

Abstract
Background:
LHX1 is an important transcription factor for the HDAC8 gene. The aim of this study was to investigate the effect of Sodium Butyrate (SB), as a histone deacetylase inhibitor, on the expression of LHX1 gene in colorectal cancer cell lines.
Methods:
HT-29 and HCT-116 cell lines were treated with 6.25 to 200 mM concentrations of SB at 24, 48, and 72 hr. The cytotoxicity effect on cell viability was evaluated by MTT assay. The 50% Inhibiting Concentration (IC50) was determined graphically. Quantitative real-time PCR was performed to investigate the LHX1 mRNA expression level.
Results:
Our study revealed that SB inhibited the proliferation of these cell lines in a concentration and time-dependent manner. The IC50 values for HT-29 cell line were 65, 18.6, and 9.2 mM after 24, 48, and 72 hr of treatment, respectively. The IC50 values for HCT-116 cell line were 35.5, 9.6, and 10 mM after 24, 48, and 72 hr of treatment, respectively. Furthermore, real-time PCR findings demonstrated that the LHX1 mRNA expression in treated HT-29 cell line significantly increased in comparison with untreated cells (p<0.05). However, in treated HCT-116 cell line, SB led to a significant decrease in the level of LHX1 mRNA (p<0.05), as compared to untreated cells.
Conclusion:
In this study, different effects of SB on LHX1 mRNA expression level were revealed in two distinct human colorectal cancer cell lines.
Keywords: Colorectal cancer, HCT-116 cells, Histone deacetylase inhibitors, Humans, Transcription factors
Introduction
Colorectal cancer is one of the most common cancers in the world including 9% of all cancers 1. This cancer is the second common cancer and the fourth cause of death due to cancer globally 2. Dysregulation in the epigenetic mechanisms, including histone acetylation, is one of the main factors contributing to the colorectal cancer 3–5. Acetylation, a process in which the chromatin structure and gene expression 6 are modified, is controlled by two types of enzymes, Histone Acetylases (HAT) and Histone Deacetylases (HDACs) 7. The change in acetylation status in cancer cells such as prostate 8, colon 9, and gastric 10 cancers has been linked to the increased expression of certain HDAC in indefinite patterns.
HDACs directly interact with transcription factors and can regulate the expression of a large number of genes 11. LHX1 (LIM Homeobox1) protein is one of the transcription factors involved in the transcription of HDAC8 gene 12. Moreover, it has different functions including regulation of cell fate, cellular skeleton organization, and tumor formation 13–16. The LHX1 expression has been reported in human cancers such as ovarian cancer, kidney carcinoma, leukemia cells, and epithelial cells 17.
Histone Deacetylase Inhibitors (HDACi) can change the balance between HAT and HDAC, and also lead to the acetylation of histone and non-histone proteins that induce transcription and related molecular effects 18. Some processes involved in the inhibition of HDAC are apoptosis, necrosis, growth inhibition, and differentiation 19–21. One of the HDACi is Sodium Butyrate (SB) 22,23. The produced butyrate in the colon may inhibit the development of colon cancer and protect against colon cancer 24,25. One of the functions of butyrate is its anti-inflammatory effect that plays a crucial role in inhibiting the histone deacetylase 26. In addition, SB influences the gene expression through binding to the transcription factors. Epigenetic regulation orchestrates various physiological procedures, comprising transcription, replication, and repair from developmental to differentiated stages and emerges with a pivotal role in the process of tumorigenesis 27–29. The understanding of these mechanisms might contribute to the optimization of prognostic and diagnostic systems, as well as the generation of novel and targeted therapeutic approaches. In the present study, the effect of SB on LHX1 mRNA expression, as a transcription factor of the HDAC8 gene, in HT-29 and HCT116 human colorectal cell lines was investigated. It is expected that the expression of LHX1 in treated cells would be decreased, in comparison with untreated cells. Our results showed that in HCT-116 cells, the expression of LHX1 was decreased; however, in HT-29 cells this expression level was increased, compared with untreated cells. One of the explanations for this may be the different tissue origin of these two cell lines given the fact that HT-29 is adenocarcinoma and HCT-116 is carcinoma. Furthermore, these cell lines represent a wide range of cancer characteristics; HCT-116 has a wild-type p53 response while being deficient in mismatch repair, whereas the HT-29 is p53 deficient and an unstable cell line 30. Molecular mechanisms may affect the underlying function in each cell line.
Materials and Methods
Cell culture
HT-29 and HCT116 human colorectal cell lines were purchased from Pasteur Institute of Iran (Tehran, Iran). HT-29 and HCT116 cells were cultured in RPMI 1640 and DMEM (Dulbecco’s Modified Eagle’s Medium) (Gibco, Germany), respectively, which was supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Gibco, Germany) and 1% penicillin-streptomycin (100 IU/ml and 100 μg/ml, respectively) (Dacell, Iran). Cells were incubated at 37°C under a humidified atmosphere of 95% air and 5% CO2 (v/v). Monolayer cells were harvested by 0.25% trypsin-EDTA (Gibco, Germany).
SB treatment
Optimization of cell numbers in 96-well plates (Spl life sciences, Korea) was performed for 24, 48, and 72 hr of incubation time. A total of 50×103 cells per well (The optimized cell number) were seeded in 96-well plates and incubated for 24 hr. SB was dissolved in sterile water with a 1 molar concentration of stock solution for in vitro studies, which was further diluted to the working concentration (6.25 to 200 mM) in culture media. All cell lines were then treated with SB at the concentrations ranging from 6.25 to 200 mM for 24, 48, and 72 hr. Untreated cells (0 Mm) and cells treated with dimethyl sulfoxide (DMSO) 20% were considered as negative and positive controls, respectively.
Cytotoxicity assay
The cytotoxic effect of SB (Biobasic, Canada Inc.) in HT-29 and HCT-116 colorectal cell lines was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma, USA) and was compared with the untreated cells (0 Mm) as a control group. Briefly, 100 μl of the MTT stock solution (5 mg/ml in PBS) was added to each well to attain a final concentration of 0.5 mg/ml in RPMI-1640 without phenol red culture (Biosera, France). After 4 hr of incubation, the supernatants were aspirated; the formazan crystals in each well were dissolved in 50 μl DMSO and the absorbance was measured at 546 nm using an ELISA reader (Garni Medical Eng. Co., Tehran, Iran). Each SB concentration was assayed in separated wells and each experiment was repeated at least 3 times. Cell viabilities were calculated using the following formula:
Afterwards, the half-maximal growth inhibitory concentration (IC50) values were estimated from dose response curves by applying linear regression analysis via the JavaScript version of PolySolve (07.20.2013) software.
RNA extraction and cDNA synthesis
A total of 3×106 HT-29 and HCT-116 human colorectal cells were seeded in 6-well plates (Spl life sciences, Korea) in 2 ml of RPMI-1640 and DMEM medium supplemented with 2% FBS, respectively, and were treated with different concentrations of SB (6.25 to 200 mM) for 24 and 48 hr. After the end of incubation time, total cellular RNA was extracted from the cancer cells treated with SB and untreated cells using RNX-Plus Solution (Sinaclon, Iran). The quality and quantity of extracted RNA were measured with agarose gel electrophoresis and a spectrophotometer (Eppendorf, Germany). Complementary DNA (cDNA) was synthesized with 2000 ng total RNA using a cDNA synthesis kit (Yektatajhiz, Iran) according to the manufacturer's protocol.
Quantitative real-time PCR (qRT-PCR)
The qRT-PCR analysis was carried out for LHX1 gene using RQ-PCR SYBR Green I system Light Cycler 96 (Roche Diagnostics, Germany). The GAPDH (Housekeeping gene) was used as an internal control. Reactions were prepared in duplicate using 2X SYBR Green Supermix (Pishgam, Iran) according to manufacturer’s instructions to a final volume of 20 μl. The following conditions were used: 95°C for 15 min, followed by 40 cycles of denaturation at 95°C for 15 s, annealing, and extension at 60°C for 60s. Quality of PCR products was evaluated by generating a melting curve, which was also used to verify the absence of PCR artifacts (Primer-dimers) or nonspecific PCR products. Variations in relative gene expressions between treated cells and control group (Untreated cells) cDNA samples were identified with Relative Expression Software Tool 9 (REST 9, Qiagen) using the 2−ΔΔCT method. The primers (10 pmol) are listed in table 1.
Table 1.
Primer sequences used in quantitative polymerase chain reaction (qRT-PCR)
| Name | Forward primer sequence (5′–3′) | Reverse primer sequence (5′–3′) | Accession number |
|---|---|---|---|
| GAPDH | GAAGGTGAAGGTCGGAGTC | GAAGATGGTGATGGGATTTC | NM_001289745.2 |
| LHX1 | TCTCCAGGGAAGGCAAACT | CGAAACACCGGAAGAAGTC | NM_005568.4 |
Data analysis
Ct values were adjusted, taking into account primer efficiencies for each gene when calculating 2−ΔΔCT values. Expression data for each target gene was also normalized to the housekeeping gene (GAPDH) and fold change calculations were made based on Schmittgen and Livak’s method by using REST 9 and LinRegPCR softwares. The level of statistical significance was set at p<0.05.
Results
The effect of SB on the cell viability of HT-29 and HCT-116 human colorectal cancer cell lines
To investigate the role of HDAC on the proliferation of colorectal cancer cells, HT-29, and HCT-116 human colorectal cell lines were treated with various concentrations of SB (From 6.25 to 200 mM) for 24, 48, and 72 hr. Then, the cytotoxicity effect of SB on cancer cells was investigated with MTT assay. The viability of HT-29 and HCT-116 cells was further decreased by higher doses of SB (6.25 to 200 mM). Our study revealed that SB could inhibit the proliferation of HT-29 (Figure 1A) and HCT-116 (Figure 1B) cell lines in a concentration and time-dependent manner.
Figure 1.
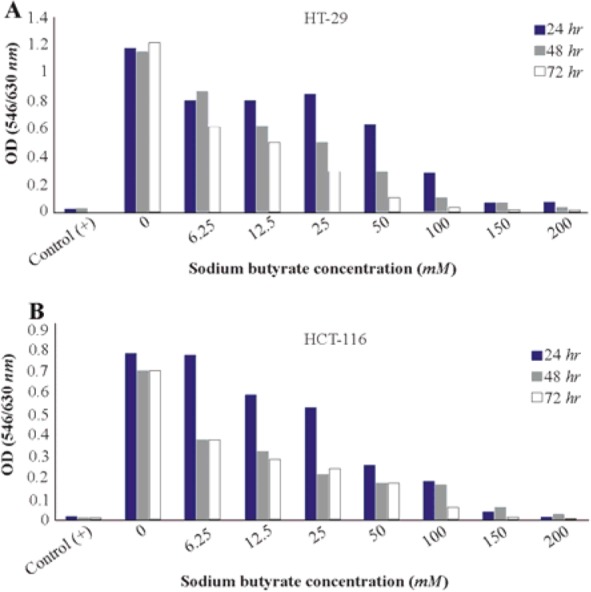
Cell viability in cancer cells treated with sodium butyrate (SB). A) HT-29 colorectal cell line was treated with 6.25 to 200 mM concentrations of SB at 37°C for 24, 48, and 72 hr of incubation. B) HCT-116 colorectal cell line was treated with 6.25 to 200 mM concentrations of SB at 37°C for 24, 48, and 72 hr of incubation. Cell viabilities were evaluated using MTT assay and calculated as a ratio of the control. Control (+): cells treated with dimethyl sulfoxide (DMSO) 20% and untreated cells (0 mM) as negative control. All experiments were performed in triplicate.
The IC50 calculated for SB
The effective concentration of SB for the determination of the half-maximal inhibitory concentration (IC50) value was obtained by regression analyses of concentration-inhibition curves. The IC50 value for HT-29 human colorectal cell line was achieved as 65 mM for the 24 hr of SB treatment, 18.6 mM for 48 hr of SB treatment, and 9.2 mM for 72 hr of SB treatment (Figure 2). As well, the IC50 value for HCT-116 human colorectal cell line was 35.5 mM for 24 hr of SB treatment, 9.6 mM for 48 hr of SB treatment, and 10 mM for 72 hr of SB treatment (Figure 3). The IC50 of SB in HT-29 and HCT-116 human colorectal cancer cell lines was significantly decreased in 24, 48, and 72 hr in a time-dependent manner.
Figure 2.
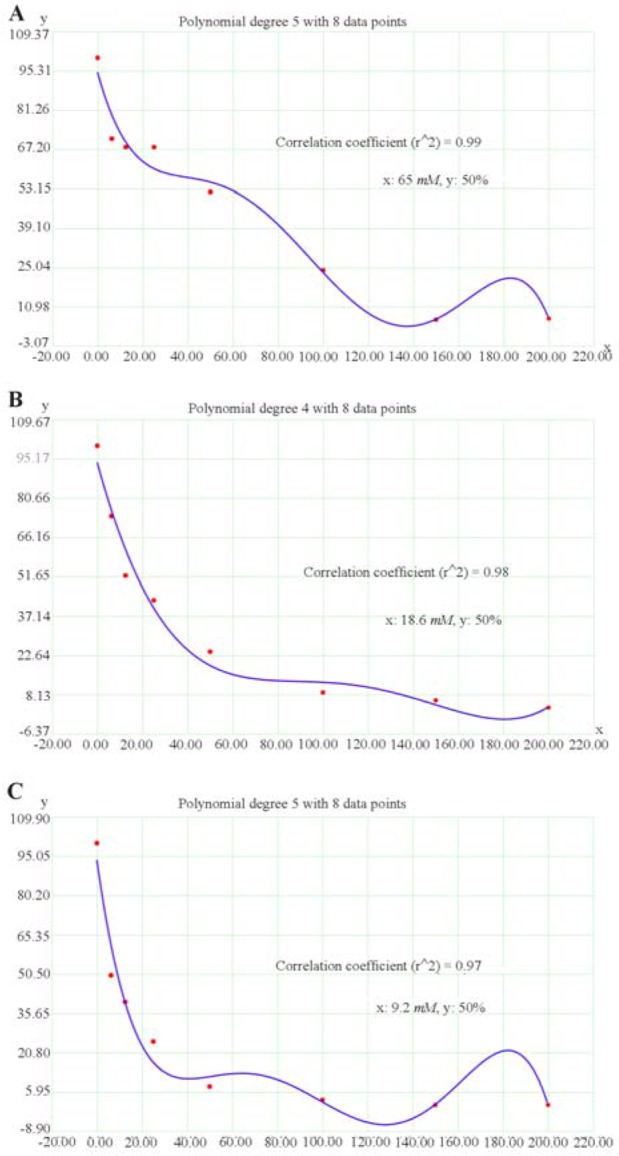
Regression analyses to calculate the 50% inhibiting concentration (IC50) values for effect of sodium butyrate (SB) on HT-29 human colorectal cell line. The horizontal axis (x) represents the concentration (mM) and the vertical axis (y) represents the percentage of the cell viability. A) The IC50 value was 65 mM for 24 hr after treatment, B) 18.6 mM for 48 hr after treatment, and C) 9.2 mM for 72 hr after treatment.
Figure 3.
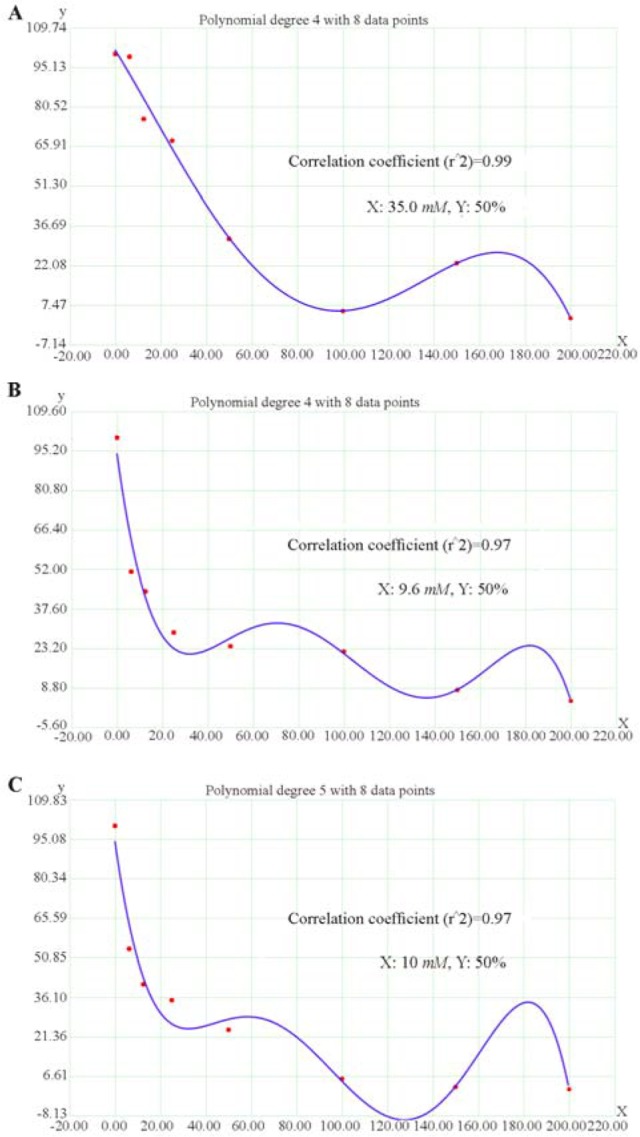
Regression analyses to calculate the 50% inhibiting concentration (IC50) values for effect of sodium butyrate (SB) in HCT-116 human colorectal cell line. The horizontal axis (x) represents the concentration (mM) and the vertical axis (y) represents the percentage of the cell viability. A) The IC50 value was 35.5 mM for 24 hr after treatment, B) 9.6 mM for 48 hr after treatment, and C) 10 mM for 72 hr after treatment.
Quantitative real-time PCR
HT-29 cell line:
The effect of SB was examined on LHX1 mRNA expression in HT-29 human colorectal cancer cell line in vitro by incubating the cells in 6.25, 12.5, 25, 50, and 100 mM concentrations of SB for 24 and 48 hr. The concentrations of 150 and 200 mM were found to be toxic. After 24 hr of incubation with 6.25 to 100 mM concentrations of SB, LHX1 mRNA expression significantly increased in all concentrations, compared with untreated cells as a control group (p<0.05) (Figure 3A); however, in higher concentration of SB, this fold change decreased in comparison with 6.25 mM concentration. This is probably owing to the very low numbers of cells at higher concentrations of SB treatment causing a denominator effect. The increased SB concentrations in the treatment were found to result in reduced cell numbers and enhanced cell death. After 48 hr of incubation, LHX1 mRNA expression was significantly enhanced at concentrations of 6.25, 25, 50, and 100 mM SB, compared with untreated cells as a control group (p<0.05). Nonetheless, there was no significant increase in the concentration of 12.5 mM (p>0.05) (Figure 4).
Figure 4.
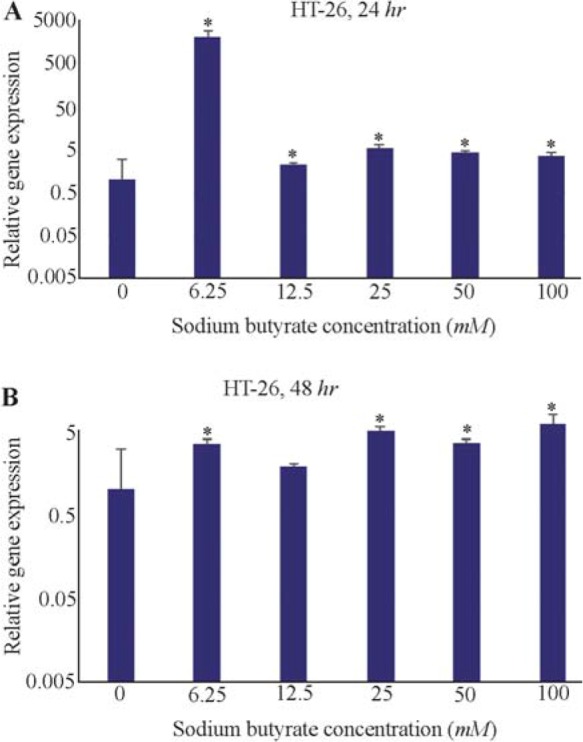
The effect of sodium butyrate (SB) on the LHX1 mRNA expression in HT-29 cell line. A) Cells were cultured for 24 hr with 6.25 to 100 mM concentrations of SB at 37°C. B) Cells were cultured for 48 hr with 6.25 to 100 mM concentrations of SB at 37°C. LHX1 mRNA expression was investigated using qRT-PCR. GAPDH was used as the internal control. LHX1 mRNA expression increased in treated cells compared to control (0 mM). * Indicates a significant increase (p<0.05) vs. controls. All experiments were performed in duplicate.
HCT-116 cell line
Also, the effects of SB on LHX1 mRNA expression in HCT-116 human colorectal cancer cell line were investigated in vitro by incubating the cells in 6.25, 12.5, 25, 50, and 100 mM concentrations of SB for 24 and 48 hr. 24 hr after treatment with SB, LHX1 mRNA expression significantly decreased at concentrations of 6.25, 12.5, 50, and 100 mM SB, compared with untreated cells as a control group (p<0.05). However, there was no significant decrease at the concentration of 25 mM (p>0.05) (Figure 5A). Likewise, 48 hr after treatment with SB, LHX1 mRNA expression was significantly down-regulated in all concentrations of 6.25 to 100 mM SB, compared with untreated cells as a control group (p<0.05) (Figure 5B).
Figure 5.
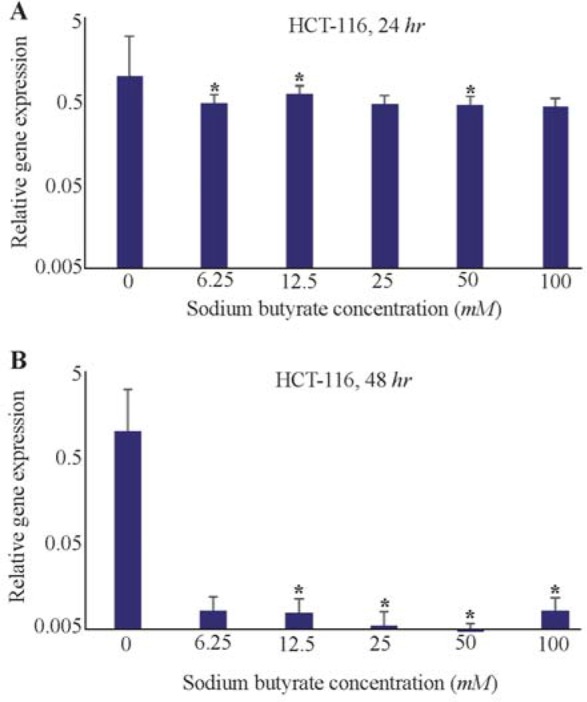
The Effect of sodium butyrate (SB) on LHX1 mRNA expression in HCT-116 cell line. A) Cells were cultured for 24 hr with 6.25 mM to 100 mM of SB at 37°C. B) Cells were cultured for 48 hr with 6.25 mM 100 mM of SB at 37°C. LHX1 mRNA expression investigated using qRT-PCR. GAPDH was used as an internal control. LHX1 mRNA expression decreased in treated cells compared to control (0 mM). * Indicates a significant reduction (p<0.05) vs. controls. All experiments were performed in duplicate.
Discussion
Acetylation is a chief part of the gene expression regulation 31,32 and is controlled by the opposite function of the HAT and HDAC enzymes 7. The dysregulated expression of HDAC enzymes is often seen in cancers 31,32. HDAC can regulate the expression of a large number of genes by direct interaction with transcription factors such as P53, E2f, Stat3, NF-KB, retinoblastoma protein, and TFIIE 11 affecting angiogenesis, cell cycle arrest, apoptosis, and the differentiation of different cell types 33,34. LHX1 is one of the transcription factors involved in the transcription of HDAC8 gene 12. Despite the normal expression of HDAC8 in healthy organs, its expression in tumor tissues is up-regulated 35,36. The selective pharmacological inhibition of HDACi represents a novel treatment for cancer therapy 18,33,34, 37,38. One of the HDACi is SB 22,23. In 2010, Ooi et al examined the effects of SB and their analogs in HT-29 cancer cells and observed that the 5 mmol/L concentration of SB resulted in decreased proliferation, increased apoptosis, and the reduction of HDAC activity 39. The findings of the presents study are consistent with the above information. The cytotoxicity of SB in HT-29 and HCT-116 human colorectal cancer cell lines was examined by using MTT assay. Our results revealed that SB could inhibit the proliferation of both HT-29 and HCT-116 cell lines in a concentration- and time-dependent manner. In HT-29 cell line, the viability of cells decreased to 52, 52, and 50% after 24, 48, and 72 hr of treatment, respectively. Besides, in HCT-116 cell line, the cell viability was diminished to 68, 51 and 54% after 24, 48, and 72 hr of treatment, respectively.
In the present study, for the first time, the effect of SB on the LHX1 mRNA expression was investigated. In 2009, Haberland et al examined the relationship between HDAC8 and homeobox transcription factors of LHX1 and Otx2 using PCR techniques in mice and concluded that the inappropriate expression of these transcription factors suppressed HDAC8 40. In 2011, Dormoy et al reviewed the transcription factor of LHX1 as a new oncogene in kidney cancer cells. They showed LHX1 gene was re-expressed in kidney cancer and it is expressed in large quantities in kidney cancer cells, whereas in the normal kidney cells, it appears with a low expression level. On the other hand, they identified that the reduction of LHX1 expression can lead to an increase in apoptosis and a decrease in cell proliferation after 72 hr 41. In addition, Saha et al have assessed the effects of an HDAC8 inhibitor on the transcription factors of Otx2 and LHX1 in mice. Their results depicted that HDAC8 suppresses the inappropriate expression of Otx2 and LHX1 and these two transcription factors are adjusted by HDAC8 42. Also, according to the literature, it was found that butyrate is able to stop cell cycle, differentiation, and apoptosis in a number of cell lines by inhibiting HDAC 43–45. SB affects the expression of genes by binding to the transcription factors. In this study, the effect of SB on LHX1 as a transcription factor of HDAC8 in colorectal cancer cell lines was investigated. Existing documents have shown the inappropriate expression of LHX1 in cancers that leads to the increased transcription, growth, and proliferation, as well as inhibition of cancer cell apoptosis 17,41. In the current study, it was expected that SB would act as a drug reducing the expression of LHX1. Our findings showed that treatments with SB significantly decreased the expression of LHX1 in HCT-116 cells in comparison with untreated cells (p<0.05). However, to our surprise, the expression of LHX1 significantly increased in HT-29 cell line, compared with untreated cells. Our results are well in line with that of Rocha et al that observed different effects of SB, as HDACi, on the expression of Estrogen Receptor (ERα). They expected that SB would lead to an increase in the ERα expression, while the opposite was found and the ERα expression was reduced 46. They suggested that treatment duration time and used concentrations may be critical in these effects 46. According to our results, Wang et al showed that HDACi could, via HDAC8/YY1, cause suppression of mutant P53 in breast cancer. HDAC8 reacts with YY1 transcription factor and adjusts the transcriptional activity. They figured out that treatment with SAHA and SB can inhibit the HDAC8 and YY1 association, enhance the YY1 acetylation, and eventually suppress the YY1-induced transcription of p53. They, also, determined that the network of HDAC8 and YY1 prevents the proliferation of breast cancer cells 47.
Conclusion
The current study indicated that SB had anticancer activities and inhibits the growth of HT-29 and HCT-116 human colorectal cancer cell lines. Moreover, the results of this study showed that LHX1 mRNA expression level was significantly different between two human colorectal cancer cell lines (HT-29 and HCT-116) due to SB treatment. In HT-29 human colorectal cell line, the significant increase of LHX1 mRNA expression was observed after 24 and 48 hr of incubation time. On the contrary, SB led to a significantly down-regulated LHX1 expression level at 24 and 48 hr of incubation time in HCT-116 human colorectal cell line. Altogether, these results indicated that there is no similar effect of SB on these different cell lines. Worthy of note, the histopathology origins of the used human colorectal cell lines in this study are distinguished. HT-29 is a cell line with adenocarcinoma origin derived from colon ascendens and colon with Dukes’ C stage (Involvement of lymph nodes) 48,49. HCT-116, on the other hand, has a carcinoma tissue origin and is derived from colon ascendens with Dukes’ D stage (Widespread metastases) 50–52. Moreover, the molecular features of these colon cancer cell lines are different 53; thus, their response to drugs is supposed to be distinct. SB might be capable of both repressing and inducing the expression of different genes. In this study, the expression of LHX1 gene was investigated in untreated and treated colorectal cells and different effects of SB on LHX1 mRNA expression were revealed in two different human colorectal cancer cell lines. Future studies are needed to evaluate the effect of SB on LHX1 mRNA expression in other human colorectal cancer cell lines as well as other cancer cell lines.
Acknowledgement
This paper has been resulted from MSc thesis of Mahsa Ghiaghi, student at Faculty of Advanced Science and Technology, Tehran Medical Sciences, Islamic Azad University, Tehran, Iran.
References
- 1.Haggar FA, Boushey RP. Colorectal cancer epidemiology: incidence, mortality, survival, and risk factors. Clin Colon Rect Surg 2009;22(4):191–197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rafiemanesh H, Mohammadian-Hafshejani A, Ghoncheh M, Sepehri Z, Shamlou R, Salehiniya H, et al. Incidence and mortality of colorectal cancer and relationships with the human development index across the world. Asian Pac J Cancer Prev 2016;17(5):2465–2473. [PubMed] [Google Scholar]
- 3.Esteller M. CpG island hypermethylation and tumor suppressor genes: a booming present, a brighter future. Oncogene 2002;21(35):5427–5440. [DOI] [PubMed] [Google Scholar]
- 4.Johnstone RW. Histone-deacetylase inhibitors: novel drugs for the treatment of cancer. Drug Discov 2002;1 (4):287–299. [DOI] [PubMed] [Google Scholar]
- 5.Iizuka M, Smith MM. Functional consequences of histone modifications. Curr Opin Genet Dev 2003;13(2): 154–160. [DOI] [PubMed] [Google Scholar]
- 6.Clayton AL, Hazzalin CA, Mahadevan LC. Enhanced histone acetylation and transcription: a dynamic perspective. Mol Cell 2006;23(3):289–296. [DOI] [PubMed] [Google Scholar]
- 7.Bannister AJ, Kouzarides T. Regulation of chromatin by histone modifications. Cell Res 2011;21(3):381–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Halkidou K, Gaughan L, Cook S, Leung HY, Neal DE, Robson CN. Upregulation and nuclear recruitment of HDAC1 in hormone refractory prostate cancer. Prostate 2004;59(2):177–189. [DOI] [PubMed] [Google Scholar]
- 9.Wilson AJ, Byun DS, Popova N, Murray LB, L'Italien K, Sowa Y, et al. Histone deacetylase 3 (HDAC3) and other class I HDACs regulate colon cell maturation and p21 expression and are deregulated in human colon cancer. J Biol Chem 2006;281(19):13548–13558. [DOI] [PubMed] [Google Scholar]
- 10.Choi JH, Kwon HJ, Yoon BI, Kim JH, Han SU, Joo HJ, et al. Expression profile of histone deacetylase 1 in gastric cancer tissues. Jpn J Cancer Res 2001;92(12):1300–1304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lin HY, Chen CS, Lin SP, Weng JR, Chen CS. Targeting histone deacetylase in cancer therapy. Med Res Rev 2006;26(4):397–413. [DOI] [PubMed] [Google Scholar]
- 12.Micelli C, Rastelli G. Histone deacetylases: structural determinants of inhibitor selectivity. Drug Discov Today 2015;20(6):718–735. [DOI] [PubMed] [Google Scholar]
- 13.Way JC, Chalfie M. mec-3, a homeobox-containing gene that specifies differentiation of the touch receptor neurons in C. elegans. Cell 1988;54(1):5–16. [DOI] [PubMed] [Google Scholar]
- 14.Sánchez-García I, Rabbitts TH. The LIM domain: a new structural motif found in zinc-finger-like proteins. Trends Genet 1994;10(9):315–320. [DOI] [PubMed] [Google Scholar]
- 15.Bridwell JL, Price JR, Parker GE, Schiller AM, Sloop KW, Rhodes SJ. Role of the LIM domains in DNA recognition by the Lhx3 neuroendocrine transcription factor. Gene 2001;277(1):239–250. [DOI] [PubMed] [Google Scholar]
- 16.Yaden BC, Savage JJ, Hunter CS, Rhodes SJ. DNA recognition properties of the LHX3b LIM homeodomain transcription factor. Mol Biol Rep 2005;32(1):1–6. [DOI] [PubMed] [Google Scholar]
- 17.Bowen NJ, Walker LD, Matyunina LV, Logani S, Totten KA, Benigno BB, et al. Gene expression profiling supports the hypothesis that human ovarian surface epithelia are multipotent and capable of serving as ovarian cancer initiating cells. BMC Med Genomics 2009;2:71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bolden JE, Peart MJ, Johnstone RW. Anticancer activities of histone deacetylase inhibitors. Nat Rev Drug Discov 2006;5(9):769–784. [DOI] [PubMed] [Google Scholar]
- 19.Lehrman G, Hogue IB, Palmer S, Jennings C, Spina CA, Wiegand A, et al. Depletion of latent HIV-1 infection in vivo: a proof-of-concept study. Lancet 2005;366(9485):549–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Williams JA, Barreiro CJ, Nwakanma LU, Lange MS, Kratz LE, Blue ME, et al. Valproic acid prevents brain injury in a canine model of hypothermic circulatory arrest: a promising new approach to neuroprotection during cardiac surgery. Ann Thorac Surg 2006;81(6):2235–2242. [DOI] [PubMed] [Google Scholar]
- 21.Almeida AM, Murakami Y, Baker A, Maeda Y, Roberts IA, Kinoshita T, et al. Targeted therapy for inherited GPI deficiency. New Engl J Med 2007;356(16):1641–1647. [DOI] [PubMed] [Google Scholar]
- 22.Hinnebusch BF, Meng S, Wu JT, Archer SY, Hodin RA. The effects of short-chain fatty acids on human colon cancer cell phenotype are associated with histone hyper-acetylation. J Nutr 2002;132(5):1012–1017. [DOI] [PubMed] [Google Scholar]
- 23.Emenaker NJ, Calaf GM, Cox D, Basson MD, Qureshi N. Short-chain fatty acids inhibit invasive human colon cancer by modulating uPA, TIMP-1, TIMP-2, mutant p53, Bcl-2, Bax, p21 and PCNA protein expression in an in vitro cell culture model. J Nutr 2001;131(11 Suppl): 3041S–3046S. [DOI] [PubMed] [Google Scholar]
- 24.Butler LM, Webb Y, Agus DB, Higgins B, Tolentino TR, Kutko MC, et al. Inhibition of transformed cell growth and induction of cellular differentiation by pyroxamide, an inhibitor of histone deacetylase. Clin Cancer Res 2001;7(4):962–970. [PubMed] [Google Scholar]
- 25.He LZ, Tolentino T, Grayson P, Zhong S, Warrell RP, Jr, Rifkind RA, et al. Histone deacetylase inhibitors induce remission in transgenic models of therapy-resistant acute promyelocytic leukemia. J Clin Invest 2001;108(9):1321–1330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cummings JH, Macfarlane GT. The control and consequences of bacterial fermentation in the human colon. J Appl Microbiol 1991;70(6):443–459. [DOI] [PubMed] [Google Scholar]
- 27.Van Engeland M, Derks S, Smits KM, Meijer GA, Herman JG. Colorectal cancer epigenetics: complex simplicity. J Clin Oncol 2011;29(10):1382–1391. [DOI] [PubMed] [Google Scholar]
- 28.Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell 2012;150(1):12–27. [DOI] [PubMed] [Google Scholar]
- 29.You JS, Jones PA. Cancer genetics and epigenetics: two sides of the same coin? Cancer cell 2012;22(1):9–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Danny CW, Waby JS, Chirakkal H, Staton CA, Corfe BM. Butyrate suppresses expression of neuropilin I in colorectal cell lines through inhibition of Sp1 transactivation. Mol Cancer 2010;9(1):276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cress WD, Seto E. Histone deacetylases, transcriptional control, and cancer. J Cell Physiol 2000;184(1):1–6. [DOI] [PubMed] [Google Scholar]
- 32.Timmermann S, Lehrmann H, Polesskaya A, Harel-Bellan A. Histone acetylation and disease. Cell Mol Life Sci 2001;58(5):728–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hildmann C, Riester D, Schwienhorst A. Histone deacetylases—an important class of cellular regulators with a variety of functions. Appl Microbiol Biotechnol 2007;75 (3):487–497. [DOI] [PubMed] [Google Scholar]
- 34.Riester D, Hildmann C, Schwienhorst A. Histone deacetylase inhibitors—turning epigenic mechanisms of gene regulation into tools of therapeutic intervention in malignant and other diseases. Appl Microbiol Biotechnol 2007;75(3):499–514. [DOI] [PubMed] [Google Scholar]
- 35.Song S, Wang Y, Xu P, Yang R, Ma Z, Liang S, et al. The inhibition of histone deacetylase 8 suppresses proliferation and inhibits apoptosis in gastric adenocarcinoma. Int J Oncol 2015;47(5):1819–1828. [DOI] [PubMed] [Google Scholar]
- 36.Wang Y, Xu P, Yao J, Yang R, Shi Z, Zhu X, et al. RETRACTED ARTICLE: MicroRNA-216b is down-regulated in human gastric adenocarcinoma and inhibits proliferation and cell cycle Progression by targeting oncogene HDAC8. Target Oncol 2016;11(2):197–207. [DOI] [PubMed] [Google Scholar]
- 37.Minucci S, Pelicci PG. Histone deacetylase inhibitors and the promise of epigenetic (and more) treatments for cancer. Nat Rev Cancer 2006;6(1):38–51. [DOI] [PubMed] [Google Scholar]
- 38.J Shuttleworth SJ, G Bailey SG, Townsend PA. Histone deacetylase inhibitors: new promise in the treatment of immune and inflammatory diseases. Curr Drug Targets 2010;11(11):1430–1438. [DOI] [PubMed] [Google Scholar]
- 39.Ooi CC, Good NM, Williams DB, Lewanowitsch T, Cosgrove LJ, Lockett TJ, et al. Efficacy of butyrate analogues in HT-29 cancer cells. Clin Exp Pharmacol Physiol 2010;37(4):482–489. [DOI] [PubMed] [Google Scholar]
- 40.Haberland M, Mokalled MH, Montgomery RL, Olson EN. Epigenetic control of skull morphogenesis by his-tone deacetylase 8. Genes Dev 2009;23(14):1625–1630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dormoy V, Beraud C, Lindner V, Thomas L, Coquard C, Barthelmebs M, et al. LIM-class homeobox gene Lim1, a novel oncogene in human renal cell carcinoma. Oncogene 2011;30(15):1753–1763. [DOI] [PubMed] [Google Scholar]
- 42.Saha A, Pandian GN, Sato S, Taniguchi J, Hashiya K, Bando T, et al. Synthesis and biological evaluation of a targeted DNA-binding transcriptional activator with HDAC8 inhibitory activity. Bioorg Med Chem 2013;21 (14):4201–4209. [DOI] [PubMed] [Google Scholar]
- 43.Davie JR. Inhibition of histone deacetylase activity by butyrate. J Nurt 2003;133(7 Suppl):2485S–2493S. [DOI] [PubMed] [Google Scholar]
- 44.Danny CW Yu, Waby JS, Chirakkal H, Staton CA, Corfe BM. Butyrate suppresses expression of neuropilin I in colorectal cell lines through inhibition of Sp1 transactivation. Mol Cancer 2010;9(1):276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Waby JS, Chirakkal H, Yu C, Griffiths GJ, Benson RS, Bingle CD, et al. Sp1 acetylation is associated with loss of DNA binding at promoters associated with cell cycle arrest and cell death in a colon cell line. Mol Cancer 2010;9(1):275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rocha W, Sanchez R, Deschênes J, Auger A, Hebert E, White JH, et al. Opposite effects of histone deacetylase inhibitors on glucocorticoid and estrogen signaling in human endometrial Ishikawa cells. Mol Pharmacol 2005; 68(6):1852– [DOI] [PubMed] [Google Scholar]
- 47.Wang ZT, Chen ZJ, Jiang GM, Wu YM, Liu T, Yi YM, et al. Histone deacetylase inhibitors suppress mutant p53 transcription via HDAC8/YY1 signals in triple negative breast cancer cells. Cell Signal 2016;28(5):506–515. [DOI] [PubMed] [Google Scholar]
- 48.Fogh J, (eds). Human Tumor Cell in Vitro. New York, USA: Plenum Press; 1975. 550 p. [Google Scholar]
- 49.Cajot JF, Sordat I, Silvestre T, Sordat B. Differential display cloning identifies motility-related protein (MRP1/CD9) as highly expressed in primary compared to metastatic human colon carcinoma cells. Cancer Res 1997;57(13):2593–2597. [PubMed] [Google Scholar]
- 50.Brattain MG, Brattain DE, Fine WD, Khaled FM, Marks ME, Kimball PM, et al. Initiation and characterization of cultures of human colonic carcinoma with different biological characteristics utilizing feeder layers of confluent fibroblasts. Oncodev Biol Med 1981;2(5):355–366. [PubMed] [Google Scholar]
- 51.Brattain MG, Fine WD, Khaled FM, Thompson J, Brattain DE. Heterogeneity of malignant cells from a human colonic carcinoma. Cancer Res 1981;41(5):1751–1756. [PubMed] [Google Scholar]
- 52.Eshleman JR, Lang EZ, Bowerfind GK, Parsons R, Vogelstein B, Willson JK, et al. Increased mutation rate at the hprt locus accompanies microsatellite instability in colon cancer. Oncogene 1995; 10(1):33–37. [PubMed] [Google Scholar]
- 53.Ahmed D, Eide PW, Eilertsen IA, Danielsen SA, Eknaes M, Hektoen M, et al. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013;2(9):e71. [DOI] [PMC free article] [PubMed] [Google Scholar]