

Abstract
Myelodysplastic syndrome (MDS) represents a heterogeneous hematopoietic disorder in which mature blood cells are derived from an abnormal multipotent progenitor cell. The current therapy for MDS involves repeated cycles of DNA methyltransferase (DNMT) inhibitors, particularly the demethylation drug 5-azacytidine (5-azaC) which has been shown to increase the survival of patients with high-risk MDS. The mechanisms behind the therapeutic effects of 5-azaC are not yet clear. In this study the effect of 5-azaC on the development of regulatory T cells (Tregs) and T-helper 17 (Th17) cells was investigated. The numbers of CD4+ T-cell subsets in 30 patients with intermediate-2/high-risk MDS were serially assessed at diagnosis and following 5-azaC treatment. The number of FoxP3+ Tregs was significantly higher after 3 months of therapy. However, there was no statistical difference in the number of Th17 cells following treatment. In vitro, 5-azaC enhanced the overall proportion of Tregs, but not Th17, in CD4+ T cells from patients with MDS. Addition of 5-azaC reduced the proliferative capacity of Tregs, suggesting that the increase in Tregs was due to conversion of conventional CD25− cells, rather than proliferation of CD25+FoxP3+ cells. The FoxP3 expression in 5-azaC-treated T effectors was also increased. Interestingly, while Tbet and RORγT mRNA transcription had no obvious changes, due to the demethylation of the FoxP3 promoter, these findings are important in associating the induction of DNA hypomethylation and the clinical response to 5-azaC.
Keywords: myelodysplastic syndrome, 5-azacytidine, regulatory T cells, T-helper 17 cells
Introduction
Treatment with demethylating agents, such as 5-azacytidine (5-azaC), not only prolongs the survival of patients with intermediate 2 and high-risk myelodysplastic syndrome (MDS) (1,2), but also leads to improvements in acute myeloid leukemia (AML) and multiple myeloma (MM) (3–5). However, the mechanisms behind the therapeutic effects of 5-azaC are not yet clear. No definite evidence exists that the clinical response is related to the induction and magnitude of DNA hypomethylation (6).
A number of studies have pointed out the immunoregulatory effects of 5-azaC, which are proven to be important in the pathogenesis of MDS (7), though the mechanisms that underlie these changes are not fully understood. 5-azaC treatment has been shown to augment regulatory T cells (Tregs) after bone marrow transplants, and thereby, to alleviate the graft-versus-host disease (GvHD) (8). 5-azaC may expand Tregs and contribute to the management of GvHD after allogeneic bone marrow transplantation (8). However, 5-azaC or decitabine have been shown to possess an intact graft-versus-leukemia (GVL) effect in transplanted mice, with an increase in the number of FoxP3+ T cells (9). In clinical practice, high-risk MDS patients benefit from 5-azaC treatment with significant prolongation of survival. For these reasons our study focused on 5-azaC in the development of CD4+ T cells, particularly Tregs and T-helper 17 (Th17).
Previous studies have shown that the number of Tregs is significantly increased in high-risk and intermediate-2 MDS, whereas in ‘low-risk’ MDS interleukin (IL)-17-producing CD4+ T cells (Th17 cells) is increased, suggesting a correlation between the number of Tregs and Th17 with the severity of the disease (10,11). The promoter of FoxP3, a key gene for the function of Tregs, is methylated in conventional CD4+CD25− T cells (12). Treatment with DNA methyltransferase (DNMT) inhibitors induces conversion of conventional CD4+CD25− T cells into CD25+FoxP3+ Tregs. This suggests that the expansion of Tregs is associated with increased FoxP3 expression due to FoxP3 promoter demethylation.
In order to understand the effects of 5-azaC on CD4+ T cells, the serial peripheral blood and bone marrow samples from patients with intermediate-2/high-risk MDS were analyzed, including the number profile of T cells prior to and following 5-azaC treatment. The effect of 5-azaC on the expression and methylation status of FoxP3 in vitro was also investigated.
Patients and methods
Patient samples
Thirty patients with intermediate-2 and high risk MDS were risk-classified according to the International Prognostic Scoring System (IPSS). The study was approved by the Ethics Committee of Shanghai East Hospital, Tongji University School of Medicine (Shanghai, China; research no. 136, 2018). Patients who participated in this research signed an informed consent and had complete clinical data. MDS patients with a median age of 62 years were treated with 5-azaC subcutaneously at a dose of 75 mg/m2/day on the first 7 days of a 28-day cycle. The median time of treatment with 5-azaC was 3 months. After informed consent, all 30 patients provided peripheral blood and bone marrow samples for analysis, prior to treatment, and at 1, 2 and 3 months while on 5-azaC treatment (Table I).
Table I.
Clinical data of MDS patients before 5-azaC treatment.
| Variables | Cases | Normal chromosome karyotype n (%) | Complex abnormal karyotype n (%) | P-value |
|---|---|---|---|---|
| Age (years) | 0.484 | |||
| <65 | 16 | 6 (37.5) | 10 (62.5) | |
| ≥65 | 14 | 8 (57.1) | 6 (42.9) | |
| Sex | 0.569 | |||
| Male | 17 | 5 (29.4) | 12 (70.6) | |
| Female | 13 | 6 (46.1) | 7 (53.8) | |
| WHO classification | 0.732 | |||
| RA | 2 | 1 (50.0) | 1 (50.0) | |
| RARS | 3 | 1 (33.3) | 2 (66.7) | |
| RCMD | 7 | 2 (28.5) | 5 (71.5) | |
| RAEB-1 | 8 | 4 (50.0) | 4 (50.0) | |
| RAEB-2 | 5 | 3 (60) | 2 (40) | |
| MDS-U | 3 | 2 (66.7) | 1 (33.3) | |
| CMML | 2 | 1 (50.0) | 1 (50.0) | |
| IPSS grouping | 0.594 | |||
| Intermediate-2 | 12 | 6 (50.0) | 6 (50.0) | |
| High risk | 18 | 7 (38.9) | 11 (61.1) |
MDS, myelodysplastic syndrome; 5-azaC, 5-azacytidine; RA, refractory anemia; RARS, refractory anemia with ring sideroblasts; RCMD, refractory cytopenia with multilineage dysplasia; RAEB-1, refractory anemia with excess blasts-1; RAEB-2, refractory anemia with excess blasts-2; MDS-U, myelodysplastic syndrome-unclassifiable; CMML, chronic myelomonocytic leukemia; IPSS, International Prognostic Scoring System.
Mononuclear cell separation
Mononuclear cells were separated from peripheral blood (PBMC) by density gradient sedimentation. For the in vitro assays, CD4+ T cells were subsequently isolated by magnetic-activated cell sorting (MACS) using the CD4+ isolation kit from Miltenyi Biotec, GmbH. To obtain CD3+CD4+CD25+FoxP3+ Tregs and CD3+CD4+IL-17+ Th17 cells, PBMC were first enriched for CD4+ T cells using a negative isolation kit (Miltenyi Biotec, Inc.) and were stained with anti-human CD4, CD25 and FoxP3. Purified Tregs and Th17 cells, defined as CD3+CD4+CD25+FoxP3+ and CD3+CD4+IL-17+, were sorted using a FACSAria sorter (BD Biosciences).
Antibodies, reagents, and flow cytometry
Peripheral blood CD4+ T cells (1×106/ml) of patients were stimulated with 500 ng/ml phorbol 12-myristate 13-acetate (PMA) and ionomycin in complete medium for 4 h, After further 4 h, CD4+ T cells were harvested and washed with PBS. To analyze the proportion of Th17 cells, CD4+ T cells were first stained with FITC-conjugated anti-human CD4 antibody at 4°C for 30 min. Then, they were fixed and permeabilized with fixation/permeabilization buffer and were intracellularly stained with APC-conjugated anti-human IL-17A antibody at room temperature in the dark for 30 min. To analyze the proportion of Tregs, CD4+ T cells were simultaneously stained with FITC-conjugated anti-human CD4 antibody and PC7-conjugated anti-human CD25 antibody, then they were fixed and permeabilized, and intracellularly stained with PE-conjugated anti-human FoxP3 antibody at room temperature in the dark for 30 min. Isotype-matched control antibodies were used in all staining processes. Flow cytometry was performed on a FACSCanto II system using FACSDiva software (BD Biosciences). Data were analyzed on FlowJo software (Tree Star, Inc.). Antibodies: CD3-ECD (mouse, monoclonal, dilution: 5 µl/test, cat. no. A07748, Beckman Coulter, Inc.), CD4-PE (mouse, monoclonal, dilution: 5 µl/test, cat. no. 347327; Becton, Dickinson and Company), CD4-FITC (mouse, monoclonal, dilution: 5 µl/test, cat. no. A07750; Beckman Coulter, Inc.), CD25-PC7 (mouse, monoclonal, dilution: 5 µl/test, cat. no. A52882; Beckman Coulter, Inc.), FoxP3-PE (mouse, monoclonal, dilution: 10 µl/test, cat. no. B46031; Beckman Coulter, Inc.), IL-17-488A (rabbit, monoclonal, dilution: 10 µl/test, cat. no. ab217359; Abcam).
Stimulation of isolated CD4+ T-cell subset
5-azaC (Sigma- Aldrich: Merck KGaA) was dissolved in acetic acid to a concentration of 20 mM and was used at 1 µM. CD4+ T cells (20×106/ml) were treated by freshly dissolved and diluted 5-azaC at a concentration of 1 µM or an equal volume of vehicle (every 24 h for 96 h). Carboxyfluorescein diacetate, succinimidyl ester (CFDA-SE or CFSE) dilution was determined by flow cytometry and the proliferation index was calculated by ModFit software (Verity Software House, Inc.).
Immunohistochemistry of bone marrow
Bone marrow was collected from patients with MDS after treatment with 5-azaC or vehicle for immunohistochemical staining and the samples were fixed with formaldehyde. FoxP3, RORγt and Tbet staining was carried out to determine the expression of transcription factors in bone marrow. The results of immunohistochemistry were obtained by a double-blind method. Five high-power fields were selected, and the results were converted into mm−2. The average value was selected as the final result. Fixative (4%) was used at room temperature for 12 h, and the samples were embedded in paraffin. The thickness of the paraffin sections was approximately 6 µm. Bovine serum albumin (BSA; 5%) was used for blocking at 37°C for 30 min. The sections were incubated with rabbit anti-FoxP3 monoclonal antibody [SP97] (cat. no. ab99963; Abcam), rabbit anti-RORγt monoclonal antibody (cat. no. ab219496; Abcam), and goat anti-rabbit IgG H&L (HRP) (cat. no. ab205718; Abcam) at 37°C for 30 min. Olympus CX41 (×400, ×200; Olympus, Corp.) was used.
Methylation assay
Genomic DNA was extracted with a blood and cell culture DNA mini kit (Qiagen, Inc.) and was digested with an EpiTect Methyl DNA Restriction kit (SABiosciences: Qiagen, Inc.). The digested product was used to quantify the remaining DNA using quantitative SYBR Green PCR. Primer sets used for amplification of human FoxP3, RORγT and Tbet promoter CpG islands were purchased from SABiosciences (Qiagen, Inc.). Quantitative PCR was performed on an ABI Fast 7500 sequence detection system (Applied Biosystems: Thermo Fisher Scientific, Inc.). Anti-mouse FoxP3 monoclonal antibody (PE) (cat. no. E-AB-F1208I; Elabscience Biotechnology, Inc.) was used. FoxP3 forward, 5′-TAGCCTCGATGTACGT-3′ and reverse, 5′-AGCCTGACGACCTAGCTCG-3′. β-actin was the reference gene; forward, 5′-TCACAGACACTGTGCTCATCTACGA-3′ and reverse, 5′-TAGCGTAACCGCTCGTTGCCAATGG-3′. Thermocycling conditions: 94°C (hot start) for 10 min, followed by 30 cycles at 94°C for 30 sec, at 55°C for 30 sec, and at 72°C for 30 sec, with a final extension at 72°C for 10 min. 2−ΔΔCq was the quantification method used in this study (13).
Statistical analysis
GraphPad Prism 5.0 (GraphPad Software, Inc.) and SPSS 19.0 (IBM Corp.) software programs were used for statistical analysis. Data are presented as n (%) or the mean ± standard deviation (SD). The measurement data were compared using the independent sample t-test, the enumeration data were compared by χ2 test, and the paired t-test was used for comparisons between two groups. P<0.05 was considered to indicate a statistically significant difference.
Results
In vivo effect of 5-azaC on Tregs
Tregs isolated in vivo showed 98.2% cell viability with 90.8% cell purity. Administration of 5-azaC increased the absolute number of peripheral blood Tregs within 3 months [1.405±0.213 vs. 2.521±0.187 (×107/l)] (mean ± SD, n=30) (P=0.021) (Fig. 1A). A modest temporary increase in the number of Tregs arose following 5-azaC treatment in the first month, although this difference was not statistically significant. The absolute numbers of peripheral blood Tregs from an MDS patient are presented in Fig. 1B and C as a representative example. An increase was observed from 2.416% of CD4+ T lymphocytes (Fig. 1B) to 5.239% of CD4+ T lymphocytes (Fig. 1C) after 3 months of therapy.
Figure 1.

Effect of 5-azaC on the number of Tregs. (A) The absolute number of Tregs was significantly higher after 3 months of therapy and the difference was statistically significant [1.405±0.213 vs. 2.521±0.187 (×107/l)] (mean ± SD, n=30; P=0.021). Notably, after 1 month of treatment the number of Tregs was slightly increased (left panel). In addition, the number of CD4+ T lymphocytes was significantly increased after 3 months (P=0.041) (right panel). (B) Representative example of an MDS patient. The number of Tregs at diagnosis was 2.416% of CD4+ lymphocytes. (C) After 3 months of therapy with 5-azaC, the number of Tregs was increased to 5.239% of CD4+ lymphocytes. 5-azaC, 5-azacytidine; Tregs, regulatory T cells; MDS, myelodysplastic syndrome.
Following treatment with 5-azaC, the absolute number of Th17 cells after 3 months of treatment showed no statistically significant difference [0.518±0.012 vs. 0.509±0.014 (×107/l)] (mean ± SD, n=30) (P=0.783) (Fig. 2A). A representive example of an MDS patient is presented in Fig. 2B and C. The number of Th17 cells at diagnosis was 0.25% of CD4+ T lymphocytes (Fig. 2B). After 3 months of therapy with 5-azaC, the number of Th17 cells was increased to 0.36% of CD4+ T lymphocytes (Fig. 2C).
Figure 2.

Effect of 5-azaC on the number of Th17 cells. (A) Following treatment with 5-azaC, the absolute number of Th17 cells after 3 months showed no statistically significant difference [0.518±0.012 vs. 0.509±0.014 (×107/l)] (mean ± SD, n=30; P=0.783) (left panel). Also, the number of CD4+ T lymphocytes showed no statistically significant difference after 3 months of treatment (P=0.217) (right panel). (B) Representative example of an MDS patient. The number of Th17 cells at diagnosis was 0.25% of CD4+ lymphocytes. (C) After 3 months of therapy with 5-azaC, the number of Th17 cells was increased to 0.36% of CD4+ lymphocytes. 5-azaC, 5-azacytidine; Th17, T-helper 17; MDS, myelodysplastic syndrome.
In vitro effect of 5-azaC on the development of Tregs
According to a dose-response experiment, examining T-cell proliferation (8,9), a dose previously shown to be non-toxic was used. The overall proportion of Tregs in CD4+ T cells from thirty patients was increased by 5-azaC at a concentration of 1 µM (every 24 h for 96 h) (0.491±0.157 vs. 2.912±0.403%) (P=0.014) (Fig. 3A), while the overall proportion of Th17 cells remained unchanged (0.480±0.169 vs. 0.583±0.238%) (P=0.308) (Fig. 3B). These findings raise the question as to whether the increase in Tregs among CD4+ T cells is due to conversion of conventional CD25− cells or proliferation of CD25+FoxP3+ cells. To address this question, 5-azaC (1 µM) was added to Tregs and Th17 cells separately (every 24 h for 96 h). Addition of 5-azaC caused a significant reduction in the absolute numbers of Tregs (0.417±0.193 vs. 4.064±0.345%) (P=0.018) (Fig. 3C). The proliferation of Th17 cells was reduced following 5-azaC addition (Fig. 3D). However, this difference was not statistically significant (0.614±0.127 vs. 0.924±0.142%) (P=0.351). Thus, Tregs ceased to proliferate when cultured in the presence of 5-azaC, indicating that the increase in Tregs among proliferating CD4+ T cells in the presence of 5-azaC is due to conversion of conventional CD25− cells, rather than proliferation of CD25+FoxP3+ cells.
Figure 3.

In vitro effect of 5-azaC on the development of Tregs and Th17 cells. (A and B) The proportion of Tregs was enhanced by 5-azaC in vitro in 96 h (0.491±0.157 vs. 2.912±0.403%) (*P<0.05, P=0.014), while the overall proportion of Th17 cells remained unchanged (0.480±0.169 vs. 0.583±0.238%) (n.s., P>0.05; P=0.308). (C) Tregs from MDS patients were significantly reduced in the presence of 5-azaC in 96 h, compared with the untreated patients (0.417±0.193 vs. 4.064±0.345%) (*P<0.05, P=0.018). (D) 5-azaC had little or no effect on the absolute numbers of similarly treated Th17 cells in 96 h (0.614±0.127 vs. 0.924±0.142%) (P=0.351). 5-azaC, 5-azacytidine. Tregs, regulatory T cells; Th17, T-helper 17; MDS, myelodysplastic syndrome.
Effect of 5-azaC on the expression of transcription factors
To study whether the conversion was due to a change in the expression of transcription factors, immunohistochemistry was performed on CD4+ T cells in bone marrow. No obvious changes in Tbet and RORγT mRNA transcription were observed, while FoxP3 mRNA transcription was enhanced significantly (80.631±10.489 vs. 30.541±5.815%) (P=0.028) (Fig. 4A). Previous studies have reported that methylation status of the FoxP3 gene promoter plays an important role in the regulation of FoxP3 expression, demonstrating an upregulation of FoxP3 expression in CD4+ T cells upon stimulation by 5-azaC (14,15). Tregs from MDS patients were stimulated by 5-azaC or vehicle for 96 h. The methylation status of CpG islands in the promoter of the FoxP3 gene was assessed. In Tregs treated by 5-azaC, the percentage of moderately methylated promoter decreased from 56.782±6.021% to 28.541±10.815% (P=0.026) (Fig. 4B), whereas the percentage of unmethylated promoter increased from 25.127±10.315% to 53.619±12.614% (P=0.032) (Fig. 4B).
Figure 4.
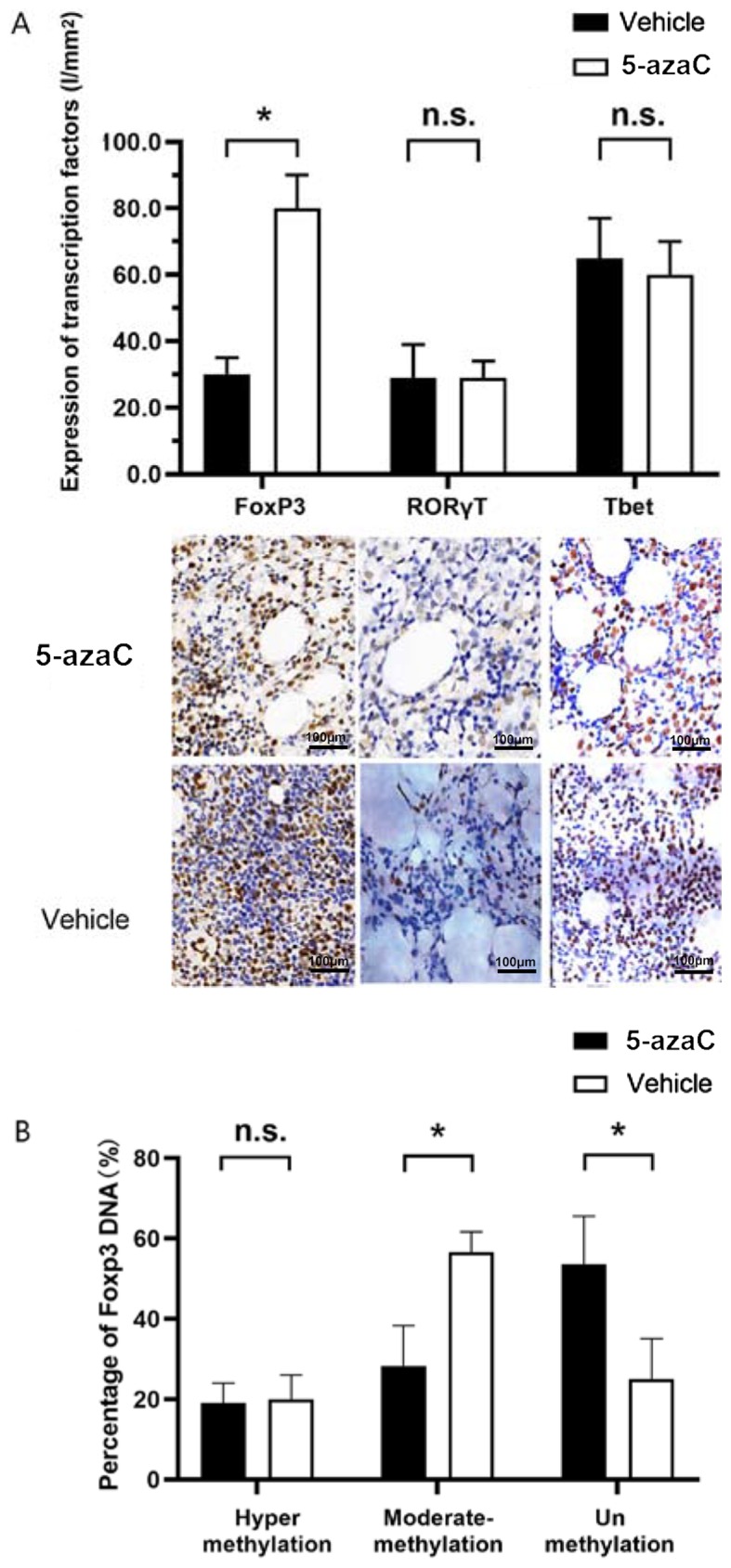
(A) Transcription factor expression of FoxP3 with 5-azaC was significantly reduced (80.631±10.489 vs. 30.541±5.815%) (*P<0.05, P=0.028), in CD4+ T cells in bone marrow immunohistochemistry, while the expression of RORγT and Tbet was not significantly changed (n.s., P>0.05; P=0.631 and 0.319, respectively). (B) In Tregs treated with 5-azaC, the percentage of moderately methylated promoter decreased from 56.782±6.021% to 28.541±10.815% (*P<0.05, P=0.026). The percentage of unmethylated promoter increased from 25.127±10.315% to 53.619±12.614% (*P<0.05, P=0.032). 5-azaC, 5-azacytidine; Tregs, regulatory T cells.
Discussion
5-azaC prolongs overall survival, in comparison to the conventional care regimens, inducing hematologic responses in up to 56.5% of MDS patients and a prolongation of patients' survival of ~10 months (2). The advantage of 5-azaC therapy has also been verified in numerous other studies. Goodyear et al (5) have shown that 5-azaC treatment induces CD4+ T-cell responses with an increase in the number of Tregs. Choi et al (9) have shown an intact GVL effect in transplanted mice following treatment with 5-azaC or decitabine, also with an increase in the number of FoxP3+ T cells. Costantini et al (16) have shown that Treg numbers increase in responders after 9 months of treatment. All these findings suggest that post-treatment Tregs do actually exert a GVL effect, rather than function as ‘suppressors’.
The transcription factor FoxP3, produced by Tregs, usually exists in the form of hypermethylation, and its expression and methylation are dissimilar in different diseases. Thymic regulatory T cell (nTreg) is the main Treg in immune tolerance of healthy people or patients after haematopoietic stem cell transplantation (HSCT). While in tumor diseases, such as MDS, it is replaced by induced T cell (iTreg) (17). Because the degree of demethylation of FoxP3 in iTreg cells is relatively weak and the stability is poor, the number of Tregs in MDS which have an immune tolerance effect is lower than that of HSCT. According to the results of this study, some Tregs were transformed from T cells of CD25− and transformed from naïve T cells in the presence of cytokines. The mechanism of Tregs in MDS patients is different from that in HSCT patients. So the occurrence of GVL is not only related to the induction of CD4+ T cells, but also to the demethylation of FoxP3 transcription factor promoter.
We recruited 30 patients with intermediate/high-risk MDS for this study and serial peripheral blood samples were tested for CD4+ T-cell subsets. Intriguingly, Treg numbers increased steadily during the 3 months of treatmnet. Initially, we compared the effect of 5-azaC on Tregs and Th17 cells from MDS patients. While 5-azaC significantly inhibited the proliferation of Tregs, the proportion of FoxP3+ Tregs was enhanced in vitro. This suggests that the increase in the percentage of Tregs is due to conversion of conventional CD4+CD25− T cells, rather than proliferation of CD25+FoxP3+ cells.
Tregs play an important role in maintaining the balance of immune response between human health and disease. While MDS patients benefit from demethylation, the number of Tregs increases, inhibiting antitumor immune response and promoting tumor cell escape. On the other hand, Tregs can downregulate excessive inflammation by producing adenosine (Ado), and protect patients from tissue injury and tumor development. Its plasticity is controlled and evolved based on the microenvironment. Therefore, it is very important to monitor the number and function of Tregs and detect the effect of demethylation on Treg transcription factor FoxP3 in MDS.
5-azaC sequesters and promotes degradation of DNMT, inducing DNA hypomethylation, thereby causing re-expression of genes, leading to differentiation and/or apoptosis of the myeloid leukemia cells (18–21). Methylation of the FoxP3 promoter plays an important role in the regulation of FoxP3 expression. 5-azaC sequesters and promotes degradation of DNMT, inducing DNA hypomethylation, thereby causing upregulation of genes (8,14,15). FoxP3 expression acts as the master switch for Tregs and expansion of Tregs is associated with increased FoxP3 expression due to FoxP3 promoter demethylation (22,23). In the present study we confirmed these results and demonstrate that during T-cell activation 5-azaC increases the transcription of FoxP3. Moreover, the reduction in methylation was specific to FoxP3 in the sorted Tregs from thirty patients and then was stimulated for 96 h in the presence of 5-azaC. The reduction in FoxP3 promoter methylation was shown to be associated with increased FoxP3 expression.
The effect of 5-azaC on the function Tregs needs further investigation. Costantini et al (16) have shown that the 5-azaC-treated Tregs have reduced suppressive function. The expanded FoxP3+ cells do not have a regulatory function, unable to suppress the secretion of pro-inflammatory cytokines. However, co-culture of 5-azaC-treated Tregs and T-effectors lead to higher levels of IL-17 secretion in comparison with the T effectors alone. Thus, the expanded Tregs no longer function as ‘suppressors’. Examining the effect of 5-azaC on the function of Tregs will be the aim of our future research.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science Foundation of China (grant nos. 81570190 and 81529001).
Availability of data and materials
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' contributions
XJ and WY conceived and designed the study. XZ acquired the patients' data. LH analyzed and interpreted the data regarding the numbers of CD4+ T-cell subsets in the 30 patients with intermediate-2/high-risk MDS. JS performed quantitative PCR. XJ was a major contributor in writing the manuscript. JS reviewed the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of Shanghai East Hospital, Tongji University School of Medicine (Shanghai, China; research no. 136, 2018). Patients who participated in this research signed an informed consent and had complete clinical data.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Fenaux P, Mufti GJ, Hellstrom-Lindberg E, Santini V, Finelli C, Giagounidis A, Schoch R, Gattermann N, Sanz G, List A, et al. International Vidaza High-Risk MDS Survival Study Group Efficacy of azacitidine compared with that of conventional care regimens in the treatment of higher-risk myelodysplastic syndromes: A randomised, open-label, phase III study. Lancet Oncol. 2009;10:223–232. doi: 10.1016/S1470-2045(09)70003-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fenaux P, Mufti GJ, Hellström-Lindberg E, Santini V, Gattermann N, Germing U, Sanz G, List AF, Gore S, Seymour JF, et al. Azacitidine prolongs overall survival compared with conventional care regimens in elderly patients with low bone marrow blast count acute myeloid leukemia. J Clin Oncol. 2010;28:562–569. doi: 10.1200/JCO.2009.23.8329. [DOI] [PubMed] [Google Scholar]
- 3.Pinto A, Zagonel V. 5-Aza-2-deoxycytidine (Decitabine) and 5-azacytidine in the treatment of acute myeloid leukemias and myelodysplastic syndromes: Past, present and future trends. Leukemia. 1993;7(Suppl 1):51–60. [PubMed] [Google Scholar]
- 4.Garcia-Manero G, Fenaux P. Hypomethylating agents and other novel strategies in myelodysplastic syndromes. J Clin Oncol. 2011;29:516–523. doi: 10.1200/JCO.2010.31.0854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Goodyear O, Agathanggelou A, Novitzky-Basso I, Siddique S, McSkeane T, Ryan G, Vyas P, Cavenagh J, Stankovic T, Moss P, et al. Induction of a CD8+ T-cell response to the MAGE cancer testis antigen by combined treatment with azacitidine and sodium valproate in patients with acute myeloid leukemia and myelodysplasia. Blood. 2010;116:1908–1918. doi: 10.1182/blood-2009-11-249474. [DOI] [PubMed] [Google Scholar]
- 6.Fandy TE, Herman JG, Kerns P, Jiemjit A, Sugar EA, Choi SH, Yang AS, Aucott T, Dauses T, Odchimar-Reissig R, et al. Early epigenetic changes and DNA damage do not predict clinical response in an overlapping schedule of 5-azacytidine and entinostat in patients with myeloid malignancies. Blood. 2009;114:2764–2773. doi: 10.1182/blood-2009-02-203547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Aggarwal S, van de Loosdrecht AA, Alhan C, Ossenkoppele GJ, Westers TM, Bontkes HJ. Role of immune responses in the pathogenesis of low-risk MDS and high-risk MDS: Implications for immunotherapy. Br J Haematol. 2011;153:568–581. doi: 10.1111/j.1365-2141.2011.08683.x. [DOI] [PubMed] [Google Scholar]
- 8.Sánchez-Abarca LI, Gutierrez-Cosio S, Santamaría C, Caballero-Velazquez T, Blanco B, Herrero-Sánchez C, García JL, Carrancio S, Hernández-Campo P, González FJ, et al. Immunomodulatory effect of 5-azacytidine (5-azaC): Potential role in the transplantation setting. Blood. 2010;115:107–121. doi: 10.1182/blood-2009-03-210393. [DOI] [PubMed] [Google Scholar]
- 9.Choi J, Ritchey J, Prior JL, Holt M, Shannon WD, Deych E, Piwnica-Worms DR, DiPersio JF. In vivo administration of hypomethylating agents mitigate graft-versus-host disease without sacrificing graft-versus-leukemia. Blood. 2010;116:129–139. doi: 10.1182/blood-2009-12-257253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kordasti SY, Afzali B, Lim Z, Ingram W, Hayden J, Barber L, Matthews K, Chelliah R, Guinn B, Lombardi G, et al. IL-17-producing CD4(+) T cells, pro-inflammatory cytokines and apoptosis are increased in low risk myelodysplastic syndrome. Br J Haematol. 2009;145:64–72. doi: 10.1111/j.1365-2141.2009.07593.x. [DOI] [PubMed] [Google Scholar]
- 11.Kordasti SY, Ingram W, Hayden J, Darling D, Barber L, Afzali B, Lombardi G, Wlodarski MW, Maciejewski JP, Farzaneh F, et al. CD4+CD25high Foxp3+ regulatory T cells in myelodysplastic syndrome (MDS) Blood. 2007;110:847–850. doi: 10.1182/blood-2007-01-067546. [DOI] [PubMed] [Google Scholar]
- 12.Polansky JK, Kretschmer K, Freyer J, Floess S, Garbe A, Baron U, Olek S, Hamann A, von Boehmer H, Huehn J. DNA methylation controls Foxp3 gene expression. Eur J Immunol. 2008;38:1654–1663. doi: 10.1002/eji.200838105. [DOI] [PubMed] [Google Scholar]
- 13.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 14.Lal G, Zhang N, van der Touw W, Ding Y, Ju W, Bottinger EP, Reid SP, Levy DE, Bromberg JS. Epigenetic regulation of Foxp3 expression in regulatory T cells by DNA methylation. J Immunol. 2009;182:259–273. doi: 10.4049/jimmunol.182.1.259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nagar M, Vernitsky H, Cohen Y, Dominissini D, Berkun Y, Rechavi G, Amariglio N, Goldstein I. Epigenetic inheritance of DNA methylation limits activation-induced expression of FOXP3 in conventional human CD25−CD4+ T cells. Int Immunol. 2008;20:1041–1055. doi: 10.1093/intimm/dxn062. [DOI] [PubMed] [Google Scholar]
- 16.Costantini B, Kordasti SY, Kulasekararaj AG, Jiang J, Seidl T, Abellan PP, Mohamedali A, Thomas NS, Farzaneh F, Mufti GJ. The effects of 5-azacytidine on the function and number of regulatory T cells and T-effectors in myelodysplastic syndrome. Haematologica. 2013;98:1196–1205. doi: 10.3324/haematol.2012.074823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Whiteside TL. Regulatory T cell subsets in human cancer: Are they regulating for or against tumor progression? Cancer Immunol Immunother. 2014;63:67–72. doi: 10.1007/s00262-013-1490-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mund C, Brueckner B, Lyko F. Reactivation of epigenetically silenced genes by DNA methyltransferase inhibitors: Basic concepts and clinical applications. Epigenetics. 2006;1:7–13. doi: 10.4161/epi.1.1.2375. [DOI] [PubMed] [Google Scholar]
- 19.Raj K, John A, Ho A, Chronis C, Khan S, Samuel J, Pomplun S, Thomas NS, Mufti GJ. CDKN2B methylation status and isolated chromosome 7 abnormalities predict responses to treatment with 5-azacytidine. Leukemia. 2007;21:1937–1944. doi: 10.1038/sj.leu.2404796. [DOI] [PubMed] [Google Scholar]
- 20.Pinto A, Attadia V, Fusco A, Ferrara F, Spada OA, Di Fiore PP. 5-Aza-2-deoxycytidine induces terminal differentiation of leukemic blasts from patients with acute myeloid leukemias. Blood. 1984;64:922–929. doi: 10.1182/blood.V64.4.922.922. [DOI] [PubMed] [Google Scholar]
- 21.Yang AS, Doshi KD, Choi SW, Mason JB, Mannari RK, Gharybian V, Luna R, Rashid A, Shen L, Estecio MR, et al. DNA methylation changes after 5-aza-2-deoxycytidine therapy in patients with leukemia. Cancer Res. 2006;66:5495–5503. doi: 10.1158/0008-5472.CAN-05-2385. [DOI] [PubMed] [Google Scholar]
- 22.Kim HP, Leonard WJ. CREB/ATF-dependent T cell receptor-induced FoxP3 gene expression: A role for DNA methylation. J Exp Med. 2007;204:1543–1551. doi: 10.1084/jem.20070109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wilson CB, Rowell E, Sekimata M. Epigenetic control of T-helper-cell differentiation. Nat Rev Immunol. 2009;9:91–105. doi: 10.1038/nri2487. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.