

Abstract
Background
Moringa oleifera Lam. is a commonly used plant in herbal medicine and has various reported bioactivities such as antioxidant, antimicrobial, anticancer and antidiabetes. It is rich in nutrients and polyphenols. The plant also has been traditionally used for alleviating allergic conditions. This study was aimed to examine the anti-allergic activity of M. oleifera extracts and its isolated compounds.
Method
M. oleifera leaves, seeds and pods were extracted with 80% of ethanol. Individual compounds were isolated using a column chromatographic technique and elucidated based on the nuclear magnetic resonance (NMR) and electrospray ionisation mass spectrometry (ESIMS) spectral data. The anti-allergic activity of the extracts, isolated compounds and ketotifen fumarate as a positive control was evaluated using rat basophilic leukaemia (RBL-2H3) cells for early and late phases of allergic reactions. The early phase was determined based on the inhibition of beta-hexosaminidase and histamine release; while the late phase was based on the inhibition of interleukin (IL-4) and tumour necrosis factor (TNF-α) release.
Results
Two new compounds; ethyl-(E)–undec-6-enoate (1) and 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) together with six known compounds; quercetin (3), kaempferol (4), β-sitosterol-3-O-glucoside (5), oleic acid (6), glucomoringin (7), 2,3,4-trihydroxybenzaldehyde (8) and stigmasterol (9) were isolated from M. oleifera extracts. All extracts and the isolated compounds inhibited mast cell degranulation by inhibiting beta-hexosaminidase and histamine release, as well as the release of IL-4 and TNF-α at varying levels compared with ketotifen fumarate.
Conclusion
The study suggested that M. oleifera and its isolated compounds potentially have an anti-allergic activity by inhibiting both early and late phases of allergic reactions.
Keywords: Moringa oleifera, Anti-allergic, Isolation, RBL-2H3, Histamine, Beta-hexosaminidase, IL-4, TNF- α, New compound
Background
Moringa oleifera Lam. (M. oleifera) is well known for its nutritional and medicinal values [1]. It is mainly cultivated in the tropical and subtropical continents due to its high tolerance to extreme weather and as a nutritional source [2–4]. The plant is also commonly known as kelor, drumstick tree, horseradish tree, marunggay and Sajna [1, 5].
M. oleifera has been reported to contain seven times more vitamin C than oranges, four times more vitamin A than carrots, four times more calcium than milk, two times more protein than yogurt and three times more potassium than bananas [6]. Its immature pods, flowers and leaves contain high percentage of essential amino acids [7]. The whole plant has high content of unsaturated and essential fatty acid such as linolenic acid, linoleic acid and oleic acid; as well as micronutrients such as iron, zinc, vitamin A, calcium and β-carotene [8]. The ability of the nutritious plant to rapidly grow in harsh climates with high yield helps in combatting severe malnutrition of the populations in the subtropical and tropical regions [9].
M. oleifera also contains high content of polyphenols that contributes to its high total antioxidant capacity and could be medicinally useful for diseases such as cancer, hypertension and hyperglycaemia [8, 10]. The plant has been reported to have antimicrobial activity against waterborne [11], oral [12] and coliform [13] bacteria, as well as dermatophytes [14]. It has anti-diabetic and anti-inflammatory activities that could be associated with high isothiocyanates content [15–17]. Its polyphenols such as chlorogenic acid, vicenin-2 and quercetin were also found to contribute to the wound healing activity [18].
Allergic reaction cases are increasing by years. The reactions can be as mild as a rash and as severe as an anaphylactic shock that can lead to death. Subsequent exposure of allergens will trigger mast cell degranulation, releasing mediators that exhibit allergic symptoms. Early phase of an allergic reaction usually occurs minutes after sensitisation, triggering the release of histamine, that will induce bronchoconstriction, vasodilation and vascular permeability [19]. Meanwhile, the late phase of an allergic reaction occurring hours after sensitisation will trigger the release of cytokines such as IL-4, IL-5, IL-9, IL-13, TNF-α and IFN-γ that will recruit inflammatory cells and further bring about mast cell degranulation.
Ayurvedic practitioners have been using M. oleifera to alleviate allergic conditions but scientific evidences are still lacking [20]. Due to the high content of flavonoids, the plant could be associated with the anti-allergic activity via inhibition of histamine and IL-4 release [10, 21]. In this study, the anti-allergic activity of M. oleifera extracts from various plant parts and its isolated compounds was evaluated using basophil cells to determine the inhibition of mast cell degranulation and cytokines production.
Methods
Materials and equipment
Pre-coated aluminium plate of thin layer chromatography (TLC) with silica gel 60 GF254 of 20 × 20 cm dimension (Merck, Darmstadt, Germany) was used to visualize separated spots of compounds upon chromatographic development under ultraviolet light (λ254, λ366, model UVGL-58). The compounds isolation process was conducted using a column chromatographic technique with silica gel 60 (230–400 mesh ASTM) (Merck, Darmstadt, Germany) and Sephadex LH20 (GE Healthcare, Uppsala, Sweden) as the stationary phases. Chemical structures of the compounds were elucidated using a nuclear magnetic resonance (NMR) spectrometer (1H NMR 600 MHz and 13C NMR 151 MHz, Cryoprobe, Bruker, Basel, Switzerland). Mass of the compounds was analysed using a high-resolution electrospray ionization mass spectrometer (HRESIMS) (Micro TOF-Q, Bruker, Basel, Switzerland). A 24 well-plate, a 96 well-plate and a 75cm2 cell culture flask were obtained from NEST, China. An incubator (IR 230 Forma, Thermo Scientific, Massachusetts, USA) and an ELISA microplate reader (Tecan Infinite M200 Pro, Mannedorf, Switzerland) were used throughout the bioassays studies.
Chemicals and reagents
The TLC plate was sprayed with 10% sulfuric acid and Dragendorff reagents to visualise the separated spots of compounds obtained. Organic solvents used for the isolation of compounds were of analytical grade (Merck, Darmstadt, Germany). Deuterated chloroform (CDCl3) and methanol (MeOD) were used in the NMR spectroscopy. Ketotifen fumarate (Abcam, Cambridge, United Kingdom) was used as a positive control for the bioassay experiments. Minimum essential media (MEM) with Earle’s salts and L-glutamine, foetal bovine serum (FBS) from Biowest (Missouri, USA) and penicillin-streptomycin (10,000 U/mL) (Gibco, New Hampshire, USA) were used as cell media.
Rat basophilic leukemic (RBL)-2H3 cells were obtained from the Japanese Collection of Research Bioresources Cell Bank (JCRB, Japan). Dimethyl sulfoxide (DMSO) (Merck, New Jersey, USA) was used for dissolving test samples. The concentration of DMSO was kept below 0.25% as it was reported safe for RBL-2H3 cell [22]. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Goldbio Technology, Missouri, USA) was used to determine cell viability. Monoclonal Anti-Dinitrophenyl (DNP) IgE antibody produced in mouse (Sigma-Aldrich, New Hampshire, USA) were used for cell sensitisation. Bovine serum albumin (BSA) solution (Sigma-Aldrich, New Hampshire, USA), dinitrophenyl-bovine serum albumin (DNP-BSA) (Santa Cruz, USA) and p-nitrophenyl-N-acethyl-β-D-glucosaminide (Merck, New Jersey, USA) were used for the bioassays. Histamine, TNF-α and IL-4 Elisa kits were obtained from Elabscience, China.
Plant materials
Leaves, seeds and pods of M. oleifera were obtained from Terengganu, Malaysia. The taxonomy of the specimens was confirmed by Dr. Shamsul Khamis, a botanist from Universiti Kebangsaan Malaysia (UKM). The specimens were deposited at the Universiti Kebangsaan Malaysia Herbarium in Bangi with a voucher specimen UKMB40408.
Preparation of extracts and isolation of compounds
The plant materials were air-dried and ground before the extraction process. The leaves (848.9 g), seeds (1999.2 g) and pods (2000.0 g) were individually macerated with 80% of ethanol for three days and filtered. The residue was extracted twice. Filtrates of each plant part were collected and evaporated using a rotary evaporator. The remaining water content was removed using a freeze-drying technique. The yield of leaf, seed and pod extracts were 42.43 g, 65.05 g and 59.56 g respectively. The extracts were sequentially fractionated using hexane, ethyl acetate and acetone as eluting solvents with various compositions.
Structural elucidation of isolated compounds
Ethyl-(E)-undec-6-enoate (1)
Elution of the hexane leaf fraction (18.91 g) with hexane-ethyl acetate (8:2, v/v) yielded compound 1. White amorphous liquid (34.2 mg), TLC: Rf 0.83 (hexane: EtOAc, 8:2), HRESIMS (+ve mode) m/z: 235.1295 [M + Na]+(calculated for C13H24O2, 212.33 g/mol). 1H NMR (CDCl3, 600 MHz): δH 0.85 (t, 3H, J = 7.1 Hz, H-13), 1.25 (m, 21H, H-1, H-12), 1.32 (m, 21H, H-6), 1.35 (m, 21H, H-11), 1.62 (m, 2H, H-5), 2.02 (m, 2H, H-10), 2.31 (t, 2H, J = 7.7 Hz, H-4), 2.79 (m, 1H, H-7), 4.15 (q, 2H, J = 7.1 Hz, H-2), 5.4 (m, 1H, H-8, H-9). 13C NMR (CDCl3, 150 MHz): δc 14.3 (C-1, C-13), 22.8 (C-12), 24.9 (C-6), 29.1 (C-5), 29.7 (C-11), 31.9 (C-10), 35 (C-4, C7), 60.1 (C-2), 130 (C-8, C-9), 174.3 (C-3).
3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2)
The ethyl acetate leaf fraction (3.38 g) was loaded onto Sephadex LH-20 using methanol-water (1:1, v/v) yielding four fractions. The third fraction was further eluted using Sephadex LH-20 with chloroform-dichloromethane-methanol (4:1:1, v/v) as a mobile phase giving four fractions. The last fraction was further eluted in silica gel column chromatography with ethyl acetate-chloroform (7:3, v/v) as a mobile phase yielding compound 2. Reddish-brown liquid (3.9 mg), TLC: Rf 0.50 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z: 176.9835 [M + 2H]2+(calculated for C15H10O10, 350.24 g/mol) .1H NMR (CDCl3, 600 MHz): δH 6.83 (d, 1H, J = 8.6 Hz, H-7), 7.89 (d, 1H, J = 8.7 Hz, H-8). 13C NMR (CDCl3, 150 MHz): δc 113.3 (C-10), 114.3 (C-7), 122.1 (C-1′), 123.0 (C-6), 128.5 (C-3′), 128.5 (C-5′), 131.4 (C-8), 138.5 (C-3), 138.5 (C-4′), 147.5 (C-2′), 147.5 (C-6′), 161.8 (C-9), 162.9 (C-5), 168.9 (C-2), 173.8 (C-4).
Quercetin (3)
The ethyl acetate leaf fraction (3.38 g) was loaded onto Sephadex LH-20 using methanol-water (1:1, v/v) yielding four fractions. The fourth fraction was further eluted using Sephadex LH-20 with chloroform-dichloromethane-methanol (4:1:1, v/v) as a mobile phase yielding compound 3. Yellow powder (5.9 mg), m.p 312–314 °C. TLC: Rf 0.49 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z: 325.0294 [M + Na]+(calculated for C15H10O7, 302.24 g/mol). 1H NMR (CDCl3, 600 MHz): δH 6.20 (d, 1H, J = 1.8 Hz, H-6), 6.41 (d, 1H, J = 1.8 Hz, H-8), 6.91 (d, 1H, J = 8.4 Hz, H-5′), 7.66 (dd, 1H, J = 8.4 Hz, H-6′), 7.76 (d, 1H, J = 2.4 Hz, H-2′). 13C NMR (CDCl3, 150 MHz): δc 93.3 (C-8), 98.2 (C-6), 103.3 (C-10), 114.9 (C-2′), 115.1 (C-5′), 120.6 (C-6′), 123.0 (C-1′), 136.1 (C-3), 145.1 (C-3′), 146.9 (C-2), 147.6 (C-4′), 157.1 (C-9), 161.3 (C-5), 164.5 (C-7), 176.2 (C-4).
Kaempferol (4)
The ethyl acetate leaf fraction (3.38 g) was loaded onto Sephadex LH-20 using methanol-water (1:1, v/v) yielding four fractions. The fourth fraction was further eluted using Sephadex LH-20 with chloroform-dichloromethane-methanol (4:1:1, v/v) as a mobile phase giving another six fractions. The fifth fraction was subjected to silica gel column chromatography using chloroform-methanol (9:1, v/v) as a mobile phase to obtain compound 4. Yellow powder (1.3 mg), m.p 276–279 °C. TLC: Rf 0.68 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z: 309.0334 [M + Na]+(calculated for C15H10O6, 286.24 g/mol). 1H NMR (CDCl3, 600 MHz): δH 6.19 (s, 1H, H-6), 6.41 (s, 1H, H-8), 6.91 (d, 1H, J = 9.0 Hz, H-3′), 6.91 (d, 1H, J = 9.0 Hz, H-5′), 8.10 (d, 1H, J = 8.4 Hz, H-6′), 8.74 (s, 1H, H-2′).13C NMR (CDCl3, 150 MHz): δc 93.1 (C-8), 97.9 (C-6), 103.2 (C-10), 114.9 (C-3′), 114.9 (C-5′), 122.3 (C-1′), 129.3 (C-2′), 129.3 (C-6′), 135.7 (C-3), 146.7 (C-2), 156.9 (C-9), 159.2 (C-4′), 161.1 (C-5), 164.2 (C-7), 176.0 (C-4).
β-Sitosterol-3-O-glucoside (5)
The acetone leaf fraction (0.95 g) was eluted in the silica gel column chromatography using ethyl acetate-chloroform (7:3, v/v) as a mobile phase to obtain seven fractions. The sixth fraction was separated using ethyl acetate (8:2, v/v) yielding four fractions. The fourth fraction was then purified using chloroform-methanol (9:1, v/v) to give compound 5. White powder (8.5 mg), m.p 209–211 °C. TLC: Rf 0.67 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z: 599.4190 [M + Na]+ (calculated for C35H60O6, 576.86 g/mol). 1H NMR (CDCl3, 600 MHz): δH 0.68 (s, 3H, H-18), 0.83 (m, 7H, H-9), 0.83 (m, 7H, H-14), 0.83 (m, 7H, H-24), 0.83 (m, 7H, H-29), 0.92 (d, 4H, J = 6.6 Hz, H-21), 0.92 (d, 4H, J = 6.6 Hz, H-26), 0.92 (d, 4H, J = 6.6 Hz, H-27), 0.96 (s, 6H, H-19), 1.00 (m, 6H, H-1), 1.00 (m, 6H, H-15), 1.17 (m, 5H, H-12), 1.17 (m, 5H, H-17), 1.25 (m, 4H, H-22), 1.25 (m, 4H, H-23), 1.25 (m, 4H, H-28), 1.37 (m, 6H, H-8), 1.38 (m, 6H, H-20), 1.42 (m, 6H, H-11), 1.52 (m, 5H, H-2), 1.52 (m, 5H, H-15), 1.53 (m, 5H, H-12), 1.65 (m, 3H, H-7), 1.65 (m, 3H, H-25), 1.82 (m, 4H, H-16), 1.95 (m, 4H, H-7), 1.98 (m, 4H, H-4), 2.63 (m, 1H, H-3), 2.90 (m, 2H, H-2′), 3.00 (td, 2H, J = 5.2 Hz, H-4′), 3.08 (dd, 2H, J = 2 Hz, H-5′), 3.12 (td, 2H, J = 4.8 Hz, H-3′), 3.48 (m, 2H, H-2′(OH)), 3.48 (m, 2H, H-3′(OH)), 3.48 (m, 2H, H-4′(OH)), 3.68 (m, 2H, H-6′(OH)), 4.25 (d, 1H, J = 7.8 Hz, H-6′), 4.45 (d, 1H, J = 5.8 Hz, H-6′), 4.91 (d, 3H, J = 6.1 Hz, H-1′), 5.35 (m, 1H, H-6). 13C NMR (CDCl3, 150 MHz): δc 12.2 (C-18), 12.3 (C-29), 19.1 (C-21), 19.4 (C-27), 19.6 (C-19), 20.2 (C-26), 23.1 (C-28), 24.3 (C-15), 25.9 (C-23), 28.3 (C-16), 29.1 (C-25), 29.7 (C-2), 31.8 (C-7), 31.8 (C-8), 31.8 (C-22), 36.0 (C-10), 36.0 (C-20), 36.7 (C-1), 37.3 (C-12), 38.8 (C-4), 42.3 (C-13), 45.6 (C-24), 50.0 (C-9), 55.9 (C-17), 56.6 (C-14), 61.6 (C-6′), 70.6 (C-4′), 73.9 (C-2′), 77.2 (C-5′), 77.3 (C-3), 77.3 (C-3′), 101.2 (C-1′), 121.7 (C-6), 140.9 (C-5).
Oleic acid (6)
The ethyl acetate seed fraction (2.10 g) was eluted in the silica gel column chromatography using hexane-ethyl acetate (7:3, v/v) as a mobile phase yielding three fractions. The second fraction was further eluted using hexane-ethyl acetate (8:2, v/v) yielding compound 6. White amorphous liquid (24.5 mg), TLC: Rf 0.74 (hexane: EtOAc, 7:3), HRESIMS (+ve mode) m/z: 603.4867 [2 M + K]+ (calculated for C18H34O2, 282.47 g/mol). 1H NMR (CDCl3, 600 MHz): δH 0.90 (t, 3H, J = 7.2 Hz, H-18), 1.30 (m, 20H, H-4, H-5, H-6, H-7, H-12, H-13, H-14, H-15, H-16, H-17), 1.65 (m, 2H, H-3), 2.03 (m, 1H, H-8), 2.03 (m, 1H, H-11), 2.37 (t, 2H, J = 7.2 Hz, H-2), 5.37 (m, 1H, H-9), 5.37 (m, 1H, H-9). 13C NMR (CDCl3, 150 MHz): δc 14.1 (C-18), 22.7 (C-17), 24.7 (C-3), 27.2 (C-8), 27.2 (C-11), 29.0 (C-4), 29.0 (C-5), 29.1 (C-14), 29.1 (C-15), 29.4 (C-13), 29.5 (C-6), 29.7 (C-12), 29.8 (C-7), 31.9 (C-16), 34.3 (C-2), 129.8 (C-10), 130.0 (C-9), 179.8 (C-1).
Glucomoringin (7)
The acetone seed fraction (2.21 g) was eluted in the silica gel column chromatography using chloroform-methanol (9:1, v/v) as a mobile phase yielding five fractions. The fourth fraction was eluted using the same mobile phase to obtain compound 7. Yellow liquid (28.1 mg), TLC: Rf 0.36 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z: 572.3460 [M + H]+ (calculated for C20H29NO14S2, 571.57 g/mol). 1H NMR (CDCl3, 600 MHz): δH 1.23 (q, 4H, J = 4.8 Hz, H-6″), 3.20 (m, H-6′), 3.49 (m, H-4″), 3.64 (m, 1H, H-5″), 3.83 (s, 1H, H-7), 3.86 (dt, 1H, J = 1.8, 8.7 Hz, H-3″), 4.03 (m, 1H, H-2″), 4.10 (m, H-5′), 4.23 (m, H-4′), 4.34 (m, H-3′), 4.47 (d, 1H, J = 7.2 Hz, H-1′), 4.64 (m, H-2′), 5.44 (dd, 1H, J = 1.8, 17.4 Hz, H-1′), 7.02 (d, 1H, J = 4.8 Hz, H-1), 7.09 (d, 1H, J = 4.8 Hz, H-6), 7.27 (d, 1H, J = 3 Hz, H-5), 7.30 (d, 1H, J = 8.4 Hz, H-3). 13C NMR (CDCl3, 150 MHz): δc 16.7 (C-6″), 21.2 (C-7), 43.0 (C-4′), 45.5 (C-3′), 47.1 (C-2′), 47.9 (C-6′), 59.8 (C-5′), 65.9 (C-1′), 69.2 (C-5″), 69.3 (C-2″), 70.6 (C-3″), 72.3 (C-4″), 98.4 (C-1″), 116.1 (C-6), 116.7 (C-2), 128.7 (C-3), 129.0 (C-5), 131.7 (C-4), 155.7 (C-1), 156.0 (N).
2,3,4-trihydroxybenzaldehyde (8)
The hexane pod fraction (19.2 g) was eluted on the silica gel column chromatography using hexane-ethyl acetate (8:2, v/v) as a mobile phase yielding four fractions. The second fraction was further eluted using hexane-ethyl acetate (9:1, v/v) giving five fractions. The third fraction was purified using hexane-ethyl acetate (7:3, v/v) yielding compound 8. Reddish brown liquid (1.8 mg), TLC: Rf 0.73 (CHCl3: MeOH, 9:1), HRESIMS (+ve mode) m/z193.0489[M + K] +(calculated for C7H6O4, 154.12 g/mol). 1H NMR (CDCl3, 600 MHz): δH 6.98 (d, 1H, J = 12 Hz, H-5), 7.83 (d, 1H, J = 12 Hz, H-6), 9.89 (s, 1H, H-7). 13C NMR (CDCl3, 150 MHz): δc 116.0 (C-1), 116.0 (C-5), 130.2 (C-3), 132.4 (C-2), 132.4 (C-4), 161.2 (C-6), 190.8 (C-7).
Stigmasterol (9)
The ethyl acetate pod fraction (3.0 g) was eluted in the silica gel column chromatography using chloroform-ethyl acetate (6:4, v/v) as a mobile phase eluting fourteen fractions. The second fraction was further subjected to silica gel column chromatography using hexane-ethyl acetate (8:2, v/v) to obtain compound 9. White crystal (28.1 mg), m.p 166–168 °C. TLC: Rf 0.39 (Hexane: EtOAc, 9:1), HR ESIMS (+ve mode) m/z: 451.3425 [M + K] + (calculated for C29H48O, 412.70 g/mol). 1H NMR (CDCl3, 600 MHz): δH 0.70 (s, 1H, H-18), 0.82 (m, 3H, H-27), 0.83 (m, 3H, H-29), 0.87 (m, 3H, H-26), 0.95 (m, 3H, H-21), 0.95 (m, 3H, H-9), 1.02 (s, 3H, H-19), 1.05 (m, 2H, H-14), 1.08 (m, 1H, H-15), 1.12 (m, 2H, H-1), 1.18 (m, 2H, H-17), 1.28 (s, 1H, H-12), 1.37 (m, 2H, H-28), 1.46 (m, 1H, H-8), 1.48 (m, 2H, H-11), 1.49 (m, 1H, H-2), 1.52 (m, 1H, H-24), 1.53 (m, 1H, H-25), 1.55 (m, 1H, H-15), 1.58 (m, 2H, H-16), 1.71 (m, 1H, H-2), 1.85 (m, 1H, H-1), 1.88 (m, 2H, H-7), 1.98 (m, 1H, H-12), 2.25 (m, 1H, H-20), 2.32 (m, 1H, H-4), 3.55 (m, 1H, H-3), 5.04 (dd, 1H, J = 15 Hz, H-23), 5.16 (dd, 1H, J = 18 Hz, H-22), 5.38 (m, 1H, H-6). 13C NMR (CDCl3, 150 MHz): δc 12.0 (C-18), 12.3 (C-29), 19.4 (C-19), 19.9 (C-27), 21.1 (C-11), 21.1 (C-26), 21.3 (C-22), 24.3 (C-15), 25.6 (C-28), 29.0 (C-16), 29.8 (C-2), 31.9 (C-7), 31.9 (C-8), 31.9 (C-25), 36.5 (C-10), 37.8 (C-1), 39.8 (C-12), 40.0 (C-20), 42.3 (C-13), 44.0 (C-4), 50.1 (C-9), 51.2 (C-24), 55.9 (C-17), 56.8 (C-14), 71.8 (C-3), 121.8 (C-6), 129.3 (C-23), 138.3 (C-22), 140.8 (C-5).
Cell viability assay
RBL-2H3 cells from the Japanese Collection of Research Bioresources Cell Bank (JCRB, Japan) in enriched media (100 μL) were seeded in 96 well plates with cell concentrations of 2.5 × 105 cells/mL. The cells were incubated for 24 h at 37 °C in a humidified incubator (5% CO2, 95% air) prior to reconstitution with 100 μL of test sample. Then the cells were incubated for another 24 h. 10 μL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (5 mg/mL in PBS buffer) was added into the mixture and was incubated further for 4 h. Upon the appearance of purple formazan crystal, the medium was removed and 100 μL of DMSO was added. After shaking the plates for 10 min, the absorbance was immediately measured using a microplate reader at 570 nm. Percentage of cell viability was determined as using the formula below, whereby OD means optical density:
Preparation of RBL-2H3 cells for degranulation assessment
RBL-2H3 cells were cultured in MEM containing 10% FBS and 1% penicillin-streptomycin at 37 °C with 5% CO2 and 95% air. The medium was changed every two days and the cells were sub-cultured upon 80% confluency. Once the cells reached the third passage with stable morphology and growth, the cells were used for the bioassay experiment. Only cells from third to fifth passages were used in this study.
RBL-2H3 cells (2 × 105 / 400 μL) in enriched medium were seeded and allowed to adhere onto 24 well plates in the humidified incubator (37 °C, 5% CO2, 95% air) for two hours. The cells were then sensitized with anti-DNP IgE (0.45 μg/mL, 100 μL) overnight at 37 °C in 5% CO2.
Inhibitory effect of beta-hexosaminidase release
The inhibitory effect of beta-hexosaminidase release from RBL-2H3 cells were measured using the method described by Shahari et al. [23]. The sensitised cells were washed twice with 500 μL Siraganian buffer (119 mM NaCl, 5 mM KCl, 5.6 mM glucose, 0.4 mM MgCl2, 1 mM CaCl2, 25 mM piperazine-N, N′-bis (2-ethanesulfonic acid) (PIPES), 40 mM NaOH, 0.1% BSA, pH 7.2) and were reconstituted with 160 μL Siraganian buffer. After 10 min of incubation at 37 °C in 5% CO2 and 95% air, the cells were treated with 20 μL of test samples (7.81, 15.62 and 31.25 μg/mL) or 20 μL of ketotifen fumarate (7.81, 15.62 and 31.25 μg/mL) and incubated for another 10 min. Cells degranulation was then stimulated by the addition of 20 μL DNP-BSA (10 mg/mL) allergen for 30 min at 37 °C in 5% CO2 and 95% air. Aliquot of the supernatant (50 μL) was then transferred into a 96 well plate and incubated with a substrate (1 μM p-nitrophenyl-N-acetyl-β-D-glucosaminide in 0.1 M citrate buffer, pH 4.5) in 1:1 ratio at 37 °C for 1 h. The reaction was quenched with the addition of 200 μL of stop solution (0.1 M Na2CO3/ NaHCO3, pH 10.0). The absorbance was measured at 405 nm using a microplate reader. Cells that were exposed to allergen were used as negative control. While cells that did not exposed to allergen were used as normal. The positive control used was ketotifen fumarate. The percentage inhibition was determined using the equation below.
Inhibitory effects on histamine and cytokines release
The inhibitory effects on histamine and cytokines release from RBL-2H3 cells were measured using the previously described methods [24, 25]. The sensitised cells were washed twice with 500 μL enriched medium and were reconstituted with 320 μL of enriched medium. After 10 min of incubation, the cells were treated with 40 μL of test samples (7.81, 15.62 and 31.25 μg/mL) or 40 μL of ketotifen fumarate (7.81, 15.62 and 31.25 μg/mL). For early-phase allergic reaction assay, the cells were stimulated using 40 μL of DNP-BSA (10 mg/mL) allergen for 30 min at 37 °C in 5% CO2 and 95% air. Aliquot of the supernatant (50 μL) was then centrifuged at 1000 x g at 8 °C for 20 min and the concentration of histamine released was measured using a Histamine ELISA Kit (Elabscience, China). For late-phase allergic reaction assay, the cells were incubated with 40 μL of DNP-BSA (10 mg/mL) allergen for 4 h at 37 °C in 5% CO2 and 95% air. The supernatant (100 μL) was centrifuged at 1000 x g at 8 °C for 20 min and the concentrations of IL-4 and TNF-α were measured using the respective ELISA kits (Elabscience, China).
Statistical analysis
The data obtained were reported as mean value ± standard error of mean (SEM) representing triplicate measurements (n = 3). The IC50 values with 95% confidence intervals were determined using a GraphPad Prism 5 software. Statistical significance of the data (p-value < 0.05) was analysed using one-way ANOVA with GraphPad Prism 5.
Results
Structure elucidation
The elution of hexane fraction of M. oleifera leaf extract yielded a new compound (compound 1). Compound 1 was obtained as white amorphous powder (34.2 mg) with HRESIMS molecular ionisation at m/z 235.1295 corresponding to [M + Na] +. Based on the value, chemical formula of the compound was deduced as C13H24O2 with a molecular weight of 212.33 g/mol. 1H and 13C NMR spectral data of the compound (Table 1) suggest the presence of a long chain skeleton with an olefinic signal at δ 5.41 (H-8, H-9) and one carbonyl signal at δ 174.13 (C-3). The carbonyl group was positioned at C-3 and the olefin at H-8 as evidenced by the 3J correlation measurement between the carbon at δ 24.9 (C-6) and deshielded methylene at δ 2.3 (H-4), and 3J correlation between the carbon at δ 24.9 (C-6) and olefin at δ 5.4 (H-8) (Fig. 1 and 2). 4J correlation between the neighbouring methylene at δ 2.02 (C-10) of olefin and terminal methyl at δ 0.85 (H-13) confirmed the position of olefin at C-8 and C-9. Although the olefin resonated as a multiplet, the compound was deduced to be in E conformation which was more stable compared to Z conformation. The spectroscopical data analysis suggest the identity of compound 1 as ethyl-(E)-undec-6-enoate.
Table 1.
1H NMR (CDCl3, 600 MHz) and 13C NMR (CDCl3, 151 MHz) spectral data of compound 1
| Position | δH (ppm) | δC(ppm) | HMBC |
|---|---|---|---|
| 1 | 1.25 (21H, m) | 14.3 | C-2 |
| 2 | 4.15 (2H, q, J = 7.1 Hz) | 60.1 | C-1, C-3 |
| 3 | – | 174.3 | – |
| 4 | 2.31 (2H, t, J = 7.7 Hz)) | 35.0 | C-3, C-5, C-6 |
| 5 | 1.60 (2H, m) | 29.1 | C-4 |
| 6 | 1.32 (21H, m) | 24.9 | C-5 |
| 7 | 2.79 (2H, m) | 35.0 | C-8 |
| 8 | 5.41 (1H, m) | 130.0 | C-6 |
| 9 | 5.41 (1H, m) | 130.0 | – |
| 10 | 2.02 (2H, m) | 31.9 | C-11, C-9 |
| 11 | 1.35 (21H, m) | 29.7 | C-13 |
| 12 | 1.25 (21H, m) | 2.8 | C-11 |
| 13 | 0.85 (4H, t, J = 7.1 Hz)) | 14.3 | C-11, C-12 |
Fig. 1.

Chemical structure of isolated compounds from M. oleifera. Ethyl-(E)-undec-6-enoate (1), 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), Quercetin (3), Kaempferol (4), β-sitosterol-3-O-glucoside (5), Oleic acid (6), Glucomoringin (7), Stigmasterol (9)
Fig. 2.
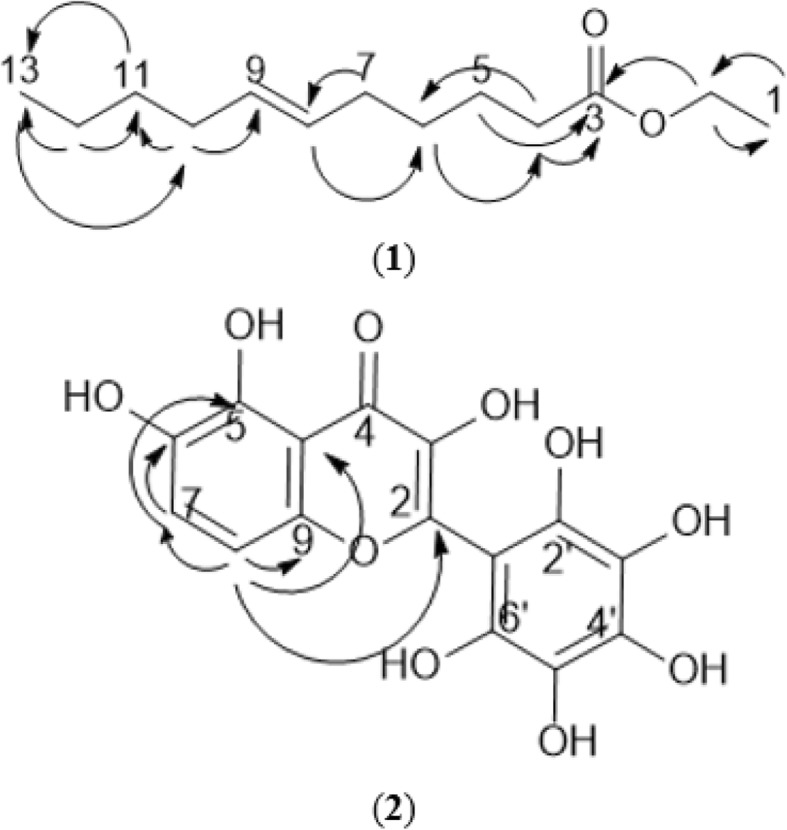
HMBC of new isolated compounds (1 and 2)
Compound 2 was obtained as reddish-brown crystals (3.9 mg) from the ethyl acetate fraction of M. oleifera leaves with HRESIMS molecular ionisation at m/z 176.9835 that correspond to [M + 2H]2+. Based on the MS data, the compound was deduced to have a molecular weight of 350.24 g/mol with a molecular formula of C15H10O10. From the NMR spectral data (Table 2), compound 2 was found to have a flavanol skeleton with eight hydroxylated carbons, one carbonyl carbon, five quaternary carbons and one methine. Only two doublets signals at δ 6.83 (H-7) and δ 7.89 (H-8) were observed on the 1H NMR spectrum. The doublets peaks were ortho-positioned to each other with J coupling value of 8.6 at A ring of the flavanol. 3J correlation between the doublets proved the ortho-position of the hydrogen. As no other hydrogen signals were observed, it was deduced that all carbons in ring B were attached to hydroxyl groups. In addition, 3J correlation between δ7.89 (H-8) and δ 168.9 (C-2) confirmed the position of the hydrogen at ring A of the flavanol structure. Based on the spectral data obtained, compound 2 was concluded to be 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one.
Table 2.
1H NMR (CD3OD, 600 MHz) and 13C NMR (CD3OD, 151 MHz) spectral data of compound 2
| Position | δH (ppm) | δc (ppm) | COSY | HSQC | HMBC |
|---|---|---|---|---|---|
| 2 | – | 168.9 | – | – | – |
| 3 | – | 138.5 | – | – | – |
| 4 | – | 173.8 | – | – | – |
| 5 | – | 162.9 | – | – | – |
| 6 | – | 123.0 | – | – | – |
| 7 | 6.83 (1H, d, J = 8.6 Hz) | 114.3 | H-8 | 114.3 | C-5, C-6, C-8 |
| 8 | 7.89 (1H, d, J = 8.7 Hz) | 131.4 | H-7 | 131.4 | C-2, C-7, C-9, C-10 |
| 9 | – | 161.8 | – | – | – |
| 10 | – | 113.3 | – | – | – |
| 1’ | – | 122.1 | – | – | – |
| 2’ | – | 147.5 | – | – | – |
| 3’ | – | 128.5 | – | – | – |
| 4’ | – | 138.5 | – | – | – |
| 5’ | – | 128.5 | – | – | – |
| 6’ | – | 147.5 | – | – | – |
Other seven isolated compounds were identified as quercetin (3) [26], kaempferol (4) [27, 28], β-sitosterol-3-O-glucoside (5) [29], oleic acid (6) [30], glucomoringin (7) [31], 2,3,4-trihydroxybenzaldehyde (8) [32] and stigmasterol (9) [33, 34] based on the MS and NMR comparison with the respective literatures (Fig. 1). 2,3,4-trihydroxybenzaldehyde (8) was not included in the bioassay study as the compound was isolated in minute quantity.
Cell viability
Cell viability assay of M. oleifera extracts and the isolated compounds at concentrations of 7.81, 15.62 and 31.25 μg/mL were found not cytotoxic towards the RBL-2H3 cells as presented in Table 3.
Table 3.
Cell viability of crude extracts on RBL-2H3 cells
| 7.81 μg/mL | 15.63 μg/mL |
31.25 μg/mL |
|
|---|---|---|---|
| M. oleifera leaves | 100.37 ± 1.61 | 101.65 ± 4.45 | 92.79 ± 6.50 |
| M. oleifera seed | 96.52 ± 5.50 | 103.22 ± 2.90 | 95.07 ± 1.77 |
| M. oleifera pod | 93.32 ± 13.17 | 95.72 ± 10.01 | 92.30 ± 5.18 |
| Ethyl-(E)–undec-6-enoate (MOLH 1) | 94.29 ± 3.73 | 90.78 ± 7.36 | 84.30 ± 9.38 |
| Quercetin (MOLE 1) | 93.96 ± 6.95 | 82.15 ± 19.95 | 85.66 ± 17.00 |
| Kaempferol (MOLE 2) | 108.82 ± 3.69 | 100.06 ± 9.25 | 100.18 ± 7.82 |
| 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (MOLE 3) | 95.36 ± 5.55 | 86.50 ± 3.68 | 83.15 ± 6.98 |
| β-sitosterol-3-O-glucoside (MOLA 1) | 69.54 ± 6.74 | 64.39 ± 6.29 | 62.60 ± 11.26 |
| Stigmasterol (MOPE 1) | 68.78 ± 2.32 | 54.39 ± 7.15 | 58.32 ± 5.99 |
| Oleic acid (MOSE 1) | 82.03 ± 3.24 | 66.95 ± 4.33 | 63.63 ± 3.45 |
| Glucomoringin (MOSA 1) | 77.87 ± 35.88 | 75.29 ± 10.24 | 58.63 ± 24.19 |
| Ketotifen fumarate (positive control) | 112.74 ± 22.95 | 100.81 ± 17.00 | 88.00 ± 3.13 |
Inhibitory effect of beta-hexosaminidase release
All extracts (Fig. 3a) and isolated compounds (Fig. 4a) significantly inhibited beta hexosaminidase release in a concentration dependent manner (7.81, 15.62 and 31.25 μg/mL) compared with the positive control, ketotifen fumarate (15.62 μg/mL). Amongst the extracts, the leaves exhibited significantly higher activity with IC50 value of 7.17 ± 1.69 μg/mL than ketotifen fumarate with IC50 value of 15.19 ± 1.10 μg/mL (Table 4). Four of the isolated compounds from the leaf extract, 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), quercetin (3), kaempferol (4) and β-sitosterol-3-O-glucoside (5) exhibited significantly higher inhibitory activity than ketotifen fumarate at varying concentrations of 15.62 and 31.25 μg/mL, and p-values (p < 0.01 – p < 0.001, Fig. 4a). Based on the IC50 values presented in Table 4, glucomoringin (7) (IC50: 10.43 ± 1.51 μM) and quercetin (3) (IC50: 19.07 ± 7.19 μM) were found to be significantly more active than ketotifen fumarate (IC50: 35.70 ± 2.59 μM).
Fig. 3.
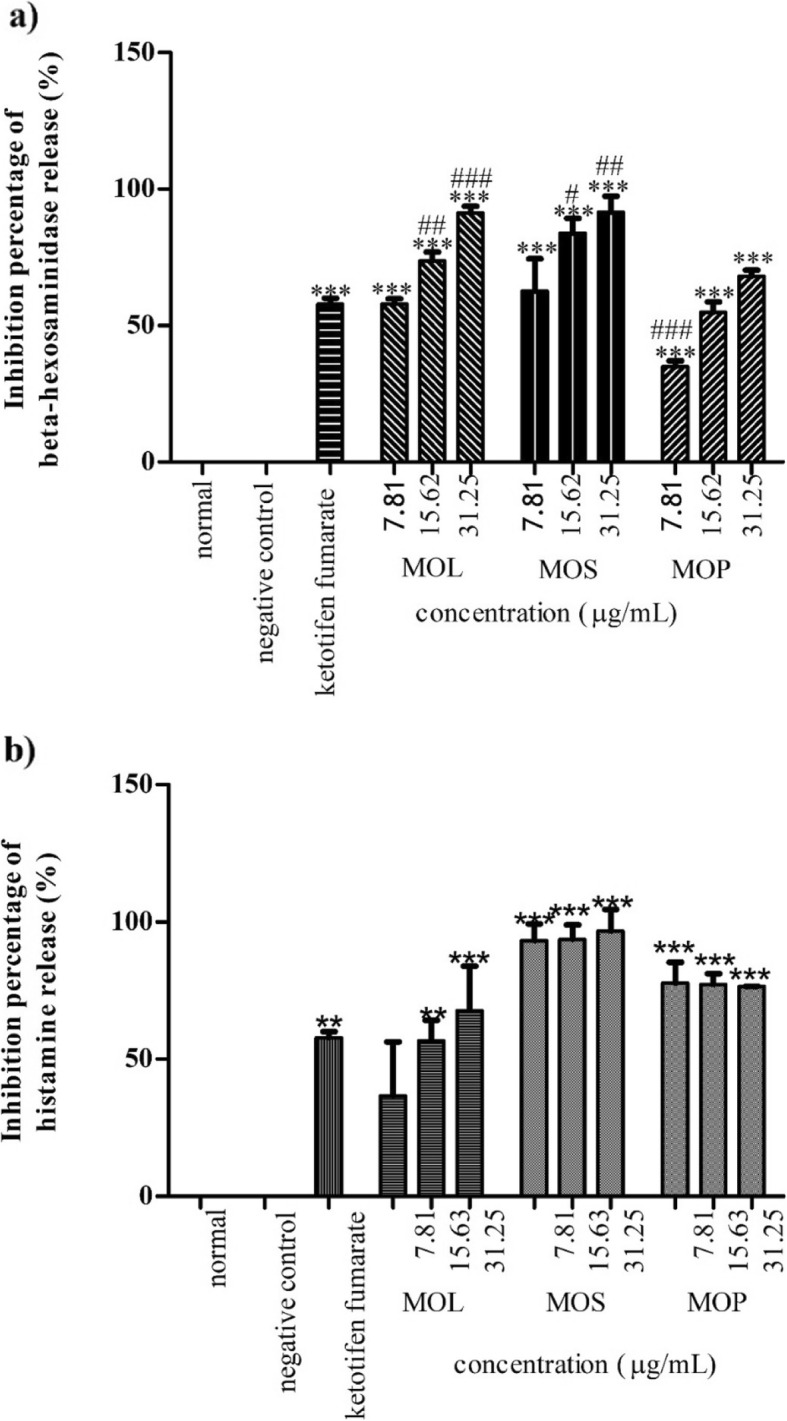
Percentage inhibition of M. oleifera crude extracts on a) beta-hexosaminidase and b) histamine releases. Data are presented as mean ± SEM (n = 3) with significant value of **p < 0.01, ***p < 0.001 as compared to positive control and #p < 0.05, ##p < 0.01, ###p < 0.001 as compared to ketotifen fumarate (15.62 μg/mL). MOL: M. oleifera leaves, MOS: M. oleifera seeds, MOP: M. oleifera pods
Fig. 4.

Percentage inhibition of M. oleifera test compounds on a) beta-hexosaminidase and b) histamine releases. Data are presented as mean ± SEM (n = 3) with significant value of *p < 0.05, **p < 0.01, ***p < 0.001 as compared to positive control and and #p < 0.05, ##p < 0.01, ###p < 0.001 as compared to ketotifen fumarate. Ethyl-(E)-undec-6-enoate (1), 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), Quercetin (3), Kaempferol (4), β-sitosterol-3-O-glucoside (5), Oleic acid (6), Glucomoringin (7), Stigmasterol (9)
Table 4.
IC50 values of inhibition towards beta-hexosaminidase and histamine release
| IC50 values of inhibition | ||
|---|---|---|
| Beta-hexosaminidase | Histamine | |
| M. oleifera leaf extract | 7.17 ± 1.69 μg/mL** | 11.66 ± 1.26 μg/mL |
| M. oleifera seed extract | 10.68 ± 0.63 μg/mL | 5.97 ± 0.84 μg/mL |
| M. oleifera pod extract | 14.89 ± 1.25 μg/mL | 7.43 ± 1.01 μg/mL |
| Ethyl-(E)–undec-6-enoate (1) | 82.68 ± 1.98 μM | 82.07 ± 13.75 μM |
| 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) | 17.70 ± 23.29 μM | 44.87 ± 2.13 μM |
| Quercetin (3) | 19.07 ± 7.19 μM* | 7.77 ± 27.86 μM |
| Kaempferol (4) | 29.39 ± 6.26 μM | 46.94 ± 10.26 μM |
| β-sitosterol-3-O-glucoside (5) | 24.93 ± 2.10 μM | – |
| Oleic acid (6) | 53.76 ± 8.95 μM | 56.05 ± 37.11 μM |
| Glucomoringin (7) | 10.43 ± 1.51 μM* | 27.22 ± 18.62 μM |
| Stigmasterol (9) | 75.92 ± 4.66 μM | 38.27 ± 24.48 μM |
| Ketotifen fumarate (positive control) |
15.19 ± 1.10 μg/mL (35.70 ± 2.59 μM) |
6.97 ± 0.04 μg/mL (16.38 ± 0.10 μM) |
Data are presented as mean ± SEM (n = 3) with significance values of *p < 0.05 and **p < 0.01 as compared to positive control.
Inhibitory effect of histamine release
All extracts significantly inhibited histamine release (Fig. 3b) compared with ketotifen fumarate. All isolated compounds, except β-sitosterol-3-O-glucoside (5), showed significant inhibitory activity at varying concentrations (Fig. 4b). M. oleifera seed extract had better activity with an IC50 value of 5.97 ± 0.84 μg/mL than ketotifen fumarate (IC50: 6.97 ± 0.04 μg/mL, Table 4). Amongst the isolated compounds, ethyl-(E)-undec-6-enoate (1), quercetin (3) and kaempferol (4) significantly inhibited histamine release in a concentration dependent manner (p < 0.01 – p < 0.001, Fig. 4b). Oleic acid (6) and glucomoringin (7) showed significant inhibitory activity at higher concentrations (15.62 and 31.25 μg/mL) (p < 0.05 – p < 0.001, Fig. 4b), whereas 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) and stigmasterol (9) significantly inhibited only at the highest concentration (31.25 μg/mL, p < 0.05, Fig. 4b). However, only quercetin (3) (IC50: 7.77 ± 27.86 μM) was found to be more active than ketotifen fumarate (IC50: 16.38 ± 0.10 μM) in inhibiting histamine release.
Inhibitory effect on cytokines release
Generally, all extracts (Fig. 5a) and the isolated compounds (Fig. 6a) inhibited IL-4 release at varying concentrations, compared with ketotifen fumarate. M. oleifera leaf and pod extracts showed higher inhibitory activity with IC50 values of 2.32 ± 0.08 and 2.74 ± 0.17 μg/mL, respectively, than ketotifen fumarate (IC50: 3.08 ± 0.49 μg/mL) (Table 5). All isolated compounds, except 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) and glucomoringin (7), exhibited higher inhibition percentage of IL-4 release at 31.25 μg/mL than ketotifen fumarate (Fig. 6a). However, based on the IC50 values (Table 5), β-sitosterol-3-O-glucoside (5) (IC50: 7.33 ± 1.76 μM) and glucomoringin (7) (IC50: 7.59 ± 0.39 μM) showed the most comparable inhibitory activity to ketotifen fumarate (IC50: 7.24 ± 1.16 μM).
Fig. 5.
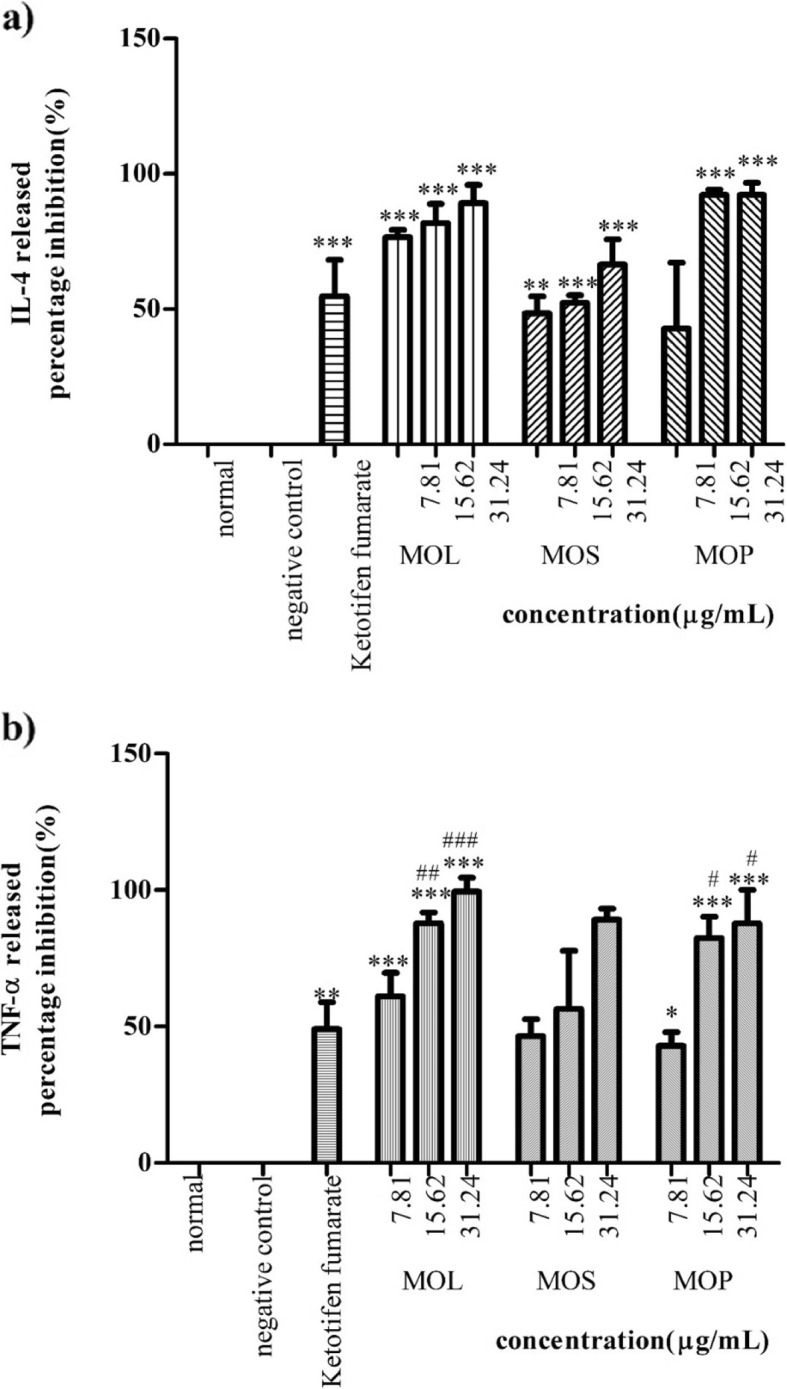
Percentage inhibition of M. oleifera crude extracts on a) IL-4 and b) TNF-α releases. Data are presented as mean ± SEM (n = 3) with significant value of *p < 0.05, **p < 0.01, ***p < 0.001 as compared to positive control and #p < 0.05, ##p < 0.01, ###p < 0.001 as compared to ketotifen fumarate (15.62 μg/mL). MOL: M. oleifera leaves, MOS: M. oleifera seeds, MOP: M. oleifera pods
Fig. 6.

Percentage inhibition of M. oleifera test compounds on a) IL-4 and b) TNF-α releases. Data are presented as mean ± SEM (n = 3) with significant value of *p < 0.05, **p < 0.01, ***p < 0.001 as compared to positive control and and #p < 0.05, ##p < 0.01 as compared to ketotifen fumarate. Ethyl-(E)-undec-6-enoate (1), 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), Quercetin (3), Kaempferol (4), β-sitosterol-3-O-glucoside (5), Oleic acid (6), Glucomoringin (7), Stigmasterol(9)
Table 5.
IC50 value of inhibition towards IL-4 and TNF-α release
| IC50 value of inhibition | ||
|---|---|---|
| IL-4 | TNF-α | |
| M. oleifera leaf extract | 2.32 ± 0.08 μg/mL | 1.20 ± 0.29 μg/mL |
| M. oleifera seed extract | 4.28 ± 0.05 μg/mL | 2.52 ± 0.33 μg/mL |
| M. oleifera pod extract | 2.74 ± 0.17 μg/mL | 2.52 ± 0.15 μg/mL |
| Ethyl-(E)–undec-6-enoate (1) | 16.91 ± 0.56 μM | 9.90 ± 2.31 μM |
| 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) | 12.17 ± 0.13 μM | 6.45 ± 0.26 μM |
| Quercetin (3) | 10.42 ± 0.07 μM | 6.24 ± 0.31 μM |
| Kaempferol (4) | 14.55 ± 3.24 μM | 8.73 ± 0.59 μM |
| β-sitosterol-3-O-glucoside (5) | 7.33 ± 1.76 μM | 5.26 ± 0.00 μM |
| Oleic acid (6) | 8.10 ± 0.26 μM | 10.11 ± 1.49 μM |
| Glucomoringin (7) | 7.59 ± 0.39 μM | 3.86 ± 8.26 μM |
| Stigmasterol (9) | 10.99 ± 1.57 μM | 6.05 ± 16.54 μM |
| Ketotifen fumarate (positive control) |
3.08 ± 0.49 μg/mL (7.24 ± 1.16 μM) |
2.91 ± 0.33 μg/mL (6.85 ± 0.77 μM) |
For the inhibitory activity of TNF-α release, the extracts of leaves, seeds and pods were found to be more active with IC50 values of 1.20 ± 0.29, 2.52 ± 0.33 and 2.52 ± 0.15 μg/mL, respectively, than ketotifen fumarate (IC50: 2.91 ± 0.33 μg/mL) (Table 5). Nonetheless, all the isolated compounds also showed higher percentage of inhibitory activity at concentrations of 15.62 and 31.25 μg/mL compared with ketotifen fumarate (Fig. 6b). However, glucomoringin (7) (IC50: 3.86 ± 8.26 μM) and β-sitosterol-3-O-glucoside (5) (IC50: 5.26 ± 0.00 μM) were found to be even more active than ketotifen fumarate (IC50: 6.85 ± 0.77 μM) (Table 5).
Discussion
Moringa oleifera is a commonly used plant that has high nutritional and medicinal values. However, scientific report on its anti-allergic property to support the traditional use is still inadequate. High content of flavonoids and flavanol glucosides in M. oleifera such as quercetin, astragalin, kaempferol and rutin could be associated with the anti-allergic potential of the plant [10].
From this study, nine compounds were isolated from the M. oleifera ethanol (80%) extracts of various plant parts. Ethyl-(E)-2-undec-6-enoate (1), 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), quercetin (3), kaempferol (4) and β-sitosterol-3-O-glucoside (5) were isolated from the leaves extract. Oleic acid (6) and glucomoringin (7) were isolated from the seeds extract; while 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), β-sitosterol-3-O-glucoside (5), 2,3,4-trihydroxybenzaldehyde (8) and stigmasterol (9) were isolated from the pod extract. Amongst the compounds isolated, ethyl-(E)-2-undec-6-enoate (1) and 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) were new compounds isolated in natural product chemistry, whereas 2,3,4-trihydroxybenzaldehyde (8) is the first to be reported of its presence in Moringaceae species.
RBL-2H3 cells derived from rat basophils were used to determine the allergic responses in vitro. Basophils have similarities with mast cells based on the composition of the granules and reactions towards allergens [35]. The mast cell degranulation was measured using beta-hexosaminidase and histamine as markers [36]. Upon exposure, the allergens will crosslink with the high-affinity IgE receptor to form IgE-FcƐRI complex triggering the basophils to degranulate and release mediators such as beta-hexosaminidase and histamine [19]. Histamine is a well-known mediator to induce bronchoconstriction and vasodilation in an allergic reaction. However, no significant involvement of beta-hexosaminidase is reported on allergic responses [37].
Early phase of an allergic reaction occurs minutes after allergen sensitisation, releasing histamine and other mediators [19]. Whereas, late phase of the allergic reaction is recognized by the release of cytokines such as IL-13, IL-4, IL-9, IL-5, IFN-γ and TNF-α after 2 to 6 h of sensitisation. The most pivotal cytokines in an allergic reaction are IL-4 and TNF-α. In the presence of IL-4, it will trigger B cell activation and the production of IgE. High amount of IgE will increase the binding site of the allergen on the mast cells, hence induce allergic reactions. TNF-α is responsible for the recruitment of inflammatory cells such as eosinophils, neutrophils, monocytes and macrophages to the site of allergen invasion. This will lead to inflammatory-related allergic responses.
In this study, M. oleifera leaf, seed and pod extracts exhibited an anti-allergic activity by inhibiting the early and late phases of allergic reaction. The leaves extract particularly suppressed the release of beta-hexosaminidase, IL-4 and TNF-α more than the positive control, ketotifen fumarate. Nevertheless, it also showed better activity in the late phase of allergic reaction (Table 5). Previous studies also found that this plant suppressed TNF-α level of atopic dermatitis of human keratinocyte cells [38] and LPS-mediated RAW 264.7 macrophage cells [39]. 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), quercetin (3), kaempferol (4) and β-sitosterol-3-O-glucoside (5) isolated from the leaves extract inhibited beta-hexosaminidase more actively than ketotifen fumarate. In the early phase of allergic reaction, quercetin (3) was found to be more potent than ketotifen fumarate in inhibiting histamine release. In the late phase, 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2) and quercetin (3) inhibited release of TNF-α, while β-sitosterol-3-O-glucoside (5) inhibited both IL-4 and TNF-α release more actively than ketotifen fumarate. Flavonoids were reported to have anti-allergic properties by inhibiting histamine and IL-4 release [21]. Three flavonoids isolated from the leaves extract; 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), quercetin (3) and kaempferol (4), might be responsible for the anti-allergic property due to their ability to stabilise mast cell from degranulation and inhibit the release of IL-4 and TNF-α from IgE-mediated RBL-2H3 cells. Quercetin (3) and kaempferol (4) have been reported to inhibit mast cells from degranulation by suppressing the calcium influx from extracellular medium [40, 41]. The specific pathway of inhibition was reported to be via the Syk pathway that attenuated the ERK activation and hence inhibited the release of IL-4 and TNF-α release [42, 43]. In addition, quercetin (3) and kaempferol (4) also inhibited PLCγ and PKC pathways of the IgE-mediated allergic reaction by reducing the expression of 5-lipoxygenase, cyclooxygenase-2 and the production of leukotriene B4 and prostaglandin E2 [43]. An in-vivo study showed that quercetin (3) and kaempferol (4) reduced mast-cell count of passive cutaneous anaphylaxis in mice [43]. From a structural activity relationship study, higher number of hydroxyls on ring B of flavanol was found to increase the inhibitory activity of histamine release in RBL-2H3 cells [24]. Quercetin (3) with two hydroxyl groups (3′4’-trihydroxy) at the flavanol ring B has higher activity than kaempferol (4) with one hydroxyl group (4′-hydroxy group). In contrast, the current study proposed that flavanol with saturated hydroxy group on ring B of 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen4-one (2) decreased the inhibitory activity of the compound.
The findings from this study showed that the extract of M. oleifera pods had higher inhibitory activity against beta-hexosaminidase and at the late phase of allergic reaction than ketotifen fumarate. The activity could be associated with its isolated compounds; 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), β-sitosterol-3-O-glucoside (5) and stigmasterol (9). Stigmasterol (9) only inhibited TNF-α release in RBL-2H3 cells more significantly than ketotifen fumarate. This is supported by the fact that stigmasterol (9) did not inhibit beta-hexosaminidase release from A23187-induced RBL-2H3 cells [44]. However, the compound was found to inhibit scratching and mast cell trafficking in compound 48/80-induced ICR mice [45]. It also decreased TNF-α concentration, ear skin oedema and neutrophil count of Wistar rats induced by 12-O-tetradecanoylphorbol-13-acetate (TPA). With an addition of a sugar moeity on the sterol, β-sitosterol-3-O-glucoside (5) inhibited beta-hexosaminidase, IL-4 and TNF-α release but did not show any significant activity on histamine release. β-sitosterol-3-O-glucoside (5) might directly inhibit beta-hexosaminidase activity instead of inhibiting mast cell degranulation. In previous studies, β-sitosterol-3-O-glucoside (5) showed only 7.0% reduction of beta-hexosaminidase release from A23187-induced RBL-2H3 cells [44] and reduced inflammation of xylene-induced oedema in mice [46].
From this study, the M. oleifera seeds extract inhibited the release of beta-hexosaminidase, histamine and TNF-α more actively than ketotifen fumarate. It could be attributed to the presence of the isolated compound, glucomoringin (7). To the best of the authors’ knowledge, this is the first report of glucomoringin (7) and its effect on mast cell stabilising activity and its potential use in inhibiting the late phase of allergic response.
Long chain-containing chemical structure as seen in ethyl-(E)-undec-6-enoate (1) and oleic acid (6) exhibited the weakest mast cell stabilizing activity. Oleic acid (6) was reported to weakly inhibit histamine and beta-hexosaminidase release by antigen- and A23187-induced RBL-2H3 cells [47, 48]. In addition, oleic acid only slightly increased intracellular Ca2+ that induced cell degranulation [49]. In an in vivo study, peanuts containing high oleic acid was also found to be less allergenic than normal peanuts [50].
Conclusion
The leaves extract exhibited the highest inhibition against the release of beta-hexosaminidase, IL-4 and TNF-α, while the seed extract inhibited the histamine release the most. Further study revealed that isolated compound such as glucomoringin (7) displayed the highest inhibitory activity on beta-hexosaminidase and TNF-α release. Meanwhile, quercetin (3) was the most potent against histamine release inhibition while, β-sitosterol-3-O-glucoside (5) exhibited the highest inhibition on IL-4 release. Overall, this study suggested the anti-allergic property of ethanol (80%) extract of M. oleifera leaves, seeds and pods, as well as the isolated compounds by stabilising mast cell from degranulation and inhibiting the early and late phases of allergic reaction in anti-DNP IgE sensitised RBL-2H3 cells induced by DNP-BSA. Two new compounds; ethyl-(E)-2-undec-6-enoate (1) and 3,5,6-trihydroxy-2-(2,3,4,5,6-pentahydroxyphenyl)-4H-chromen-4-one (2), were isolated from the leaves extract. However, only the latter compound was active against the beta-hexosaminidase and TNF-α release.
Acknowledgements
The authors would like to thank the Center for Research and Instrumentation (CRIM), Universiti Kebangsaan Malaysia for performing the NMR and ESIMS analyses, and staff of Faculty of Pharmacy, Universiti Kebangsaan Malaysia for the technical assistance given throughout the study.
Authors’ contributions
KH, EK, MJ, JAJ and LKW designed the study and revised the manuscript. NZAR performed the study, conducted data collection and prepared the manuscript. All the authors have read and approved the manuscript.
Funding
The study was funded by the Ministry of Agriculture and Agro-Based Industry, Malaysia with a grant number of NH1015D074. The funding body did not contribute to the design of the study and collection, analysis, interpretation of data and in writing the manuscript.
Availability of data and materials
All-important data regarding the study is included in this published article and any supplementary data is available from the corresponding author upon request.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declared that there is no conflict of interest regarding the publication of this research paper.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Nur Zahirah Abd Rani, Email: zahira.rani@gmail.com.
Endang Kumolosasi, Email: e_kumolosasi@ukm.edu.my.
Malina Jasamai, Email: malina@ukm.edu.my.
Jamia Azdina Jamal, Email: jamia@ukm.edu.my.
Kok Wai Lam, Email: david_lam@ukm.edu.my.
Khairana Husain, Email: khairana@ukm.edu.my.
References
- 1.Fahey JW. Moringa oleifera: A Review of the Medical Evidence for Its Nutritional, Therapeutic, and Prophylactic Properties. Part 1. Trees for Life. 2005;1.
- 2.Leone Alessandro, Spada Alberto, Battezzati Alberto, Schiraldi Alberto, Aristil Junior, Bertoli Simona. Moringa oleifera Seeds and Oil: Characteristics and Uses for Human Health. International Journal of Molecular Sciences. 2016;17(12):2141. doi: 10.3390/ijms17122141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mahmood KT, Mugal T. Haq IU. Natl Gift- Rev J Pharm Sci Res. 2010;2:775–781. [Google Scholar]
- 4.Hossain MK, Sturm B, Quadt A, Hensel O. A Sustainable Natural Resource (Moringa oleifera) in Tropical and Subtropical Areas: An Intensive Literature Review. International Research on Food Security, Natural Resource Management and Rural Development; Bonn, Germany 2017.
- 5.Csurhes S, Navie S. Invasive plant risk assessment: Moringa oleifera. In: Queensland DoAaFB, editor. Australia: State of Queensland; 2010. [Google Scholar]
- 6.Mathur BS. Moringa Book. St. Louis, MI: Trees for Life International; 2005.
- 7.Sanchez-Machado DI, Nunez-Gastelum JA, Reyes-Moreno C, Ramirez-Wong B, Lopez-Cervantes J. Nutritional quality of edible parts of Moringa oleifera. Food Anal Methods. 2010;3:175–180. doi: 10.1007/s12161-009-9106-z. [DOI] [Google Scholar]
- 8.Leone A, Fiorillo G, Criscuoli F, Ravasenghi S, Santagostini L, Fico G, et al. Nutritional characterization and phenolic profiling of Moringa oleifera leaves grown in Chad. Sahrawi Refugee Camps, and Haiti International Journal of Molecular Sciences. 2015;16:18923–18937. doi: 10.3390/ijms160818923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Thurber MD, Fahey JW. Adoption of Moringa oleifera to combat under-nutrition viewed through the lens of the “diffusion of innovations” theory. Ecol Food Nutr. 2009;48:212–225. doi: 10.1080/03670240902794598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Abd Rani NZ, Husain K, Kumolosasi E. Moringa genus: a review of phytochemistry and pharmacology. Frontier Pharmacology. 2018;9:26. doi: 10.3389/fphar.2018.00026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Walter A, Samuel W, Peter A, Jospeh O. Antibacterial activity of Moringa oleifera and Moringa stenopetala methanol and n-hexane seed extracts on bacteria implicated in waterborne diseases. Afr J Microbiol Res. 2011;5:153–157. [Google Scholar]
- 12.Elgamily H, Moussa A, Elboraey A, El-Sayed H, Al-Moghazy M, Abdalla A. Microbiological assessment of Moringa oleifera extracts and its incorporation in novel dental remedies against some oral pathogens. Open Access Macedonian Journal of Medical Sciences. 2016;4:585–590. doi: 10.3889/oamjms.2016.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rahman MM, Rahman MM, Akhter S, Jamal MA, Pandeya DR, Haque MA. Control of coliform bacteria detected from diarrhea associated patients by extracts of Moringa oleifera. Nepal Medical College journal. 2010;12. [PubMed]
- 14.Chuang PH, Lee CW, Chou JY, Murugan M, Shieh BJ, Chen HM. Anti-fungal activity of crude extracts and essential oil of Moringa oleifera lam. Bioresour Technol. 2007;98:232–236. doi: 10.1016/j.biortech.2005.11.003. [DOI] [PubMed] [Google Scholar]
- 15.Galuppo M, Giacoppo S, De Nicola GR, Iori R, Navarra M, Lombardo GE. Antiinflammatory activity of glucomoringin isothiocyanate in a mouse model of experimental autoimmune encephalomyelitis. Fitoterapia. 2014;95:160–174. doi: 10.1016/j.fitote.2014.03.018. [DOI] [PubMed] [Google Scholar]
- 16.Cheenpracha S, Park EJ, Yoshida WY, Barit C, Wall M, Pezzuto JM. Potential anti-inflammatory phenolic glycosides from the medicinal plant Moringa oleifera fruits. Bioorg Med Chem. 2010;18:6598–6602. doi: 10.1016/j.bmc.2010.03.057. [DOI] [PubMed] [Google Scholar]
- 17.Waterman C, Rojas-Silva P, Tumer TB, Kuhn P, Richard AJ, Wicks S. Isothiocyanate-rich Moringa oleifera extract reduces weigh gain, insulin resistance and hepatic gluconeogenesis in mice. Mol Nutr Food Res. 2015;59:1013–1024. doi: 10.1002/mnfr.201400679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chin CY, Jalil J, Ng PY, Ng SF. Development and formulation of Moringa oleifera standardised leaf extract film dressing for wound healing application. J Ethnopharmacol. 2018;212:188–199. doi: 10.1016/j.jep.2017.10.016. [DOI] [PubMed] [Google Scholar]
- 19.Akdis CA, Agache I. Global Atlas of Allergy: European Academy of Allergy and Clinical Immunology; 2014.
- 20.Chopra RN, Chopra IC, Handa KL, Kapur ID. Chopra's indigenous drugs of India. UN Dhur and Sons: Calcutta, India; 1938. [Google Scholar]
- 21.Kawai M, Hirano T, Higa S, Arimitsu J, Maruta M, Kuwahara Y, et al. Flavanoids and related compounds as anti-allergic substances. Allergol Int. 2007;56:113–123. doi: 10.2332/allergolint.R-06-135. [DOI] [PubMed] [Google Scholar]
- 22.Pinho BR, Sousa C, Valentao P, Oliveira JMA, Andrade PB. Modulation of basophils’ degranulation and AllergyRelated enzymes by monomeric and Dimeric Naphthoquinones. PLoS One. 2014;9:1–10. doi: 10.1371/journal.pone.0090122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shahari MS, Husain K, Kumolosasi E, Rajab NF. Histamine and Beta-Hexosaminidase inhibitory effects of crude alkaloid from Kopsia arborea Blume in RBL-2H3 cell lines. Nat Prod: Ind J. 2017;13.
- 24.Mastuda H, Morikawa T, Ueda K, Managi H, Yoshikawa M. Structural requirements of flavonoids for inhibition of antigen-induced degranulation, TNF-α and IL-4 production from RBL-2H3 cells. Bioorg Med Chem. 2002;10:3123–3128. doi: 10.1016/S0968-0896(02)00227-4. [DOI] [PubMed] [Google Scholar]
- 25.Itoh T, Ohguchi K, Iinuma M, Nozawa Y, Akao Y. Inhibitory effect of xanthones isolated from the pericarp of Garcinia mangostana L. on rat basophilic leukemia RBL-2H3 cell degranulation. Bioorg Med Chem. 2008;16:4500–4508. doi: 10.1016/j.bmc.2008.02.054. [DOI] [PubMed] [Google Scholar]
- 26.Husain K, Zakaria SM, Lajis NH, Shaari K, Ismail IS, Israf DA, et al. Novel sesquiterpene and copyrine alkaloids from Anaxagorea javanica Blume. Phytochem Lett. 2012;5:788–702. doi: 10.1016/j.phytol.2012.09.003. [DOI] [Google Scholar]
- 27.Cornelius MTF, de Carvalho MG, da Silva TMS, Alves CCF, Siston APN, Alves KZ, et al. Other Chemical Constituents Isolated from Solanum crinitum Lam. (Solanaceae) J Braz Chem Soc. 2010;21:2211–2219. doi: 10.1590/S0103-50532010001200007. [DOI] [Google Scholar]
- 28.Wahab A, Tahira, Begum S, Ayub A, Mahmood I, Mahmood T, et al. Luteolin and kaempferol from Cassia alata, antimicrobial and antioxidant activity of its methanolic extracts. FUUAST Journal Biology. 2014;4:1–5. [Google Scholar]
- 29.Mouffok S, Haba H, Lavaud C, Long C, Benkhaled M. Chemical constituents of Centaurea omphalotricha Coss. & Durieu ex Batt. & Trab. Rec Nat Prod. 2012;6:292–295. [Google Scholar]
- 30.Abe M, Ito Y, Suzuki A, Onoue S, Noguchi H, Yamada S. Isolation and pharmacological characterization of fatty acids from saw palmetto extract. Anal Sci. 2009;25:553–557. doi: 10.2116/analsci.25.553. [DOI] [PubMed] [Google Scholar]
- 31.de Graaf RM, Krosse S, Swolfs AE, te Brinke E, Prill N, Leimu R, et al. Isolation and identification of 4-α-rhamnosyloxy benzyl glucosinolate in Noccaea caerulescens showing intraspecific variation. Phytochemistry. 2015;110:166–171. doi: 10.1016/j.phytochem.2014.11.016. [DOI] [PubMed] [Google Scholar]
- 32.Mulabagal V, Alexander-Lindo RL, Dewitt DL, Nair MG. Health-beneficial phenolic aldehyde in Antigonon leptopus tea. Evid Based Complement Alternat Med. 2011;2011:6. doi: 10.1093/ecam/nep041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mawa Shukranul, Jantan Ibrahim, Husain Khairana. Isolation of Terpenoids from the Stem of Ficus aurantiaca Griff and their Effects on Reactive Oxygen Species Production and Chemotactic Activity of Neutrophils. Molecules. 2016;21(1):9. doi: 10.3390/molecules21010009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Husain K, Jamal JA, Jalil J. Phytochemical study of Cananga odorata Hook, F. & Thoms (Annonaceae) Int J Pharm Pharm Sci. 2012;4:465–467. [Google Scholar]
- 35.Miyake K, Karasuyama H. Emerging roles of basophils in allergic inflammation. Allergol Int. 2017;66:382–391. doi: 10.1016/j.alit.2017.04.007. [DOI] [PubMed] [Google Scholar]
- 36.Tewtrakul S, Itharat A. Anti-allergic substances from the rhizomes of Dioscorea membranacea. Bioorg Med Chem. 2006;14:8707–8711. doi: 10.1016/j.bmc.2006.08.012. [DOI] [PubMed] [Google Scholar]
- 37.Fukuishi N, Murakami S, Ohno A, Yamanaka N, Matsui N, Fukutsuji K, et al. Does β-hexosaminidase function only as a degranulation indicator in mast cells? The primary role of β-hexosaminidase in mast cell granules. J Immunol. 2014;193:1886–1894. doi: 10.4049/jimmunol.1302520. [DOI] [PubMed] [Google Scholar]
- 38.Choi EJ, Debnath T, Tang Y, Ryu YB, Moon SH, Kim EK. Topical application of Moringa oleifera leaf extract ameliorates experimentally induced atopic dermatitis by the regulation of Th1/Th2/Th17 balance. Biomed Pharmacother. 2016;84:870–877. doi: 10.1016/j.biopha.2016.09.085. [DOI] [PubMed] [Google Scholar]
- 39.Muangnoi C, Chingsuwanrote P, Praengamthanachoti P, Svasti S, Tuntipopipat S. Moringa oleifera pod inhibits inflammatory mediator production by lipopolysaccharide-stimulated RAW 264.7 murine macrophage cell lines. Inflamm. 2012;35:445–455. doi: 10.1007/s10753-011-9334-4. [DOI] [PubMed] [Google Scholar]
- 40.Itoh T, Ninomiya M, Yasuda M, Koshikawa K, Deyashiki Y, Nozawa Y, et al. Inhibitory effects of flavonoids isolated from Fragaria ananassa Duch on IgE-mediated degranulation in rat basophilic leukemia RBL-2H3. Bioorg Med Chem. 2009;17:5374–5379. doi: 10.1016/j.bmc.2009.06.050. [DOI] [PubMed] [Google Scholar]
- 41.Mlcek Jiri, Jurikova Tunde, Skrovankova Sona, Sochor Jiri. Quercetin and Its Anti-Allergic Immune Response. Molecules. 2016;21(5):623. doi: 10.3390/molecules21050623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee EJ, Ji GE, Sung MK. Quercetin and kaempferol suppress immunoglobulin E-mediated allergic inflammation in RBL-2H3 and Caco-2 cells. Inflamm Res. 2010;59:847–854. doi: 10.1007/s00011-010-0196-2. [DOI] [PubMed] [Google Scholar]
- 43.Kim M, Lim SJ, Kang SW, Um BH, Nho CW. Aceriphyllum rossii extract and its active compounds, Quercetin and Kaempferol inhibit IgE-mediated mast cell activation and passive cutaneous anaphylaxis. J Agric Food Chem. 2014;62:3750–3758. doi: 10.1021/jf405486c. [DOI] [PubMed] [Google Scholar]
- 44.Thabet AA, Youssef FS, Korinek M, Chang FR, Wu YC, Chen BH, et al. Study of the anti-allergic and anti-inflammatory activity of Brachychiton rupestris and Brachychiton discolor leaves (Malvaceae) using in vitro models. BMC Complement Altern Med. 2018;18. [DOI] [PMC free article] [PubMed]
- 45.Antwi AO, Obiri DD, Osafo N, Essel LB, Forkuo AD, Atobiga C. Stigmasterol alleviates cutaneous allergic responses in rodents. Biomed Res Int. 2018;2018:1–13. doi: 10.1155/2018/3984068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Huang LJ, Gao WY, Li X, Zhao WS, Huang LQ, Liu CX. Evaluation of the in Vivo anti-inflammatory effects of extracts from Pyrus bretschneideri Rehd. J Agric Food Chem. 2010;58:8983–8987. doi: 10.1021/jf101390q. [DOI] [PubMed] [Google Scholar]
- 47.Teshima R, Amano F, Nakamura R, Tanaka Y, Sawada J. Effects of polyunsaturated fatty acids on calcium response and degranulation from RBL-2H3 cells. Int Immunopharmacol. 2007;7:205–210. doi: 10.1016/j.intimp.2006.09.017. [DOI] [PubMed] [Google Scholar]
- 48.Korinek M, Tsai Y-H, El-Shazly M, Lai K-H, Backlund A, Wu S-F, et al. Anti-allergic Hydroxy fatty acids from Typhonium blumei explored through ChemGPS-NP. Front Pharmacol. 2017;8. [DOI] [PMC free article] [PubMed]
- 49.Kim MC, Kim MG, Jo YS, Song HS, Eom TI, Sim SS. Effects of C18 fatty acids on intracellular Ca2+ mobilization and histamine release in RBL-2H3 cells. Korean J Physiol Pharmacol. 2014;18:241–247. doi: 10.4196/kjpp.2014.18.3.241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Nguyen TTH, Sakurai H, Miyahara M, Yoshihashi T, Uematsu T, Yamaki T, et al. Comparative study on allergenic properties of high-oleic and conventional peanuts. Pol J Food Nutr Sci. 2003;12:88–95. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All-important data regarding the study is included in this published article and any supplementary data is available from the corresponding author upon request.