

Recently we 1 reported experiments on the revival of the dog's heart from ventricular fibrillation. Briefly summarized, it was shown that ventricular fibrillation produced by faradic stimulation can be abolished by intracardiac injection of a 5% KCl solution in doses of 1 cc per kilogram. Massage was not required in order to produce these effects, but injections into both ventricular cavities appeared to be essential for prompt action. After every trace of fibrillation had disappeared, a subsequent injection into both ventricular chambers of a 5% CaCl2 and 0.1% heparin solution restored a coordinated beat, only, however, when supplemented by cardiac massage.
This article is chiefly concerned with detailed studies of the fibrillary process, the alterations effected by potassium before complete inhibition occurred, and the mechanisms concerned in revival. Apart from their purely scientific interest, these and subsidiary problems are of practical importance as well, because the basic changes involved in the cardiac mechanism supply scientific proof that the reported resuscitations could by no chance have been a matter of fortunate spontaneous revival.
THE LIMITATIONS AND POSSIBILITIES OF GRAPHIC METHODS APPLIED TO FIBRILLATING VENTRICLES
The hopelessness of following the incoordinate movements over individual portions of the ventricle by inspection has led investigators to adopt myocardiographic and electrocardiographic methods in their study. But the older mechanical methods were often employed without their inadequacy being realized, and the interpretations of electrocardiograms have not always been tempered with the necessary caution.
In our investigations on ventricular fibrillation, we endeavored to employ the newer aids in graphic registration; hence it was important to be oriented to the significance of results that may be obtained by their use.
Accurate mechanical registration of the fibrillating ventricles has proved impossible in the past because efficient and responsive apparatus could not be built. Now that the difficulties in the construction of adequate mechanical recorders have largely been overcome, we are still confronted with the quandary that any mechanical appliance, no matter how efficient or whether it takes the form of surface myograph, cardiometer, or intraventricular pressure recorder, constitutes an instrument that records not the ultimate contractions of the ventricle but the resultants of many simultaneous phenomena occurring in successive intervals of time. Our understanding of the intimate nature of ventricular fibrillation has undoubtedly been impeded by our inability to fathom what is taking place in these ultimate fractions of cardiac muscle. The difficulty exists to a certain extent even when the heart is beating in a coordinate manner. Thus, one of us 2 , 3 has recently pointed out that intraventricular pressure curves can be used as graphic evidence of what the ultimate fractions are doing only because their sequence and orderliness cannot easily be changed under physiological experimental conditions. When, as apparently occurs in fibrillation, the excitation is irregular, disordered, and unpredictable from moment to moment; intraventricular pressure curves give no direct evidence as to the character of the fibrillary process. Their value in understanding the mechanisms of fibrillation is only indirect in nature. We may illustrate our meaning by an actual observation: The intraventricular pressure curves of Figure 1 show that after induction of fibrillation the intraventricular pressure curves alter to a remarkable degree. The pressures that normally varied between 5 and 100 mmIIg with each cardiac cycle suddenly fall to a mean value of about 10 or 20 mmIIg. The large, regular, characteristic pressure pulses are replaced by very small, uneven, irregular undulations that bear no direct relation to the fibrillary movements or, as in this instance, to no variations at all.
Figure 1.
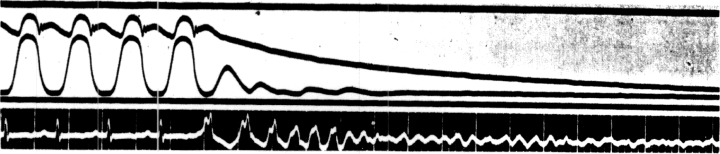
Aortic and left ventricular pressure curves and electrocardiogram (Lead II) showing the effects introducing ventricular fibrillation due to faradic excitation. 1 to 2—Initial tachysystole stage. 2 to 3—Stage of consecutive incoordination. Time 0.04 and 0.2 seconds.
While it is clear that intraventricular pressure records cannot be employed to interpret the variations in the strength and frequency of fractionate ventricular contractions, they are of service in another direction. They demonstrate that fibrillation, which follows faradic stimulation, rapidly passes into such a degree of incoordination that any semblance of regular pressure curves ceases within a second after applying a faradic current. As far as the pressure curves are concerned, the preliminary stage in which a state of tachysystole can be said to exist is extremely short.
Volume curves of the ventricles are of less value than inspection. They show an immediate increase in the size of the heart with the onset of fibrillation, an increase that progresses as fibrillation continues. Inspection gives the same evidence of dilatation but shows in addition that it is due to the predominant dilatation of the right ventricle. Indeed, as fibrillation continues, the ballooning effect due to returning blood becomes so extreme on the right side that the left ventricle may have the appearance of a small rounded tumor protruding from the right ventricle. Such an increase in size has frequently been misinterpreted as evidence of an atonic condition of ventricular muscle or of a state equivalent to paralysis. As already pointed out, pressure curves from the left as well as from the right ventricle show that for a long time a pressure range of 10–20 mmIIg persists, a finding inconsistent with the idea of atonia. Myographic records from the left ventricle give additional evidence that the mean effect of the fibrillating contraction is to hold ventricular musculature in a position approximately midway between that of systole and diastole. In the right ventricle, however, the venous blood, which is pumped into the right ventricle by the continued contractions of the right auricle, soon develops sufficient pressure to overcome the tendency to diminish the size of the right ventricle. Moreover, myographic appliances attached to the surface of the heart give no direct evidence as to the character or rate of fibrillary contractions occurring in the ventricle unless they are attached in great numbers and are applied so as to record contractions between points not more than 2 or 3 mm apart.
With the introduction of the electrocardiograph for practical use, it was generally believed that it offered a method far superior to any mechanical appliance for studying the ultimate character of the fibrillary process. Convenient as the procedure has become, it is not without drawbacks, both in its application and in the interpretation of the graphs. When the records are derived by one of the customary three leads, it should be kept in mind that the electrocardiographic oscillations correspond in neither amplitude, regularity, nor frequency to the ultimate contractions occurring in any portion of the ventricle. Like the normal electrocardiogram deflections, they represent the first differential quotient of the algebraic sum of potential differences that occur in the heart from moment to moment, oriented in the direction of the lead. There are these added difficulties in the case of fibrillating ventricles, namely, (1) that the plane of orientation is continually changing as the heart progressively dilates, and (2) that the amplitude and frequency of electrocardiographic deflections due essentially to excitation may not correspond in magnitude or number to the sums of fractionated contractions simultaneously operating in the ventricles.
Studies by Means of Moving Pictures
These difficulties in the utilization of graphic methods make it necessary to continue placing considerable emphasis upon the description of the incoordinate movements actually witnessed on the surface of the ventricles. As already emphasized, the two chief difficulties in the method of direct observation are (1) the impossibility of noting simultaneously the movements that occur in diverse portions of the ventricular surface and (2) the difficulty of following the extremely rapid movements in any given area. The use of moving pictures taken at close range and at a more rapid speed than that at which they are subsequently projected obviates both of these difficulties and in addition allows considerable magnification of the quiverings occurring in individual areas of the ventricle. Such films or their isolated portions can be repeatedly thrown upon a screen and reviewed again and again. The observer focuses his attention first upon one and then upon another area or feature of the film itself. By doing this repeatedly in many experiments the authors feel that their conception of the character of the fibrillary process has been greatly extended.
THE NATURAL COURSE OF VENTRICULAR FIBRILLATION
Fibrillation in the anesthetized dog continues from 15 to 50 minutes (average 24 minutes) before complete standstill takes place. The characteristics of the fibrillating ventricles were studied in 18 dogs by taking moving pictures and electrocardiograms during the natural course of such fibrillation.
The method of experimentation was briefly as follows: Dogs were anesthetized with small doses of morphine and sufficient quantities of sodium barbital administered intravenously. The chest was opened, the heart exposed, the pericardium severed and stitched to the lateral walls of the chest so as to form a cradle for the heart. Moving pictures were taken by means of a Bell‐Howell camera on 16 mm/film at a rate of 32 exposures per second. The camera supplied with a Zeiss 25 mm F‐2.7 lens was mounted at a distance of 12 to 18 inches from the anterior surface of the heart. Adequate illumination was furnished by three 1000 watt lamps, so that pictures could be taken with the diaphragm stop between 2.7 and 4. Orthochromatic film was used in the first experiments, but great improvement resulted from the subsequent use of panchromatic film. In many experiments, special methods were adopted for locating specific areas upon the ventricular surface. In some cases, small beads were fastened with horsehair by a delicate stitch through the epicardium. In others, white stitches with large knots were similarly placed. In still other experiments, the expedient of thrusting small white‐headed pins into the heart itself was tried, and finally the epicardium was bespeckled with small dots of white Duco paint. Finally, good use was made of the reflections of the lamp filaments from the glistening surface of the epicardium. These produced mirror phenomena that frequently brought out the peculiar characteristics of the fibrillating movements to the best advantage.
Coincident with the moving pictures, electrocardiographic records were taken by standard leads. The moments when such records were being inscribed were signaled by a marker operating at the side of the heart. In this way comparisons between the electrocardiographic deflections (usually taken by Lead II) with the actual movements shown in cinematographic records could be made at any time.*
TRANSITIONAL CHANGES IN THE CHARACTER OF FIBRILLARY CONTRACTIONS
From the onset of fibrillation until the ventricles naturally become quiescent, many variations occur in the appearance of the ventricles and in the electrocardiograms. While these changes are transitional in nature, there is sufficient abruptness in the changes to make it permissible for descriptive purposes to divide the course of fibrillation into stages:
(a) The Initial Tachysystolic Phenomena. Fibrillation of the ventricles usually begins with a very few (2 to 8) peristaltic waves that sweep rapidly over the ventricular surfaces in the general direction of the superficial muscle bundles. Frequently they seem to arise from a single focus located either in the central region or pulmonary conus of the right ventricle and in this case are fairly regular in period and direction. This has been ascertained by counting the number of photographic exposures between the completion of the separate waves. In other cases, however, the waves appear to spread in several directions, beginning sometimes on one area and at other times on several surface regions. The larger contraction waves sweep over both ventricles; e.g., a contraction may originate in the central region of the right ventricle, move in both directions, reaching the base of the right ventricle and the apex of the heart approximately simultaneously; or the wave may start at the pulmonary conus of the right ventricle, sweep over the anterior surface of the right ventricle, apparently hurdle the coronary vessels and pass on uninterruptedly to the left ventricular apex. These first movements are accompanied by characteristic deflections in the electrocardiograms that are illustrated in Figures 1, 2B, and 3A, between the points labeled 1 and 2. They are of large size, rise and fall rapidly, and occupy an interval of 0.08 seconds or more. Beat by beat their amplitude decreases, their gradients become more gradual, and the periods of the entire beat decrease, i.e., the calculated ventricular rate becomes progressively faster. When the periods are reduced to 0.1 or 0.08 seconds, i.e., corresponding if continued to a ventricular rate of 600 to 750 per minute, the process abruptly passes into the next phase.
Figure 2.
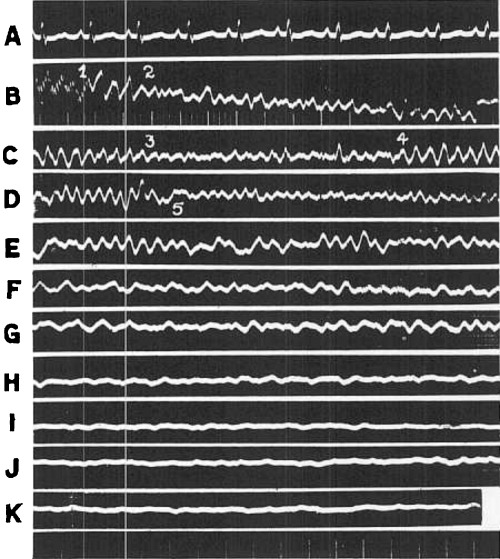
Segments of electrocardiogram (Lead II) taken at various stages during the course of fibrillation (043), A, control; B, and C, Immediate effects; D, after thirty seconds; E, after 1 minute; F, after minute; G, after 2 minute; H, minute; I, after 16 minute; J, after 20 minute; K, after 21 minute. Time 0.04 and 0.2 seconds.
Figure 3.
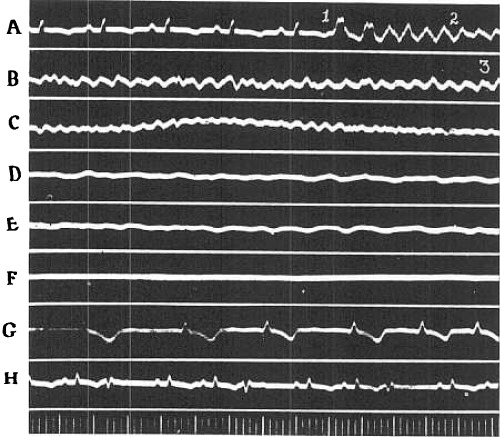
Segments of electrocardiogram (Lead II). showing the early stages of natural fibrillation, the effects of KCl and of revival following use of CaCl2 (O‐42). A, control and initial tachysystolic stage; B, coarse incoordination, minutes later; C, tremulous incoordination after 2 minutes; D, 2 minutes; after use of KCl; E, 12 minutes after; F, minutes; after; G, initial idioventricular beats, recorded 19 minutes after fibrillation first started; H, 30 minutes after record; H. Time 0.04 and 2 seconds.
As shown in Figure 1. each electrical deflection is accompanied by a small but distinct elevation of intraventricular pressure, though the aortic pressure declines steadily because no blood is expelled. All the evidence, therefore, favors the view that these beats are still coordinated in the sense that they sweep over the ventricles in regular paths and develop sufficient force to elevate intraventricular pressure.
In the dog's heart this taehysystolic stage usually lasts less than 1 second; in fact, it usually sets in before faradie stimulation has ceased. This is not surprising in view of the high contraction frequencies reached.
(b) Stage of Conculsive Incoordination. The initial stage already described leads into a second in which waves of contraction with distinctly different rhythms and sequence pass over the ventricles. This stage ordinarily continues for 15 to 40 seconds, but in several instances it was shorter.
When any small circular area 2 to 3 cm in diameter is carefully observed in projected moving pictures, it can easily be seen that the frequency has increased and that the sequence of contractions is less regular. The localized contractions appear to be rather forceful, but successive contraction waves do not spread over identical paths or involve the same surface groups of muscle tissue. By comparing several such areas, it is noted that the frequencies differ and bear no relation one to the other. Owing to the extreme incoordination, no pressure oscillations occur within the ventricles.
This stage of incoordination is characterized by large, rather violent oscillations of the galvanometer string. They are illustrated in Figures 1 and 3B, between points 2 and 3. In these instances the successive waves vary considerably in contour, amplitude, and period, but the frequency, calculated over a considerable portion of the curve, may be placed between 600 and 660 per minute. Occasionally, the electrocardiographic oscillations become even more rapid while this form of incoordination is in progress. This is shown in Figure 2B between points 3 and 4 where they attain the alarming frequency of 1560 per minute. Careful comparisons of electrocardiograms and cinematographic records during such intervals have clearly demonstrated, however, that these rapid electrocardiographic deflections do not portray the contraction phenomena visible on the surface of the heart, for the latter are always much slower during this stage of fibrillation. In the instance shown in Figure 2 the stage terminated at point 5.
When the ventricles are held in the palm of the hand, a fluttering, undulatory, convulsive sensation is experienced, no doubt due to the fact that blocks of myocardium are contracting with different strengths and at different rates and rhythms. From the convulsive appearance of the heart and the fact that no periodic changes of intraventricular pressure are produced, we have designated it as the stage of convulsive incoordination. The terms coarse fibrillation, flutter, undulation, ondulation, Wühlen und Wogen, etc., used by others might be applied but are avoided since we are not certain that they were always limited to the particular stage we desire to describe.
(c) Stage of Tremulous Incoordination. This type of incoordination ordinarily follows upon the coarse, convulsive type and continues for 2 or 3 minutes. The entire surface of the ventricles presents the appearance of multitudes of shivering and shimmering motions of a very rapid sort. The contraction waves spread very rapidly and apparently over only short distances. We have seen many instances where reflected light beams from the epicardium give the impression of darting wriggling worms or of contraction waves chasing each other in circles. When the heart is palpated, it gives the impression that has been so aptly described as similar to that obtained when the hand is thrust into a can of worms. This stage is undoubtedly comparable to the types of fibrillation described as delirium cordis, mouvement fibrillaire, Herz‐zittern, fibrillary twitchings, etc., by others, but for reasons already given we have purposely avoided previously used terms.
Viewed as slow‐motion picture films, the process is seen to consist of localized contraction waves spreading over short distances or circling around in very limited areas. Their speed of propagation is not actually increased. The frequencies of these visible contractions in different regions of the ventricular surface are apparently unrelated. We have studied one instance carefully in which the rate varied between 600 and 1800 per minute in different portions of the right ventricle. We have tried numerous and laborious expedients for interrelating these rapid contraction circles in selected regions, but after diligent attempts we have found this impossible.
During this stage the electrocardiographic deflections increase in frequency and diminish in amplitude. They are shown in Figure 2D after point 5 where their frequency is about 1140 per minute and in Figure 3C where they average about 1520 per minute. Counts in other experiments range from 1100 to 1700 per minute. Pressure curves recorded from the ventricular cavities show that these rapidly executed incoordinate contractions of extremely small muscle units not only maintain the previous existing intraventricular tension but sometimes actually increase it somewhat. The pressure curves, however, continue as a straight line and show no evidence of oscillations except occasionally those due to continuing auricular contractions.
(d) Stage of Progressive Atonic Incoordination. The stage of rapid incoordinate contractions just described is gradually replaced by a stage of coarse atonic incoordination, so designated because the visible waves not only become coarser and slower, but they no longer maintain the previous small even elevation of intraventricular pressure. We are not aware of any previous description of this stage; most observers limit themselves to the phenomena characterizing the earlier stages.
The ventricular surfaces show characteristic changes in appearance. During the early portion of this coarse atonic stage of fibrillation, wavelets of contraction are seen to spread over small areas of the ventricle. The rhythm is obviously different in different regions. The rate of propagation slows down materially, the frequency of movement is reduced, and the contractions are obviously less vigorous. At first such movements occur over the entire ventricular surface, but gradually they decrease in intensity until certain areas display none. The quiescent areas increase in number until finally only the slightest movements remain in a few regions. The areas that usually pulsate longest in this manner are those that lie near the coronary vessels, usually to the right side of the septal demarcation. The last movements to be seen are mere quivers, perceptible only when beams of light from a lamp are reflected from the moist and glistening epicardium. Frequently, when all visible movement has apparently ceased and no sensation can be felt by the finger or heard on auscultation, the electrocardiograph still continues to show minute irregular oscillations. Whether these are accompained by contractions in the deeper layers of the ventricle or whether conduction occurs without mechanical shortening under such circumstances, cannot be decided with finality.
The nature of the electrocardiographic oscillations up to the final cessation of all movement is shown in Figure 2, segments E to K. At first, the oscillations are fairly large; indeed they resemble those occurring during the early stage of convulsive incoordination. Their frequency ranges from 540 to 720 in Figure 2. The amplitude of the deflections gradually decreases until toward the end only small oscillations remain. The frequency then also diminishes, e.g., in Figure 2K the frequency is about 360. These changes in rate correspond reasonably well with movements visible in cinematographic records, but obviously the amplitudes of the deflections in the early part of this phase bear no relation to the intensity of fibrillary contractions.
(e) Auricular Contractions During the Course of Ventricular Fibrillation. The auricles maintain a regular rhythm for a variable length of time. Sometimes their contractions terminate before ventricular fibrillation ceases and at other times they outlast the fibrillation. The changes in the auricles are those common to other forms of asphyxia. The auricular rate soon decreases, the contractions first increase but soon decrease in vigor. It should be borne in mind that the electrical records of ventricular activities may be periodically modified by electrical variations due to auricular contraction. Very early the rate of propagation is reduced. Peristaltic waves sweep over the auricle at slow rates, giving a beautiful visible demonstration of the successive contractions of adjoining fractions. Frequently heart block develops between the right and left auricles. On several occasions independent rhythms have been observed, suggesting that a rhythmic center becomes active in the left auricle.
THE CHANGES DURING POTASSIUM INHIBITION
The earlier manifestations of fibrillation transpire so quickly that potassium salts cannot be administered easily until the stage of tremulous incoordination has supervened. Cinematographic and electrocardiographic records from eight experiments were carefully studied. They consistently showed that potassium salts do not modify the stages of the fibrillary process: they merely hasten the process. The 15 to 50 minutes naturally required for fibrillation to run its course are often shortened to 1 or 2 minutes. Projected moving pictures show that the fine fibrillary waves in any region recur less frequently and spread more slowly without involving larger areas. On the contrary, the active regions gradually diminish, leaving motionless areas over the entire surface of the heart. The vigor of the localized contractions also diminishes visibly, until large areas are entirely quiescent. Feeble, slow contractions finally remain only in very restricted areas, the anterior region of the right ventricle near the interventricular sulcus being the usual area in which feeble contractions persist longest.
The electrocardiographic curves reveal no new features, but the entire process normally present runs its course more rapidly. The portion of the atonic stage that is characterized by large deflections is particularly affected. It is either very short or does not appear at all. Figure 3 is an example of the characteristic electrocardiographic changes. Segments A, B and C illustrate the course of events during the first 2 minutes of natural fibrillation. The record shown as segment D was taken 1 minute later, i.e., 15 seconds after injection of KCl. The records reproduced as segments E and F were taken at additional intervals of 20 and 25 seconds, respectively. Almost immediately the amplitude of oscillations greatly diminished and their frequency was reduced to 240 per second in segment E. Fibrillation was completely abolished (segment F) 1 minute after completion of the injections.
It should be added that visible auricular contractions that normally persist for many minutes also promptly stopped. The auricles quickly came to a complete standstill. This demonstrates that potassium diffuses quickly through fluids in the cavities of the heart.
THE COURSE OF REVIVAL AFTER USE OF CALCIUM AND MASSAGE
After intraventricular injection of calcium salts and after the ventricles have been massaged for several minutes in such a way that each compression causes a slight elevation of aortic pressure, it is noticed that the ventricles respond to mechanical stimuli or compression. Similar responses may be obtained by gentle taps or single electric shocks. The ventricles recover their functions of conduction and contraction prior to regaining their ability to generate spontaneous impulses. The auricles are not yet contracting automatically; the nodal structures likewise have not regained their automaticity. Within a comparatively short time—a matter of 10 seconds in some experiments and 1 to 2 minutes in others—spontaneous ventricular beats begin. Projected moving pictures show clearly the broad waves of contraction spreading over the ventricles. Beginning over the central region of the right ventricle they usually sweep upward toward the base of the right ventricle, although downward moving waves of contraction were witnessed in two instances. In the case of the left ventricle, they start at the apex, move upward along the coronary vessels, then to the left and toward the base. One instance was found in which the contraction waves had the ratio of 3:52, but, as a rule, they have the same tempo. The waves appearing over the surface of the two ventricles are not quite synchronous, however; sometimes the right and sometimes the left ventricular waves are in the lead.
As these slightly asynchronous waves gradually become stronger, the ventricles undergo a curious deformation, each beat being not unlike that seen when rapid ventricular tachycardia is produced by stimulating one ventricle artificially. Since each of these contractions elevates mean arterial pressure, it must be accompanied by elevations of intraventricular pressure. Unfortunately, we have not succeeded in recording reliable intraventricular pressure curves of this event.
At the same time that the idioventricular rhythm develops or slightly thereafter, the auricular beats return. Frequently the left auricle beats alone; again both auricles beat at different rhythms, and finally a coordinated rhythm is established. Impulses do not yet reach the ventricles, which continue to beat with an idioventricular rhythm. This is also demonstrated by electrocardiographic tracings. Segment G of Figure 3 shows a series of such idioventricular beats of increasing frequency that were recorded after an initial period of less regular action. They indicate that a single ventricular focus dominates the rhythm. This is not always the case, however, as is shown in the tracings of Figures 4 and 5. Comparison of segments A, B, and C of Figure 4 shows clearly that the pacemaker shifted. This is partly due to the fact that a more rhythmic center functionates but not necessarily so. Thus the rate was 58 per minute in segment C and 120 in segment B. Dynamically, beats originating in different centers were equally effective, for the blood pressure altered with the heart rate. Frequently, also, successive stimuli arise from different foci; a struggle for supremacy in the idioventricular rhythm apparently takes place. This is illustrated in segments A, B, and C of Figure 5, which shows records obtained on recovery from a second fibrillation that were induced approximately 1 hour later in the same animal.
Figure 4.
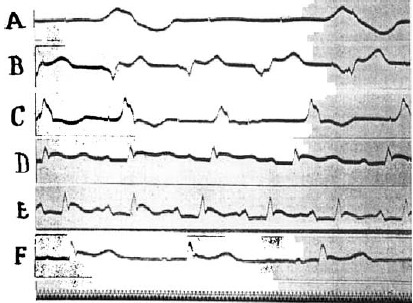
Electrocardiograms (Lead II) showing mechanisms in recovery from fibrillation. Description in text. Time 0.02 seconds (O‐70. VI–XI).
Figure 5.
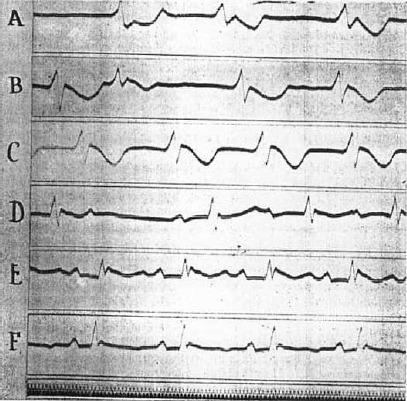
Electrocardiograms (Lead II), showing mechanisms of recovery from a second fibrillation in the same dog. Time 0.02 seconds. Description in text (O‐70. XV–XX).
The development of this idioventricular rhythm is of greatest importance in tiding the animal over the last few critical minutes still remaining before irreparable damage is done to the central nervous system. It often precedes the redevelopment of a sinus rhythm for the auricles and, at all events, establishes effective beats capable of maintaining a reasonable blood pressure level during the interval that complete A‐V block persists.
The fact that revival is inaugurated by an idioventricular rhythm is additional positive evidence that our recoveries were not spontaneous or due to chance. Spontaneous recovery, such as occurs frequently in the cat's heart and occasionally as a special phenomenon in sensitized dogs' hearts, is different in nature. Fibrillation ceases promptly, a long postundulatory pause follows, and then a regular supraventricular beat reappears immediately.
After revival by calcium, associated with massage, such a rhythm only develops subsequent to an idioventricular rhythm of several minutes' duration. The change in the pacemaker is quite abrupt. Previous to its development an auricular beat dominated by the sinus node makes its appearance, auricles and ventricles beating with independent rhythms. Quite suddenly the total block is lifted, and the ventricles begin to beat in sequence with the auricles. The electrocardiographic changes typical of this condition are shown in the lower records of Figures 4 and 5. The normal P, R, and T sequence obtains. The records taken immediately after development of a supraventricular rhythm differ from the normal in several ways. Usually, as in the record of Figure 5D, the P‐R interval is prolonged, but this is always reduced to normal values within 5 or 6 minutes or less (cf. Figures 4F and 5F). The deflection is often abnormal in contour, reminding one of the variable changes that follow coronary ligation. Frequently the descending level of R does not return to the base line but grades into it by a broad declining plateu or is followed by a rounded hump. These features are often more pronounced than in the case of segments reproduced as Figures 4E and 5E. After these abnormalities disappear, the T wave may become accentuated as in Figure 4F, but frequently it is negative, as shown in Figures 3H and 5F. The normal rhythm once regained becomes permanent. A relapse to an idioventricular type was not observed. Not infrequently it is interrupted by premature systoles of auricular, nodal, or ventricular origin, as in Figure 3H. Gradually the normal characteristics of the electrocardiogram reappear. The P‐R interval is reduced to normal values, and the ventricular waves regain their natural appearance. That the heart has not been injured permanently, either by the concentrated salt solutions employed or by the period of temporary anemia, is thus conclusively established.
CONCLUSIONS
1. The natural course of events from the onset of fibrillation following faradic excitation to complete diastolic rest was studied by recording electrocardiograms and moving pictures simultaneously. In addition, intraventricular pressure curves were optically recorded during a number of experiments. The changes taking place when fibrillation was inhibited by intraventricular injections of 5% KCl and the process of recovery following similar injections of 5% CaCl2 solutions combined with massage, were investigated by the same means.
2. Fibrillation induced by faradic stimulation continues naturally for 15 to 50 minutes and may be divided into four stages, on the basis of surface changes, electrocardiographic deflections and intraventricular pressure variations.
3. The initial stage of tachysystole lasts less than 1 second and is characterized by the spread of rapidly recurring but coordinated contraction waves, by large electrocardiographic deflections with steep gradients, and by definite if small intraventricular pressure variations.
4. The second stage of convulsive incoordination ordinarily lasts 15 to 40 seconds and is characterized by rapid irregular localized contractions that spread to short and variable distances over the heart. They are accompanied by large electrical deflections, 600 or more per minute, which vary considerably in size, amplitude, and contour.
5. The third stage of tremulous incoordination ordinarily continues for 2 or 3 minutes and is characterized by multitudes of irregular yet forceful shivering or trembling motions, each spreading to very short distances and with highly variable frequencies over different surface regions. They give rise to small irregular electrocardiographic oscillations having frequencies between 1100 and 1700 per minute, and are capable of increasing the intraventricular pressure level slightly.
6. The fourth stage of atonic incoordination is characterized by feeble wavelets of contraction spreading irregularly and at slow rates over small areas until more and more areas become quiescent, and finally the very slightest movements remain in a few areas only. The electrical deflections perhaps become slightly more regular in contour and spacing, but their amplitude becomes progressively smaller, and their frequency is gradually reduced to 400 per minute or less.
7. Potassium chloride injected into both ventricular cavities does not modify the stages through which fibrillation naturally passes; it merely hastens the process so that fibrillation stops within an average period of 2.4 minutes.
8. Intraventricular injections of CaCl2 after potassium inhibition combined with massage, first inaugurate a coordinated idioventricular rhythm, characterized by slow waves of contraction sweeping over the two ventricles asynchronously but in coordinated fashion. After a short interval, a supraventricular rhythm is reestablished, the electrocardiogram regaining all its normal characteristics.
Published by the Physiology Committee on Electric Shock.
Footnotes
*Cinematographic films demonstrating the changes to be described have been exhibited before the American Physiological Society (1928). Cleveland Academy of Medicine (1928). Pacific Northwest Medical Assn. (1928). Buffalo University Medical School Alumni Association (1929), and International Congress of Physiology (1929).
REFERENCES
- 1. Wiggers, CJ . Am J Physiol February, 1930. [Google Scholar]
- 2. Wiggers, CJ . Am J Physiol 80: 1, 1927. [Google Scholar]
- 3. Wiggers CJ. Pressure Pulses in the Cardiovascular System. London/New York : Longmans, Green and Co., 1928,p. 146. [Google Scholar]