

Abstract
Motion information is essential in daily life because it provides cues to depth, timing, object identification, and self-motion, as well as input to the oculomotor system. As the peripheral visual field is exquisitely sensitive to motion, we investigated the periphery of individuals with central visual field loss (CFL) to determine whether speed and direction discrimination are intact in this population. We compared CFL participants' (N = 8), older (N = 6), and young controls' (N = 6) ability to discriminate motion speed and direction in a two-spatial-alternative forced-choice design. Participants viewed moving dots on the left and right of a fixation marker and judged which side had the faster speed or more clockwise direction. For the young control group, we repeated the experiment with the stimulus limited to thin strips of fixed width at eccentricities of 5°, 10°, and 15°. There was no significant difference in mean speed or direction discrimination thresholds of CFL participants and older controls for either velocity. Young controls had significantly lower thresholds than the CFL group for both tasks. We did not find an effect of visual acuity, viewing eccentricity, or scotoma location on individuals' ability to discriminate speed or direction. Our results indicate that for high-visibility stimuli moving at 5°–10°/s, speed and direction discrimination are intact in the periphery of individuals with CFL.
Keywords: speed discrimination, direction discrimination, motion perception, macular degeneration, peripheral vision
Introduction
Age-related macular degeneration (AMD) is the most common cause of vision loss in the developed world and affects nearly 7% of individuals over age 40 in the United States alone (Klein et al., 2011). Individuals with macular degeneration often experience a loss of their central visual field, with only peripheral vision remaining. Because of the prevalence of macular degeneration and its potentially devastating effects on the central visual field, it is important to understand how different aspects of visual function are affected in this population. One aspect important in daily life is the perception of motion (Nakayama, 1985). Some studies suggest that individuals with central field loss (CFL) are adept at using peripheral motion information, as in the case of vection (Tarita-Nistor, González, Markowitz, Lillakas, & Steinbach, 2008) and heading perception (Odom, Mali, & Leys, 2010). This view is consistent with a body of work that demonstrates the periphery's sensitivity to motion processing, especially at faster speeds (McKee & Nakayama, 1984).
In contrast to these findings, a study (Eisenbarth, MacKeben, Poggel, & Strasburger, 2008) aimed specifically at testing motion sensitivity in CFL found elevated contrast thresholds to judge motion direction. Specifically, they reported that motion contrast sensitivity was significantly reduced for eccentricities up to 20° in individuals with CFL, compared to age-matched and young controls. Thus, it appears that motion sensitivity is impaired well beyond the macula, which extends to less than 10° eccentricity (Strasburger, Rentschler, & Jüttner, 2011). However, the actual eccentricities of the stimulus in Eisenbarth et al. (2008) are uncertain, as the authors did not measure the eccentricity of the peripheral retinal locus (PRL), or monitor eye movements during the task. Although it is possible that their findings reveal deficits beyond the macula for stimuli close to contrast threshold, other work has found no differences in absolute detection thresholds at eccentricities greater than 10° in individuals with AMD (Sunness, Massof, Johnson, Finkelstein, & Fine, 1985). Thus, the combination of low stimulus contrasts and unknown stimulus eccentricity does not quite address whether motion perception for high contrast stimuli is impaired beyond the region of the scotoma.
Our own studies show that smooth pursuit in the CFL population is impaired compared to age-matched controls (Shanidze, Fusco, Potapchuk, Heinen, & Verghese, 2016); (Shanidze, Heinen, & Verghese, 2017). In these studies, we examined smooth pursuit during monocular and binocular viewing, and found that performance was impaired under all conditions, regardless of scotoma size or binocular overlap. Since smooth pursuit is a velocity-driven behavior (Beutter and Stone, 1998; Churchland & Lisberger, 2001), one might hypothesize that deficiencies in the perception of speed or direction might lead to this impairment. It is also possible that the lower gains may simply be due to the periphery's insensitivity to the lower speeds (McKee & Nakayama, 1984) characteristic of retinal slip. Therefore, the question remains whether this deficiency is due to other CFL-related limitations, such as a limited oculomotor range (Stahl, 2001; Whittaker, Budd, & Cummings, 1988), or an impairment of peripheral motion processing.
To probe potential impairments of motion processing in CFL, we set out to systematically measure sensitivity to speed and direction of motion. With a wealth of evidence suggesting that normal aging causes an impairment of motion perception and eye movements (Eisenbarth et al., 2008; Odom et al., 2010; Sharpe & Sylvester, 1978), we compared speed and direction discrimination in individuals with CFL to age-matched and young controls. We asked individuals to make speed and direction discrimination judgments while fixating a central target. To draw a direct parallel to our smooth pursuit experiments (Shanidze et al., 2017), we chose to stay in a similar velocity range of 5°/s and 10°/s. Although most studies of visual function in macular degeneration have measured visual function monocularly and related it to disease progression in that eye, we wanted to understand if motion perception is impaired under real-world circumstances. Therefore, we used a full-field, high-contrast stimulus that participants viewed binocularly, better approximating real-world conditions. We mapped out the binocular scotoma of our participants to understand the relationship of scotoma extent to motion perception under binocular viewing conditions. We hypothesized that under full-field conditions, individuals with CFL would perform similarly to age-matched controls, as in other studies comparing the two groups (e.g., Odom et al., 2010). Consistent with our hypotheses, we found that individuals with CFL and age-matched controls had similar performance in speed and direction discrimination, but were significantly worse than the young control group.
Having measured the eccentricity of the preferred retinal locus, we ensured viewing eccentricity from fixation (foveal or eccentric) by monitoring the eye movements of our participants. As scotomas varied significantly in size and location across participants, we examined whether restricting the information to a narrow region of the periphery affected motion perception. We varied the eccentricity (up to 15°) of narrow bands of the motion stimulus, and had our young control participants perform the task. Our results show that performance across tested eccentricities remained constant for the young control group.
We also used a binocular perimetry technique to measure the extent of the binocular scotoma on the same screen on which motion stimuli were presented. As we used a two-spatial alternative paradigm, with the stimuli to be discriminated on the left and right of fixation, we wondered whether individuals with CFL, whose scotoma fell more on one side than the other, had impaired motion perception on that side. Examination of the pattern of correct responses to the test stimulus on each side suggests that performance is not related to binocular scotoma size or location. Furthermore, we did not find a relationship between speed and direction discrimination thresholds and visual acuity or PRL distance of the better eye, suggesting that motion perception in the periphery is not affected by loss of central visual field.
Methods
Participants
All research was performed in accordance with the Declaration of Helsinki and was approved by the Institutional Review Board of the Smith-Kettlewell Eye Research Institute. We explained the nature and possible consequences of the study to each participant and obtained their signed consent. We recruited eight participants with central field loss (age range: 57–91, four females) and six age-matched controls (age range: 60–77, four females). Additionally, we recruited six young controls (YC, age range: 26–48, five females). All control participants had no eye movement disorders and had acuity that was normal, or corrected to normal. Seven CFL participants had macular degeneration (six with age-related macular degeneration and one with Stargardt disease) in one or both eyes, and one had macular edema in one eye and was blind in the other (Table 1). We were not able to complete an eye tracker calibration on the participant with macular edema (CFL1) and therefore did not perform binocular perimetry or monitor his fixation. His performance was close to the mean of the CFL group for both speed and direction discrimination. Therefore, we have included his data in the figures, but they are excluded from all statistical analyses. Participant YC3 dropped out of the study prior to completing the direction discrimination condition. Her speed data are included for analysis.
Table 1.
Participant information. Notes: Binocular scotoma maps: green dots represent seen flashes; red dots are missed flashes while fixating a center fixation cross with PRL/fovea (yellow asterisk). White Xs indicate the closest seen location within the motion discrimination stimulus on each side. For individuals with non-foveal PRLs, the distance of these points to the fovea is given in column 8, assuming PRL location of the dominant eye. White vertical lines show the location of the inner edges of the motion stimulus. Fuchsia triangles are original fovea locations (based on the dominant eye). §The grid spacing for CFL4 was smaller than 2°; therefore, the Xs show the first seen locations within the motion stimulus. However, we assume the individual could see all of the motion stimulus because he has a scotoma only in the left eye. *Participant AMC4 has vision corrected to 0.04 visual acuity and 1.8 contrast sensitivity; however, participant's corrective lenses did not allow for eye tracking. After confirming the participant could do the task comparably without correction, data was collected with gaze-monitoring but not corrective lenses.
| ID |
Age |
Sex |
Dx |
Binocular acuity (LogMAR) |
Binocular MARS contrast sensitivity |
Binocular fixation stability (deg2) |
Dominant eye PRL eccentricity (°), (x,y) |
Binocular scotoma |
| CFL1 | 71 | M | Macular edema | 0.9 | N/A | |||
| CFL2 | 76 | F | AMD (monocular) | 0 | 0.92 | 0.474 | 0 (0, 0) | |
| CFL3 | 86 | F | AMD | 0.4 | 1.40 | 3.239 | 6.24 (−6.2, −0.7) | |
| Closest viewing eccentricity: R = 0.83°, L = 9.01° | ||||||||
| CFL4 | 74 | M | AMD (monocular) | 0.1 | 1.32 | 10.966 | 0 (0, 0)§ | |
| CFL5 | 57 | M | Stargardt | 0.1 | 1.68 | 18.716 | 5.55 (−2.4, −5.0) | |
| Closest viewing eccentricity: R = 3.91, L = 6.04 | ||||||||
| CFL6 | 76 | M | AMD | 1.3 | 1.36 | 25.948 | 14.72 (−6.9, −13.0) | |
| Closest viewing eccentricity: R = 9.68, L = 14.51 | ||||||||
| CFL7 | 91 | F | AMD | 0.1 | 1.24 | 18.752 | 4.33 (−2.8, −3.3) | |
| Closest viewing eccentricity: R = 2.15, L = 6.98 | ||||||||
| CFL8 | 76 | F | AMD | 0.3 | 1.36 | 2.628 | 0.95 (−0.9, 0.3) | |
| Closest viewing eccentricity: R = 1.84, L = 3.71 | ||||||||
| AMC1 | 77 | M | 0.1 | 1.80 | 0.193 | 0 | ||
| AMC2 | 62 | F | 0.0 | 1.80 | 0.478 | 0 | ||
| AMC3 | 77 | F | −0.1 | 1.80 | 1.302 | 0 | ||
| AMC4 | 60 | M | 0.5* | 1.72 | 0.478 | 0 | ||
| AMC5 | 73 | F | 0.0 | 1.72 | 0.566 | 0 | ||
| AMC6 | 73 | F | 0.26 | 1.80 | 6.034 | 0 | ||
| YC1 | 30 | F | −0.1 | 1.70 | 0.102 | 0 | ||
| YC2 | 34 | F | −0.18 | 1.84 | 0.604 | 0 | ||
| YC3 | 29 | F | 0.0 | 1.84 | 0.304 | 0 | ||
| YC4 | 38 | F | 0.12 | 1.84 | 0.372 | 0 | ||
| YC5 | 26 | F | 0.0 | 1.92 | 2.219 | 0 | ||
| YC6 | 48 | M | −0.08 | 1.84 | 0.293 | 0 |
Prior to testing, all participants were screened using a standard battery of tests to measure acuity and contrast sensitivity (acuity inclusion criterion: 20/600 or higher). In the CFL group, monocular scotomas were mapped using standard microperimetry approaches, at 0 dB, with a scanning laser ophthalmoscope (Optos OCT/SLO). PRL eccentricity was measured from the fovea using the procedure outlined in Verghese, Tyson, Ghahghaei, and Fletcher (2016). Briefly, when possible, the foveal pit location was estimated from optical coherence tomography, and if the foveal pit could not be visualized, the location of the fovea was estimated using normative data of foveal distance from the center of the optic disc (Kabanarou et al., 2006). For individuals with central field loss, we assume similar eccentricity for binocular viewing as their better eye (Kabanarou et al., 2006; Tarita-Nistor, Brent, Steinbach, & González, 2012). For a summary of participant information, see Table 1.
Stimuli
Stimuli were presented at a distance of 1 m on a large projection screen (75.5 × 55 cm, or 41.36 × 30.75° of visual angle, see Figure 1A in Janssen & Verghese, 2014), in a two-spatial-alternative forced-choice design. Stimuli were created using MATLAB (MathWorks, Natick, MA) and Psychtoolbox-3.
Figure 1.

Full-field Stimulus: (A) For speed discrimination, participants judged which side of fixation had faster-moving dots (longer arrow, right). Screen, central blank region, and target sizes are denoted in white text. (B) For direction discrimination, participants judged which side had dots moving more downward (downward arrow, left).
Each participant's head was restrained comfortably in a chin and forehead rest, and eye movements were recorded monocularly using an EyeLink 1000 infrared eye tracker (SR Research, Ontario, Canada), placed in the tower mount configuration. All viewing was binocular, with participants' better (CFL) or dominant (control) eye being tracked. Data were sampled at 1000 Hz. The EyeLink 3-point calibration procedure was performed at the beginning of each block. Each trial presentation lasted 500 ms. To ensure viewing eccentricity, fixation was monitored throughout the trial. If the eye left the tolerance window (5° diameter) in the center of the screen at any time prior to the end of the trial, the trial was aborted and subsequently repeated at the end of the block (# aborted/block - CFL: median = 2, semi-interquartile range = 6.25, AMC: median = 1, semi-interquartile range = 1, YC: median = 0, semi-interquartile range = 0). The participant with the largest number of aborted trials was the oldest participant (CFL7, 91), who had frequent blinks and had trouble suppressing the impulse to look at the stimulus after onset.
Full-field speed and direction discrimination
On each trial, participants fixated a central fixation target (1° diameter) and viewed fields of moving dots on both sides of fixation (Figure 1). Dots were 0.5° in diameter and there were 1,000 dots per side (dot luminance: 3.6 cd/m2, background luminance: 259.2 cd/m2). Previous work has shown that spatial configuration of the stimulus has an effect on speed discrimination, with motion that appears to continue across a border being most difficult to discriminate (Verghese & McKee, 2006). The study further showed that separating the two parts of a display by a gap improved performance. Therefore, we had dots move in a direction 45° clockwise of horizontal on both sides of the screen and separated the two sides by a blank central region, 4° in width (Figure 1).
For speed-discrimination trials, dots on one side, chosen at random, moved at a reference speed of 5°/s or 10°/s and the dots on the other side moved at a faster test speed, selected from one of five possible speed increments. Speed increment values ranged from 1% to 50% of the reference speed, depending on the observer. Participants were asked to report whether the faster speed was on the left or the right side.
For direction-discrimination trials, dots on both sides of fixation moved at the same speed (either 5°/s or 10°/s, in separate blocks). On one side, chosen at random, the dots moved in the reference direction (45° clockwise from horizontal), while on the other, the test direction was rotated further clockwise by one of five possible increments (1°–30°), so that it had a more downward direction. Participants were asked to report whether the dots with a more vertical, downward direction of motion were on the left or right side.
Each block consisted of 50 trials. The average time to complete a block of trials varied in length across participant groups (CFL: 370.3 ± 204.1 s; AMC: 230.5 ± 43.5 s; YC: 189.3 ± 20.5 s), depending on the number of aborted trials and time to attain fixation. All participants were given the opportunity to practice prior to the first block for both the speed and direction conditions. After practice, participants in all three groups (CFL, age-matched control, and YC) performed a minimum of 50 trials of speed and direction discrimination, at each reference speed. Trials at each reference speed were presented within a single block. Eye tracker calibration was repeated at the beginning of each stimulus block. Several participants were asked to complete additional blocks in a speed or direction condition if their data were nonmonotonic or poorly fit by the Weibull psychometric function. The range of increment values was adjusted in the subsequent blocks to obtain a more reliable estimate of threshold. If a participant was too fatigued to complete a speed or direction condition on a given day, additional blocks were tested in a subsequent session. To report their responses, participants were asked to press the arrow key on a keyboard that corresponded to the faster/more downward direction (left and right arrows to indicate choices on the left and right, respectively).
Eccentricity-dependent speed and direction discrimination
The YC group performed additional trials where they viewed a more spatially restricted version of the stimulus already noted. For the eccentricity-dependent blocks, participants fixated as already mentioned, but were shown a narrow strip of moving dots on each side of fixation. The strip was centered at 5°, 10°, and 15° eccentricity and its width was m-scaled to compensate for cortical magnification for each eccentricity (2°, 3.25°, and 4.5°, respectively; Figure 2; Virsu & Rovamo, 1979). For eccentricity control trials, blocks consisted of 100 trials and were blocked by eccentricity. For each velocity, the full set of eccentricity blocks was preceded and followed by blocks of full-field stimulus trials, which were then compared to account for fatigue or learning effects. Participants were tested with dots moving at 5°/s, 10°/s, and 20°/s. The order of velocities and eccentricities at each velocity tested was randomized for each participant.
Figure 2.

Eccentricity-dependent stimulus. Moving dots were shown in strips centered at 5° (A), 10° (B), and 15° (C) eccentricities. Strips were 2°, 3.25°, and 4.5° in width, respectively.
Binocular perimetry and fixational stability
To determine the extent and location of binocular scotomas in our CFL group, we mapped their visual field. Individuals were seated 1 m from the same tangent screen and were asked to fixate the center, after performing a standard 5-point calibration. Fixation was monitored using the EyeLink 1000, with gaze being restricted to a central tolerance window of 3° in radius. Individuals were asked to respond every time they saw a flash (0.5°) on the screen. Flashes were displayed in an initial grid of 5 × 5 points that was centered on the display screen and spanned a 16.7° × 6.6° area. This area was centered on the fixation locus for each individual and spanned only a fraction of the total motion stimulus extent (represented by the black area in the perimetry maps in Table 1). Flashes were presented at a Weber contrast of ∼1. Each flash location was chosen at random and was repeated twice. Misses were counted and if a location was missed on both repetitions it was considered a potential binocular scotoma location. To probe this and neighboring locations at a finer scale, we used a finer grid of points—with half the spacing, in every quadrant with a potential scotoma. The finer grid was also repeated twice (maps are shown in the last column of Table 1; red dots indicate missed, and green dots indicate seen flash locations). As described in Section 2.1, for our CFL participants we determined the foveal location relative to fixation using the method of Verghese et al. (2016). Using this value we used the binocular perimetry map to calculate the closest visible location that was visible on the left and right sides of the motion stimulus (listed in column 8 and marked with white Xs in the last column of Table 1).
To determine the fixational stability of our participants during the binocular task, we aggregated 10 s of eye-position data during pretrial fixation periods for each participant for the first full block of trials. We then computed the 95% bivariate contour ellipse area (BCEA; Crossland, Sims, Galbraith, & Rubin, 2004) for these fixations to determine if fixation during the task was within the tolerance window of 28.3 deg2 (circular window, 5° diameter). The values are reported in column 7 of Table 1 for each participant.
Data analysis
For each participant, repeated blocks were grouped for analysis. The proportion correct versus speed (or direction) increments were fit with a Weibull function (Psychtoolbox-3) and threshold and slope fit parameters were recorded for further analysis. While thresholds can be reliably determined with as little as 50 trials, determining the slope estimates with the same reliability requires an order of magnitude more trials (Kontsevich & Tyler, 1999), so we do not discuss slope estimates further. The same analysis was performed for each eccentricity condition in young controls. Thresholds for each eccentricity and velocity combination were grouped across participants for comparison.
Comparison of right versus left visual field advantage
For individuals with CFL who have eccentric fixation, the binocular scotoma may be larger on one side of fixation. As the task in our study was to pick the side with the faster or more vertically moving test stimulus, it is possible that proportion of correct responses will be lower when the test stimulus (with the speed or direction change) occurs on the side with the larger scotoma. To specifically test this possibility, we analyzed proportion correct when the test was on the left or right side of the display. To do so, we selected all the difficult trials, defined as those below the estimated threshold of a given block, and calculated the proportion of correct responses when the test stimulus was on the left or the right side. For this analysis, we combined all full-field speed discrimination blocks together and all full-field direction discrimination blocks together for each participant. To determine if performance was unbiased (equal proportion correct on the left and right), we calculated the 95% confidence interval for each value of proportion correct, assuming a Binomial Error Distribution: sqrt[%Corr(1-%Corr)/N], where N is the number of trials at each increment value.
Results
Speed discrimination
Figure 3A is a summary of threshold values for the full-field condition, across the three participant groups and two velocities (blue: 5°/s; red: 10°/s) We used a two-way analysis of variance (ANOVA; factors: participant group and stimulus velocity) with repeated measures and a Tukey adjustment for multiple comparisons to look at discrimination thresholds. When we compared the data between groups, we found no significant difference between individuals with CFL and age-matched controls for both tested velocities (5°/s: p = 0.376, 10°/s: p = 0.647). For each group, performance was not significantly different between the two tested velocities (p = 0.693). There was a significant difference between CFL and young control groups' thresholds for both 5°/s (p = 0.004; meanCFL = 0.237 ± 0.087, meanControl = 0.181 ± 0.039, meanYC = 0.096 ± 0.040) and 10°/s (p = 0.012; meanCFL = 0.237 ± 0.100, meanControl = 0.207 ± 0.067, meanYC = 0.125 ± 0.061). We repeated the analysis with participant CFL1 included and arrived at the same outcome.
Figure 3.
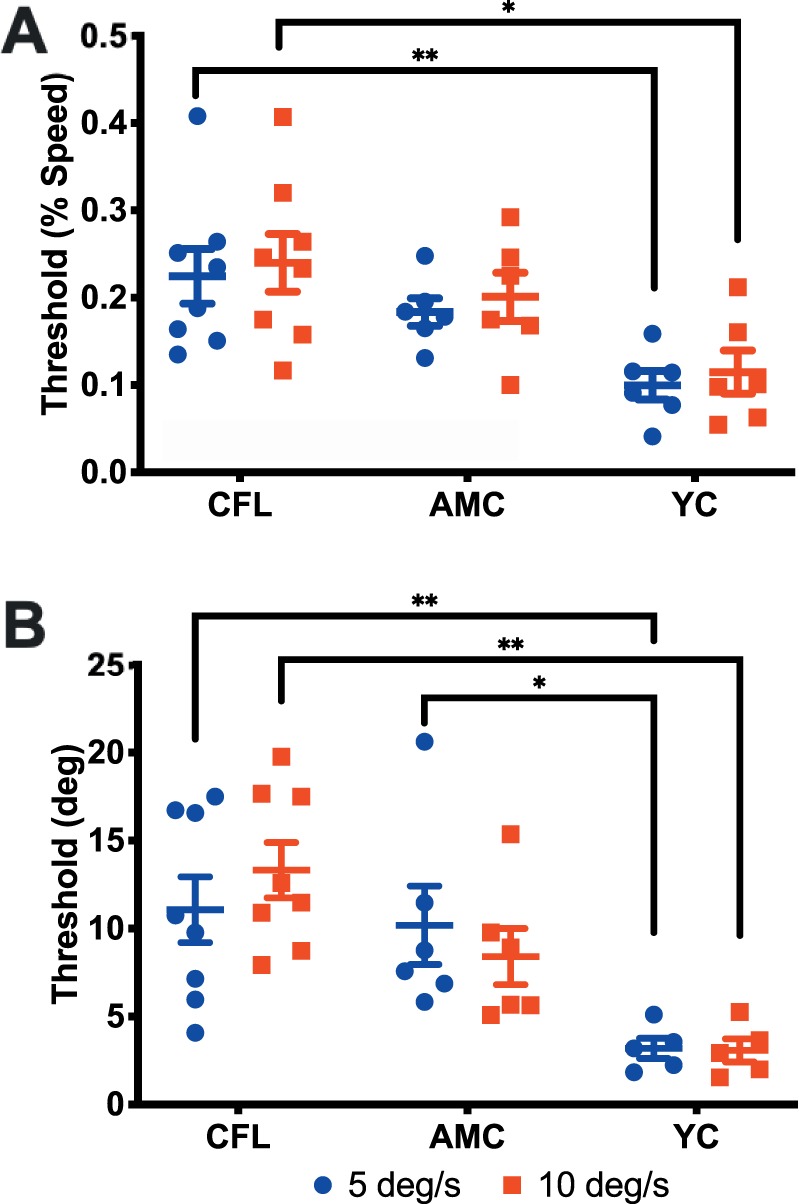
Speed (A) and direction (B) discrimination thresholds for the three tested groups. Blue and red points represent individual data for the 5°/s and 10°/s conditions, respectively. Error bars: standard error of the mean.
Direction discrimination
We saw a very similar pattern of results for the direction discrimination condition (Figure 3B). There was a significant difference between the CFL and young control groups for both 5°/s (p = 0.009; meanCFL = 11.249 ± 5.682, meanControl = 10.711 ± 5.466, meanYC = 3.185 ± 1.282) and 10°/s (p = 0.001; meanCFL = 13.583 ± 4.724, meanControl = 8.958 ± 3.915, meanYC = 2.930 ± 1.655). Additionally, there was a significant difference between performance in age-matched and young controls in the 5°/s condition (p = 0.033). Outcomes were the same when participant CFL 1 was included in the analysis.
Eccentricity-dependent speed and direction discrimination
To test whether stimulus eccentricity alone might affect performance, we examined speed and direction discrimination performance when the stimulus was restricted to narrow strips at each of three different eccentricities. In addition to the two velocities tested previously (5°/s and 10°/s), we also included a velocity of 20°/s. Because this experiment involved a significant number of additional testing blocks, only the young control (YC) group participated. We found no significant difference in speed or direction discrimination thresholds (Figure 4) across eccentricities for all tested velocities (p > 0.1, two-way ANOVA with repeated measures and a Tukey adjustment for multiple comparisons).
Figure 4.

Speed (A) and direction (B) discrimination thresholds at the three tested eccentricities in young controls. Stimulus velocities were 5°/s, 10°/s, and 20°/s (blue, red, and green, respectively). Thin, dashed lines represent each participant. Thick lines are group means; error bars: standard error of the mean.
Additionally, because each participant did an extensive number of eccentricity-restricted trials, we wanted to determine if there were any improvements in discrimination performance over time due to learning. Specifically, we measured full-field speed and direction discrimination before and after we tested the three eccentricities at a given speed. This procedure was repeated for each of the three velocities tested (5°/s, 10°/s, 20°/s). For speed discrimination, there was no significant difference in full-field thresholds between the pre- and posteccentricity-restricted blocks, at any of the three velocities. For direction discrimination, the 5°/s and 10°/s conditions showed no significant difference in full-field thresholds before and after the eccentricity-restricted trials. Only the 20°/s condition showed an improvement for the full-field block at the end of the eccentricity trials compared to the beginning, (p = 0.008, mean1 = 4.76, mean2 = 2.24, two-way ANOVA with repeated measures, and Sidak correction for multiple comparisons). This finding is unlikely to be indicative of an overall improvement in performance over time because of the counterbalanced design in which the three stimulus velocities were tested. Participants for whom we saw this improvement did not all do the 20°/s block in the same order. In fact, for the three individuals for whom the effect was evident, one did the 20°/s block first, one did it second, and one did it as the last block.
To determine if there was a difference in discrimination performance between viewing the full-field version and narrow strips of the stimulus at any of the eccentricities, we compared each eccentricity condition to the initial full-field test. For all three tested velocities, we found no difference in performance across conditions (p > 0.05, two-way ANOVA, Tukey correction for multiple comparisons).
Acuity and PRL distance do not affect motion perception
To examine whether individual variations in speed and direction discrimination were related to vision loss in the participants with CFL, we analyzed speed and direction discrimination thresholds with respect to binocular visual acuity and eccentricity of the PRL in the better eye. Data, along with the regression fits, are plotted for each observer in Figure 5. An inspection of the aggregate data for both participants with CFL and age-matched controls does not show a significant relationship between either speed or direction discrimination thresholds and binocular visual acuity (p > 0.5) or PRL eccentricity of the better eye (p > 0.15). To ensure that these results were not skewed by the control data, we repeated the analysis only for those with CFL. The outcome of this analysis was consistent with the aggregate data—there was no significant relationship for any of the comparisons (p > 0.2).
Figure 5.

Speed and direction discrimination thresholds are plotted as a function of binocular visual acuity (A–B) and PRL eccentricity of the better eye (C–D). CFL and age-matched control participants are shown in maroon and orange, respectively. Black lines indicate linear regressions done for the combined data set, none of which is significantly different from zero.
No influence of scotoma location on motion perception
Individuals with CFL sometimes use a PRL that places the scotoma on one side of fixation. In our experiments, we wanted to determine whether individuals for whom the scotoma was larger on one side of fixation relative to the other (left vs. right) would have an impaired ability to perceive motion on that side. To this end, we examined whether CFL participants were more likely to be correct when the test stimulus was on a given side, and if so, if there was a correlation between scotoma size/location and size/direction of the response bias.
To test this hypothesis, we analyzed each group's tendency for errors on each side (Figure 6). Figure 6A and B shows proportion correct responses when the test stimulus was on the right versus the left side for speed (A) and direction (B) discrimination. Error bars are the 95% confidence intervals to indicate if there is a significant deviation from equal performance to test stimuli on the two sides (black diagonal line). For all three groups, we saw a tendency for more correct responses when the test stimulus was on the left side of the display. To determine if this tendency was different across groups, we calculated the ratio of proportion correct responses on the left versus the right, for each participant. These data are shown in Figure 6C (speed discrimination) and D (direction discrimination). Consistent with our observation of a left-side preference, the majority of the points for all groups are located above the y = 1 line, which indicates equal proportion correct on both sides. Although this tendency is particularly apparent for the CFL group, the distributions of the three groups are not significantly different from each other (Kruskal-Wallis one-way analysis of variance, speed: p = 0.054; direction: p = 0.782).
Figure 6.

Proportion correct on the right versus left side for speed (A) and direction (B) discrimination trials for CFL (maroon), age-matched control (orange) and young control (teal) participants. Number labels refer to CFL participants in Table 1. Black line: equivalent performance. Error bars: 95% CI. (C–D) Summary of proportion correct data in (A) and (B), respectively. Each point represents a ratio of proportion correct responses on the left versus the right. The horizontal, black line indicates equal correct responses on the left and right. Horizontal lines in maroon, orange, and teal indicate the median ratios for the three groups.
We observe that for three individuals with CFL the proportion correct was significantly greater when the test stimulus was on the left side of the display both speed and direction discrimination (the 95% confidence intervals do not intersect the diagonal line that indicates equal thresholds on the left and right). These participants' data are numbered in the first column of Figure 6A and B.
One possibility is that under conditions of eccentric viewing when individuals are fixating with a peripheral PRL, the binocular scotoma may be larger on one side of fixation than the other. The task in our study was to pick the side with the faster or more vertically moving test stimulus. The reference speed (or direction) was always fixed across a block of trials, while the test speed (or direction) varied. Previous studies have shown that in cases where the reference is fixed, observers internalize this reference value, and are able to base their discrimination on the test stimulus alone (Morgan, Watamaniuk, & McKee, 2000). If the test stimulus with the variable velocity or speed fell on the side with the larger scotoma, it may explain why proportion of correct responses was lower on that side. An examination of the binocular scotoma location for the three individuals with the significant bias for speed and direction discrimination shows that the biased performance is unrelated to scotoma location or size (Table 1, CFL). One individual has no binocular scotoma (4 in Figure 6, blue points), the second has a moderately sized right binocular scotoma (labeled 3 in Figure 6, blue points), and the third individual (5 in Figure 6, blue points) has a binocular scotoma in the upper visual field that is roughly symmetrical on the right and left.
Discussion
Motion perception in central field loss
Our data indicate that for high-visibility stimuli, individuals with central field loss are able to make speed and direction discrimination judgments as well as their age-matched controls at speeds as slow as 5°/s. It is of note, that despite significantly worse fixational stability, the CFL group performed comparably on the motion discrimination tasks to the age-matched controls. Our finding that sensitivity to fine changes in speed and direction is intact in individuals with CFL is consistent with previous data that show that when cortical magnification is taken into account, velocity discrimination is as precise in the periphery as it is at the fovea (McKee & Nakayama, 1984). Furthermore, a study looking at heading precision in AMD found that in the absence of added noise, individuals with AMD could locate the focus of expansion with comparable precision to their age-matched controls (Odom et al., 2010). The results of our study with high visibility stimuli, taken together with the results of Odom et al. (2010) indicate that as long as the stimulus is visible to observers with CFL, their ability to discriminate small changes in speed, direction, and heading is similar to age-matched controls.
However, our findings are somewhat at odds with previous work that suggested that motion sensitivity is decreased due to damage from macular degeneration as far as 20° out in the periphery (Eisenbarth et al., 2008). Eisenbarth and colleagues found that contrast thresholds for direction discrimination were significantly elevated in AMD for eccentricities up to 20°, for stimuli moving at 5.7°/s (Eisenbarth et al., 2008). The implication from this study is that it is not just the macula, which extends to ∼9° eccentricity, but the periphery beyond that is affected in macular degeneration. This result is at odds with the report from Sunness et al. (1985) that thresholds for absolute detection of briefly flashed stimuli were unimpaired beyond 10° eccentricity in individuals with AMD. Furthermore, Watson and Robson (1981) showed that thresholds for judging the direction of motion are close to thresholds for absolute detection, making the Sunness et al. finding on absolute thresholds particularly relevant to the contrast thresholds for judging motion direction in the Eisenbarth et al. study.
The fact that Eisenbarth and colleagues found elevated thresholds for motion direction judgment well outside the macula suggests two possibilities. The first is that there are subtle changes in sensitivity that can be picked up only with more sensitive measurements near contrast threshold. The Sunness et al. (1985) result argues against this possibility as thresholds for absolute detection were found to be normal in individuals with CFL for eccentricities beyond 10°. Eisenbarth et al. make the case that unlike the Sunness et al. result, their results indicate “deficits of dynamic visual field properties” in age-related macular degeneration, as they also find elevated thresholds for double-pulse resolution and lower values for critical flicker frequency at eccentricities beyond the scotoma. However, as the stimuli for both of these experiments (double pulse threshold and critical flicker frequency) were not full-field, but small (∼1°) and local, it still leaves open the possibility that their exact eccentricity was unknown.
Thus, the second possibility is that the difference between the Eisenbarth findings and ours is that the true eccentricity of the small (3.8° diameter) direction-discrimination stimulus was unknown in their study. While their stimuli were presented at specific eccentricities from the fixation marker and performance was compared at these stimulus eccentricities for all participant groups, the actual viewing eccentricity was unknown as they did not measure the eccentricity of the PRL in individuals with macular degeneration. Thus, it is unclear what part of the retina was used to fixate the cross and how far this locus was from the original fovea. Additionally, eye movements were not monitored with an eye tracker, making it difficult to estimate the true eccentricity of the stimulus. Accordingly, the difference in our finding and theirs could well be due to a misestimation of true stimulus eccentricity in the study by Eisenbarth and colleagues. In our experiments, all observers were presented with a large, full-field stimulus and a central fixation marker. As we know the approximate location of the PRL from the original fovea in individuals with CFL, we can estimate directly the eccentricity of the two fixed inner edges of the motion stimulus. For our participants with CFL, we find the largest eccentricity for the inner left and right edges of the stimulus to be 14.51° and 9.68°, respectively (CFL6, Table 1). CFL participants' performance at these eccentricities is consistent with our results in young controls where motion discrimination performance did not change for thin stimulus strips up to 15° eccentricity.
There is additional, indirect evidence of spared peripheral visual function from the oculomotor domain. For optokinetic nystagmus, a large study of individuals with AMD who had scotomas ranging from none to 30° in diameter, showed that scotoma size did not impact the gain of the optokinetic response at a stimulus velocity of 15°/s. At faster speeds, only scotomas greater than 20° in diameter had significant impact on the OKN response (Valmaggia, Charlier, & Gottlob, 2001). In another set of studies, the authors found that high speed motion (45°/s–60°/s) induced vection responses that were as strong in CFL observers as they were in controls (Tarita-Nistor et al., 2008).
Effects of aging
We do find, however, that age is a significant factor in motion perception, with our group of younger controls having significantly better performance in both tasks. This data adds to a mounting set of evidence that suggests that age plays a significant role in motion perception and related oculomotor behaviors. Eisenbarth and colleagues found that individuals in their young control group (mean age: 27) performed significantly better than age-matched controls across all tested eccentricities (Eisenbarth et al., 2008). It is significant to point out that since both control groups used the fovea for fixation, the comparison across eccentricities was consistent for these groups. Odom and colleagues found a similar pattern, with younger controls (ages 21–59) having lower thresholds for heading judgments than the older control group. Bennett and colleagues looked at motion detection in a group of observers whose ages ranged from 23 to 81 years (Bennett, Sekuler, & Sekuler, 2007). Participants viewed random dot cinematograms and had to discriminate whether the stimulus had a coherent direction or was random noise. They also had to identify the direction of motion of the coherently moving stimulus. Individuals older than age 70 were significantly worse than all other age groups at determining whether a signal was present and at identifying the direction of motion.
In the oculomotor domain, a study that compared a group of younger individuals (ages 19–32) to an older group (ages 65–77) found a significant decrement in smooth pursuit performance in the older group (Sharpe & Sylvester, 1978). The authors tested a large range of pursuit velocities (5°/s–50°/s) and found a significant difference in performance between the two age groups for all but the slowest velocity. Combined, these studies raise the question as to whether the decrease in smooth pursuit gain with age is due to an age-related decline in motion perception. In our own studies of smooth pursuit (Shanidze et al., 2016; Shanidze et al., 2017), we observed comparable gains in older, healthy-sighted participants to those reported by Sharpe and Sylvester. However, we found that pursuit gains in CFL were lower still. We explore the relationship between pursuit gain and motion perception in CFL in the section in this paper titled, “Smooth pursuit gain and motion perception in CFL.”
Since age is a significant factor in outcomes on our speed and direction discrimination tasks, we wanted to explore whether the imperfect age-match between our CFL and older control groups has bearing on our results. Although both groups have a very similar lower-bound in their age ranges, we were not able to find a nonagenarian control without vision deficits (glaucoma, diabetic retinopathy, or macular disease). Based on our findings, as well as those discussed already, one might expect that the higher mean age in the CFL group might lead to higher speed and direction discrimination thresholds in the CFL group compared to the older controls. However, despite a higher mean age in the CFL group, we do not see significant differences between the two older participant groups. To ensure that the age discrepancy did not affect our results, we repeated our analyses comparing speed and direction discrimination thresholds across groups without including the oldest CFL participant (CFL7). The outcomes of our analyses were in complete agreement with the trends reported already.
Effects of scotoma size and location
Central field loss is associated with a loss of visual information that can vary in extent and location. In our group of participants, individuals had damage that ranged from no binocular field loss, to a binocular scotoma of nearly 25° in diameter. Furthermore, depending on the placement of the PRL relative to scotoma, the eccentricity of the visual field loss could vary considerably. There are two questions that arise. First, does limiting the amount of visible motion information or its eccentricity affect performance? Second, does scotoma location in the field affect performance?
To address the first question, we asked individuals in our young control group to perform the speed and direction discrimination tasks with only thin strips of moving dots visible on each trial. We varied the eccentricity of these strips between 5° and 15° to reflect our CFL population and scaled the strip width to account for cortical magnification. For velocities of 5°/s, 10°/s, and 20°/s we did not see an effect of viewing eccentricity on performance. Furthermore, when we compared full-field versus restricted-field conditions, we found speed and direction discrimination performance to be comparable. This latter finding is unsurprising, given previous research that shows that speed discrimination at a given eccentricity is roughly independent of stimulus size in the absence of noise (Verghese & Stone, 1996). In that study, Verghese and Stone showed that increasing the size of a peripheral moving grating by a factor of six caused no improvement in speed discrimination thresholds.
To address the second question, we examined whether individuals with CFL had impaired performance at the location of their scotoma. Specifically, we measured performance for conditions when the test stimulus was on the left or the right of the screen. As the reader might recall, in our experiments the reference speed and direction were fixed within each block and observers were asked to determine which side had the faster test speed or more vertical direction. Morgan and colleagues have shown that when the reference is fixed, it is superfluous and observers make judgements based on an implicit reference (Morgan et al., 2000). For this reason, we analyzed performance relative to the test side. Our initial prediction was that individuals with greatest and most asymmetrical damage to their visual field would show the greatest imbalance in performance. We did find that three individuals with CFL had significantly lower proportion correct on the right side compared to the left for both speed and direction discrimination tasks. However, contrary to our prediction, these individuals with the strongest bias had either no, small, or symmetrical binocular scotomas. This analysis, combined with our analysis of thresholds as a function of binocular visual acuity and PRL eccentricity of the better eye suggests that there is no clear relationship between motion discrimination and scotoma location or fixational eccentricity.
In fact, the left-visual advantage is not unique to the CFL group; we found that all three participant groups showed an overall trend towards higher proportion correct on the left (Figure 6C and D). One possible explanation could be a known left visual-field advantage in motion perception. Bosworth, Petrich, and Dobkins, (2012) showed that the left visual-field advantage for motion discrimination was apparent only under demanding dual-task conditions when observers had to perform a shape counting task at fixation, as well as discriminate the direction of a peripheral stimulus. On the other hand, no left versus right hemifield effects were found when observers directed their full attention to the peripheral task in the absence of a central task. A left-visual field advantage was also found for a temporal order judgment task in the periphery (Matthews & Welch, 2015) under conditions of high attentional load with a demanding letter identification task at fixation. While we did not have an explicit task at fixation, we wondered whether the requirement to maintain fixation was more demanding for individuals with CFL because of their poor fixation stability. An inspection of fixation stability shows that the individuals with CFL who showed the largest left bias had good to moderate fixation stability (see Table 1). Furthermore, individuals with the largest fixation instability (and largest scotoma) had no bias. Although maintaining fixation may be more challenging for those individuals with poor fixational stability it may not rise to the level of difficulty to the central task in Bosworth et al. (2012) and Matthews and Welch (2015). As such, it is unlikely that fixation instability can explain the left bias in our task.
Smooth pursuit gain and motion perception in CFL
The deficits in smooth pursuit gain in individuals with CFL do not appear to be due to deficits in motion perception. Specifically, the retinal slip velocities resulting from the lower gains in CFL are likely too low to be detected by the periphery, which is sensitive to faster speeds. Therefore, the additional decrement that we see in smooth pursuit performance may be due to the loss of the central visual field, which is sensitive to slower speeds. Four of the CFL participants in this study (CFL4, CFL5, CFL6, CFL7) were also tested on the binocular smooth pursuit task described in Shanidze et al. (2017). When we examined retinal slip speeds for targets moving at 5°/s and 10°/s for these four individuals compared to the average retinal slip for controls in that same experiment, we found significantly higher retinal slip speeds in the CFL group. However, we did not find greater retinal slip to be correlated with these individuals' ability to discriminate speed or direction in the present study.
The deficits in smooth pursuit can be caused by perceptual and or oculomotor factors. This study examines whether there is a deficit in motion perception and shows that individuals with CFL are able to get accurate motion information about speed and direction for stimuli moving at speeds of 5°/s or faster, suggesting that motion perception is intact for high contrast, full-field stimuli. Therefore, we believe that oculomotor factors may account for the deficits in smooth pursuit. For example, when the PRL (and therefore eye position in the orbit) is eccentric, individuals have a limited oculomotor range that may contribute to lower smooth pursuit gains (Guitton & Volle, 1987; Shanidze, Ullman, Badler, & Verghese, 2018; Stahl, 2001; Whittaker et al., 1988). Additionally, learning to use the PRL as a reference for eye movements may contribute to oculomotor deficits in CFL (White & Bedell, 1990). Examining the contribution of oculomotor deficits to smooth pursuit is outside the scope of this study and further research is needed to understand what other factors cause the deficits in smooth pursuit gain in CFL.
Limitations
There are several limitations that must be considered. Our study uses a relatively small group of individuals with CFL. However, our small participant group includes a range of scotoma sizes and overlap, from people with no binocular scotoma, to those with a large binocular scotoma. Additionally, we included individuals with overlapping and nonoverlapping scotomas of varying sizes and individuals who used eccentric fixational loci and those who are still able to use the fovea in one eye. For this diverse CFL participant group and the control participants, we found that age was the single determining factor of performance—a highly robust finding despite limited participant numbers. Due to the very nature of progression of central field loss, we were not able to equate fixation eccentricity across participants for CFL. Instead, we utilized a large, full-field stimulus that could recruit a large portion of the residual retina. Our peripheral eccentricity result with the young control group further supports our finding that viewing eccentricity does not affect performance for speed or direction discrimination under conditions of high stimulus visibility.
Conclusions
Central field loss has been shown to extensively affect performance of daily tasks that require high acuity vision (Mangione, Gutierrez, Lowe, Orav, & Seddon, 1999). This decrement is the direct consequence of damage to the central portion of the retina. However, less is known about how CFL might affect tasks such as motion processing, for which the normal periphery is exquisitely sensitive. In this study we set out to assess speed and direction discrimination performance in CFL under real-world visibility conditions, and found that performance of observers with CFL is comparable to that of their age-matched controls. While our study does not address the possibility of subtler deficits near the threshold of visibility, it was designed to emulate motion in the real world, where information tends to be redundant and of high contrast. Our findings suggest that under these conditions, individuals with CFL should be able to perform motion-related tasks comparably to their contemporaries with normal vision.
Acknowledgments
This work was supported by National Institutes of Health (NIH) grant R00 EY026994 to NS and R01 EY027390 to PV.
Commercial relationships: none.
Corresponding author: Natela Shanidze.
Email: natela@ski.org.
Address: Smith-Kettlewell Eye Research Institute, San Francisco, CA, USA.
Contributor Information
Natela Shanidze, natela@ski.org, https://www.ski.org/users/natela-shanidze.
Preeti Verghese, preeti@ski.org, https://www.ski.org/users/preeti-verghese.
References
- Bennett P. J, Sekuler R, Sekuler A. B. The effects of aging on motion detection and direction identification. Vision Research. (2007);47(6):799–809. doi: 10.1016/j.visres.2007.01.001. [DOI] [PubMed] [Google Scholar]
- Beutter B. R, Stone L. S. Human motion perception and smooth eye movements slow similar directional biases for elongated apertures. Vision Research. (1998);38(9):1273–1286. doi: 10.1016/S0042-6989(97)00276-9. [DOI] [PubMed] [Google Scholar]
- Bosworth R. G, Petrich J. A. F, Dobkins K. R. Effects of spatial attention on motion discrimination are greater in the left than right visual field. Vision Research. (2012);52(1):11–19. doi: 10.1016/j.visres.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churchland M. M, Lisberger S. G. Experimental and computational analysis of monkey smooth pursuit eye movements. Journal of Neurophysiology. (2001);86(2):741–759. doi: 10.1152/jn.2001.86.2.741. [DOI] [PubMed] [Google Scholar]
- Crossland M. D, Sims M, Galbraith R. F, Rubin G. S. Evaluation of a new quantitative technique to assess the number and extent of preferred retinal loci in macular disease. Vision Research. (2004);44(13):1537–1546. doi: 10.1016/j.visres.2004.01.006. [DOI] [PubMed] [Google Scholar]
- Eisenbarth W, MacKeben M, Poggel D. A, Strasburger H. Characteristics of dynamic processing in the visual field of patients with age-related maculopathy. Graefe's Archive for Clinical and Experimental Ophthalmology. (2008);246(1):27–37. doi: 10.1007/s00417-007-0641-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guitton D, Volle M. Gaze control in humans: Eye-head coordination during orienting movements to targets within and beyond the oculomotor range. Journal of Neurophysiology. (1987);58(3):427–459. doi: 10.1152/jn.1987.58.3.427. [DOI] [PubMed] [Google Scholar]
- Janssen C. P, Verghese P. Stop & think: Looking into a scotoma. Journal of Vision. (2014);14(10):222. https://doi.org/10.1167/14.10.222 Abstract. [Google Scholar]
- Kabanarou S. A, Crossland M. D, Bellmann C, Rees A, Culham L. E, Rubin G. S. Gaze changes with binocular versus monocular viewing in age-related macular degeneration. Ophthalmology. (2006);113(12):2251–2258. doi: 10.1016/j.ophtha.2006.06.028. [DOI] [PubMed] [Google Scholar]
- Klein R, Chou C.-F, Klein B. E. K, Zhang X, Meuer S. M, Saaddine J. B. Prevalence of age-related macular degeneration in the US population. Archives of Ophthalmology. (2011);129(1):75–80. doi: 10.1001/archophthalmol.2010.318. [DOI] [PubMed] [Google Scholar]
- Kontsevich L. L, Tyler C. W. Bayesian adaptive estimation of psychometric slope and threshold. Vision Research. (1999);39(16):2729–2737. doi: 10.1016/s0042-6989(98)00285-5. [DOI] [PubMed] [Google Scholar]
- Mangione C. M, Gutierrez P. R, Lowe G, Orav E. J, Seddon J. M. Influence of age-related maculopathy on visual functioning and health-related quality of life. International Congress Series. (1999);128(1):45–53. doi: 10.1016/S0002-9394(99)00169-5. [DOI] [PubMed] [Google Scholar]
- Matthews N, Welch L. Left visual field attentional advantage in judging simultaneity and temporal order. Journal of Vision. (2015);15(2):1–13.:7. doi: 10.1167/15.2.7. https://doi.org/10.1167/15.2.7 PubMed] [ Article. [DOI] [PubMed] [Google Scholar]
- McKee S. P, Nakayama K. The detection of motion in the peripheral visual field. Vision Research. (1984);24(1):25–32. doi: 10.1016/0042-6989(84)90140-8. [DOI] [PubMed] [Google Scholar]
- Morgan M. J, Watamaniuk S. N, McKee S. P. The use of an implicit standard for measuring discrimination thresholds. Vision Research. (2000);40(17):2341–2349. doi: 10.1016/S0042-6989(00)00093-6. [DOI] [PubMed] [Google Scholar]
- Nakayama K. Biological image motion processing: a review. Vision Research. (1985);25(5):625–660. doi: 10.1016/0042-6989(85)90171-3. [DOI] [PubMed] [Google Scholar]
- Odom J. V, Mali J. O, Leys M. J. Heading precision in age-related macular degeneration (AMD): Effects of directional noise. Investigative Ophthalmology & Visual Science. (2010);51(13):3619–3619. [Google Scholar]
- Shanidze N, Fusco G, Potapchuk E, Heinen S, Verghese P. Smooth pursuit eye movements in patients with macular degeneration. Journal of Vision. (2016);16(3):1–14.:1. doi: 10.1167/16.3.1. https://doi.org/10.1167/16.3.1 PubMed] [ Article. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shanidze N, Heinen S, Verghese P. Monocular and binocular smooth pursuit in central field loss. Vision Research. (2017);141:181–190. doi: 10.1016/j.visres.2016.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shanidze N, Ullman E, Badler J, Verghese P. Interaction of eye and head movements during smooth pursuit in macular degeneration. Investigative Ophthalmology & Visual Science. (2018);59(9):4412. [Google Scholar]
- Sharpe J. A, Sylvester T. O. Effect of aging on horizontal smooth pursuit. Investigative Ophthalmology & Visual Science. (1978);17(5):465–468. Retrieved from http://eutils.ncbi.nlm.nih.gov/entrez/eutils/elink.fcgi?dbfrom=pubmed&id=640792&retmode=ref&cmd=prlinks. [PubMed] [Google Scholar]
- Stahl J. S. Eye-head coordination and the variation of eye-movement accuracy with orbital eccentricity. Experimental Brain Research. (2001);136:200–210. doi: 10.1007/s002210000593. [DOI] [PubMed] [Google Scholar]
- Strasburger H, Rentschler I, Jüttner M. Peripheral vision and pattern recognition: A review. Journal of Vision. (2011);11(5):1–82.:13. doi: 10.1167/11.5.13. https://doi.org/10.1167/11.5.13 PubMed] [ Article. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sunness J. S, Massof R. W, Johnson M. A, Finkelstein D, Fine S. L. Peripheral retinal function in age-related macular degeneration. Archives of Ophthalmology. (1985);103(6):811–816. doi: 10.1001/archopht.1985.01050060071029. [DOI] [PubMed] [Google Scholar]
- Tarita-Nistor L, Brent M. H, Steinbach M. J, González E. G. Fixation patterns in maculopathy: From binocular to monocular viewing. Optometry and Vision Science. (2012);89(3):277–287. doi: 10.1097/OPX.0b013e318244e8b1. [DOI] [PubMed] [Google Scholar]
- Tarita-Nistor L, González E. G, Markowitz S. N, Lillakas L, Steinbach M. J. Increased role of peripheral vision in self-induced motion in patients with age-related macular degeneration. Investigative Ophthalmology & Visual Science. (2008);49(7):3253–3258. doi: 10.1167/iovs.07-1290. [DOI] [PubMed] [Google Scholar]
- Valmaggia C, Charlier J, Gottlob I. Optokinetic nystagmus in patients with central scotomas in age related macular degeneration. The British Journal of Ophthalmology. (2001);85(2):169–172. doi: 10.1136/bjo.85.2.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verghese P, McKee S. P. Motion grouping impairs speed discrimination. Vision Research. (2006);46(8-9):1540–1546. doi: 10.1016/j.visres.2005.07.029. [DOI] [PubMed] [Google Scholar]
- Verghese P, Stone L. S. Perceived visual speed constrained by image segmentation. Nature. (1996);381(6578):161–163. doi: 10.1038/381161a0. [DOI] [PubMed] [Google Scholar]
- Verghese P, Tyson T. L, Ghahghaei S, Fletcher D. C. Depth perception and grasp in central field loss. Investigative Ophthalmology & Visual Science. (2016);57(3):1476–1487. doi: 10.1167/iovs.15-18336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virsu V, Rovamo J. Visual resolution, contrast sensitivity, and the cortical magnification factor. Experimental Brain Research. (1979);37(3):475–494. doi: 10.1007/BF00236818. [DOI] [PubMed] [Google Scholar]
- Watson A. B, Robson J. G. Discrimination at threshold: Labelled detectors in human vision. Vision Research. (1981);21(7):1115–1122. doi: 10.1016/0042-6989(81)90014-6. [DOI] [PubMed] [Google Scholar]
- White J. M, Bedell H. E. The oculomotor reference in humans with bilateral macular disease. Investigative Ophthalmology & Visual Science. (1990);31(6):1149–1161. [PubMed] [Google Scholar]
- Whittaker S. G, Budd J, Cummings R. W. Eccentric fixation with macular scotoma. Investigative Ophthalmology & Visual Science. (1988);29(2):268–278. [PubMed] [Google Scholar]