspring over time despite partial sterility, allowing for persistence of these hybrid lineages (Ellstrand et al. 1996). We also expected higher rates of hybridization in plant groups with traits that increase the likelihood of interspecific mating, either by reducing barriers to gene flow or promoting outbreeding. These include traits such as pollination syndrome (contrasting evidence for increased hybridization with both biotic: Rieseberg and Wendel 1993, or abiotic: Ellstrand et al. 1996, pollination syndromes), bilaterally symmetrical flowers (Stebbins 1959; Sargent 2004), reproductive systems that require cross‐breeding (higher outcrossing rates: Stace 1975; Grant 1981), sexual breeding systems (Grant 1981), and generative/nonvegetative reproductive systems (Ellstrand et al. 1996). Some groups may be genetically predisposed to hybridize, for instance lineages with few chromosomal translocations that allow for greater fertility in hybrids (Grant 1981), smaller genome sizes (as reported in Bureš et al. 2004), or less variable genome sizes that may allow for greater interspecific compatibility. Finally, hybridization may be the product of opportunity, where greater opportunity might be conferred via having agricultural relatives that by nature are abundant and widespread, being less threatened by extinction, or being found in more disturbed environments where contact with relatives might be initiated (Anderson and Stebbins 1954; Grant 1981; Guo 2014). Given the large number of the above‐described potential drivers of hybridization, and because hybridization is an emergent phenomenon rather than a simple morphological or physiological trait, we expect a priori that any one correlate may have little explanatory power.
Table 1.
A review of the potential traits associated with hybridization in plants, as identified by a literature search, with further information on data types and sources in our analysis
| Category | Trait | Prediction | Prediction type | Data description | Data source |
|---|---|---|---|---|---|
| Life history | Perenniality | + | Empirical1–5 Theoretical2 | Mean score (0 = annual, 0.5 = annual/biennial/perennial, 1 = perennial) | Floras |
| Woodiness | + | Empirical5 , 6 | Mean score (0 = herbaceous, 0.5 = either, 1 = woody) | Floras | |
| Reproductive | Pollination syndrome | ± | Empirical4 , 7 | Mean score (0 = abiotic, 0.5 = both, 1 = biotic) | TRY (Kattge et al. 2011) |
| Floral symmetry | + | Theoretical6 , 8 | Mean score (0 = actinomorphic, 0.5 = both, 1 = zygomorphic) | TRY (Kattge et al. 2011) | |
| Outcrossing | + | Empirical2 , 4 Theoretical1–3 | Mean outcrossing rate (t) | Goodwillie et al. 2005; Moeller et al. 2017 | |
| Breeding system | + | Theoretical2 | Mean score (0 = asexual, 0.5 = both, 1 = sexual) | TRY (Kattge et al. 2011) | |
| Reproductive system | – | Empirical4 Theoretical1 | Mean score (0 = vegetative, 0.5 = both, 1 = generative) | TRY (Kattge et al. 2011) | |
| Genetic predisposition | C‐value | – | Empirical9 | Mean C‐value (genome size in picogram) | Bennett and Leitch (2005) |
| C.V. C‐value | ± | Theoretical | Mean coefficient of variation of C‐value | Bennett and Leitch (2005) | |
| Chromosomal translocations | – | Theoretical2 | Not analyzed | ||
| Genetic divergence | ± | Empirical10 , 11 Reviewed12 , 13 | Not analyzed | ||
| Opportunity/environment | Agricultural status | + | Theoretical14 | Mean score (0 = noncrop species, 1 = crop species) | SINGER |
| Red List | – | Theoretical15 | Mean score (0 = LC, 0.5 = NT, LR/nt, 1 = LR/cd, 2 = VU, 3 = EN, 4 = CR, 5 = EX, EW) | Baillie et al. (2004) |
The “Prediction” column gives the predicted sign of the association between the trait and hybridization propensity, relative to the orientation in the “Data Description” column. “Prediction Type” distinguishes whether predictions from the literature are based on a theoretical argument or simply on an observed (but not phylogenetically corrected) empirical association. We expand on proposed mechanisms in Table S1. Data used in analyses were mean scores across all species within the group of interest (family or genus). When we did not have data to test the potential relationship, the “Data Source” column is blank. Descriptions of traits, how they were scored for this study, predictions (empirical or theoretical) from the literature (see superscripts for sources), and sources for the data used in this study.
1Grant (1958); 2Grant (1981); 3Stace (1975); 4Ellstrand et al. (1996); 5Beddows and Rose (2018); 6Stebbins (1959); 7Rieseberg and Wendel (1993); 8Sargent (2004); 9Bureš et al. (2004); 10Paun et al. (2009); 11Stelkens and Seehausen (2009); 12Mallet (2005); 13Mavarez and Linares (2008); 14Allard (1999); 15Allendorf et al. (2001).
LC, least concern; NT, near threatened; LR/nt, lower risk/near threatened; LR/cd, lower risk/conservation dependent; VU, vulnerable; EN, endangered; CR, critically endangered; EX, extinct; EW, extinct in the wild.
At the regional scale, measures of hybridization have been empirically linked to various plant attributes. Beddows and Rose (2018) performed a case study on the flora of Michigan, a single state in the United States. They surveyed the published flora for interspecific hybrids and several plant attributes, including life history and life form, and used multiple logistic regressions to determine what factors were correlated with various measures of hybridization. Although taxonomic order was included in the analysis, they did not explicitly account for the phylogenetic nonindependence of the taxa analyzed. In their analysis, hybridization was positively correlated with perenniality, woodiness, habitat disturbance, and number of herbarium records, and they additionally detected significant effects of taxonomic order (Beddows and Rose 2018).
Thus far, there has been no comprehensive analysis of the potential correlates of hybridization in plants at the global scale, nor has there been an analysis accounting for phylogenetic nonindependence among taxa. Here, we build on the work of Whitney et al. (2010), which quantified hybridization across the globe in 282 different plant families and 3212 genera using data from eight regional floras. We expanded this dataset and combined it with trait data collected from the regional floras and additional external datasets to ask whether hybridization in plants (quantified using two metrics) is statistically associated with 11 different traits at both the family and genus levels, while simultaneously accounting for the phylogenetic nonindependence of the taxa analyzed.
Methods
EXTENT OF HYBRIDIZATION
To characterize the extent of hybridization across vascular plant families, we analyzed eight floras: the Great Plains of the United States (McGregor and Barkley 1986); the British Isles (Stace 1997); Hawai'i (Wagner et al. 1999); the Intermountain Region of the western United States (Cronquist et al. 1972); the Northeastern United States (Magee and Ahles 1999); California (Hickman 1993); Europe (Tutin et al. 1964); and Victoria, Australia (Walsh and Entwisle 1994) (Fig. 1). These floras are the same as those used in Whitney et al. (2010), with the exception that we have here included the final published volume of the Intermountain Region (volume 2A, 2012). Floras were chosen nonrandomly to include those that contained multiple mentions of hybrids, and are therefore a biased subset reflecting regions where hybrids are common or, more likely, reflecting authors interested in hybridization and attuned to recording instances of it.
Figure 1.
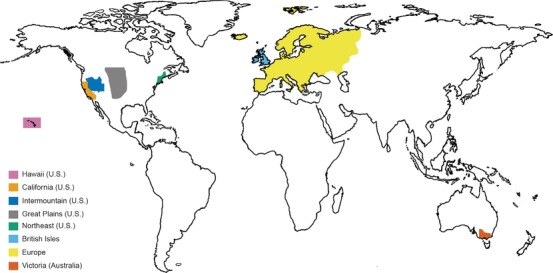
World map indicating the coverage areas of the floras used in this study. Pink = Hawaii (USA), light orange = California (USA), dark blue = Intermountain (USA), gray = Great Plains (USA), green = Northeast (USA), light blue = British Isles, yellow = Europe, dark orange = Victoria (Australia).
For each vascular plant family in each flora, the numbers of interspecific hybrids and the numbers of nonhybrid species were determined as in Whitney et al. (2010). For counting purposes, we follow Ellstrand et al. (1996) in defining a “hybrid” as a hybrid type derived from a unique combination of two parental species. Thus, in each flora, each pair of hybridizing species was counted as generating a single hybrid, even if there was evidence that the pair had hybridized multiple times. Our recognition of an interspecific hybrid does not imply that it was formally or taxonomically recognized in the flora (though some were), nor does it imply processes such as hybrid speciation or the formation of a hybrid population that is stable over the long term. It simply is an observation that a pair of parental species has interbred and resulted in hybrid offspring that have persisted in the wild long enough to be noted by an author of a flora. Hybrids may be unnamed or denoted with an “ × ” (Turland et al. 2018), a species name, or a subspecies name, but the naming process is idiosyncratic with respect to degree of reproductive isolation and time since hybrid formation. Therefore, we do not attempt to distinguish among different types or ages of hybrids, instead using the previous practice of considering all hybrid types together (see Ellstrand et al. 1996). We assume that the traits of interest (e.g., woodiness and perenniality) are likely conserved across the time scale that any hybrid formation occurs, and thus that associations between trait values and even well‐established, reproductively isolated hybrids (e.g., allopolyploid species) are informative in our analyses.
Only native and naturalized taxa were considered. Taxa clearly resulting from anthropogenic crosses (e.g., “garden hybrids”) and taxa only in cultivation were ignored. We tallied intra‐ and intergeneric hybrids separately, and the latter were split between genera (e.g., half of each hybrid was assigned to each contributing genus). We did not count hybrids among subspecies or probable primary intergradation (diverging subpopulations maintaining genetic connections, Stebbins 1959). We counted naturalized hybrids mentioned in a flora that apparently arose outside the region covered by the flora. Finally, in some floras, particular groups were described as producing multiple hybrids without detailed specification of their numbers or the parental species involved. In these few cases, we estimated the number of hybrids as either 2 hybrids or 20% of the number of species present, whichever was greater. We analyzed all floras at the generic level and reassigned those genera (with their associated counts of species and hybrids) to families based on The Plant List (http://www.theplantlist.org/) to accommodate taxonomic changes since the publication of the floras.
We collected hybridization data on 282 plant families and 3229 different genera. Observations of genera with a single nonhybrid species identified in a single flora were then eliminated to avoid including groups with no chance for hybridization, and a single family that could not be placed phylogenetically with confidence (Capparaceae, see below) was also excluded. This resulted in a final sample size of 195 families for the family‐level analysis. For the genus‐level analysis, we were unable to place 34 genera in the phylogeny (see below), resulting in a final sample size of 1772 genera (Table S2).
For completeness and comparability, we characterized hybridization for each family or group using two metrics: hybridization propensity and hybrid ratio. Hybridization propensity reflects the realized percentage of all possible hybrid combinations and is calculated as in Whitney et al. (2010). For a taxonomic group (genus) of n nonhybrid species:
| (1) |
Although it is unrealistic that every pair of species within a group hybridizes (so the denominator of eq. (1) is perhaps unrealistically large), incorporating bounds on the percentage of species that could potentially hybridize would require additional information beyond the scope of this study. Hybrid ratio, employed by Beddows and Rose (2018), reflects the number of hybrid combinations relative to all nonhybrid taxa. N is used as the denominator as a correction for opportunity, as the presence of more nonhybrid species means more opportunities to form both intra‐ and intergeneric hybrids. For a taxonomic group (genus) of n nonhybrid species:
| (2) |
We calculated and analyzed both to enable comparisons of our findings to previous studies. Note the scale difference: by convention, hybridization propensity is a percentage bounded between 0 and 100, while hybrid ratio is unbounded (in practice, it ranges from 0 to 0.15 with outliers up to 1.2). For each genus, numbers of both nonhybrids and hybrids were calculated by summing hybrid counts across all floras analyzed. No attempt was made to avoid “double counting” of hybrids formed from the same parents in different regions. Thus, each metric incorporates information on both the number of hybridizing taxa and the frequency with which they hybridize in different regions. Genus‐level metrics were calculated based on the observations across all floras, while family‐level metrics were weighted means of metrics of the component genera (weighted by species number in each genus). Both hybridization propensity and hybrid ratio measures were log‐transformed prior to analysis to more closely match assumptions of normality.
TRAITS OF PLANT GROUPS
The number of annual, biennial, and perennial species, and the number of herbaceous versus woody species were summed for each genus in each flora. The floras provided remarkably complete data on these variables (>95% species covered), but missing data on perenniality and woodiness of the species were determined from other sources (e.g., USDA plants database). Species described as intermediate (e.g., “annual/biennial”) were split between categories (e.g., counted as 0.5 annuals and 0.5 biennials). Species were considered woody if they were characterized by substantial aboveground woody biomass, for example, “trees,” “shrubs,” “subshrubs,” “woody vines,” and “lianas.” Species with rootstocks as the only woody parts were considered herbaceous. For genus and family‐level analyses, we used the percentage of species scored as perennial and the percentage of species scored as perennial as our trait data (Table 1).
Data for several traits were downloaded from the TRY database (Kattge et al. 2011). These included information on pollination syndrome (abiotic or biotic: Fitter and Peat 1994; Koike 2001; Ogaya and Peñuelas 2003; Diaz et al. 2004; Kühn et al. 2004; Gachet et al. 2005; Moretti and Legg 2009; Onstein et al. 2014; de Frutos et al. 2015; Giroldo 2016; S. Chapin, unpubl. data; M. Leishman, unpubl. data), breeding system (asexual or sexual: Kühn et al. 2004), floral symmetry (actinomorphic or zygomorphic: Dressler et al. 2014), and reproductive system (vegetative or generative: Fitter and Peat 1994; Kühn et al. 2004; Klimešová and de Bello 2009). For each species in the TRY dataset, trait values were simplified to be either 0, 0.5 (for mixed or combined), or 1 (see Table 1 for coding schemes for individual traits). We used genus or family‐level means for pollination syndrome, breeding system, floral symmetry, and reproductive system as trait data in subsequent analyses.
We compiled additional trait data from other sources. We assessed agricultural status by calculating the percentage of species in each family that were listed as crop species as defined in the System‐wide Information Network for Genetic Resources database (http://singer.cgiar.org/Search/SINGER/search.htm , downloaded July 2009). We assessed threatened status using data from the Red List (Baillie et al. 2004). We assigned numeric values representing each species’ threatened status (see Table 1 for scoring categories) and used genus‐ or family‐level means. We estimated genus‐ and family‐level mean outcrossing rates from Goodwillie et al. (2005) and Moeller et al. (2017). Finally, genome size estimates (both “Prime Estimates” and others) were downloaded from the Plant DNA C‐values database (Bennett and Leitch 2005). We calculated the mean genome size per species (including all ploidy level variants, if present in the database) and then calculated genus and family‐level means. C‐value was log‐transformed prior to analysis. We also estimated the coefficient of variation for genome size by calculating mean C‐values for each ploidy level of each species, then calculating the coefficient of variation across these means for each genus and family levels. See Table 1 for full information on the traits assessed.
COMPOSITE TREE CONSTRUCTION AND PHYLOGENETIC SIGNAL
Subsequent analyses were conducted in R version 3.3.3 (R Core Development Team 2016). To account for the phylogenetic nonindependence of our observations, we used phylogenetic generalized least squares regression (PGLS regression: Grafen 1989; Martins and Hansen 1997). The family‐level seed plant phylogeny was imported from the tree of Qian and Jin (2016) (an updated and corrected version of Zanne et al. 2014) into R using the “ape” package (Paradis et al. 2004). The phylogeny was trimmed and resolved to include only the seed plant families for which we had data using the S.Phylomaker function from Qian and Jin (2016). To include nonseed plants, we manually constructed phylogenies in Mesquite version 3.40 (Maddison and Maddison 2018) based on their position in the literature for ferns (Smith et al. 2006) and fern allies (Pryer et al. 2004) and combined them in R. To construct a genus‐level phylogeny, we used S.Phylomaker and added within‐family relationships for the ferns and their allies by hand based on the literature (Hauk et al. 2003; Pryer et al. 2004; Schneider et al. 2004a; Schneider et al. 2004b; Ebihara et al. 2006; Liu et al. 2007; He and Zhang 2012; Sundue et al. 2014; de Gasper et al. 2017).
We estimated phylogenetic signal via Pagel's λ separately for each measure of hybridization and each trait using the phylopars() function with model set to “lambda” in the “Rphylopars” package (Goolsby et al. 2017). We compared this model to a star phylogeny with λ = 0 using likelihood ratio tests. Although the “Rphylopars” package allows imputation of missing trait values (Bruggeman et al. 2009; Goolsby et al. 2017), we had high amounts of missing data (for a given trait, up to 61% in families and 89% in genera) so chose not to impute and instead pruned trees to exclude taxa with missing data before each analysis.
ANALYSES OF HYBRIDIZATION VERSUS POTENTIAL CORRELATES
We calculated the raw correlations between all 11 traits and the two hybridization metrics at both the family and generic levels using the corr.test() function in the R package “psych” (Revelle 2017). However, raw correlations do not account for phylogenetic nonindependence among taxa (Felsenstein 1985), so we report these only for frame of reference.
PGLS regression provides a flexible framework for detecting associations among traits under different evolutionary models (Grafen 1989; Martins and Hansen 1997). PGLS was conducted using the phylopars.lm() function in the R package “Rphylopars” (Goolsby et al. 2017). We performed univariate PGLS regressions for each of our traits on both metrics of hybridization at the family and generic levels, subsetting the data and phylogenies to prevent imputation (see above for explanation). Note that we were missing values for some traits due to lack of available data and for other traits because they were not applicable to all taxonomic groups (e.g., only seed plants have pollination syndromes, and only flowering plants have floral symmetry). Regressions were performed under the Brownian motion (BM), Ornstein–Uhlenbeck (OU), and early burst (EB) models of evolution, and then compared using AIC and BIC. As either BM or EB was the best model across all traits, and as all models were within 2 AIC, we report BM results as representative. We corrected for multiple comparisons using the Benjamini–Hochberg procedure (Benjamini and Hochberg 1995) within each hybridization measure and taxonomic level combination (11 total tests per combination) using a false discovery rate of 0.05.
PHYLOGENETIC PATH ANALYSIS
A potential multivariate analysis including all 11 traits as predictors of hybridization was not practical, because of missing trait data. However, we did have nearly complete information for woodiness and perenniality. To simultaneously estimate the relationships between hybridization and both perenniality and woodiness, we used the “phylopath” package (van der Bijl 2018) to run phylogenetic path analyses. Although causal relationships cannot be determined from correlational evidence, path analysis allows for an understanding of direct and indirect relationships under proposed causal models (von Hardenberg and Gonzalez‐Voyer 2013; Kennedy et al. 2018). We used these models to determine the relative strength of these two highly correlated predictors of hybridization when present in the same model. We tested five path structures for each combination of taxonomic level and measure of hybridization (Fig. S1). The fit of models was estimated using the C statistic, which provides an estimate of goodness of fit of the model to the data (Shipley 2013). We report results from the best model using CICc, the C statistic information criterion (von Hardenberg and Gonzalez‐Voyer 2013).
Results
HYBRIDIZATION METRICS AND PHYLOGENETIC SIGNAL
In the 195 plant families analyzed, 112 contained hybrids and 83 did not. The mean value for family‐level hybridization propensity was 2.55% (range = 0–100%) and for hybrid ratio was 0.086 (range = 0–1.196) (Fig. 2, Table S3). At the family level, the log‐transformed values for hybridization propensity and hybrid ratio were significantly correlated (corr = 0.701, P < 0.001) (Table S4). There was significant phylogenetic signal in hybridization propensity (λ = 0.30, P < 0.001) and a lower, but still significant, measure of phylogenetic signal in hybrid ratio (λ = 0.14, P < 0.01) (Table 2). Eight out of 11 traits had significant phylogenetic signal at the family level (perenniality, woodiness, percent agricultural, floral symmetry, pollination syndrome, reproductive system, C‐value, and coefficient of variation in C‐value; see Table 2).
Figure 2.
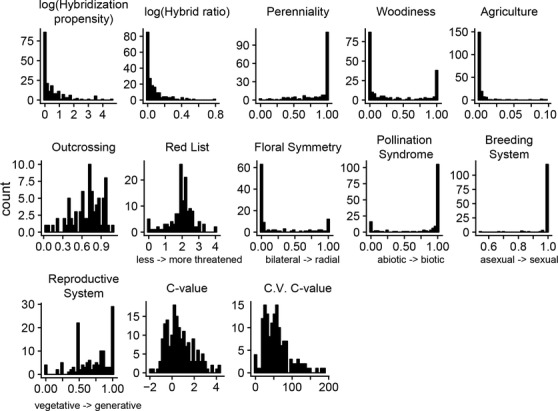
Distributions of family‐level hybridization metrics and family‐average traits. See Table 1 for trait descriptions and units. Nonintuitive trait values have brief descriptions on the x‐axes.
Table 2.
Phylogenetic signal (Pagel's λ and associated chi‐square statistics and P‐values) of hybridization measures and potential predictors at different taxonomic levels
| Family level | Genus level | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trait | N observed | Pagel's λ | Chi‐Square | DF | P‐value | N observed | Pagel's λ | Chi‐Square | DF | P‐value |
| Hybridization propensity | 195 | 0.30 | 32.31 | 1 | 0.000 | 1772 | 0.11 | 52.28 | 1 | 0.000 |
| Hybrid ratio | 195 | 0.14 | 8.06 | 1 | 0.005 | 1772 | 0.13 | 52.05 | 1 | 0.000 |
| Perenniality | 195 | 0.22 | 10.34 | 1 | 0.001 | 1754 | 0.47 | 314.01 | 1 | 0.000 |
| Woodiness | 195 | 0.47 | 40.87 | 1 | 0.000 | 1767 | 0.80 | 968.73 | 1 | 0.000 |
| Percent agricultural | 195 | 0.26 | 3.90 | 1 | 0.048 | 1772 | 1.00 | 6738.41 | 1 | 0.000 |
| Outcrossing | 76 | 0.01 | 0.01 | 1 | 0.943 | 158 | 0.24 | 3.72 | 1 | 0.054 |
| Red List | 138 | 0.00 | ‐0.01 | 1 | 1.000 | 374 | 0.25 | 21.45 | 1 | 0.000 |
| Floral symmetry | 114 | 0.51 | 13.33 | 1 | 0.000 | 235 | 0.76 | 124.51 | 1 | 0.000 |
| Pollination syndrome | 164 | 0.79 | 70.89 | 1 | 0.000 | 878 | 0.93 | 1208.71 | 1 | 0.000 |
| Breeding system | 130 | 0.03 | 0.17 | 1 | 0.678 | 639 | 0.09 | 8.87 | 1 | 0.003 |
| Reproductive system | 133 | 0.32 | 18.48 | 1 | 0.000 | 655 | 0.46 | 135.09 | 1 | 0.000 |
| C‐value | 177 | 0.66 | 57.11 | 1 | 0.000 | 761 | 0.74 | 476.77 | 1 | 0.000 |
| C.V. C‐value | 144 | 0.37 | 7.04 | 1 | 0.008 | 522 | 0.00 | –0.00 | 1 | 1.000 |
We analyzed 1772 different plant genera, of which 492 contained hybrids and 1280 did not. The mean value for genus‐level hybridization propensity was 2.885% (range = 0–300%) and for hybrid ratio was 0.060 (range = 0–1.609) (Table S3). At the genus level, the log‐transformed values for hybridization propensity and hybrid ratio were significantly correlated (corr = 0.846, P < 0.001) (Table S4). We also detected low but significant phylogenetic signal in hybridization propensity (λ = 0.11, P < 0.001) and hybrid ratio (λ = 0.13, P < 0.001) at the genus level (Table 2). Nine out of 11 traits had significant phylogenetic signal at the genus level (all but outcrossing and the coefficient of variation of C‐value; see Table 2).
PLANT TRAITS
We assessed 11 potential correlates of hybridization using data from the floras as well as other sources (Table 1). The dataset was dominated by perennial and herbaceous taxa as well as by taxa with radially symmetric flowers, biotic pollination syndromes, sexual breeding systems, and generative reproductive systems (Fig. 2, Table S3).
CORRELATES OF HYBRIDIZATION
Using univariate regressions at the family level, we detected significant associations (P < 0.05) linking abiotic pollination syndrome to increased hybridization propensity and a trend (0.05 < P < 0.10) for links between both higher outcrossing rates and larger genome sizes and hybridization propensity (Fig. 3, Table 3). We detected associations between perenniality, woodiness, and more abiotic pollination syndromes with hybrid ratio, although only the latter was significant (Fig. 3, Table 3). However, after correcting for multiple comparisons, none of these family level associations were significant. Adjusted R 2 values were very low, with a maximum of 0.034.
Figure 3.
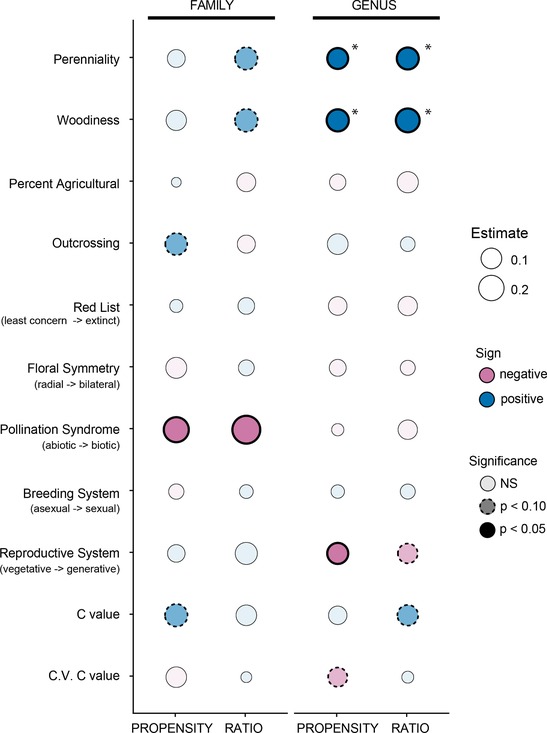
Predictors of hybridization propensity and hybrid ratio at the family (left) and genus (right) levels from PGLS univariate regressions. Sizes of the circles indicate the absolute value of the strength of the estimate. Color indicates the sign (positive = blue, negative = pink) of the estimate. The transparency and border indicate the significance of the estimate: lightest shaded circles were not significant (P > 0.10), medium shading with dashed borders indicates a trend (P < 0.10), and darkest shading with solid bold borders indicates statistical significance (P < 0.05). Asterisks indicate that the relationship is significant after a Benjamini–Hochberg procedure.
Table 3.
Univariate PGLS results at different taxonomic levels
| Family level | ||||||
|---|---|---|---|---|---|---|
| Hybridization propensity | Hybrid ratio | |||||
| Trait | Estimate | P‐value | Adjusted‐R 2 | Estimate | P‐value | Adjusted‐R 2 |
| Perenniality | 0.057 | 0.391 | –0.001 | 0.135 | 0.061 | 0.013 |
| Woodiness | 0.093 | 0.206 | 0.003 | 0.141 | 0.077 | 0.011 |
| Percent agricultural | 0.004 | 0.951 | –0.005 | –0.072 | 0.322 | 0.000 |
| Outcrossing | 0.125 | 0.083 | 0.027 | –0.060 | 0.498 | –0.007 |
| Red List | 0.013 | 0.857 | –0.007 | 0.043 | 0.606 | –0.005 |
| Floral symmetry | –0.105 | 0.167 | 0.008 | 0.034 | 0.744 | –0.008 |
| Pollination syndrome | –0.191 | 0.019 | 0.028 | –0.267 | 0.010 | 0.034 |
| Breeding system | –0.029 | 0.610 | –0.006 | 0.017 | 0.817 | –0.007 |
| Reproductive system | 0.053 | 0.392 | –0.002 | 0.127 | 0.106 | 0.012 |
| C‐value | 0.136 | 0.084 | 0.011 | 0.099 | 0.288 | 0.001 |
| C.V. C‐value | –0.099 | 0.183 | –0.006 | 0.005 | 0.958 | –0.007 |
| Genus level | ||||||
|---|---|---|---|---|---|---|
| Perenniality | 0.103 | 0.000 * | 0.007 | 0.123 | 0.000 * | 0.011 |
| Woodiness | 0.126 | 0.000 * | 0.007 | 0.161 | 0.000 * | 0.011 |
| Percent agricultural | –0.039 | 0.840 | –0.001 | –0.109 | 0.568 | 0.000 |
| Outcrossing | 0.101 | 0.171 | 0.006 | 0.024 | 0.752 | –0.006 |
| Red List | –0.067 | 0.295 | 0.000 | –0.078 | 0.248 | 0.001 |
| Floral Symmetry | –0.045 | 0.515 | –0.002 | –0.026 | 0.698 | –0.004 |
| Pollination syndrome | –0.009 | 0.904 | –0.001 | –0.079 | 0.314 | 0.000 |
| Breeding system | 0.015 | 0.671 | –0.001 | 0.026 | 0.515 | –0.001 |
| Reproductive system | –0.106 | 0.011 * | 0.008 | –0.085 | 0.074 | 0.003 |
| C‐value | 0.065 | 0.226 | 0.001 | 0.101 | 0.081 | 0.003 |
| C.V. C‐value | –0.077 | 0.064 | 0.005 | 0.008 | 0.871 | –0.002 |
*Relationships significant after Benjamini–Hochberg procedure; raw P‐values < 0.05 are in bold.
At the genus level, increased perenniality and woodiness were associated with increased hybridization in both metrics. These relationships were still significant after a Benjamini–Hochberg correction (Table 3). There was a slight association (0.05 < P < 0.10) between less variable genome sizes and increased hybridization propensity and a significant association (after correcting for multiple comparisons) between more vegetative reproductive systems and hybridization propensity. There were trends for genera with more vegetative reproductive systems and larger genome sizes to have higher values of hybrid ratio (Fig. 3, Table 3). Adjusted R 2 values were also very low, with a maximum of 0.011. Family‐ and genus‐level relationships were generally in consensus, in that there were no instances where a well‐supported association at one taxonomic level was well supported in the opposite direction at the other taxonomic level (Fig. 3, Table 3).
PHYLOGENETIC PATH ANALYSIS
To account for the high correlations among two traits with detectable associations with hybridization in the univariate regressions), we examined relationships between hybridization and both perenniality and woodiness using phylogenetic path analyses (Fig. S1). At both the family and genus levels, the best models indicate that woodiness does not have a direct link to hybridization, but instead has an indirect association via a pathway including perenniality and perenniality's direct association with hybridization (Fig. 4, Table S5). The estimated path coefficients were all positive and above 0 ± standard error (Table S6).
Figure 4.
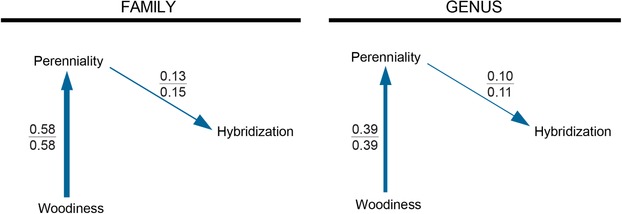
Path coefficients associated with hybridization propensity (top value) and hybrid ratio (bottom value) at the family (left) and genus (right) levels from phylogenetic path analysis using two predictors with large sample sizes that are also highly correlated: perenniality and woodiness. Final models were chosen via CICc from five candidate models (see Fig. S1). Widths of the arrows approximately indicate the strength of the coefficient and the direction of the relationship. A lack of an arrow indicates that a relationship was not included in the best model.
RAW CORRELATIONS
For comparative purposes, we present raw correlations in the Supporting Information table. Several relationships between traits and hybridization rate or propensity were detected in the raw analyses that were not detected in the phylogenetically corrected analyses, emphasizing the importance of examining these relationships in a phylogenetic context (Table S4).
Discussion
Hybridization is not evenly distributed across the phylogenetic tree of life (Ellstrand et al. 1996), nor is it evenly distributed within plants, as we have documented here and elsewhere (Whitney et al. 2010). We detected several associations between hybridization rates and plant traits (perenniality, woodiness, outcrossing rate, pollination syndrome, reproductive system, genome size, and genome size variation) across the globe. Below, we organize our discussion of these associations sequentially, first discussing traits that may allow the formation of hybrids, followed by traits that may allow for the persistence of hybrids.
CORRELATES OF HYBRIDIZATION: FACTORS THAT MAY ALLOW FOR HYBRID FORMATION
Lineages may have detectable associations with specific factors that allow for the more frequent formation of hybrids. These associations may be direct or indirect in nature. For example, there may be a direct association between outcrossing and high levels of hybridization. High levels of outcrossing (or obligate outcrossing, as an extreme) mean that plants need to reproduce with another individual, necessitating the transfer of pollen, and increasing the odds of contacting and reproducing with another species when compared to selfing (Stace 1975; Ellstrand et al. 1996). Supporting this idea, we detected a trend for a positive association at the family level between outcrossing rate and hybridization propensity (Fig. 3, Table 3).
Other factors may be indirectly associated with hybridization. Grant (1958) hypothesized that associations between perenniality/woodiness and increased hybridization rates were actually indirect associations via outcrossing. He observed that perennial outcrossers were the most likely category of plants to participate in interspecific breeding and that autogamous or selfing plants were the least likely. We found associations between hybridization metrics and both woodiness and perenniality (Fig. 3, Table 3), and these traits were also correlated with outcrossing (Table S4). Our findings match previous hypotheses and nonphylogenetically corrected associations between hybridization and woodiness and/or perenniality (Stebbins 1959; Stace 1975; Ellstrand et al. 1996; Beddows and Rose 2018). In our analyses, the links between perenniality/woodiness and our hybridization measures were stronger than links with outcrossing rate (which had only a moderate association with hybridization propensity across families), but this discrepancy may be due to the restricted number of taxa for which we had outcrossing rate data (outcrossing data for 76 families and 158 genera, compared with perenniality and woodiness data for 195 families and 1754 and 1767 genera, respectively, Table 2 and Table S3). Perenniality and woodiness are positively correlated in plants; our evidence suggests that perenniality may be driving the association with hybridization, as there was more evidence for models including a direct path from perenniality to hybridization than a direct path from woodiness to hybridization (Fig. 4, Tables S5 and S6).
Factors not associated with outcrossing directly may also increase the chances of mating with heterospecifics and forming hybrids. Abiotic pollination syndromes may reduce prezygotic barriers to reproduction by allowing for promiscuous transfer of pollen, independent of biotic vectors. We found associations between abiotic pollination and hybridization at the family level, but not the genus level (Fig. 3, Table 3). We believe this is the first empirical dataset used to explicitly test for this association while correcting for phylogenetic nonindependence (see Rieseberg and Wendel 1993; Ellstrand et al. 1996 for raw correlations, in both directions), and our results suggest that perhaps the less‐discriminant abiotic pollination mode may lead to more hybridization. Additionally, low variation in genome size within a taxonomic group (which may signal the absence of ploidy variation) may be associated with the formation of hybrids, because ploidy barriers may block hybridization.
Interestingly, we failed to detect associations between hybridization and several hypothesized drivers. We (and others, Table 1, Table S1) posited that many of these traits would enable increased formation of hybrids via opportunity in sheer numbers or wide distributions (agricultural status, Red List status), or via reduced prezygotic barriers to hybrid formation (floral symmetry, breeding system). We note that the lack of detected associations could either be biologically real, or due to small sample sizes for some traits (Table S3). Further, other potential correlates not tested in this study could also promote the formation of hybrids (e.g., disturbance and low genetic divergence; Table 1).
CORRELATES OF HYBRIDIZATION: FACTORS THAT MAY ALLOW FOR HYBRID PERSISTENCE
Lineages may also have detectable associations with specific factors that allow for the persistence of hybrids once they have been formed. Early‐generation hybrids are generally thought to exhibit either decreased fitness (hybrid breakdown) or, conversely, increased fitness (heterosis). The persistence of a hybrid lineage could be linked to either overcoming the latter or sustaining the former (stabilized heterosis). Long lifespans (associated with our traits perenniality and woodiness) may allow hybrid individuals with partial sterility to still have high levels of lifetime fitness, as a small number of viable seeds produced over multiple seasons can result in many offspring over time (Ellstrand et al. 1996). Thus, the association we detected between perenniality/woodiness and hybridization rate could be driven by effects on both hybrid formation (via outcrossing, see above) and persistence.
At the other extreme, heterosis due to heterozygosity at loci throughout the genome is expected to decline as sexual recombination results in the pairing of homozygous alleles in offspring (Conner and Hartl 2004). Stabilized heterosis is the preservation of the increase in fitness through time. Stabilized heterosis can be achieved through vegetative propagation, where early‐generation fitness is maintained via the production of new individuals with a genetic composition identical to that of the parent. Consistent with this idea, we found that genera with more hybrids tended to have more vegetative reproductive systems (vs. generative) (Fig. 3, Table 3). There are several examples of clonal hybrids, for instance in Tamarix (Gaskin and Schaal 2002) and Myriophyllum (Moody and Les 2002), and in many crop plants (reviewed in McKey et al. 2010).
Not all reproduction without outcrossing, however, is capable of preserving stabilized heterosis. For example, selfing (autogamy) should result in acceleration of the loss of heterosis due to a rapid reduction in heterozygosity (e.g., Johansen‐Morris and Latta 2006). If a hybrid forms and then reproduces by selfing rather than outcrossing, it will not have the benefit of stabilized heterosis and the hybrid lineage may fail to persist. We found higher outcrossing rates in plant groups with more hybrids, perhaps reflecting this lack of hybrid persistence in selfing groups.
Some previous work in the genus Cirsium suggests that species with smaller genome sizes are more likely to form hybrids (Bureš et al. 2004). Although only marginally statistically significant, our evidence suggests a trend that groups with larger genomes can be associated with higher levels of hybridization propensity, contrary to this previous work. The association between larger genome sizes and higher hybridization rates could be due to the presence of numerous allopolyploids (hybrids produced from complete genomes of different species) within the group. Allopolyploidy could contribute to both high estimates of hybridization rates and large genome sizes for a given plant group, resulting in the observed associations. Further study is needed to investigate this pattern.
EFFECTS OF TAXONOMIC SCALE
Lineages that are more distantly related (longer time since divergence) tend to have stronger reproductive barriers between them than lineages that are more closely related (less time since divergence) (Coyne and Orr 1989, 1997; Moyle and Nakazato 2010), although there are exceptions and this pattern may be dependent on other aspects of taxonomic scale (Moyle et al. 2004; Scopece et al. 2008; Nosrati et al. 2011). The majority of plant hybridization takes places within genera (Whitney et al. 2010), although instances of intergeneric hybridization have been observed, especially in nonflowering plants (Wagner et al. 1992; Wagner 1993; Fraser‐Jenkins 1997; Garland and Moore 2012; Arrigo et al. 2013; Larson et al. 2014; Rothfels et al. 2015). We collected data at the generic level and analyzed these data at both the family (weighted) and genus taxonomic levels. Regressions tended to be more well supported at the generic level after accounting for multiple comparisons (Fig. 3, Table 3). We found no well‐supported relationship at one taxonomic level that was well supported in the opposite direction at the other taxonomic level. Relationships found at the generic level and not found at the family level (for instance, between hybridization rate and reproductive system) could be due either to sample size differences (a statistical explanation) or the facts that genera within families differ with respect to specific traits, and that hybridization largely takes place within genera (a biological explanation). Relationships supported at the family level and not found at the genus level (for instance, between hybridization and pollination syndrome) could be due to increased precision in estimating both trait values and hybridization metrics within families, as the latter contain greater numbers of species than do genera.
MEASURES OF HYBRIDIZATION
Our measures of hybridization were based on the number of unique hybrid combinations produced, either as a proportion of potential hybrid combinations or simply using the number of nonhybrid species as a denominator. Our findings using both hybridization propensity and hybrid ratio were largely consistent. Not only were they significantly correlated at both the family and genus levels (Table S4) but their relationships with our proposed plant attributes were largely consistent. There were differences in significance when examining one or the other, but the trends were similar (Fig. 3, Table 3). We note that there is another metric that we did not employ, hybridization frequency, which takes into account the fraction of hybridizing parental species rather than their resultant taxa (Mallet 2005; Beddows and Rose 2018). Our database was constructed following Ellstrand et al. 1996 in a way that does not allow for the implementation of this metric, as we did not keep track of parental species. However, we note that the three hybridization metrics can be highly correlated (e.g., Beddows and Rose 2018) and thus suggest that analyses using hybridization frequency may not detect patterns different from those we report.
LIMITATIONS
Although this study examines published floras that span three different continents and one island group, our conclusions may be limited and biased by the geographic extent examined. All but two of our floras are from Europe and mainland North America, with the Victoria, Australia and Hawai'i floras representing the Pacific Region. Four of the floras are from mainland North America, and these include almost half of all species observations (Table S2). To expand this dataset to other regions, we need comprehensive regional floras that specifically record instances of hybridization. Such floras are difficult to find, as they require both interest in hybrids by the authors and the decision to include information on them in the floristic treatment.
We collected data on hybridization using a method suited to their detection in regional floras. There is increasing evidence for instances of hybridization that are not necessarily morphologically apparent but are inferred using genetic or molecular evidence (i.e., Cronn and Wendel 2004; Soltis et al. 2007; McIntosh et al. 2014; Mitchell and Holsinger 2018). At present, a comprehensive analysis including cryptic hybrids is not feasible, but as molecular methods become increasingly common (reviewed in Taylor and Larson 2019), a reanalysis incorporating expanded means of detecting hybrids would surely provide further insights.
Conclusions
We found several phylogenetically informed associations between hybridization rates and plant attributes. Perenniality and woodiness across taxonomic levels, higher outcrossing rates and abiotic pollination syndromes at the family level, and less variable genome sizes at the genus level all associated with increased hybridization metrics and may be acting by increasing the formation of hybrids. Additionally, the associations between increased hybridization and perenniality, woodiness, outcrossing, and genome size, as well as more vegetative reproductive systems at the genus level, may be due to these factors increasing the persistence of hybrids that have already formed. We recognize that this evidence is correlational in nature and does not provide any causal inferences. Moreover, the explanatory power of our models was extremely low (as measured by adjusted R 2 values, Table 3). We caution that while we detected significant statistical associations, the vast majority of variation in hybridization rates remains unexplained. Future work is needed to experimentally test the nature of the relationships that we present here on a global scale. For instance, experiments comparing the evolutionary trajectories and population dynamics of closely related species pairs that are either abiotically or biotically pollinated (or both, such as ambophilous plants) could detect differences in rates of hybrid formation, and thus could support our correlative data. Our findings provide strong hypotheses for further investigating the drivers of hybridization and will aid in not only understanding hybridization as a stand‐alone phenomenon, but also its role in invasion, range expansion, speciation, radiation, and diversification.
Associate Editor: S. Wright
Supporting information
Figure S1.
Table S1. Potential mechanisms underlying proposed associations between traits and hybridization.
Table S2. Summary hybridization data collected from floras.
Table S3. Summary information for trait data used in this study.
Table S4. Raw correlations among traits and measures of hybridization.
Table S5. Phylogenetic path analysis model summaries.
Table S6. Phylogenetic path analysis model coefficients.
AUTHOR CONTRIBUTIONS
KDW and LGC conceived of the original study. KDW, LGC, NM, JRA, and KCP collected data, and ABG contributed data through the TRY database. NM performed the analyses. NM and KDW wrote the manuscript. All authors contributed to revisions.
ACKNOWLEDGMENTS
Funding for the study has been provided by NSF DEB 1257965 and UNM startup funds (both to KDW). Thanks to Loren Albert for assistance with trait scoring and to the Whitney‐Rudgers lab members and four anonymous reviewers for feedback. This study has been supported by the TRY initiative on plant traits (http://www.try-db.org). The TRY initiative and database is hosted, developed, and maintained by J. Kattge and G. Bönisch (Max Planck Institute for Biogeochemistry, Jena, Germany). TRY is currently supported by the DIVERSITAS/Future Earth and the German Centre for Integrative Biodiversity Research (iDiv) Halle‐Jena‐Leipzig.
DATA ARCHIVING
All hybridization data and phylogenetic trees are available from the Dryad digital repository https://doi.org/10.5061/dryad.q83bk3jdd.
LITERATURE CITED
- Abbott, R. , Albach D., Ansell S., Arntzen J. W., Baird S. J. E., Bierne N. et al. 2013. Hybridization and speciation. J. Evol. Biol. 26:229–246. [DOI] [PubMed] [Google Scholar]
- Abbott, R. J. 2017. Plant speciation across environmental gradients and the occurrence and nature of hybrid zones. J. Syst. Evol. 55:238–258. [Google Scholar]
- Albertin, W. , and Marullo P.. 2012. Polyploidy in fungi: evolution after whole‐genome duplication. Proc. R. Soc. Lond., B, Biol. Sci. 279:2497–2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allard, R. W. 1999. Principles of plant breeding. John Wiley & Sons, New York, NY. [Google Scholar]
- Allendorf, F. W. , Leary R. F., Spruell P., and Wenburg J. K.. 2001. The problems with hybrids: setting conservation guidelines. Trends Ecol. Evol. 16:613–622. [Google Scholar]
- Anderson, E. , and Stebbins G. L.. 1954. Hybridization as an evolutionary stimulus. Evolution 8:378–388. [Google Scholar]
- Arnold, M. L. 2006. Evolution through genetic exchange. Oxford Univ. Press, Oxford, U.K. [Google Scholar]
- Arrigo, N. , Therrien J., Anderson C. L., Windham M. D., Haufler C. H., and Barker M. S.. 2013. A total evidence approach to understanding phylogenetic relationships and ecological diversity in Selaginella subg. Tetragonostachys. Am. J. Bot. 100:1672–1682. [DOI] [PubMed] [Google Scholar]
- Baillie, J. , Hilton‐Taylor C., and Stuart S. N.. 2004. 2004 IUCN Red List of threatened species: a global species assessment. IUCN, Gland. [Google Scholar]
- Barton, N. H. 2001. The role of hybridization in evolution. Mol. Ecol. 10:551–568. [DOI] [PubMed] [Google Scholar]
- Beddows, I. , and Rose L. E.. 2018. Factors determining hybridization rate in plants: a case study in Michigan. Perspect. Plant Ecol. Evol. Syst. 34:51–60. [Google Scholar]
- Benjamini, Y. , and Hochberg Y.. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B. Stat. Methodol. 57:289–300. [Google Scholar]
- Bennett, M. D. , and Leitch I. J.. 2005. Plant DNA C‐values database. Royal Botanic Gardens Kew, Kew, England.
- Bruggeman, J. , Heringa J., and Brandt B. W.. 2009. PhyloPars: estimation of missing parameter values using phylogeny. Nucleic Acids Res. 37:W179–W184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bureš, P. , Wang Y. F., Horová L., and Suda J.. 2004. Genome size variation in Central European species of Cirsium (Compositae) and their natural hybrids. Ann. Bot. 94:353–363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell, L. G. , Blanchette C. M., and Small E.. Risk analysis of gene flow from cultivated, addictive, social‐drug plants to wild relatives. Bot. Rev. 85:149–185. [Google Scholar]
- Conner, J. K. , and Hartl D. A.. 2004. A primer of ecological genetics. Sinauer Associates Inc., Sunderland, MA. [Google Scholar]
- Coyne, J. A. , and Orr H. A.. 1989. Patterns of speciation in Drosophila . Evolution 43:362–381. [DOI] [PubMed] [Google Scholar]
- Coyne, J. A. , and Orr H. A. 1997. Patterns of speciation in Drosophila . Evolution 51:295–303. [DOI] [PubMed] [Google Scholar]
- Cronn, R. , and Wendel J. F.. 2004. Cryptic trysts, genomic mergers, and plant speciation. New Phytol. 161:133–142. [Google Scholar]
- Cronquist, A. , Holmgren S., Holmgren N., Reveal J., and Holmgren P.. 1972. Intermountain Flora vols. 1, 2A, 2B, 3A, 3B, 4, 5, 6. Hafner, New York, NY. [Google Scholar]
- de Frutos, Á. , Navarro T., Pueyo Y., and Alados C. L.. 2015. Inferring resilience to fragmentation‐induced changes in plant communities in a semi‐arid Mediterranean ecosystem. PLoS One 10:e0118837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Gasper, A. L. , Almeida T. E., Dittrich V. A. O., Smith A. R., and Salino A.. 2017. Molecular phylogeny of the fern family Blechnaceae (Polypodiales) with a revised genus‐level treatment. Cladistics 33:429–446. [DOI] [PubMed] [Google Scholar]
- Diaz, S. , Hodgson J., Thompson K., Cabido M., Cornelissen J., Jalili A., et al. 2004. The plant traits that drive ecosystems: evidence from three continents. J. Veg. Sci. 15:295–304. [Google Scholar]
- Dressler, S. , Schmidt M., and Zizka G.. 2014. Introducing African plants—a photo guide—an interactive photo data‐base and rapid identification tool for continental Africa. Taxon 63:1159–1161. [Google Scholar]
- Ebihara, A. , Dubuisson J. Y., Iwatsuki K., Hennequin S., and Ito M.. 2006. A taxonomic revision of Hymenophyllaceae. Blumea 51:221–280. [Google Scholar]
- Ellstrand, N. C. , and Hoffman C. A.. 1990. Hybridization as an avenue of escape for engineered genes. Bioscience 40:438–442. [Google Scholar]
- Ellstrand, N. C. , and Schierenbeck K. A.. 2000. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc. Natl. Acad. Sci. USA 97:7043–7050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellstrand, N. C. , Whitkus R., and Rieseberg L. H.. 1996. Distribution of spontaneous plant hybrids. Proc. Natl. Acad. Sci. USA 93:5090–5093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felsenstein, J . 1985. Phyogenies and the comparative method. Amer. Nat. 125:1–15. [Google Scholar]
- Fitter, A. H. , and Peat H. J.. 1994. The ecological flora database. J. Ecol. 82:415–425. [Google Scholar]
- Focke, W. O. 1881. Die Pflanzen‐Mischlinge: Ein Beitrag zur Biologie der Gewächse. Gebr. Borntraeger, Berlin.
- Fraser‐Jenkins, C. R. 1997. New syndrome in Indian pteridology and the ferns of Nepal. International Book Distributors, Dehradun, India. [Google Scholar]
- Gachet, S. , Véla E., and Tatoni T.. 2005. BASECO: a floristic and ecological database of Mediterranean French flora. Biodivers. Conserv. 14:1023–1034. [Google Scholar]
- Garland, M. A. , and Moore G.. 2012. Hesperotropsis, a new nothogenus for intergeneric crosses between Hesperocyparis and Callitropsis (Cupressaceae), and a review of the complicated nomenclatural history of the Leyland cypress. Taxon 61:667–670. [Google Scholar]
- Gaskin, J. F. , and Schaal B. A.. 2002. Hybrid Tamarix widespread in US invasion and undetected in native Asian range. Proc. Natl Acad. Sci. USA 99:11256–12259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giroldo, A. B. 2016. Pequenas plantas, grandes estratégias: adaptações e sobrevivência no Cerrado. Universidade de Brasília, Brazil. [Google Scholar]
- Goodwillie, C. , Kalisz S., and Eckert C. G.. 2005. The evolutionary enigma of mixed mating systems in plants: occurrence, theoretical explanations, and empirical evidence. Annu. Rev. Ecol. Evol. Syst. 36:47–79. [Google Scholar]
- Goolsby, E. W. , Bruggeman J., and Ané C.. 2017. Rphylopars: fast multivariate phylogenetic comparative methods for missing data and within‐species variation. Methods Ecol. Evol. 8:22–27. [Google Scholar]
- Grafen, A. 1989. The phylogenetic regression. Philos. Trans. R. Soc. Lond., Biol. Sci. 326:119–157. [DOI] [PubMed] [Google Scholar]
- Grant, V. 1958. The regulation of recombination in plants. Cold Spring Harb. Symp. Quant. Biol. 23:337–363. [DOI] [PubMed] [Google Scholar]
- Grant, V. 1981. Plant speciation. Columbia Univ. Press, New York, NY. [Google Scholar]
- Guo, Q. 2014. Plant hybridization: the role of human disturbance and biological invasion. Divers. Distrib. 20:1345–1354. [Google Scholar]
- Hauk, W. D. , Parks C. R., and Chase M. W.. 2003. Phylogenetic studies of ophioglossaceae: evidence from rbcL and trnL‐F plastid DNA sequences and morphology. Mol. Phylogenet. Evol. 28:131–151. [DOI] [PubMed] [Google Scholar]
- He, L. J. , and Zhang X. C.. 2012. Exploring generic delimitation within the fern family thelypteridaceae. Mol. Phylogenet. Evol. 65:757–764. [DOI] [PubMed] [Google Scholar]
- Hickman, J. C. 1993. The Jepson manual. Univ. of California Press, Berkeley, CA. [Google Scholar]
- Hovick, S. M. , and Whitney K. D.. 2014. Hybridisation is associated with increased fecundity and size in invasive taxa: meta‐analytic support for the hybridisation‐invasion hypothesis. Ecol. Lett. 17:1464–1477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hovick, S. M. , Campbell L. G., Snow A. A., and Whitney K. D.. 2012. Hybridization alters early life‐history traits and increases plant colonization success in a novel region. Amer. Nat. 179:192–203. [DOI] [PubMed] [Google Scholar]
- Johansen‐Morris, A. D. , and Latta R. G.. 2006. Fitness consequences of hybridization between ecotypes of Avena barbata: hybrid breakdown, hybrid vigor, and transgressive segregation. Evolution 60:1585–1595. [PubMed] [Google Scholar]
- Kattge, J. , Diaz S., Lavorel S., Prentice I. C., Leadley P., Bönisch G. et al. 2011. TRY–a global database of plant traits. Global Change Biol. 17:2905–2935. [Google Scholar]
- Kennedy, J. D. , Borregaard M. K., Marki P. Z., Machac A., Fjedså J., and Rahbek C.. 2018. Expansion in geographical and morphological space drives continued lineage diversification in a global passerine radiation. Proc. Roy. Soc. B. 285:2181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klimešová, J. , and de Bello F.. 2009. CLO‐PLA: the database of clonal and bud bank traits of Central European flora. J. Veg. Sci. 20:511–516. [DOI] [PubMed] [Google Scholar]
- Koike, F. 2001. Plant traits as predictors of woody species dominance in climax forest communities. J. Veg. Sci. 12:327–336. [Google Scholar]
- Kühn, I. , Durka W., and Klotz S.. 2004. BiolFlor: a new plant‐trait database as a tool for plant invasion ecology. Divers. Distrib. 10:363–365. [Google Scholar]
- Larson, E. L. , White T. A., Ross C. L., and Harrison R. G.. 2014. Gene flow and the maintenance of species boundaries. Mol. Ecol. 23:1668–1678. [DOI] [PubMed] [Google Scholar]
- Liu, H. M. , Zhang X. C., Wang W., Qiu Y. L., and Chen Z. D.. 2007. Molecular phylogeny of the fern family Dryopteridaceae inferred from chloroplast rbcL and atpB genes. Int. J. Plant Sci. 168:1311–1323. [Google Scholar]
- Maddison, W. , and Maddison D.. 2018. Mesquite: a modular system for evolutionary analysis v. 3.40.
- Magee, D. W. , and Ahles H. E.. 1999. Flora of the northeast: a manual of the vascular flora of New England and adjacent New York. Univ. of Massachusetts Press, Amherst, MA. [Google Scholar]
- Mallet, J. 2005. Hybridization as an invasion of the genome. Trends Ecol. Evol. 20:229–237. [DOI] [PubMed] [Google Scholar]
- Mallet, J. 2007. Hybrid speciation. Nature 446:279–283. [DOI] [PubMed] [Google Scholar]
- Marques, D. A. , Meier J. I., and Seehausen O.. 2019. A combinatorial view on speciation and adaptive radiation. Trends Ecol. Evol. 34:531–544. [DOI] [PubMed] [Google Scholar]
- Martins, E. P. , and Hansen T. F.. 1997. Phylogenies and the comparative method: a general approach to incorporating phylogenetic information into the analysis of interspecific data. Amer. Nat. 149:646–667. [Google Scholar]
- Mavarez, J. , and Linares M.. 2008. Homoploid hybrid speciation in animals. Mol. Ecol. 17:4181–4185. [DOI] [PubMed] [Google Scholar]
- McGregor, R. L. , and Barkley T. M.. 1986. Flora of the great plains. Univ. Press of Kansas, Lawrence, KS. [Google Scholar]
- McKey, D. , Elias M., Pujol B., and Duputié A.. 2010. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 186:318–332. [DOI] [PubMed] [Google Scholar]
- McIntosh, E. J. , Rossetto M., Weston P. H., and Wardle G. M.. 2014. Maintenance of strong morphological differentiation despite ongoing natural hybridization between sympatric species of Lomatia (Proteaceae). Ann. Bot. 113:861–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell, N. , and Holsinger K. E.. 2018. Cryptic natural hybridization between two species of Protea . S. Afr. J. Bot. 118:306–314. [Google Scholar]
- Moeller, D. A. , Briscoe Runquist R. D., Moe A. M., Geber M. A., Goodwillie C., Cheptou P. O. et al. 2017. Global biogeography of mating system variation in seed plants. Ecol. Lett. 20:375–384. [DOI] [PubMed] [Google Scholar]
- Moody, M. L. , and Les D. H.. 2002. Evidence of hybridity in invasive watermilfoil (Myriophyllum) populations. Proc. Natl Acad. Sci. USA 99:14867–14871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moretti, M. , and Legg C.. 2009. Combining plant and animal traits to assess community functional responses to disturbance. Ecography 32:299–309. [Google Scholar]
- Moyle, L. C. , and Nakazato T.. 2010. Hybrid incompatibility “snowballs” between Solanum species. Science 329:1521–1523. [DOI] [PubMed] [Google Scholar]
- Moyle, L. C. , Olson M. S., and Tiffin P.. 2004. Patterns of reproductive isolation in three angiosperm genera. Evolution 58:1195–1208. [DOI] [PubMed] [Google Scholar]
- Nosrati, H. , Price A. H., and Wilcox C. C.. 2011. Relationship between genetic distances and postzygotic reproductive isolation in diploid Fragaria (Rosaceae). Bot. J. Linn. Soc. 104:510–526. [Google Scholar]
- Ogaya, R. , and Peñuelas J.. 2003. Comparative field study of Quercus ilex and Phillyrea latifolia: photosynthetic response to experimental drought conditions. Environ. Exper. Bot. 50:137–148. [Google Scholar]
- Onstein, R. E. , Carter R. J., Xing Y., and Linder H. P.. 2014. Diversification rate shifts in the cape floristic region: the right traits in the right place at the right time. Perspect. Plant Ecol. Evol. Syst. 16:331–340. [Google Scholar]
- Paradis, E. , Claude J., and Strimmer K.. 2004. APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20:289–290. [DOI] [PubMed] [Google Scholar]
- Paun, O. , Forest F., Fay M. F., and Chase M. W.. 2009. Hybrid speciation in angiosperms: parental divergence drives ploidy. New Phytol. 182:507–518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pryer, K. M. , Schuettpelz E., Wolf P. G., Schneider H., Smith A. R., and Cranfill R.. 2004. Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences. Am. J. Bot. 91:1582–1598. [DOI] [PubMed] [Google Scholar]
- Qian, H. , and Jin Y.. 2016. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant. Ecol. 9:233–239. [Google Scholar]
- R Core Development Team . 2016. R: a language and environment for statistical computing. Vienna, Austria.
- Revelle, W. R. 2017. psych: procedures for personality and psychological research.
- Rhymer, J. M. , and Simberloff D.. 1996. Extinction by hybridization and introgression. Annu. Rev. Ecol. Evol. Syst. 27:83–109. [Google Scholar]
- Rieseberg, L. H. 2003. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 301:1211–1216. [DOI] [PubMed] [Google Scholar]
- Rieseberg, L. H. , and Wendel J. F.. 1993. Introgression and its consequences in plants Pp. 70–109 in Harrison R.G., ed. Hybrid zones and the evolutionary process. Oxford Univ. Press, Oxford, U.K. [Google Scholar]
- Rieseberg, L. H. , Kim S. C., Randell R. A., Whitney K. D., Gross B. L., Lexer C. et al. 2007. Hybridization and the colonization of novel habitats by annual sunflowers. Genetica 129:149–165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothfels, C. J. , Johnson A. K., Hovenkamp P. H., Swofford D. L., Roskam H. C., Fraser‐Jenkins C. R. et al. 2015. Natural hybridization between genera that diverged from each other approximately 60 million years ago. Amer. Nat. 185:433–442. [DOI] [PubMed] [Google Scholar]
- Sargent, R. D. 2004. Floral symmetry affects speciation rates in angiosperms. Proc. R. Soc. Lond., B, Biol. Sci. 271:603–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schierenbeck, K. A. , and Ellstrand N. C.. 2009. Hybridization and the evolution of invasiveness in plants and other organisms. Biol. Invasions 11:1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider, H. , Russell S. J., Cox C. J., Bakker F., Henderson S., Rumsey F. et al. 2004a. Chloroplast phylogeny of asplenioid ferns based on rbcL and trnL‐F spacer sequences (Polypodiidae, Aspleniaceae) and its implications for biogeography. Syst. Bot. 29:260–274. [Google Scholar]
- Schneider, H. , Smith A. R., Cranfill R., Hildebrand T. J., Haufler C. H., and Ranker T. A.. 2004b. Unraveling the phylogeny of polygrammoid ferns (Polypodiaceae and Grammitidaceae): exploring aspects of the diversification of epiphytic plants. Mol. Phylogenet. Evol. 31:1041–1063. [DOI] [PubMed] [Google Scholar]
- Schwenk, K. , Brede N., and Streit B.. 2008. Introduction. Extent, processes and evolutionary impact of interspecific hybridization in animals. Philos. Trans. R. Soc. Lond., Biol. Sci. 363:2805–2811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scopece, G. , Widmer A., and Cozzolino S.. 2008. Evolution of postzygotic reproductive isolation in a guild of deceptive orchids. Amer. Nat. 171:315–326. [DOI] [PubMed] [Google Scholar]
- Seehausen, O. 2004. Hybridization and adaptive radiation. Trends Ecol. Evol. 19:198–207. [DOI] [PubMed] [Google Scholar]
- Shipley, B. 2013. The AIC model selection method applied to path analytic models compared using a d‐separation test. Ecology 94:560–564. [DOI] [PubMed] [Google Scholar]
- Smith, A. R. , Pryer K. M., Schuettpelz E., Korall P., Schneider H., and Wolf P. G.. 2006. A classification for extant ferns. Taxon 55:705–731. [Google Scholar]
- Soltis, D. E. , Soltis P. S., Schemske D. W., Hancock J. F., Thompson J. N., Husband B. C. et al. 2007. Autopolyploidy in angiosperms: have we grossly underestimated the number of species? Taxon 56:13–30. [Google Scholar]
- Soltis, P. S. , and Soltis D. E.. 2009. The role of hybridization in plant speciation. Annu. Rev. Plant Biol. 60:561–588. [DOI] [PubMed] [Google Scholar]
- Stace, C. 1997. New flora of the British Isles. 2nd ed. Cambridge Univ. Press, Cambridge, U.K. [Google Scholar]
- Stace, C. A. 1975. Hybridization and the Flora of the British Isles. Academic Press, London. [Google Scholar]
- Stebbins, G. L. 1959. The role of hybridization in evolution. Proc. Am. Philos. Soc. 103:231–251. [Google Scholar]
- Stelkens, R. , and Seehausen O.. 2009. Genetic distance between species predicts novel trait expression in their hybrids. Evolution 63:884–897. [DOI] [PubMed] [Google Scholar]
- Sundue, M. A. , Parris B. S., Ranker T. A., Smith A. R., Fujimoto E. L., Zamora‐Crosby D. et al. 2014. Global phylogeny and biogeography of grammitid ferns (Polypodiaceae). Mol. Phylogenet. Evol. 81:195–206. [DOI] [PubMed] [Google Scholar]
- Taylor, S. A. , and Larson E. L.. 2019. Insights from genomes into the evolutionary importance and prevalence of hybridization in nature. Nature Ecol. Evol. 3:170–177. [DOI] [PubMed] [Google Scholar]
- Todesco, M. , Pascual M. A., Owens G. L., Ostevik K. L., Moyers B. T., Hübner S. et al. 2016. Hybridization and extinction. Evol. Appl. 9:892–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turland, N. J. , Wiersema J. H., Barrie F. R., Greuter W., Hawksworth D. L., Herendeen P. S. et al. 2018. International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Koeltz Botanical Books, Glashütten. [Google Scholar]
- Tutin, T. G. , Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M. et al. 1964. Flora Europaea, vols. 1–5 Cambridge Univ. Press, Cambridge, U.K. [Google Scholar]
- van der Bijl, W. 2018. Phylopath: easy phylogenetic path analysis in R. PeerJ 6:e4718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Hardenberg, A. , and Gonzalez‐Voyer A.. 2013. Disentangling evolutionary cause‐effect relationships with phylogenetic confirmatory path analysis. Evolution 67:378–387. [DOI] [PubMed] [Google Scholar]
- Wagner, W. H. 1993. New species of Hawaiian pteridophytes. Contrib. Univ. Mich. Herb 19:63–82. [Google Scholar]
- Wagner, W. H. , Wagner F. S., Reznicek A. A., and Werth C. R.. 1992. xDryostichum singulare (Dryopteridaceae), a new fern nothogenus from Ontario. Can. J. Bot. 70:245–253. [Google Scholar]
- Wagner, W. L. , Herbst D. R., and Sohmer S. H.. 1999. Manual of the Flowering Plants of Hawai'i, Vols. 1 and 2 Univ. of Hawai'i and Bishop Museum Press, Honolulu, HI. [Google Scholar]
- Walsh, N. G. , and Entwisle T. J.. 1994. Flora of Victoria, Vols. 2–4 Inkata Press, Melbourne, Australia. [Google Scholar]
- Whitney, K. D. , Ahern J. R., Campbell L. G., Albert L. P., and King M. S.. 2010. Patterns of hybridization in plants. Perspect. Plant Ecol. Evol. Syst. 12:175–182. [Google Scholar]
- Wolf, J. B. , Bayer T., Haubold B., Schilhabel M., Rosenstiel P., and Tautz D.. 2010. Nucleotide divergence vs. gene expression differentiation: comparative transcriptome sequencing in natural isolates from the carrion crow and its hybrid zone with the hooded crow. Mol. Ecol. 19(Suppl 1):162–175. [DOI] [PubMed] [Google Scholar]
- Yakimowski, S. B. , and Rieseberg L. H.. 2014. The role of homoploid hybridization in evolution: a century of studies synthesizing genetics and ecology. Am. J. Bot. 101:1247–1258. [DOI] [PubMed] [Google Scholar]
- Zanne, A. E. , Tank D. C., Cornwell W. K., Eastman J. M., Smith S. A., FitzJohn R. G. et al. 2014. Three keys to the radiation of angiosperms into freezing environments. Nature 506:89. [DOI] [PubMed] [Google Scholar]
- Zapiola, M. L. , Campbell C. K., Butler M. D., and Mallory‐Smith C. A.. 2008. Escape and establishment of transgenic glyphosate‐resistant creeping bentgrass Agrostis stolonifera in Oregon, USA: a 4‐year study. J. Appl. Ecol. 45:486–494. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1.
Table S1. Potential mechanisms underlying proposed associations between traits and hybridization.
Table S2. Summary hybridization data collected from floras.
Table S3. Summary information for trait data used in this study.
Table S4. Raw correlations among traits and measures of hybridization.
Table S5. Phylogenetic path analysis model summaries.
Table S6. Phylogenetic path analysis model coefficients.