

Abstract
Under physiological and pathological conditions, elastin is degraded to produce elastin-derived peptides (EDPs). EDPs are detected in the healthy human brain, and its concentration significantly increases after ischemic stroke. Both elastin and EDPs contains replications of the soluble VGVAPG hexapeptide, which has a broad range of biological activities. Effects of VGVAPG action are mainly mediated by elastin-binding protein (EBP), which is alternatively spliced, enzymatically inactive form of the GLB1 gene. This study was conducted to elucidate the activation and role of the N-methyl-D-aspartate receptor (NMDAR) in elastin-derived VGVAPG peptide-dependent calcium homeostasis in mouse cortical astrocytes in vitro. Cells were exposed to 10 nM VGVAPG peptide and co-treated with MK-801, nifedipine, verapamil, or Src kinase inhibitor I. After cell stimulation, we measured Ca2+ level, ROS production, and mRNA expression. Moreover, the Glb1 and NMDAR subunits (GluN1, GluN2A, and GluN2B) siRNA gene knockdown were applied. We found the VGVAPG peptide causes Ca2+ influx through the NMDA receptor in mouse astrocytes in vitro. Silencing of the Glb1, GluN1, GluN2A, and GluN2B gene prevented VGVAPG peptide-induced increase in Ca2+. Nifedipine does not completely reduce VGVAPG peptide-activated ROS production, whereas MK-801, verapamil, and Src inhibitor reduce VGVAPG peptide-activated Ca2+ influx and ROS production. These data suggest the role of Src kinase signal transduction from EBP to NMDAR. Moreover, the VGVAPG peptide affects the expression of NMDA receptor subunits.
Subject terms: Biochemistry, Molecular biology, Medical research, Molecular medicine, Pathogenesis
Introduction
Elastin is an insoluble, amorphous, hydrophobic protein with widespread presence in the walls of blood vessels as well as in the microvasculature of the normal human brain1. Under physiological and pathological conditions, elastin is degraded to generate elastin-derived peptides (EDPs)2,3. EDPs are detected in the healthy human brain, and its levels significantly increase after an ischemic stroke4,5. Elastin and EDPs contains replications of the soluble VGVAPG hexapeptide, which has a broad range of biological activities, such as increasing cell proliferation and migration, affecting chemotaxis, or causing protease release6–8. Effects of VGVAPG action are mainly mediated by elastin-binding protein (EBP), which is a alternatively spliced, enzymatically inactive form of the GLB1 gene (β-galactosidase), known as S-Gal9.
In the central nervous system (CNS), astrocytes are responsible for the synthesis of extracellular matrix (ECM), supply of nutrients and oxygen to neurons, removal of dead neurons, and playing a role in neurotransmission and synaptic connections10,11. To date, it is known that astrocytes do not have action potentials. However, they display spontaneous and pharmacologically evoked increases in intracellular Ca2+ concentration ([Ca2+]i), which is a sign of its excitability12. In astrocytes, an increase in the Ca2+ level can result in mitochondrial dysfunction, increased production of free radicals, and activation of degenerative process controlled by Ca2+-activated proteases and phospholipases13. Therefore, it has been suggested that intracellular Ca2+ influx after trauma, ischemia, or stroke leads to cell damage13,14.
The N-methyl-D-aspartate (NMDA) receptor (NMDAR) is the most important excitatory receptor, permeable to Ca2+, Na+, and K+. However, its permeability to ions is strongly dependent on the composition of the subunit15. NMDAR are heterotetramers composed of two GluN1 subunits and two GluN2 A-D or GluN3 A-B subunits16. The di-heteromeric GluN1/GluN2B and GluN1/GluN2A receptors are an important fraction of juvenile and adult NMDARs. Moreover, in the adult CNS, particularly in structures such as the hippocampus and cortex, GluN2A and GluN2B are the predominant subunits17,18. It is well known that the NMDAR is involved in excitotoxicity-induced cell death in human embryonic stem cell-derived neurons, neuroblasts, and neuroblastoma cells19–21. NMDAR-mediated excitotoxicity is Ca2+-dependent and is typical of the nervous system22.
Studies to date have described that κ-elastin increases Ca2+ influx in human monocytes and fibroblasts, as well as in smooth muscle cells from pig aorta23. Similar, tropoelastin, EDPs, and the VGVAPG peptide increase the Ca2+ level in human umbilical venous endothelial cells (HUVEC)24,25. Moreover, in different glioma cell lines (C6, CB74, CB109, and CB191) κ-elastin or the (VGVAPG)3 peptide provoked a pronounced and dose-dependent increase in [Ca2+]i26.
It is well known that different Ca2+ signaling pathways interact with other cellular signaling systems such as reactive oxygen species (ROS)27. However, data on the ROS production in relation to EDPs are limited. To date, EDPs decrease ROS production in human neutrophils28. On the other hand, it has been described that EDPs induce ROS production in murine monocytes and human fibroblasts29–31. However, such data suggest that the effects of EDPs are cell and/or tissue dependent. In a previous study, we found that the VGVAPG peptide increases ROS production in an EBP-dependent manner both in mouse astrocytes in vitro32 and a human neuroblastoma (SH-SY5Y) cell line33. It is well known that calcium influx the NMDAR stimulates ROS production34. Moreover, signaling pathways transduced through the elastin receptor can use G protein and, thus, calcium channels are opened35. EBP-mediated cell proliferation has been described to involve the activation of Gi proteins with the opening of calcium L-type channels and resultant increase in Ca2+ levels in porcine smooth muscle cells36.
The present study was undertaken to elucidate the activation and role of the NMDAR in elastin-derived VGVAPG peptide-dependent calcium homeostasis in mouse cortical astrocytes in vitro.
Results
Ca2+ determination
After 15-min exposure of mouse astrocytes to 10 nM VGVAPG peptide, the level of cellular Ca2+ increased by 24.92% compared to control. VVGPGA does not cause influx of Ca2+ in to cells. MK-801, nifedipine, verapamil, and Src kinase inhibitor I prevented Ca2+ influx induced by the VGVAPG peptide (Fig. 1A).
Figure 1.
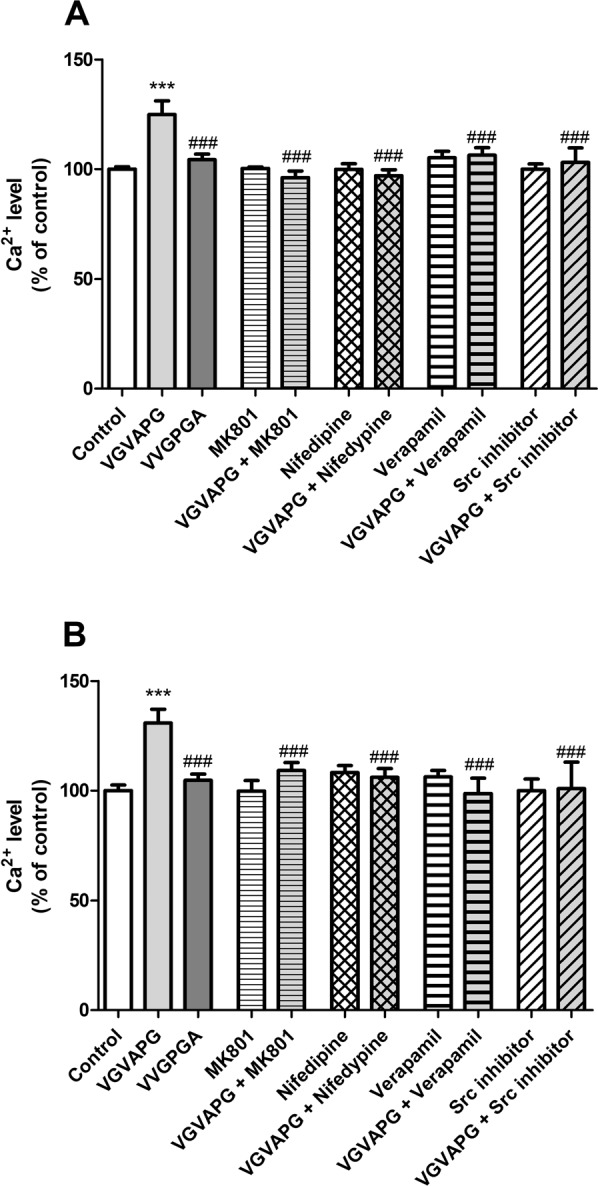
The effect of 10 nM VGVAPG peptide, 10 nM VVGPGA peptide, and co-treatment with 1 µM MK-801, 10 µM nifedipine, 10 µM verapamil, or 1 µM Src kinase inhibitor I on Ca2+ level in primary mouse astrocytes; (A) after 15 min and (B) after 30 min of exposure. Each point represents the mean ± SD of three independent experiments, each of which comprised six replicates per treatment group. *P < 0.05, **P < 0.01, and ***P < 0.001, versus the control cells. ###P < 0.001, versus the VGVAPG peptide-treated group.
After 30-min exposure of cells to VGVAPG peptide, the Ca2+ influx remained at a similar level as after 15-min exposure. The VGVAPG peptide increased of cellular Ca2+ level by 30.33% compared to control. VVGPGA does not cause influx of Ca2+ in to cells. MK-801, nifedipine, verapamil, and Src kinase inhibitor I prevented Ca2+ influx induced by VGVAPG peptide (Fig. 1).
ROS measurement
After 3-h exposure of mouse astrocytes to 10 nM VGVAPG peptide, we observed an increase in ROS production by 48.70% compared to the control. VVGPGA does not stimulate ROS production in astrocytes. MK-801 alone reduced ROS production by 16.36%, compared to control. Interestingly, nifedipine alone increased ROS production by 27.53%, compared to control. MK-801, verapamil, and Src kinase inhibitor I reduced ROS production stimulated by VGVAPG peptide to vehicle control levels. Nifedipine slightly but significantly reduced ROS production stimulated by the VGVAPG peptide by 19.65% as compared to the VGVAPG peptide-stimulated group (Fig. 2A).
Figure 2.
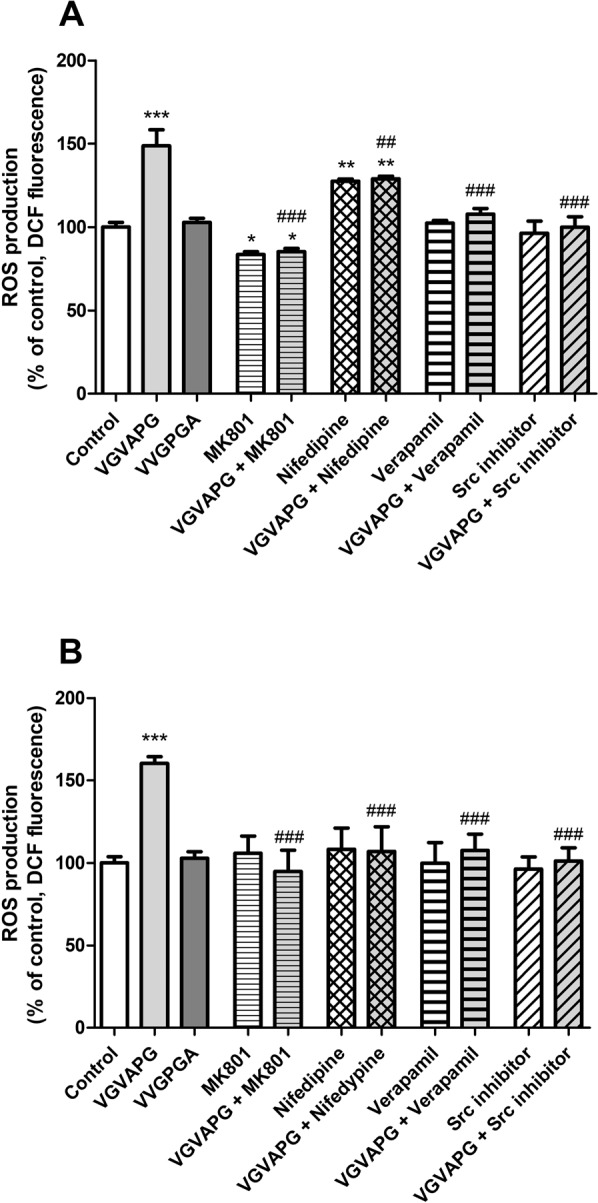
Effect of 10 nM VGVAPG peptide, 10 nM VVGPGA peptide, and co-treatment with 1 µM MK-801, 10 µM nifedipine, 10 µM verapamil, or 1 µM Src kinase inhibitor I on ROS production in primary mouse astrocytes; (A) after 3-h exposure and (B) after 6-h exposure. Each point represents the mean ± SD of three independent experiments, each of which comprised six replicates per treatment group. *P < 0.05, **P < 0.01, and ***P < 0.001, versus the control cells. ##P < 0.01 and ###P < 0.001, versus the VGVAPG peptide-treated group.
After 6 h of astrocyte exposure to VGVAPG peptide, we observed an increase in ROS production by 60.28% compared to control. VVGPGA does not stimulate ROS production in astrocytes. MK-801, verapamil, nifedipine, and Src kinase inhibitor I reduce ROS production stimulated by the VGVAPG peptide (Fig. 2B).
Ca2+ level and ROS production in astrocytes after gene silencing procedure
Ca2+ level
In cells transfected with scrambled siRNA, 15-min exposure to 10 nM VGVAPG peptide increased Ca2+ influx by 30.16% compared to control. The Glb1 gene-silencing procedure protects cells by an increase in Ca2+ influx. Likewise, silencing of NMDAR subunits (GluN1, GluN2A, and GluN2B) protects cells by an increase in Ca2+ influx (Fig. 3A).
Figure 3.
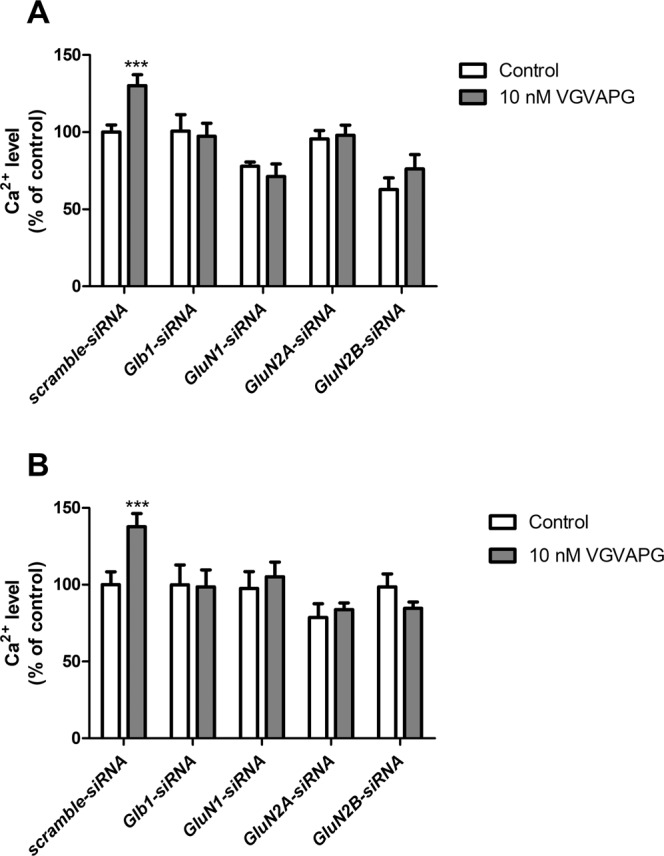
Effect of GluN1, GluN2A, and GluN2B siRNA on Ca2+ level after stimulation of primary mouse astrocytes with 10 nM VGVAPG peptide in primary mouse astrocytes; (A) after 15 min and (B) after 30 min of exposure. Each point represents the mean ± SD of three independent experiments, each of which comprised six replicates per treatment group. **P < 0.01 versus the control cells.
After 30-min exposure to VGVAPG peptide, results similar to those previously described were obtained. The VGVAPG peptide increased Ca2+ influx (increase by 37.69%) compared to control. Silencing of NMDAR subunits (GluN1, GluN2A, and GluN2B) protects cells by increasing Ca2+ influx (Fig. 3B).
ROS production
In cells transfected with scrambled siRNA, 3-h exposure to 10 nM VGVAPG peptide increased ROS production by 34.65% compared to the control. The GluN1, GluN2A, and GluN2B gene-silencing procedure protects cells by increasing ROS production (Fig. 4A).
Figure 4.
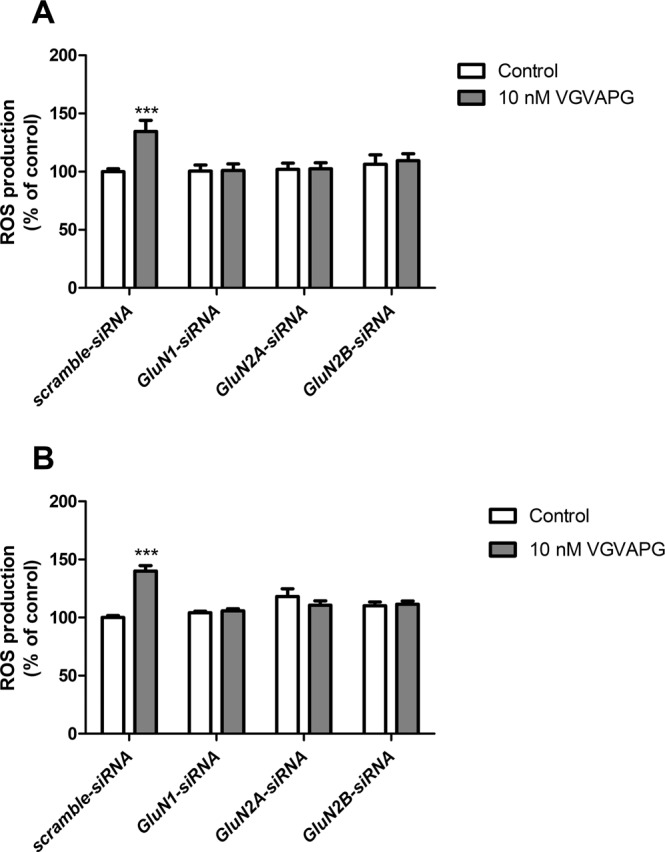
Effect of GluN1, GluN2A, and GluN2B siRNA on ROS production after stimulation of primary mouse astrocytes with 10 nM VGVAPG peptide; (A) after 3-h exposure and (B) after 6-h exposure. Each point represents the mean ± SD of three independent experiments, each of which comprised six replicates per treatment group. *P < 0.05, **P < 0.01, and ***P < 0.001, versus the control cells.
After 6-h exposure to VGVAPG peptide, we obtained results similar to those that were previously described. VGVAPG peptide increases ROS production by 40.16% compared to the control. The GluN1, GluN2A, and GluN2B gene-silencing procedure protects cells by increased ROS production in astrocytes (Fig. 4B).
mRNA expression of GluN1, GluN2A, and GluN2B NMDAR subunits
In cells transfected with scrambled siRNA, 6-h exposure to 10 nM VGVAPG increased mRNA expression of the GluN2A NMDAR subunit by 21.66%. Silencing of the Glb1 gene protects cells through the VGVAPG peptide, and we did not observe changes in mRNA expression of the GluN1 and GluN2A genes. However, silencing of the Glb1 gene causes 10 nM VGVAPG to induce an increase in mRNA expression of the GluN2B gene (increase by 24.16% compared to control; Fig. 5).
Figure 5.
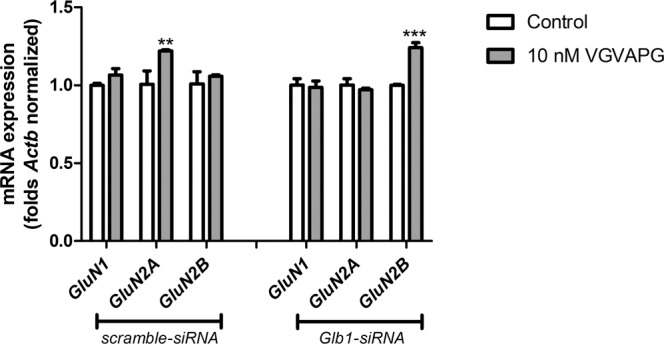
Effect of Glb1 siRNA transfection on GluN1, GluN2A, and GluN2B mRNA expression after stimulation of primary mouse astrocytes with 10 nM VGVAPG peptide in vitro; after 6 h of exposure. Each point represents the mean ± SD of three independent experiments, each of which comprised six replicates per treatment group. **P < 0.01 and ***P < 0.001, versus the control cells.
Discussion
Our experiments are the first to show that the VGVAPG peptide increases Ca2+ influx in to mouse primary astrocytes in vitro. The experiments showed that MK-801 (a noncompetitive antagonist of the NMDAR) and Src kinase inhibitor I completely prevent Ca2+ influx after VGVAPG stimulation. Calcium-channel antagonists are a heterogeneous group of compounds. Nifedipine acts primarily on the smooth muscle cells of blood vessels, and has little effect on the heart. On the other hand, verapamil is cardioselective, and mainly affects the heart by blocking voltage-gated calcium channels37. Therefore, we chose nifedipine and verapamil for further research. Moreover, our study shows that nifedipine and verapamil prevent Ca2+ influx stimulated by the VGVAPG peptide in mouse astrocytes in vitro, although not as strongly as MK-801 or Src kinase inhibitor. Moreover, after silencing of both the Glb1 gene and NMDAR subunits (GluN1, GluN2A, and GluN2B genes), the VGVAPG peptide does not activate Ca2+ influx. The data we obtained suggest that the activation of EBP results in opening of the NMDAR and other types of calcium channels are less often blocked by nifedipine and verapamil. To date, it is well documented that EDPs and/or the VGVAPG peptide initiate Ca2+ influx in different cell types such as HUVEC, SMC, fibroblasts, and gliomas23–26,38. Moreover, numerous reports have shown that the EDP-activated influx of Ca2+ into cells requires EBP activation23–25,38. Some authors have suggested that G proteins and c-Src as well as ERK1/2 or MEK1/2 kinases are involved in signal transduction, although the molecular mechanism mediating the opening of Ca2+ channels remains unclear36,38,39.
The next step of our study was to determine ROS production as a consequence of the influx of Ca2+ ions into cells. We found that stimulation of primary astrocytes by the VGVAPG peptide increased ROS production in both of the time intervals studied. MK-801, verapamil, and Src inhibitor prevent ROS production that was stimulated by the VGVAPG peptide. Similarly, after silencing of NMDAR subunits (GluN1, GluN2A, and GluN2B genes), the VGVAPG peptide did not cause ROS production. Interestingly, nifedipine does not completely prevent the ROS production stimulated by the VGVAPG peptide. Our results suggest that the binding of the VGVAPG peptide to EBP leads to increased ROS production, which mainly could be NMADR dependent. To date, reports concerning ROS production after EDP and/or VGVAPG stimulation are inconsistent. Our previous studies have shown that the VGVAPG peptide increases ROS production in an EBP-dependent manner in mouse primary astrocytes in vitro32. Similarly, an increase in ROS production after EDP stimulation has been reported in monocytes, human fibroblasts, and a neuroblastoma (SH-SY5Y) cell line29,30,33. Moreover, EDPs have been described to enhance the activities of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), or glutathione peroxidase (GPx), and increase lipid peroxidation in human fibroblasts40. Furthermore, the VGVAPG peptide increases the expression and activity of GPx in SH-SY5Y cells33. On the other hand, the VGVAPG peptide reduced ROS production in neutrophils in controls and patients with stable chronic obstructive pulmonary disease28.
Phosphorylation regulates many cellular processes, including protein activity, localization, mobility, and protein–protein interactions. Direct phosphorylation of ionotropic glutamate receptors is a key mechanism that regulates channel function and receptor localization at synapses41,42. The proto-oncogene tyrosine-protein kinase Src is an important modulator of NMDARs. It has been described that the NR2A, NR2B, and NR2D subunits can be tyrosine phosphorylated41. Moreover, phosphorylation of the abovementioned subunits results in increased NMDAR activity43. Our data are the first to show that, in astrocytes in vitro, the Src kinase inhibitor I prevents Ca2+ influx and ROS production stimulated by the VGVAPG peptide. These data suggest a role of Src kinase in signal transduction from EBP to NMDAR and, probably, in other calcium channels.
It is well known that, in cells of the nervous system, the NMDAR is one of the most important ion channels19–21. NMDAR are responsible for Ca2+ influx and cellular ROS production that can result in excitotoxic cell death. However, our previous studies have shown that VGVAPG is nontoxic, and does not activate the apoptotic process in mouse astrocytes in vitro44. Therefore, we decided to determine the involvement of NMDAR subunits in the mechanism of action of VGVAPG in astrocytes in vitro. Our experiment with silencing of the GluN1, GluN2A, and GluN2B genes confirmed the experiments with the NMDAR antagonist MK-801. Moreover, we determined that the VGVAPG peptide affects the mRNA expression of GluN2A NMDAR subunits. In cells transfected with scramble siRNA, 10 nM VGVAPG peptide increased the mRNA expression of the GluN2A NMDAR subunit. On the other hand, after the silencing of the Glb1 gene, we did not observe changes in the mRNA expression of GluN2A genes. However, silencing of the Glb1 gene ensured the 10 nM VGVAPG could increase the mRNA expression of the GluN2B gene. The proportion of the GluN2B and GluN2A subunits changes with age and indicates the maturation of the nervous system20,45. In fetal life, strong expression of the GluN2B subunit is observed whereas, after birth, this expression decreases in favor of the GluN2A subunit46,47. However, a heterotetramer containing GluN2B subunits is mainly responsible for excitotoxicity and the intensity of neuronal apoptosis, whereas the opposite effect is caused by the presence of GluN2A subunits promoting cell survival48,49. Our data suggest that the VGVAPG peptide changes the NMDAR expression profile to one similar to the adult profile. Increased expression of GluN2A can promote cell survival and supports our hypothesis postulated in our other papers that the VGVAPG peptide promotes regeneration and the healing process in astrocytes32,44,50. However, after Glb1 silencing, the NMDAR expression profile changed to similar to an embryonic one, and could promote cell apoptosis. Furthermore, these data suggest the involvement of other EDP receptors.
Conclusion
Our study shows, for the first time, that the VGVAPG peptide causes Ca2+ influx through NMDAR in mouse astrocytes in vitro. Silencing of the Glb1, GluN1, GluN2A, and GluN2B genes prevented a VGVAPG peptide-induced increase in the Ca2+ level. Nifedipine does not completely reduce VGVAPG peptide-activated ROS production, whereas MK-801, verapamil, and Src kinase inhibitor I reduce VGVAPG peptide-activated Ca2+ influx and ROS production. There appears to be a potential role of Src kinase in signal transduction from EBP to NMDAR. Moreover, the VGVAPG peptide induced an increase in the expression of GluN2A NMDAR subunits, which promote cell survival in adult life. However, due to the lack of sufficient data explaining the molecular mechanism of action of the VGVAPG peptide in the nervous system, more studies on this topic are necessary.
Materials and Methods
Reagents
We purchased Dulbecco’s Modified Eagle’s Medium/Hams F-12 (DMEM/F12) without phenol red, along with trypsin, rosiglitazone, pioglitazone, penicillin, streptomycin, amphotericin B, nifedipine, verapamil, MK-801, and dimethyl sulfoxide (DMSO) from Sigma–Aldrich (St. Louis, MO, USA). Glb1 gene siRNA (sc-61342), NMDAζ1 (GluN1) (sc-36082), NMDAε1 (GluN2A) (sc-36084), and NMDAε2 (GluN2B) (sc-36086) gene siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The INTERFERin siRNA transfection reagent was purchased from Polyplus-transfection (Illkirch, France). The VGVAPG and VVGPGA peptides were synthesized by LipoPharm.pl (Gdańsk, Poland). Charcoal/dextran-treated fetal bovine serum (FBS) was purchased from EURx (Gdańsk, Poland). The Src kinase inhibitor I (14592) and Fluo-3 AM (14960) were purchased from the Cayman Chemical Company (Michigan, USA). The High Capacity cDNA – Reverse Transcription Kit and the TaqMan probes corresponding to specific genes encoding Actb (Mm00607939_s1), GluN1 (Mm00433800_m1), GluN2A (Mm00433802_m1), and GluN2B (Mm00433820_m1) were obtained from Life Technologies Applied Biosystems (Foster City, CA, USA). Stock solutions of the VGVAPG and VVGPGA peptides were prepared in DMSO, and then added to the DMEM/F12 medium. The final concentration of DMSO in the culture medium was always 0.1%.
Astrocyte-enriched cell culture
All experiments were conducted on mouse astrocyte cell cultures, which were prepared from the fetuses of pregnant female Swiss mice according to a previously described method that generates an almost pure culture of astrocytes (>98% astrocytes)44(supplementary data). Cells were cultured in DMEM/F12 1:1 without phenol red that was supplemented with 10% FBS, 100 U/mL penicillin, 0.10 mg/mL streptomycin, and 250 ng/mL amphotericin B, maintained at 37 °C in a humidified atmosphere containing 5% CO2. The astrocytes were seeded at a density of 5 × 105 per well in a 96-well plate, for colorimetric and fluorometric analysis, and 60 × 105 per well in a 12-well plate for mRNA analysis. The culture medium was changed prior to the experiment with the VGVAPG peptide and a tool compound was selected for this study.
siRNA gene-silencing procedure
Glb1, GluN1, GluN2A, and GluN2B siRNA were used to inhibit the expression of EBP and NMDAR subunits in mouse primary astrocytes. The experiment was conducted in accordance with a previously described procedure51. Briefly, appropriate siRNA was applied for 7 h at a final concentration of 50 nM in antibiotic-free medium containing the siRNA transfection reagent INTERFERin. After transfection, the astrocytes were cultured for 12 h prior to commencing the experiment. Vehicle controls included positive siRNA and scramble siRNA containing a scrambled sequence that did not lead to specific degradation of any known cellular mRNA. Knockdown of the Glb1 gene was estimated at 39% of the control mRNA and 60.05% of the control protein as previously described44,50. Knockdown of the GluN1, GluN2A, or GluN2B genes were estimated at 22.02, 25.21 and 24.32%, respectively of the control mRNA.
Ca2+ determination in astrocytes
To measure the calcium level, we used the Fluo-3/AM indicator, which exhibits increased fluorescence emission intensity on binding to Ca2+ and, therefore, is used to detect increases in the level of intracellular calcium52. Briefly, a final concentration of 4 μM Fluo-3/AM dye and 0.04% Pluronic F-127 in the loading medium (DMEM/F12 without FBS) was used for 60 min before compound treatment. Thereafter, the cells were washed three times with 200 μL PBS without Ca2+ supplemented with HEPES. After 15 and 30 min of incubation of the cells with 10 nM VGVAPG or VVGPGA peptide (5% CO2 at 37 °C), the fluorescence of Fluo-3 was measured with a microplate reader (FilterMax F5) at maximum excitation and emission spectra of 485 and 535 nm, respectively.
ROS measurement
The fluorogenic dye H2DCFDA was used to detect intracellular ROS according to a previously described method53. Briefly, a final concentration of 5 μM H2DCFDA was used. The cells were incubated in H2DCFDA in a serum-free and phenol red-free DMEM/F12 medium for 45 min before compound treatment. After 3- and 6-h incubation of the cells with 10 nM VGVAPG or VVGPGA peptide (5% CO2 at 37 °C), the culture medium was replaced with fresh medium to remove the extracellular residual DCF and the studied compound to reduce the fluorescence background. Cells treated with 0.3% hydrogen peroxide (H2O2) were used as a positive control (results not shown). DCF fluorescence was measured with a microplate reader (FilterMax F5) at maximum excitation and with emission spectra of 485 and 535 nm, respectively.
Real-time PCR analysis of mRNAs specific to genes encoding GluN1, GluN2A, and GluN2B
The experiment was conducted through a previously described procedure54. For the real-time PCR assay, astrocytes were seeded onto 12-well plates and initially cultured for 24 h. After 6-h exposure to 10 nM VGVAPG peptide, the samples were collected, and total RNA was extracted from the astrocytes using a Qiagen RNeasy Mini Kit according to the manufacturer’s instructions. Both the quality and quantity of the RNA were determined spectrophotometrically at 260 and 280 nm, respectively (ND/1000 UV/Vis; Thermo Fisher NanoDrop, USA). Two-step real-time reverse transcription (RT)-PCR was conducted with both the RT reaction and the quantitative PCR (qPCR) run using the CFX Real Time System (BioRad, USA). The RT reaction was carried out at a final volume of 20 μL with 180 ng RNA (as a cDNA template) using the cDNA reverse transcription kit in accordance with the manufacturer’s instructions. Products from the RT reaction were amplified using the FastStart Universal Probe Master (Rox) kit with TaqMan probes as primers for specific genes encoding Actb, Glb1, GluN1, GluN2A, and GluN2B. Amplification was carried out in a total volume of 20 μL containing 1× FastStart Universal Probe Master (Rox) and 1.5 μL of the RT product, which was used as the PCR template. Standard qPCR procedures were carried out as follows: 2 min at 50 °C and 10 min at 95 °C, followed by 45 cycles of 15 s at 95 °C and 1 min at 60 °C. The threshold value (Ct) for each sample was set during the exponential phase, and the ΔΔCt method was used for data analysis. Actb was used as the reference gene. Random-peptide-sequence VVGPGA was used as control and did not affect GluN1, GluN2A, and GluN2B mRNA expression (data not shown).
Statistical analysis
Data are presented as means ± SD of three independent experiments. Each treatment was repeated six times (n = 6) and measured in triplicate. Data were analyzed using Multi-Mode Analysis STATISTICA 13.0 software and normalized to fluorescence in the vehicle-treated cells (% of control). One-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison procedure was applied, with ***p < 0.001, **p < 0.01, and *p < 0.05 indicative of statistical significance versus the control.
Ethical approval
Animal care followed official governmental guidelines, and all efforts were made to minimize the number of animals used and their suffering. All procedures were performed in accordance with the National Institutes of Health Guidelines for the Care. Moreover, all procedures were approved by a Bioethics Commission (no. 46/2014, First Local Ethical Committee on Animal Testing at the Jagiellonian University in Krakow), as compliant with the European Union law.
Acknowledgements
This work was supported through statutory funds from the University of Information Technology and Management in Rzeszow, Poland (Grant No. DS MN 503-05-02-03).
Author contributions
K.A.S. designed the research. K.A.S. performed the cell culture research. K.A.S. analyzed the data. K.A.S. and J.G. wrote the manuscript. K.A.S. obtained financial support for the project. All authors read and approved the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Jung S, Rutka JT, Hinek A. Tropoelastin and Elastin Degradation Products Promote Proliferation of Human Astrocytoma Cell Lines. J. Neuropathol. Exp. Neurol. 1998;57:439–448. doi: 10.1097/00005072-199805000-00007. [DOI] [PubMed] [Google Scholar]
- 2.Gmiński J, Mykała-Cieśla J, Machalski M, Dróżdż M. Anti-elastin antibodies in patients with lung cancer. Immunol. Lett. 1992;33:211–215. doi: 10.1016/0165-2478(92)90049-T. [DOI] [PubMed] [Google Scholar]
- 3.Antonicelli F, Bellon G, Debelle L, Hornebeck W. Elastin-elastases and inflamm-aging. Curr. Top. Dev. Biol. 2007;79:99–155. doi: 10.1016/S0070-2153(06)79005-6. [DOI] [PubMed] [Google Scholar]
- 4.Tzvetanov P, Nicoloff G, Rousseff R, Christova P. Increased levels of elastin-derived peptides in cerebrospinal fluid of patients with lacunar stroke. Clin. Neurol. Neurosurg. 2008;110:239–244. doi: 10.1016/j.clineuro.2007.11.003. [DOI] [PubMed] [Google Scholar]
- 5.Nicoloff G, Tzvetanov P, Christova P, Baydanoff S. Detection of elastin derived peptides in cerebrospinal fluid of patients with first ever ischaemic stroke. Neuropeptides. 2008;42:277–282. doi: 10.1016/j.npep.2008.02.003. [DOI] [PubMed] [Google Scholar]
- 6.Hornebeck W, Emonard H, Monboisse J-C, Bellon G. Matrix-directed regulation of pericellular proteolysis and tumor progression. Semin. Cancer Biol. 2002;12:231–41. doi: 10.1016/S1044-579X(02)00026-3. [DOI] [PubMed] [Google Scholar]
- 7.Maquart FX, Bellon G, Pasco S, Monboisse JC. Matrikines in the regulation of extracellular matrix degradation. Biochimie. 2005;87:353–360. doi: 10.1016/j.biochi.2004.10.006. [DOI] [PubMed] [Google Scholar]
- 8.Adair-Kirk TL, Senior RM. Fragments of extracellular matrix as mediators of inflammation. Int. J. Biochem. Cell Biol. 2008;40:1101–1110. doi: 10.1016/j.biocel.2007.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Privitera S, Prody Ca, Callahan JW, Hinek A. The 67-kDa enzymatically inactive alternatively spliced variant of β- galactosidase is identical to the elastin/laminin-binding protein. J. Biol. Chem. 1998;273:6319–6326. doi: 10.1074/jbc.273.11.6319. [DOI] [PubMed] [Google Scholar]
- 10.Verkhratsky Alexei, Nedergaard Maiken, Hertz Leif. Why are Astrocytes Important? Neurochemical Research. 2014;40(2):389–401. doi: 10.1007/s11064-014-1403-2. [DOI] [PubMed] [Google Scholar]
- 11.Verkhratsky A, Kirchhoff F. NMDA Receptors in glia. Neuroscientist. 2007;13:28–37. doi: 10.1177/1073858406294270. [DOI] [PubMed] [Google Scholar]
- 12.Shigetomi E, Bowser DN, Sofroniew MV, Khakh BS. Two Forms of Astrocyte Calcium Excitability Have Distinct Effects on NMDA Receptor-Mediated Slow Inward Currents in Pyramidal Neurons. J. Neurosci. 2008;28:6659–6663. doi: 10.1523/JNEUROSCI.1717-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rzigalinski BA, Liang S, McKinney JS, Willoughby KA, Ellis EF. Effect of Ca2+ on In Vitro Astrocyte Injury. J. Neurochem. 2002;68:289–296. doi: 10.1046/j.1471-4159.1997.68010289.x. [DOI] [PubMed] [Google Scholar]
- 14.Maneshi MM, Sachs F, Hua SZ. A Threshold Shear Force for Calcium Influx in an Astrocyte Model of Traumatic Brain Injury. J. Neurotrauma. 2014;32:1020–1029. doi: 10.1089/neu.2014.3677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dzamba D, Honsa P, Anderova M. NMDA Receptors in Glial Cells: Pending Questions. Curr. Neuropharmacol. 2013;11:250–262. doi: 10.2174/1570159X11311030002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stroebel D, Casado M, Paoletti P. Triheteromeric NMDA receptors: from structure to synaptic physiology. Curr. Opin. Physiol. 2018;2:1–12. doi: 10.1016/j.cophys.2017.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Watanabe M, Inoue Y, Sakimura K, Mishina M. Developmental changes in distribution of NMDA receptor channel subunit mRNAs. Neuroreport. 1992;3:1138–40. doi: 10.1097/00001756-199212000-00027. [DOI] [PubMed] [Google Scholar]
- 18.Monyer H, Burnashev N, Laurie DJ, Sakmann B, Seeburg PH. Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron. 1994;12:529–40. doi: 10.1016/0896-6273(94)90210-0. [DOI] [PubMed] [Google Scholar]
- 19.Gupta K, Hardingham GE, Chandran S. NMDA receptor-dependent glutamate excitotoxicity in human embryonic stem cell-derived neurons. Neurosci. Lett. 2013;543:95–100. doi: 10.1016/j.neulet.2013.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sun Z-W, Zhang L, Zhu S-J, Chen W-C, Mei B. Excitotoxicity effects of glutamate on human neuroblastoma SH-SY5Y cells via oxidative damage. Neurosci. Bull. 2010;26:8–16. doi: 10.1007/s12264-010-0813-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nato G, et al. Striatal astrocytes produce neuroblasts in an excitotoxic model of Huntington’s disease. Development. 2015;142:840–845. doi: 10.1242/dev.116657. [DOI] [PubMed] [Google Scholar]
- 22.Dong X, Wang Y, Qin Z. Molecular mechanisms of excitotoxicity and their relevance to pathogenesis of neurodegenerative diseases. Acta Pharmacol. Sin. 2009;30:379–387. doi: 10.1038/aps.2009.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jacob MP, Fülöp T, Foris G, Robert L. Effect of elastin peptides on ion fluxes in mononuclear cells, fibroblasts, and smooth muscle cells. Proc. Natl. Acad. Sci. USA. 1987;84:995–999. doi: 10.1073/pnas.84.4.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Faury G, Usson Y, Robert-Nicoud M, Robert L, Verdetti J. Nuclear and cytoplasmic free calcium level changes induced by elastin peptides in human endothelial cells. Proc. Natl. Acad. Sci. USA. 1998;95:2967–2972. doi: 10.1073/pnas.95.6.2967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Faury G, et al. Action of tropoelastin and synthetic elastin sequences on vascular tone and on free Ca2+ level in human vascular endothelial cells. Circ. Res. 1998;82:328–36. doi: 10.1161/01.RES.82.3.328. [DOI] [PubMed] [Google Scholar]
- 26.Coquerel B, et al. Elastin-derived peptides: Matrikines critical for glioblastoma cell aggressiveness in a 3-D system. Glia. 2009;57:1716–1726. doi: 10.1002/glia.20884. [DOI] [PubMed] [Google Scholar]
- 27.Görlach A, Bertram K, Hudecova S, Krizanova O. Calcium and ROS: A mutual interplay. Redox Biol. 2015;6:260–271. doi: 10.1016/j.redox.2015.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dupont A, et al. Impairment of neutrophil reactivity to elastin peptides in COPD. Thorax. 2013;68:421–8. doi: 10.1136/thoraxjnl-2012-201833. [DOI] [PubMed] [Google Scholar]
- 29.Robert L, Jacob MP, Frances C, Godeau G, Hornebeck W. Interaction between elastin and elastases and its role in the aging of the arterial wall, skin and other connective tissues. A review. Mech. Ageing Dev. 1984;28:155–66. doi: 10.1016/0047-6374(84)90015-0. [DOI] [PubMed] [Google Scholar]
- 30.Scandolera A, et al. Uncoupling of Elastin Complex Receptor during In Vitro Aging Is Related to Modifications in Its Intrinsic Sialidase Activity and the Subsequent Lactosylceramide Production. PLoS One. 2015;10:e0129994. doi: 10.1371/journal.pone.0129994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gayral S, et al. Elastin-derived peptides potentiate atherosclerosis through the immune Neu1-PI3K pathway. Cardiovasc. Res. 2014;102:118–127. doi: 10.1093/cvr/cvt336. [DOI] [PubMed] [Google Scholar]
- 32.Szychowski KA, Gmiński J. The VGVAPG Peptide Regulates the Production of Nitric Oxide Synthases and Reactive Oxygen Species in Mouse Astrocyte Cells In Vitro. Neurochem. Res. 2019;44:1127–1137. doi: 10.1007/s11064-019-02746-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Szychowski KA, Rombel-Bryzek A, Dołhańczuk-Śródka A, Gmiński J. Antiproliferative Effect of Elastin-Derived Peptide VGVAPG on SH-SY5Y Neuroblastoma Cells. Neurotox. Res. 2019;36:503–514. doi: 10.1007/s12640-019-00040-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hernández-Fonseca K, Cárdenas-Rodríguez N, Pedraza-Chaverri J, Massieu L. Calcium-dependent production of reactive oxygen species is involved in neuronal damage induced during glycolysis inhibition in cultured hippocampal neurons. J. Neurosci. Res. 2008;86:1768–1780. doi: 10.1002/jnr.21634. [DOI] [PubMed] [Google Scholar]
- 35.Duca L, et al. Matrix ageing and vascular impacts: Focus on elastin fragmentation. Cardiovasc. Res. 2016;110:298–308. doi: 10.1093/cvr/cvw061. [DOI] [PubMed] [Google Scholar]
- 36.Mochizuki S, Brassart B, Hinek A. Signaling pathways transduced through the elastin receptor facilitate proliferation of arterial smooth muscle cells. J. Biol. Chem. 2002;277:44854–44863. doi: 10.1074/jbc.M205630200. [DOI] [PubMed] [Google Scholar]
- 37.van Zwieten PA, Pfaffendorf M. Similarities and differences between calcium antagonists: pharmacological aspects. J. Hypertens. Suppl. 1993;11:S3–11. doi: 10.1097/00004872-199303001-00002. [DOI] [PubMed] [Google Scholar]
- 38.Maurice P, et al. Elastin fragmentation and atherosclerosis progression: The elastokine concept. Trends Cardiovasc. Med. 2013;23:211–221. doi: 10.1016/j.tcm.2012.12.004. [DOI] [PubMed] [Google Scholar]
- 39.Fahem A, et al. Elastokine-mediated up-regulation of MT1-MMP is triggered by nitric oxide in endothelial cells. Int. J. Biochem. Cell Biol. 2008;40:1581–1596. doi: 10.1016/j.biocel.2007.11.022. [DOI] [PubMed] [Google Scholar]
- 40.Gmiński J, Wȩglarz L, Dróżdż M, Goss M. Pharmacological modulation of the antioxidant enzymes activities and the concentration of peroxidation products in fibroblasts stimulated with elastin peptides. Gen. Pharmacol. Vasc. Syst. 1991;22:495–497. doi: 10.1016/0306-3623(91)90012-U. [DOI] [PubMed] [Google Scholar]
- 41.Chen BS, Roche KW. Regulation of NMDA receptors by phosphorylation. Neuropharmacology. 2007;53:362–368. doi: 10.1016/j.neuropharm.2007.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee HK. Synaptic plasticity and phosphorylation. Pharmacol. Ther. 2006;112:810–832. doi: 10.1016/j.pharmthera.2006.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chen T, Zhang B, Li G, Chen L, Chen L. Simvastatin enhances NMDA receptor GluN2B expression and phosphorylation of GluN2B and GluN2A through increased histone acetylation and Src signaling in hippocampal CA1 neurons. Neuropharmacology. 2016;107:411–421. doi: 10.1016/j.neuropharm.2016.03.028. [DOI] [PubMed] [Google Scholar]
- 44.Szychowski KA, Wójtowicz AK, Gmiński J. Impact of Elastin-Derived Peptide VGVAPG on Matrix Metalloprotease-2 and -9 and the Tissue Inhibitor of Metalloproteinase-1, -2, -3 and -4 mRNA Expression in Mouse Cortical Glial Cells In Vitro. Neurotox. Res. 2019;35:100–110. doi: 10.1007/s12640-018-9935-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Turman JE, Lee OK, Chandler SH. Differential NR2A and NR2B expression between trigeminal neurons during early postnatal development. Synapse. 2002;44:76–85. doi: 10.1002/syn.10059. [DOI] [PubMed] [Google Scholar]
- 46.Ritter LM, Vazquez DM, Meador-Woodruff JH. Ontogeny of ionotropic glutamate receptor subunit expression in the rat hippocampus. Brain Res. Dev. Brain Res. 2002;139:227–36. doi: 10.1016/S0165-3806(02)00572-2. [DOI] [PubMed] [Google Scholar]
- 47.Zhang XY, Liu AP, Ruan DY, Liu J. Effect of developmental lead exposure on the expression of specific NMDA receptor subunit mRNAs in the hippocampus of neonatal rats by digoxigenin-labeled in situ hybridization histochemistry. Neurotoxicol. Teratol. 2002;24:149–160. doi: 10.1016/S0892-0362(01)00210-0. [DOI] [PubMed] [Google Scholar]
- 48.Liu Y, et al. NMDA receptor subunits have differential roles in mediating excitotoxic neuronal death both in vitro and in vivo. J. Neurosci. 2007;27:2846–57. doi: 10.1523/JNEUROSCI.0116-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Vizi ES, Kisfali M, Lőrincz T. Role of nonsynaptic GluN2B-containing NMDA receptors in excitotoxicity: evidence that fluoxetine selectively inhibits these receptors and may have neuroprotective effects. Brain Res. Bull. 2013;93:32–8. doi: 10.1016/j.brainresbull.2012.10.005. [DOI] [PubMed] [Google Scholar]
- 50.Szychowski KA, Gmiński J. Impact of elastin-derived VGVAPG peptide on bidirectional interaction between peroxisome proliferator-activated receptor gamma (Pparγ) and beta-galactosidase (β-Gal) expression in mouse cortical astrocytes in vitro. Naunyn. Schmiedebergs. Arch. Pharmacol. 2019;392:405–413. doi: 10.1007/s00210-018-1591-4. [DOI] [PubMed] [Google Scholar]
- 51.Szychowski KA, et al. Triclosan-Evoked Neurotoxicity Involves NMDAR Subunits with the Specific Role of GluN2A in Caspase-3-Dependent Apoptosis. Mol. Neurobiol. 2019;56:1–12. doi: 10.1007/s12035-018-1083-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zieminska E, Lenart J, Diamandakis D, Lazarewicz JW. The Role of Ca2+ Imbalance in the Induction of Acute Oxidative Stress and Cytotoxicity in Cultured Rat Cerebellar Granule Cells Challenged with Tetrabromobisphenol A. Neurochem. Res. 2017;42:777–787. doi: 10.1007/s11064-016-2075-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Szychowski KA, Rybczyńska-Tkaczyk K, Leja ML, Wójtowicz AK, Gmiński J. Tetrabromobisphenol A (TBBPA)-stimulated reactive oxygen species (ROS) production in cell-free model using the 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) assay—limitations of method. Environ. Sci. Pollut. Res. 2016;23:12246–12252. doi: 10.1007/s11356-016-6450-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Szychowski KA, et al. Anticancer properties of 4-thiazolidinone derivatives depend on peroxisome proliferator-activated receptor gamma (PPARγ) Eur. J. Med. Chem. 2017;141:162–168. doi: 10.1016/j.ejmech.2017.09.071. [DOI] [PubMed] [Google Scholar]