

Abstract
Background: To provide novel insight into the development of new therapeutic strategies to combat breast cancer, differentiation-inducing activity of clinically achievable concentrations of arsenite (AsIII) and tetrandrine (Tetra) was investigated in breast cancer cell lines MDA-MB-231 and MCF-7. Methods: Differentiation induction of cancer cells was analyzed by flow cytometer. Alterations of genes related to differentiation, and proliferation of human normal peripheral blood mononuclear cells (PBMCs) were analyzed using western blotting and cell viability assay, respectively. Results: Exposure to Tetra alone or in combination with AsIII induced differentiation of both cells characterized by upregulation of ICAM-1, downregulation of Her2/neu. In comparison with MCF-7, the combination of lower concentrations of AsIII and Tetra induced differentiation of MDA-MB-231, indicating that MDA-MB-231 cells were highly susceptible to differentiation. The differentiation occurred in parallel with activation of Erk signaling pathway, and was abolished by PD98059, a potent Erk inhibitor. Consistent with in vitro experimental results, the upregulation of ICAM-1 and the activation of Erk signaling pathway were also observed in MDA-MB-231 breast tumors in xenograft mouse obtained from our previous study. No obvious proliferation inhibition of PBMCs was observed following the exposure to AsIII combined with Tetra at the concentrations capable of inducing differentiation of MDA-MB-231 cells. Conclusion: The Erk signaling pathway may be crucially involved in the differentiation induction of breast cancer cells in vitro and in vivo. Collectively, our results suggest that the combination can probably serve as promising candidates for the development of novel therapeutic approaches for different types of breast cancer.
Keywords: Arsenite, tetrandrine, breast cancer cells, differentiation, combination therapy, Treg cells
Introduction
Despite advances in early detection, diagnosis, and targeted treatment options, breast cancer is still the most common cancer among women worldwide and persists as one of the leading causes of cancer-related deaths in women [1,2]. The need for novel therapeutic strategies remains paramount given the sustained development of drug resistance, tumor recurrence, and metastasis [1,2]. In this regard, remarkable clinical results achieved with arsenic trioxide (As2O3, a trivalent arsenic derivative) in the treatment of acute promyelocytic leukemia (APL) has encouraged more researchers to explore its potential treatment application for other malignancies, including solid tumors [3,4]. Several research groups including us have demonstrated that arsenic compounds such as trivalent arsenic (arsenite, AsIII) exhibit inhibitory effects against breast cancer cells [5-11]. We recently demonstrated a clear cytotoxic effect of AsIII against estrogen receptor (ER)-positive human breast cancer cell line MCF-7, and further clarified that tetrandrine (Tetra), a bis-benzylisoquinoline alkaloid isolated from the root of Stephania tetrandra S. Moore, significantly enhanced the cytotoxicity of AsIII in a synergistic manner [7]. In addition, we recently demonstrated antitumor activity of AsIII in combination with Tetra against human triple-negative breast cancer (TNBC) cell line MDA-MB-231 in vitro and in vivo [8]. These previous findings thus raised the possibility of utilizing arsenic compounds to treat patients with breast cancer.
The aim of differentiation therapy is to induce the differentiation of malignant cells, consequently cause them to cease proliferation, ultimately control their tumorigenic and malignant potential [12,13]. In general, differentiation therapy possesses the obvious characteristics of relatively low toxicity compared with conventional chemotherapy [12,13]. Use of all-trans retinoic acid (ATRA) and/or As2O3 in the treatment of APL has acquired a therapeutic niche, representing one of the most successful model of differentiation therapy [4]. In this respect, we have demonstrated that granulocyte colony-stimulating factor potentiates differentiation induction by ATRA and As2O3 and enhances arsenic uptake in an APL cell line HT93A [14]. We also demonstrated that not only ATRA but also valproic acid induced differentiation in NB4, another APL cell line, and their combination further augmented the differentiation activity, in which expression of transcription factors, CCAAT/enhancer-binding proteins (CEBPα, β) and PU.1 were closed involved [15]. More recently, we clarified that dasatinib, an inhibitor for Src family kinases, enhanced the differentiation-inducing activity of ATRA and dihydroxyvitamin D3 (VD3) in HL-60 cells [16]. Despite this, the effect of AsIII and Tetra on breast cancer cells in terms of differentiation induction remains largely unexplored.
In this study, in order to provide novel insights into the development of new therapeutic strategies to combat breast cancer using a combined regime of AsIII and Tetra, differentiation-inducing activity of the two drugs, each alone or in combination, was investigated in two different types of human breast cancer cell lines, MDA-MB-231 and MCF-7. As mentioned above, we recently demonstrated that long-term co-administration of AsIII and Tetra significantly reduced tumor volume and weight in MDA-MB-231 mouse xenografts [8]. Whether similar differentiation occurred in tumor tissues obtained from our previous in vivo study was further investigated in order to confirm the in vitro findings of the current study. The effects of AsIII and Tetra, each alone or in combination, on the population of CD4+ T cells and CD4+CD25+Foxp3+ regulator T (Treg) cells in mitogen-activated human normal peripheral blood mononuclear cells (PBMCs) were also evaluated, based on the fact that Treg cells have been suggested to play critical role in limiting antitumor immune response and promoting immunological ignorance of cancer cells [17-20].
Materials and methods
Materials
Sodium arsenite (NaAsO2, AsIII) and tetrandrine (Tetra) were purchased from Tri Chemical Laboratories (Yamanashi, Japan) and National Institutes for Food and Drug Control (Beijing, China), respectively. Fetal bovine serum (FBS) was purchased from Nichirei Biosciences (Tokyo, Japan). Dulbecco’s modified Eagle’s medium (DMEM), RPMI-1640 medium, phenazine methosulfate (PMS) and dimethyl sulfoxide (DMSO) were obtained from Wako Pure Chemical Industries (Osaka, Japan). 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)carbony]-2H-tetrazolium hydroxide (XTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Concanavalin A (ConA), a conventionally known T-cell mitogen, was obtained from Seikagaku Kogyo Co., Ltd. (Tokyo, Japan). ERK inhibitor PD98059 was purchased from Calbiochem (La Jolla, CA, USA). Can Get Signal® Immunoreaction Enhancer Solution were purchased from Toyobo Co., Ltd. (Osaka, Japan).
Cell culture and treatment
Breast cancer cell lines, MDA-MB-231 and MCF-7, were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and the RIKEN Cell Bank (Ibaraki, Japan), respectively. MDA-MB-231 cells were cultured in DMEM medium, and MCF-7 cells were cultured in RPMI-1640 medium, both of which were supplemented with 10% heat-inactivated FBS and 100 U/ml of penicillin and 100 µg/ml of streptomycin, in a humidified 5% CO2 atmosphere at 37°C. PBMCs were isolated from healthy volunteers using lymphocyte separation solution (d=1.077) (Nakarai Co., Kyoto, Japan) according to the method previously described [20,21]. Briefly, 3 ml of heparinized blood was loaded on 3 ml of lymphocyte separation solution. After centrifugation at 400×g for 30 min at room temperature, the opaque interface containing PBMCs was transferred to a clean centrifuge tube and washed three times with PBS. PBMCs were also cultured in RPMI-1640 medium supplemented with 10% heat-inactivated FBS, 100 U/ml of penicillin and 100 μg/ml of streptomycin in a humidified 5% CO2 atmosphere at 37°C. For experiments, the cell density was adjusted to 5×105 cells/ml prior to the treatments. This study has been approved by the IRB committee of Tokyo University of Pharmacy and Life Sciences. An informed consent was obtained from all healthy volunteers.
Differentiation analysis
Intercellular adhesion molecule-1 (ICAM-1), also referred to as CD54, is a cell surface glycoprotein member of the immunoglobulin superfamily responsible for regulating cell/cell contacts [22]. It is also known as a differentiation marker which is present in normal mammary epithelium but frequently downregulated in breast cancer cells [23]. Immunohistochemical examination of resected tumors from female patients with invasive breast cancer revealed that ICAM-1 expression had negative correlation to tumor size and infiltration, and that patients with ICAM-1 positive tumors had better relapse-free and overall survival than those with negative tumors [24]. The expression level of ICAM-1 was assessed using a FACSCanto flow cytometer (Becton Dickinson, San Jose, CA, USA) according to a method previously described with slight modifications [25,26]. Briefly, after the treatment with various concentrations of AsIII and Tetra, alone or in combination, for 96 h, ~1×106 cells were collected and washed once with PBS (pH 7.4) (Gibco®; Thermo Fisher Scientific, Waltham, MA, USA), followed by the addition of 10 μl of monoclonal mouse anti-human CD54 PE-conjugated antibody (BioLegend San Diego, CA, USA). After the incubation for 20 min in the dark at 37°C, the cells were washed with PBS and resuspended in 0.4 ml staining buffer [0.4% (v/v) formaldehyde neutral buffer solution in PBS (pH 7.4); Nacalai Tesque, Inc., Kyoto, Japan], and then analyzed by flow cytometry. The results were shown as mean fluorescence intensity (MFI) with linear scaling and calculated by subtracting the mean fluorescence of the unstained cells from that of the stained cells, as described previously [15,16,27]. In order to evaluate the correlation between the activation of Erk and the differentiation induced by AsIII and Tetra, breast cancer cell lines were treated with 10 µM PD98059 for 30 min prior to the treatment with various concentrations of AsIII and Tetra, alone or in combination, in the presence or absence of 10 µM PD98059 for an additional 96 h, followed by differentiation analysis as described above.
Proliferation assay of PBMCs
The effect of AsIII and Tetra, alone or in combination, on the proliferation of PBMCs was measured by XTT dye-reduction assay according to the method previously described with slight modifications [7,8,21]. Briefly, PBMCs was adjusted to 5×105 cells/ml, and 186 μl of the cell suspension was inoculated into 96-well plates, followed by the addition of 10 μl of ConA (0.1 mg/ml in PBS) at final concentrations of 5.0 μg/ml. Subsequently, 4 μl of each drug or their combination solution was added to give final indicated concentrations. Cells in sixplicate were treated with various concentrations of AsIII and Tetra, alone or in combination, for 72 h. Following the treatment, XTT and PMS were added into each well at final concentrations of 0.2 mg/ml and 1 mM, respectively. After incubation at 37°C for 4 h, the plates were mixed and the absorbance at 450 nm was measured with a microplate reader (EMax Plus®, Molecular Devices, CA, USA). The relative cell viability was expressed as the ratio of the absorbance of each treatment group against those of the corresponding untreated control group. Data are shown as means ± standard deviation (SD) from more than three independent experiments.
Analysis of CD4+ T cells and Treg cells
After treatment with 0.5 μM AsIII, 0.5 μg/ml Tetra, each alone or in combination, for 72 h, alterations of CD4+ T cells and Treg cells population in mitogen-activated PBMCs were analyzed using a FACSCalibur™ II flow cytometer (BD Biosciences, Mountain View, CA, USA) according to the method previously described with slight modifications [19,20]. Briefly, after treatment with the two drugs, ~1×106 cells were collected and washed once with PBS (pH 7.4) (Gibco®; Thermo Fisher Scientific, Waltham, MA, USA), followed by the addition of 10 μl of monoclonal mouse anti-human CD4 PerCP-Cy5.5-conjugated antibody and 10 μl of monoclonal mouse anti-human CD25 PE-conjugated antibody (BD Biosciences), respectively. To exclude the amount of non-specific binding, 10 μl of PerCP-Cy5.5-conjugated and 10 μl of PE-conjugated mouse IgG1 κ isotype control (BD Biosciences) were used and evaluated as background, respectively. After the incubation for 20 min in the dark at 37°C, the cells were washed with PBS and resuspended in diluted Foxp3 buffer A (BD Biosciences), followed by the incubation for 10 min in the dark at room temperature. Then, the cells were washed once with PBS and resuspended in 0.2 ml of Foxp3 buffer C composed of 49 parts Foxp3 buffer A and 1 part Foxp3 buffer B (BD Biosciences), and incubated for 30 min in the dark at room temperature. After the incubation, the cells were washed once with PBS and 10 μl of mouse anti-human Foxp3 Alexa Fluor® 488-conjugated antibody or 10 μl of Alexa Fluor® 488-conjugated mouse IgG1 isotype control to re-suspend the pellet was added, and incubation for 30 min in the dark at 37°C followed. After washing once with PBS, the cells were resuspended in 0.4 ml staining buffer [0.4% (v/v) formaldehyde neutral buffer solution in PBS (pH 7.4); Nacalai Tesque, Inc., Kyoto, Japan] and analyzed by flow cytometry, and then the data were further analyzed using CellQuest software (BD Biosciences). CD4+ T cells in the lymphocyte fraction were gated, and the percentages of CD4+ T cells and Treg cells in the CD4+ T cells were calculated, respectively.
Western blot analysis
For protein samples preparation, cell pellets (approximately 1-2×106 cells per 110 μl Laemmli buffer) were suspended in lysis buffer (Laemmli buffer containing 100 mM DTT, 2 μg/ml leupeptin, 2 μg/ml aprotinin, 1 μg/ml pepestain, 1 mM PMSF). The suspensions of cells were sonicated using a sonicator (Qsonica, LLC, CT, USA) with 10 short bursts of 2 sec followed by intervals of 2 sec for cooling. The suspensions were kept at all times in an ice bath. Sonicated cells were heated in 95°C for 5 min, and then centrifuged at 13,000 g for 15 min at 4°C. Protein concentrations of the supernatant were determined according to Bradford’s method using the protein assay dye reagent (Bio-Rad, CA, USA) according to the manufacturer’s instructions, and using BSA as the standard. Western blot analysis was carried out according to the methods previously described [28]. Briefly, after separation of proteins on a sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis, followed by transferring to a polyvinylidene difluoride (PVDF) membrane (Millipore Corp, MA, USA), protein bands were detected using the following primary antibodies and dilution ratios: mouse anti-human β-actin (1:5000 dilution; cat. no. A-5441; Sigma-Aldrich, MO, USA), mouse anti-human ICAM-1 (G-5) (1:1000 dilution; cat. no. sc-8439; Santa Cruz Biotechnology, CA, USA), rabbit anti-human HER2/ErbB2 (29D8) (1:1000 dilution; cat. no. 2165), rabbit anti-human phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (1:2000 dilution; cat. no. 4695) and p44/42 MAPK (Erk1/2) (137F5) (1:1000 dilution; cat. no. 4370) (Cell Signaling Technology, MA, USA). Blotted protein bands were detected with respective horseradish peroxidase-conjugated secondary antibody and an enhanced chemiluminescence (ECL) Western blot analysis system (Amersham Pharmacia Biotech, Buckinghamshire, UK). In order to confirm the in vitro findings of the current study, the alteration of the expression levels of ICAM-1, phospho-Erk and Erk was also evaluated in previous stocked tumor tissues obtained from MDA-MB-231 mouse xenografts, which have been used to clarify that co-administration of AsIII and Tetra significantly reduced tumor volume and weight in the mouse xenografts in our previous in vivo study [8]. Similarly to the protein samples preparation from cancer cell lines, the previous stocked tumor tissues (at a ratio of approximately 1 g of tissue per 10 ml Laemmli buffer) obtained from MDA-MB-231 mouse xenografts were also suspended in lysis buffer, and then sonicated, followed by the same procedures described as above.
Statistical analysis
Experiments were independently repeated three times, and the results are presented as the means ± standard deviation (SD) of the three assays. Statistical analysis was performed using GraphPad Prism® 6 software. The Student’s t-test was used to compare sample means between two groups, and one-way analysis of variance followed by Dunnett’s post hoc test was used to compare sample means among three or more groups. A probability level of P<0.05 was considered statistically significant.
Results
Induction of the expression of ICAM-1 in breast cancer cells treated with AsIII and Tetra, alone or in combination
ICAM-1 has been identified and used as a marker of differentiation for human mammary epithelial cells and human breast cancer cells in several studies [25,26,29,30]. Furthermore, ATRA, which has been successfully used as a differentiation agent in the treatment of APL, has been reported to induce differentiation of human breast cancer cell lines by upregulating ICAM-1 expression [26]. Consistent with the previous report, both flow cytometry (FCM) and western blot (WB) analysis demonstrated that 1 μM ATRA, served as positive control in the current study, prominently upregulated the expression of ICAM-1 in MDA-MB-231 cells (Figures 1A, 1C and 2A), although similar phenomena were not observed in MCF-7 cells (Figures 1B, 1D and 2B).
Figure 1.

Induction of the expression of ICAM-1 in breast cancer cells treated with AsIII and Tetra, alone or in combination. The expression level of ICAM-1 was determined using a FACSCanto flow cytometer after the treatment with various concentrations of AsIII alone (0.125, 0.25, 0.5, 1 and 2 µM), Tetra alone (0.125, 0.25, 0.5, 1 and 2 µg/ml), their combination in constant ratio (0.125 µM AsIII+0.125 µg/ml Tetra, 0.25 µM AsIII+0.25 µg/ml Tetra, 0.5 µM AsIII+0.5 µg/ml Tetra, 1 µM AsIII+1 µg/ml Tetra and 2 µM AsIII+2 µg/ml Tetra) for 96 h. The expression levels of ICAM-1 in breast cancer cells were analyzed and presented as histograms plotted by FACS as described in methods section. Representative FACS histograms of ICAM-1 expression in MDA-MB-231 (A) and MCF-7 (B) are shown. Untreated and treated cells are represented by filled histograms and open histograms, respectively. Relative expression levels of ICAM-1 in MDA-MB-231 cells (C) and MCF-7 cells (D) were calculated, respectively, based on FACS profiles shown in (A) and (B). Experiments were independently repeated at least three times and results are shown as means ± SD. *, P<0.05 vs. control; #, P<0.05 vs. each alone. As, AsIII; Tetra, tetrandrine; ATRA, all-trans retinoic acid. MFI mean fluorescence intensity. ATRA (1 µM) used as a positive control for differentiation.
Figure 2.
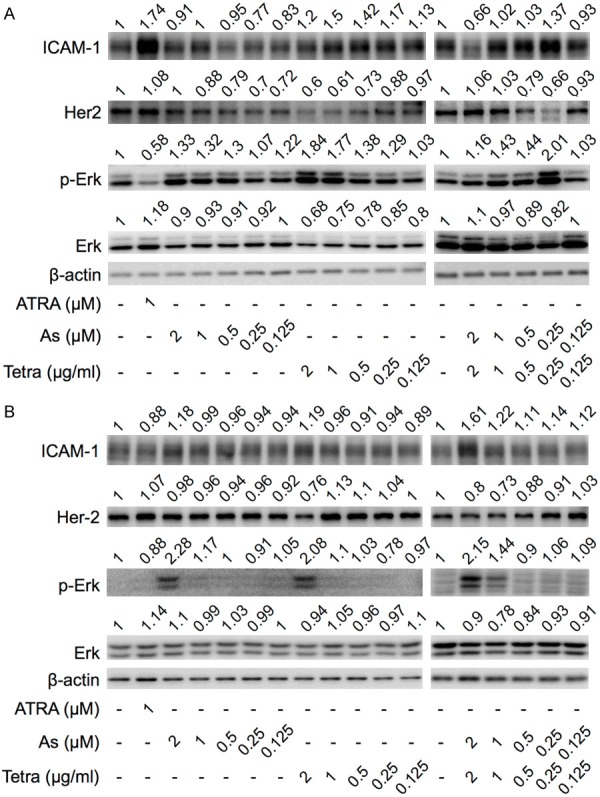
Expression profile of differentiation-related proteins in breast cancer cells treated with AsIII and Tetra, alone or in combination. After treatment with indicated concentrations of AsIII and Tetra, alone or in combination, for 96 h, the expression profiles of differentiation-related proteins in MDA-MB-231 cells (A) and MCF-7 cells (B) were analyzed using western blot as described in material and methods. Representative image of the expression profile of each protein is shown from three independent experiments. The densitometry of protein bands was analyzed using a program, NIH ImageJ 1.52a. The values above each image represent the ratios between each key molecule and β-actin protein expression levels, which were further compared with those of control group (untreated cells). p-Erk, phosphorylated Erk; As, AsIII; Tetra, tetrandrine; ATRA, all-trans retinoic acid. ATRA (1 µM) used as a positive control for differentiation.
Since it has been established that the plasma concentrations of AsIII were generally believed to be maintained between 0.1-2 μM in APL patients treated with As2O3 [4], clinically achievable concentrations of AsIII (under 2 μM) were used throughout the study. As shown in Figures 1A, 1C and 2A, FCM results showed that relatively high concentrations of Tetra (0.5, 1 and 2 μg/ml) slightly but significantly induced the expression of ICAM-1 in MDA-MB-231 cells, which is in good agreement with the WB experimental results, indicating its differentiation-inducing activity in the cells. Whereas almost no alteration of ICAM-1 expression was observed in the cells treated with different concentrations of AsIII regardless of analytical approaches. Intriguingly, both FCM and WB results showed that the combination of relatively low concentrations of AsIII and Tetra, especially 0.25 μM AsIII+0.25 μg/ml Tetra, slightly but significantly upregulated the expression level of ICAM-1, although similar upregulation was not observed in the cells when treated with either drug alone. In contrast, the combination of 1 μM AsIII+1 μg/ml Tetra failed to upregulate the expression level of ICAM-1 in MDA-MB-231 cells, although 1 μg/ml Tetra alone exhibited activity to induce ICAM-1 expression. In addition, exposure to the combination of the highest concentrations of 2 μM AsIII+2 μg/ml Tetra resulted in a significant downregulation of the expression of ICAM-1, which might be attributed to the cytotoxicity of the long-term combination treatment.
In comparison with MDA-MB-231 cells, only the highest concentrations of AsIII (2 μM) and Tetra (2 μg/ml) alone upregulated the expression level of ICAM-1 in MCF-7 cells (Figures 1B, 1D and 2B). FCM results also demonstrated that the combination of relatively high concentrations of AsIII and Tetra (1 μM AsIII+1 μg/ml Tetra and 2 μM AsIII+2 μg/ml Tetra) slightly but significantly upregulated the expression level of ICAM-1 when compared to control group and each drug alone (Figure 1B and 1D), which were largely consistent with WB results (Figure 2B), indicating that the two drugs exhibit a stronger differentiation-inducing activity than ATRA in MCF-7 cells.
Effects of AsIII and Tetra, alone or in combination, on the expression level of Her2/neu and the activation of Erk pathway in breast cancer cells
Downregulation of human epidermal growth factor receptor type 2 (Her2/neu) has been reported to be associated with differentiation of breast cancer cells including MCF-7 [31,32]. As shown in Figure 2A, consistent with the alteration profiles of ICAM-1, a clear dose-dependent decrease in the expression level of Her2/neu was observed in MDA-MB-231 treated with Tetra, especially at its relatively high concentrations of 1 and 2 μg/ml. Similarly, the expression of Her2/neu was also reduced by approximate 40% following the combinatorial treatment with 0.25 μM AsIII+0.25 μg/ml Tetra. On the other hand, a slight downregulation of Her2/neu expression was observed in the cells after treatment with relatively low, but not high concentrations of AsIII, which might not be directly linked to differentiation. In the case of MCF-7, as shown in Figure 2B, the expression level of Her2/neu was downregulated by approximately 20-30% following the treatment with 2 μg/ml Tetra, and the combination of two drugs (1 μM AsIII+1 μg/ml Tetra and 2 μM AsIII+2 μg/ml Tetra), almost in parallel with the alteration profiles of ICAM-1.
In addition, extracellular-regulated kinase (Erk) signaling pathway has been demonstrated to be involved in the differentiation of human breast cancer cells [32,33]. As shown in Figure 2, the expression levels of phosphorylated Erk (phospho-Erk) were obviously upregulated by the treatment with AsIII and Tetra, each alone or in combination, at the concentrations of which significantly upregulated the expression levels of ICAM-1 in both breast cancer cells, indicating the activation of Erk signaling pathway. In accordance with the upregulation of phospho-Erk, a slight downregulation of total-Erk was also observed in MDA-MB-231 cells (Figure 2A).
Upregulation of ICAM-1 and activation of Erk signaling pathway in MDA-MB-231 mouse xenografts treated with AsIII and Tetra, alone or in combination
We recently demonstrated growth inhibition of MDA-MB-231 by AsIII and Tetra in vitro and in vivo, in which long-term (10 weeks) co-administration of AsIII (2 mg/kg/day) and Tetra (20 mg/kg/day) significantly reduced tumor volume and weight, directly supporting their in vitro antitumor activity [8]. In order to explore whether differentiation is also involved in the tumor growth inhibition in vivo, the expression levels of ICAM-1, phospho-Erk as well as total Erk were evaluated in previous stocked tumor tissues obtained from the above-mentioned MDA-MB-231 mouse xenografts. As shown in Figure 3A and 3B, in comparison with vehicle-control group, a significant upregulation of ICAM-1 was observed in the isolated tumors derived from mice treated with either drug alone or their combination. Intriguingly, alteration of phospho-Erk/Erk ratio was further strengthened by the combination as compared to either drug alone, indicating the activation of Erk signaling pathway (Figure 3A and 3C).
Figure 3.
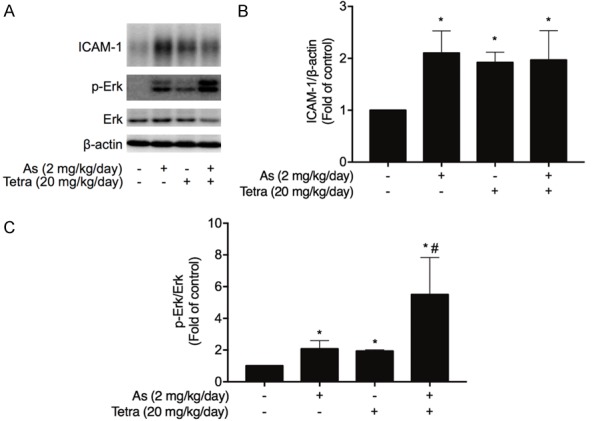
Upregulation of ICAM-1 and activation of Erk signaling pathway in MDA-MB-231 mouse xenografts treated with AsIII and Tetra, alone or in combination. MDA-MB-231 tumors in the mice xenograft model treated with AsIII and Tetra, alone or in combination, were obtained from our previous study, in which long-term (10 weeks) co-administration of AsIII (2 mg/kg/day) and Tetra (20 mg/kg/day) significantly reduced tumor volume and weight [8]. (A) The expression profiles of ICAM-1, phospho-Erk and total Erk were analyzed using western blot as described in material and methods. Results are representatives of three independent experiments. The relative expression levels of ICAM-1 (B) and phospho-Erk/Erk (C) were expressed as the ratios between each target gene protein and β-actin protein expression levels, and compared with those of untreated control group, respectively. Results are shown as the means ± SD from three independent experiments. p-Erk, phosphorylated Erk; As, AsIII; Tetra, tetrandrine. The procedures for establishment of MDA-MB-231 mouse xenografts, and the following administration of AsIII and Tetra, each alone or in combination, and the preparation of tumor tissues are shown in Supplementary Materials.
Effect of PD98059 on the expression profile of ICAM-1 induced by AsIII and Tetra, alone or in combination, in breast cancer cells
In order to clarify the correlation between the activation of Erk signaling pathway and the differentiation of breast cancer cells, the effect of PD98059, a potent Erk inhibitor, on the expression level of ICAM-1 induced by AsIII and Tetra was further investigated. Considering the concept of minimizing the side effects of AsIII, and the capacity of two drugs to induce differentiation in different breast cancer cells, differential concentrations of AsIII and Tetra were used. As shown in Figure 4A, similar to the results in Figure 1C, exposure to the combination of 0.25 μM AsIII+0.25 μg/ml Tetra and 1 μg/ml Tetra significantly upregulated ICAM-1 expression in MDA-MB-231 cells, respectively. The upregulation of ICAM-1 was also observed in the cells when treated with 0.25 μg/ml Tetra alone, whereas there was only a modest upregulation of ICAM-1 following the same treatment in Figure 1C, the slight discrepancy of which might be attributed to batch cultures. Of note, the addition of PD98059 completely abrogated the upregulation of ICAM-1 triggered by the two drugs. Intriguingly, the addition of PD98059 also resulted in a slight but significant downregulation of endogenous level of ICAM-1 in MDA-MB-231 cells, indicating the critical role of Erk signaling pathway in regulating ICAM-1 expression. Similar to MDA-MB-231 cells, the upregulation of ICAM-1 expression triggered by 2 μM AsIII and 2 μg/ml Tetra, each alone or in combination, was also obviously reversed by the addition of PD98059 in MCF-7 cells (Figure 4B).
Figure 4.
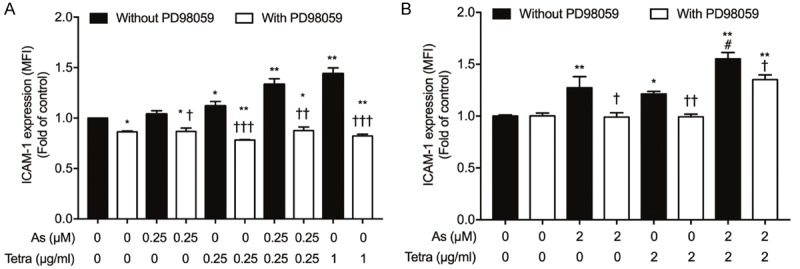
Effect of PD98059 on the expression profile of ICAM-1 induced by AsIII and Tetra, alone or in combination, in breast cancer cells. After treatment with indicated concentrations of AsIII and Tetra, alone or in combination, in the presence or absence of 10 μM PD98059, for 96 h, the expression profiles of ICAM-1 were determined using a FACSCanto flow cytometer as described in material and methods. Relative expression levels of ICAM-1 in MDA-MB-231 (A) and MCF-7 (B) were calculated based on their respective FACS profiles. *, P<0.01, **, P<0.0001 vs. control; †, P<0.05, ††, P<0.001, †††, P<0.0001 group with PD98059 vs. respective group without PD98059; #, P<0.001 vs. each alone. As, AsIII; Tetra, tetrandrine.
Effect of AsIII and Tetra, each alone or in combination, on the proliferation of PBMCs
As shown in Figure 5, an approximately 20% proliferation inhibition of PBMCs was observed following exposure to various concentrations of AsIII (0.125-2 μM) alone for 72 h, reflecting its good tolerance in clinical use. Moreover, almost no proliferation inhibition was observed in the cells after treatment with a relatively low concentration of Tetra (0.125-0.5 μg/ml), whereas a considerable inhibitory effect of a relatively high concentration of Terta (1 and 2 μg/ml) was observed. Of note, the combination of a relatively low concentration of the two drugs (0.125 μM AsIII+0.125 μg/ml Tetra, 0.25 μM AsIII+0.25 μg/ml Tetra, 0.5 μM AsIII+0.5 μg/ml Tetra) did not result in a clear proliferation inhibition, indicating that Tetra was capable of reversing the inhibitory effect of AsIII on PBMCs. On the other hand, approximately 30% and 60% proliferation inhibition of PBMCs were observed following the exposure to the combination of 1 μM AsIII+1 μg/ml Tetra and 2 μM AsIII+2 μg/ml Tetra, respectively.
Figure 5.
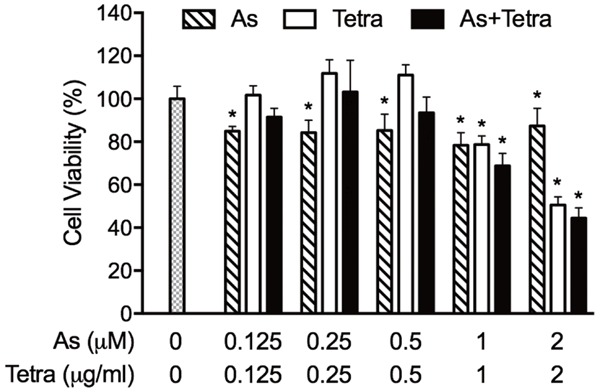
Effects of AsIII and Tetra, alone or in combination, on the proliferation of PBMCs. Following the treatment with various concentrations of AsIII alone (0.125, 0.25, 0.5, 1, 2 μM), Tetra alone (0.125, 0.25, 0.5, 1, 2 μg/mL), or their combination (0.125 μM+0.125 μg/mL, 0.25 μM+0.25 μg/mL, 0.5 μM+0.5 μg/mL, 1 μM+1 μg/mL, 2 μM+2 μg/mL) for 72 h, cell viability was determined by XTT assay as described under materials and methods. Relative cell viability was calculated as the ratio of the absorbance at 450 nm of each treatment group against those of the corresponding untreated control group. Data are shown as the means ± SD from more than three independent experiments. *P<0.05, vs control. As, AsIII; Tetra, tetrandrine.
Effects of AsIII and Tetra, each alone or in combination, on the populations of CD4+ T cells and Treg cells in mitogen-activated human PBMCs
The populations of CD4+ T cells and Treg cells in mitogen-activated human PBMCs were analyzed by flow cytometry following the treatment with two drugs for 72 h. In the experiment, 0.5 μM AsIII and 0.5 μg/ml Tetra were selected since each of them are their respective highest concentrations with limited or no proliferation inhibition, and their combination exhibited no obvious cytotoxicity against PBMCs (Figure 5). As shown in Figure 6B and 6C, exposure to 0.5 μg/ml Tetra tended to decrease the population of Treg cells, whereas 0.5 μM AsIII had little effect in this respect. Similar to the effect of Tetra alone, a slight downregulation of the population of Treg cell was also observed when treated with the combination of 0.5 μM AsIII+0.5 μg/ml Tetra, although the changes were not statistically significant. On the other hand, no influence of two drugs, each alone or in combination, on the population of CD4+ T cells was observed (Figure 6D).
Figure 6.

Effects of AsIII and Tetra, each alone or in combination, on the population of CD4+ T cells and Treg cells in mitogen-activated human PBMCs. After treatment with 0.5 μM AsIII and 0.5 μg/ml Tetra, alone or in combination, for 72 h, alterations of CD4+ T cell and Treg cell populations in ConA-activated PBMCs were analyzed by a flow cytometer as described in materials and methods. (A) Lymphocyte fraction of PBMCs (a1) were gated for CD4+ T cells (a2), and the percentage of Treg cells in the CD4+ T cells fraction in each treatment were calculated (B). Treg cells (C) and CD4+ T cells (D) in ConA-activated PBMCs treated with 0.5 μM AsIII and 0.5 μg/ml Tetra, alone or in combination, for 72 h were calculated, respectively. Data are presented as means ± SD from more than three independent experiments. As, AsIII; Tetra, tetrandrine.
Discussion
Previous studies have demonstrated that certain chemicals such as ATRA and VD3 are capable of inducing the differentiation of malignant cells, consequently causing them to cease proliferation instead of killing them through cytotoxicity [4,14-16]. Generally, differentiation therapy possesses the obvious characteristics of relatively low toxicity compared with conventional chemotherapy [12,13]. Although the differentiation-inducing activity of Tetra has been demonstrated in leukemic cells [34,35], whether differentiation induction occurs in breast cancer cells has not yet been investigated. Results from this study demonstrated for the first time that relatively high concentrations of Tetra (>0.5 μg/ml) alone clearly induced differentiation in different types of breast cancer cells regardless of their estrogen dependency, as evidenced by the upregulation of ICAM-1 and downregulation of Her2/neu proteins. Indeed, similar alterations of both ICAM-1 and Her2/neu proteins have been reported to be associated with differentiation of human breast cancer cells [25,26,29-32]. Consistent with the findings in the current study, previous reports also showed that similar concentrations (2 μg/ml) of Tetra forced leukemic cells including HL-60, NB4 and K562 to differentiate [34,35]. In comparison to previous report showing 0.5 μM arsenic trioxide (As2O3) induced differentiation of breast cancer cell lines including MDA-MB-231 and MCF-7 [25], obvious features of differentiation were not observed in the current study. The discrepancy might be explained by the following facts: 1) NaAsO2 was used in the current study, while As2O3 was used in the previous study, 2) different sources of breast cancer cells in two researches. In fact, the breast cancer cells used in the previous study were obtained from ATCC, whereas in the current study MCF-7 were obtained from the RIKEN Cell Bank (Ibaraki, Japan). In agreement with the previous report [25], our in vivo study experimentally demonstrated that long-term administration of AsIII induced differentiation of tumor tissue obtained from previous stocked MDA-MB-231 mouse xenografts, suggesting its differentiation-inducing activity in vivo.
We further demonstrated that exposure to the combination of AsIII and Tetra at their respective clinically acceptable concentrations induced differentiation in both cancer cells. Again, the combination-triggered differentiation was also confirmed in tumor tissues in MDA-MB-231 mouse xenografts, suggesting that two drugs-mediated differentiation partially contributed to the achievement of long-term co-administration of AsIII and Tetra to reduce tumor volume and weight in our recent report [8]. Of note, the combination of relatively low concentrations of AsIII and Tetra (0.25 μM AsIII+0.25 μg/ml Tetra) significantly induced differentiation in MDA-MB-231 cells. In comparison, only relatively high concentrations of two drugs (1 μM AsIII+1 μg/ml Tetra and 2 μM AsIII+2 μg/ml Tetra) exhibited similar differentiation-inducing activity in MCF-7, suggesting that MDA-MB-231, known as a TNBC cell line, were highly susceptible to differentiation. Considering TNBC is one of the most difficult subtypes of breast cancer to treat due to its aggressive, metastatic behavior, and a lack of a targeted therapy [36], the combination of AsIII and Tetra, which have long been successfully used in clinic, can probably serve as promising candidates to differentiate cancer cells, ultimately cease their uncontrolled proliferation in patients with TNBC. Previous studies have demonstrated that VES, a derivate of vitamin E (RRR-α-tocopheryl) induced differentiation in human breast cancer cells including MCF-7 in parallel with the downregulation of Her2/neu [31,32]. Furthermore, it has been clarified that antibodies reacting with Her2/neu receptor can induce differentiation of human breast cancer cells [31,37]. In agreement with these previous reports, following exposure to the combination of AsIII and Tetra, a clear downregulation of Her2/neu protein was observed in MDA-MB-231 as well as MCF-7 cells. In addition, it has been demonstrated that the ability of VES to induce MCF-7 cells differentiation does depend on Erk signaling pathway, based on the fact that the addition of PD98059 blocked the differentiation-inducing activity of VES [32]. Similarly, Liu et al. also demonstrated that the activation of Erk signaling pathway was involved in genistein-induced differentiation of breast cancer stem/progenitor cells [33]. In line with previous reports, coincidentally with differentiation induction, the activation of Erk signaling pathway was observed in MDA-MB-231, MCF-7 cells as well as tumor tissue derived from MDA-MB-231 mouse xenografts following the treatment with the combination of AsIII and Tetra. Additionally, the differentiation of both cancer cells was significantly reversed by the addition of PD98059, suggesting the critical role of Erk signaling pathway in the differentiation induction.
We also demonstrated no obvious proliferation inhibition of PBMCs following the exposure to the combination of AsIII and Tetra (0.125 μM AsIII+0.125 μg/ml Tetra, 0.25 μM AsIII+0.25 μg/ml Tetra, 0.5 μM AsIII+0.5 μg/ml Tetra) for 72 h, which clearly induced differentiation of MDA-MB-231 cells. In clinic, the plasma concentrations of AsIII were generally believed to be maintained between 0.1-2 μM in APL patients treated with As2O3 [4]. Collectively, these results suggest the potential therapeutic efficacy of the combination to patients with TNBC. Our results also showed a slight downregulation of the population of Treg cells following treatment with 0.5 μM AsIII combined with 0.5 μg/ml Tetra, consistent with our recent report [38]. Given that Treg cells play a critical role in tumor immunotolerance by suppressing antitumor immunity [17,18,39], the clinic use of Tetra, as an adjuvant therapeutic agent, might provide benefits to patients treated with AsIII, although accurate effects of the combination on the population of Treg cells warrants further investigation in a breast cancer xenograft model.
Conclusions
Our results demonstrated that clinically achievable concentrations of AsIII combined with Tetra clearly induced differentiation in different types of breast cancer cells regardless of their estrogen dependency, and that MDA-MB-231 cells were more susceptible to differentiation than MCF-7 cells. Although further investigations are needed to clarify the mechanisms underlying the differentiation, our results suggest that the activation of Erk signaling pathway and downregulation of Her2/neu are closely implicated in the differentiation. We further demonstrated that exposure to the two drugs at the concentrations inducing differentiation of MDA-MB-231 cells did not result in a clear proliferation inhibition of PBMCs, whereas caused a slight downregulation of the population of Treg cells. Collectively, our results suggest that the combination can probably serve as promising candidates for the development of novel therapeutic approaches for different types of breast cancer, especially TNBC.
Acknowledgements
This work was partially supported by The Japan Society for the Promotion of Science (JSPS) KAKENHI Grant to Bo Yuan (Grant Numbers 26460233) (Grant Numbers 17K08465). This work was partially supported by National Natural Science Foundation of China (NSFC) to Xiaohua Pei (Grant Number 81774319). This work was also partially supported by Beijing Municipal Natural Science Foundation (Grant Number 7182098).
Disclosure of conflict of interest
None.
Supporting Information
References
- 1.Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63:11–30. doi: 10.3322/caac.21166. [DOI] [PubMed] [Google Scholar]
- 2.Taylor S, Lam M, Pararasa C, Brown JE, Carmichael AR, Griffiths HR. Evaluating the evidence for targeting FOXO3a in breast cancer: a systematic review. Cancer Cell Int. 2015;15:1. doi: 10.1186/s12935-015-0156-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dilda PJ, Hogg PJ. Arsenical-based cancer drugs. Cancer Treat Rev. 2007;33:542–564. doi: 10.1016/j.ctrv.2007.05.001. [DOI] [PubMed] [Google Scholar]
- 4.Yuan B, Yoshino Y, Kaise T, Toyoda H. Application of arsenic trioxide therapy for patients with leukaemia. In: Sun H, editor. Biological Chemistry of Arsenic, Antimony and Bismuth. Chichester: John Wiley Sons, Ltd.; 2010. pp. 263–292. [Google Scholar]
- 5.Liu W, Gong Y, Li H, Jiang G, Zhan S, Liu H, Wu Y. Arsenic trioxide-induced growth arrest of breast cancer MCF-7 cells involving FOXO3a and IkappaB kinase beta expression and localization. Cancer Biother Radiopharm. 2012;27:504–512. doi: 10.1089/cbr.2012.1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wang X, Gao P, Long M, Lin F, Wei JX, Ren JH, Yan L, He T, Han Y, Zhang HZ. Essential role of cell cycle regulatory genes p21 and p27 expression in inhibition of breast cancer cells by arsenic trioxide. Med Oncol. 2011;28:1225–1254. doi: 10.1007/s12032-010-9552-x. [DOI] [PubMed] [Google Scholar]
- 7.Yao M, Yuan B, Wang X, Sato A, Sakuma K, Kaneko K, Komuro H, Okazaki A, Hayashi H, Toyoda H, Pei X, Hu X, Hirano T, Takagi N. Synergistic cytotoxic effects of arsenite and tetrandrine in human breast cancer cell line MCF-7. Int J Oncol. 2017;51:587–598. doi: 10.3892/ijo.2017.4052. [DOI] [PubMed] [Google Scholar]
- 8.Yuan B, Yao M, Wang X, Sato A, Okazaki A, Komuro H, Hayashi H, Toyoda H, Pei X, Hu X, Hirano T, Takagi N. Antitumor activity of arsenite in combination with tetrandrine against human breast cancer cell line MDA-MB-231 in vitro and in vivo. Cancer Cell Int. 2018;18:113. doi: 10.1186/s12935-018-0613-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhao Y, Onda K, Sugiyama K, Yuan B, Tanaka S, Takagi N, Hirano T. Antitumor effects of arsenic disulfide on the viability, migratory ability, apoptosis and autophagy of breast cancer cells. Oncol Rep. 2019;41:27–42. doi: 10.3892/or.2018.6780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhao Y, Onda K, Yuan B, Tanaka S, Kiyomi A, Sugiyama K, Sugiura M, Takagi N, Hirano T. Arsenic disulfideinduced apoptosis and its potential mechanism in two and threedimensionally cultured human breast cancer MCF7 cells. Int J Oncol. 2018;52:1959–1971. doi: 10.3892/ijo.2018.4357. [DOI] [PubMed] [Google Scholar]
- 11.Zhao Y, Yuan B, Onda K, Sugiyama K, Tanaka S, Takagi N, Hirano T. Anticancer efficacies of arsenic disulfide through apoptosis induction, cell cycle arrest, and pro-survival signal inhibition in human breast cancer cells. Am J Cancer Res. 2018;8:366–386. [PMC free article] [PubMed] [Google Scholar]
- 12.de The H. Differentiation therapy revisited. Nat Rev Cancer. 2018;18:117–127. doi: 10.1038/nrc.2017.103. [DOI] [PubMed] [Google Scholar]
- 13.Talukdar S, Emdad L, Das SK, Sarkar D, Fisher PB. Evolving strategies for therapeutically targeting cancer stem cells. Adv Cancer Res. 2016;131:159–191. doi: 10.1016/bs.acr.2016.04.003. [DOI] [PubMed] [Google Scholar]
- 14.Iriyama N, Yuan B, Hatta Y, Horikoshi A, Yoshino Y, Toyoda H, Aizawa S, Takeuchi J. Granulocyte colony-stimulating factor potentiates differentiation induction by all-trans retinoic acid and arsenic trioxide and enhances arsenic uptake in the acute promyelocytic leukemia cell line HT93A. Oncol Rep. 2012;28:1875–1882. doi: 10.3892/or.2012.2006. [DOI] [PubMed] [Google Scholar]
- 15.Iriyama N, Yuan B, Yoshino Y, Hatta Y, Horikoshi A, Aizawa S, Takei M, Takeuchi J, Takagi N, Toyoda H. Enhancement of differentiation induction and upregulation of CCAAT/enhancer-binding proteins and PU.1 in NB4 cells treated with combination of ATRA and valproic acid. Int J Oncol. 2014;44:865–873. doi: 10.3892/ijo.2013.2236. [DOI] [PubMed] [Google Scholar]
- 16.Iriyama N, Yuan B, Hatta Y, Takagi N, Takei M. Lyn, a tyrosine kinase closely linked to the differentiation status of primary acute myeloid leukemia blasts, associates with negative regulation of all-trans retinoic acid (ATRA) and dihydroxyvitamin D3 (VD3)-induced HL-60 cells differentiation. Cancer Cell Int. 2016;16:37. doi: 10.1186/s12935-016-0314-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Facciabene A, Motz GT, Coukos G. T-regulatory cells: key players in tumor immune escape and angiogenesis. Cancer Res. 2012;72:2162–2171. doi: 10.1158/0008-5472.CAN-11-3687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sakaguchi S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nat Immunol. 2005;6:345–352. doi: 10.1038/ni1178. [DOI] [PubMed] [Google Scholar]
- 19.Song MM, Fang S, Tanaka S, Sugiyama K, Kiyomi A, Kato R, Onda K, Yuan B, Takagi N, Hu X, Hirano T. Effects of arsenic disulfide on proliferation, cytokine production, and frequencies of CD4(+), CD8(+), and regulatory T cells in mitogen-activated human peripheral blood mononuclear cells. Int Immunopharmacol. 2015;29:832–838. doi: 10.1016/j.intimp.2015.08.034. [DOI] [PubMed] [Google Scholar]
- 20.Yuan B, He J, Kisoh K, Hayashi H, Tanaka S, Si N, Zhao HY, Hirano T, Bian B, Takagi N. Effects of active bufadienolide compounds on human cancer cells and CD4+CD25+Foxp3+ regulatory T cells in mitogen-activated human peripheral blood mononuclear cells. Oncol Rep. 2016;36:1377–1384. doi: 10.3892/or.2016.4946. [DOI] [PubMed] [Google Scholar]
- 21.Yoshino Y, Yuan B, Okusumi S, Aoyama R, Murota R, Kikuchi H, Takagi N, Toyoda H. Enhanced cytotoxic effects of arsenite in combination with anthocyanidin compound, delphinidin, against a human leukemia cell line, HL-60. Chem Biol Interact. 2018;294:9–17. doi: 10.1016/j.cbi.2018.08.008. [DOI] [PubMed] [Google Scholar]
- 22.Springer TA. Adhesion receptors of the immune system. Nature. 1990;346:425–434. doi: 10.1038/346425a0. [DOI] [PubMed] [Google Scholar]
- 23.Budinsky AC, Brodowicz T, Wiltschke C, Czerwenka K, Michl I, Krainer M, Zielinski CC. Decreased expression of ICAM-1 and its induction by tumor necrosis factor on breast-cancer cells in vitro. Int J Cancer. 1997;71:1086–1090. doi: 10.1002/(sici)1097-0215(19970611)71:6<1086::aid-ijc27>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- 24.Ogawa Y, Hirakawa K, Nakata B, Fujihara T, Sawada T, Kato Y, Yoshikawa K, Sowa M. Expression of intercellular adhesion molecule-1 in invasive breast cancer reflects low growth potential, negative lymph node involvement, and good prognosis. Clin Cancer Res. 1998;4:31–36. [PubMed] [Google Scholar]
- 25.Baj G, Arnulfo A, Deaglio S, Mallone R, Vigone A, De Cesaris MG, Surico N, Malavasi F, Ferrero E. Arsenic trioxide and breast cancer: analysis of the apoptotic, differentiative and immunomodulatory effects. Breast Cancer Res Treat. 2002;73:61–73. doi: 10.1023/a:1015272401822. [DOI] [PubMed] [Google Scholar]
- 26.Baj G, Arnulfo A, Deaglio S, Tibaldi E, Surico N, Malavasi F. All-trans retinoic acid inhibits the growth of breast cancer cells by up-regulating ICAM-1 expression. J Biol Regul Homeost Agents. 1999;13:115–122. [PubMed] [Google Scholar]
- 27.Iriyama N, Yuan B, Yoshino Y, Hatta Y, Horikoshi A, Aizawa S, Takeuchi J, Toyoda H. Aquaporin 9, a promising predictor for the cytocidal effects of arsenic trioxide in acute promyelocytic leukemia cell lines and primary blasts. Oncol Rep. 2013;29:2362–2368. doi: 10.3892/or.2013.2388. [DOI] [PubMed] [Google Scholar]
- 28.Yuan B, Ohyama K, Takeichi M, Toyoda H. Direct contribution of inducible nitric oxide synthase expression to apoptosis induction in primary smooth chorion trophoblast cells of human fetal membrane tissues. Int J Biochem Cell Biol. 2009;41:1062–1069. doi: 10.1016/j.biocel.2008.09.031. [DOI] [PubMed] [Google Scholar]
- 29.Bacus SS, Gudkov AV, Zelnick CR, Chin D, Stern R, Stancovski I, Peles E, Ben-Baruch N, Farbstein H, Lupu R, et al. Neu differentiation factor (heregulin) induces expression of intercellular adhesion molecule 1: implications for mammary tumors. Cancer Res. 1993;53:5251–5261. [PubMed] [Google Scholar]
- 30.Constantinou AI, Krygier AE, Mehta RR. Genistein induces maturation of cultured human breast cancer cells and prevents tumor growth in nude mice. Am J Clin Nutr. 1998;68:1426s–1430s. doi: 10.1093/ajcn/68.6.1426S. [DOI] [PubMed] [Google Scholar]
- 31.Bacus SS, Kiguchi K, Chin D, King CR, Huberman E. Differentiation of cultured human breast cancer cells (AU-565 and MCF-7) associated with loss of cell surface HER-2/neu antigen. Mol Carcinog. 1990;3:350–362. doi: 10.1002/mc.2940030607. [DOI] [PubMed] [Google Scholar]
- 32.You H, Yu W, Sanders BG, Kline K. RRR-alpha-tocopheryl succinate induces MDA-MB-435 and MCF-7 human breast cancer cells to undergo differentiation. Cell Growth Differ. 2001;12:471–480. [PubMed] [Google Scholar]
- 33.Liu Y, Zou T, Wang S, Chen H, Su D, Fu X, Zhang Q, Kang X. Genistein-induced differentiation of breast cancer stem/progenitor cells through a paracrine mechanism. Int J Oncol. 2016;48:1063–1072. doi: 10.3892/ijo.2016.3351. [DOI] [PubMed] [Google Scholar]
- 34.Liu T, Men Q, Wu G, Yu C, Huang Z, Liu X, Li W. Tetrandrine induces autophagy and differentiation by activating ROS and Notch1 signaling in leukemia cells. Oncotarget. 2015;6:7992–8006. doi: 10.18632/oncotarget.3505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wu G, Liu T, Li H, Li Y, Li D, Li W. c-MYC and reactive oxygen species play roles in tetrandrine-induced leukemia differentiation. Cell Death Dis. 2018;9:473. doi: 10.1038/s41419-018-0498-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Carey L, Winer E, Viale G, Cameron D, Gianni L. Triple-negative breast cancer: disease entity or title of convenience? Nat Rev Clin Oncol. 2010;7:683–692. doi: 10.1038/nrclinonc.2010.154. [DOI] [PubMed] [Google Scholar]
- 37.Bacus SS, Stancovski I, Huberman E, Chin D, Hurwitz E, Mills GB, Ullrich A, Sela M, Yarden Y. Tumor-inhibitory monoclonal antibodies to the HER-2/Neu receptor induce differentiation of human breast cancer cells. Cancer Res. 1992;52:2580–2589. [PubMed] [Google Scholar]
- 38.Xu W, Meng K, Tu Y, Tanaka S, Onda K, Sugiyama K, Hirano T, Yamada H. Tetrandrine potentiates the glucocorticoid pharmacodynamics via inhibiting P-glycoprotein and mitogen-activated protein kinase in mitogen-activated human peripheral blood mononuclear cells. Eur J Pharmacol. 2017;807:102–108. doi: 10.1016/j.ejphar.2017.04.007. [DOI] [PubMed] [Google Scholar]
- 39.Maruyama T, Kono K, Mizukami Y, Kawaguchi Y, Mimura K, Watanabe M, Izawa S, Fujii H. Distribution of Th17 cells and FoxP3(+) regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer. Cancer Sci. 2010;101:1947–1954. doi: 10.1111/j.1349-7006.2010.01624.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.