

Respiratory tract infections due to community-acquired respiratory viruses including respiratory syncytial virus, Influenza, parainfluenza virus 3 and human metapneumovirus are detected in up to 40% of allogeneic hematopoietic stem cell transplant recipients in whom they cause severe symptoms including pneumonia and bronchiolitis and can be fatal. Given the lack of effective antivirals and the data from our group demonstrating that adoptively transferred ex vivo-expanded virus-specific T cells can be clinically beneficial for the treatment of both latent (Epstein-Barr virus, cytomegalovirus, BK virus, human herpesvirus 6) and lytic (adenovirus) viruses, we investigated the potential for extending this immunotherapeutic approach to respiratory viruses. We now describe a system that rapidly generates a single preparation of polyclonal (CD4+ and CD8+) virus-specific T cells reactive against 12 antigens derived from four viruses (respiratory syncytial virus, Influenza, parainfluenza virus 3 and human metapneumovirus) that commonly cause post-transplant morbidity and mortality. With a single in vitro stimulation we consistently generated Th1-polarized T-cell lines that produced multiple effector cytokines and were selectively reactive against viral-expressing targets, with no evidence of “off target” auto- or allo-reactivity. Finally, we demonstrated the clinical relevance of these virus-specific T cells by measuring responses in transplant recipients who successfully controlled active infections. These results support the clinical importance of T-cell immunity in mediating protective antiviral effects against community-acquired respiratory viruses and demonstrate the feasibility of utilizing a broad-spectrum immunotherapeutic in immunocompromised patients with uncontrolled infections.
Introduction
Acute upper and lower respiratory tract infections (RTI) due to community-acquired respiratory viruses (CARV) including respiratory syncytial virus (RSV), influenza, parainfluenza virus (PIV) and human metapneumovirus (hMPV) are a major public health problem.1 For example, RSV-induced bronchiolitis is the most common reason for hospital admission in children under one year of age,2-4 while the Center for Disease Control (CDC) estimates that, annually, Influenza accounts for up to 35.6 million illnesses worldwide, between 140,000 and 710,000 hospitalizations, annual costs of approximately $87.1 billion in disease management in the US alone, and between 12,000 and 56,000 deaths.
Thus, CARV are a leading cause of morbidity and mortality worldwide, with individuals whose immune systems are naïve (e.g. young children) or compromised being the most vulnerable. For example, in allogeneic hematopoietic stem cell transplant (HSCT) recipients, the incidence of CARV-related respiratory viral diseases is as high as 40%.5 While most patients initially present with rhinorrhea, cough and fever, in approximately 50% of cases, infections progress to the lower respiratory tract and are characterized by severe symptoms including pneumonia and bronchiolitis and mortality rates of 23-50%.6-9 There are neither approved preventative vaccines nor antiviral drugs for hMPV10 and PIV11 and for Influenza the preventative vaccine is not indicated unless patients are at least six months post-HSCT.12 Aerosolized ribavirin (RBV) has been approved by the US Food and Drug Administration (FDA) for the treatment of RSV, but it is extremely costly (5-day course = $149,756) and logistically difficult to administer, requiring a specialized nebulization device that connects to an aerosol tent surrounding the patient.13-16 Thus, the lack of approved antiviral agents for many clinically problematic CARV, and the high cost and complexity of administering aerosolized RBV, underscores the need for alternative treatment strategies.
Our group has previously demonstrated that the adoptive transfer of in vitro-expanded virus-specific T cells (VST) can safely and effectively prevent and treat infections associated with both latent [Epstein-Barr virus (EBV), cytomegalovirus (CMV), BK virus (BKV), human her-pesvirus 6 (HHV6)] and lytic [adenovirus (AdV)] viruses in allogeneic HSCT recipients.17,18 Given that susceptibility to CARV is associated with underlying cellular immune deficiency,1,5,6 in the current study, we explored the feasibility of extending the therapeutic range of VST therapy to include Influenza, RSV, hMPV and PIV-3.
We here describe a mechanism by which we can rapidly generate a single preparation of polyclonal (CD4+ and CD8+) VST with specificity for 12 immunodominant antigens derived from our four target viruses using Good Manufacturing Practices (GMP)-compliant manufacturing methodologies. The viral proteins used for stimulation were chosen on the basis of both their immunogenicity to T cells and their sequence conservation.19-21 The expanded cells are Th1-polarized, polyfunctional and selectively able to react to and kill, viral antigen-expressing target cells with no activity against non-infected autologous or allogeneic targets, attesting to both their selectivity for viral targets and their safety for clinical use. We anticipate such multi-respiratory virus-targeted cells (multi-R-VST) will provide broad spectrum benefit to immunocompromised individuals with uncontrolled CARV infections.
Methods
Donors and cell lines
Peripheral blood mononuclear cells (PBMC) were obtained from healthy volunteers and HSCT recipients with informed consent using Baylor College of Medicine institutional review board-approved protocols (H-7634, H-7666) and were used to generate phytohemagglutinin (PHA) blasts and multi-R-VST. PHA blasts were generated as previously reported20 and cultured in VST medium [45% RPMI 1640 (HyClone Laboratories, Logan, UT, USA), 45% Click’s medium (Irvine Scientific, Santa Ana, CA, USA), 2 mM GlutaMAX TM-I (Life Technologies, Grand Island, New York, NY, USA), and 10% human AB serum (Valley Biomedical, Winchester, VA, USA)] supplemented with interleukin 2 (IL2) (100 U/mL; NIH, Bethesda, MD, USA), which was replenished every two days.
Multi-respiratory virus-targeted cell generation
Pepmixes
Peripheral blood mononuclear cells were stimulated with peptide libraries (15mers overlapping by 11aa) spanning Influenza A (NP1, MP1), RSV (N, F), hMPV (F, N, M2-1, M) (JPT Peptide Technologies, Berlin, Germany) and PIV-3 antigens (M, HN, N, F) (Genemed Synthesis, San Antonio, TX, USA). Lyophilized pep-mixes were reconstituted in dimethyl sulfoxide (DMSO) (Sigma-Aldrich) and stored at -80°C.
Generation of virus-specific T cells
To generate multi-R-VST, PBMC (2.5x107) were transferred to a G-Rex10 (Wilson Wolf Manufacturing Corporation, St. Paul, MN, USA) with 100 mL of VST medium supplemented with IL7 (20 ng/mL), IL4 (800 U/mL) (R&D Systems, Minneapolis, MN, USA) and pepmixes (2 ng/peptide/mL) and cultured for 10-13 days at 37°C, 5% CO2.
Flow cytometry
Immunophenotyping
Multi-R-VST were surface-stained with monoclonal antibodies to: CD3, CD25, CD28, CD45RO, CD279 (PD-1) [Becton Dickinson (BD), Franklin Lakes, NJ, USA], CD4, CD8, CD16, CD62L, CD69 (Beckman Coulter, Brea, CA, USA) and CD366 (TIM-3) (BioLegend, San Diego, CA, USA). Cells were acquired on a Gallios™ Flow Cytometer and analyzed with Kaluza® Flow Analysis Software (Beckman Coulter). See Online Supplementary Appendix for details.
Intracellular cytokine staining
Multi-R-VST were harvested, resuspended in VST medium (2x106/mL) and 200 μL added per well of a 96-well plate. Cells were incubated overnight with 200 ng of individual test or control (irrelevant non-viral, e.g. SURVIVIN, WT1) pepmixes, along with Brefeldin A (1 μg/mL), monensin (1 μg/mL), CD28 and CD49d (1 μg/mL) (BD). Intracellular cytokine staining (ICS) for IFN© and TNF〈 was performed as described in the Online Supplementary Appendix.
FoxP3 staining
FoxP3 staining was performed using the eBioscience FoxP3 kit (Thermo Fisher Scientific, Waltham, MA, USA), per manufacturers’ instructions and as detailed in the Online Supplementary Appendix.
Functional studies
Enzyme-linked immunospot
Enzyme-linked immunospot (ELIspot) analysis was used to quantitate the frequency of IFN© and Granzyme B-secreting cells. PBMC and multi-R-VST were resuspended at 5x106 and 2x106 cells/mL, respectively, in VST medium and 100 μL of cells added to each ELIspot well. Antigen-specific activity was measured as previously described after direct stimulation (500 ng/peptide/mL) with the individual test or control pepmixes. See the Online Supplementary Appendix for more details.
Multiplex
The multi-R-VST cytokine profile was evaluated using the MIL-LIPLEX High Sensitivity Human Cytokine Panel (Millipore, Billerica, MA, USA), per manufacturer’s instructions (see the Online Supplementary Appendix).
Chromium release assay
A standard 4-hour chromium (Cr51) release assay was used to measure the specific cytolytic activity of multi-R-VST with autologous antigen-loaded PHA blasts as targets (20 ng/pep-mix/1x106 target cells). Effector:Target (E:T) ratios of 40:1, 20:1, 10:1, and 5:1 were used to analyze specific lysis. The percentage of specific lysis was calculated [(experimental release -spontaneous release)/(maximum release - spontaneous release)] x 100. In order to measure the autoreactive and alloreactive potential of multi-R-VST lines, autologous and allogeneic PHA blasts alone were used as targets.
Results
Generation of polyclonal multi-respiratory virus-targeted cells from healthy donors
To investigate the feasibility of generating VST-specific T-cell lines containing sub-populations of cells reactive against Influenza, RSV, hMPV, and PIV-3 we utilized a pool of overlapping peptide libraries spanning immunogenic antigens from each of the target viruses (Influenza – NP1 and MP1; RSV – N and F; hMPV – F, N, M2-1 and M; PIV-3 – M, HN, N and F) to stimulate PBMC before culture in a G-Rex10 in cytokine-supplemented VST medium (Figure 1A). Over 10-13 days we achieved an average 8.5-fold increase in cells (Figure 1B) [increase from 0.25x107 PBMC/cm2 to mean 1.9±0.2x107 cells/cm2 (median: 2.05x107, range: 0.6-2.82x107 cells/cm2; n=12)], which were comprised almost exclusively of CD3+ T cells (96.2±0.6%; mean±SEM), with a mixture of cytotoxic (CD8+; 18.1±1.3%) and helper (CD4+ ; 74.4±1.7%) T cells (Figure 1C), with no evidence of regulatory T-cell outgrowth, as assessed by CD4/CD25/FoxP3+ staining (Online Supplementary Figure S1). Furthermore, the expanded cells displayed a phenotype consistent with effector function and long-term memory as evidenced by upregulation of the activation markers CD25 (50.2±3.8%), CD69 (52.8±6.3%), CD28 (85.8±2%) as well as expression of central (CD45RO+/CD62L+: 61.4±3%) and effector memory markers (CD45RO+/CD62L−: 20.3±2.3%), with minimal PD1 (6.9±1.4%) or Tim3 (13.5±2.3%) surface expression (Figure 1C and D).
Figure 1.

Generation of polyclonal multi-respiratory virus-targeted cells (multi-R-VST) from healthy donors. (A) A schematic representation of the multi-R-VST generation protocol. (B) Fold expansion achieved over a 10-13 day period based on cell counting using trypan blue exclusion (n=12). (C and D) Phenotype of the expanded cells (mean±Standard Error of Mean, n=12) (SEM).
Anti-viral specificity of multi-respiratory virus-targeted cells
To next determine whether the expanded populations were antigen-specific, we performed an IFN© ELIspot assay using each of the individual stimulating antigens as an immunogen. All 12 lines generated proved to be reactive against all of the target viruses (Table 1 and Online Supplementary Figure S2). Figure 2A summarizes the magnitude of activity against each of the stimulating antigens, while Online Supplementary Figure S3 shows the response of our expanded VST to titrated concentrations of viral antigen. Of note, over the 10-13 days in culture we achieved an enrichment in VST of between 14.6±4.3-fold (PIV-3-HN) and 50.4±9.9-fold (RSV-N) (Figure 2B). The precursor frequencies of CARV-reactive T cells within donor PBMC are summarized in Online Supplementary Figures S4 and S5. Taken together these data suggest that respiratory VST reside in the memory pool and can be readily amplified ex vivo using GMP-compliant manufacturing methodologies.
Table 1.
Reactivity of expanded virus-specific T cells lines against individual stimulating antigens.

Figure 2.

Specificity and enrichment of multi-respiratory virus-targeted cells (multi-R-VST). (A) The specificity of virus-reactive T cells within the expanded T-cell lines following exposure to individual stimulating antigens from each of the target viruses. Data are presented as mean± Standard Error of Mean (SEM) SFC/2x105 (n=12). (B) Fold enrichment of specificity [peripheral blood mononuclear cels (PBMC) vs. multi-R-VST; n=12]. (C) IFN© production, as assessed by ICS from CD4 helper (top) and CD8 cytotoxic T cells (bottom) after viral stimulation in one representative donor (dot plots were gated on CD3+ cells). (D) Summary results for nine donors screened (mean±SEM).
To next evaluate whether viral specificity was contained within the CD4+ or CD8+ or both T-cell subsets we performed ICS, gating on CD4+ and CD8+ IFN©-producing cells. Figure 2C shows representative results from one donor with activity against all four viruses detected in both T-cell compartments [(CD4+: Influenza – 5.28%; RSV – 11%; hMPV – 6.57%; PIV-3 – 3.37%), (CD8+: Influenza – 2.26%; RSV – 4.36%; hMPV – 2.69%; PIV-3 – 2.16%)] while Figure 2D shows a summary of results for nine donors screened, confirming that our multi-R-VST are polyclonal and poly-specific.
Functional characterization of multi-respiratory virus-targeted cells
The production of multiple proinflammatory cytokines and expression of effector molecules has been shown to correlate with enhanced cytolytic function and improved in vivo T-cell activity. Hence, we next examined the cytokine profile of our multi-R-VST following antigen exposure. The majority of IFN©-producing cells also produced TNF〈 (see Figure 3A for detailed ICS results from 1 donor, and Figure 3B, for summary results for 9 donors), in addition to GM-CSF, as measured by Luminex array (Figure 3C, left panel) with baseline levels of prototypic Th2/suppressive cytokines (Figure 3C, right panel). Furthermore, upon antigenic stimulation our cells produced the effector molecule Granzyme B, suggesting the cytolytic potential of these expanded cells (Figure 3D, n=9). Taken together, these data demonstrate the Th1-polarized and polyfunctional characteristics of our multi-R-VST.
Figure 3.

Multi-respiratory virus-targeted cells (multi-R-VST) are polyclonal and polyfunctional. (A) Dual IFN© and TNF〈 production from CD3+ T cells as assessed by intracellular cytokine staining (ICS) in one representative donor. (B) Summary of results from nine donors screened [mean±Standard Error of Mean (SEM)]. (C) Cytokine profile of multi-R-VST as measured by multiplex bead array. (D) Assessment of the production of Granzyme B by ELIspot assay. Results are reported as SFC/2x105 input VST (mean±SEM, n=9).
Multi-respiratory virus-targeted cells are cytolytic and kill virus-loaded targets
To investigate the cytolytic potential of these expanded cells in vitro, we co-cultured multi-R-VST with autologous Cr51-labeled PHA blasts, which were loaded with viral pepmixes with unloaded PHA blasts serving as a control. Viral antigen-loaded targets were specifically recognized and lysed by our expanded multi-R-VST (40:1 E:T -Influenza: 13±5%, RSV: 36±8%, hMPV: 26±7%, PIV-3: 22±5%, n=8) (Figure 4A and Online Supplementary Figure S6). Finally, even though these VST had received only a single stimulation, there was no evidence of activity against non-infected autologous targets nor of alloreactivity (graft-versus-host potential) using HLA-mismatched PHA blasts as targets (Figure 4B), an important consideration if these cells are to be administered to allogeneic HSCT recipients.
Figure 4.
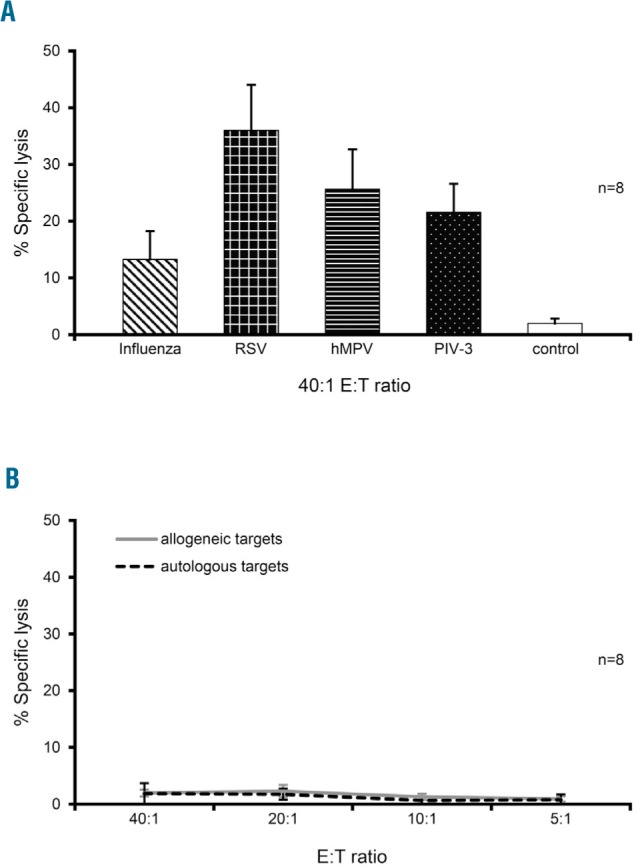
Multi-respiratory virus-targeted cells (multi-R-VST) are exclusively reactive against virus-infected targets. (A) Cytolytic potential of multi-R-VST evaluated by standard 4-hour Cr51 release assay using autologous pepmix-pulsed PHA blasts as targets (E:T 40:1; n=8) with unloaded phytohemagglutinin (PHA) blasts as a control. Results are presented as percentage of specific lysis (mean±SEM). (B) Demonstration that multi-R-VST show no activity against either non-infected autologous or allogeneic PHA blasts, as assessed by Cr51 release assay.
Detection of CARV-specific T cells in hematopoietic stem cell transplant recipients
Finally, to assess the potential clinical relevance of multi-R-VST we investigated whether allogeneic HSCT recipients with active/recent CARV infections exhibited elevated levels of reactive T cells during/following an active viral episode. Figure 5A shows the results of Patient #1, a 64-year old male with acute myeloid leukemia (AML) who received a matched related donor (MRD) transplant with reduced intensity conditioning. The patient developed a severe upper respiratory tract infection (URTI) nine months post-HSCT that was confirmed to be RSV-related by polymerase chain reaction (PCR) analysis. He was not on any immunosuppression at the time of infection but was placed on prednisone the day of infection diagnosis to control pulmonary inflammation. Within four weeks his symptoms resolved without specific antiviral treatment. To assess whether T-cell immunity contributed to viral clearance, we analyzed the circulating frequency of RSV-specific T cells over the course of his infection. Immediately prior to infection this patient exhibited a very weak response to the RSV antigens N and F (6.5 SFC/5x105 PBMC). However, within a month of viral exposure, RSV-specific T cells had expanded in vivo (527 SFC/5x105 PBMC), representing an 81-fold increase in reactive cells (Figure 5A) which declined thereafter, coincident with viral clearance. Of note, the observed RSV-specific responses did not follow the overall increase in lymphocyte/CD4+ counts, thus indicating that T-cell expansion was virus-driven and not due to general immune reconstitution. Similarly, Patient #2, a 23-year old male with acute lymphoblastic leukemia (ALL) who received a matched unrelated donor (MUD) transplant with myeloablative conditioning, and developed a severe RSV-related URTI five months post HSCT while on tapering doses of tacrolimus. His infection symptomatically resolved within one week, coincident with the administration of ribavirin. To investigate whether endogenous immunity also played a role in viral clearance we monitored reactive T-cell numbers over time. Viral clearance was accompanied by an increase in the circulating frequency of RSV-specific T cells (peak 93 SFC/5x105 PBMC) with subsequent return to baseline levels (Figure 5B). The same patient was hospitalized seven months post transplant for a subsequent pneumococcal pneumonia with concurrent detection (by PCR) of hMPV in sputum. His pneumonia was treated with antibiotics with subsequent resolution of disease and viral clearance, coincident with a marked expansion of hMPV-specific T cells (reactive against F, N, M2-1 and M), which increased from 4 SFC to a peak of 70 SFC and subsequent decline to baseline levels (Figure 5C). Again, the observed RSV-and hMPV-specific responses were independent of the overall increase in lymphocyte/CD4+ counts.
Figure 5.

Detection of respiratory virus-targeted (RSV)- and human metapneumovirus (hMPV)-specific T cells in the peripheral blood of hematopoietic stem cell transplant (HSCT) recipients. Peripheral blood mononuclear cells (PBMC) isolated from two HSCT recipients with three infections were tested for specificity against the infecting viruses, using IFN© ELIspot as a readout. (A and B) Results from two patients with RSV-associated upper respiratory tract infection (URTI) which was controlled, coincident with a detectable rise in endogenous RSV-specific T cells. (C) Clearance of an hMPV-lower respiratory tract infection (LRTI) with expansion of endogenous hMPV-specific T cells. ALC: absolute lymphocyte count.
Online Supplementary Figure S7 shows the results of three additional HSCT recipients who developed CARV infections. Patient #3 is a 15-year old female with AML who received a haplo-identical transplant with reduced intensity conditioning, and developed an RSV-induced URTI and LRTI while on tacrolimus five weeks post transplant. The patient was administered ribavirin and the infection resolved within four weeks. We monitored RSV-reactive T cells over time and viral clearance coincided with a striking increase in the frequency of RSV-specific T cells (from 0 to 506 SFC/5x105 PBMC) (Online Supplementary Figure S7A). Similarly, Patient #4, a 10-year old male patient with ALL who received a MUD transplant with myeloablative conditioning, developed a PIV3-related URTI and LRTI one month after HSCT while on tacrolimus. His infection symptomatically resolved within five weeks, coincident with the administration of rib-avirin. To investigate whether endogenous immunity also played a role in viral clearance, we monitored PIV3-reactive T-cell numbers over time. Viral clearance was accompanied by an increase in the circulating frequency of T cells specific for the PIV-3 antigens M, HN, N and F (peak 38 SFC/5x105 PBMC) with subsequent decline (Online Supplementary Figure S7B). Finally, we show Patient #5, a 3-year old male with chronic granulomatous disease who received a MRD transplant with myeloablative conditioning and developed a severe PIV-3-related URTI four months post HSCT while on cyclosporine. The patient received ribavirin but (at last timepoint assessed) continued to exhibit disease symptoms and failed to demonstrate PIV-3-specific T cells (Online Supplementary Figure S7C). Taken together, these data suggest the in vivo rele vance of CARV-specific T cells in the control of viral infections in immunocompromised patients.
Discussion
In the present study, we explored the feasibility of targeting multiple clinically problematic respiratory viruses using ex vivo expanded T cells. We have now shown that we can rapidly generate polyclonal, CD4+ and CD8+ T cells with specificities directed to a total of 12 antigens derived from four seasonal CARV [Influenza, RSV, hMPV and PIV-3] that are responsible for upper and lower respiratory tract infections in the immunocompromised host. These broad spectrum VST, generated using GMP-compliant methodologies, were Th1-polarized, produced multiple effector cytokines upon stimulation, and killed virus-infected targets without auto- or allo-reactivity. Finally, the detection of reactive T-cell populations in the peripheral blood of allogeneic HSCT recipients who successfully cleared active CARV infections suggests the potential for clinical benefit following the adoptive transfer of such multi-R-VST.
Community-acquired respiratory virus-associated acute upper and lower RTI are a major public health problem with young children, the elderly and those with suppressed or compromised immune systems being the most vulnerable.1-3 These infections are associated with symptoms including cough, dyspnea, and wheezing, and dual/multiple co-existing infections are common, with frequencies that may exceed 40% among children under 5-years of age and are associated with increased risk of morbidity and hospitalization.22-26 Among immunocompromised allogeneic HSCT recipients up to 40% experience CARV infections that can range from mild (associated symptoms including rhinorrhea, cough and fever) to severe (bronchiolitis and pneumonia) with associated mortality rates as high as 50% in those with LRTI.5-9 The therapeutic options are limited. For hMPV and PIV-3, there are currently no approved preventative vaccines nor therapeutic antiviral drugs, while the off-label use of the nucleoside analog RBV and the investigational use of DAS-181 (a recombinant sialidase fusion protein) have had limited clinical impact.10,11,27,28 The preventative annual Influenza vaccine is not recommended for allogeneic HSCT recipients until at least six months post transplant (and excluded in recipients of intensive chemotherapy or anti-B-cell antibodies), while neuraminidase inhibitors are not always effective for the treatment of active infections.12 For RSV, aerosolized RBV is FDA-approved for the treatment of severe bronchiolitis in infants and children, and it is also used off-label for the prevention of upper or lower RTI and treatment of RSV pneumonia in HSCT recipients.13,15,16 However, its widespread use is limited by the cumbersome nebulization device and ventilation system required for drug delivery, as well as the considerable associated cost. For example, in 2015, aerosolized RBV cost $29,953 per day, with five days representing a typical treatment course.14 Thus, the lack of approved treatments combined with the high cost of antiviral agents led us to explore the potential for using adoptively-transferred T cells to prevent and/or treat CARV infections in this patient population.
The pivotal role of functional T-cell immunity in mediating viral control of CARV has only recently attracted attention. For example, a retrospective study of 181 HSCT patients with RSV URTI, reported lymphopenia (defined as ALC ≤100/mm3) as a key determinant in identifying patients whose infections would progress to LRTI, while RSV neutralizing antibody levels were not significantly associated with disease progression.29 Furthermore, in a recent retrospective analysis of 154 adult patients with hematologic malignancies with or without HSCT treated for RSV LRTI, lymphopenia was significantly associated with higher mortality rates.30 Both of these studies are suggestive of the importance of cellular immunity in mediating protective immunity in vivo.
Our group has previously demonstrated the feasibility and clinical utility of ex vivo-expanded VST to treat a range of clinically problematic viruses including the latent viruses CMV, EBV, BKV, HHV-6 and AdV.17,31-33 Our initial studies (and those of others)34-37 explored the safety and activity of donor-derived T-cell lines, but more recently we have developed an “off the shelf” universal T-cell platform whereby VST specific for all five viruses (CMV, EBV, BKV, HHV-6, AdV) were prospectively generated and banked, thus ensuring their immediate availability for administration to immunocompromised patients with uncontrolled infections. Indeed, in our recent phase II clinical trial, we administered these partially HLA-matched VST to 38 patients with a total of 45 infections that had proven refractory to conventional antiviral agents and achieved an overall response rate of 92%, with no significant toxicity.18 This precedent of clinical success using adoptively transferred T cells, as well as the absence of effective therapies for a range of CARV, prompted us to explore the potential for extending the therapeutic scope of VST therapy to Influenza, RSV, hMPV and PIV-3 infections post HSCT. In this context, one could consider the option of prophylactic VST administration seasonally to high-risk patients [e.g. young (<5 years) and elderly adults, patients with impaired immune systems]. Alternatively, these cells could be used therapeutically in patients with URTI who have failed conventional antiviral medications in order to prevent LRT progression.
Thus, using our established, GMP-compliant VST manufacturing methodology, we demonstrated the feasibility of generating VST reactive against a spectrum of CARV-derived antigens chosen on the basis of both their immunogenicity to T cells and their sequence conservation [Influenza – NP1 and MP1;20,38,39 RSV – N and F;15,16,20 hMPV – F, N, M2-1 and M;21 PIV3 – M, HN, N and F19] from 12 donors with diverse haplotypes. The expanded cells were polyclonal (CD4+ and CD8+), Th1-polarized and polyfunctional, and were able to lyse viral antigen-expressing targets while sparing non-infected autologous or allogeneic targets, attesting to both their virus specificity and their safety for clinical use. Finally, to assess the clinical significance of these findings we examined the peripheral blood of five allogeneic HSCT recipients with active RSV, hMPV and PIV-3 infections. Four of these patients successfully controlled the viruses within 1-5 weeks, coincident with an amplification of endogenous reactive T cells and subsequent return to baseline levels upon viral clearance, while one patient failed to mount an immune response against the infecting virus and has equally failed to clear the infection to date. These data suggests that the adoptive transfer of ex vivo-expanded cells should be clinically beneficial in patients whose own cellular immunity is lacking.
In conclusion, we have shown that it is feasible to rapidly generate a single preparation of polyclonal multi-respiratory (multi-R)-VST with specificities directed to Influenza, RSV, hMPV and PIV-3 in clinically relevant numbers using GMP-compliant manufacturing method ologies. These data provide the rationale for a future clinical trial of adoptively transferred multi-R-VST for the prevention or treatment of CARV infections in immunocom-promised patients.
Footnotes
Check the online version for the most updated information on this article, online supplements, and information on authorship & disclosures: www.haematologica.org/content/105/1/235
Funding
This work was supported by the Flow Cytometry and Cell and Vector Production shared resources in the Dan L. Duncan Comprehensive Cancer Center (support grant P30 CA125123).
SV was funded in part by an educational grant from the Hellenic Foundation of Hematology. PL is supported by the American Society of Hematology Junior Faculty Scholar grant and the Leukemia Texas Research grant. J. F. V. is supported by a Mentored Research Scholars Grant in Applied and Clinical Research (grant number MRSG-14-197-01-LIB) from the American Cancer Society.
References
- 1.Hodinka RL. Respiratory RNA Viruses. Microbiol Spectr. 2016;4(4) [DOI] [PubMed] [Google Scholar]
- 2.Gill PJ, Richardson SE, Ostrow O, et al. Testing for Respiratory Viruses in Children: To Swab or Not to Swab. JAMA Pediatr. 2017;171(8):798-804. [DOI] [PubMed] [Google Scholar]
- 3.Nair H, Simoes EA, Rudan I, et al. Global and regional burden of hospital admissions for severe acute lower respiratory infections in young children in 2010: a systematic analysis. Lancet. 2013;381(9875):1380-1390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shi T, McAllister DA, O'Brien KL, et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet. 2017; 390(10098):946-958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Paulsen GC, Danziger-Isakov L. Respiratory Viral Infections in Solid Organ and Hematopoietic Stem Cell Transplantation. Clin Chest Med. 2017; 38(4):707-726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Abbas S, Raybould JE, Sastry S, et al. Respiratory viruses in transplant recipients: more than just a cold. Clinical syndromes and infection prevention principles. Int J Infect Dis. 2017;62:86-93. [DOI] [PubMed] [Google Scholar]
- 7.Hutspardol S, Essa M, Richardson S, et al. Significant Transplantation-Related Mortality from Respiratory Virus Infections within the First One Hundred Days in Children after Hematopoietic Stem Cell Transplantation. Biol Blood Marrow Transplant. 2015;21(10):1802-1807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lin R, Liu Q. Diagnosis and treatment of viral diseases in recipients of allogeneic hematopoietic stem cell transplantation. J Hematol Oncol. 2013;6:94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Renaud C, Xie H, Seo S, et al. Mortality rates of human metapneumovirus and respiratory syncytial virus lower respiratory tract infections in hematopoietic cell transplantation recipients. Biol Blood Marrow Transplant. 2013;19(8):1220-1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shah DP, Shah PK, Azzi JM, et al. Human metapneumovirus infections in hematopoi-etic cell transplant recipients and hemato-logic malignancy patients: A systematic review. Cancer Lett. 2016;379(1):100-106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Shah DP, Shah PK, Azzi JM, et al. Parainfluenza virus infections in hematopoietic cell transplant recipients and hematologic malignancy patients: A systematic review. Cancer Lett. 2016; 370(2):358-364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chemaly RF, Shah DP, Boeckh MJ. Management of respiratory viral infections in hematopoietic cell transplant recipients and patients with hematologic malignancies. Clin Infect Dis. 2014;59 Suppl 5:S344-351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Beaird OE, Freifeld A, Ison MG, et al. Current practices for treatment of respiratory syncytial virus and other non-influenza respiratory viruses in high-risk patient populations: a survey of institutions in the Midwestern Respiratory Virus Collaborative. Transpl Infect Dis. 2016; 18(2):210-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chemaly RF, Aitken SL, Wolfe CR, et al. Aerosolized ribavirin: the most expensive drug for pneumonia. Transpl Infect Dis. 2016;18(4):634-636. [DOI] [PubMed] [Google Scholar]
- 15.Griffiths C, Drews SJ, Marchant DJ. Respiratory Syncytial Virus: Infection, Detection, and New Options for Prevention and Treatment. Clin Microbiol Rev. 2017;30(1):277-319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Walsh EE. Respiratory Syncytial Virus Infection: An Illness for All Ages. Clin Chest Med. 2017;38(1):29-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Papadopoulou A, Gerdemann U, Katari UL, et al. Activity of broad-spectrum T cells as treatment for AdV, EBV, CMV, BKV, and HHV6 infections after HSCT. Sci Transl Med. 2014;6(242):242ra83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tzannou I, Papadopoulou A, Naik S, et al. Off-the-Shelf Virus-Specific T Cells to Treat BK Virus, Human Herpesvirus 6, Cytomegalovirus, Epstein-Barr Virus, and Adenovirus Infections After Allogeneic Hematopoietic Stem-Cell Transplantation. J Clin Oncol. 2017;35(31):3547-3557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Aguayo-Hiraldo PI, Arasaratnam RJ, Tzannou I, et al. Characterizing the cellular immune response to parainfluenza virus 3. J Infect Dis. 2017; 216(2):153-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gerdemann U, Keirnan JM, Katari UL, et al. Rapidly generated multivirus-specific cytotoxic T lymphocytes for the prophylaxis and treatment of viral infections. Mol Ther. 2012;20(8):1622-1632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tzannou I, Nicholas SK, Lulla P, et al. Immunologic Profiling of Human Metapneumovirus for the Development of Targeted Immunotherapy. J Infect Dis. 2017;216(6):678-687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Goka E, Vallely P, Mutton K, et al. Influenza A viruses dual and multiple infections with other respiratory viruses and risk of hospitalisation and mortality. Influenza Other Respir Viruses. 2013;7(6):1079-1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Goka EA, Vallely PJ, Mutton KJ, et al. Single, dual and multiple respiratory virus infections and risk of hospitalization and mortality. Epidemiol Infect. 2015;143(1):37-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kouni S, Karakitsos P, Chranioti A, et al. Evaluation of viral co-infections in hospital-ized and non-hospitalized children with respiratory infections using microarrays. Clin Microbiol Infect. 2013;19(8):772-777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lim FJ, de Klerk N, Blyth CC, et al. Systematic review and meta-analysis of respiratory viral coinfections in children. Respirology. 2016;21(4):648-655. [DOI] [PubMed] [Google Scholar]
- 26.Stefanska I, Romanowska M, Donevski S, et al. Co-infections with influenza and other respiratory viruses. Adv Exp Med Biol. 2013;756:291-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Salvatore M, Satlin MJ, Jacobs SE, et al. DAS181 for treatment of parainfluenza Virus infections in hematopoietic stem cell Transplant recipients at a single center. Biol Blood Marrow Transplant. 2016;22(5):965-970. [DOI] [PubMed] [Google Scholar]
- 28.Zenilman JM, Fuchs EJ, Hendrix CW, et al. Phase 1 clinical trials of DAS181, an inhaled sialidase, in healthy adults. Antiviral Res. 2015;123:114-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kim YJ, Guthrie KA, Waghmare A, et al. Respiratory syncytial virus in hematopoietic cell transplant recipients: factors determining progression to lower respiratory tract disease. J Infect Dis. 2014;209(8):1195-1204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vakil E, Sheshadri A, Faiz SA, et al. Risk factors for mortality after respiratory syn-cytial virus lower respiratory tract infection in adults with hematologic malignancies. Transpl Infect Dis. 2018;20(6):e12994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gerdemann U, Katari UL, Papadopoulou A, et al. Safety and clinical efficacy of rapidly-generated trivirus-directed T cells as treatment for adenovirus, EBV, and CMV infections after allogeneic hematopoietic stem cell transplant. Mol Ther. 2013;21(11):2113-2121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Heslop HE, Slobod KS, Pule MA, et al. Long-term outcome of EBV-specific T-cell infusions to prevent or treat EBV-related lymphoproliferative disease in transplant recipients. Blood. 2010;115(5):925-935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Leen AM, Christin A, Myers GD, et al. Cytotoxic T lymphocyte therapy with donor T cells prevents and treats aden-ovirus and Epstein-Barr virus infections after haploidentical and matched unrelated stem cell transplantation. Blood. 2009; 114(19):4283-4292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Doubrovina E, Oflaz-Sozmen B, Prockop SE, et al. Adoptive immunotherapy with unselected or EBV-specific T cells for biopsy-proven EBV+ lymphomas after allogeneic hematopoietic cell transplantation. Blood. 2012;119(11):2644-2656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Feucht J, Opherk K, Lang P, et al. Adoptive T-cell therapy with hexon-specific Th1 cells as a treatment of refractory adenovirus infection after HSCT. Blood. 2015; 125(12):1986-1994. [DOI] [PubMed] [Google Scholar]
- 36.Feuchtinger T, Opherk K, Bethge WA, et al. Adoptive transfer of pp65-specific T cells for the treatment of chemorefractory cytomegalovirus disease or reactivation after haploidentical and matched unrelated stem cell transplantation. Blood. 2010; 116(20):4360-4367. [DOI] [PubMed] [Google Scholar]
- 37.Peggs KS, Verfuerth S, Pizzey A, et al. Cytomegalovirus-specific T cell immunotherapy promotes restoration of durable functional antiviral immunity following allogeneic stem cell transplantation. Clin Infect Dis. 2009;49(12):1851-1860. [DOI] [PubMed] [Google Scholar]
- 38.Chen L, Zanker D, Xiao K, et al. Immunodominant CD4+ T-cell responses to influenza A virus in healthy individuals focus on matrix 1 and nucleoprotein. J Virol. 2014;88(20):11760-11773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Grant EJ, Quinones-Parra SM, Clemens EB, et al. Human influenza viruses and CD8(+) T cell responses. Curr Opin Virol. 2016; 16:132-142. [DOI] [PubMed] [Google Scholar]