
Figure 3. Heart-specific transcriptional profiling of DM1 flies identifies deregulation of genes controlling cellular calcium level.
(A) Venn diagrams of genes deregulated in Hand>UPRT;MblRNAi and in Hand>UPRT;Bru-3 contexts (FC > 1.7) followed by heatmap of commonly deregulated genes. (B) Genemania interaction network of conserved candidates including stj, Rgk2, Syn1 and inaD known to be involved in Ca2 regulation. A scheme presenting the structure of the voltage-gated calcium channel and its regulatory component Stj/α2δ3 is included. Color code in genemania network represents up and down regulation according to the heatmap. Cyan/Blue intensity code indicates fold change of genes expression in the heatmap.
Figure 3—source data 1. A list of Drosophila genes and their Human orthologs deregulated in both Hand>Bru-3 and Hand>MblRNAi contexts.
Orthology scores were obtained from DIOPT tool (DRSC Integrative Ortholog Prediction Tool) (Hu et al., 2011) which predicts orthologs, based on sequence homology, phylogenetic trees, and functional similarity (www.flyrnai.org/cgi-bin/DRSC_orthologs.pl).
elife-51114-fig3-data1.xlsx (31.1KB, xlsx)
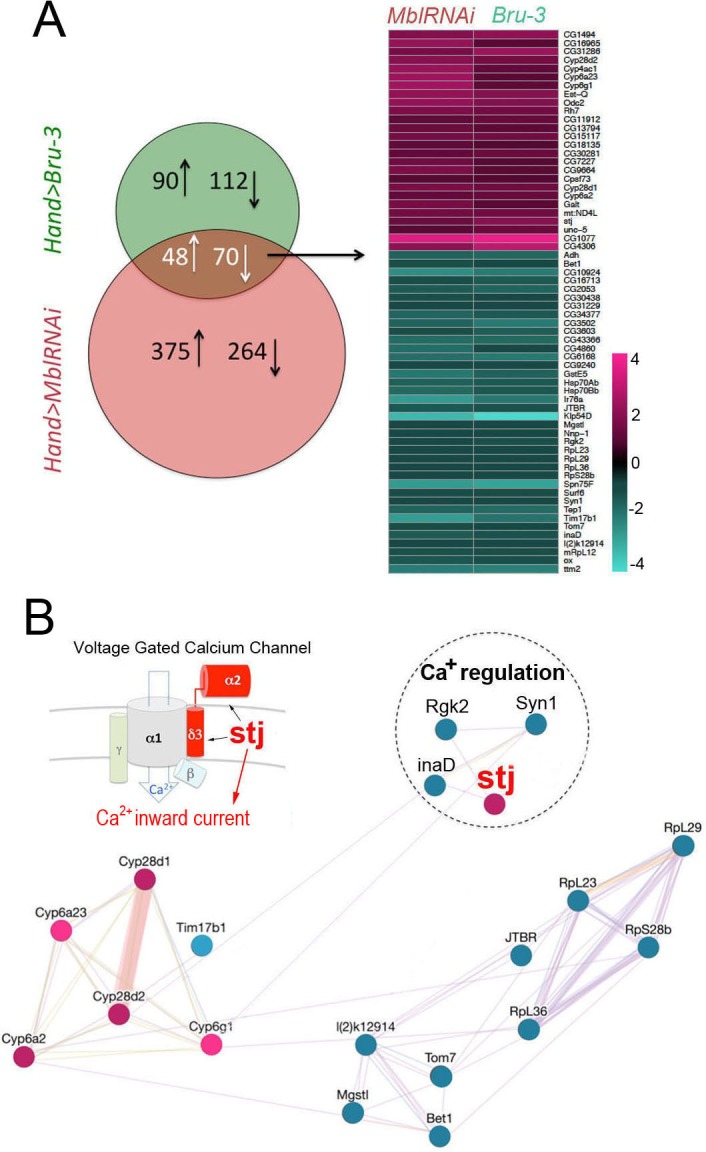
Figure 3—figure supplement 1. Attenuation of inaD, Syn1 and Rgk2 all lead to an increase of diastolic (DD) and systolic (SD) heart diameters.

For each candidate gene effects of attenuation were tested using two different UAS-RNAi lines crossed with Hand-Gal4 driver. More than 20 flies for each genotype were analysed by SOHA. Statistical significance of DD and SD changes was assessed using one-way ANOVA, Kruskal-Wallis Dunn’s multiple comparison post-test (* - p<0.05; ** - p<0.01; *** - p<0.001).
Figure 3—figure supplement 2. The expression levels of CG42617 and CG16868, two additional α2δ protein-coding genes are not affected in the heart of DM1 fly models.

Normalized RNAseq IGV tracks in control (Hand>lacZ) and in pathogenic DM1 contexts (Hand>MblRNAi and Hand>Bru-3) are shown aligned with genomic exon/intron organization of CG42617 (A) and CG16868 (B).