

Abstract
Estrogens protect against diet-induced obesity in women and female rodents. For example, a lack of estrogens in postmenopausal women is associated with an increased risk of weight gain, cardiovascular diseases, low-grade inflammation, and cancer. Estrogens act with leptin to regulate energy homeostasis in females. Leptin-deficient mice (ob/ob) exhibit morbid obesity and insulin resistance. The gut microbiome is also critical in regulating metabolism. The present study investigates whether estrogens and leptin modulate gut microbiota in ovariectomized ob/ob (obese) or heterozygote (lean) mice fed high-fat diet (HFD) that received either 17β-Estradiol (E2) or vehicle implants. E2 attenuated weight gain in both genotypes. Moreover, both obesity (ob/ob mice) and E2 were associated with reduced gut microbial diversity. ob/ob mice exhibited lower species richness than control mice, while E2-treated mice had reduced evenness compared with vehicle mice. Regarding taxa, E2 was associated with an increased abundance of the S24-7 family, while leptin was associated with increases in Coriobacteriaceae, Clostridium and Lactobacillus. Some taxa were affected by both E2 and leptin, suggesting these hormones alter gut microbiota of HFD-fed female mice. Understanding the role of E2 and leptin in regulating gut microbiota will provide important insights into hormone-dependent metabolic disorders in women.
Subject terms: Endocrine system and metabolic diseases, Microbiome
Introduction
Estrogens profoundly influence energy homeostasis1–3, as well as reproductive physiology and behavior4–6. Estrogens reduce food intake, attenuate body weight gain and adiposity, and increase physical activity in humans and rodents1,2. Postmenopausal women have lower levels of circulating estrogens and an increased tendency to gain fat weight, which increases their risk for obesity, cardiovascular disease, stroke, and type 2 diabetes7–9. Similarly, in mice on a high-fat diet (HFD), ovariectomy increases energy intake and obesity, while estradiol (E2) treatment prevents weight gain2,10–13, indicating that estrogens protect against HFD-induced obesity.
Leptin is a peptide hormone secreted primarily by adipocytes, which acts primarily in the brain to stimulate metabolism, promote satiety, and regulate fat storage14–16. A mutation in the ob gene that encodes leptin results in mice lacking the hormone (ob/ob)17. While phenotypically normal at birth, ob/ob mice quickly develop obesity and diabetes18. Additionally, ob/ob mice exhibit increased food intake and decreased physical activity, energy metabolism, and body temperature compared to lean controls, making ob/ob mice an excellent genetic model of obesity19–21. Administering leptin to adult ob/ob mice reverses these effects by decreasing food intake, increasing energy output and decreasing circulating levels of glucose and insulin19,22.
Leptin and estrogen signaling pathways interact to influence reproduction and energy metabolism. High levels of E2 are associated with increased leptin sensitivity in both male and female rodents23. In contrast, decreased estrogens in ovariectomized mice and postmenopausal women increases leptin resistance, resulting in obesity24,25. While female ob/ob mice do not have an estrous cycle, leptin administration restores fertility, including successful ovulation, pregnancy, and birth, indicating the profound effects of leptin on reproduction26. While these findings indicate functional interactions between E2 and leptin, primarily through the convergence of their central pathways25,27, these hormonal effects on peripheral regulators of energy metabolism are not well understood.
The gut microbiome, which is composed primarily of the bacteria in the intestinal tract and their metabolites, has profound effects on energy metabolism28. The gut microbiome aids in digestion and absorption of macro- and micro-nutrients from food29. Gut microbiota are also essential modulators of host immune homeostasis. Protective polysaccharides produced from the break-down of dietary fibers attenuate inflammation30,31. Gut microbiota can also synthesize and metabolize neurotransmitters and hormones to alter host physiology32–34. Furthermore, changes in body weight have been associated with changes in gut microbial diversity in humans and rodents35. For example, the transfer of gut microbiota from obese human or mice donors results in an obese phenotype in recipient mice36–38. Similarly, depletion of gut microbiota using antibiotics increases adiposity in mice39.
A variety of factors, including host genetics, diet, stress, and gonadal hormones can alter the gut microbiome33,34,40–42. Sex differences in gut microbiota have been reported in humans and rodents41,43. In a European population, higher levels of Bacteroidetes and Prevotella were observed in men compared to women44. Male mice exhibited higher abundances of Lachnospiraceae (phylum Firmicutes) and Parabacteroides spp. (phylum Bacteroidetes) and Proteobacteria than female mice33,45. Additionally, testosterone administration to female neonatal rats decreased gut microbial diversity during adulthood, and increased the ratio of the two most abundant phyla, Firmicutes and Bacteroidetes40. The hormone-dependent changes in gut microbiota were more robust compared to the diet-induced changes in these rats40. Ovariectomy also alters gut microbial diversity in adult mice33,45,46. While obesity and ob/ob genotype are associated with a reduction in Bacteroidetes/Firmicutes ratio in humans and male mice, respectively47–50, the effects of leptin on gut microbiota have not been studied in females. Since estrogens and leptin interact to profoundly affect female metabolic homeostasis10,24, it is important to understand the effects of leptin and its interaction with estrogens. Using a mouse model of obesity (ob/ob), the present study investigated if estradiol and leptin associate with longitudinal changes in gut microbiota and energy homeostasis in female mice on a HFD.
Results
Estradiol and ob/ob genotype altered weight gain and food intake in female mice on a HFD
Body weights were collected every four days from the four experimental groups: control E2 (n = 13), control Veh (n = 12), ob/ob E2 (n = 8), and ob/ob Veh (n = 8). Longitudinal analysis of weight change showed that E2 decreased weight gain (F = 15.67, p < 0.001; ANOVA) in female mice fed a HFD. E2 treatment attenuated weight gain from day 11 through the end of the study (Fig. 1A,B). Within-genotype comparison showed that E2 attenuated weight gain in Het mice from days 11–35, compared to Veh mice. For ob/ob mice, E2 reduced weight gain on days 11, 15 and 27–35, and showed strong trends on days 19 (p = 0.059) and 23 (p = 0.063). Because the Het and ob/ob mice had different weights at the beginning of the study, the % weight gain was also analyzed to remove the bias due to existing differences in weights prior to hormone and diet manipulation. Similar to the effects on body weight, E2 treatment reduced % weight gain from days 11–35. In particular, E2-treated Het mice gained less % weight than Veh Het mice on days 11–35 (Fig. 1B). Within the ob/ob mice, E2 decreased % weight gain on days 7–15 and 27–35.
Figure 1.

Estradiol and leptin alter weight gain and food intake in adult female ob/ob and lean control mice. (A) Average weight, (B) percent weight gain and (C) food intake (2 mice/cage) of heterozygote controls (Het) E2 (n = 13), Het Veh (n = 12), ob/ob E2 (n = 8) and ob/ob Veh (n = 7) with arrow indicating the start of high fat diet. The effects of genotype on average weight (A) and food intake (C) were present on all days. Days with effects of E2 are denoted by *(p < 0.05; ANOVA). Error bars indicate ± SEM.
Genotype also affected weight gain (F = 1119.79, p < 0.001; ANOVA) and % weight gain on longitudinal measures (F = 4.23, p = 0.047; ANOVA). ob/ob mice weighed more than control Het mice at all time points regardless of E2 treatment. At the start of the study, ob/ob mice weighed approximately twice that of the Het mice, which persisted throughout the study. Finally, there was an interaction of E2 and genotype on body weight (F = 4.21, p = 0.048; ANOVA), with E2-treated Het mice gaining more % weight than E2-treated ob/ob mice on days 4 and 7 (Fig. 1B). The Veh mice did not differ in % weight gain between the two genotypes.
E2 treatment reduced HFD consumption (F = 11.04, p = 0.002; ANOVA) from days 7 to 19 (Fig. 1C). Analysis within each genotype showed that E2 attenuated food intake in ob/ob mice, but not in Hets, on days 7, 11 and 15. Comparison between lean Hets and ob/ob mice showed that obesity profoundly increased food intake (F = 110.5, p < 0.001, ANOVA). Het mice consumed less calories than ob/ob mice throughout the study. Within the E2-treated groups, an increase in food intake in ob/ob groups compared to E2 Het mice was observed only at the beginning (days 7 and 11) and end (days 27, 31 and 35) of the study. There was an interaction between genotype and treatment on food intake (F = 5.52, p = 0.029; ANOVA). We also observed a decrease in food intake on day 19 in ob/ob Veh mice, but not other groups, following surgery on day 16 for removal of BrdU osmotic pumps.
Estradiol and ob/ob genotype alter gut microbial diversity during HFD
To assess the impact of E2 treatment and obesity on α-diversity of the gut microbiota, 16S rDNA from fecal samples from ob/ob and Het mice were analyzed. The data set contained 137 samples after removing samples with less than 2,000 reads. The 16S rDNA targeted sequencing yielded 25,399 reads/sample on average (range 11,154–82,733). Clustering of these 16S sequence tags produced 473 OTUs at 97% similarity level. The identified OTUs belonged to 10 phyla, 42 families and 62 genera based on the Greengenes database. During HFD, ob/ob mice had a lower number of total identified OTUs than Het mice, indicating that ob/ob mice had lower species richness than control mice (Fig. 2A; p = 0.002, linear regression). In addition, E2 treatment was associated with lower species evenness in both genotypes, suggesting that E2-treated mice have a more heterogeneous distribution of gut microbial communities than Veh mice (Fig. 2B; p = 0.0008).
Figure 2.

Estradiol and obesity reduce gut microbiota alpha diversity. (A) ob/ob genotype is associated with lower species richness as measured by Observed Taxonomic Units. (B) E2 treatment is associated with lower species evenness as measured by the Pielou’s evenness index. “a” and “b” indicate groups with different species richness while “c” and “d” indicate groups with different species evenness (p < 0.05, linear regression). Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6).
The ordination plot based on Bray-Curtis distance, which measures differences due to relative abundances and composition, showed distinct clustering of microbial communities from the four experimental groups (Fig. 3; p = 0.004, PERMANOVA). The first and second principal coordinates (PCo1 and PCo2) explain 29.4% and 17.5% of the variation in the microbial communities, respectively. Genotype, most likely through obesity, accounted for most of the difference observed as reflected by PCo1 (p = 4E-8, t-test, Fig. 3). There was also a clear effect of E2 treatment, represented by PCo2 (p = 0.001, t-test, Fig. 3).
Figure 3.
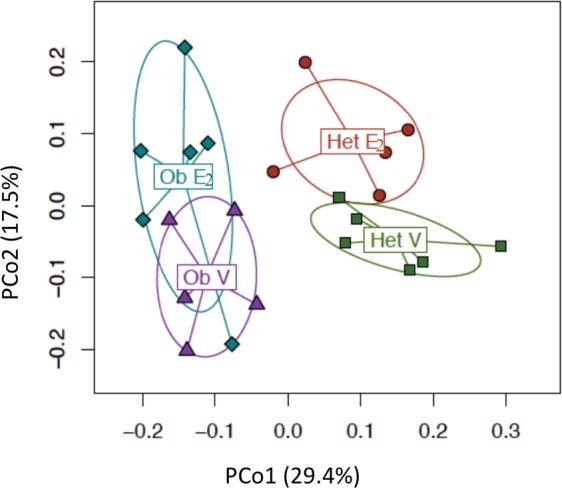
Gut microbial communities distinctly cluster as an effect of estradiol treatment and leptin. PCo1 and PCo2 clustering of each group. Bray-Curtis distance was used to calculate principal coordinates 1 and 2 and PERMANOVA, to calculate the association with leptin or E2 on aggregate data over all days. Ob E2 = E2-treated ob/ob (n = 6), Ob V = Vehicle ob/ob (n = 6), Het E2 = E2-treated Het (n = 6), and Het V = Vehicle Het mice (n = 6).
Similar analyses were performed using Bray-Curtis distance measures on each day to characterize the temporal changes in gut microbiota. The effect of E2 on β-diversity was robust and continuously observed from the 2nd week of OVX (days 15–35). The effect of genotype was also present on days 7,15, 23 and 35, and a trend was observed on the remaining two days (4 and 31) (Table 1).
Table 1.
PERMANOVA P-values for individual days based on Bray-Curtis distance.
| Day | Day 4 | Day 7 | Day 15 | Day 23 | Day 31 | Day 35 |
|---|---|---|---|---|---|---|
| Treatment | 0.095 | 0.234 | 0.011* | 0.044* | 0.023* | 0.022* |
| Genotype | 0.081 | 0.004* | 0.009* | 0.001* | 0.06 | 0.011* |
*Indicate a p-value of <0.05.
Estradiol treatment alters relative abundances of intestinal microbiota
Gut microbial community composition was further analyzed on aggregate data from all days to identify the relative abundances at phylum, family and genus levels (Fig. 4). E2-mediated shifts in relative abundances was evident on many of these taxa levels. In all four experimental groups, the most prevalent phyla (>90%) were Firmicutes, Bacteroidetes, Actinobacteria and Tenericutes (Fig. 4A). The two most abundant families were S24-7 and Lachnospiraceae, within Bacteroidetes and Firmicutes, respectively (Fig. 4B). At the genus level, Coprococcus and Ruminococcus (both within phylum Firmicutes) were the most abundant (Fig. 4C).
Figure 4.

Gut microbiota associate with estradiol treatment and obesity at multiple taxa levels. Microbiota community structure at the (A) phylum, (B) family and (C) genus level, separated by estradiol treatment and ob/ob genotype. Data are shown as relative proportion of the taxa identified. Taxa with prevalence of >10% or with a maximum proportion of >0.2% were included. Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6), and ob/ob Veh mice (n = 6). An overdispersed Poisson regression was used to calculate associations of taxa with leptin and E2.
To identify whether the differences in community structure were driven by differences in the relative abundances of particular microbial taxa, a differential abundance analysis based on overdispersed Poisson regression model was run on aggregate data to uncover the effects of treatment (Fig. 5A) and genotype (Fig. 5B). In the cladogram, the nodes in the first circle represent phyla, and the extending outer nodes in each level represent lower taxa within each phylum. A total of 26 taxa were differentially associated between E2 and Veh groups (Fig. 6A). Firmicutes were more associated with Veh treatment, while Bacteroidetes were associated with E2 treatment (Figs. 5A and 6A). Within Firmicutes, the class Erysipelotrichi, including Allobaculum and Coprobacillus spp., were more abundant in Veh than E2 mice. Similarly, the relative abundances of the class Bacilli, including its families Lactobacillaceae and Peptostreptococcaceae, were greater in Veh than E2 groups. Another family, Streptococcaceae, including Lactococcus spp., were also more associated with Veh mice. In contrast, the family Ruminococcaceae was more abundant in E2 mice. Within Bacteroidetes, the class Bacteroidia, and its lower taxa Bacteroidales and S24-7, were more abundant in E2 than Veh mice (Figs. 5A and 6A).
Figure 5.

Estradiol and obesity associate with changes in the gut microbial composition. (A) Phyla and lower level taxa associated with estradiol (E2) or vehicle (Veh) treatment. (B) Phyla and lower level taxa associated with ob/ob or Het genotype. Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6).
Figure 6.

Gut microbiota associate with estradiol treatment and obesity at multiple taxa levels. Boxplots showing microbial taxa that are significantly altered by (A) Estradiol (E2) treatment and (B) genotype (overdispersed Poisson regression). Analysis was done on aggregate data from all 6 day points. Data are shown as relative proportion of the taxa identified. Taxa with prevalence of >10% or with a maximum proportion of >0.2% were included. E2-treated ob/ob (n = 6), E2-treated Het (n = 6), Veh ob/ob (n = 6), vehicle Het mice (n = 6).
For the taxa that differed across groups as detected from the aggregated data, the effect of E2 treatment on relative abundances was analyzed for each day. A comprehensive list of the taxa, adjusted p-values (q-values) and fold changes between E2 and Veh mice for each day are provided in Supplemental Table 1. E2-treated mice resisted a decrease in relative abundances of Bacteroidetes compared to Veh mice on days 23 (q = 0.007) and 31 (q = 0.09) (Fig. 7A). A main driver of the shifts in Bacteroidetes was the family S24-7. E2-treated mice resisted a decrease in S24-7 abundances compared to Veh mice on day 23 (q = 0.015) (Fig. 7B).
Figure 7.

Estradiol and obesity associate with changes in the relative abundances of Bacteroidetes (phylum) and S24-7 (family) following the start of HFD. Relative abundances of the (A) the phylum Bacteroidetes and (B) its family S24-7 over time. *Indicates effects of E2 and # indicates effects of genotype (q < 0.1, overdispersed Poisson regression). Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6). Error bars indicate ± SEM.
Compared to Veh mice, E2-treated animals resisted changes in relative abundances of the phylum Firmicutes, with lower relative abundances of Firmicutes on days 23 (q = 0.01), 31 (q = 0.06) and 35 (q = 0.04) compared to Veh mice (Fig. 8A). Within Firmicutes, Lactobacillus spp. showed a significant reduction on day 31 (q = 0.05) in E2-treated mice (Fig. 8B).
Figure 8.

Estradiol and obesity associate with changes in the relative abundances of the phylum Firmicutes and its lower taxa. Relative abundances over time of the (A) phylum Firmicutes and genera (B) Clostridium, (C) Lactobacillus and (D) Coprococcus. *Indicates effects of E2 and # indicates effects of genotype (q < 0.1, overdispersed Poisson regression). Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6). Error bars indicate ± SEM.
Estradiol also altered the relative abundance of the phylum Actinobacteria from days 15 to 35. On these days, E2-treated mice resisted an increase in the relative abundances of Actinobacteria compared to Veh mice (Fig. 9A). The E2-mediated effect on Actinobacteria was due mostly to the family Coriobacteriaceae which resisted this increase also on days 15 (q = 0.001), 23 (q < 0.001) and 35 (q = 0.03) (Fig. 9B).
Figure 9.

Estradiol and obesity resist increases in the relative abundances of the phylum Actinobacteria and its lower taxa. Relative abundances of the (A) phylum Actinobacteria and (B) family Coriobacteriaceae over time. *Indicates effects of E2 and # indicates effects of genotype (q < 0.1 overdispersed Poisson regression). Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6). Error bars indicate ± SEM.
Obesity is associated with changes in relative abundances of gut microbiota
As the gut microbial communities of the female mice clustered separately based on obesity (in ob/ob genotype) (Fig. 3), we further analyzed the effects of obesity on relative abundances at the taxa levels (Fig. 5B). A total of 26 identified taxa showed a differential association between ob/ob and Het mice (Fig. 6B). Comparison of the gut microbial communities between genotypes revealed that Firmicutes, including its families Lactobacillaceae, Turibacteraceae, Peptostreptococcaceae, Dehalobacteriaceae, and Ruminococcaceae were more abundant in Het mice than ob/ob mice. At the genus level, Lactobacillus, Turibacter, Dehalobacterium, Dorea, Ruminococcus, and Oscillospira were more abundant in Het controls than ob/ob. Similarly, the phylum Actinobacteria and its family Coriobacteriaceae showed a greater abundance in control Het mice. In contrast, Enterococcus, Coprococcus, Lachnospira, Anaerotruncus and Coprobacillus spp. of the phylum Firmicutes were more abundant in ob/ob mice compared to Het mice (Figs. 5B and 6B).
Differential abundance analysis across days showed that genotype affected the relative abundances of multiple taxa (Supplemental Table 2). Leptin was associated with increases in the relative abundances of Bacteroidetes and its family S24-7 in Het mice at the beginning (day 4; q = 0.04), but by day 23 (after 19 days on HFD; q = 0.03), these were lower than in ob/ob mice (Fig. 7). Within Firmicutes, Clostridium, Lactobacillus and Lactococcus spp. were altered by genotype (Fig. 8). Lactobacillus and Clostridium spp. were more abundant in Het mice. The relative abundance of Lactobacillus increased in Het mice compared to ob/ob mice on days 4 (q = 0.01), 7 (q < 0.001) and 23 (q = 0.001) (Fig. 8B). Similarly, Clostridium was higher in Het mice on days 4 (q = 0.04), 7 (q = 0.01) and 15 (q < 0.001) than ob/ob mice (Fig. 8C). In contrast, Lactococcus was more abundant in ob/ob mice on days 7 (q = 0.01), 15 (q = 0.002) and 35 (q = 0.007) (Fig. 8D).
Obesity was also associated with altered relative abundance of the phylum Actinobacteria over time. Compared to ob/ob mice, Het mice had an increase in the relative abundance of Actinobacteria from days 7 to 35 (Fig. 9A). This leptin-dependent increase in Actinobacteria in Hets was due mostly to Coriobacteriaceae (family), also from days 7 to 35 (Fig. 9B), suggesting this microbial family is influenced by leptin or obesity status during HFD.
A subset of microbial taxa was affected by both E2 and leptin. In particular, Streptococcaceae, Lactococcus and Coprobacillus were decreased, whereas Ruminococcaceae were increased by E2 and leptin. In addition, these two hormones exerted opposing effects on other families, such that E2 decreased, and leptin (in Het controls) increased, abundances of Coriobacteriaceae, Lactobacillaceae and Peptostreptococcaceae.
Discussion
Mounting evidence indicates profound effects of estrogens, leptin and gut microbiota on energy homeostasis10,25,51,52. The present study investigated if estradiol and leptin-mediated energy regulation is associated with temporal changes in gut microbiota of females. Using leptin-deficient ob/ob female mice fed a HFD, we investigated the effects of estradiol and obesity (due to leptin deficiency) on body weight, energy intake, and gut microbiota in the present study. We found that E2 treatment decreases food intake and protects against HFD-induced weight gain in both ob/ob (obese) and Het (lean) females. ob/ob mice had increased HFD intake and greater body weight, compared to the Het controls. The differences in food intake and body weight between the two genotypes are reflected in their changes in gut microbiota. Both E2 treatment and genotype were associated with altered gut microbiota diversity. ob/ob female mice exhibited lower species richness, which supports previous studies that link obesity with a reduced gut microbiota diversity in humans and male mice53–55. Interestingly, E2 treatment was associated with lower species evenness compared to Veh mice. At the phylum level, E2 treatment slowed down a HFD-induced decrease in Bacteroidetes and increases in Firmicutes and Actinobacteria compared to Veh controls. Leptin also was associated with changes in Bacteroidetes and Firmicutes and a profound increase in Actinobacteria. Many taxa were also associated with both leptin and E2. These findings suggest that E2 and leptin can act independently, or interact together, to modulate gut microbiota to mediate energy regulation during HFD intake in females.
We and others have shown that E2 protects female mice from HFD-induced obesity10,11,13,56–59. Estrogens exert protective effects by acting directly on brain10–12,60, pancreas61, liver56,57,59,62, adipose tissue59, and muscle63,64 to regulate energy production and utilization. Moreover, long-term E2 treatment improves glucose tolerance and insulin sensitivity and attenuates lipid synthesis in liver in ob/ob female mice65. The present findings suggest that the modulation of gut microbiota is another mechanism by which E2 mediates energy homeostasis. While E2 was associated with a decrease in the gut microbial evenness in the current study, an increase in microbial diversity has been reported in cycling rats and E2-treated female mice40,66. These differences across studies could be due to fluctuating estrogens and other ovarian hormones in the cycling rats and/or species differences40, and a much higher dose of estradiol (2.5 mg/day, consistent with levels at pregnancy) used in the mice66. Alternatively, while a lower microbial diversity is usually associated with obesity and metabolic disorders67,68, a change in relative abundance without any changes in microbial richness or evenness can also alter microbial homeostasis47.
E2 attenuated a longitudinal shift in the two major phyla Bacteroidetes and Firmicutes, compared to the Veh controls. In particular, the order Bacteroidales, and its families S24-7 and Ruminococcaceae were positively associated with E2 treatment, whereas Allobaculum spp. were negatively associated. Many microbes that belong to S24-7 produce short chain fatty acids (SCFA; fermented products of dietary fibers) that protect against inflammation69,70. In particular, SCFA, including butyrate, maintain a low pH in the gastrointestinal tract and aid in nutrient absorption and pathogen inactivation71. Interestingly, S24-7 is downregulated in inflammatory conditions, including Crohn’s disease, colitis and type I diabetes72–74. While the current findings suggest that E2 provides protection against diet-induced metabolic disorders in females by maintaining healthy levels of microbes belonging to S24-7, it will be important in future studies to selectively manipulate these microbes to test their functions in energy regulation.
In the present study, E2 was associated with changes in Firmicutes, which is generally associated with obesity in rodents and humans75. Studies in males have shown that a high caloric diet profoundly affects Firmicutes36,76,77. Consistent with these findings in males, in the present study, Veh female mice on a HFD gained weight and had an increase in relative abundances of Firmicutes. However, E2-treated mice resisted this HFD-induced increase, suggesting that modulation of these microbes contributes to the regulation of energy homeostasis by estrogens. Firmicutes contain more OTUs that are efficient energy producers than Bacteroidetes, leading to increased calorie absorption and weight gain52. Additionally, some members of Firmicutes promote lipid droplet formation increasing fatty acid absorption and weight gain78. In the current study, E2 was associated with changes in relative abundances of Firmicutes in both Het and ob/ob mice, suggesting that E2 affects this phylum independent of body weight and leptin levels.
The obese leptin-deficient (ob/ob) mice had less diverse microbiota as observed by species richness, suggesting that leptin is required for an enriched microbial ecosystem. In support, obesity is associated with a decreased microbial diversity in humans53. Furthermore, a Danish study on men and women found that higher levels of leptin in obese populations were associated with a lower richness of the gut microbial communities, suggesting that optimum levels of leptin are associated with metabolic health and a more diverse gut microbiota54. Taken together, the present results suggest that changes in leptin levels are associated with disruptions in metabolic and microbial homeostasis.
While ob/ob mice weighed almost twice that of Het mice in the beginning, the percent weight gain was greater in Veh Hets compared to Veh ob/ob mice. Interestingly, the relative abundance of the phylum Actinobacteria was greater in Hets compared to ob/ob mice, primarily due to an increase in its family Coriobacteraceae. Actinobacteria is increased in obese humans and is associated with ulcerative colitis53,68. More than two-thirds of the obesity-related human gut microbes belong to Actinobacteria53,79. In particular, Coriobacteriaceae have been positively associated with increased cholesterol absorption in hamsters and obese humans80,81. A profound increase in the relative abundance of Coriobacteriaceae in Veh Het mice, the group that showed the highest percent weight gain, but not in E2 Hets, suggests this OTU is associated with HFD-induced weight gain.
Estrogens elicit many effects on physiology and behavior by binding to their intracellular and membrane receptors82–85. While estrogen receptor-α (ERα) and ERβ have been implicated in the effects of estrogens on metabolism86–88, ERα appears to be the primary contributor to energy balance86,89–91. ERα knock-out mice exhibit increased visceral adiposity, impaired glucose tolerance and elevated insulin levels91. Furthermore, systemic activation of ERα, but not ERβ, decreases food intake and body weight in female rats90. While the effects of ERα on gut microbiota have not been investigated directly, a study using ERβ knock-out female mice suggests that ERβ influences gut microbiota in a diet-specific manner92. In support of this finding, ERβ is expressed in human and mouse colon epithelium93,94. Alternatively, given that estrogens reduce food intake10, it is also possible that E2 influences gut microbiota by altering nutrition availability.
Estrogens may also act as direct substrates for gut microbiota. For example, microbes with β-glucuronidase and β-glucosidase enzymes, including Lactobacillus, Bifidobacterium, and Clostridium spp., convert inactive estrogens into their active forms through deconjugation95–97. In addition, estrogens may alter metabolism and immune responses through direct actions on gut microbial metabolites. For example, E2 treatment protects against HFD-induced metabolic disorders by blocking the activation of lipopolysaccharides (LPS), the endotoxins produced by gram-negative microbes98–100. Additionally, E2 upregulates intestinal alkaline phosphatase, a protective enzyme in the gut epithelium that functions by attenuating pro-inflammatory signals45. These studies, taken together with the present findings, suggest that estrogens can alter gut microbiota through multiple direct and indirect mechanisms to protect from metabolic disorders.
The mechanisms by which leptin affects gut microbiota are not well understood. Intestinal epithelium expresses leptin receptors (LepR), but ablation of these receptors does not alter gut microbiota or body weight in male mice101. In females, leptin may act in concert with ERß in the gut epithelium93,94. While genotype effects may be primarily dependent on leptin, it is important to note that ob/ob mice are obese and Het mice are lean. Obesity is an independent modulator of gut microbiota. Obese and lean twins exhibit differences in gut microbial diversity53, and the obese phenotype can be transferred to lean recipients through the transplant of gut microbiota from obese donors36,37. Alternately, HFD and obesity directly alter the host transcriptome and epigenome by differentially activating enhancers and promoters in the intestinal epithelium102. In addition, female ob/ob mice have reduced estrogen levels and are sterile, and thus have altered hormone-dependent development26,103. These findings suggest that leptin, acting directly, or indirectly via alteration of obesity status, affects host physiology, including gut microbial homeostasis. In future studies, it will be important to test the effects of leptin administration on the gut microbiota in HFD-fed ovariectomized ob/ob mice.
This study provides further insights into the potential roles of E2 and leptin in HFD-induced obesity and gut microbiota. While the identification of taxa that differ between treatments is the first step, their potential actions on energy metabolism will need to be explored in functional studies that manipulate the microbiota using fecal microbiota transplant, selective administration, or depletion of the target microbes. Given that many metabolic diseases are characterized by disturbances in the gut microbiome, investigating how estradiol and leptin-dependent energy regulation associates with disruption in microbial homeostasis provides a foundation for future functional studies on the role of these hormones and the gut microbiome in metabolic disorders in women.
Materials and Methods
Animals
Seven week-old lean heterozygote (Het) and obese ob/ob (leptin-deficient) mice (24 mice/genotype) were purchased from Jackson Laboratory (Bar Harbor, Maine), kept on a 12:12 light:dark cycle and allowed to acclimate at the Wellesley College animal facility for a week. Het mice, which have a metabolic phenotype similar to wildtype mice as evidenced by normal serum glucose and insulin, body temperature, and energy expenditure19, were used as controls21 to account for genetic background. While phenotypically very similar, Het mice have lower leptin levels than wildtype mice104. All mice were ovariectomized (OVX) and subcutaneously implanted with a silastic capsule containing 17 -Estradiol (E2, #E8875, Sigma; 50 µg in 25 µl of 5% ethanol/sesame oil) or Veh (5% ethanol/sesame oil) resulting in the following four experimental groups: control E2, 2) control Veh, 3) ob/ob E2, and 4) ob/ob Veh. Following surgery, mice were pair-housed with a mouse of the same genotype and treatment.
As part of another study to identify newborn cells, on day 7 mice underwent intracranial surgery for the implantation of an intraventricular cannula attached to a subcutaneous osmotic pump filled with bromodeoxyuridine. The osmotic pumps were removed after 10 days. On day 35, animals received an intraperitoneal injection of leptin (5 mg/kg) 45 minutes prior to being euthanized for assessment of acute leptin response in the brain, unrelated to the present study. While possible, it is unlikely that leptin administration 45 minutes prior to sacrifice would have effects on gut microbiota. All procedures were approved by the Institutional Animal Care and Use Committee of Wellesley College and performed in accordance with National Institutes of Health Animal Care and Use Guidelines.
Food intake and body weight measurements and fecal sample collection
Following OVX (day 0), all mice were maintained on a standard diet for three days (13.5% calories from fat, Purina, #5001) before switching to a HFD (58.4% calories from fat, Teklad, #03584) on day 4 and maintained on a HFD for the remainder of the study. Throughout the experiment, food intake and body weights were recorded every four days from heterozygote control (Het) E2 (n = 13), Het Veh (n = 12), ob/ob E2 (n = 8) and ob/ob Veh (n = 7) mice. Food intake was calculated per cage, from a total of Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6) cages, with each cage containing two mice of the same treatment. For cages in which one mouse died, the amount of food eaten by the remaining mouse was doubled to match with cages containing two mice. Similarly, for microbiota analysis, fecal samples from each cage was counted as n = 1, resulting in Het E2 (n = 6), Het Veh (n = 6), ob/ob E2 (n = 6) and ob/ob Veh (n = 6). Fecal samples were collected on days 4, 7, 15, 23, 31, and 35 and immediately stored at −80 °C.
DNA extraction from fecal samples, 16S rDNA sequencing and bioinformatics processing
DNA was extracted from fecal samples using a MO BIO PowerSoil DNA Isolation Kit (Valencia, CA) with minor adjustments to the manufacturer’s protocol. A 5-minute incubation with the elution buffer before centrifugation was added to increase the DNA yield. The quality and quantity of the DNA samples were measured using Nanodrop (Thermo Scientific, Waltham, MA). The samples were stored at −20 °C until sequencing.
The V3-V4 region of the 16S rDNA was amplified using the following universal 16S rDNA primers: forward 341F (5′-CCTACGGGAGGCAGCAG-3′) and reverse 806R (5′-GGACTACHVGGGTWTCTAAT-3′) with sequence adapters on both primers and sample-specific Golay barcodes on the reverse primer105. The PCR products were quantified by PicoGreen (Invitrogen, Carlsbad, CA) using a plate reader. After quantification, amplicons were pooled in equal concentrations, cleaned up using UltraClean PCR Clean-Up Kit (MO BIO, Carlsbad, CA), and again quantified using the Qubit (Invitrogen, Carlsbad, CA). The pooled samples were then sequenced using paired-end v2 chemistry using Illumina Miseq sequencing technology (Illumina, San Diego, CA) at the Microbiome Core, Mayo Clinic (Rochester, Minnesota).
Paired R1 and R2 sequence reads were then processed via the hybrid-denovo bioinformatics pipeline, which clustered a mixture of good-quality paired-end and single-end reads into operational taxonomic units (OTUs) at 97% similarity level106. OTUs were assigned taxonomy using the RDP classifier trained on the Greengenes database (v13.5)107. A phylogenetic tree based on FastTree algorithm was constructed based on the OTU representative sequences108. Singleton OTUs as well as samples with less than 2,000 reads were removed from downstream analysis.
Statistical analysis
Food intake and body weight data analysis
Repeated measures ANOVA (SPSS, v.24) was performed to examine the effects of treatment and genotype on food intake and body weight over time. After a main effect was confirmed, ANOVA without corrections was performed on measures from each day to identify the days with effects of one or both variables. One–way ANOVAs were then conducted to identify differences between specific groups when effects of treatment or genotype were present. Differences were considered statistically significant at p < 0.05.
16S rDNA sequence analysis
Analyses were first performed on the aggregated data, in which sequence reads from each mouse across all days were aggregated. To study specific longitudinal trends, stratified analyses on individual days were also performed if needed.
Diversity analysis
Both α-diversity and β-diversity were analyzed on the rarefied OTU data. α-diversity (within-sample diversity) reflects species richness and evenness within the microbial populations. Two representative α-diversity measures were investigated: the observed number of OTUs, an index of the species richness, and the Pielou’s evenness index109. A multiple linear regression model (“lm” function in R) was used to test the association between α-diversity (outcome) and treatment/genotype (covariates, both included in the model).
β-diversity (between-sample diversity) reflects the shared diversity between bacterial populations in terms of ecological distances; pair-wise distance measure allows quantification of the overall compositional difference between samples. Different β-diversity measures provide distinctive views of the community structure. The β-diversity measures were calculated using Bray-Curtis dissimilarity, which measures differences in bacterial composition based on taxa abundances (“vegdist” function in the R “vegan” package, v2.4-3). To test the association between β-diversity measures and treatment or genotype, we used PERMANOVA (999 permutations, “adonis” function in the R “vegan” package, v2.4-3) when adjusting the effect of the other covariate. Ordination plots were generated using principal coordinate analysis (PCoA) on the distance matrix (“cmdscale” function in R)110.
Taxa analysis
Differential abundance analyses were performed at the phylum, class, order, family and genus levels. Taxa with prevalence less than 10% or with a maximum proportion less than 0.2% were excluded from analysis to reduce the number of tests. An overdispersed Poisson model was fitted to the taxa counts with treatment and genotype as covariates (“glm” function in R)111. Wald test was used to assess significance. To account for variable sequencing depths, the GMPR size factor was estimated and used as an offset (log scale) in the regression model112. False discovery rate (FDR) control (B-H procedure, ‘p.adjust’ in R) was used to correct for multiple testing at each taxonomical level, and FDR-adjusted p-values (q-values) < 0.1 were considered significant113. The differential taxa were visualized on a cladogram using GraPhlAn114. All statistical analyses were performed in R (v. 3.3.2, R Development Core Team).
Supplementary information
Acknowledgements
This work was funded by NIH R01 DK61935 (M.J.T.) and a Re-Entry award (E.P.B.) from the National Institute of Diabetes and Digestive and Kidney Diseases. The authors thank Dr. Cassandra Pattanayak, Director of the Quantitative Analysis Institute at Wellesley College, for input on statistical analysis.
Author contributions
K.D.A. conducted the study, analyzed data, wrote the manuscript. X.G. conducted the study, analyzed data, wrote the manuscript. E.P.B. designed and conducted the study and analyzed data. J.C. analyzed data and wrote the manuscript. M.J.T. designed the study, analyzed data, and wrote the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-019-56723-1.
References
- 1.Asarian L, Geary N. Modulation of appetite by gonadal steroid hormones. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006;361(1471):1251–1263. doi: 10.1098/rstb.2006.1860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wade GN, Gray JM. Gonadal effects on food intake and adiposity: a metabolic hypothesis. Physiol. Behav. 1979;22(3):583–593. doi: 10.1016/0031-9384(79)90028-3. [DOI] [PubMed] [Google Scholar]
- 3.Clegg DJ. Minireview: the year in review of estrogen regulation of metabolism. Mol. Endocrinol. 2012;26(12):1957–1960. doi: 10.1210/me.2012-1284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.McCarthy MM. Estradiol and the developing brain. Physiol. Rev. 2008;88(1):91–124. doi: 10.1152/physrev.00010.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pfaff D, Waters E, Khan Q, Zhang X, Numan M. Minireview: estrogen receptor-initiated mechanisms causal to mammalian reproductive behaviors. Endocrinology. 2011;152(4):1209–1217. doi: 10.1210/en.2010-1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Blaustein JD. Neuroendocrine regulation of feminine sexual behavior: lessons from rodent models and thoughts about humans. Annu. Rev. Psychol. 2008;59:93–118. doi: 10.1146/annurev.psych.59.103006.093556. [DOI] [PubMed] [Google Scholar]
- 7.Roquer J, Campello AR, Gomis M. Sex differences in first-ever acute stroke. Stroke. 2003;34(7):1581–1585. doi: 10.1161/01.STR.0000078562.82918.F6. [DOI] [PubMed] [Google Scholar]
- 8.Carr MC. The emergence of the metabolic syndrome with menopause. J. Clin. Endocrinol. Metab. 2003;88(6):2404–2411. doi: 10.1210/jc.2003-030242. [DOI] [PubMed] [Google Scholar]
- 9.Guo SS, Zeller C, Chumlea WC, Siervogel RM. Aging, body composition, and lifestyle: the Fels Longitudinal Study. Am. J. Clin. Nutr. 1999;70(3):405–411. doi: 10.1093/ajcn/70.3.405. [DOI] [PubMed] [Google Scholar]
- 10.Bless EP, Reddy T, Acharya KD, Beltz BS, Tetel MJ. Oestradiol and diet modulate energy homeostasis and hypothalamic neurogenesis in the adult female mouse. J. Neuroendocrinol. 2014;26(11):805–816. doi: 10.1111/jne.12206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bless EP, et al. Adult neurogenesis in the female mouse hypothalamus: Estradiol and high-fat diet alter the generation of newborn neurons expressing estrogen receptor alpha. eNeuro. 2016;3(4):1–11. doi: 10.1523/ENEURO.0027-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brown LM, Clegg DJ. Central effects of estradiol in the regulation of food intake, body weight, and adiposity. J. Steroid. Biochem. Mol. Biol. 2010;122(1-3):65–73. doi: 10.1016/j.jsbmb.2009.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xu Y, et al. Distinct hypothalamic neurons mediate estrogenic effects on energy homeostasis and reproduction. Cell. Metab. 2011;14(4):453–465. doi: 10.1016/j.cmet.2011.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Erickson JC, Hollopeter G, Palmiter RD. Attenuation of the obesity syndrome of ob/ob mice by the loss of neuropeptide Y. Science. 1996;274(5293):1704–1707. doi: 10.1126/science.274.5293.1704. [DOI] [PubMed] [Google Scholar]
- 15.Vaisse C, et al. Leptin activation of Stat3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat. Genet. 1996;14(1):95–97. doi: 10.1038/ng0996-95. [DOI] [PubMed] [Google Scholar]
- 16.Sahu A. Leptin signaling in the hypothalamus: emphasis on energy homeostasis and leptin resistance. Front. neuroendocrinology. 2003;24(4):225–253. doi: 10.1016/j.yfrne.2003.10.001. [DOI] [PubMed] [Google Scholar]
- 17.Green ED, et al. The human obese (OB) gene: RNA expression pattern and mapping on the physical, cytogenetic, and genetic maps of chromosome 7. Genome Res. 1995;5(1):5–12. doi: 10.1101/gr.5.1.5. [DOI] [PubMed] [Google Scholar]
- 18.Coleman DL. Diabetes-Obesity Syndromes in Mice. Diabetes. 1982;31:1–6. doi: 10.2337/diab.31.1.S1. [DOI] [PubMed] [Google Scholar]
- 19.Pelleymounter MA, et al. Effects of the obese Gene Product on Body Weight Regulation in ob/ob Mice. Science. 1995;269:540–543. doi: 10.1126/science.7624776. [DOI] [PubMed] [Google Scholar]
- 20.Mistry AM, Andrew G, Swick AG, Romsos DR. Leptin Rapidly Lowers Food Intake and Elevates Metabolic Rates in Lean and ob/ob Mice. J. Nutr. 1997;127(10):2065–2072. doi: 10.1093/jn/127.10.2065. [DOI] [PubMed] [Google Scholar]
- 21.Shimomura K, et al. Is leptin a key factor which develops obesity by ovariectomy? Endocr. J. 2002;49(4):417–423. doi: 10.1507/endocrj.49.417. [DOI] [PubMed] [Google Scholar]
- 22.Steppan CM, Crawford DT, Chidsey-Frink KL, Ke H, Swick AG. Leptin is a potent stimulator of bone growth in ob/ob mice. Regul. Pept. 2000;92(1–3):73–78. doi: 10.1016/S0167-0115(00)00152-X. [DOI] [PubMed] [Google Scholar]
- 23.Clegg DJ, Riedy CA, Smith KA, Benoit SC, Woods SC. Differential sensitivity to central leptin and insulin in male and female rats. Diabetes. 2003;52(3):682–687. doi: 10.2337/diabetes.52.3.682. [DOI] [PubMed] [Google Scholar]
- 24.Clegg DJ, Brown LM, Woods SC, Benoit SC. Gonadal hormones determine sensitivity to central leptin and insulin. Diabetes. 2006;55(4):978–987. doi: 10.2337/diabetes.55.04.06.db05-1339. [DOI] [PubMed] [Google Scholar]
- 25.Gao Q, Horvath TL. Cross-talk between estrogen and leptin signaling in the hypothalamus. Am. J. Physiol. Endocrinol. Metab. 2008;294(5):E817–826. doi: 10.1152/ajpendo.00733.2007. [DOI] [PubMed] [Google Scholar]
- 26.Chehab FF, Lim ME, Lu R. Correction of the sterility defect in homozygous obese female mice by treatment with the human recombinant leptin. Nat. Genet. 1996;12(3):318–320. doi: 10.1038/ng0396-318. [DOI] [PubMed] [Google Scholar]
- 27.Ainslie DA, et al. Estrogen deficiency causes central leptin insensitivity and increased hypothalamic neuropeptide Y. Int. J. Obes. Relat. Metab. Disord. 2001;25(11):1680–1688. doi: 10.1038/sj.ijo.0801806. [DOI] [PubMed] [Google Scholar]
- 28.Soto, M. et al. Gut microbiota modulate neurobehavior through changes in brain insulin sensitivity and metabolism. Mol. Psychiatry (2018). [DOI] [PMC free article] [PubMed]
- 29.Ley RE, et al. Evolution of mammals and their gut microbes. Science. 2008;320(5883):1647–1651. doi: 10.1126/science.1155725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Round JL, Mazmanian SK. The gut microbiome shapes intestinal immune responses during health and disease. Nat. Rev. Immunology. 2009;9(5):313–323. doi: 10.1038/nri2515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kang, C. et al. Gut Microbiota Mediates the Protective Effects of Dietary Capsaicin against Chronic Low-Grade Inflammation and Associated Obesity Induced by High-Fat Diet. mBio. 8(3) (2017). [DOI] [PMC free article] [PubMed]
- 32.Clarke G, et al. Minireview: Gut microbiota: the neglected endocrine organ. Mol. Endocrinol. 2014;28(8):1221–1238. doi: 10.1210/me.2014-1108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Markle JG, et al. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science. 2013;339(6123):1084–1088. doi: 10.1126/science.1233521. [DOI] [PubMed] [Google Scholar]
- 34.Tetel, M. J., de Vries, G. J., Melcangi, R. C., Panzica, G. & O’Mahony, S. M. Steroids, Stress, and the Gut Microbiome-Brain Axis. J. Neuroendocrinol (2017). [DOI] [PMC free article] [PubMed]
- 35.Cani PD. Human gut microbiome: hopes, threats and promises. Gut. 2018;67(9):1716–1725. doi: 10.1136/gutjnl-2018-316723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Turnbaugh PJ, et al. The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009;1:6ra14. doi: 10.1126/scitranslmed.3000322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ridaura VK, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013;341(6150):1241214. doi: 10.1126/science.1241214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ellekilde M, et al. Transfer of gut microbiota from lean and obese mice to antibiotic-treated mice. Sci. Rep. 2014;4:5922. doi: 10.1038/srep05922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cho I, et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature. 2012;488(7413):621–626. doi: 10.1038/nature11400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Moreno-Indias I, et al. Neonatal Androgen Exposure Causes Persistent Gut Microbiota Dysbiosis Related to Metabolic Disease in Adult Female Rats. Endocrinology. 2016;157(12):4888–4898. doi: 10.1210/en.2016-1317. [DOI] [PubMed] [Google Scholar]
- 41.Org, E. et al. Sex differences and hormonal effects on gut microbiota composition in mice. Gut. microbes 0 (2016). [DOI] [PMC free article] [PubMed]
- 42.Foster JA, McVey Neufeld KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36(5):305–312. doi: 10.1016/j.tins.2013.01.005. [DOI] [PubMed] [Google Scholar]
- 43.Jasarevic E, Morrison KE, Bale TL. Sex differences in the gut microbiome-brain axis across the lifespan. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016;371(1688):20150122. doi: 10.1098/rstb.2015.0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mueller S, et al. Differences in fecal microbiota in different European study populations in relation to age, gender, and country: a cross-sectional study. Appl. Env. Microbiol. 2006;72(2):1027–1033. doi: 10.1128/AEM.72.2.1027-1033.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kaliannan K, et al. Estrogen-mediated gut microbiome alterations influence sexual dimorphism in metabolic syndrome in mice. Microbiome. 2018;6(1):205. doi: 10.1186/s40168-018-0587-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yurkovetskiy L, et al. Gender bias in autoimmunity is influenced by microbiota. Immunity. 2013;39(2):400–412. doi: 10.1016/j.immuni.2013.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ley RE, et al. Obesity alters gut microbial ecology. Proc. Natl Acad. Sci. USA. 2005;102(31):11070–11075. doi: 10.1073/pnas.0504978102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Murphy EF, et al. Composition and energy harvesting capacity of the gut microbiota: relationship to diet, obesity and time in mouse models. Gut. 2010;59(12):1635–1642. doi: 10.1136/gut.2010.215665. [DOI] [PubMed] [Google Scholar]
- 49.Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006;444(7122):1022–1023. doi: 10.1038/4441022a. [DOI] [PubMed] [Google Scholar]
- 50.Yang M, et al. Gut Microbiota Composition and Structure of the Ob/Ob and Db/Db Mice. Int. J. Endocrinol. 2019;2019:1394097. doi: 10.1155/2019/1394097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Blaustein JD, Wade GN. Ovarian influences on the meal patterns of female rats. Physiol. Behav. 1976;17(2):201–208. doi: 10.1016/0031-9384(76)90064-0. [DOI] [PubMed] [Google Scholar]
- 52.Turnbaugh PJ, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–1031. doi: 10.1038/nature05414. [DOI] [PubMed] [Google Scholar]
- 53.Turnbaugh PJ, et al. A core gut microbiome in obese and lean twins. Nature. 2009;457(7228):480–484. doi: 10.1038/nature07540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Le Chatelier E, et al. Richness of human gut microbiome correlates with metabolic markers. Nature. 2013;500(7464):541–546. doi: 10.1038/nature12506. [DOI] [PubMed] [Google Scholar]
- 55.Ravussin Y, et al. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obes. (Silver Spring, Md.) 2012;20(4):738–747. doi: 10.1038/oby.2011.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Camporez JP, et al. Cellular mechanism by which estradiol protects female ovariectomized mice from high-fat diet-induced hepatic and muscle insulin resistance. Endocrinology. 2013;154(3):1021–1028. doi: 10.1210/en.2012-1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Riant E, et al. Estrogens protect against high-fat diet-induced insulin resistance and glucose intolerance in mice. Endocrinology. 2009;150(5):2109–2117. doi: 10.1210/en.2008-0971. [DOI] [PubMed] [Google Scholar]
- 58.Mamounis, K. J., Hernandez, M. R., Margolies, N., Yasrebi, A. & Roepke, T. A. Interaction of 17beta-estradiol and dietary fatty acids on energy and glucose homeostasis in female mice. Nutr. Neurosci .1–14 (2017). [DOI] [PMC free article] [PubMed]
- 59.Bryzgalova G, et al. Mechanisms of antidiabetogenic and body weight-lowering effects of estrogen in high-fat diet-fed mice. Am. J. Physiol. Endocrinol. Metab. 2008;295(4):E904–912. doi: 10.1152/ajpendo.90248.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Gao Q, Horvath TL. Neurobiology of feeding and energy expenditure. Annu. Rev. Neurosci. 2007;30:367–398. doi: 10.1146/annurev.neuro.30.051606.094324. [DOI] [PubMed] [Google Scholar]
- 61.Tiano JP, Mauvais-Jarvis F. Molecular mechanisms of estrogen receptors’ suppression of lipogenesis in pancreatic beta-cells. Endocrinology. 2012;153(7):2997–3005. doi: 10.1210/en.2011-1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yasrebi A, Rivera JA, Krumm EA, Yang JA, Roepke TA. Activation of Estrogen Response Element-Independent ERalpha Signaling Protects Female Mice From Diet-Induced Obesity. Endocrinology. 2017;158(2):319–334. doi: 10.1210/en.2016-1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ribas, V. et al. Skeletal muscle action of estrogen receptor a is critical for the maintenance of mitochondrial function andmetabolic homeostasis in females. Sci. Transl. Med (2016). [DOI] [PMC free article] [PubMed]
- 64.Campbell, S. E., Mehan, K. A., Tunstall, R. J., Febbraio, M. A. & Cameron-Smith, D. 17beta-estradiol upregulates the expression of peroxisome proliferator-activated receptor and lipid oxidative genes in skeletal muscle. J. Mol. Endocrinol (2003). [DOI] [PubMed]
- 65.Gao H, et al. Long-term administration of estradiol decreases expression of hepatic lipogenic genes and improves insulin sensitivity in ob/ob mice: a possible mechanism is through direct regulation of signal transducer and activator of transcription 3. Mol. Endocrinol. 2006;20(6):1287–1299. doi: 10.1210/me.2006-0012. [DOI] [PubMed] [Google Scholar]
- 66.Benedek G, et al. Estrogen protection against EAE modulates the microbiota and mucosal-associated regulatory cells. J. Neuroimmunol. 2017;310:51–59. doi: 10.1016/j.jneuroim.2017.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature. 2012;489(7415):220–230. doi: 10.1038/nature11550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Peters BA, et al. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018;8(1):9749. doi: 10.1038/s41598-018-28126-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ormerod KL, et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome. 2016;4(1):36. doi: 10.1186/s40168-016-0181-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ríos-Covián, D. et al. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front Microbiol. 7 (2016). [DOI] [PMC free article] [PubMed]
- 71.Macfarlane GT, Macfarlane S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012;95(1):50–60. doi: 10.5740/jaoacint.SGE_Macfarlane. [DOI] [PubMed] [Google Scholar]
- 72.Krych L, Nielsen DS, Hansen AK, Hansen CH. Gut microbial markers are associated with diabetes onset, regulatory imbalance, and IFN-gamma level in NOD mice. Gut microbes. 2015;6(2):101–109. doi: 10.1080/19490976.2015.1011876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Rooks MG, et al. Gut microbiome composition and function in experimental colitis during active disease and treatment-induced remission. ISME J. 2014;8(7):1403–1417. doi: 10.1038/ismej.2014.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Morgan XC, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012;13(9):R79. doi: 10.1186/gb-2012-13-9-r79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bäckhed F, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl Acad. Sci. USA. 2004;101(44):15718–15723. doi: 10.1073/pnas.0407076101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hildebrandt MA, et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology. 2009;137(5):1716–1724 e1711-1712. doi: 10.1053/j.gastro.2009.08.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhang C, et al. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 2012;6(10):1848–1857. doi: 10.1038/ismej.2012.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Semova I, et al. Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe. 2012;12(3):277–288. doi: 10.1016/j.chom.2012.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Lepage P, et al. Twin study indicates loss of interaction between microbiota and mucosa of patients with ulcerative colitis. Gastroenterology. 2011;141(1):227–236. doi: 10.1053/j.gastro.2011.04.011. [DOI] [PubMed] [Google Scholar]
- 80.Martínez, I. et al. Diet-Induced Alterations of Host Cholesterol Metabolism Are Likely To Affect the Gut Microbiota Composition in Hamsters. Appl. Environ. Microbiol (2013). [DOI] [PMC free article] [PubMed]
- 81.Zhang H, et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl Acad. Sci. USA. 2009;106:2365–2370. doi: 10.1073/pnas.0812600106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Jensen EV, et al. A two-step mechanism for the interaction of estradiol with rat uterus. Proc. Natl Acad. Sci. USA. 1968;59(2):632–638. doi: 10.1073/pnas.59.2.632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl Acad. Sci. USA. 1996;93(12):5925–5930. doi: 10.1073/pnas.93.12.5925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tetel MJ, Pfaff DW. Contributions of estrogen receptor-alpha and estrogen receptor-ss to the regulation of behavior. Biochim. Biophys. Acta. 2010;1800(10):1084–1089. doi: 10.1016/j.bbagen.2010.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mani SK, Oyola MG. Progesterone signaling mechanisms in brain and behavior. Front. Endocrinol. (Lausanne) 2012;3:7. doi: 10.3389/fendo.2012.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Frank A, Brown LM, Clegg DJ. The role of hypothalamic estrogen receptors in metabolic regulation. Front. Neuroendocrinol. 2014;35(4):550–557. doi: 10.1016/j.yfrne.2014.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Qiu J, et al. A G-protein-coupled estrogen receptor is involved in hypothalamic control of energy homeostasis. J. Neurosci. 2006;26(21):5649–5655. doi: 10.1523/JNEUROSCI.0327-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Park CJ, et al. Genetic rescue of nonclassical ERalpha signaling normalizes energy balance in obese Eralpha-null mutant mice. J. Clin. Invest. 2011;121(2):604–612. doi: 10.1172/JCI41702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Santollo J, Katzenellenbogen BS, Katzenellenbogen JA, Eckel LA. Activation of ERalpha is necessary for estradiol’s anorexigenic effect in female rats. Horm. Behav. 2010;58(5):872–877. doi: 10.1016/j.yhbeh.2010.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Santollo J, Wiley MD, Eckel LA. Acute activation of ER alpha decreases food intake, meal size, and body weight in ovariectomized rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007;293(6):R2194–2201. doi: 10.1152/ajpregu.00385.2007. [DOI] [PubMed] [Google Scholar]
- 91.Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc. Natl Acad. Sci. USA. 2000;97(23):12729–12734. doi: 10.1073/pnas.97.23.12729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Menon R, et al. Diet complexity and estrogen receptor beta status affect the composition of the murine intestinal microbiota. Appl. Env. Microbiol. 2013;79(18):5763–5773. doi: 10.1128/AEM.01182-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Enmark E, et al. Human estrogen receptor beta-gene structure, chromosomal localization, and expression pattern. J. Clin. Endocrinol. Metab. 1997;82(12):4258–4265. doi: 10.1210/jcem.82.12.4470. [DOI] [PubMed] [Google Scholar]
- 94.Campbell-Thompson, M., Lynch, I. J., Bhardwaj, B. Expression of estrogen receptor (ER) subtypes and ERbeta isoforms in colon cancer. Cancer Res (2011). [PubMed]
- 95.McIntosh FM, et al. Phylogenetic distribution of genes encoding beta-glucuronidase activity in human colonic bacteria and the impact of diet on faecal glycosidase activities. Env. Microbiol. 2012;14(8):1876–1887. doi: 10.1111/j.1462-2920.2012.02711.x. [DOI] [PubMed] [Google Scholar]
- 96.Kwa, M., Plottel, C. S., Blaser, M. J. & Adams, S. The Intestinal Microbiome and Estrogen Receptor-Positive Female Breast Cancer. J. Natl. Cancer Inst. 108(8) (2016). [DOI] [PMC free article] [PubMed]
- 97.Flores R, et al. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: a cross-sectional study. J. Transl. Med. 2012;10:253. doi: 10.1186/1479-5876-10-253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Cani PD, et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008;57(6):1470–1481. doi: 10.2337/db07-1403. [DOI] [PubMed] [Google Scholar]
- 99.Cani PD, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007;56(7):1761–1772. doi: 10.2337/db06-1491. [DOI] [PubMed] [Google Scholar]
- 100.Blasco-Baque V, et al. High-fat diet induces periodontitis in mice through lipopolysaccharides (LPS) receptor signaling: protective action of estrogens. PLoS One. 2012;7(11):e48220. doi: 10.1371/journal.pone.0048220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Rajala MW, et al. Leptin acts independently of food intake to modulate gut microbial composition in male mice. Endocrinology. 2014;155(3):748–757. doi: 10.1210/en.2013-1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Qin Y, et al. An obesity-associated gut microbiome reprograms the intestinal epigenome and leads to altered colonic gene expression. Genome Biol. 2018;19(1):7. doi: 10.1186/s13059-018-1389-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ng KY, Yong J, Chakraborty TR. Estrous cycle in ob/ob and ovariectomized female mice and its relation with estrogen and leptin. Physiol. Behav. 2010;99(1):125–130. doi: 10.1016/j.physbeh.2009.11.003. [DOI] [PubMed] [Google Scholar]
- 104.Chung, W. K. et al. Heterozygosity for Lepob or Leprdb affects body composition and leptin homeostasis in adult mice. Am. J. Physiol (1998). [DOI] [PubMed]
- 105.Caporaso JG, et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA. 2011;108(Suppl 1):4516–4522. doi: 10.1073/pnas.1000080107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Chen X, et al. Hybrid-denovo: a de novo OTU-picking pipeline integrating single-end and paired-end 16S sequence tags. GigaScience. 2018;7(3):1–7. doi: 10.1093/gigascience/gix129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Cole JR, et al. Ribosomal Database Project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014;42(Database issue):D633–642. doi: 10.1093/nar/gkt1244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Price MN, Dehal PS, Arkin AP. FastTree: computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009;26(7):1641–1650. doi: 10.1093/molbev/msp077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Pielou EC. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966;13:131–144. doi: 10.1016/0022-5193(66)90013-0. [DOI] [Google Scholar]
- 110.McArdle BH, Anderson MJ. Fitting multivariate models to community data: a comment on distance‐based redundancy analysis. Ecology. 2001;82(1):290–297. doi: 10.1890/0012-9658(2001)082[0290:FMMTCD]2.0.CO;2. [DOI] [Google Scholar]
- 111.Chen J, et al. Altered gut microbiota in female mice with persistent low body weights following removal of post-weaning chronic dietary restriction. Genome Med. 2016;8(1):103. doi: 10.1186/s13073-016-0357-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Chen L, et al. GMPR: A robust normalization method for zero-inflated count data with application to microbiome sequencing data. PeerJ. 2018;6:e4600. doi: 10.7717/peerj.4600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to MultipleTesting. J. R. Stat. Soc. B (Methodol.) 1995;57:289–300. [Google Scholar]
- 114.Asnicar F, Weingart G, Tickle TL, Huttenhower C, Segata N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ. 2015;3:e1029. doi: 10.7717/peerj.1029. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.