

Abstract
The type VI secretion system (T6SS) is used by many bacteria to engage in social behavior and can affect the health of its host plant or animal. Because activities associated with T6SSs are often costly, T6SSs must be tightly regulated. However, our knowledge regarding how T6SS assembly and contraction are regulated remains limited. Using the plant pathogen Agrobacterium tumefaciens, we show that effectors are not just passengers but also impact on T6SS assembly. The A. tumefaciens strain C58 encodes one T6SS and two Tde DNase toxin effectors used as major weapons for interbacterial competition. Here, we demonstrate that loading of Tde effectors onto their cognate carriers, the VgrG spikes, is required for active T6SS secretion. The assembly of the TssBC contractile sheath occurs only in the presence of Tde effectors. The requirement of effector loading for efficient T6SS secretion was also validated in other A. tumefaciens strains. We propose that such a mechanism is used by bacteria as a strategy for efficacious T6SS firing and to ensure that effectors are loaded onto the T6SS prior to completing its assembly.
Keywords: Agrobacterium tumefaciens, effector, interbacterial competition, type VI secretion system, VgrG
Subject Categories: Microbiology, Virology & Host Pathogen Interaction; Structural Biology
Bacterial effectors impact on the assembly of the type VI secretion machinery. Their attachment to VgrG spikes is required for active secretion, which ensures that cargo is loaded onto the T6SS prior to completing its assembly.
Introduction
The type VI secretion system (T6SS) is a versatile secretion system that has been implicated in virulence, antagonism, nutrient acquisition, and horizontal gene transfer 1, 2, 3. Contact‐dependent interbacterial competition appears to be the major function of T6SS, a function that can influence the composition of microbial communities 4, 5. A T6SS machine resembles a contractile phage tail‐like structure 6, 7, 8. Effectors are loaded onto the T6SS either via non‐covalent interactions (cargo effectors) or as carboxy‐terminal extensions to either of the three core structural components (Hcp, VgrG, or PAAR) 2, 9. Upon contraction, the puncturing device loaded with effectors is fired and propelled, carrying effectors, across the cell membrane into the extracellular milieu or into targeted bacterial or eukaryotic cells. In general, cargo effectors are considered as accessory components of the secretion apparatus. This is based on the repeated observations that Hcp and/or VgrG, key markers for T6SS secretion, can be detected in the extracellular milieu of mutants lacking effector genes 10, 11, 12, 13, 14.
The plant pathogenic bacterium Agrobacterium tumefaciens strain C58 encodes one T6SS main cluster consisting of the imp and hcp operons and vgrG2 operon distal to the main cluster 15. Three T6SS toxin effectors were identified, in which secretion of Tde1 and Tde2 DNases is governed specifically by VgrG1 and VgrG2, respectively, and secretion of Tae amidase is likely mediated by Hcp 16, 17. Tde effectors are major weapons deployed by A. tumefaciens for interbacterial competition in planta 10. Each of the effector genes is genetically linked to a cognate immunity gene, which form three toxin–immunity pairs (i.e., tae‐tai and tde1‐tdi1 located in the main gene cluster and tde2‐tdi2 encoded downstream of vgrG2 distal to the main cluster; Fig 1A). Self‐intoxication is prevented in the toxin‐producing cells by the cognate immunity proteins.
Figure 1. Presence of Tde effectors in the cell is critical for secretion of the cognate VgrG.
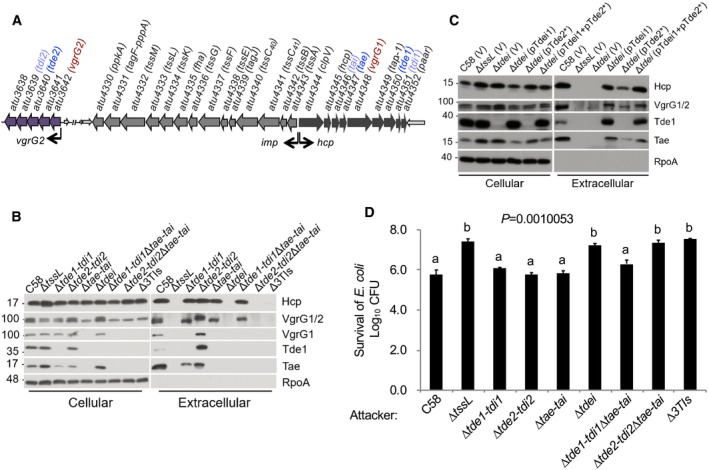
- The structure of the imp and hcp operons and vgrG2 operon in Agrobacterium tumefaciens C58 10. The arrows represent coding sequencing, with arrowheads depicting the direction of expression. The wider arrows represent genes in the T6SS‐associated operons. The vgrG2 operon is located distal to imp operon and hcp operon. The vgrG and toxin–immunity gene pairs are highlighted in colors.
- T6SS secretion assay of A. tumefaciens strains: wild‐type C58, various mutants lacking one, two, or three toxin–immunity gene pairs, and a mutant lacking tssL.
- T6SS secretion assay of various A. tumefaciens strains: wild‐type C58, ΔtssL, the tde double deletion mutant (Δtdei) containing pRL662 and pTrc200 empty vector (V) only, or expression of pTdei1 (tde1‐tdi1 expressed from pTrc200), pTde2* (catalytic site‐mutated tde2 expressed on pRL662), or pTdei1+ pTde2*.
- Agrobacterium tumefaciens antibacterial activity assay against Escherichia coli. The strains of A. tumefaciens were co‐cultured at a ratio of 30:1 with E. coli DH10B (+ pRL662) on LB agar. The survival of target E. coli cells was quantified by counting CFUs on gentamicin‐containing LB agar plates. Data represent mean ± SEM of 6 biological replicates from three independent experiments. One‐way ANOVA followed by Turkey HSD test was used for statistical analysis. Two groups with significant differences (P = 0.0010053) are indicated with different letters (a and b).
Data are from one independent experiment and reproduced in at least two independent experiments. Source data are available online for this figure.
Here, we show that the loading of cargo effectors onto their cognate VgrG spike proteins is required for efficient T6SS‐dependent secretion by A. tumefaciens. We demonstrate that in the absence of VgrG cargo effector genes, the levels of secretion of Hcp and VgrG and formation of the TssBC sheath are significantly reduced in A. tumefaciens. Given the prevalence of T6SS‐encoding loci in host‐associated bacteria, these findings inform on mechanisms that may influence the composition of microbial communities.
Results
Tde effector loading onto VgrG is required for secretion of cognate VgrG
Despite evidence suggesting that effectors are not components of the secretion apparatus 10, 11, 12, 13, 14, our previous study showed that A. tumefaciens strains with variants of VgrG1 lacking the Tde1‐binding domain are able to secrete Hcp but at slightly lower levels 16. This led us to hypothesize that effector‐loaded VgrG is more efficiently recruited for T6SS assembly and/or secretion. To test this hypothesis, we first examined whether the secretion of Hcp and VgrG proteins, a hallmark of T6SS firing, is affected by the presence or absence of effector genes. We found that the secretion of Hcp and VgrG proteins is affected by the presence or absence of Tde effector genes in A. tumefaciens. In wild‐type C58, but not ΔtssL, a secretion‐deficient mutant, Hcp, VgrG1/2, Tde1, and Tae were detected in the medium, referred as extracellular fraction, confirming their T6SS‐dependent secretion (Fig 1B). However, VgrG1 and VgrG2 proteins were not detected in the extracellular fraction of mutant strains in which their cognate effector gene was absent. VgrG1 was not detectable in the extracellular fraction of any mutant minimally lacking the tde1‐tdi1 toxin–immunity gene pair (i.e., Δtde1‐tdi1, Δtdei lacking both tde1‐tdi1 and tde2‐tdi2, and Δ3TIs lacking all three toxin–immunity pairs). Similarly, VgrG2 is inferred to behave in a similar manner in mutants with tde2‐tdi2 toxin–immunity gene pair deleted. Complementing the Δtdei mutant with tde1‐tdi1 restored its ability to secrete VgrG1, while the mutant expressing tde2* (encoding Tde2 variant with catalytic site mutation) failed to secrete VgrG1. In contrast, the mutant carrying tde2*, but not tde1‐tdi1, restored the secretion of VgrG2 (Fig 1C), which also suggested that DNase activity of Tde effectors is not required for T6SS assembly and secretion. The levels of Hcp were similar in the extracellular fractions of the wild‐type strain as well as that from each of the mutants lacking a single toxin–immunity gene pair. Interbacterial competition assays showed that A. tumefaciens C58 can only kill Escherichia coli when at least one Tde effector is delivered (Fig 1D). The reliance on Tde DNases but not Tae amidase as the primary effectors against E. coli is consistent with previous finding that Tde but not Tae influences the in planta interbacterial competition activity of A. tumefaciens 10.
Tde1 and Tde2 require specific adaptor/chaperone proteins to be loaded onto their cognate VgrG spike 16. Thus, we next examined whether secretion of the two VgrG spike proteins requires the cognate adaptor/chaperones: Tap‐1 (DUF4123‐containing protein) for VgrG1 and Atu3641 (DUF2169‐containing protein) for VgrG2. Secretion was assayed in Δtap‐1 and Δatu3641, mutants deleted of genes encoding the adaptor/chaperones for Tde1 and Tde2, respectively (Fig 2A) 16. Non‐secreted T6SS proteins (TssB and ClpV) as well as secreted proteins (Hcp, VgrG1/2, Tde1, and Tae) accumulated to detectable levels in the cellular fractions of all strains (Figs EV1A and 2A). VgrG1 and Tde1 were only detected in the extracellular fraction of cells that encoded Tap‐1. Likewise, VgrG2 was only detected in the extracellular fraction of cells that encoded Atu3641. Hcp and Tae were detected in the extracellular fraction when at least one VgrG variant was secreted from cells. Importantly, Hcp, VgrG1/2, Tde1, and Tae were not detected in the extracellular fraction of the Δtap‐1Δatu3641 mutant. Secretion of VgrG1 or VgrG2 could be restored when the mutant was complemented with tap‐1 or atu3641, respectively (Fig 2B). As controls, relevant T6SS proteins were detected in the cellular fractions (Figs 2B and EV1B).
Figure 2. Tde effector loading onto VgrG is required for secretion of the cognate VgrG.
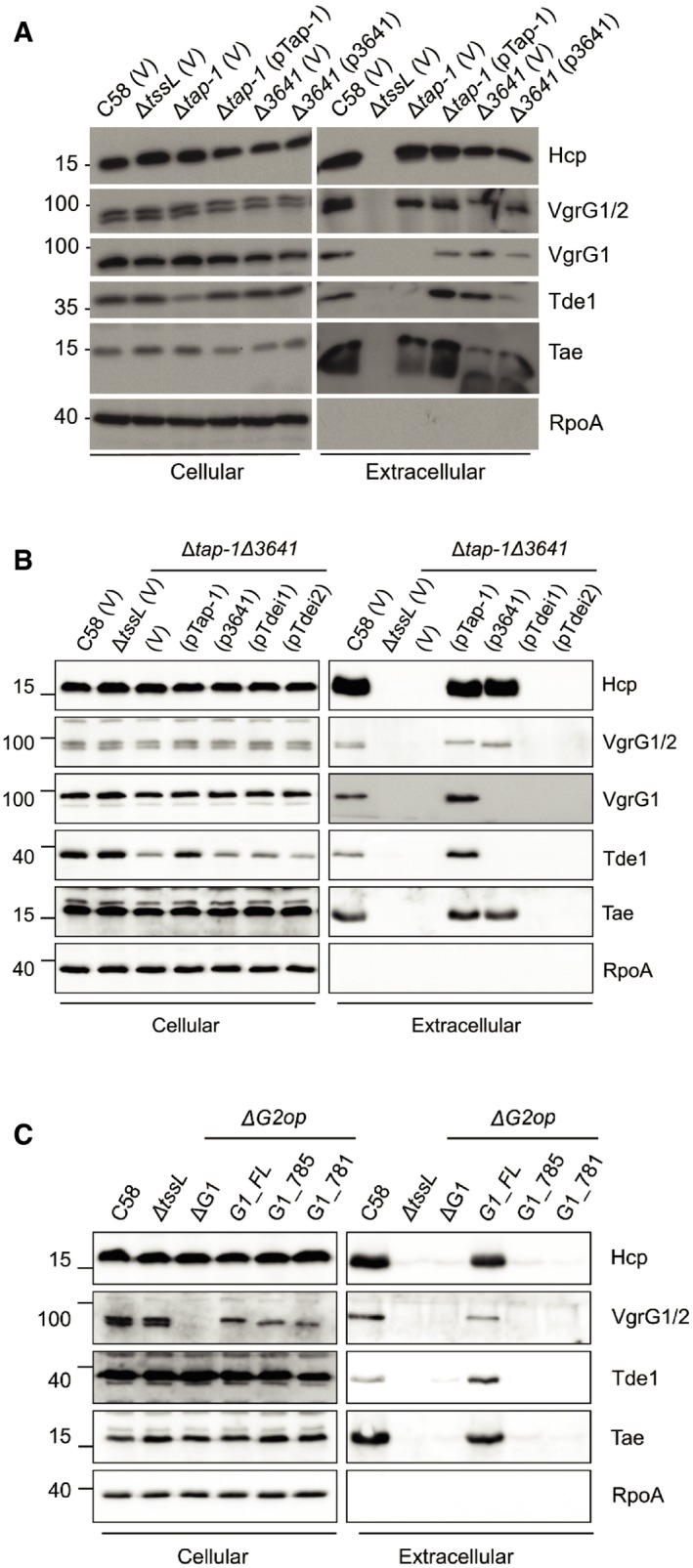
- T6SS secretion assay of Agrobacterium tumefaciens strains: wild‐type C58, ΔtssL, Δtap‐1, and Δatu3641 harboring a pTrc200 vector (V) or derivatives pTap‐1 (tap‐1 expressed from pTrc200), or p3641 (atu3641 expressed from pTrc200).
- T6SS secretion assay of A. tumefaciens C58 Δtap‐1Δatu3641 containing an empty pTrc200 vector (V) or its derivatives expressing tap‐1, atu3641, tde1‐tdi1, or tde2‐tdi2.
- T6SS secretion assay of A. tumefaciens C58ΔG2op mutant encoding full‐length or truncated VgrG1 proteins.
Data are from one independent experiment and reproduced in at least two independent experiments. Source data are available online for this figure.
Figure EV1. Western blots of cellular fractions of representative T6SS proteins from Fig 2 .
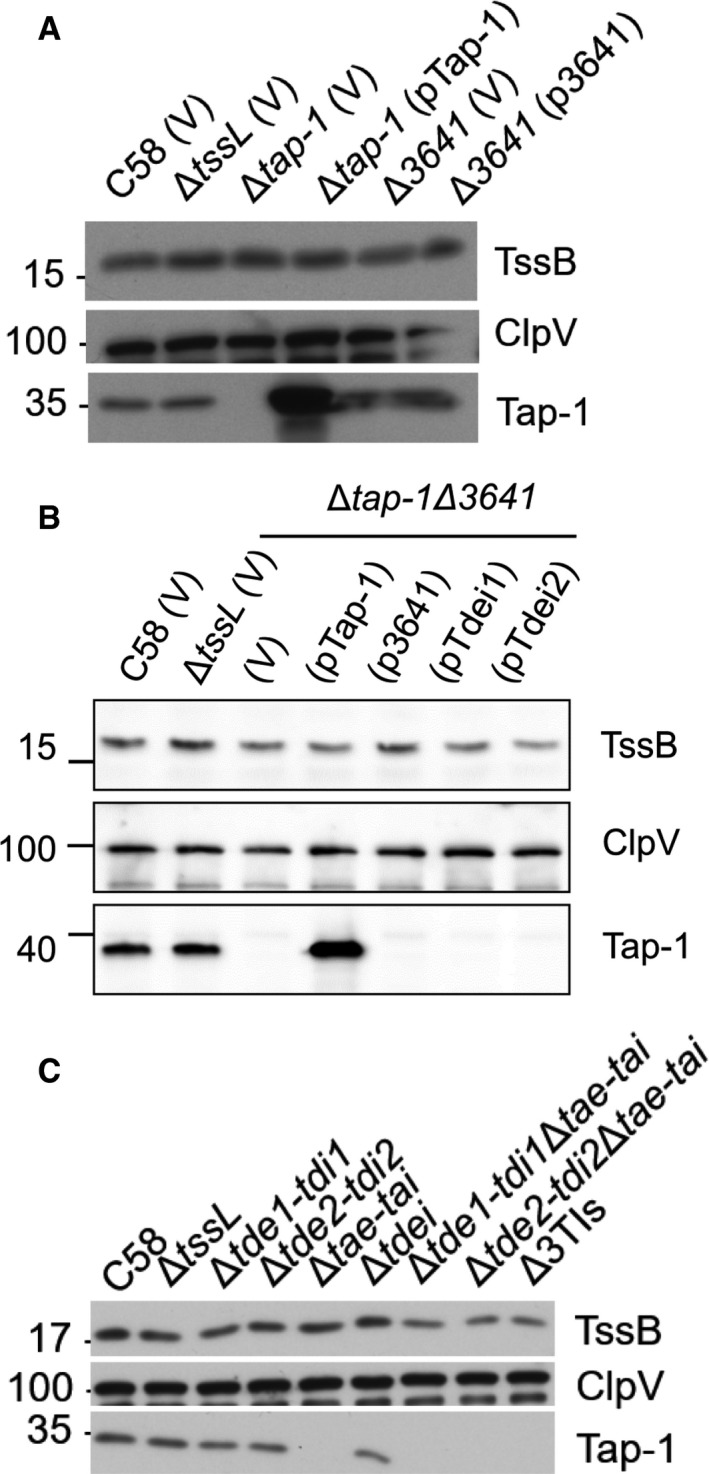
- Aliquots of the same cellular fractions that were analyzed in Fig 2A were probed with a set of different antibodies.
- Aliquots of the same cellular fractions that were analyzed in Fig 2B were probed with a set of different antibodies.
- Aliquots of the same cellular fractions that were analyzed in Fig 1B were probed with a set of different antibodies.
As reported 10, Tde1 accumulates to lower levels in the absence of tap‐1 (Fig 2A and B). Use of the trc promoter in an attempt to overexpress Tde1‐Tdi1 or Tde2‐Tdi2 from a plasmid in Δtap‐1Δatu3641 was unable to restore secretion of cognate VgrG variants (Fig 2B). Because Tde1 always accumulated at lower levels in the absence of tap‐1 regardless of whether it is expressed endogenously or from a plasmid, we could not exclude the possibility that the deficiency of VgrG1 secretion in Δtap‐1 might be due to the lower amounts of cellular Tde1 in the absence of Tap‐1. However, the data are consistent with the role of Tap‐1 in stabilizing Tde1 as reported earlier 10 and demonstrated that the adaptor/chaperone is required for secretion of the cognate VgrG spike proteins.
In our previous study, when VgrG1 variants lacking the Tde1‐binding region were expressed in a vgrG1vgrG2 double deletion mutant (ΔG1ΔG2) and at higher levels than that of native VgrG1 in wild‐type C58, they were still able to mediate secretion of Hcp and Tae effector, albeit at slightly lower levels 16. We then hypothesized that overexpression of VgrG in the absence of a cognate effector may be sufficient to initiate the assembly of the T6SS. A polymutant strain (Δtde1‐tdi1ΔG1ΔG2op) lacking both Tde1/2 effectors and VgrG1/2 was generated and used for overexpression of VgrG1 and its variants. As expected, Hcp did not accumulate in the extracellular fraction of this mutant (Fig EV2A). However, when wild‐type VgrG1 or truncated variants (G1_812, G1_804, and G1_785) that are abrogated in their ability to interact with Tde1 were overexpressed in this mutant, Hcp could be detected at high levels in the extracellular fraction. The shortest variant of VgrG1 (G1_781) previously shown to be incapable of restoring Hcp secretion in ΔG1ΔG2 16 was unable to restore Hcp secretion in Δtde1‐tdi1ΔG1ΔG2op (Fig EV2A). The data demonstrated that overexpression of VgrG overrides the absence of effector genes for initiation of T6SS assembly. However, when the chromosome‐encoded vgrG1 coding sequence was truncated, variants G1_785 and G1_781 were barely detectable in the extracellular fraction while maintaining cellular Tde1 at wild‐type levels (Figs 2C and EV2B). Furthermore, the defect in the secretion of G1_785 and G1_781 is correlated with the lack of detectable extracellular Hcp, Tae, and Tde1. The data altogether demonstrated that Tde1 effector loading onto VgrG1 rather than the abundance of cellular Tde1 is critical for VgrG1 secretion and further supports the importance of Tde loading onto cognate VgrG in T6SS assembly.
Figure EV2. Expression levels of truncated VgrG1 variants affected Hcp secretion.
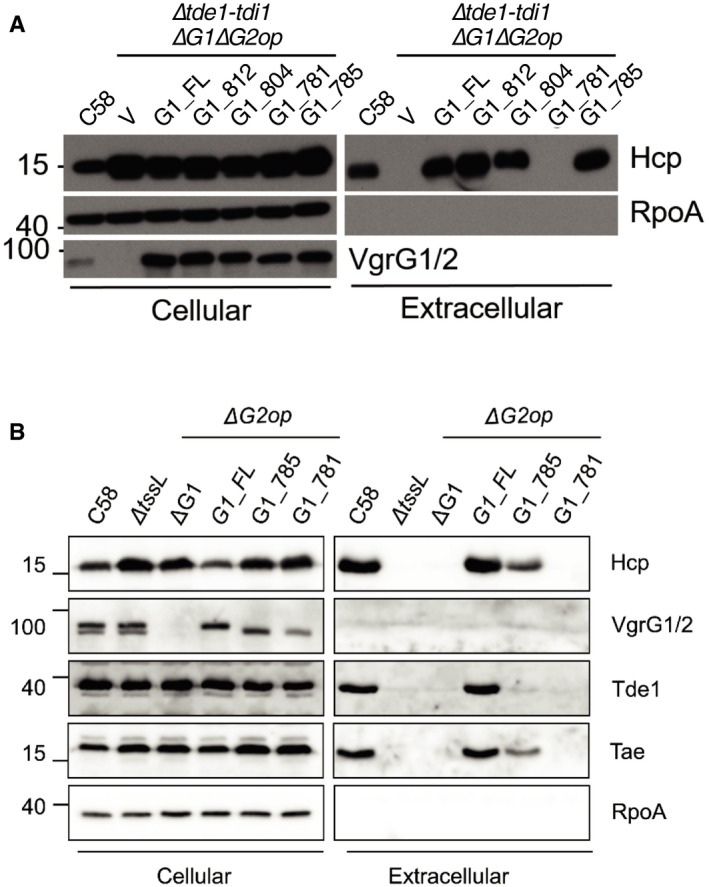
- T6SS secretion assay of Agrobacterium tumefaciens C58 ΔtdeiΔvgrG1ΔvgrG2 (ΔtdeiΔG1ΔG2op mutant) harboring a pRL662 vector (V) or its derivatives overexpressing full‐length and truncated VgrG1 proteins.
- T6SS secretion assay of A. tumefaciens C58ΔG2op mutant encoding full‐length and truncated VgrG1 proteins.
The absence of Tde effectors may have affected VgrG stability and compromised the efficiency of T6SS assembly because the intensities of the signals for the truncated VgrG1 (G1_785 and G1_781) variants are slightly lower than that of full‐length VgrG1 (Fig 2C). Thus, we quantified the change in levels of VgrG1/2 in wild‐type and Δtdei strains after adding chloramphenicol, an inhibitor of translation. The levels of VgrG1/2 decayed at similar rates in the two strains (Figs EV3A and B). Therefore, the stability and steady‐state levels of VgrG1 and VgrG2 proteins do not appear to be affected by the presence or absence of their cognate Tde effector or their cognate adaptor/chaperone (Figs 1 and 2). The data above demonstrated that loading cargo effectors onto cognate VgrG proteins is important for T6SS assembly and for ejecting the Hcp tube as well as VgrG spike proteins from A. tumefaciens strain C58.
Figure EV3. VgrG protein stability is not affected by the absence of Tde‐Tdi pairs.
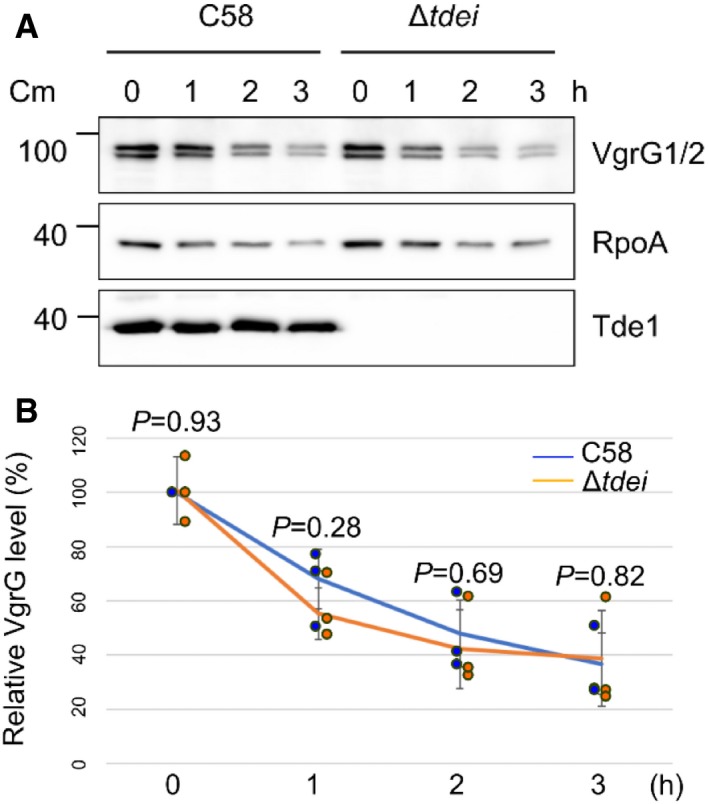
- At times indicated, total protein fractions were collected from wild‐type C58 or Δtdei grown in 523 medium with chloramphenicol and analyzed in Western blots probed with antibodies for indicated proteins.
- The intensities of bands corresponding to VgrG were quantified using ImageJ software. Intensity values were normalized to those of VgrG of wild‐type C58 at 0 h. Data are mean ± SD of three independent experiments. P values were calculated using unpaired two‐sided t‐test, and no significant difference was detected between C58 and Δtdei as indicated (P value > 0.1).
Source data are available online for this figure.
The presence of Tde effectors is required for efficient TssBC sheath assembly
Given these findings, we next tested whether the presence of Tde effectors is critical to initiate Hcp polymerization and assembly of the TssBC sheath. TssB fused to a fluorescent protein has been shown to assemble into the TssBC sheaths that were observed as fluorescent foci or streaks across the cell width 6, 18. The sheaths were reported to be dynamical contractile structures that were able to contract and disassemble after the extended sheaths were formed 6, 18. Thus, these fluorescent structures under the microscope can be used as an indicator of the sheath polymerization and T6SS assembly. Therefore, a plasmid expressing TssB with a C‐terminal fusion to green fluorescence protein (TssB‐GFP) was expressed in the ΔtssB, ΔtssLΔtssB, and ΔtdeiΔtssB mutant strains of A. tumefaciens C58. TssB‐GFP in ΔtssB partially restored T6SS‐mediated secretion, as determined on the basis of the amount of secreted Hcp relative to that of the ΔtssB expressing an unmodified variant of TssB (Fig EV4). The protein abundance of TssB‐GFP in different strains was similar, as determined on the basis of band intensities of TssB‐GFP and its truncated form (Fig EV4). Similar to the previous reports 6, the sheaths were often observed as foci or streaks across the cell width. Moreover, these fluorescent structures were observed in the vast majority of the ΔtssB (TssB‐GFP) cells (Fig 3A). We then counted the number of fluorescent foci in each strain (Fig 3B). The results showed that the ΔtssB (TssB‐GFP) strain had slightly more than one fluorescent structure per cell, and the ΔtssBΔtdei (TssB‐GFP) strain had very few fluorescent structures (2–5 foci out of 100 cells). The fluorescent streaks were rarely seen in the later strain (Fig 3A and B). In contrast, no streaks were found in the negative control strain, ΔtssBΔtssL (TssB‐GFP; Fig 3A and B).
Figure EV4. C‐terminal GFP‐tagged TssB partially complemented Hcp secretion in ΔtssB .
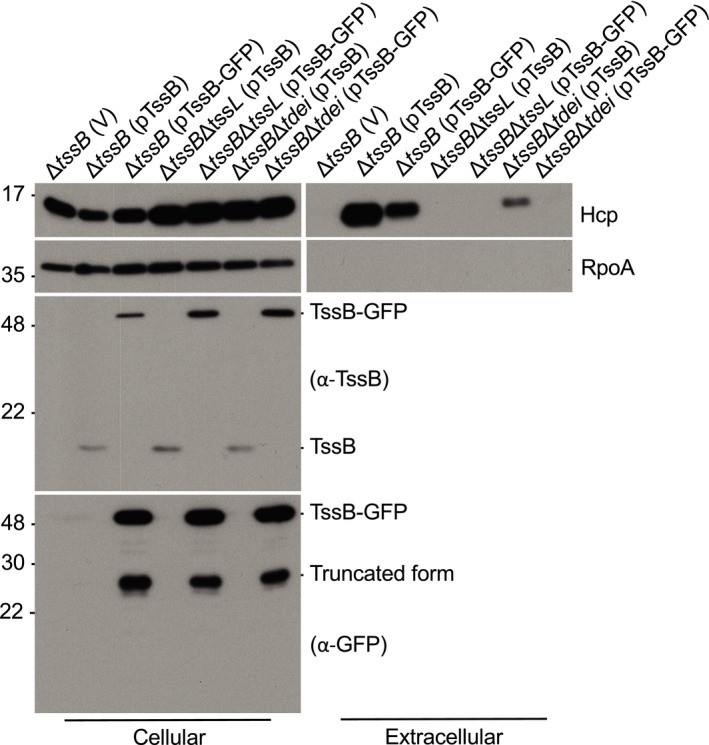
Various Agrobacterium tumefaciens strains were grown in I‐medium (pH5.5), and the cellular and extracellular (S) fractions were collected for Western blotting probed with antibodies for indicated proteins. Molecular weight markers (in kDa) are indicated on the left. Data are from one independent experiment and reproduced in at least two independent experiments.Source data are available online for this figure.
Figure 3. The presence of Tde effectors is required for efficient TssBC sheath assembly.
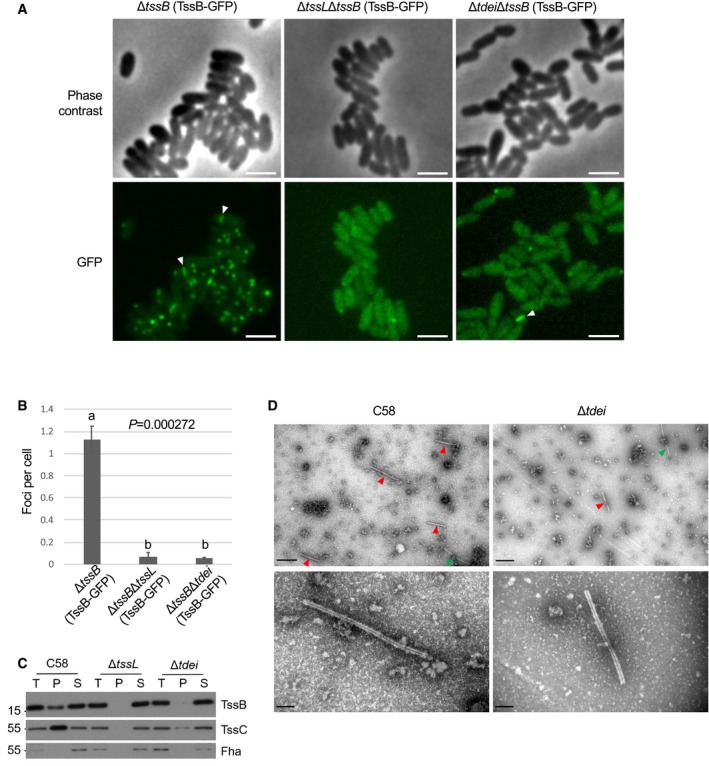
- Representative images of cells of Agrobacterium tumefaciens C58 strains ΔtssB, ΔtssLΔtssB, and ΔtdeiΔtssB cells each expressing TssB‐GFP from pTrc200. Upper panel: phase contrast images; lower panel: green fluorescence images. Examples of GFP streaks were indicated by white arrowheads. Scale bar: 2 μm.
- Quantification of TssB‐GFP foci in different genetic background. The number of the fluorescent foci was counted using the “analysis particle” function in Fiji with a manually set threshold. The number of cells was counted manually. For each strain, a total of three images were obtained from one independent experiment and each image contained more than 300 cells for quantification and statistical analysis. One‐way ANOVA followed by Turkey HSD test was used for statistical analysis. Data are mean ± SEM, and two groups with significant differences (P = 0.000272) are indicated with different letters (a and b). Similar results are reproduced in two independent experiments.
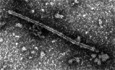
- Western blots of the isolated sheaths, which were prepared via ultracentrifugation of cell lysates from A. tumefaciens C58 wild‐type, ΔtssL, and Δtdei. T: total proteins from the cell lysate. P: pellet samples containing TssBC sheaths after ultracentrifugation. S: supernatant after ultracentrifugation. Proteins were analyzed in Western blots probed with indicated antibodies. Fha serves as a cytoplasmic control. Molecular weight markers (in kDa) are indicated on the left. Similar results were obtained from at least two independent experiments. Data are from one independent experiment and reproduced in at least two independent experiments.
- Pellet samples of wild type and Δtdei were visualized with transmission electron microscopy (TEM). Sheaths are indicated by red arrowheads, and flagella, which are distinguishable on the basis of smoother, more solid, and thinner structures (˜ 13 nm in width), are indicated by green arrowheads 33. Data are from one independent experiment and reproduced in at least two independent experiments. Scale bar: 0.5 μm (upper panel), 100 nm (upper panel).
Source data are available online for this figure.
We also isolated and characterized T6SS sheaths, which are tubular structures formed by TssB and TssC proteins 6. Pellets and supernatants from crude protein extracts after sheath preparation procedure of the wild‐type, ΔtssL, and Δtdei strains of A. tumefaciens C58 were first analyzed by Western blots (Fig 3C). The presence of TssB and TssC proteins in the pellet indicates that they have polymerized into a sheath, whereas monomers/non‐polymerized subunits remain in the supernatant. TssB and TssC proteins were detected in the pellet fraction of the wild type but not of the ΔtssL mutant. The amounts of TssB and TssC detected in the pellet fractions of Δtdei were substantially lower than those detected in the wild type, suggesting that the TssBC sheaths were inefficiently formed in the absence of Tde effector‐encoding genes.
The pellet fractions were further characterized via transmission electron microscopy (TEM). Sheath‐like structures were frequently detected in fractions from C58, and only very few were observed in fractions from Δtdei (Fig 3D). No sheath structures were identified from the fraction from ΔtssL. Regardless of the source, the sheaths were similar in structure. The diameter of the sheath structure was calculated to be ~ 30 nm. They have a hollow lumen, suggesting these were contracted sheathes which had ejected the Hcp tube. Relative to sheaths of other bacteria, those from A. tumefaciens have a similar morphology and diameter with the ones reported in other bacteria (25–33 nm) 6, 19, 20, 21, 22. The data together suggest that effector‐loaded VgrG is an important trigger for efficient TssBC sheath assembly. Such a mechanism may be employed by A. tumefaciens to prevent the assembly of the T6SS machine when effectors are not present and/or loaded.
Effector loading onto cognate VgrG carrier is critical for Hcp/VgrG secretion in different A. tumefaciens genomospecies
Agrobacterium tumefaciens is recognized as genetically diverse and subdivided into genomospecies, which are equivalent to species‐level groups 23. We therefore tested whether loading of cargo effector onto VgrG to activate T6SS is also used by other A. tumefaciens genomospecies/strains. Several strains of A. tumefaciens encode functional T6SSs that are necessary for interbacterial competition 24. From these, A. tumefaciens strains 1D1108, 15955, and 12D1, which belong to different genomospecies with only ~ 80% gene content similarity to C58, were selected to test whether effector loading onto VgrG to regulate T6SS occurs in genetically diverse A. tumefaciens 24. These three strains each carry a T6SS gene cluster similar to C58 but each has only a single vgrG module that circumscribe several downstream genes (v1–6 for 12D1, v1–9 for 1D1108, v1–7 for 15955; Fig 4A) 24. Strains 12D1 and 15955 contain Tap‐1 homolog (encoded by v1 in 12D1 and v5 in 15955) and 1D1108 v4 encodes a DUF2169‐containing protein, the three of which are located upstream of known or predicted toxin–immunity gene pairs (v3–4 in 12D1, v6–7 in 1D1108, and v6–7 in 15955) distinct among them and C58.
Figure 4. Impacts of vgrG‐associated genes on T6SS secretion and antibacterial activity of Agrobacterium tumefaciens strains other than C58.
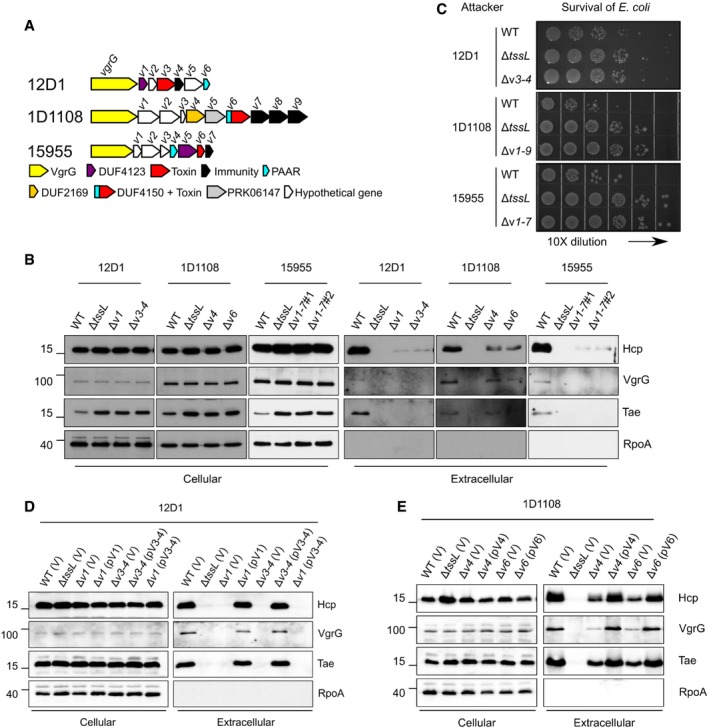
- The vgrG genetic modules of the tested A. tumefaciens strains. Genes are color‐coded according to the predicted function or results of functional assays 24. The vgrG‐associated genes are v1–v6 for 12D1, v1–v9 for 1D1108, and v1–v7 for 15955.
- Secretion assay of wild type and various mutants of 12D1, 1D1108, and 15955.
- Antibacterial activity assay of wild type and mutants of 12D1, 1D1108, and 15955 was carried out in a ratio of 30:1 against Escherichia coli harboring the plasmid pRL662. The target E. coli cells were serially diluted and grown overnight on gentamicin‐containing LB agar prior to photographing. Each competition was done at least four times and reproduced in three independent experiments.
- Secretion assay of wild‐type 12D1, ΔtssL, Δv1, Δv3–4 containing pRL662 empty vector (V) or its derivatives with v1 or v3–4 were analyzed for secretion.
- Secretion assay of wild‐type 1D1108, ΔtssL, Δv4, Δv6 containing pRL662 empty vector (V) or its derivatives with v4 or v6.
Mutants lacking genes encoding predicted toxin and/or adaptor/chaperone had substantially reduced levels of Hcp and VgrG proteins in extracellular fractions, as compared to that of their respective wild‐type strains (Fig 4B). Consistent with the secretion results, interbacterial competition assays showed that these mutants were as compromised as their corresponding ΔtssL mutants in competing with E. coli (Fig 4C). Expression of the adaptor or toxin/immunity genes in their respective mutants restored the ability of the cells to secrete Hcp, VgrG, and Tae to that of wild‐type levels (Fig 4D and E). Expression of the v3–4 toxin–immunity pair in the Δv1 adaptor/chaperone mutant of 12D1 did not restore any secretion activity. These results support the conclusion that the efficient assembly of T6SS requires an adaptor/chaperone. Thus, effector loading is a common mechanism for regulating T6SS across the A. tumefaciens group of bacteria and perhaps also in other bacterial species.
Effector loading mechanisms are differentially regulated by the conserved Tae toxin
The highly diminished Hcp/VgrG secretion levels in Δtdei, Δtde2‐tdi2Δtae‐tai, and Δ3TIs are surprising because in a previous study, the level of secreted Hcp in the Δ3TIs mutant was similar to that of wild‐type C58 10. A key difference between this previous study and the one presented here is that, in the previous study, the secretion assay was conducted in an acidic minimal medium (I‐medium, pH 5.5), whereas here we used a rich medium (523) for unambiguous detection of VgrG secretion. Therefore, we carried out secretion assays for single, double, and triple toxin–immunity deletion strains in I‐medium (pH 5.5). As reported previously, extracellular Hcp levels in Δ3TIs are similar to those from wild‐type C58. However, Hcp was barely detected in Δtdei lacking both tde1‐tdi1 and tde2‐tdi2 (Fig 5A). Expression of Tae alone or the Tae‐Tai pair from a plasmid in Δ3TIs reduced Hcp secretion levels, while Tai expression in Δ3TIs did not impact levels of extracellular Hcp in Δ3TIs (Fig 5B). These results suggested a role of Tae in regulating Hcp secretion levels when grown in different growth conditions.
Figure 5. Effect of tae on T6SS secretion and antibacterial activity in the C58 Δtdei mutant grown in minimal medium.
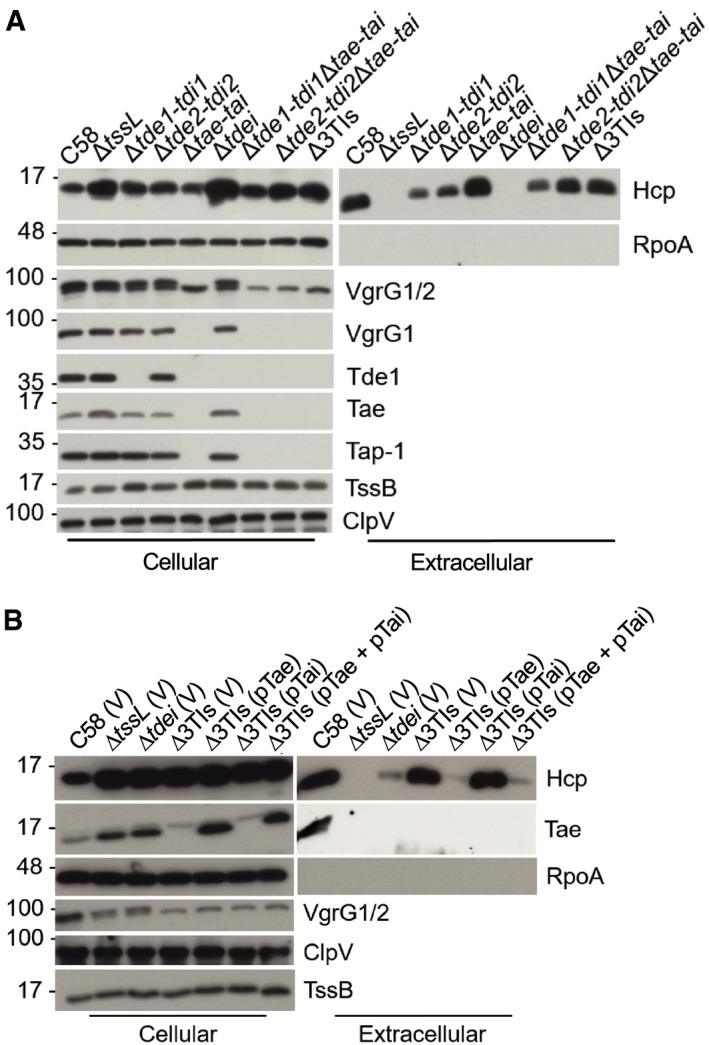
- T6SS secretion assay of Agrobacterium tumefaciens wild‐type C58, ΔtssL, and various single, double, and triple toxin–immunity deletion strains.
- T6SS secretion assay of A. tumefaciens wild‐type C58, ΔtssL, Δtdei, and Δ3TIs harboring the indicated plasmids.
During the course of this study, we also noticed that in Δtae‐tai, downstream‐encoded proteins (VgrG1, Tap‐1, and Tde1 proteins) were not detected in either of the cellular or extracellular fractions, while imp operon‐encoded TssB and ClpV‐encoded upstream of tae‐tai were detected in all analyzed strains (Figs 1B and EV1C). This suggests that Δtae‐tai has a polar effect and explains why only VgrG2 but not VgrG1 was detected in Δtae‐tai and Δtde2‐tdi2Δtae‐tai mutants. To investigate whether the differential role of Tae in regulating Hcp secretion in different growth media also occurs in another A. tumefaciens strain, we made a deletion of tae‐tai gene pair in both WT and Δv3–v4 of 12D1 to determine whether the loss of tae‐tai can impact Hcp secretion in I‐medium. As expected, Hcp secretion was highly reduced in Δv3–v4 as compared to that of WT and Δtae‐tai, which supports the requirement of VgrG cargo effector for efficient Hcp secretion in both rich and minimal media (Fig 6A). To our surprise, deletion of tae‐tai from Δv3–v4 (Δv3–v4Δtae‐tai) of 12D1 did not restore Hcp secretion, whereas complementing v3–v4 in both Δv3–v4 and Δv3–v4Δtae‐tai strains with v3–v4 restored Hcp secretion to wild‐type levels (Fig 6B). Accordingly, antibacterial activity against E. coli was correlated with active and wild‐type levels of Hcp secretion (Fig 6C). In addition, the Δtap‐1Δ3641 mutant of C58 with deletions of both adaptor genes for Tde1 and Tde2 effectors also remained deficient in Hcp secretion when grown in minimal medium (Fig 6D). These results reinforced the requirement of cargo effector loading onto VgrG for assembling the T6SS and ejecting toxin effector for killing competing bacterial cells in different growth environments.
Figure 6. Impact of tae‐tai on T6SS secretion and antibacterial activity of Agrobacterium tumefaciens strains 12D1 and C58 grown in minimal medium.
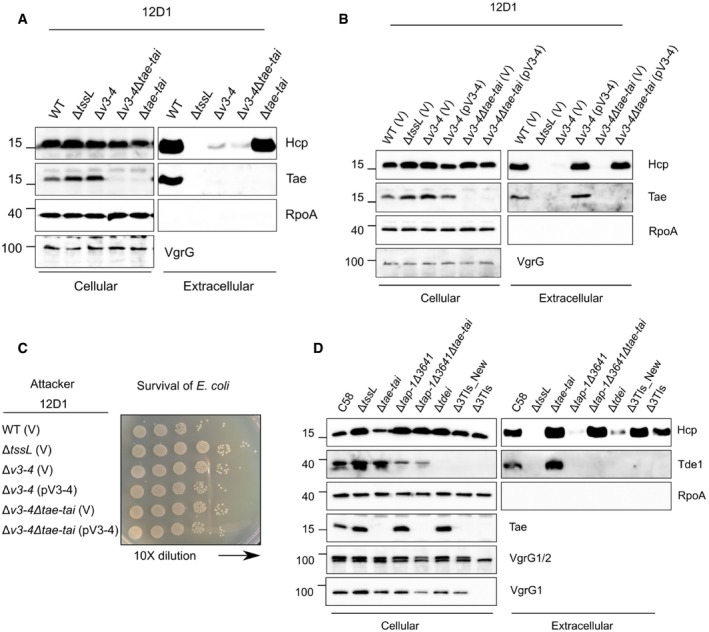
- T6SS secretion assay of A. tumefaciens 12D1 wild‐type, ΔtssL, and various toxin–immunity deletion strains.
- T6SS secretion assay of A. tumefaciens 12D1 wild‐type, ΔtssL, and various toxin–immunity deletion strains harboring the indicated plasmids.
- Antibacterial activity assay of A. tumefaciens 12D1 wild‐type, ΔtssL, and various toxin–immunity deletion strains harboring the indicated plasmids was carried out in a ratio of 30:1 against Escherichia coli harboring the plasmid pRL662. The target E. coli cells were serially diluted and grown overnight on gentamicin‐containing LB agar prior to photographing. Each competition was done at least four times and reproduced in two independent experiments.
- T6SS secretion assay of A. tumefaciens C58 wild‐type, ΔtssL, and various mutant strains.
Interestingly, deletion of the tae‐tai gene pair did not cause a polar effect in 12D1 as the VgrG protein was detected at wild‐type levels in Δtae‐tai and Δv3–v4Δtae‐tai mutants (Fig 6A and B). This result motivated us to revisit the polar effect of Δtae‐tai and Δ3TIs observed in C58; tae was truncated in these strains and left only 20 nucleotides upstream of the 3′ vgrG gene (Fig EV5). We generated a new tae‐tai deletion in the WT, Δtdei, and Δtap‐1Δ3641 background of C58, designated as Δtae‐tai*, Δ3TIs*, and Δtap‐1Δ3641Δtae‐tai*, respectively. This mutant was truncated in tae and left 77 nucleotides upstream of vgrG (Fig EV5). The newly constructed truncation in Δtae‐tai*, Δ3TIs*, and Δtap‐1Δ3641Δtae‐tai* did not have polar effects as VgrG1 and Tde1 proteins were detected, which, however, retained the ability to secrete Hcp (Fig 6D).
Figure EV5. Design of the non‐polar tae‐tai deletion mutation in C58.
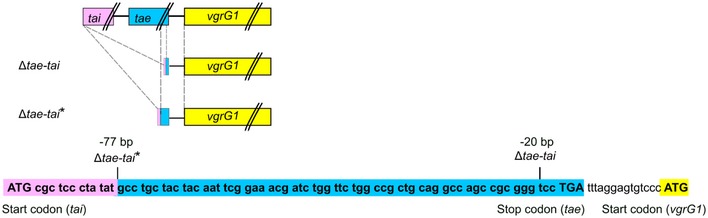
The C‐terminal coding portion of the tai‐tae locus and N‐terminal coding portion of vgrG1 (vgrG) are presented. A deletion mutant of tae‐tai was constructed to retain 77 nucleotides upstream of the start codon of the vgrG1 coding sequence (highlighted in blue). This is predicted to generate an in‐frame deletion of tai‐tae. The original tae‐tai deletion retained only 20 nucleotides upstream of the start codon of the vgrG1 coding sequence.
The above evidence indicates that the presence of Tae toxin alone or Tae‐Tai could have different impacts on T6SS assembly and subsequent Hcp secretion under different scenario. In C58, when A. tumefaciens is grown in rich medium, T6SS is only assembled when the cargo effector is loaded onto its cognate VgrG regardless of the presence or absence of Tae effector. In minimal medium, the loading of cargo effector onto its VgrG is irrelevant for T6SS assembly if Tae is absent (e.g., Δ3TIs). However, in the presence of Tae or Tae‐Tai pairs, which are highly conserved across A. tumefaciens genomospecies 24, the Tae effector ensures that the T6SS is fully assembled only when VgrG is loaded with effectors. In 12D1, the T6SS is only assembled when cargo effector is loaded onto its cognate VgrG regardless of the presence or absence of Tae‐Tai pair either grown in rich or minimal media. Thus, effector loading mechanisms are differentially regulated by the conserved Tae toxin. The rationale of Tae effector with different impacts in Hcp secretion under different scenario and the underlying mechanisms of how Tae effector controls T6SS assembly are beyond the scope of this study and await future investigation.
Discussion
In this study, we describe a new mechanism that applies across the A. tumefaciens group of bacteria. Cargo effector loading onto its cognate VgrG is critical for T6SS assembly and ejecting toxin effectors to kill competitor bacterial cells. Such a mechanism may be an energy‐saving strategy deployed by bacteria to ensure efficacious T6SS firing. However, this mechanism may have been overlooked in many other species of bacteria, because of the lack of mutants deleted of all effector‐encoding genes. Likewise, in other species, the two functions of VgrG, delivering effectors and being a structural feature of the T6SS, have not been uncoupled. A recent study in Vibrio cholerae independently reported that recruiting effector onboard is crucial for T6SS assembly 25. Thus, regulation of the T6SS via effector loading onto VgrG is potentially a widespread mechanism, which may be deployed by many T6SS‐possessing bacteria to influence their fitness and composition of their communities. In addition, the presence of Tde2 variant with catalytic site mutation retains the ability to trigger Hcp secretion in this study is consistent with the findings that physical presence but not the enzymatic activity of three V. cholerae T6SS effectors is required for T6SS assembly 25. These studies together demonstrated it is the effector loading rather than its enzymatic activity is the trigger in T6SS assembly.
Previous studies provided evidence that VgrG interacts with components of the T6SS baseplate and the interactions are critical for assembly of the T6SS 7, 8, 26, 27. Our findings further suggest that effector loading onto VgrG spike is the key in completing T6SS assembly. The current knowledge led us to propose hypothetical models illustrating the effector loading mechanisms (Fig 7). Model I depicts that effector‐loaded VgrG exhibits higher affinity than VgrG itself for recruitment onto the T6SS baseplate complex therefore having a higher chance to trigger the T6SS assembly. Alternatively, effector loading onto the VgrG‐baseplate complexes may be the trigger for Hcp polymerization and/or TssBC sheath assembly (Model II). In the absence of either cargo effectors or adaptors, VgrG may not be loaded onto the baseplate or the VgrG‐baseplate complex is incapable of triggering T6SS assembly. This new concept opens future research to investigate how widespread of the T6SS effector loading mechanisms across different bacterial groups and the underlying mode of action.
Figure 7. Proposed models of T6SS effector loading mechanisms.
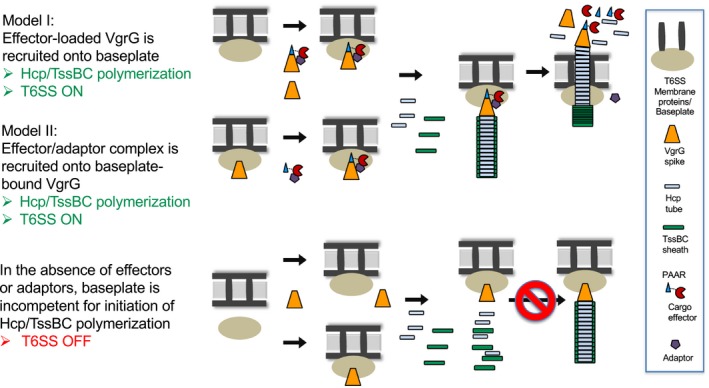
Model I: the VgrG‐PAAR–effector–adaptor complex forms first before binding to the membrane‐associated baseplate complex. Model II: VgrG binds to baseplates first prior to be loaded with the effector–adaptor complex. In both, the formation of an effector‐loaded VgrG‐baseplate complex is critical for initiating the polymerization of the Hcp and TssBC sheath and ejecting the effector complex outward. In the absence of either cargo effectors or adaptors, VgrG may not bind to the baseplate efficiently or the VgrG‐baseplate complex is incompetent for triggering T6SS assembly. The effector shown in this model represents a PAAR/DUF4150‐fused effector (such as Tde2 of C58, V6 of 1D1108) but also applicable to those where the effector and PAAR are encoded in separate genes (such as Tde1 of C58, V3 of 12D1, V6 of 15955), based on the evidence of VgrG‐PAAR‐Tde1‐Tap‐1 complex formation shown previously 16. The adaptor protein is depicted as being released from the associated effector but the step which happens remains to be determined. The proposed models do not consider the role of Tae that may differentially regulate the outcome of effector loading in T6SS assembly.
Materials and Methods
Bacterial strains, growth conditions, and molecular techniques
Bacterial strains and sequences of primers used in this study are listed in Appendix Tables S1 and S2, respectively. Agrobacterium tumefaciens was grown in 523 medium at 25°C, and E. coli was grown in LB medium at 37°C, unless otherwise indicated 10, 16, 17. Antibiotics and concentrations used were as follows: gentamycin (50 μg/ml for A. tumefaciens and 30 μg/ml for E. coli) and spectinomycin (200 μg/ml).
DNA preparation and plasmids construction
Plasmid DNA was extracted using Presto Mini Plasmids Kit (Geneaid, Taiwan). 2× manufacturer instructions were followed in using Ready Mix A (Zymeset, Taiwan) for polymerase chain reactions (PCRs). For the construction of plasmids used for mutant generation, ~ 500 bp of the upstream and downstream sequence of gene targeted for deletion/truncation were amplified from genomic DNA with the primer sets and cloned into XbaI‐ and BamHI‐digested pJQ200KS plasmid. For the construction of pTrc‐TssB‐GFP, A. tumefaciens tssB and the upstream sequences corresponding to the ribosome binding site were amplified from pTrc‐TssB (EML4043; Appendix Table S1) with primers tssB_XmaI_F and tssB‐GFP_R (Appendix Table S2). The gfp gene was amplified from pBBR1‐GFP (EML3) 28 using primers GFP_HindIII_R and tssB‐GFP_F. The two amplified products were fused together in a second PCR that used primers tssB_XmaI_F and GFP_HindIII_R, which include restriction site sequences for XmaI and HindIII. The purified fusion product and pTrc200 plasmid were digested with XmaI and HindIII‐HF (New England Biolabs, Ipswich, USA) and subsequently ligated together using T4 DNA ligase (New England Biolabs, Ipswich, USA). For plasmid pV3–4 (12D1), the PCR product of v3–4 from 12D1 was digested with EcoRI and BamHI, ligated to pRL662 digested with the same restriction enzymes, and transformed into E. coli strain DH10B. The following plasmids were generated by Gateway cloning. The PCR products of tde2‐tdi2, v1 from 12D1, and v4 and v6 from 1D1108 were recombined to pDONR222 vector via Gateway BP reaction to generate entry clones (Invitrogen, Carlsbad, CA). Constructs were verified via PCR. The genes from the entry clones were recombined via Gateway LR reaction into pRL662_RfC.1 or pTrc200_RfC.1 and transformed into E. coli strain DH10B. All the plasmid construct was confirmed via colony PCR, enzyme digestion, sequencing, and Western blot of A. tumefaciens cells harboring the plasmid.
Mutant construction
The pJQ200KS suicide plasmid 29 and double crossover method were used to generate in‐frame deletions of A. tumefaciens genes 30. In brief, cells were electroporated with suicide plasmids; transformants were selected on 523 agar plates containing gentamycin without sucrose. The Gm‐resistant colonies were cultured overnight in LB broth without Gm, serially diluted and spread onto 523 agar plates containing 5% sucrose without Gm to enrich for bacterial cells that had undergone a second crossover event. The deletion mutants were confirmed via colony PCR and Western blot analyses.
Secretion assay
Secretion assays were performed as described 16. Briefly, A. tumefaciens strains were cultured in 523 medium overnight and subcultured, using an initial cell density of OD600 nm = 0.2, in I‐medium (pH 5.5) or 523 medium, depending on the design of the experiments. After 6 h of subculturing, the samples were centrifuged at 10,000 g for 10 min to separate the extracellular and the cellular fractions. The cell pellets were adjusted to OD600 = 5.0, and the respective extracellular fractions were filtered through low protein‐binding 0.22 μm sterilized filter units (Millipore, Tullagreen, Ireland) and proteins were precipitated in trichloroacetic acid 15. Western blot analyses were done as previously described 10, 16, 17.
Interbacterial competition assay
Methods for interbacterial competition assays were previously described 10. Briefly, A. tumefaciens strains were co‐cultured at with E. coli K‐12 cells harboring the plasmid pRL662 (conferring gentamycin resistance), at a ratio of 30:1 on LB agar. The surviving E. coli cells were serially diluted, spotted, or quantified by counting colony‐forming units (CFUs) on gentamycin‐containing LB agar plates. Statistics were calculated using one‐way ANOVA and Tukey's honestly significance difference (HSD) test (http://astatsa.com/OneWay_Anova_with_TukeyHSD/).
Protein stability assay
For VgrG protein stability analysis, A. tumefaciens cells were resuspended at OD600 ∼ 0.5 in 523 medium containing chloramphenicol (100 μg/ml) to inhibit protein synthesis. Cells were harvested after 1, 2, and 3 h and adjusted to OD600 nm = 5.0 for Western blot analysis 31. Intensities of specific protein bands were quantified using ImageJ software.
T6SS sheath preparation and transmission electron microscopy (TEM)
Isolation of T6SS sheaths was performed by the following methods previously described 6. In brief, A. tumefaciens cells were cultured overnight in 5 ml 523 and subcultured into 50 ml I‐medium at 25°C for 6 h. The cells were harvested and lysed for 15 min at 37°C in 4 ml of buffer containing 0.5× CelLytic B (Sigma‐Aldrich, St. Louis, USA), 150 mM NaCl, 50 mM Tris pH 7.4, lysozyme (500 μg/ml), DNase I (50 μg/ml), 1 mM phenylmethylsulfonyl‐fluoride (PMSF), and 0.05% Triton X‐100. Cell debris was removed via centrifugation at 10,000 g, 10 min at 4°C. The high‐molecular‐weight complexes were separated from the clear cell lysate via ultracentrifugation at 150,000 g for 1 h at 4°C. Pellet fractions were resuspended in 150 μl buffer containing 150 mM NaCl, 50 mM Tris pH 7.4, and 0.75 μl Protease Inhibitor Cocktail Set III (EMD Millipore, Pacific Center Court San Diego, USA). The samples were analyzed via Western analyses. For TEM, 10 μl of the samples was deposited on copper grids with carbon‐formvar film support. After 3 min, samples were negative stained for 90 seconds with 25% uranyl acetate. A Tecnai G2 Spirit TEM (FEI, Hillsboro, USA) set at 80 kV was used to visualize samples.
Fluorescence microscopy
Cells were cultured in 523 medium overnight followed by subculturing in I‐medium for 3 h. A total of 1 ml of each bacterial culture was fixed with 8 μl 25% glutaraldehyde and 125 μl 37% formaldehyde for 20 min and washed twice with PBS (Biomate, Taiwan) with additional 0.5% Tween‐20 (PBST). Bacterial cells were resuspended with 30 μl PBST. Three microliter of the fixed cell suspension was deposited onto agarose pad (PBST with 2% agarose) to reduce random movement of cells. An upright microscope BX61 (Olympus, Tokyo, Japan) equipped with a CMOS camera (C11440 ORCA‐R2 Flash 2.8, Hamamatsu, Japan), objective lens (UPlanFLN 100×/1.30, Olympus, Tokyo, Japan), and a GFP filter set (Part number: FITC‐3540C‐000, Semwork, New York, USA) were used. Images were acquired and processed using Improvision Volocity 6.3 software (Perkin Elmer, Waltham, USA) and Fiji 32, respectively.
Author contributions
C‐FW, Y‐WL, and E‐ML conceived and designed the experiments. Y‐WL, C‐FW, and J‐SL performed the experiments and analyzed the data. DB, MP, and Y‐LS provided the tools. JHC, Y‐LS, MP, and E‐ML supervised the execution of the experiments. Y‐WL, E‐ML, and JHC, with contributions from C‐FW, J‐SL, MP, and Y‐LS, wrote the paper. All authors read and approved the paper.
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting information
Appendix
Expanded View Figures PDF
Source Data for Expanded View Figures
Review Process File
Source Data for Figure 1
Source Data for Figure 2
Source Data for Figure 3
Source Data for Figure 4
Source Data for Figure 5
Source Data for Figure 6
Acknowledgements
The authors thank Romain Kooger in the Institute of Molecular Biology & Biophysics, ETH Zürich, for discussions and providing the protocol for preparing TssBC sheaths. We acknowledge the assistance by Claudia Parada of the Institute of Biological Chemistry and staff in the Plant Cell Biology Core Laboratory as well as DNA Sequencing Core Laboratory at the Institute of Plant and Microbial Biology, Academia Sinica, Taiwan. The authors also thank Lay‐Sun Ma, Chih‐Horng Kuo, and Romain Kooger for critically reading the manuscript, Nia Santo and Manda Yu for assistance in preparing the revision, and the members of Lai laboratory for their discussions and suggestions. Funding for this project was provided by the Ministry of Science and Technology of Taiwan (MOST; grant no. 104‐2311‐B‐001‐025‐MY3 and 107‐2311‐B‐001‐019‐MY3 to E‐ML and 106‐2311‐B‐001‐009 to Y‐LS). Work in the Chang laboratory is supported in part by the National Institute of Food and Agriculture, US Department of Agriculture award 2014‐51181‐22384. MP is supported by the Swiss National Science Foundation (grant no. 179255) and the European Research Council (grant no. 679209). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
EMBO Reports (2020) 21: e47961
References
- 1. Russell AB, Peterson SB, Mougous JD (2014) Type VI secretion effectors: poisons with a purpose. Nat Rev Microbiol 12: 137–148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Lien Y‐W, Lai E‐M (2017) Type VI secretion effectors: methodologies and biology. Front Cell Infect Microbiol 7: 254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Ringel PD, Hu D, Basler M (2017) The role of type VI secretion system effectors in target cell lysis and subsequent horizontal gene transfer. Cell Rep 21: 3927–3940 [DOI] [PubMed] [Google Scholar]
- 4. Bernal P, Llamas MA, Filloux A (2018) Type VI secretion systems in plant‐associated bacteria. Environ Microbiol 20: 1–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Verster AJ, Ross BD, Radey MC, Bao Y, Goodman AL, Mougous JD, Borenstein E (2017) The landscape of type VI secretion across human gut microbiomes reveals its role in community composition. Cell Host Microbe 22: 411–419.e414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Basler M, Pilhofer M, Henderson GP, Jensen GJ, Mekalanos JJ (2012) Type VI secretion requires a dynamic contractile phage tail‐like structure. Nature 483: 182–186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Brunet YR, Zoued A, Boyer F, Douzi B, Cascales E (2015) The type VI secretion TssEFGK‐VgrG phage‐like baseplate is recruited to the TssJLM membrane complex via multiple contacts and serves as assembly platform for tail tube/sheath polymerization. PLoS Genet 11: e1005545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Zoued A, Durand E, Brunet YR, Spinelli S, Douzi B, Guzzo M, Flaugnatti N, Legrand P, Journet L, Fronzes R et al (2016) Priming and polymerization of a bacterial contractile tail structure. Nature 531: 59–63 [DOI] [PubMed] [Google Scholar]
- 9. Cianfanelli FR, Monlezun L, Coulthurst SJ (2016) Aim, load, fire: the type VI secretion system, a bacterial nanoweapon. Trends Microbiol 24: 51–62 [DOI] [PubMed] [Google Scholar]
- 10. Ma L‐S, Hachani A, Lin J‐S, Filloux A, Lai E‐M (2014) Agrobacterium tumefaciens deploys a superfamily of type VI secretion DNase effectors as weapons for interbacterial competition in planta. Cell Host Microbe 16: 94–104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Flaugnatti N, Le TTH, Canaan S, Aschtgen M‐S, Nguyen VS, Blangy S, Kellenberger C, Roussel A, Cambillau C, Cascales E et al (2016) A phospholipase A1 antibacterial Type VI secretion effector interacts directly with the C‐terminal domain of the VgrG spike protein for delivery. Mol Microbiol 99: 1099–1118 [DOI] [PubMed] [Google Scholar]
- 12. Cianfanelli FR, Alcoforado Diniz J, Guo M, De Cesare V, Trost M, Coulthurst SJ (2016) VgrG and PAAR proteins define distinct versions of a functional type VI secretion system. PLoS Pathog 12: e1005735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. English G, Trunk K, Rao VA, Srikannathasan V, Hunter WN, Coulthurst SJ (2012) New secreted toxins and immunity proteins encoded within the Type VI secretion system gene cluster of Serratia marcescens . Mol Microbiol 86: 921–936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Hood RD, Singh P, Hsu F, Güvener T, Carl MA, Trinidad RRS, Silverman JM, Ohlson BB, Hicks KG, Plemel RL et al (2010) A type VI secretion system of Pseudomonas aeruginosa targets a toxin to bacteria. Cell Host Microbe 7: 25–37 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Wu H‐Y, Chung P‐C, Shih H‐W, Wen S‐R, Lai E‐M (2008) Secretome analysis uncovers an Hcp‐family protein secreted via a type VI secretion system in Agrobacterium tumefaciens . J Bacteriol 190: 2841–2850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Bondage DD, Lin J‐S, Ma L‐S, Kuo C‐H, Lai E‐M (2016) VgrG C terminus confers the type VI effector transport specificity and is required for binding with PAAR and adaptor–effector complex. Proc Natl Acad Sci USA 113: E3931–E3940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Lin J‐S, Ma L‐S, Lai E‐M (2013) Systematic dissection of the agrobacterium type VI secretion system reveals machinery and secreted components for subcomplex formation. PLoS ONE 8: e67647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Basler M, Mekalanos JJ (2012) Type 6 secretion dynamics within and between bacterial cells. Science (New York, N.Y.) 337: 815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lossi NS, Manoli E, Förster A, Dajani R, Pape T, Freemont P, Filloux A (2013) The HsiB1C1 (TssB‐TssC) complex of the Pseudomonas aeruginosa type VI secretion system forms a bacteriophage tail sheathlike structure. J Biol Chem 288: 7536–7548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wang J, Brackmann M, Castaño‐Díez D, Kudryashev M, Goldie KN, Maier T, Stahlberg H, Basler M (2017) Cryo‐EM structure of the extended type VI secretion system sheath–tube complex. Nat Microbiol 2: 1507–1512 [DOI] [PubMed] [Google Scholar]
- 21. Salih O, He S, Planamente S, Stach L, MacDonald JT, Manoli E, Scheres SHW, Filloux A, Freemont PS (2018) Atomic structure of type VI contractile sheath from Pseudomonas aeruginosa . Structure 26: 329–336.e323 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Clemens DL, Ge P, Lee BY, Horwitz MA, Zhou ZH (2015) Atomic structure of T6SS reveals interlaced array essential to function. Cell 160: 940–951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Costechareyre D, Bertolla F, Nesme X (2009) Homologous recombination in Agrobacterium: potential implications for the genomic species concept in bacteria. Mol Biol Evol 26: 167–176 [DOI] [PubMed] [Google Scholar]
- 24. Wu CF, Santos MNM, Cho ST, Chang HH, Tsai YM, Smith DA, Kuo CH, Chang J, Lai EM (2019) Plant pathogenic Agrobacterium tumefaciens strains have diverse type VI effector‐immunity pairs and vary in in planta competitiveness. Mol Plant Microbe Interact 32: 961–971 [DOI] [PubMed] [Google Scholar]
- 25. Liang X, Kamal F, Pei TT, Xu P, Mekalanos JJ, Dong TG (2019) An onboard checking mechanism ensures effector delivery of the type VI secretion system in Vibrio cholerae . Proc Natl Acad Sci USA 116: 23292–23298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Planamente S, Salih O, Manoli E, Albesa‐Jové D, Freemont PS, Filloux A (2016) TssA forms a gp6‐like ring attached to the type VI secretion sheath. EMBO J 35: 1613–1627 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Dix SR, Owen HJ, Sun R, Ahmad A, Shastri S, Spiewak HL, Mosby DJ, Harris MJ, Batters SL, Brooker TA et al (2018) Structural insights into the function of type VI secretion system TssA subunits. Nat Commun 9: 4765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Ouahrani‐Bettache S, Porte F, Teyssier J, Liautard J‐P, Köhler S (1999) pBBR1‐GFP: a broad‐host‐range vector for prokaryotic promoter studies. Biotechniques 26: 620–622 [DOI] [PubMed] [Google Scholar]
- 29. Quandt J, Hynes MF (1993) Versatile suicide vectors which allow direct selection for gene replacement in gram‐negative bacteria. Gene 127: 15–21 [DOI] [PubMed] [Google Scholar]
- 30. Ma L‐S, Lin J‐S, Lai E‐M (2009) An IcmF family protein, ImpLM, is an integral inner membrane protein interacting with ImpKL, and its walker a motif is required for type VI secretion system‐mediated Hcp secretion in Agrobacterium tumefaciens . J Bacteriol 191: 4316–4329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Wu C‐F, Lin J‐S, Shaw G‐C, Lai E‐M (2012) Acid‐induced type VI secretion system is regulated by ExoR‐ChvG/ChvI signaling cascade in Agrobacterium tumefaciens . PLoS Pathog 8: e1002938 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Schindelin J, Arganda‐Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B et al (2012) Fiji: an open‐source platform for biological‐image analysis. Nat Methods 9: 676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Lai EM, Chesnokova O, Banta LM, Kado CI (2000) Genetic and environmental factors affecting T‐pilin export and T‐pilus biogenesis in relation to flagellation of Agrobacterium tumefaciens . J Bacteriol 182: 3705–3716 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix
Expanded View Figures PDF
Source Data for Expanded View Figures
Review Process File
Source Data for Figure 1
Source Data for Figure 2
Source Data for Figure 3
Source Data for Figure 4
Source Data for Figure 5
Source Data for Figure 6