

Abstract
The placenta plays a critical role in mammalian reproduction. Although it is a transient organ, its function is indispensable to communication between the mother and fetus, and supply of nutrients and oxygen to the growing fetus. During pregnancy, the placenta is vulnerable to various intrinsic and extrinsic conditions which can result in increased risk of fetal neurodevelopmental disorders as well as fetal death. The placenta controls the neuroendocrine secretion in the brain as a means of adaptive processes to safeguard the fetus from adverse programs, to optimize fetal development and other physiological changes necessary for reproductive success. Although a wealth of information is available on neuroendocrine functions in pregnancy, they are largely limited to the regulation of hypothalamus–pituitary–adrenal/gonad (HPA/ HPG) axis, particularly the oxytocin and prolactin system. There is a major gap in knowledge on systems-level functional interaction between the brain and placenta. In this review, we aim to outline the current state of knowledge about the brain-placental axis with description of the functional interactions between the placenta and the maternal and fetal brain. While describing the brain-placental interactions, a special emphasis has been given on the therapeutics and pharmacology of the placental receptors to neuroligands expressed in the brain during gestation. As a key feature of this review, we outline the prospects of integrated pharmacogenomics, single-cell sequencing and organ-on-chip systems to foster priority areas in this field of research. Finally, we remark on the application of precision genomics approaches to study the brain-placental axis in order to accelerate personalized medicine and therapeutics to treat placental and fetal brain disorders.
Keywords: pregnancy, neuroendocrinology, uterus, fetal brain, placenta, pharmacogenomics
Graphical Abstract
1. Introduction and overview of placenta
Pregnancy establishment and success involve complex cellular, molecular and physiological processes. The fertilized egg divides and develops into a blastocyst which then adheres and nidates into the endometrium. For implantation to occur, an implantation competent blastocyst must be superimposed on a receptive endometrium. Then a precise orchestration of apposition and attachment processes ensues invasion of the trophoblast cells to penetrate the endometrial epithelium and invade into the endometrial stroma [1-2]. Human placental development starts with attachment of the blastocyst to the endometrium epithelium on days 6 to 7 after conception. At this stage, the blastocyst consists of an outer single-layered epithelial cover and an inner cell mass. The trophoblast which is the outer layered epithelial of blastocyst acts as the direct precursor of the epithelial parts of the placenta which develops into a functional barrier between mother and fetus. The inner cell mass of the blastocyst contributes to the development of placental mesenchyme and the fetal vascular system.
The placenta develops to a fully functional fetomaternal organ in which the blastocyst that forms the fetus forms the fetal placenta (Chorion frondosum), and the maternal uterine tissue contributes to the formation of the maternal (Decidua basalis) component [3]. While maternal blood comes into direct contact with the fetal chorion, fetal blood (deoxygenated) comes in contact with the placenta via umbilical arteries. Besides metabolizing and transferring nutrients to the growing fetus, the placenta also functions as an organ to mediate the removal of waste products such as urea, uric acid, and creatinine from the fetus to the maternal blood. The placenta is the only organ that is formed by the interaction of the maternal and fetal tissues. Both the mouse and human have hemochorial placentation, the maternal blood comes in direct contact with the fetal chorion [4]. The main functional unit within the placenta is the labyrinth zone and the villus compartment in the mouse and human, respectively. As gestation advances, the placenta continues to develop and adapt to support fetal health and development. The adaptive ability of the placenta is evident from its function that ensures the development of the fetus in stressful conditions such as hypoxia and undernutrition [5]. Various physiologic stressors around conception and during pregnancy can manifest in pregnancy complications and miscarriage. Among pregnancy disorders associated with placental dysfunction, preeclampsia is distinctive because of its impact on both maternal and fetal well-being. Preeclampsia is associated with fetal growth restriction as well as damage to several organs. Several studies have suggested that secreted placental products, such as soluble VEGF receptor (sFlt) and placental growth factor (PlGF), have a role in preeclampsia progression [6].
The placenta also protects the fetus by preventing transmission of infectious diseases from the uterus. In addition, the placenta also acts as a pivotal endocrine organ to produce different hormones, monoamines, and steroids. Interaction of specific corticotropin-releasing hormones with oxytocin, neurosteroids, and prostaglandins influences activities during early as well as late in pregnancy. The brain not only plays a central role in controlling all aspects of homeostatic processes [7], but also functions with endocrine organs to regulate diverse physiological and metabolic processes relating to development, reproduction, aging and diseases [8]. The hypothalamus acts as the central part of the brain to orchestrate coordinated interaction with different endocrine systems: the hypothalamic–pituitary–adrenal axis (HPA), hypothalamic–pituitary–thyroid axis (HPT), and hypothalamic–pituitary–gonadal axis (HPG). They control various secretions involved in diverse physiological processes [9-10]. Dysregulation of neuroendocrine signaling during pregnancy and parturition can affect the activities of the placenta and fetal membranes during gestation [11]. Thus, understanding the functional links between the brain and reproductive system is essential to obtain better insights into the molecular complexity of pregnancy and to develop novel therapeutic targets to protect the developing fetus from environmental stressors.
2. Placenta and maternal brain connection
During pregnancy, the maternal brain undergoes temporal changes at different level. Pregnancy changes the maternal brain for at least two years following birth, including enhancement of key hippocampal functions in human [12-13]. Study also suggests that changes in hormone levels during late pregnancy and early postpartum influence specific cognitive abilities in women [14]. During pregnancy, many women experience varying symptoms relating to brain, often referred to as ‘pregnancy brain’, including forgetfulness, increased absentmindedness, and memory disturbances [15]. These findings have suggested that pregnancy may lead to temporal alternation in the maternal cognitive function and behavior in women but it is not known if those changes are dependent upon the gestational stages. In rodents, study shows that changes in the maternal brain during pregnancy regulate maternal behaviors such as pup retrieval and nest building in mice [16]. Furthermore, transcriptome analysis shows that the expression level of specific genes is altered in the maternal brain during pregnancy [17-18]. However, a direct functional link between placenta and maternal brain has not been demonstrated yet. Our recent studies based on gene expression data found evidence for functional links between the placenta and maternal brain [17]. Correlated expression of genes, including those encoding different ligands and receptors, have been identified between placenta and the maternal brain in that study. Similarly, it has been shown that placental dysfunction alters the gene expression in the maternal brain, and those changes have potential to affect maternal behavior [18].
2.1. Neuroendocrine signaling
The placenta forges a physical link between the mother and fetus. Though its primary function is to deliver nutrients and oxygen to the fetus as well as to remove waste products, placental functions are far more than we think. The placenta acts as an endocrine organ and secretes endocrine, paracrine and autocrine neuroactive signaling molecules [19] that regulate the function of other endocrine glands to effect maternal adaptation to pregnancy. Several neuroactive hormones, such as oxytocin, melatonin, serotonin, and thyrotropin-releasing hormone (TRH) target the maternal brain [20]. The oxytocin targets the maternal brain to stimulate maternal nursing behavior [21]. Serotonin and melatonin play major roles in determining the maternal mood and behavior during pregnancy and postpartum period [22]. TRH stimulates the release of thyrotropin hormone from the anterior pituitary gland. The excess amount of thyrotropin during pregnancy is critical to stimulate the secretion of the prolactin-like hormone [23].
The placenta produces different protein hormones (human chorionic gonadotropin, placental lactogen), growth factors (IGF-1, TGFB, EGF, VEGF), steroids (Estrogens, Progesterone), peptide hormones (CRH, GnRH, NPY, Kisspeptin, Oxytocin, adrenomedullin), and adipokines (Leptin, Adiponectin, Resistin, Ghrelin). In addition, different ligand-receptor pairs are coordinately expressed between the placenta and maternal brain in pregnant mice including the brain-derived neurotrophic factor (Bdnf) and its corresponding receptor (TrkB) [17]. BDNF stimulates the growth and survival of trophectoderm cells in the placenta and has a biologic role in placental response to adverse maternal effects such as obesity and fine particle air pollution [24]. Efna1 ligand gene is expressed in the placenta with Eph receptor genes such as Epha3, Epha4, Epha5, Epha6, Epha7, Epha8, Ephb1, and Ephb6 are expressed in the maternal brain [17]. Likewise, Efna3 and Efna5 ligand genes are expressed in the maternal brain, and their corresponding receptor (Eph1) is expressed in the placenta. The Eph receptors are protein-tyrosine kinases that have a role in both healthy placental development and placental pathologies, such as preeclampsia [25-26]. The RNA-seq data [17] showed a set of 23 pairs ligand genes and their cognate receptor genes which are coordinately regulated in the placenta relative to the maternal brain and fetal brain on gestation day 15 in mice (Figure 1). These ligands and receptors may be expressed in other tissues and physiological conditions. But there is no study to support if any other tissue regulates those genes in a coordinated manner as seen between the placenta and brain during pregnancy. Based on hierarchical cluster analysis of gene expression level (accession# GSE121799), the receptors in the placenta are expressed in more closely manner with the fetal brain than the maternal brain. On the other hand, the expression of ligands is highly variable and also distinct in the placenta relative to both the maternal and fetal brain. This supports the idea that differential regulation of ligand and receptor genes between placenta and brain may be involved in placental communication with the mother and fetus.
Figure 1.

A) Expression of ligand and receptor pairs between placenta and brain in mice. The ligands are shown as ‘O’ and receptors as ‘)’. B) The expression of placental (P) expression relative to the maternal brain (MB) and fetal brain (FB) is shown.
The differentially regulated genes between the placenta and maternal brain are associated with regulation of immune system process, neuron development and differentiation, cell surface receptor signaling, and metabolic regulation [17]. A mouse model [18] was developed to study placental function by crossing the house mouse and the Algerian mouse. The maternal brain of house mouse females carrying hybrid litters after mating with Algerian mouse males showed a similar gene expression (Drd3 and placenta-specific gene families) to that of Algerian mouse females. Also, significant changes in the gene expression of Formin 1, Dopamine receptor 3 (Drd3), Calneuron 1 and Cathepsin R was observed in the maternal brain [18]. These findings have demonstrated the influence of the placental genome on maternal brain and have confirmed the interplay between the placenta and maternal brain.
2.2. Balancing the energy equation
Placental adaptation and positive energy balance are two major mechanisms in pregnancy that ensure energy requirement to meet the enhanced physiological and metabolic activities in gestating mothers. Placental adaptation includes structural changes such as enlargement of the labyrinth exchange zone and increase in the rate of nutrient transportation [27-29]. Increase in food intake and nutrient absorption as well as depositing fat in the body is used as means to generate a positive energy balance during late pregnancy for optimal fetal growth and also during lactation [30-31]. How the normal homeostatic mechanisms are modified in pregnant females is still largely unknown. But, mitochondria function plays a major role in energy metabolism and other homeostatic requirements by the placenta and brain during pregnancy [32-33]. In that regard, it has been shown that mitochondrial gene expression in placenta is significantly associated with different maternal psychological stresses during pregnancy [34]. Besides the role of mitochondria in placental function, mitochondria also play a role in the aging of brain [35]. It has also been shown that lifetime stress is associated with the copy number of mitochondrial DNA (mtDNA) in the placenta [36].
The placenta is also involved in the pregnancy-induced resistance to the anorectic actions of leptin. Placental lactogen and prolactin play an important role to regulate this resistance during pregnancy. Pseudopregnant rats exhibit prolactin surges at early pregnancy, but they lack the chronic increase of placental lactogen [37]. Although food intake is increased in pseudopregnant animals due to the effect of early prolactin surges, they are able to develop a leptin-induced anorectic effect. Importantly, chronic intracerebroventricular infusion of prolactin in these pseudopregnant rats results in acquiring the resistance to leptin-induced decrease in food intake [38]. This evidence demonstrates that the placenta plays a key role in balancing the energy equation through its communication with the maternal brain.
2.3. Uterine influence on the brain-placental axis
Conditional knockout of Foxa2 in the mouse uterus altered gene expression of the placenta on gestation day 15 suggesting uterine influence on the brain-placental axis [17]. Thus, it is imperative to discuss different uterine changes during pregnancy, and relate those to the placental development and function. The uterus is a complex organ consisting of three major tissue compartments (epithelium, stroma, and myometrium) and two epithelial cell types; luminal epithelium (LE) and glandular epithelium (GE) (1-3). Dynamic and cell-type specific actions of ovarian estrogen (E2) and progesterone (P4) regulate endometrial function, blastocyst implantation, stromal cell decidualization and placentation. In humans and rodents, the establishment of pregnancy requires different processes in the uterus such as ultra-structural changes of the LE and GE, secretory transformation of the uterine glands, differentiation of the stroma into decidual cells, recruitment of specialized immune cells, vascular remodeling, and placenta development. Insufficiencies in any of these processes may compromise pregnancy and impact fetal-maternal health [39-40].
In mice, the initiation of the attachment reaction, occurring between 2200-2300 h on day 4 of pregnancy, is followed by extensive stromal cell proliferation surrounding the implanting blastocyst which is evident by the morning of day 5. The epithelial cells surrounding the implanting blastocysts are removed through the process of entosis and apoptosis beginning on the night of day 5. Between day 5 and 6 of pregnancy, stromal cells, adjacent to the implanted blastocyst at the antimesometrial pole of the uterus, cease proliferating and begin differentiation. These cells form the primary decidual zone (PDZ), an area that is avascular and epithelioid (40-42). Stromal cells bordering the PDZ continue to proliferate and differentiate into cells forming the secondary decidual zone (SDZ), which are polyploid and located in the antimesometrial pole [43-44]. Placental and embryonic growth begin to replace the SDZ after day 8 of pregnancy, reducing it to a thin layer of cells referred to as the decidua capsularis. The decidual cells formed on the mesometrial side of the uterus are termed the decidua basalis and remain throughout pregnancy [43-44]
Although many of the molecular mechanisms involved in the processes of establishment of a successful pregnancy are similar in the human compared to the mouse, many noteworthy differences are observed. In human, nidation of the blastocyst begins about day 7 and is complete by days 10 to 12 following fertilization [45-47]. As the trophectoderm of the blastocyst begins to attach and adhere to the epithelium, the cytotrophoblast begins to differentiate and form a syncytiotrophoblast that then penetrates between the LE cells into the stroma. The implanted conceptus becomes encapsulated within the superficial endometrium and is surrounded by decidualizing stromal cells, providing a conducive environment for placental development [45, 48].
2.4. Coping with external conditions
In pregnancy, high levels of chronic stress may cause health problems to both the mother and fetus. It has been shown in cattle that heat stress during pregnancy can perturb endocrine dynamics and decreases the birth weight of calves [49]. In sheep, the increase of maternal stress hormones during pregnancy elevates postnatal blood pressure [50]. In human, maternal stress during pregnancy increases the risk of neurodevelopmental disorders such as autism spectrum disorders, hyperactivity disorder and schizophrenia [51-55]. Normally, such harmful impacts of pregnancy-associated stress are responded by activation of an allostatic mechanism that mitigate neuroendocrine stress responses [56]. Although the placenta could be a target of maternal stress, its neuroendocrine function plays a role in reducing the responsiveness of maternal HPA axis to acute stressors [57]. The placental secretions help the mother to prepare, accommodate and provide the care for the fetus against stress [20]. Prolactin-like protein A (Plpa) is produced by the chorioallantoic placenta. In contrast to wild-type mice, which can adapt hypoxic stress and maintain pregnancy, Plpa knockout mice are sensitive to hypoxic stress. The placenta of Plpa null mutant mice is underdeveloped and show aberrant vasculature with subsequent inability to support and maintain pregnancy [58]. These findings have shown that the placenta is not an isolated organ to provide the contact between the mother and her fetus, but it acts at different levels to ensure the ability of the mother to support pregnancy and to provide the care for the newborn.
2.5. Relationship between parity and the maternal brain
Studies have also shown that parity may have a functional link with learning or memory of brain during pregnancy, possibly due to the prior steroid exposure during pregnancy [59]. Paris and Frye 2008 [59] assessed cognitive function of mice brain to object placement and recognition tasks between nulliparous (no prior pregnancy) and multiparous (with previous pregnancies) rats, and found that multiparous animals performed better in object placement and recognition tasks compared to rats that were never pregnant. This finding suggested that parity does influence learning and memory of pregnant females, and such behaviors are further found to be correlated with a lower corticosterone but higher estrogen levels in the multiparous relative nulliparous rats in the same study. A review of data from rodent and human mothers suggested that reproductive experience may influence cognition and anxiety in a different manner than primiparous or nulliparous females [60]. This review suggested that reproductive experience may benefit pregnant women to control the anxiety/stress response and enhance certain aspects of memory similar to results observed from rodent studies. Although the underlying mechanism of how parity influences cognitive functions in response to pregnancy remains poorly understood, it has been suggested that changes in the hippocampal neurogenesis during the peripartum period may have a functional link to the parity vs. cognition relationship [61].
3. Placenta and the fetal brain connection
Signaling plays a key role in regulating placental function to the communication between mother and fetus. The classical insulin-like growth factor (IGF) system is a well-studied example of placental role in maternal-fetal communication [62]. Also, the placenta contains a unique microbiome [63] that may have functional roles in pregnancy outcomes. However, several studies have demonstrated that placental regulation of Igf2, Cortisol, Serotonin, and T4/T3 influences neurodevelopmental processes of the fetus. In mice, knockout of Igf2-P0 transcript in the placenta shows intrauterine growth restriction (IUGR) leading to dysregulated placental delivery of nutrients to the fetus, and anxiogenic effects in the offspring [64]. Evidences further suggest that placental control of OGT (O-linked n-acetylglucosamine transferase) influences fetal development in a sex-specific manner [65-66]. In addition, expression of Ppara, Igfbp1, Hifa, and Glut4 in the placenta influences fetal programming in a sex-specific manner [67]. Similarly, placental control of proinflammatory cytokines (Il6 and Il1b) has been shown to alter the neural expression of dopamine D1 and D2 receptors in brain [68]. Studies have also shown that dysregulation of the triiodothyronine (T3) and thyroxine (T4) hormones in the thyroid gland and serotonin in the brain can cause neurodevelopmental defects in the growing fetus [69-72].
3.1. Development of the fetal brain
A fundamental question in biology is how a brain develops. It is known that the central nervous system (CNS) arises from the ectodermal tissue called the neural tube that differentiates into the spinal cord and the brain. Neural tube genesis occurs at gestation day (gd) 9-9.5 in mice that has a gestational length of 19-21 days. In human, the process starts between gd 24-28 where the length of gestation is 266-280 days [73-74]. Development of CNS is a highly coordinated spatiotemporal process that includes the proliferation of glia and neurons and their migration, followed by programmed cell death, formation of synapses, myelination, and establishment of neuronal circuits. There is a remarkable similarity among the rodents and humans in the process of cortical development during fetal brain formation [75] given the fact that the placenta of human and rodents are both hemochorial in nature [76]. Any perturbation during the brain development leads to immediate or postnatal adult disease consequence, collectively known as Developmental Origin of Health and Diseases (DOHaD). Rest (repressor element 1 silencing transcription factor) is a key transcription factor that acts as a master regulator of neurogenesis [77]. In our earlier study [17], we conducted iRegulon [78] analysis to predict transcription factors that control correlated up-regulation or down-regulation of genes in the fetal brain relative to the placenta in mice on day 15 of pregnancy. The analysis showed that Rest is a key regulator of gene expression crosstalk between the placenta and fetal brain. Determining role of Rest in the genome regulation of placenta and developing fetal brain can provide new information if placenta plays a role in neurogenesis during brain formation of the fetus.
3.2. Infections
Placenta has been traditionally believed to a protective organ and its environment had been considered as sterile. Now it is well established that placenta has its own microbiome [63]. The placenta, fetal membranes, and fetus exist in close proximity to several microbial organisms of the lower urogenital tract. In normal pregnancy, the placenta harbors nonpathological commensal microbiota and variation in this composition can be associated with pregnancy disorders [79]. Study suggests that the placenta can acquire the infection not only from the urogenital route but from the oral route through the hematogenous access [80]. Pathogens can either directly cross the placental barrier and colonize the fetus, or they can elicit an inflammatory response in the placenta [81]. Infections activate inflammatory pathways, causing the release of various proinflammatory biomarkers such as cytokines, interleukins, and other molecules. The inflammatory signals passing through the placenta from the maternal side can have adverse effects on the fetal brain development. Such insults result in both physical and behavioral phenotypes in juvenile, adult or aged offspring. Well recognized neurodevelopmental disorders like schizophrenia and autism have been associated with maternal infection [82-82]. The location and the timing of the insult due to infections have an important role in cerebral development and, ultimately, function [84]. The effects of such prenatal insults underlie some cognitive, behavioral, and psychiatric disorders [85]. Viral infections that are capable of crossing the placental barrier and reaching the fetus can have detrimental effects on fetal development. Zika virus infection outbreaks in pregnant mothers caused microencephaly in the fetuses [86]. The effect on the fetal brain is due to the vertical transmission of the virus via transplacental route. Similarly, prenatal infection with human influenza virus is known to cause long-term structural and functional changes in the pup brains [87]. Cytokines and inflammatory cells are thought to regulate many common pathways associated with perinatal brain injury due to infection. Inflammatory activation of microglia (brain macrophages) attack and injure white matter oligodendrocyte progenitor cells (pre-oligodendrocytes) and derail their differentiation process during the brain development process [88]. These effects on the fetal brain ultimately can result in psychologic and cognitive disease in later life.
3.3. Chorioamnionitis
Chorioamnionitis is a pathological condition characterized by the presence of acute inflammatory infiltrate in membranes and chorion of the placenta. It can be triggered either by the infection of the placenta and its membranes or by the activated maternal immune system. In early human gestation, prior to the 20th week of gestation, the leukocytes are mainly of maternal origin. By mid-trimester, the fetus begins to be capable of producing leukocytes that participate in the inflammatory response. In the presence of infectious agents, chorioamnion elicits inflammatory response characterized by the release of a combination of proinflammatory and inhibitory cytokines and chemokines in the maternal and fetal compartments [89]. Cytokines can activate the HPA axis resulting in an increased level of glucocorticoid in the peripheral bloodstream that can permanently modify the brain response to stress [84]. Chorioamnionitis is the leading cause of preterm birth and neonatal complications and is the most frequent cause of fetal death in the second half of pregnancy [90]. Preterm birth affects around 10% of deliveries and is a leading cause of neurodevelopmental impairment. Chorioamnionitis affects around 40–80% of preterm deliveries and it can initiate a fetal inflammatory response that is injurious to the developing brain and other organs. Epidemiological evidence has indicated the association of several adult neurodegenerative diseases to the immune activation in mother’s womb during pregnancy [91]. Inflammatory signals increase the permeability of blood brain barrier and expose the brain to the cytotoxic protein. Factors that lead to these effects include epigenetic modification of the stem cell population, microglial activation, oligodendrocyte dysmaturation, reactive oxygen and nitrogen species accumulation, sensitization of the brain to hypoxia-ischemia and the activation of coagulation cascade. The stage of pregnancy with distinct phase of the neonatal brain development can have differential neurodevelopmental vulnerability depending upon the timing of onset of chorioamnionitis [84, 92].
3.4. Drug transfer and xenobiotics
The development of the full-grown fetus from a fertilized egg occurs through the process of lineage-specific cell growth and differentiation. This process occurs at different rates which is dependent on species and gestation length. This energy dependent process depends on the placenta to extract the nutrients from the mother’s circulation. In the placenta, the fetal cells are in close proximity to the maternal cells to facilitate this transfer. In humans, the fetoplacental-maternal circulation is established around the tenth week of pregnancy [93]. Transfer of nutrients across the placenta can occur by simple diffusion, facilitated diffusion, active transport and receptor-mediated endocytosis. The transport process applies equally to drugs and xenobiotics. The placenta is considered to have a barrier function to protect the developing fetus. But most drugs cross the placenta. Nonpolar fat-soluble drugs that have low molecular weight with non-protein binding properties can cross placenta easily and quickly. Alcohol is one example. The placenta expresses a variety of xenobiotic-metabolizing enzymes but it can also catalyze the formation of reactive metabolites that are toxic to the fetus [94].
Over the years, the pharmaceutical intervention has increased to combat pregnancy complications arising from the intake of different prescription and non-prescription drugs by the pregnant mothers. Most widely used types of drugs are diuretics, antihistaminics, entiemetics, antibacterials, tranquilizers and analgesics [95]. Substance abuses are increasing concerns of the modern lifestyle that puts the fetal development at risk. Alcohol and nicotine can produce severe deficiencies in brain development. Nicotine binds to nicotinic acetylcholine receptors, ligand-gated ion channels that are widely expressed in the fetal nervous system [96]. Smoking exposure during pregnancy can lead to decreased birthweight, increase risk of preterm birth and have teratogenic effect on the fetus. Animal studies have shown that fetal brain nicotine level reaches much greater than maternal blood levels, and the exposure can lead to long-term disruption of brain architecture and chemistry [97-98]. Pharmacologically, alcohol is a depressant that blocks N-methyl-D-aspartate receptor activity and increases GABAergic activity. There is overwhelming basic and clinical evidence regarding the negative neurodevelopmental consequences of prenatal alcohol exposure broadly known as fetal alcohol syndrome (FAS), which is characterized by growth deficiencies, craniofacial dysmorphologies and CNS damage [99]. FAS is recognized as a mental disorder by the International Classification of Diseases and the Diagnostic and Statistical Manual of Mental Disorders. With the modern lifestyle, there are increasing cases of psychiatric disorders. Antipsychotic medications like dopamine and serotonin receptor antagonists are prescribed in such cases. It is now clear that placenta produces serotonin that plays an important function in fetal brain development [100]. Further studies on understanding the long-term functional implication of such antipsychotic drugs and substance abuses in fetal brain development are needed for policy formulation to safeguard pregnancies.
4. Animal models and clinical studies
Since the advent of the Barker Hypothesis in 1990, which posits that the intrauterine environment has direct and long-lasting effects on the fetus [101], much attention has been directed towards researching the fetal origins of adult disease. In this pursuit, the brain-placental axis is being dissected to understand the association between the placenta and the fetal brain and more recently the association of the placenta with the maternal brain. Animal models have proven invaluable for studying pregnancy and in particular the interaction between the placenta and the maternal/ fetal brain.
4.1. Basic biology
Animal models of placenta have provided valuable information about its role as a multifunctional organ for fetal growth and survival. However, no perfect model of the human placenta exists. Although obvious anatomical differences exist between mice and humans, at the basic level both species form a hemochorial, discoid placenta during pregnancy. The hemochorial placenta represents the most intimate association between the fetal cells and the maternal circulation and as such is also the most invasive form of placentation [102]. In both mouse and human, the trophoblast cells undergo a series of proliferation and differentiation events and invade the uterine decidua followed by the maternal vasculature. These specialized trophoblast cells replace the maternal endothelium creating a trophoblast lined conduit to funnel maternal blood into the placenta. In humans, the placental villi are analogous to the labyrinth, and also contain syncytiotrophoblast cells that are in direct contact with the maternal blood [103-104]. Although differences exist, such as the presence of the junctional zone in the mouse placenta which is absent in the human, analogies can be drawn between mouse and human in placental cell types and genes controlling placental development. In a comparison of near-term placentae, over 7000 ortholog genes were detected between mouse and human with 70% being expressed in both species [105]. Interestingly out of 170 genes known to cause placental abnormalities in mice, 138 (81%) were expressed in human placentae supporting the idea that such candidate genes can be evaluated in mice as models as a means to study human placental disorders.
4.2. Fetal health
Maternal environment is one of the major factors that determine fetal health. Mother’s weight, nutritional state, anemia, smoking, substance abuse, and uterine blood flow are some of the conditions that can have detrimental effects on fetal growth. Preterm birth is the most common cause of infant deaths throughout the world [106]. It is estimated that approximately 15 million babies are delivered as preterm. Placental malperfusion and functional insufficiency, among others, is a leading cause of preterm birth [107]. Development of the blood-brain barrier and regulation of barrier transport systems contribute to protecting the fetal brain from exposure of pregnant mothers to harmful drugs. Our understanding of how intrauterine conditions influence the development of fetal brain remains poor. Understanding the systems-level regulation of maternal effects on fetal brain development is important as it would make a significant impact on studying fetal health, and brain disorders in the offspring later in life.
Because the mouse is a genetically tractable model system, it has provided great insight into placental function and its influence on fetal development. In particular, much attention has been directed towards understanding how inadequate invasion and vascular remodeling by trophoblast cells results in impaired placental blood flow and leads to pregnancy complications such as preeclampsia and IUGR [108]. IUGR is a pathological condition in which the fetus fails to attain its full genetic growth potential. Several genes have been linked to the invasive phenotype of trophoblast cells, such as Htra1 and Notch signaling members. Using a mouse model, loss of Htra1 resulted in decreased numbers and differentiation of the precursors of invasive trophoblast cells, decreased invasion and defective vascular remodeling leading to an IUGR phenotype [109]. Members of the Notch signaling pathway were shown to be important for trophoblast invasion and vascular remodeling in mice and humans, and their perturbation was associated with placental defects [110].
Fetuses impacted by IUGR suffer from immediate as well as long-term physiological complications. An IUGR infant has an increased risk of adverse health outcomes including poor neurological development and postnatal growth, immune deficiencies and adulthood risk of cardiovascular disease, diabetes mellitus and hyperinsulinemia, among others [111]. It is well established that placental defects/insufficiency, such as those observed in IUGR, can impact fetal brain development [71,112]. Additional animal models using guinea pigs, rats and sheep have provided valuable information on IUGR impacts on neurodevelopment which show varying degrees of “brain-sparing” and other similarities to human IUGR [113]. Interestingly, in an attempt to protect the brain, the fetus will adapt its circulation to preserve oxygen and nutrient supply to the brain (brain-sparing) [114] and will alter neuro and placental gene expression in favor of the brain [115] at the cost of the placenta. However, some have indicated that brain-sparing is associated with worse neurological outcomes than IUGR babies without brain-sparing [114]. Perhaps it is a matter of duration of brain-sparing, with prolonged brain-sparing resulting in further placental complications, which further impact the fetus. The placenta can convert maternal tryptophan into serotonin and serves as the primary source of serotonin for the developing fetal forebrain [116]. Several ligand-receptor interactions have been predicted between the placenta and fetal/maternal brain from gene expression data [17] some of which included Itgb8, Itgav, Notch1, Rspo3, and Vcam1 which were previously implicated in placental defects [3, 17]. Together, this research demonstrates the need to expand our view of the role of the placenta from a barrier and transport organ to one that is actively involved in synthesizing factors important for neurodevelopment and possibly beyond.
4.3. Therapeutics
Placental insufficiencies such as those seen in IUGR are multifactorial conditions with an array of fetal, neonatal and adult consequences. Therefore, different treatments are need to cure the variable clinical symptoms. In regards to neurological outcomes, interventions include taurine and melatonin supplementation, Newborn Individualized Developmental and Assessment Program System (NIDCAP) and targeted medical rehabilitation [111]. Some research suggests that exogenous supplementation of growth hormone (GH) may improve neuro deficits of IUGR children. However, the data thus far is scant and contradictory [117]. It is evident that research on mechanisms of impaired neurodevelopment in instances of placental insufficiency and possible treatments is greatly needed. The expression of different ephrin and integrin receptors in the placenta are strikingly correlated with the expression of their ligands in the fetal brain [17], Efna1 is expressed in the placenta with Eph receptors (Epha3, Epha4, Epha5, Epha6, Epha7, Epha8, Ephb1, and Ephb6) expressed in both the maternal and fetal brain. On the other hand, Efna3 and Efna5 are expressed in the brain (both maternal and fetal) with their receptor (Eph1) expressed in the placenta [17]. Integrins are key regulators of angiogenesis process that is required during pregnancy for the development of new blood vessel and maternal-fetal communication [118]. Integrins and ephrins are currently targeted for several therapeutics for human diseases [119-120]. In humans, at least three integrins (out of 24 known) based drugs are currently available to cure diseases including multiple sclerosis and Crohn’s disease. Similarly, ephrins are involved in several pathological conditions including neurological disorders and viral infections for which they are currently investigated for potential drug targets [119].
Opioid addiction is a major ongoing crisis in many parts of the world including the United States [121]. Exposure to opioids during pregnancy can influence the uterine environment causing different negative effects such as IUGR, placental abruption, preterm delivery, oligohydramnios, stillbirth, and neonatal abstinence syndrome (NAS) in human. NAS is a serious condition characterized by central nervous system hyperirritability and autonomic nervous system dysfunction [122-123]. The regulation of opioid receptors in uterus, placenta, and brain may influence these clinical outcomes. In mice, the three receptors (mu, kappa and delta) for opioid are absent in the placenta but their level of expression is significantly low in the fetal brain (mean read count = 34) compared to the maternal brain (mean read count = 422) at gestation day 15 [17]. It is likely that opioid receptors are tightly regulated by the placenta to suppress their expression in fetal brain but not in the maternal brain, possibly to safeguard the fetus from that harmful exposure during pregnancy. It will be thus worth investigating how opioid receptors are regulated to influence the brain-placental axis function. Though animal models for opioid addiction are known [124-126], to our knowledge no investigation has been conducted to identify factors that regulate opioid receptors in the maternal brain. The discovery of those factors can help develop drugs to cure opioid addiction in pregnant women.
5. Future prospects and research avenues
Understanding the architecture and hormonal links between the placenta and brain requires innovative experiments using both top-down and bottom-up approaches. Though these approaches have been used in research relating to animal physiology and metabolism [127], not much research has been conducted using these methods in the field of neuroendocrinology. An example of the top-down approach will be the establishment of genetic alteration (such as conditional knockout models) or physiological manipulation of maternal condition (nutritional and external stimuli) followed by performing assays that can identify changes in the physiological or transcriptional interaction between the brain and placenta. On the other hand, a bottom-up approach will utilize known etiological differences in placental and/or brain disorder during pregnancy, and then underpinning the genes or hormones significantly associated with those phenotypes. We believe that these approaches can be leveraged in systems biology studies by integrating contemporary research tools such as organoids, organ-on-chips and pharmacogenomics towards developing personalized therapeutics to pregnancy associated diseases.
5.1. Prospects of applying of organoids and organ-on-chips in studying placenta and fetal brain
Designing research experiments with human reproductive organs poses both technical as well as ethical challenges. In this regard, the use of cell and organoid culture methods can offer an alternative solution to study the structure and function of the placental-brain axis. Historically, cell culture has been used extensively for understanding how specific hormones such as progesterone and estradiol stimulate the proliferation of decidual cells and trophoblastic cells [128]. Culturing of endometrial stromal cells and placenta-derived stem cells have been successfully demonstrated [129-133]. The colony-forming technique, in vivo transplantation, label retention assays or three-dimensional culture systems have been applied to study stem/progenitor cells of uterus, but the organoid culture of uterine cells have been recognized as a promising technique to study uterine cell biology and identify factors for development of therapeutics [134-138]. These methods rely on growing stem cells using specialized media and culture conditions that promote self-organization of the cells through sorting them into multicellular three dimensional structures [139]. The success of these methods depends on the functionality of the resulting organoids to mimic that of the organ or tissue from which they were derived [140] (Figure 2). Organoid systems have been used in recent studies to investigate placental development and trophoblast interactions with maternal environment [141-142]. The protocol of developing placental organoids is described in details in a recent publication [143].
Figure 2.

Organoids generated in our lab from the epithelia of mouse endometrium. A) Hematoxylin and eosin staining of longitudinal section showing (arrows) the glandular epithelium (GE) and luminal epithelium (LE) of mouse endometrium. B) The 3D structures representing organoids of epithelial cells using matrigel based recombinant protein cocktail culture conditions.
A limitation of the organoid systems is the requirement to optimize specific culture protocol so that the 3D structure recapitulates the key features of structure and physiological functions of the organ. In this regard, organ-on-chip offers an alternative solution to utilize microfluidic and microfabrication technologies to develop models that resemble the structure and function of the organ. They offer new avenues to study the development of human organs and aid our ability to manipulate conditions to study physiological interactions between organs [144]. Soft lithography techniques were used to fabricate a placenta-on-chip that mimicked the architecture and function of the placenta [145]. The fabricated chip was able to maintain and proliferate JEG-3 trophoblasts and human umbilical vein endothelial cells (HUVEC) on an extracellular matrix scaffold and also recapitulate placental barrier by measuring glucose transport that was consistent with findings from in vivo studies. Furthermore, microfluidic chips in combination with cell co-culture technique was used to study physiological mechanism of placental barrier [146]. While the co-culture technique induced fusion of trophoblast cells to form a syncytialized epithelium, the microfluidic chip allowed to form dense microvilli and to replicate expression and physiological localization of membrane transport proteins of the placenta. Similar to the placenta-on-chip, researchers have also made progress in the development of brain organoids [147] and brain-on-a-chip systems [148-149]. Brain organoids have been developed to study polarized neuroepithelium, cell type heterogeneity and segregation of discrete brain regions during development of the fetal brain [150]. An integrated approach has been employed in some studies where first the brain organoids were generated and then they were used to fabricate the organoid-on-chip to recapitulate the microenvironment of the cells [151-152]. This method has been recently applied to a study where the brain organoids were exposed to nicotine to study neuronal regulation in response to prenatal exposure. Using this approach, regionalization and cortical development were found to be disrupted in the nicotine-treated brain organoids [153]. Such integrative approaches of developing placenta and brain organoids and organ-on-chips technologies have potential to study the dynamic changes that occur between the placenta and the brain throughout gestation. However, as discussed above, these approaches need extensive experiments to manipulate and standardize the co-culture conditions to recapitulate the physiological interactions between brain and placenta during pregnancy.
Organoid and organ-on-chip systems hold promises to the development of novel therapeutics and clinical interventions [154]. Recent efforts on application of organ-on-chip systems in drug target identification, target-based screening, and phenotypic screening have shown progresses in pre-clinical studies [155]. A noticeable advance in the application of bioengineered models of the placenta has focused on studying drug efflux via transporters across the human placental barrier [156]. Similarly, organ-on-chips are used to study the fetal membrane microenvironment which is relevant to research on preterm births [157]. One of the emphases of this research is to investigate how chorioamnionitis, which is linked to about 70% of preterm birth cases, may dysregulate the premature rupture of the fetal membranes that extend from the placenta and surround the developing fetus. Several studies have shown that abnormal function of fetal brain preludes pre-term births [158-159], and it is expected that such bioengineered chip models can reveal a better insight into the placenta and fetal brain links in preterm births.
5.2. Precision genomics and personalized medicine for placental and fetal disorders
Precision genomics is poised to bring a new dimension to the integration of genomics and medicine to human health care. Pregnancy is a major ongoing emphasis in this area where precision pharmacogenomics approach is increasingly used to design individualized drug treatments to pregnancy disorders [160-161]. Precision analytics of next-generation sequencing data of genes coding for hormones, neuroligands, and their receptors can help identify new drug targets for diseases relating to placental and fetal developmental abnormalities. Precision pharmacogenomics can provide valuable data towards developing personalized medicine for opioid pain management, antihypertensive and antidepressant drug, preterm labor tocolytics, and antenatal corticosteroids. The pharmacokinetics and pharmacodynamics of different drugs may vary from person to person during pregnancy for which precision genomics can be highly relevant to develop individualized medications [160]. For example, genetic variation in cytochrome P450 (CYP) genes, specifically CYP2D6, CYP2C19 and CYP2C9, predominantly account for variability in drug metabolism [162]. Enzymes produced by these three genes metabolize nearly 80% of currently available pharmaceutical drugs, and this variation in drug metabolism can be further confounded by changes in hormones produced during pregnancy [163-164]. Similarly, significant association is known between polymorphisms in serotonin transporter and differential response to antidepressant drugs [165]. To date, most emphases have been on developing safe, sensitive and non-invasive genetics and genomics assays for prenatal diagnosis [166], and recent advent in next-generation sequencing have paved new prospects in sequencing fetal genomes in a routine manner [167]. Recent advances in single-cell sequencing have shown great promise to delineate placental structure and function in more precise manner than before [168-170]. Though personalized fetal medicine is merely a concept at this time, given the rapid progress in pharmacogenomics and predictive analytics [171-173] development of personalized medicine to treat placental and fetal disorders is a realistic possibility in the years to come.
5.3. Changes in both the fetal brain and maternal brain
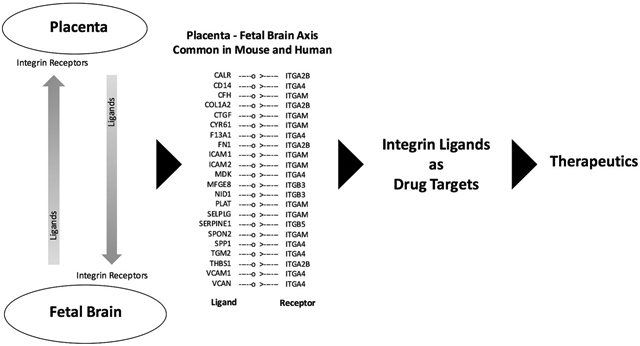
Understanding the physiological changes in both the maternal and fetal brain is critical to better understand the dynamic changes of pregnancy particularly in response to the placental ageing and function. Unfortunately, this area is poorly investigated and the literature lacks data on this topic. However, identifying cells in both maternal and fetal brain that concomitantly respond to physiological changes in the placenta can provide new insights in this regard. Recently, the single-cell RNA sequencing (scRNA-seq) technology has demonstrated unprecedented power and precision to underpin ligands and receptors in the maternal-fetal interface [174]. These ligand-receptor pairs are available at www.CellPhoneDB.org database that is expected to provide valuable resource to identify drug targets to specific interactions between decidual natural killer cells and the fetal extravillous trophoblast cells. A recent study [175] employed scRNA-seq approach to study the gene expression and differentiation pattern of different cell types of the human placenta. Single cell gene expression data was also generated to compare fetal brain of human and mouse that provided new information on molecular conservation and diversity of brain development between species [176]. In a separate study, single cells were analyzed from different mice tissues including placenta, fetal brain and adult brain to study their gene expression profiles [177]. These single cell data of human and mouse placenta, fetal brain and adult brain [publicly available at Gene Expression Omnibus database under the accession numbers GSE89497 (human placenta), GSM2906465 and GSM2906466 (mouse placenta), GSE76381 (human brain), and GSM2906405, GSM2906406, GSM2906415, GSM2906454, and GSM2906455 (mouse brain)] provide a good resource to mine for genes coding ligands and receptors that are canonically expressed between placenta and fetal/ adult brain. Employing an integrative analysis approach of scRNA-seq data based on canonical correlation method [178], our analysis of these datasets reveal that specific integrins and their ligands (see the list in the graphical abstract of this paper) are expressed in canonically correlated manner between placenta and brain both in the mouse and human. Integrin ligands hold promise in drug discovery and therapeutics for different human diseases [118-120], and may provide new avenues to investigate their application to develop therapeutics to placental and brain disorders relating to pregnancy.
6. Concluding remarks
Our current state of knowledge in placental and fetal biology, in general, requires new paradigms to pursue. Though a great deal of achievement has been made in advancing the field, we have not yet succeeded in curing placental insufficiency, hypoxia, preterm birth and fetal brain defects and other birth defects. Research needs to incorporate concepts and techniques from diverse areas including reproductive biology, systems biology, pharmacology, neuroendocrinology and chemical biology to understand factors regulating the placental development and the brain-placental axis. Though a great deal of information is available on diverse aspects relating to pregnancy establishment and success, studies on genetic underpinning of maternal factors that influence fetal brain remains limited. Integrative pharmacogenomics and single-cell analyses hold huge promise in this aspect to unravel the specific maternal-fetal communications and to identify new drug targets and therapeutics for pregnancy-associated diseases.
Acknowledgements
This work was supported in parts by grant from the University of Missouri (Research Council Grant) to S.K.B and R21 HD076347 and R01 HD096266 from the Eunice Kennedy Shriver National Institute of Child Health and Development to T.E.S.
Footnotes
Conflict of interest
The authors declare no conflict of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Su RW, Fazleabas AT. Implantation and Establishment of Pregnancy in Human and Nonhuman Primates. Adv Anat Embryol Cell Biol. 2015; 216:189–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kelleher AM, DeMayo FJ, Spencer TE. Uterine Glands: Developmental Biology and Functional Roles in Pregnancy. Endocr Rev. 2019; pii: er.2018–00281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chavatte-Palmer P, Guillomot M. Comparative implantation and placentation. Gynecol Obstet Invest. 2007; 64:166–174. [DOI] [PubMed] [Google Scholar]
- 4.Chavatte-Palmer P, Tarrade A. Placentation in different mammalian species. Ann Endocrinol (Paris). 2016; 77:67–74. [DOI] [PubMed] [Google Scholar]
- 5.Gude NM, Roberts CT, Kalionis B, King RG. Growth and function of the normal human placenta. Thromb Res. 2004; 114:397–407. [DOI] [PubMed] [Google Scholar]
- 6.Chau K, Hennessy A, Makris A. Placental growth factor and pre-eclampsia. J Hum Hypertens. 2017; 31:782–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Roh E, Song DK, Kim MS. Emerging role of the brain in the homeostatic regulation of energy and glucose metabolism. Exp Mol Med. 2016; 48:e216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Malenka RC, Nicoll RA. Long-term potentiation--a decade of progress? Science. 1999; 285:1870–1874. [DOI] [PubMed] [Google Scholar]
- 9.Duthie L, Reynolds RM. Changes in the maternal hypothalamic-pituitary-adrenal axis in pregnancy and postpartum: influences on maternal and fetal outcomes. Neuroendocrinology. 2013; 98:106–115. [DOI] [PubMed] [Google Scholar]
- 10.Brunton PJ. Resetting the dynamic range of hypothalamic-pituitary-adrenal axis stress responses through pregnancy. J Neuroendocrinol. 2010; 22: 1198–1213. [DOI] [PubMed] [Google Scholar]
- 11.Petraglia F, Imperatore A, Challis JR. Neuroendocrine mechanisms in pregnancy and parturition. Endocr Rev. 2010; 31:783–816. [DOI] [PubMed] [Google Scholar]
- 12.Hoekzema E, Barba-Muller E, Pozzobon C, Picado M, Lucco F, Garcia-Garcia D, Soliva JC, Tobena A, Desco M, Crone EA, Ballesteros A, Carmona S, Vilarroya O. Pregnancy leads to long-lasting changes in human brain structure. Nat Neurosci 2017; 20:287–296. [DOI] [PubMed] [Google Scholar]
- 13.Kinsley CH, Madonia L, Gifford GW, Tureski K, Griffin GR, Lowry C, Williams J, Collins J, McLearie H, Lambert KG. Motherhood improves learning and memory. Nature 1999; 402:137–138. [DOI] [PubMed] [Google Scholar]
- 14.Henry JF, Sherwin BB. Hormones and cognitive functioning during late pregnancy and postpartum: a longitudinal study. Behav Neurosci. 2012; 126:73–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Casey P, Huntsdale C, Angus G, Janes C. Memory in pregnancy. II: Implicit, incidental, explicit, semantic, short-term, working and prospective memory in primigravid, multigravid and postpartum women. J Psychosom Obstet Gynaecol. 1999; 20:158–164. [DOI] [PubMed] [Google Scholar]
- 16.Stolzenberg DS, Stevens JS, Rissman EF. Experience-facilitated improvements in pup retrieval; evidence for an epigenetic effect. Horm Behav 2012; 62:128–135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Behura SK, Kelleher AM, Spencer TE. Evidence for functional interactions between the placenta and brain in pregnant mice. FASEB J. 2019; 33:4261–4272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Arévalo L, Campbell P. Placental effects on maternal brain revealed by disrupted placental gene expression in mouse hybrids. bioRxiv 2019; 527143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lévy F Neuroendocrine control of maternal behavior in non-human and human mammals. Ann. Endocrinol. (Paris). 2016; 114–125. [DOI] [PubMed] [Google Scholar]
- 20.Napso T, Yong HEJ, Lopez-Tello J, Sferruzzi-Perri AN. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front Physiol. 2018; 1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bosch OJ, Neumann ID. Both oxytocin and vasopressin are mediators of maternal care and aggression in rodents: from central release to sites of action. Horm. Behav 2012; 293–303. [DOI] [PubMed] [Google Scholar]
- 22.Angoa-Pérez M, Kuhn DM. Neuronal serotonin in the regulation of maternal behavior in rodents. Neurotransmitter (Houst) 2015; e615. [PMC free article] [PubMed] [Google Scholar]
- 23.Lu CC, Chen JJ, Tsai SC, Chien EJ, Chien CH, Wang PS. Increase of thyrotropin response to thyrotropin-releasing hormone (TRH) and TRH release in rats during pregnancy. Chin. J. Physiol 1998; 211–216. [PubMed] [Google Scholar]
- 24.Kawamura K, Kawamura N, Sato W, Fukuda J, Kumagai J, Tanaka T. Brain-derived neurotrophic factor promotes implantation and subsequent placental development by stimulating trophoblast cell growth and survival. Endocrinology 2009; 150:3774–3782. [DOI] [PubMed] [Google Scholar]
- 25.Goldman-Wohl D, Greenfield C, Haimov-Kochman R, Ariel I, Anteby EY, Hochner-Celnikier D, Farhat M, Yagel S. Eph and ephrin expression in normal placental development and preeclampsia. Placenta 2004; 25:623–630. [DOI] [PubMed] [Google Scholar]
- 26.Chatzizacharias NA, Giaginis CT, Agapitos E, Theocharis SE. The role of ephrins' receptors and ephrins' ligands in normal placental development and disease. Expert Opin Ther Targets. 2014; 18:269–75. [DOI] [PubMed] [Google Scholar]
- 27.Angiolini E, Fowden A, Coan P, Sandovici I, Smith P, Dean W, Burton G, Tycko B, Reik W, Sibley C, Constância M. Regulation of placental efficiency for nutrient transport by imprinted genes. Placenta 2006; S98e102. [DOI] [PubMed] [Google Scholar]
- 28.Fowden GJ, Burton AL. Review: The placenta and developmental programming: Balancing fetal nutrient demands with maternal resource allocation. Placenta 2012; 23–27. [DOI] [PubMed] [Google Scholar]
- 29.Constancia M, Hemberger M, Hughes J, Dean W, Ferguson-Smith A, Fundele R.. Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 2002; 945e8. [DOI] [PubMed] [Google Scholar]
- 30.Grattan DR, Ladyman SR, Augustine RA. Hormonal induction of leptin resistance during pregnancy. Physiol Behav 2007; 366–374. [DOI] [PubMed] [Google Scholar]
- 31.Ladyman SR. Leptin resistance during pregnancy in the rat. J Neuroendocrinol 2008; 269–277. [DOI] [PubMed] [Google Scholar]
- 32.Belkacemi L, Desai M, Nelson DM, Ross MG. Altered mitochondrial apoptotic pathway in placentas from undernourished rat gestations. Am J Physiol Regul Integr Comp Physiol. 2011; 301:R1599–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mayeur S, Lancel S, Theys N, Lukaszewski MA, Duban-Deweer S, Bastide B, Hachani J, Cecchelli R, Breton C, Gabory A, Storme L, Reusens B, Junien C, Vieau D, Lesage J. Maternal calorie restriction modulates placental mitochondrial biogenesis and bioenergetic efficiency: putative involvement in fetoplacental growth defects in rats. Am J Physiol Endocrinol Metab. 2013; 304:E14–22. [DOI] [PubMed] [Google Scholar]
- 34.Lambertini L, Chen J, Nomura Y. Mitochondrial Gene Expression Profiles Are Associated with Maternal Psychosocial Stress in Pregnancy and Infant Temperament. PLoS One. 2015; 10:e0138929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Grimm A, Eckert A. Brain aging and neurodegeneration: from a mitochondrial point of view. J Neurochem. 2017; 143:418–431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brunst KJ, Sanchez Guerra M, Gennings C, Hacker M, Jara C, Bosquet Enlow M, Wright RO, Baccarelli A, Wright RJ. Maternal Lifetime Stress and Prenatal Psychological Functioning and Decreased Placental Mitochondrial DNA Copy Number in the PRISM Study. Am J Epidemiol. 2017; 186:1227–1236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Freeman ME, Smith MS, Nazian SJ, Neill JD. Ovarian and hypothalamic control of the daily surges of prolactin secretion during pseudopregnancy in the rat. Endocrinology 1974; 875–882. [DOI] [PubMed] [Google Scholar]
- 38.Augustine RA, Grattan DR. Induction of central leptin resistance in hyperphagic pseudopregnant rats by chronic prolactin infusion. Endocrinology 2008; 1049–1055. [DOI] [PubMed] [Google Scholar]
- 39.Kelleher AM, Milano-Foster J, Behura SK, Spencer TE. Uterine glands coordinate on-time embryo implantation and impact endometrial decidualization for pregnancy success. Nat Commun 2018; 9:2435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cha J, Sun X, Dey SK. Mechanisms of implantation: strategies for successful pregnancy. Nat Med 2012; 18:1754–1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Das SK. Regional development of uterine decidualization: molecular signaling by Hoxa-10. Molecular reproduction and development 2010; 77:387–396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhang S, Lin H, Kong S, Wang S, Wang H, Wang H, Armant DR. Physiological and molecular determinants of embryo implantation. Mol Aspects Med 2013; 34:939–980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gellersen B, Brosens JJ. Cyclic decidualization of the human endometrium in reproductive health and failure. Endocr Rev 2014; 35:851–905 [DOI] [PubMed] [Google Scholar]
- 44.Gellersen B, Brosens IA, Brosens JJ. Decidualization of the human endometrium: mechanisms, functions, and clinical perspectives. Semin Reprod Med 2007; 25:445–453 [DOI] [PubMed] [Google Scholar]
- 45.Burton GJ, Scioscia M, Rademacher TW. Endometrial secretions: creating a stimulatory microenvironment within the human early placenta and implications for the aetiopathogenesis of preeclampsia. J Reprod Immunol 2011; 89:118–125 [DOI] [PubMed] [Google Scholar]
- 46.Wooding FBP, Burton GJ. Comparative Placentation: Structures, Functions and Evolution. Berlin: Springer, c2008. [Google Scholar]
- 47.Red-Horse K, Zhou Y, Genbacev O, Prakobphol A, Foulk R, McMaster M, Fisher SJ. Trophoblast differentiation during embryo implantation and formation of the maternal-fetal interface. J Clin Invest 2004; 114:744–754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Burton GJ, Jauniaux E, Charnock-Jones DS. Human early placental development: potential roles of the endometrial glands. Placenta 2007; 28 Suppl A:S64–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Collier RJ, Doelger SG, Head HH, Thatcher WW, Wilcox CJ. Effects of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and postpartum milk yield of Holstein cows. J Anim Sci. 1982; 309–19. [DOI] [PubMed] [Google Scholar]
- 50.Dodic M, May CN, Wintour EM, Coghlan JP. An early prenatal exposure to excess glucocorticoid leads to hypertensive offspring in sheep. Clin Sci (Lond) 1998; 149–155. [DOI] [PubMed] [Google Scholar]
- 51.Beversdorf DQ, Manning SE, Hillier A, Anderson SL, Nordgren RE, Walters SE, Nagaraja HN, Cooley WC, Gaelic SE, Bauman ML. Timing of prenatal stressors and autism. J Autism Dev Disord. 2005; 471–478. [DOI] [PubMed] [Google Scholar]
- 52.Khashan AS, Abel KM, McNamee R, Pedersen MG, Webb RT, Baker PN, Kenny LC, Mortensen PB. Higher risk of offspring schizophrenia following antenatal maternal exposure to severe adverse life events. Arch Gen Psychiatry 2008; 146–152. [DOI] [PubMed] [Google Scholar]
- 53.Kinney DK, Miller AM, Crowley DJ, Huang E, Gerber E. Autism prevalence following prenatal exposure to hurricanes and tropical storms in Louisiana. J Autism Dev Disord 2008; 481–488. [DOI] [PubMed] [Google Scholar]
- 54.Li J, Olsen J, Vestergaard M, Obel C. Attention-deficit/hyperactivity disorder in the offspring following prenatal maternal bereavement: a nationwide follow-up study in Denmark. . Eur Child Adolesc Psychiatry 2010; 747–753. [DOI] [PubMed] [Google Scholar]
- 55.Rodriguez A, Bohlin G. Are maternal smoking and stress during pregnancy related to ADHD symptoms in children? J Child Psychol Psychiatry 2005; 246–254. [DOI] [PubMed] [Google Scholar]
- 56.Voltolini C, Petraglia F. Neuroendocrinology of pregnancy and parturition. Handb. Clin. Neurol 2014, 17–36. [DOI] [PubMed] [Google Scholar]
- 57.Russell John A., Brunton Paula J.. Giving a good start to a new life via maternal brain allostatic adaptations in pregnancy. Frontiers in Neuroendocrinology 2019; 100739. [DOI] [PubMed] [Google Scholar]
- 58.Ain R, Dai G, Dunmore JH, Godwin AR, Soares MJ. A prolactin family paralog regulates reproductive adaptations to a physiological stressor. Proc Natl Acad Sci U S A. 2004; 101:16543–16548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Paris JJ, Frye CA. Estrous cycle, pregnancy, and parity enhance performance of rats in object recognition or object placement tasks. Reproduction. 2008; 136:105–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Macbeth AH, Luine VN. Changes in anxiety and cognition due to reproductive experience: a review of data from rodent and human mothers. Neurosci Biobehav Rev. 2010; 34:452–467. [DOI] [PubMed] [Google Scholar]
- 61.Pawluski JL, Galea LA. Reproductive experience alters hippocampal neurogenesis during the postpartum period in the dam. Neuroscience. 2007; 149:53–67. [DOI] [PubMed] [Google Scholar]
- 62.Díaz P, Powell TL, Jansson T. The role of placental nutrient sensing in maternal-fetal resource allocation. Bio Reprod. 2014; 91(4):82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Konishi H, Fujiya M, Kohgo Y. Host-microbe interactions via membrane transport systems. Environ Microbiol. 2015; 17(4):931–937. [DOI] [PubMed] [Google Scholar]
- 64.Mikaelsson MA, Constância M, Dent CL, Wilkinson LS, Humby T. Placental programming of anxiety in adulthood revealed by Igf2-null models. Nat Commun. 2013; 4:2311. [DOI] [PubMed] [Google Scholar]
- 65.Howerton CL, Morgan CP, Fischer DB, Bale TL. O-GlcNAc transferase (OGT) as a placental biomarker of maternal stress and reprogramming of CNS gene transcription in development. Proc Natl Acad Sci U S A. 2013; 110:5169–5174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pantaleon M, Steane SE, McMahon K, Cuffe JSM, Moritz KM. Placental O-GlcNAc-transferase expression and interactions with the glucocorticoid receptor are sex specific and regulated by maternal corticosterone exposure in mice. Sci Rep. 2017; 7:2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mueller BR, Bale TL. Sex-specific programming of offspring emotionality after stress early in pregnancy. J Neurosci. 2008; 28:9055–9065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Bronson SL, Bale TL. Prenatal stress-induced increases in placental inflammation and offspring hyperactivity are male-specific and ameliorated by maternal antiinflammatory treatment. Endocrinology. 2014; 155:2635–2646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Goeden N, Velasquez J, Arnold KA, Chan Y, Lund BT, Anderson GM, Bonnin A. Maternal Inflammation Disrupts Fetal Neurodevelopment via Increased Placental Output of Serotonin to the Fetal Brain. J Neurosci. 2016; 36:6041–6049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zoeller RT. Transplacental thyroxine and fetal brain development. J Clin Invest. 2003; 111: 954–957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Shallie PD, Naicker T. The placenta as a window to the brain: A review on the role of placental markers in prenatal programming of neurodevelopment. Int J Dev Neurosci. 2019; 73:41–49. [DOI] [PubMed] [Google Scholar]
- 72.Bonnin A, Goeden N, Chen K, Wilson ML, King J, Shih JC, Blakely RD, Deneris ES, Levitt P. A transient placental source of serotonin for the fetal forebrain. Nature. 2011; 472:347–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.DeSesso JM, Scialli AR, Holson JF. Apparent lability of neural tube closure in laboratory animals and humans. Am J Med Genet. 1999; 87:143–162. [DOI] [PubMed] [Google Scholar]
- 74.Rice D, Barone S Jr. Critical periods of vulnerability for the developing nervous system: evidence from humans and animal models. Environ Health Perspect. 2000; 108 Suppl 3:511–533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Molnár Z, Clowry G. Cerebral cortical development in rodents and primates. Prog Brain Res. 2012; 195:45–70. [DOI] [PubMed] [Google Scholar]
- 76.Malassiné A, Frendo JL, Evain-Brion D. A comparison of placental development and endocrine functions between the human and mouse model. Hum Reprod Update. 2003; 9:531–539. [DOI] [PubMed] [Google Scholar]
- 77.Gao Z, Ure K, Ding P, Nashaat M, Yuan L, Ma J, Hammer RE, Hsieh J. The master negative regulator REST/NRSF controls adult neurogenesis by restraining the neurogenic program in quiescent stem cells. J Neurosci. 2011; 31:9772–9786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Janky R, Verfaillie A, Imrichová H, Van de Sande B, Standaert L, Christiaens V, Hulselmans G, Herten K, Naval Sanchez M, Potier D, Svetlichnyy D, Kalender Atak Z, Fiers M, Marine JC, Aerts S. iRegulon: from a gene list to a gene regulatory network using large motif and track collections. PLoS Comput Biol. 2014; 10:e1003731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Aagaard K, Ma J, Antony KM, Ganu R, Petrosino J, Versalovic J. The placenta harbors a unique microbiome. Sci Transl Med. 2014; 6:237ra65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Fardini Y, Chung P, Dumm R, Joshi N, Han YW. Transmission of diverse oral bacteria to murine placenta: evidence for the oral microbiome as a potential source of intrauterine infection. Infect Immun. 2010; 78:1789–1796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Robbins JR, Bakardjiev AI. Pathogens and the placental fortress. Curr Opin Microbiol. 2012; 15:36–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Atladóttir HO, Thorsen P, Østergaard L, Schendel DE, Lemcke S, Abdallah M, Parner ET. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J Autism Dev Disord. 2010; 40:1423–1430. [DOI] [PubMed] [Google Scholar]
- 83.Brown AS. Epidemiologic studies of exposure to prenatal infection and risk of schizophrenia and autism. Dev Neurobiol. 2012; 72:1272–1276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Meyer U, Nyffeler M, Engler A, Urwyler A, Schedlowski M, Knuesel I, Yee BK, Feldon J. The time of prenatal immune challenge determines the specificity of inflammation-mediated brain and behavioral pathology. J Neurosci. 2006; 26:4752–4762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Cordeiro CN, Tsimis M, Burd I. Infections and Brain Development. Obstet Gynecol Surv. 2015; 70:644–655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Wen Z, Song H, Ming GL. How does Zika virus cause microcephaly? Genes Dev. 2017; 31:849–861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Raj RS, Bonney EA, Phillippe M. Influenza, immune system, and pregnancy. Reprod Sci. 2014; 21:1434–1451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hagberg H, Mallard C Ferriero DM, Vannucci SJ, Levison SW, Vexler ZS, Gressens P. The role of inflammation in perinatal brain injury. Nat Rev Neurol. 2015; 11:192–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Tita AT, Andrews WW. Diagnosis and management of clinical chorioamnionitis. Clin Perinatol. 2010; 37:339–354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Altshuler G, McAdams AJ. The role of the placenta in fetal and perinatal pathology. Highlights of an eight months' study. Am J Obstet Gynecol. 1972; 113:616–626. [DOI] [PubMed] [Google Scholar]
- 91.Knuesel I, Chicha L, Britschgi M, Schobel SA, Bodmer M, Hellings JA, Toovey S, Prinssen EP. Maternal immune activation and abnormal brain development across CNS disorders. Nat Rev Neurol. 2014; 10:643–660. [DOI] [PubMed] [Google Scholar]
- 92.Wu YW, Escobar GJ, Grether JK, Croen LA, Greene JD, Newman TB. Chorioamnionitis and cerebral palsy in term and near-term infants. JAMA. 2003; 290: 2677–2684. [DOI] [PubMed] [Google Scholar]
- 93.van der Aa EM, Peereboom-Stegeman JH, Noordhoek J, Gribnau FW, Russel FG. Mechanisms of drug transfer across the human placenta. Pharm World Sci. 1998; 20:139–48. [DOI] [PubMed] [Google Scholar]
- 94.Pasanen M, Pelkonen O. The expression and environmental regulation of P450 enzymes in human placenta. Crit Rev Toxicol. 1994; 24:211–229. [DOI] [PubMed] [Google Scholar]
- 95.Sabo A, Stanulović M, Jakovljević V, Grujić Z. Collaborative study on drug use in pregnancy: the results of the follow-up 10 years after (Novi Sad Centre). Pharmacoepidemiol Drug Saf. 2001; 10:229–235. [DOI] [PubMed] [Google Scholar]
- 96.Rogers JM. Tobacco and pregnancy: overview of exposures and effects. Birth Defects Res C Embryo Today. 2008; 84:1–15. [DOI] [PubMed] [Google Scholar]
- 97.Gaither KH, Brunner Huber LR, Thompson ME, Huet-Hudson YM. Does the use of nicotine replacement therapy during pregnancy affect pregnancy outcomes? Matern Child Health J. 2009; 13: 497–504. [DOI] [PubMed] [Google Scholar]
- 98.Sarasin A, Schlumpf M, Muller M, Fleischmann I, Lauber ME, Lichtensteiger W. Adrenal-mediated rather than direct effects of nicotine as a basis of altered sex steroid synthesis in fetal and neonatal rat. Reprod Toxicol. 2003; 17:153–162. [DOI] [PubMed] [Google Scholar]
- 99.Levitt P Prenatal effects of drugs of abuse on brain development, Drug Alcohol Depend. 1998; 51:109–125. [DOI] [PubMed] [Google Scholar]
- 100.Bonnin A, Goeden N, Chen K, Wilson ML, King J, Shih JC, Blakely RD, Deneris ES, Levitt P. A transient placental source of serotonin for the fetal forebrain. Nature. 2011; 472:347–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Barker DJ. The fetal and infant origins of adult disease. BMJ 1990; 301: p.1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Rosenfeld CS. Introduction to Comparative Placentation, in Comparative Reproductive Biology, Schatten Hand Constatinescu GM, Editors. Blackwell Publishing: Ames, IA: 2007; p. 263–319. [Google Scholar]
- 103.Woods L, Perez-Garcia V, Hemberger M. Regulation of Placental Development and Its Impact on Fetal Growth-New Insights From Mouse Models. Front Endocrinol (Lausanne), 2018; 9: p.570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Georgiades P, Ferguson-Smith AC, Burton GJ. Comparative developmental anatomy of the murine and human definitive placentae. Placenta. 2002; 23:3–19. [DOI] [PubMed] [Google Scholar]
- 105.Cox B, Kotlyar M, Evangelou AI, Ignatchenko V, Ignatchenko A, Whiteley K, Jurisica I, Adamson SL, Rossant J, Kislinger T. Comparative systems biology of human and mouse as a tool to guide the modeling of human placental pathology. Mol Syst Biol. 2009; 5:279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Callaghan WM, MacDorman MF, Rasmussen SA, Qin C, Lackritz EM. The contribution of preterm birth to infant mortality rates in the United States. Pediatrics. 2006; 118:1566–1573. [DOI] [PubMed] [Google Scholar]
- 107.Morgan TK. Role of the Placenta in Preterm Birth: A Review. Am J Perinatol. 2016; 33:258–266. [DOI] [PubMed] [Google Scholar]
- 108.Brosens I, Pijnenborg R, Vercruysse L, Romero R. The "Great Obstetrical Syndromes" are associated with disorders of deep placentation. Am J Obstet Gynecol. 2011; 204: 193–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Hasan MZ, Ikawati M, Tocharus J, Kawaichi M, Oka C. Abnormal development of placenta in HtrA1-deficient mice. Dev Biol. 2015; 397:89–102. [DOI] [PubMed] [Google Scholar]
- 110.Hunkapiller NM, Gasperowicz M, Kapidzic M, Plaks V, Maltepe E, Kitajewski J, Cross JC, Fisher SJ. A role for Notch signaling in trophoblast endovascular invasion and in the pathogenesis of pre-eclampsia. Development. 2011; 138:2987–2998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Sharma D, Shastri S, Sharma P. Intrauterine Growth Restriction: Antenatal and Postnatal Aspects. Clin Med Insights Pediatr. 2016; 10:67–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Zeltser LM, Leibel RL. Roles of the placenta in fetal brain development. Proc Natl Acad Sci U S A. 2011; 108:15667–15668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Miller SL, Huppi PS, Mallard C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J Physiol. 2016; 594: 807–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Cohen E, Baerts W, van Bel F. Brain-Sparing in Intrauterine Growth Restriction: Considerations for the Neonatologist. Neonatology. 2015; 108:269–276. [DOI] [PubMed] [Google Scholar]
- 115.Broad KD, Keverne EB. Placental protection of the fetal brain during short-term food deprivation. Proc Natl Acad Sci U S A. 2011; 108:15237–15241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Bonnin A, Goeden N, Chen K, Wilson ML, King J, Shih JC, Blakely RD, Deneris ES, Levitt P. A transient placental source of serotonin for the fetal forebrain. Nature. 2011; 472:347–350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.de Bie HM, Oostrom KJ, Delemarre-van de Waal HA. Brain development, intelligence and cognitive outcome in children born small for gestational age. Horm Res Paediatr. 2010; 73:6–14. [DOI] [PubMed] [Google Scholar]
- 118.Avraamides CJ, Garmy-Susini B, Varner JA. Integrins in angiogenesis and lymphangiogenesis. Nat Rev Cancer. 2008; 8:604–617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Barquilla A, Pasquale EB. Eph receptors and ephrins: therapeutic opportunities. Annu Rev Pharmacol Toxicol. 2015; 55: 465–487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Ley K, Rivera-Nieves J, Sandborn WJ, Shattil S. Integrin-based therapeutics: biological basis, clinical use and new drugs. Nat Rev Drug Discov. 2016; 15:173–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Degenhardt L, Charlson F, Mathers B, Hall WD, Flaxman AD, Johns N, Vos T. The global epidemiology and burden of opioid dependence: results from the global burden of disease 2010 study. Addiction. 2014; 109:1320–1333 [DOI] [PubMed] [Google Scholar]
- 122.Krans EE, Patrick SW. Opioid Use Disorder in Pregnancy: Health Policy and Practice in the Midst of an Epidemic. Obstet Gynecol. 2016; 128:4–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Shainker SA, Saia K, Lee-Parritz A. Opioid addiction in pregnancy. Obstet Gynecol Surv. 2012; 67:817–825. [DOI] [PubMed] [Google Scholar]
- 124.Frascella J, Richardson KA, McLemore GL. Animal models of drug addiction in support of novel therapeutic strategies. ILAR J. 2011; 52:233–238. [DOI] [PubMed] [Google Scholar]
- 125.Grenald SA, Largent-Milnes TM, Vanderah TW. Animal models for opioid addiction drug discovery. Expert Opin Drug Discov. 2014; 9:1345–1354. [DOI] [PubMed] [Google Scholar]
- 126.Park EM, Meltzer-Brody S, Suzuki J. Evaluation and management of opioid dependence in pregnancy. Psychosomatics. 2012; 53:424–432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Shahzad K, Loor JJ. Application of Top-Down and Bottom-up Systems Approaches in Ruminant Physiology and Metabolism. Curr Genomics. 2012; 13:379–394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Iguchi T, Tani N, Sato T, Fukatsu N, Ohta Y. Developmental changes in mouse placental cells from several stages of pregnancy in vivo and in vitro. Biol Reprod. 1993; 48:188–196. [DOI] [PubMed] [Google Scholar]
- 129.Oliveira MS, Barreto-Filho JB. Placental-derived stem cells: Culture, differentiation and challenges. World J Stem Cells. 2015; 7:769–775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Jividen K, Movassagh MJ, Jazaeri A, Li H. Two methods for establishing primary human endometrial stromal cells from hysterectomy specimens. J Vis Exp. 2014; 87:51513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Aldarmahi A. Establishment and characterization of female reproductive tract epithelial cell culture. J Microsc Ultrastruct. 2017; 5:105–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Gargett CE. Uterine stem cells: what is the evidence? Hum Reprod Update. 2007; 13:87–101. [DOI] [PubMed] [Google Scholar]
- 133.Collas P, Noer A, Timoskainen S. Programming the genome in embryonic and somatic stem cells. J Cell Mol Med. 2007; 11:602–620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Deane JA, Cousins FL, Gargett CE. Endometrial organoids: in vitro models for endometrial research and personalized medicine. Biol Reprod. 2017; 97:781–783. [DOI] [PubMed] [Google Scholar]
- 135.Turco MY, Gardner L, Hughes J, Cindrova-Davies T, Gomez MJ, Farrell L, Hollinshead M, Marsh SGE, Brosens JJ, Critchley HO, Simons BD, Hemberger M, Koo BK, Moffett A, Burton GJ. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nat Cell Biol. 2017; 19:568–577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Boretto M, Maenhoudt N, Luo X, Hennes A, Boeckx B, Bui B, Heremans R, Perneel L, Kobayashi H, Van Zundert I, Brems H, Cox B, Ferrante M, Uji-I H, Koh KP, D'Hooghe T, Vanhie A, Vergote I, Meuleman C, Tomassetti C, Lambrechts D, Vriens J, Timmerman D, Vankelecom H Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening Nat Cell Biol., 21 (2019), pp. 1041–1051 [DOI] [PubMed] [Google Scholar]
- 137.Santamaria X, Mas A, Cervelló I, Taylor H, Simon C. Uterine stem cells: from basic research to advanced cell therapies. Hum Reprod Update. 2018; 24:673–693. [DOI] [PubMed] [Google Scholar]
- 138.Boretto M, Cox B, Noben M, Hendriks N, Fassbender A, Roose H, Amant F, Timmerman D, Tomassetti C, Vanhie A, Meuleman C, Ferrante M, Vankelecom H. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development. 2017; 144:1775–1786. [DOI] [PubMed] [Google Scholar]
- 139.Yin X, Mead BE, Safaee H, Langer R, Karp JM, Levy O. Engineering Stem Cell Organoids. Cell Stem Cell. 2016; 18:25–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Fitzgerald HC, Behura SK, Dhakal P, Schust DJ, Spencer TE. Development and Comprehensive Analysis of Hormone Responsive Human Endometrial Epithelial Organoids in 3D Culture. Reprod Sci. 2019; 26:197A–197A. [Google Scholar]
- 141.Turco MY, Gardner L, Kay RG, Hamilton RS, Prater M, Hollinshead MS, Mc Whinnie A, Esposito L, Fernando R, Skelton H, Reimann F, Gribble FM, Sharkey A, Marsh SGE, O'Rahilly S, Hemberger M, Burton GJ, Moffett A. Trophoblast organoids as a model for maternal-fetal interactions during human placentation. Nature. 2018; 564:263–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Haider S, Meinhardt G, Saleh L, Kunihs V, Gamperl M, Kaindl U, Ellinger A, Burkard TR, Fiala C, Pollheimer J, Mendjan S, Latos PA, Knöfler M. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Reports. 2018; 11:537–551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Vogt N Organoids that model the fetal placenta. Nat Methods. 2019; 16:144. [DOI] [PubMed] [Google Scholar]
- 144.Ingber DE. Developmentally inspired human 'organs on chips'. Development. 2018; 145:dev156125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Lee JS, Romero R, Han YM, Kim HC, Kim CJ, Hong JS, Huh D. Placenta-on-a-chip: a novel platform to study the biology of the human placenta. J Matern Fetal Neonatal Med. 2016; 29:1046–1054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Blundell C, Tess ER, Schanzer AS, Coutifaris C, Su EJ, Parry S, Huh D. A microphysiological model of the human placental barrier. Lab Chip. 2016; 16:3065–3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Li Y, Muffat J, Omer A, Bosch I, Lancaster MA, Sur M, Gehrke L, Knoblich JA , Jaenisch R. Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell. 2017; 20:385–396.e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Haring AP, Sontheimer H, Johnson BN. Microphysiological Human Brain and Neural Systems-on-a-Chip: Potential Alternatives to Small Animal Models and Emerging Platforms for Drug Discovery and Personalized Medicine. Stem Cell Rev. 2017; 13:381–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Kilic O, Pamies D, Lavell E, Schiapparelli P, Feng Y, Hartung T, Bal-Price A, Hogberg HT, Quinones-Hinojosa A, Guerrero-Cazares H, Levchenko A. Brain-on-a-chip model enables analysis of human neuronal differentiation and chemotaxis. Lab Chip. 2016; 16:4152–4162. [DOI] [PubMed] [Google Scholar]
- 150.Luo C, Lancaster MA, Castanon R, Nery JR, Knoblich JA, Ecker JR. Cerebral Organoids Recapitulate Epigenomic Signatures of the Human Fetal Brain. Cell Rep. 2016; 17:3369–3384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Skardal A, Shupe T, Atala A. Organoid-on-a-chip and body-on-a-chip systems for drug screening and disease modeling. Drug Discov Today. 2016; 21:1399–1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Takebe T, Sekine K, Kimura M, Yoshizawa E, Ayano S, Koido M, Funayama S, Nakanishi N, Hisai T, Kobayashi T, Kasai T, Kitada R, Mori A, Ayabe H, Ejiri Y, Amimoto N, Yamazaki Y, Ogawa S, Ishikawa M, Kiyota Y, Sato Y, Nozawa K, Okamoto S, Ueno Y, Taniguchi H. Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells. Cell Rep. 2017; 21:2661–2670. [DOI] [PubMed] [Google Scholar]
- 153.Wang Y, Wang L, Zhu Y, Qin J. Human brain organoid-on-a-chip to model prenatal nicotine exposure. Lab Chip. 2018; 18:851–860. [DOI] [PubMed] [Google Scholar]
- 154.Mittal R, Woo FW, Castro CS, Cohen MA, Karanxha J, Mittal J, Chhibber T, Jhaveri VM. Organ-on-chip models: Implications in drug discovery and clinical applications. J Cell Physiol. 2019; 234:8352–8380. [DOI] [PubMed] [Google Scholar]
- 155.Rothbauer M, Rosser JM, Zirath H, Ertl P. Tomorrow today: organ-on-a-chip advances towards clinically relevant pharmaceutical and medical in vitro models. Curr Opin Biotechnol. 2019; 55:81–86. [DOI] [PubMed] [Google Scholar]
- 156.Blundell C, Yi YS, Ma L, Tess ER, Farrell MJ, Georgescu A, Aleksunes LM, Huh D. Placental Drug Transport-on-a-Chip: A Microengineered In Vitro Model of Transporter-Mediated Drug Efflux in the Human Placental Barrier. Adv Healthc Mater. 2018; 7: 10.1002/adhm.201700786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Gnecco JS, Ding T, Smith C, Lu J, Bruner-Tran KL, Osteen KG. Hemodynamic forces enhance decidualization via endothelial-derived prostaglandin E2 and prostacyclin in a microfluidic model of the human endometrium. Hum Reprod. 2019; 34:702–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Thomason ME, Scheinost D, Manning JH, Grove LE, Hect J, Marshall N, Hernandez-Andrade E, Berman S, Pappas A, Yeo L, Hassan SS, Constable RT, Ment LR, Romero R. Weak functional connectivity in the human fetal brain prior to preterm birth. Sci Rep. 2017; 7:39286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Paton MCB, McDonald CA, Allison BJ, Fahey MC, Jenkin G, Miller SL. Perinatal Brain Injury As a Consequence of Preterm Birth and Intrauterine Inflammation: Designing Targeted Stem Cell Therapies. Front Neurosci. 2017; 11:200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Helldén A, Madadi P. Pregnancy and pharmacogenomics in the context of drug metabolism and response. Pharmacogenomics. 2013;14: 1779–1791. [DOI] [PubMed] [Google Scholar]
- 161.Haas DM. Pharmacogenetics and individualizing drug treatment during pregnancy. Pharmacogenomics. 2014; 15:69–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Zhou SF, Di YM, Chan E, Du YM, Chow VD, Xue CC, Lai X, Wang JC, Li CG, Tian M, Duan W. Clinical pharmacogenetics and potential application in personalized medicine. Curr Drug Metab. 2008; 9:738–784. [DOI] [PubMed] [Google Scholar]
- 163.Jeong H Altered drug metabolism during pregnancy: hormonal regulation of drug-metabolizing enzymes. Expert Opin Drug Metab Toxicol. 2010; 6:689–699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Hodge LS, Tracy TS. Alterations in drug disposition during pregnancy: implications for drug therapy. Expert Opin Drug Metab Toxicol. 2007; 3:557–571. [DOI] [PubMed] [Google Scholar]
- 165.Taylor MJ, Sen S, Bhagwagar Z. Antidepressant response and the serotonin transporter gene-linked polymorphic region. Biol Psychiatry. 2010; 68:536–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Dondorp W, de Wert G, Bombard Y, Bianchi DW, Bergmann C, Borry P, Chitty LS, Fellmann F, Forzano F, Hall A, Henneman L, Howard HC, Lucassen A, Ormond K, Peterlin B, Radojkovic D, Rogowski W, Soller M, Tibben A, Tranebjærg L, van El CG, Cornel MC; European Society of Human Genetics; American Society of Human Genetics. Non-invasive prenatal testing for aneuploidy and beyond: challenges of responsible innovation in prenatal screening. Eur J Hum Genet. 2015; 23:1438–1450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Bianchi DW. From prenatal genomic diagnosis to fetal personalized medicine: progress and challenges. Nat Med. 2012; 18:1041–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Pavličev M, Wagner GP, Chavan AR, Owens K, Maziarz J, Dunn-Fletcher C, Kallapur SG, Muglia L, Jones H. Single-cell transcriptomics of the human placenta: inferring the cell communication network of the maternal-fetal interface. Genome Res. 2017; 27:349–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Tsang JCH, Vong JSL, Ji L, Poon LCY, Jiang P, Lui KO, Ni YB, To KF, Cheng YKY, Chiu RWK, Lo YMD. Integrative single-cell and cell-free plasma RNA transcriptomics elucidates placental cellular dynamics. Proc Natl Acad Sci U S A. 2017; 114:E7786–E7795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Suryawanshi H, Morozov P, Straus A, Sahasrabudhe N, Max KEA, Garzia A, Kustagi M, Tuschl T, Williams Z. A single-cell survey of the human first-trimester placenta and decidua. Sci Adv. 2018; 4:eaau4788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Quinney SK, Patil AS, Flockhart DA. Is personalized medicine achievable in obstetrics? Semin Perinatol. 2014; 38:534–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Ventola CL. The role of pharmacogenomic biomarkers in predicting and improving drug response: part 2: challenges impeding clinical implementation. P T. 2013; 38:624–627. [PMC free article] [PubMed] [Google Scholar]
- 173.Kalinin AA, Higgins GA, Reamaroon N, Soroushmehr S, Allyn-Feuer A, Dinov ID, Najarian K, Athey BD. Deep learning in pharmacogenomics: from gene regulation to patient stratification. Pharmacogenomics. 2018; 19:629–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Vento-Tormo R, Efremova M, Botting RA, Turco MY, Vento-Tormo M, Meyer KB, Park JE, Stephenson E, Polański K, Goncalves A, Gardner L, Holmqvist S, Henriksson J, Zou A, Sharkey AM, Millar B, Innes B, Wood L, Wilbrey-Clark A, Payne RP, Ivarsson MA, Lisgo S, Filby A, Rowitch DH, Bulmer JN, Wright GJ, Stubbington MJT, Haniffa M, Moffett A, Teichmann SA. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature. 2018; 563:347–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Liu Y, Fan X, Wang R, Lu X, Dang YL, Wang H, Lin HY, Zhu C, Ge H, Cross JC, Wang H. Single-cell RNA-seq reveals the diversity of trophoblast subtypes and patterns of differentiation in the human placenta. Cell Res 2018; 28:819–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.La Manno G, Gyllborg D, Codeluppi S, Nishimura K, Salto C, Zeisel A, Borm LE, Stott SRW, Toledo EM, Villaescusa JC, Lönnerberg P, Ryge J, Barker RA, Arenas E, Linnarsson S. Molecular Diversity of Midbrain Development in Mouse, Human, and Stem Cells. Cell 2016; 167:566–580.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Han X, Wang R, Zhou Y, Fei L, Sun H, Lai S, Saadatpour A, Zhou Z, Chen H, Ye F, Huang D, Xu Y, Huang W, Jiang M, Jiang X, Mao J, Chen Y, Lu C, Xie J, Fang Q, Wang Y, Yue R, Li T, Huang H, Orkin SH, Yuan GC, Chen M, Guo G. Mapping the Mouse Cell Atlas by Microwell-Seq. Cell 2018; 172:1091–1107.e17. [DOI] [PubMed] [Google Scholar]
- 178.Butler A, Hoffman P, Smibert P, Papalexi E, Satija R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat Biotechnol. 2018; 36:411–420. [DOI] [PMC free article] [PubMed] [Google Scholar]