

Abstract
Inflammation is a common pathogenesis in many diseases. Salvia miltiorrhiza Bunge (Danshen), a traditional Chinese medicine, has been considered to have good anti-inflammatory effects. In the present study, we investigated the anti-inflammatory effect of diethyl blechnic (DB), a novel compound isolated from Danshen, and its possible mechanisms in lipopolysaccharide (LPS)-induced RAW264.7 macrophages. The results showed that DB can inhibit the LPS-induced pro-inflammatory cytokines release of prostaglandin E2 (PGE2) and mRNA expression of TNF-α, IL-6, and IL-1β. In addition, the results of the flow cytometry assay and the fluorometric intracellular ROS kit assay indicated that DB reduced the generation of ROS in LPS-stimualted RAW264.7 cells. DB reversed the LPS-induced loss of the mitochondrial membrane potential (MMP). Furthermore, DB suppressed the LPS-stimulated increased expression of Toll-like receptor 4 (TLR4), myeloid differential protein-88 (MyD88) and phosphorylation of TAK1, PI3K, and AKT. DB promoted NF-E2-related factor 2 (Nrf2) into the nucleus, increased the expression of heme oxygenase-1 (HO-1) and NAD(P)H dehydrogenase [quinone] 1 (NQO1) and reduced the expression of Keap1. In summary, DB may inhibit LPS-induced inflammation, which mainly occurs through TLR4/MyD88 and oxidative stress signaling pathways in RAW264.7 cells.
Keywords: diethyl blechnic, inflammation, TLR4/MyD88, NF-κB, oxidative stress
1. Introduction
Inflammation is the body’s physiological defense response to different stimuli [1,2]. Moderate inflammation can enhance the body’s autoimmune response, but excessive inflammation activates immune cells, which can damage both healthy and damaged tissues. Stimulated cells proliferate, differentiate, and then secrete inflammatory factors, chemokines, and other factors to promote the occurrence and development of tissue inflammation [3]. Macrophages have an important role in an inflammatory response and are the central cells that initiate the production of inflammatory mediators [4]. LPS is an important inflammation trigger, which induces cytokines to participate in inflammation [5]. So far, the LPS-induced RAW264.7 model has been widely used in the study of inflammation [6].
Screening of anti-inflammatory drugs based on key molecules associated with the inflammatory signaling pathway has become a hot research topic over recent years. NF-κB is a multipotent transcription factor, which participates in transcriptional regulation of various inflammatory mediators [7], including pro-inflammatory cytokines (TNF-a, IL-6, IL-1β), chemokines and immune receptors [8]. The role of TLRs in the upstream signaling molecules of inflammation and related diseases is highly implicated [9]. When induced by LPS, MyD88 is recruited to TLR4, which then activates phosphatidylinositol 3-kinase (PI3K), protein kinase B (AKT), and transforms growth factor-b-activated kinase 1 (TAK1). This process induces metastasis of NF-κB heterodimer into the nucleus, initiates the transcription of inflammation target genes, and then amplifies the inflammation signal [10,11,12]. This data suggest that the inhibition of TLR4/MyD88/PI3K and NF-κB signaling pathways could be an effective management strategy for a number of inflammation diseases.
Keap1/Nrf2/HO-1 is a classic antioxidant pathway [13], which can negatively regulate inflammatory regulators and enzymes, inhibit oxidative stress and overexpression of pro-inflammatory factors [14]. Under normal physiological conditions, NRF2 binds to Keap-1 and is then inhibited. Once oxygen free radicals are produced, the expression of Keap-1 decreases and the free Nrf2 increases. At the same time, the free Nrf2 translocates into the nucleus and binds to the antioxidant elements of antioxidant genes such as NQO1, HO-1, which initiate compensatory expression of genes and eliminate oxygen free radicals and oxidative stress through the biological activities of NQO1 and HO-1 [15]. Therefore, the nuclear translocation of NRF2 and the increased expression of HO-1 can effectively inhibit inflammation reaction.
Natural products have been reported as an important source for anti-inflammatory agent discovery [16]. Our previous study has shown that diethyl blechnic (DB), a novel compound isolated from Danshen, has significant anti-inflammatory properties and the MAPKs pathway and NF-κB pathways participated in its anti-inflammatory process [17,18], yet, its antioxidative effect has not yet been explored. In this study, following the previous study, we continued to examine the protective effect of DB and its molecular mechanisms against LPS-induced inflammation and oxidative stress in vitro.
2. Results
2.1. DB Suppresses LPS-Induced inflammatory Response in RAW264.7 Cells
Diethyl blechnic (Figure 1A), a novel natural product isolated from Salvia miltiorrhiza Bunge, showed no significant cytotoxicity in RAW264.7 cells compared with normal control (Figure 1B). In addition, DB suppressed the LPS-induced inflammatory by inhibiting nitrite level (Figure 1C), iNOS and COX-2 expression (Figure 1D), and NO production (Figure 1E,F) in LPS-induced RAW264.7 cells, which is consistent with our previous study [18].
Figure 1.


DB suppressed the inflammatory response in LPS-stimulated RAW264.7 cells. (A) The chemical structure of DB. (B) The cytotoxicity of DB (0, 5, 10, and 20 µM) analyzed by MTT assay after 24 h treatment, (n = 5). (C) RAW264.7 cells were pretreated with DB (0, 5, 10, and 20 µM) for 1 h and then stimulated with LPS (1 µg/mL) for 18 h. The levels of nitrite were determined by the Griess assay, (n = 5). (D) Cells were pretreated with DB (0, 5, 10, and 20 µM) for 1 h and co-cultured with LPS (1 µg/mL) for another 18 h. Total proteins were collected and the expressions of iNOS and COX-2 were detected by Western blotting, (n = 3). (E) Cells were stimulated with LPS (1 µg/mL) for 8 h with or without DB (20 µM) pretreatment for 1 h; the NO levels were measured by flow cytometry, (n = 3). (F) Statistical analysis of the NO per group. ** p < 0.01, and *** p < 0.001 vs. LPS group.
2.2. DB Suppresses the Release of Pro-Inflammatory Cytokines in LPS-Stimulated RAW264.7 Cells
Next, we investigated the effect of DB on the release of pro-inflammatory cytokines in LPS-induced RAW264.7 cells. The results indicated that DB decreased the levels of pro-inflammatory cytokines PGE2 (Figure 2A), TNF-α and IL-6 (Figure S1A,B). In addition, the LPS-increased mRNA expression levels of IL-1β, IL-6 and TNF-α were significantly reversed following DB pretreatment (Figure 2B). These data suggest that DB may significantly decrease the release and the gene expression of pro-inflammatory cytokines.
Figure 2.

DB suppressed the release and gene expression of pro-inflammatory cytokines PGE2 in LPS-induced RAW264.7 cells. RAW 264.7 cells were pretreated with DB for 1 h and then stimulated with LPS (1 µg/mL) for 24 h. The levels of PGE2 (A) in the culture medium were determined by ELISA kits. RAW264.7 cells pretreated with DB (20 µM) for 1 h were stimulated with LPS (1 µg/mL) for 6 h, after which IL-6, TNF-α, and IL-1β mRNA levels were determined by qRT-PCR, (B) mRNA levels were determined by qRT-PCR, (n = 3). ** p < 0.01, and *** p < 0.001vs LPS group.
2.3. DB Suppresses LPS-Induced NF-κB Nuclear Translocation in RAW264.7 Cells
NF-κB is a group of nucleoprotein factors that regulate the expression of a wide range of genes, which have a pivotal role in LPS-induced inflammatory processes [19,20]. Some studies have reported that LPS may induce the translocation of NF-κB/p65 from the cytoplasm to the nucleus, and then regulate the release of large amounts of inflammatory mediators such as TNF-α, IL-6, IL-1β, NO, and iNOS [20]. In this study, we found that LPS was able to increase the mRNA expression of NF-κB/P65 and IκBα in RAW264.7 cells, which was suppressed by DB (Figure 3A,B).
Figure 3.

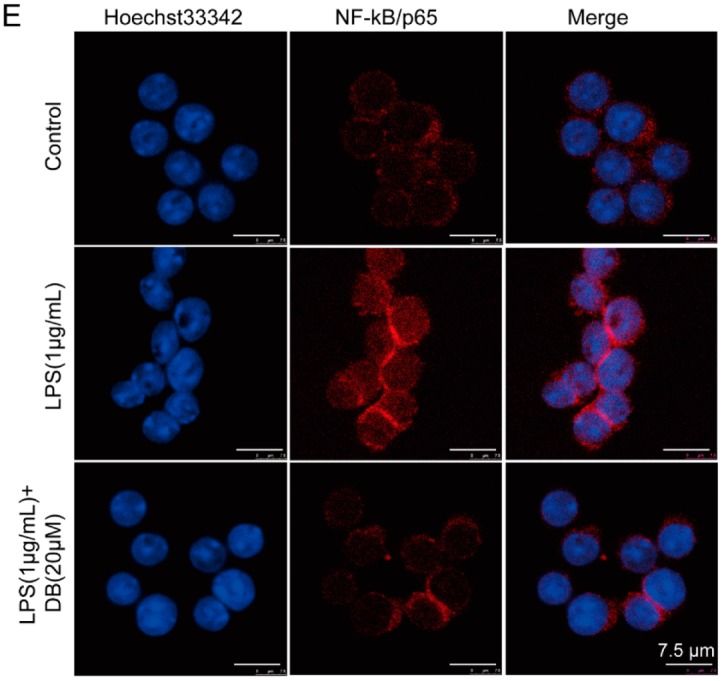
DB suppressed the LPS-induced NF-κB nuclear translocation in RAW264.7 cells. RAW264.7 cells pretreated with DB (20 µM) for 1 h were stimulated with LPS (1 µg/mL) for 6 h, after which NF-κB/p65 (A) and IκBα (B) mRNA levels were determined by qRT-PCR. RAW264.7 cells were pretreated with DB (20 µM) for 1 h and then stimulated with LPS (1 µg/mL) for 2 h, (n = 3). The protein expression of p65 in cytoplasm and nucleus was detected by western blotting, (n = 3) (C,D). The localization of p65 in the cytoplasm and nuclear was measured by immunofluorescence staining. Seven to nine cells were chosen for observation, (n = 3) (E). * p < 0.05, and *** p < 0.001vs LPS group.
In addition, LPS decreased the expression of NF-κB in the cytoplasm and increased expression of NF-κB in the nucleus, which was partially reversed by DB (Figure 3C). Furthermore, in LPS-stimulated RAW264.7 cells, immunofluorescence analysis indicated that DB suppressed the translocation of NF-κB/p65 into the nucleus (Figure 3D).
2.4. DB Suppresses the TLR4-MyD88 Pathway in LPS-Stimulated RAW264.7 Cells
TLR4, a transmembrane receptor located on the surface of many cells, has a pivotal role in inflammatory processes. The stimulation of LPS causes TLR4 to form a dimer, which then modulates the NF-κB signaling pathway, thereby spawning a pathogen-specific innate immune response through the release of pro-inflammatory cytokines [21]. To further study the specific internal mechanisms of DB, the expression of TLR4 and MyD88, phosphorylation of TAK1, PI3K, and AKT were analyzed in LPS-induced RAW264.7 cells. The results showed that LPS treatment activated the TLR4/MyD88 pathway, which was significantly suppressed by DB (Figure 4).
Figure 4.

DB suppressed the LPS-stimulated TLR4-MyD88 pathway. RAW264.7 cells were stimulated with LPS (1 µg/mL) for 2 h to 8 h with or without DB (20 µM) pretreatment for 1 h. The protein expression of TLR4, MyD88, and phosphorylation of TAK1, PI3K, AKT were measured by western blotting, (n = 3) (A). Statistical analyses were performed for each group of TLR4, MyD88 (B) and p-TAK1, p-PI3K, p-AKT (C). n = 3. ** p < 0.01, and *** p < 0.001 vs. LPS group.
2.5. DB Suppresses the Loss of MMP and LPS-Induced ROS Generation
Mitochondrial membrane potential (MMP) is formed by the asymmetric distribution of protons and other ions on both sides of the intima during respiratory oxidation and is important for maintaining the normal physiological function of cells [22,23]. LPS disrupts the stability of MMP, which is not conducive to maintaining normal physiological functions of cells [22,23,24]. The fluorescence of MMP and the fluorescence statistic data were shown in Figure 5A,B, respectively, LPS-induced increase of green fluorescence JC-1 was almost completely reversed to red fluorescence by DB, suggesting that DB restored LPS-induced MMP loss.
Figure 5.
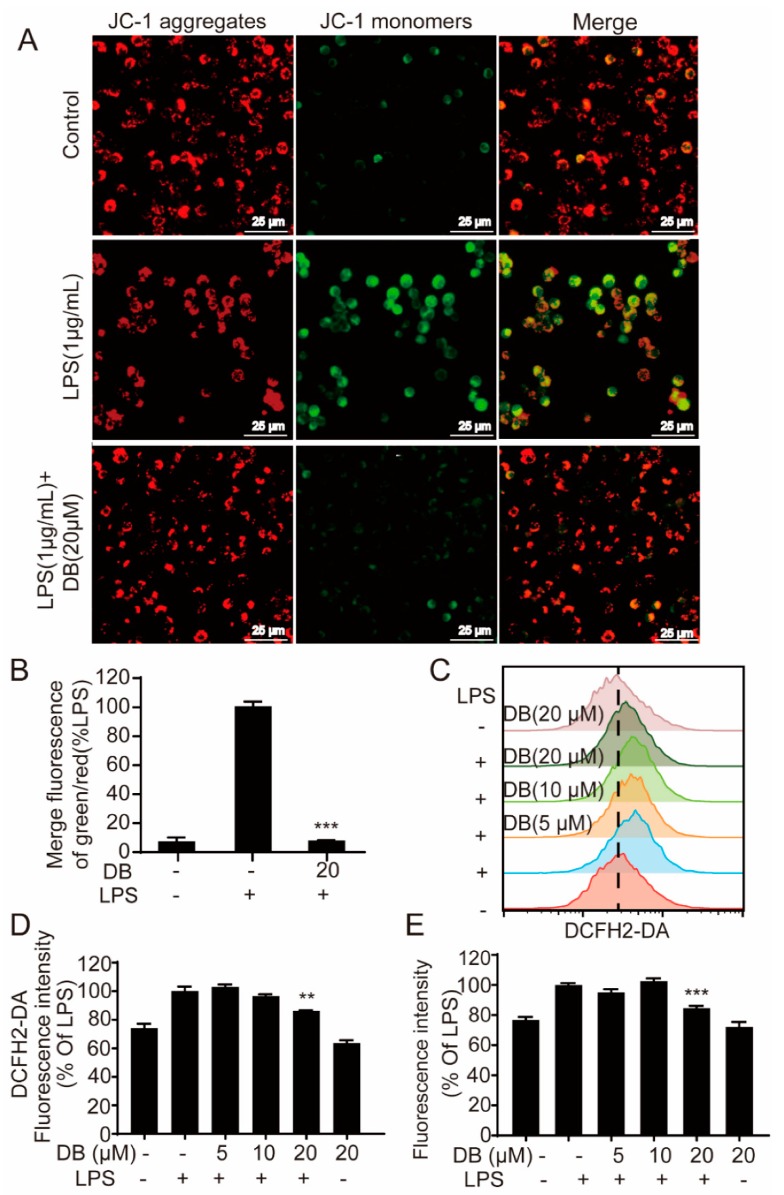
DB suppressed the loss of MMP and ROS generation. (A) Cells were stimulated with LPS (1 µg/mL) for 8 h with or without DB (20 µM) pretreatment for 1 h, the MMP was detected with JC-1 and the images were captured by fluorescence microscopy (Leica DMi8), (n = 3). (B) The statistical analysis of the ratio of green/red fluorescence for the corresponding images. (C) Cells pretreated with indicated concentrations of DB for 1 h were co-cultured with LPS (1 µg/mL) for another 6 h. Then cells labeled with DCFH2-DA (1 µM) for 30 min were investigated by flow cytometry, (n = 3). (D) The statistical analysis of fluorescence intensity of the flow cytometry results. (E) Cells pretreated with indicated concentrations of DB for 1 h were co-cultured with LPS (1 µg/mL) for another 6 h. Add 100 µL/well of Master Reaction Mix into a cell plate and incubate for 1 h in an incubator. Then the fluorescence intensity was determined by a fluorescence microplate reader (λex = 490/λem = 525 nm), (n = 4).
Mitochondria are the main sites for ROS production. In this study, we found that the LPS-stimulation of macrophages resulted in ROS increases within the cells. On the contrary, DB decreased intracellular ROS level detected by the flow cytometry assay (Figure 5C,D), ROS kit (Figure 5E) and fluorescence assay (Figure S1), respectively.
2.6. DB Suppresses the LPS-Stimulated Inflammatory Response Through the Nrf2 Pathway
An oxidative stress pathway is an important mechanism of inflammatory response. Nrf2 has an important role in oxidative stress, which normally binds to its negative regulatory protein Keapl in the cytoplasm during a resting state [25].
When Nrf2 decouples from Keapl and transfer into the nucleus, it will combine with antioxidant response element (ARE) to initiate downstream antioxidant protein gene transcription [26]. Consequently, the expression of HO-1 inhibits the production of inflammatory mediators, while NQO1 constrains the production of oxygen free radicals during oxidative stress [14,27,28]. In our study, the results indicated that DB increased the mRNA expression of Nrf2 and HO-1 in RAW264.7 cells (Figure 6A,B). Besides, LPS significantly enhanced the expression of Keap-1, which was down-regulated by DB. DB pretreatment upregulated the protein expressions of Nrf2, HO-1, and NQO1 (Figure 6C). DB treatment decreased the expression of Nrf2 in the cytoplasm and increased the expression of Nrf2 in the nucleus respectively, meaning that DB pretreatment might promote Nrf2 translocation from the cytoplasm into the nucleus (Figure 6D). Furthermore, using confocal microscopy assay, the immunofluorescence analysis indicated that DB promoted Nrf2 translocation from cytoplasm into nucleus (Figure 6E).
Figure 6.
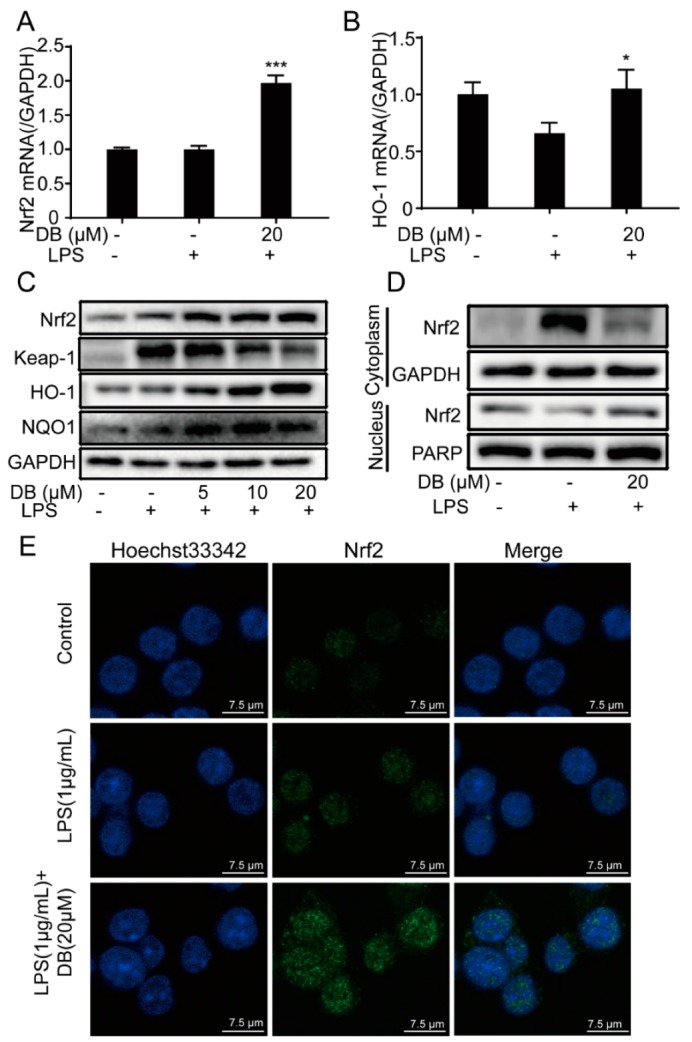
DB suppressed LPS-stimulated inflammatory response through Nrf2 pathways. RAW264.7 cells pretreated with DB (20 µM) for 1 h were stimulated with LPS (1 µg/mL) for 6 h, after which Nrf2 (A) and HO-1 (B) mRNA levels were determined by qRT-PCR, (n = 3). (C) RAW 264.7 cells were stimulated with LPS (1 µg/mL) for 18 h with or without DB (0, 5, 10, and 20 µM) pretreatment for 1 h. The protein expression of Nrf2, Keap1 HO-1, and NQO1 were measured by western blotting, (n = 3). (D) The protein expressions of Nrf2 in cytoplasm and nucleus were detected by western blotting after induced by LPS for 2 h with or without DB (20 µM) pretreatment, (n = 3). (E) The localization of Nrf2 in the cytoplasm and nucleus was measured by immunofluorescence staining, (n = 3). * p < 0.05, and *** p < 0.001 VS. LPS group.
3. Discussion
Inflammation is a defensive immune response to stimulation. Bacteria, viruses, endotoxins and so on can lead to inflammation of human tissues, organs and cells [29]. LPS is the main component of the cell wall of Gram-negative bacteria. Its toxic component is lipid A, which can induce an immune response of mammalian cells and lead to the release of pro-inflammatory factors. Macrophages are the main inflammatory cells in the process of inflammation and related diseases [30]. RAW264.7, a mouse macrophage, induced by LPS, is used as an inflammation model in vitro. LPS can induce cells to produce a variety of inflammatory mediators, cytokines, chemokines and similar, which in turn promote inflammation [31]. Therefore, this model is widely applied for drug screening and anti-inflammation evaluation. In this study, we employed RAW264.7 cells for the investigation of DB’s anti-inflammatory effect. Gao et al. illustrated the study of DB [18]. However, they mainly focused on the isolation and structure elucidation of DB and the part of anti-inflammatory activity. The mechanism need to be further studied.
INOS, which is mainly expressed under the stimulation of inflammation and LPS [32], is closely related to inflammation. COX-2 is an inducible expression enzyme, whose expression is increased in various inflammatory reactions and is related to the severity of inflammation [33]. TNF-α, IL-1β, and IL-6 are pro-inflammatory cytokines and early inflammatory markers of the body produced by LPS-induced macrophages, which can be secreted and released in large quantities under conditions of injury, infection and immune response [34,35,36]. They are the triggers of inflammatory response. In this study, DB significantly suppressed LPS-induced nitrite level in RAW264.7 cells, which was furthermore confirmed by detecting iNOS expression using western blotting and NO generation using cytometry assay, respectively. In addition, TNF-α, IL-1β, and IL-6 cytokines release and gene expression were also inhibited by DB pretreatment with LPS-stimulated RAW264.7 cells. Collectively, DB displayed an obvious anti-inflammatory effect in vitro. Compared with the previous study [18], the dosage and the activity of DB are different. However, DB both displays significant anti-inflammatory activity in two different study. We speculate the main factors to lead to the small discrepancy. The experiments are performed, in two different laboratory, meaning many different experimental conditions, including the source of cells, machines and materials will lead to the small discrepancy. In addition, although DB is relatively stable, around 5 years have passed since we obtained the compound, so the DB may have degraded a little and lost part of its activity. Fortunately, so far, the purity of DB is over 98% (Figure S3), suggesting that DB is relatively stable. DB still has a significant anti-inflammatory activity, suggesting DB indeed is a potential lead-compound as anti-inflammatory agent for further research.
The NF-κB signaling pathway is important in the pathological process of the inflammatory responses [37]. NF-κB is a key transcription factor in inflammatory responses. It exists in the cytoplasm in the form of inactive NF-κB/IκB complex at rest. When cells are stimulated by LPS, IκB is phosphorylated and degraded, and then NF-κB is transferred to the nucleus to regulate the expression of cytokines, chemokines and inflammatory response [38]. Our results suggested that DB significantly inhibited LPS-induced increased mRNA expression of NF-κB/p65 and IκBα and reversed the translocation of NF-κB from the cytoplasm into the nucleus in LPS-stimulated RAW264.7 cells. The results were confirmed by the immunofluorescence analysis and the cytoplasm and nucleus proteins separation kit. As a result, it appears that the NF-κB pathway participated in DB’s anti-inflammatory process.
Toll-like receptor 4 (TLR4) is the special pathogen pattern receptor on the cell surface, which can detect the exogenous homologous ligand LPS and activate the transcription factor NF-κB, leading to the expression of inflammatory cascade effector enzymes such as TNF-α, IL-1β, and IL-6. [39]. TLR4 can activate the downstream of myeloid differentiation factor 88 (MyD88)-dependent signaling pathway, including the PI3K/Akt and NF-κB pathways. Our results indicated that DB suppressed LPS-induced protein expression of TLR4and MyD88, and phosphorylation of TAK1, PI3K, and AKT, suggesting that DB exerts anti-inflammatory effects via TLR4/MyD88-PI3K/AKT pathway.
Reactive oxygen species (ROS) is a by-product of aerobic metabolism. It has strong chemical reactivity and an important role in cell signal transduction and body stability [40]. Once stimulated by LPS, ROS can sharply increase in RAW264.7 cells inducing inflammation through a series of signal transduction pathways via the oxidative stress pathway. In addition, excessive ROS can detriment mitochondria activity by decreasing MMP [41]. In this study, our data suggested that DB also exerts an anti-inflammatory effect by inhibiting ROS generation and MMP loss.
Oxidative stress is caused by the imbalance between the production of reactive oxygen species and the body’s antioxidant defense, which have an important role in the inflammatory response [42,43]. Nuclear factor E2-related factor 2 (NRF2)/heme oxygenase 1 (HO-1) pathway is one of the most important antioxidant stress mechanisms in vivo, which participates in the antioxidant stress response of most tissues and organs [44]. Under normal physiological conditions, NRF2 is stored in the cytoplasm combined with the inhibitory protein Keap1 [45]. When stimulated by oxidative stress, NRF2 is dissociated from Keap1, transferred to the nucleus, and combined with antioxidant response elements (ARE), where it activates the transcription of the HO-1 gene to achieve antioxidant effects [46,47,48,49]. Our results showed that DB blocked LPS-induced translocation of Nrf2 from the nucleus into the cytoplasm, which was further confirmed by immunofluorescence analysis and the nuclear and cytoplasmic protein extraction kit. In conclusion, our data demonstrates that DB exhibits anti-inflammatory and anti-oxidant effects in LPS-stimulated RAW264.7 cells via the TLR4/MyD88-PI3K/AKT NF-κB, and Nrf2/HO-1 signaling pathways.
4. Materials and Methods
4.1. Reagents and Chemicals
The purity of DB (over 98%) isolated from Danshen in our laboratory was determined by high-performance liquid chromatography (HPLC). LPS (Lipopolysaccharides from Escherichia coli O111:B4), Griess reagent (modified-G4410), 2′,7′-dichlorodihydrofluorescein diacetate (DCFH2-DA), methylthiazolyldiphenyl-tetrazolium bromide (MTT), 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolylcarbocyanine iodide (JC-1), MAK-143 intracellular ROS kit and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s modified eagle medium (DMEM), NO detector DAF-FM, fetal bovine serum (FBS) were acquired from Life Technologies/Gibco Laboratories (Grand Island, NY, USA). IL-6 and TNF-α ELISA kits were obtained from Neonbioscience (Shenzhen, China). Antibodies against iNOS (#13120), COX-2 (#4842), NF-κB/p65 (#8242T), Keap-1 (#4678), Nrf2 (#12721), HO-1 (#70081), NQO1,PARP (#9532), TAK1 (#5206), p-TAK1 (#9939), MyD88 (#4283), TLR4 (#14358), p-PI3K (#4228), PI3K (#4249), Akt (#4691), p-Akt (#4060), mTOR (#2983), p-mTOR (#5536),and GAPDH (#5174) were obtained from Cell Signaling (Beverly, MA, USA). Nuclear and Cytoplasmic Protein Extraction kit was obtained from Beyotime (Shanghai, China).
4.2. Qualtiy Control of DB
The purity of DB was determined by HPLC (Accutiy/Arc with 2998, Waters, Milford, MA, USA). An ODS column (CORTECS® C18, 2.7µm, 4.6 × 50 mm) was used. The following mobile phase: acetonitrile/water (50/50) with a flow rate of 1.0 mL/min was adopted. The absorbance wavelength of the detector was set at 220 and 430 nm.
4.3. Cell Culture
RAW264.7 macrophages cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in DMEM supplemented with 10% FBS in a humidified atmosphere containing 5%CO2/95% air at 37 °C.
4.4. MTT Assay
Cells were cultured in 96-well plates at a density of 5 × 104 per well overnight and then treated with DB (0, 5, 10, and 20 µM). After 24 h, MTT dye (5 mg/mL) was added to each well and incubated for another 3 h at 37 °C. After removal of the medium, 100 µL of DMSO was added to each well and properly mixed for another 10 min. The absorbance at 570 nm was determined using a microplate reader.
4.5. Griess Reagent Assay
RAW264.7 cells were seeded in 12-well plates at a density of 1 × 105 per well and cultured overnight. Cells were first pretreated with DB (0, 5, 10, and 20 µM) for 1 h, and then incubated in 1 µg/mL LPS for another 24 h. The medium was collected to determine the nitrite level using the Griess assay.
4.6. Assessment of Cytokine Release
TNF-α and IL-6 were investigated by ELISA following the manufacturer’s instructions. Cells were plated into 24-well plates overnight. Consequently, cells were pretreated with DB (0, 5, 10, and 20 µM) for 1 h, following incubation in DB and LPS (1 µg/mL) for 24 h. The medium was determined for the determination of the release of the cytokine.
4.7. ROS Kit
RAW264.7 cells were cultured in 96-well black plates with a clear bottom at a density of 5 × 104 per well overnight. Cells were then pretreated with DB (0, 5, 10, and 20 µM) for 1 h. Consequently, cells were cultured with or without LPS (1 µg/mL) for another 8 h. Then, MAK-143 intracellular ROS kit (Sigma-AldrichLouis, MO, USA) was used to measure the level of intracellular ROS according to the manufacturer’s instruction. In a brief, add 100 µL/well of Master Reaction Mix into cell plate and incubate for 1 h in an incubator. Then the fluorescence intensity was determined by a fluorescence microplate reader (λex = 490/λem = 525 nm).
4.8. Flow Cytometry Assay
RAW264.7 cells were seeded in 12-well plates at a density of 2 × 105 cells per well and cultured overnight. Subsequently, cells were pretreated with DB (0, 5, 10, and 20 µM) for 1 h, following treatment with or without LPS (1 µg/mL). Different probes were used to detect the corresponding indicators, including NO detector DAF-FM (1 µM, 1 h) and ROS detector DCFH2-DA (1 µM, 30 min). After probes incubation, cells were collected and tested by flow cytometry (Becton-Dickinson, Franklin Lakes, NJ, USA).
4.9. Western Blotting Analysis
RAW264.7 cells were seeded in a dish or 6-well plates and fostered overnight. The next day, cells were treated with DB (0, 5, 10, and 20 µM) and LPS for a certain period of time. Total cell proteins were extracted using RIPA (1% PMSF and 1% cocktail). The cytoplasmic extraction and nuclear protein kit (Beyontime, Shanghai, China) were used to obtain the cytoplasmic and nuclear proteins, following the manufacturer’s protocol. A BCA protein kit (Thermofisher, Waltham, MA, USA) was employed to determine protein concentrations. The denatured proteins were then separated by 8% or 10% SDS-PAGE gels and transferred to PVDF membrane (Millipore, Billerica, MA, USA). After blocking the PVDF membrane with 5% nonfat milk for 1h, the PVDF membrane was incubated with primary antibodies (1:1000) at 4 °C for more than 12 h. After wash with TBST and incubation with secondary antibody (1:5000) for 2 h at room temperature, the membranes were analyzed with ChemiDoc™ MP Imaging System (Bio-Rad, Hercules, CA, USA). GAPDH was used as a housekeeping protein.
4.10. Quantitative Real-Time PCR (qRT-PCR) Assay
RAW264.7 cells were seeded at a density of 2 × 106 cells per well into a dish and cultured overnight. Cells were pretreated with DHT (20 µM) for 1 h, following treatment with LPS (1 µg/mL) for another 2 h. Total RNA was extracted using a Trizol assay kit. Subsequently, RNA (1 µg) was subjected to qRT-PCR using a qPCR master mix kit (Applied Biosystems, PowerSYBR Green PCR Master Mix, 4367659, London, UK). PCR amplification was performed by incorporating SYBR green. The oligonucleotide primers for mouse IL-1β, IL-6, TNF-α, p65, IκBα, HO-1, NRF2, and GAPDH were synthesized by Invitrogen (Nanning, China); the gene sequences were as follows:
| IL-1β-F | GAAAGACGG CACACCCACCCT |
| IL-1β-R | GCTCTGCTTGTGAGGTGCTGATGTA |
| IL-6-F | TCCAGTTGCCTTC TTGGGAC |
| IL-6-R | GTGTAATTAAGCCTCCGACTTG |
| TNF-α-F | TTCTGTCTACTGAACTTCGGGGTGATCGGTCC |
| TNF-α-R | GTATGAGATAGCAAATCGGCTGACGGTGTGGG |
| NF-κB/p65-F | GCACGGATGACAGAGGCGTGTATAAGG |
| NF-κB/p65-R | GGCGGATGATCTCCTTCTCTCTGTCTG |
| IκB-α-F | TGCTGAGGCACTTCTGAG |
| IκB-α-R | CTGTATCCGGGTGCTTGG |
| HO-1-F | TCAGTCCCAAACCTCGCGGT |
| HO-1-R | GCTGTGCAGGTGTTAGCC |
| NRF2-F | AGCAGGACATGGAGCAAGTT |
| NRF2-R | TTCTTTTTCCAGCGAGGAGA |
| GAPDH-F | CATGACCACAGTCCATGCCATCAC |
| GAPDH-R | TGAGGTCCACCACCC TGTTGCTGT |
4.11. Immunofluorescence Assay
RAW264.7 cells were seeded in confocal dishes (SPL, Pocheon, Korea) with a density of 20 × 105 cells per well overnight. Cells were then pretreated with DB (20 µM) for 1 h and LPS-stimulated (1 µg/mL) for 1 h. Consequently, cells were fixed, punched, blocked, and then incubated with NF-κB p65 or NRF2 antibody (1:100) overnight at 4 °C. Finally, cells were incubated with Alexa Fluor 594 secondary antibody for 1 h. Nuclei were revealed by Hoechst 33342 staining. Fluorescence images were collected under a confocal microscope system (Leica, Wetzlar, Germany).
4.12. Fluorescence Assay
RAW264.7 cells were seeded in 96-well plates with a density of 2 × 105 cells per well overnight. Cells were pretreated with DB (20 µM) for 1h and then incubated with or without LPS for another 8 h. Consequently, cells were stained with JC-1 (10 µg/mL) and DCFH2-DA (100 µM) for 30 min. Fluorescence images were captured using a fluorescence microscopy (Leica, DMi8, Wetzlar, Germany).
4.13. Statistical Analysis
Data are presented as means ±SD. All experiments were repeated at least three times. Data were normally distributed and analyzed by one-way-ANOVA by Graph Pad Prism 7 software (Microsoft, Seattle, WA, USA). A P value < 0.05 was considered to be statistically significant.
Supplementary Materials
The following are available online at https://www.mdpi.com/1420-3049/24/24/4502/s1, Figure S1: DB suppressed the release of pro-inflammatory cytokines in LPS-induced RAW264.7 cells. Figure S2: DB suppressed LPS-induced ROS generation in RAW264.7 cells. Figure S3: The chromatographic of DB.
Author Contributions
H.G. and L.H. designed the research. J.H., S.H., X.-X.L., Q.-Q.W. and Y.C. (Yushun Cui) conducted anti-inflammatory experiments. Y.C. (Yangling Chen) isolated DB from Danshen. J.H wrote the manuscript. H.G and S.Y. revised the manuscript. All authors reviewed the manuscript.
Funding
This study was supported by the Young Teachers’ Basic Ability Improvement in Guangxi University project (XP019008), the National Natural Science Foundation of China (NSFC, 81803807), Guangxi Natural Science Foundation (2018JJB140265), Guangxi Science and Technology Base and Talent Special Project (2018AD19034), and the Project of Guangxi Overseas “100 persons plan” High-level Expert.
Conflicts of Interest
The authors declare no competing conflicts of interest.
Footnotes
Sample Availability: Samples of the compounds are available from the authors.
References
- 1.Tsoupras A., Lordan R., Zabetakis I. Inflammation, not Cholesterol, Is a Cause of Chronic Disease. Nutrients. 2018;10:604. doi: 10.3390/nu10050604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Panchal S.K., Brown L. Cholesterol versus Inflammation as Cause of Chronic Diseases. Nutrients. 2019;11:2332. doi: 10.3390/nu11102332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang C.C., Lin H.L., Wey S.P., Jan T.R. Areca-nut extract modulates antigen-specific immunity and augments inflammation in ovalbumin-sensitized mice. Immunopharmacol. Immunotoxicol. 2011;33:315–322. doi: 10.3109/08923973.2010.507208. [DOI] [PubMed] [Google Scholar]
- 4.PJ M., TA W. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011;11:723–737. doi: 10.1038/nri3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Raetz C.R.H., Whitfield C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002;71:635–700. doi: 10.1146/annurev.biochem.71.110601.135414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nakanishi-Matsui M., Yano S., Matsumoto N., Futai M. Lipopolysaccharide induces multinuclear cell from RAW264.7 line with increased phagocytosis activity. Biochem. Biophys. Res. Commun. 2012;425:144–149. doi: 10.1016/j.bbrc.2012.07.050. [DOI] [PubMed] [Google Scholar]
- 7.Doyle S.L., O’Neill L.A. Toll-like receptors: From the discovery of NFκB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006;72:1102–1113. doi: 10.1016/j.bcp.2006.07.010. [DOI] [PubMed] [Google Scholar]
- 8.Dinarello C.A. The IL-1 family of cytokines and receptors in rheumatic diseases. Nat. Rev. Rheumatol. 2019;15:612–632. doi: 10.1038/s41584-019-0277-8. [DOI] [PubMed] [Google Scholar]
- 9.Plociennikowska A., Hromada-Judycka A., Borzecka K., Kwiatkowska K. Co-operation of TLR4 and raft proteins in LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2015;72:557–581. doi: 10.1007/s00018-014-1762-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kong L., Ge B.X. MyD88-independent activation of a novel actin-Cdc42/Rac pathway is required for Toll-like receptor-stimulated phagocytosis. Cell Res. 2008;18:745–755. doi: 10.1038/cr.2008.65. [DOI] [PubMed] [Google Scholar]
- 11.Shim J.H., Xiao C., Paschal A.E., Bailey S.T., Rao P., Hayden M.S., Lee K.Y., Bussey C., Steckel M., Tanaka N., et al. TAK1, but not TAB1 or TAB2, plays an essential role in multiple signaling pathways in vivo. Genes Dev. 2005;19:2668–2681. doi: 10.1101/gad.1360605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Badr G., Al-Sadoon M.K., El-Toni A.M., Daghestani M. Walterinnesia aegyptia venom combined with silica nanoparticles enhances the functioning of normal lymphocytes through PI3K/AKT, NFκB and ERK signaling. Lipids Health Dis. 2012;11:27. doi: 10.1186/1476-511X-11-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Itoh K., Wakabayashi N., Katoh Y., Ishii T., Igarashi K., Engel J.D., Yamamoto M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999;13:76–86. doi: 10.1101/gad.13.1.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Loboda A., Damulewicz M., Pyza E., Jozkowicz A., Dulak J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016;73:3221–3247. doi: 10.1007/s00018-016-2223-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Karam B.S., Chavez-Moreno A., Koh W., Akar J.G., Akar F.G. Oxidative stress and inflammation as central mediators of atrial fibrillation in obesity and diabetes. Cardiovasc. Diabetol. 2017;16:120. doi: 10.1186/s12933-017-0604-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zheng J., Fan R., Wu H., Yao H., Yan Y., Liu J., Ran L., Sun Z., Yi L., Dang L., et al. Directed self-assembly of herbal small molecules into sustained release hydrogels for treating neural inflammation. Nat. Commun. 2019;10:1604. doi: 10.1038/s41467-019-09601-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gao H., Huang L., Ding F., Yang K., Feng Y., Tang H., Xu Q.M., Feng J., Yang S. Simultaneous purification of dihydrotanshinone, tanshinone I, cryptotanshinone, and tanshinone IIA from Salvia miltiorrhiza and their anti-inflammatory activities investigation. Sci. Rep. 2018;8:8460. doi: 10.1038/s41598-018-26828-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gao H., Sun W., Zhao J., Wu X., Lu J.J., Chen X., Xu Q.M., Khan I.A., Yang S. Tanshinones and diethyl blechnics with anti-inflammatory and anti-cancer activities from Salvia miltiorrhiza Bunge (Danshen) Sci. Rep. 2016;6:33720. doi: 10.1038/srep33720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Armaka M., Ospelt C., Pasparakis M., Kollias G. The p55TNFR-IKK2-Ripk3 axis orchestrates arthritis by regulating death and inflammatory pathways in synovial fibroblasts. Nat. Commun. 2018;9:618. doi: 10.1038/s41467-018-02935-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Donato A.J., Black A.D., Jablonski K.L., Gano L.B., Seals D.R. Aging is associated with greater nuclear NF κ B, reduced I κ B α, and increased expression of proinflammatory cytokines in vascular endothelial cells of healthy humans. Aging Cell. 2008;7:805–812. doi: 10.1111/j.1474-9726.2008.00438.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Shah N., Dhar D., El Zahraa Mohammed F., Habtesion A., Davies N.A., Jover-Cobos M., Macnaughtan J., Sharma V., Olde Damink S.W., Mookerjee R.P., et al. Prevention of acute kidney injury in a rodent model of cirrhosis following selective gut decontamination is associated with reduced renal TLR4 expression. J. Hepatol. 2012;56:1047–1053. doi: 10.1016/j.jhep.2011.11.024. [DOI] [PubMed] [Google Scholar]
- 22.Fu C.Y., Chen J., Lu X.Y., Zheng M.Z., Wang L.L., Shen Y.L., Chen Y.Y. Dimethyl fumarate attenuates lipopolysaccharide-induced mitochondrial injury by activating Nrf2 pathway in cardiomyocytes. Life Sci. 2019;235:116863. doi: 10.1016/j.lfs.2019.116863. [DOI] [PubMed] [Google Scholar]
- 23.Fallah M., Mohammadi H., Shaki F., Hosseini-Khah Z., Moloudizargari M., Dashti A., Ziar A., Mohammadpour A., Mirshafa A., Modanloo M., et al. Doxorubicin and liposomal doxorubicin induce senescence by enhancing nuclear factor κ B and mitochondrial membrane potential. Life Sci. 2019;232:116677. doi: 10.1016/j.lfs.2019.116677. [DOI] [PubMed] [Google Scholar]
- 24.Green D.R., Reed J.C. Mitochondria and apoptosis. Science. 1998;281:1309–1312. doi: 10.1126/science.281.5381.1309. [DOI] [PubMed] [Google Scholar]
- 25.Motohashi H., Yamamoto M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004;10:549–557. doi: 10.1016/j.molmed.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 26.Satoh T., McKercher S.R., Lipton S.A. Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic. Biol. Med. 2013;65:645–657. doi: 10.1016/j.freeradbiomed.2013.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang J., Ohta T., Maruyama A., Hosoya T., Nishikawa K., Maher J.A., Shibahara S., Itoh K., Yamamoto M. BRG1 interacts with Nrf2 to selectively mediate HO-1 induction in response to oxidative stress. Mol. Cell. Biol. 2006;26:7942–7952. doi: 10.1128/MCB.00700-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li L., Dong H., Song E., Xu X., Liu L., Song Y. Nrf2/ARE pathway activation, HO-1 and NQ01 induction by polychlorinated biphenyl quinone is associated with reactive oxygen. species and PI3K/AKT signaling. Chem.-Biol. Interact. 2014;209:56–67. doi: 10.1016/j.cbi.2013.12.005. [DOI] [PubMed] [Google Scholar]
- 29.Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454:428–435. doi: 10.1038/nature07201. [DOI] [PubMed] [Google Scholar]
- 30.Mosser D.M., Edwards J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008;8:958–969. doi: 10.1038/nri2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yao Y.D., Shen X.Y., Machado J., Luo J.F., Dai Y., Lio C.K., Yu Y., Xie Y., Luo P., Liu J.X., et al. Nardochinoid B Inhibited the Activation of RAW264.7 Macrophages Stimulated by Lipopolysaccharide through Activating the Nrf2/HO-1 Pathway. Molecules. 2019;24:2482. doi: 10.3390/molecules24132482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Surh Y.J., Chun K.S., Cha H.H., Han S.S., Keum Y.S., Park K.K., Lee S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κ B activation. Mutat. Res. 2001;480:243–268. doi: 10.1016/S0027-5107(01)00183-X. [DOI] [PubMed] [Google Scholar]
- 33.Minghetti L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004;63:901–910. doi: 10.1093/jnen/63.9.901. [DOI] [PubMed] [Google Scholar]
- 34.Zelova H., Hosek J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013;62:641–651. doi: 10.1007/s00011-013-0633-0. [DOI] [PubMed] [Google Scholar]
- 35.Martinon F., Burns K., Tschopp J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell. 2002;10:417–426. doi: 10.1016/S1097-2765(02)00599-3. [DOI] [PubMed] [Google Scholar]
- 36.Nemeth E., Rivera S., Gabayan V., Keller C., Taudorf S., Pedersen B.K., Ganz T. IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. J. Clin. Investig. 2004;113:1271–1276. doi: 10.1172/JCI200420945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Matias M.L., Gomes V.J., Romao-Veiga M., Ribeiro V.R., Nunes P.R., Romagnoli G.G., Peracoli J.C., Peracoli M.T.S. Silibinin Downregulates the NF-κB Pathway and NLRP1/NLRP3 Inflammasomes in Monocytes from Pregnant Women with Preeclampsia. Molecules. 2019;24:1548. doi: 10.3390/molecules24081548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bak M.-J., Hong S.-G., Lee J.-W., Jeong W.-S. Red Ginseng Marc Oil Inhibits iNOS and COX-2 via NF κ B and p38 Pathways in LPS-Stimulated RAW 264.7 Macrophages. Molecules. 2012;17:13769–13786. doi: 10.3390/molecules171213769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yuan R., Huang L., Du L.J., Feng J.F., Li J., Luo Y.Y., Xu Q.M., Yang S.L., Gao H., Feng Y.L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking TLR4 dimerization. Pharmacol. Res. 2019;142:102–114. doi: 10.1016/j.phrs.2019.02.017. [DOI] [PubMed] [Google Scholar]
- 40.Radi R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA. 2018;115:5839–5848. doi: 10.1073/pnas.1804932115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zorov D.B., Juhaszova M., Sollott S.J. MITOCHONDRIAL REACTIVE OXYGEN SPECIES (ROS) AND ROS-INDUCED ROS RELEASE. Physiol. Rev. 2014;94:909–950. doi: 10.1152/physrev.00026.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Reuter S., Gupta S.C., Chaturvedi M.M., Aggarwal B.B. Oxidative stress, inflammation, and cancer How are they linked? Free Radic. Biol. Med. 2010;49:1603–1616. doi: 10.1016/j.freeradbiomed.2010.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li P., Li L., Zhang C., Cheng X., Zhang Y., Guo Y., Long M., Yang S., He J. Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling. Molecules. 2019;24:2421. doi: 10.3390/molecules24132421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hu L., Tian K., Zhang T., Fan C.H., Zhou P., Zeng D., Zhao S., Li L.S., Smith H.S., Li J., et al. Cyanate Induces Oxidative Stress Injury and Abnormal Lipid Metabolism in Liver through Nrf2/HO-1. Molecules. 2019;24:3231. doi: 10.3390/molecules24183231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Francis N., Rao S., Blanchard C., Santhakumar A. Black Sorghum Phenolic Extract Regulates Expression of Genes Associated with Oxidative Stress and Inflammation in Human Endothelial Cells. Molecules. 2019;24:3321. doi: 10.3390/molecules24183321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Yang H.-L., Lin S.-W., Lee C.-C., Lin K.-Y., Liao C.-H., Yang T.-Y., Wang H.-M., Huang H.-C., Wu C.-R., Hseu Y.-C. Induction of Nrf2-mediated genes by Antrodia salmonea inhibits ROS generation and inflammatory effects in lipopolysaccharide-stimulated RAW264.7 macrophages. Food Funct. 2015;6:230–241. doi: 10.1039/C4FO00869C. [DOI] [PubMed] [Google Scholar]
- 47.Fan L., Fan Y., Liu L., Tao W., Shan X., Dong Y., Li L., Zhang S., Wang H. Chelerythrine Attenuates the Inflammation of Lipopolysaccharide-Induced Acute Lung Inflammation Through NF-κB Signaling Pathway Mediated by Nrf2. Front. Pharmacol. 2018;9:1047. doi: 10.3389/fphar.2018.01047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bao Y., Meng X., Liu F., Wang F., Yang J., Wang H., Xie G. Protective effects of osthole against inflammation induced by lipopolysaccharide in BV2 cells. Mol. Med. Rep. 2018;17:4561–4566. doi: 10.3892/mmr.2018.8447. [DOI] [PubMed] [Google Scholar]
- 49.Sawle P., Foresti R., Mann B.E., Johnson T.R., Green C.J., Motterlini R. Carbon monoxide-releasing molecules (CO-RMs) attenuate the inflammatory response elicited by lipopolysaccharide in RAW264.7 murine macrophages. Br. J. Pharmacol. 2005;145:800–810. doi: 10.1038/sj.bjp.0706241. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.