

Abstract
A new species of the Campylobacter genus is described, isolated from the preputial mucosa of bulls (Bos taurus). The five isolates obtained exhibit characteristics of Campylobacter, being Gram-negative non-motile straight rods, oxidase positive, catalase negative and microaerophilic. Phenotypic characteristics and nucleotide sequence analysis of 16S rRNA and hsp60 genes demonstrated that these isolates belong to a novel species within the genus Campylobacter. Based on hsp60 gene phylogenetic analysis, the most related species are C. ureolyticus, C. blaseri and C. corcagiensis. The whole genome sequence analysis of isolate FMV-PI01 revealed that the average nucleotide identity with other Campylobacter species was less than 75%, which is far below the cut-off for isolates of the same species. However, whole genome sequence analysis identified coding sequences highly homologous with other Campylobacter spp. These included several virulence factor coding genes related with host cell adhesion and invasion, transporters involved in resistance to antimicrobials, and a type IV secretion system (T4SS), containing virB2-virB11/virD4 genes, highly homologous to the C. fetus subsp. venerealis. The genomic G+C content of isolate FMV-PI01 was 28.3%, which is one of the lowest values reported for species of the genus Campylobacter. For this species the name Campylobacter portucalensis sp. nov. is proposed, with FMV-PI01 (= LMG 31504, = CCUG 73856) as the type strain.
Introduction
The Campylobacteraceae family of the order Campylobacterales is the largest and most diverse family in class Epsilonproteobacteria of the phylum Proteobacteria [1]. Campylobacter, the type genus of the family, contains species known to be pathogenic to humans [2] and other animals [3] as well as non-pathogenic species that colonize a large range of molluscs, reptiles, birds and mammals [4]. Presently, the Campylobacter genus comprises 31 species and 13 subspecies [5]. Cells of most Campylobacter species are motile, microaerophilic, Gram-negative, slender, spirally curved rods and 0.5–5 μm long by 0.2–0.8 μm wide. However, some species exhibit straight rod morphology [1] and C. gracilis, C. hominis, C. ureolyticus and C. blaseri are non-motile [6–9]. Seven species colonize cattle [4], of which C. coli, C. hyointestinalis, C. jejuni, C. lanienae and C. ureolyticus are found in faeces [10–12], C. sputorum is a commensal of the penile and preputial mucosae [13] and C. fetus includes two subspecies with clinical relevance in cattle. The C. fetus subspecies fetus colonizes the bovine intestinal tract, causing sporadic abortion, whereas the subspecies venerealis inhabits exclusively the genital tract of cattle, and is the etiologic agent of Bovine Genital Campylobacteriosis [14]. Herein, we describe a new species of Campylobacter isolated from a beef herd with history of reproductive failure compatible with Bovine Genital Campylobacteriosis, in the Alentejo province of Portugal.
Material and methods
Ethics statement
The in vivo samples used in this study were obtained from a bull that performed natural mating in a herd with clinical signs of Bovine Genital Campylobacteriosis. The samples were collected by a certified veterinarian, using the recommended OIE sampling methods for diagnostic purposes. No ethical approval was required as this was part of a routine veterinarian evaluation of beef herd reproductive failure. Ex vivo samples were obtained from animals slaughtered for human consumption. As sampling was performed post-mortem, in a certified slaughterhouse, no ethical approval from an Institutional Animal Care and Use Committee (IACUC) or other relevant ethics board was required.
Sampling, isolation procedures and culture conditions
Samples were obtained in vivo and post-mortem. The in vivo sample was collected from one mature Charolais bull, for laboratory diagnostic purposes. This sample was obtained from the preputial fornix using a technique combining scraping and small volume fluid washing (phosphate buffered saline; PBS) of the mucosa [15], and transported to the laboratory within 4 hours in two aliquots, one in Weybridge transport enrichment medium (TEM) and one in PBS. The bull had a normal routine breeding soundness evaluation and performed natural mating in a beef herd in the Alentejo province of Portugal. This herd showed fertility features compatible with Bovine Genital Campylobacteriosis, namely a low breeding season fertility, and late and spread calvings within the calving season. Post-mortem samples were also collected from the preputial fornix at two slaughterhouses and transported to the laboratory in PBS within 4 hours. The in vivo collected sample transported in Weybridge TEM was incubated at 37°C in a microaerobic atmosphere (Genbox Microaer, Biomérieux, France) for 48 hours, as an enrichment step. Enriched samples were plated through two different approaches: i) passively filtered onto blood agar (BA) and ii) spread in Campylobacter Skirrow Agar (CSA) [16]. In the BA approach, 0.65-μm mixed cellulose ester membrane filters (Advantec, Japan) were applied to the surface of BA plates supplemented with 5% sheep blood (Columbia agar + 5% sheep blood, Biomérieux) and inoculated with 100 μl of enriched sample for 30 minutes in aerobic conditions at room temperature; filters were then removed. In the CSA approach, 100 μl of enriched sample was spread on CSA plates. The BA and CSA plates were then incubated in microaerobic conditions at 37°C for 72 hours. The samples transported in PBS (in vivo and post-mortem collected) were diluted (ten-fold dilutions; 10−1, 10−2 and 10−3), streaked onto BA plates and incubated in a microaerobic atmosphere at 37°C for 48 hours. Colonies with Campylobacter-like morphology (small, smooth, translucent) were streaked onto BA and returned to a microaerobic atmosphere for a further incubation at 37°C for 48 hours. Before phenotypic and genotypic characterization, cells were microscopically examined using Gram staining.
Molecular identification and phylogenetic analysis
Genomic DNA from bacterial isolates was extracted using DNeasy Blood and Tissue kit (Qiagen, Germany), according to the manufacturer’s instructions. The 16S rRNA gene was amplified and sequenced using a universal set of primers–fD1 and rP1 (Table 1). Additionally, the flanking regions were amplified and sequenced with the primer sets FrAF/Vc1-2 and FrBF/FrBR (Table 1). Primers (FrAF, FrBF and FrBR) were designed using Primer-BLAST [17], based on the whole genome sequencing data. Primers FrAF and FrBR were designed to target a neighbour sequence of the 16S rRNA gene in order to obtain a full-length sequence. PCR reactions were carried out in a 50 μl mixture containing 0.3 μM of each primer, 200 μM of deoxynucleotide-triphosphates (4you4 dNTP Mix, Bioron, Germany), 1x reaction buffer (Complete NH4 reaction Buffer, 10x, Bioron), 2 units of DFS-Taq Polymerase (Bioron) and 3 μl of DNA. The thermal cycle conditions were as follows: 94°C for 2 min, followed by 30 cycles of denaturation (94°C for 30 s), annealing (30 s), and extension (72°C for 60 s), with a terminal extension step of 72°C for 5 min. The annealing temperature set for each primer pair is shown in Table 1. The amplified sequences were aligned and trimmed to create a full-length 16S rRNA gene sequence. The nucleotide sequence of FMV-PI01 isolate (GenBank accession no: MN417497) was compared with other 16S rRNA gene sequences deposited in the NCBI database, using BLASTN algorithm. To investigate the taxonomic position of the bacterial isolates, a phylogenetic tree based on 1513 nucleotide positions of 16S rRNA gene sequences was reconstructed. Available sequences of 16S rRNA gene of other Campylobacter species were retrieved from the GenBank database for phylogenetic analysis with Molecular Evolutionary Genetic Analysis (MEGA) X software [18]. Sequences were aligned with Clustal W algorithm [19] and positions with missing data were trimmed. The phylogenetic tree was reconstructed by the neighbour-joining method and stability of grouping was estimated by bootstrap analysis, set for 1000 replications. To further refine the phylogenetic analysis, a phylogenetic tree based on hsp60 gene (also known as cpn60 and groEL) sequences was also reconstructed, as described above. The amplification and sequencing of hsp60 gene were carried out using two primer pairs (hsp60_AF/hsp60_AR and hsp60_BF/hsp60_BR, Table 1) to obtain a 1472 bp long sequence.
Table 1. Primer sequences used for 16S rRNA and hsp60 genes amplification.
| Gene | Designation | Primers (5′-3′) | Annealing temperature | Amplicon size (bp) | Reference |
|---|---|---|---|---|---|
| 16S rRNA | fD1 | AGAGTTTGATCCTGGCTCAG | 52°C | 1475 | [20] |
| rP1 | ACGGTTACCTTGTTACGACTT | ||||
| FrAF | CGATTGAGCCAAGGGCTTTA | 52°C | 461 | This study | |
| Vc1-2R | ACTTAACCCAACATCTCACG | [21] | |||
| FrBF | ACACGTGCTACAATGGCATA | 53°C | 451 | This study | |
| FrBR | TCTCTGAAAACTAAACAAGGATGA | ||||
| hsp60 | hsp60_AF | AACTTTATGGTGGCGTTAAAA | 52°C | 1118 | |
| hsp60_AR | AGTTTCTGTTGCAGCACCTA | ||||
| hsp60_BF | AGCTTAATGTTGTTGAGGGA | 51°C | 1085 | ||
| hsp60_BR | TTACATCATACCACCCATAC |
Biochemical characterization and growth conditions
For biochemical characterization, bacterial cultures grown in a microaerobic atmosphere at 37°C for 48 hours were used. Oxidase activity was determined with oxidase test sticks (Liofilchem, Italy) and catalase activity was evaluated by observation of bubbling formation on a 3% peroxide hydrogen solution within 5 seconds. Urease and hydrogen sulfide (H2S) production were assessed on Christensen Urea Agar (Liofilchem) and Triple Sugar Iron (TSI) Agar (Liofilchem), respectively. Additionally, commercial tests were used to evaluate nitrate reduction, hippurate hydrolysis (Liofilchem) and indoxyl acetate hydrolysis (Indoxyl strips, Sigma-Aldrich), following the manufacturer’s instructions. The growth on BA supplemented with 1% glycine, 2% NaCl, 3.5% NaCl, 0.04% Tetrazolium chloride (TTC) and on MacConkey agar was determined according to standardized procedures, previously described [22,23]. The evaluation of growth on anaerobic and aerobic atmospheres at 37°C, and microaerobic growth at 25°C, 37°C or 42°C, after 48 to 96 hours was also performed. The bacterial motility was assessed by the hanging drop technique [24], using bacterial suspensions in PBS after 48 hours of growth on BA.
Reference strains C. fetus subsp. fetus NCTC 10842, C. fetus subsp. venerealis NCTC 10354, C. coli CNET 068, C. jejuni subsp. jejuni NCTC 11168, and isolates identified as C. sputorum bv. sputorum, and Proteus sp. were used as controls in the tests described above.
Electron microscopy
Electron micrographs were taken from a pure culture of isolate FMV-PI01. Preparations for electron microscopy were performed as previously described [25–27], followed by post-fixation in 2% osmium tetroxide and stepwise dehydration (30/50/70/90/100%) in ethanol. For transmission electron microscopy, samples were embedded in EPON812. Ultrathin sections (80 nm) were placed onto 300-mesh formvar-carbon-coated nickel grids (Plano, Wetzlar, Germany) and stained with Uranyless and lead citrate [27]. Specimens were viewed on a CM12 transmission electron microscope operating at 60 kV.
For scanning electron microscopy, fixed and dehydrated samples were resuspended in two changes of hexamethyl-disilazane (Sigma), sputter coated with gold, and inspected on a JEOL 840 scanning electron microscope operating at 25 kV.
Whole genome sequencing and comparative genomic analysis
The genomic DNA was extracted from a pure culture of the isolate FMV-PI01, grown on BA over 48 hours, using the DNeasy Blood and Tissue kit (Qiagen, Germany). After the genomic library preparation, the generated DNA fragments were sequenced using the HiSeq 4000 System (Illumina), with 150 bp paired-end reading sequences and assembled using CLC Genomics Workbench version 11.0.1 (CLC bio, Denmark), at Stabvida (Caparica, Portugal). The assembled genome was annotated with the Rapid Annotation Using Subsystem Technology (RAST) 2.0 pipeline [28,29].
The average nucleotide identity (ANI) was calculated with the webserver JspeciesWS [30] for the isolate FMV-PI01 and other Campylobacter species. The G+C content was determined based on the whole genome sequence of isolate FMV-PI01.
The identification of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) / CRISPR-associated (Cas) systems was performed using the CRISPRCasFinder webserver [31]. Additionally, the presence of putative virulence factor coding genes was evaluated based on the homology of translated sequences using BLASTP. Only sequences with query coverage >95% and identity >50% with known protein sequences of virulence factors were considered.
To estimate the pathogenic potential of this novel species, the assembled genome of isolate FMV-PI01 was analysed with the PathogenFinder Web server [32], using the automatic model option.
Results and discussion
Morphological characterization
The aliquot of the in vivo collected preputial sample transported in Weybridge TEM produced no Campylobacter-like morphology colonies on either approach (BA and CSA). In contrast, the in vivo collected preputial sample aliquot transported in PBS and streaked onto BA produced Campylobacter-like morphology colonies (isolate FMV-PI01). Post-mortem preputial samples, collected in PBS, produced four isolates (isolates FMV-PI02 to FMV-PI05) from beef bulls arising from four geographically distinct herds.
Colony morphology in BA was common to all isolates, colonies being punctiform, circular, with convex elevation, smooth margins, without pigmentation and no haemolysis observable. Bacterial cells from all isolates were Gram-negative and exhibited a straight rod shape. Electron microscopy revealed that isolate FMV-PI01 bacterial cells had an average length of 1.68±0.07 μm (n = 30) and an average width of 0.44±0.01 μm (n = 30), and were devoid of a flagellum. Occasional filamentous cells were observed displaying lengths up to 18 μm (Fig 1).
Fig 1. Electron micrographs of isolate FMV-PI01.

(A) Transmission electron microscope micrograph in longitudinal and transverse views. Scale bar, 500 nm. (B) Scanning electron microscope micrograph in detail. Scale bar, 2 μm (C) Scanning electron microscope micrograph with evident long filamentous cells (white arrow). Scale bar, 2 μm.
Phenotypic characterization
The phenotypic tests were validated by control Campylobacter species strains, whose results were all in agreement with those reported in the literature [33]. All five isolates presented identical phenotypic characteristics (Table 2). Isolates were positive for oxidase activity, a feature present in all Campylobacter species except C. gracilis and sporadic isolates of C. concisus and C. showae [1]. Isolates were negative for catalase activity and were unable to hydrolyse urea, hippurate and indoxyl acetate. From the seven Campylobacter species that colonize cattle, only C. jejuni is able to hydrolase hippurate [4,8]. Contrary to the majority of Campylobacter species, the isolates were unable to reduce nitrate. In fact, only three other species of the Campylobacter genus fail to reduce nitrate: C. hominis, C. mucosalis and C. concisus [1]. Production of H2S was not observed in TSI agar. The growth tests showed that the isolates were microaerophilic, although they could tolerate anaerobic conditions with weaker growth. The microbial growth in microaerobic conditions was similar to the majority of the Campylobacter species [1], observable at 37°C and 42°C, but not at 25°C. Further testing showed that four of the five isolates grew in the presence of 1% glycine, but none grew on BA supplemented with 2% or 3.5% NaCl. Growth was not observed either on MacConkey agar or on Muller Hinton Agar supplemented with 5% sheep blood. Unlike other Campylobacter species (e.g. C. hominis and C. ureolyticus) [34], the isolates did not require hydrogen (H2) to grow since the employed gas-generating sachet Genbox does not release hydrogen. No motility was observed on the hanging drop method preparation, which can justify the absence of colonies with Campylobacter-like morphology in the BA approach since that technique was developed to isolate motile Campylobacter spp. that can cross the 0.65-μm mixed cellulose ester membrane filters. To summarize, the five isolates were Gram-negative rods, microaerophilic and oxidase positive, which are phenotypic traits common to the genus Campylobacter [13]. However, the isolates were distinguishable from the most related species C. sputorum, C. corcagiensis, C. blaseri, C. ureolyticus and C. geochelonis (see genomic characterization), since unlike these species, the isolates could not reduce nitrate (Table 2).
Table 2. Phenotypic characteristics differentiating FMV-PI isolates from the other Campylobacter species.
Taxa: 1—FMV-PI isolates (n = 5); 2—C. hominis; 3—C. gracilis; 4—C. sputorum; 5—C. ureolyticus; 6—C. corcagiensis; 7—C. mucosalis; 8—C. concisus; 9—C. pinnipediorum subsp. pinnipediorum; 10—C. curvus; 11—C. rectus; 12—C. showae; 13—C. coli; 14—C. hyointestinalis subsp. hyointestinalis; 15—C. jejuni subsp. jejuni; 16—C. lanienae; 17- C. fetus subsp. venerealis;; 18—C. hepaticus; 19—C. avium; 20—C. canadensis; 21—C. cuniculorum; 22—C. geochelonis; 23—C. helveticus; 24—C. insulaenigrae; 25—C. lari subsp. lari; 26—C. peloridis; 27—C. subantarcticus; 28—C. upsaliensis; 29—C. volucris; 30—C. blaseri; 31—C. iguaniorum; 32—C. ornithocola. Data for reference taxa were obtained from previous species descriptions [7–9,33,35–47]. + 90–100%; (+) 75–89%; v 26–74%; (-) 11–25%; - 0–10%; nd–not determined; w—weakly positive.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Motility | - | - | - | + | - | + | + | + | nd | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | nd | nd | + | nd | - | nd | nd |
| Oxidase | + | + | - | + | + | + | + | v | + | + | + | v | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Catalase | - | - | (-) | v* | v | + | - | - | + | - | (-) | + | + | + | + | + | (+) | + | w | v | + | + | - | + | + | + | + | - | + | + | + | - |
| α-haemolysis | - | - | - | + | v | - | - | (-) | + | (-) | + | + | (-) | v | + | + | V | - | - | - | + | - | + | nd | + | nd | + | + | nd | - | + | - |
| Urease | - | - | - | v* | + | + | - | - | + | - | - | - | - | - | - | - | - | - | - | v | - | - | - | - | - | nd | nd | - | nd | + | - | + |
| Hippurate hydrolysis | - | - | - | - | - | - | - | - | - | (-) | - | - | - | - | + | - | - | (+) | + | - | - | + | - | - | - | - | - | - | - | - | - | - |
| Indoxyl acetate hydrolysis | - | - | (+) | - | v | v | - | - | - | v | + | v | + | - | + | - | - | + | + | - | + | - | + | - | - | nd | - | + | - | + | - | - |
| Nitrate reduction | - | - | (+) | (+) | + | (+) | (-) | (-) | + | + | + | + | + | + | + | + | (+) | v | + | v | + | + | + | + | + | nd | nd | + | + | + | + | + |
| H2S production | - | - | - | + | - | + | + | - | + | (-) | - | v | - | + | - | - | - | - | - | v | - | - | - | + | - | nd | - | - | - | + | + | + |
| Growth in/at/on: | ||||||||||||||||||||||||||||||||
| 1% glycine | v | + | + | + | + | + | v | (-) | v | + | + | v | (+) | + | + | - | (-) | + | - | v | - | + | v | + | + | + | (+) | + | - | + | + | - |
| 2% NaCl | - | nd | + | + | + | + | - | (-) | nd | v | v | + | - | - | - | nd | - | - | - | nd | - | + | - | - | + | (+) | + | - | - | nd | nd | nd |
| 3.5% NaCl | - | nd | - | v | + | nd | - | - | nd | - | - | - | - | - | - | nd | - | - | nd | - | nd | nd | - | - | - | nd | - | - | nd | nd | nd | nd |
| MacConkey Agar | - | - | (+) | v | v | - | (+) | - | nd | (+) | - | + | v | v | - | + | V | - | - | + | - | - | - | nd | - | nd | (-) | - | w | + | nd | nd |
| TTC 0.04% | - | - | - | - | - | - | - | - | nd | V | - | - | + | - | + | nd | - | + | - | nd | v | - | - | nd | + | nd | nd | v | - | nd | nd | nd |
| 25°C, microaerobic | - | - | - | - | - | nd | - | - | + | - | - | - | - | (-) | - | - | + | - | - | - | - | + | - | - | - | - | - | - | - | + | + | + |
| 37°C, microaerobic | + | + | - | + | + | + | + | + | + | v | - | v | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 42°C, microaerobic | + | (-) | v | + | v | + | + | (+) | - | v | (-) | v | + | + | + | + | - | + | + | + | (+) | - | + | - | + | + | + | + | + | + | - | - |
| 37°C, anaerobic | w | + | + | + | + | + | + | + | + | + | + | + | - | - | - | + | V | - | - | + | - | + | - | - | - | nd | + | - | + | + | + | + |
| 37°C, aerobic | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| H2 requirement | - | + | + | - | + | - | + | + | - | + | + | + | - | v | - | - | - | - | v | - | - | - | - | nd | - | nd | - | - | nd | - | - | - |
*test results differ between biovars.
Phylogenetic analysis
The 16S rRNA gene sequence alignment revealed that the five bacterial isolates shared 99.87% sequence similarity. This homology, associated to their similar phenotypic characteristics, confirms that they belong to the same species. The comparative analysis of the 16S rRNA gene sequence of isolate FMV-PI01, using the BLASTN algorithm, confirmed that this isolate is closely related to the genus Campylobacter. The highest identities were obtained with C. concisus and C. gracilis (100% coverage and 94.7% identity) and C. hominis (100% coverage and 94.1% identity). These 16S rRNA sequence identities are below the threshold of 97%, defined for bacteria belonging to the same species [48], which supports the identification of a novel species within the Campylobacter genus.
The phylogenetic analysis using 16S rRNA and hsp60 genes (Figs 2 and 3) demonstrated that these isolates form a robust cluster. However, the phylogenetic position of the five isolates was not clearly established based on the phylogenetic analysis. The low bootstrap values observed (< 70%) indicate that the 16S rRNA and hsp60 genes have a weak discriminatory power relatively to the group of phylogenetically related species.
Fig 2. Phylogenetic tree based on 16S rRNA gene sequences of Campylobacter species, reconstructed by the neighbour-joining method.
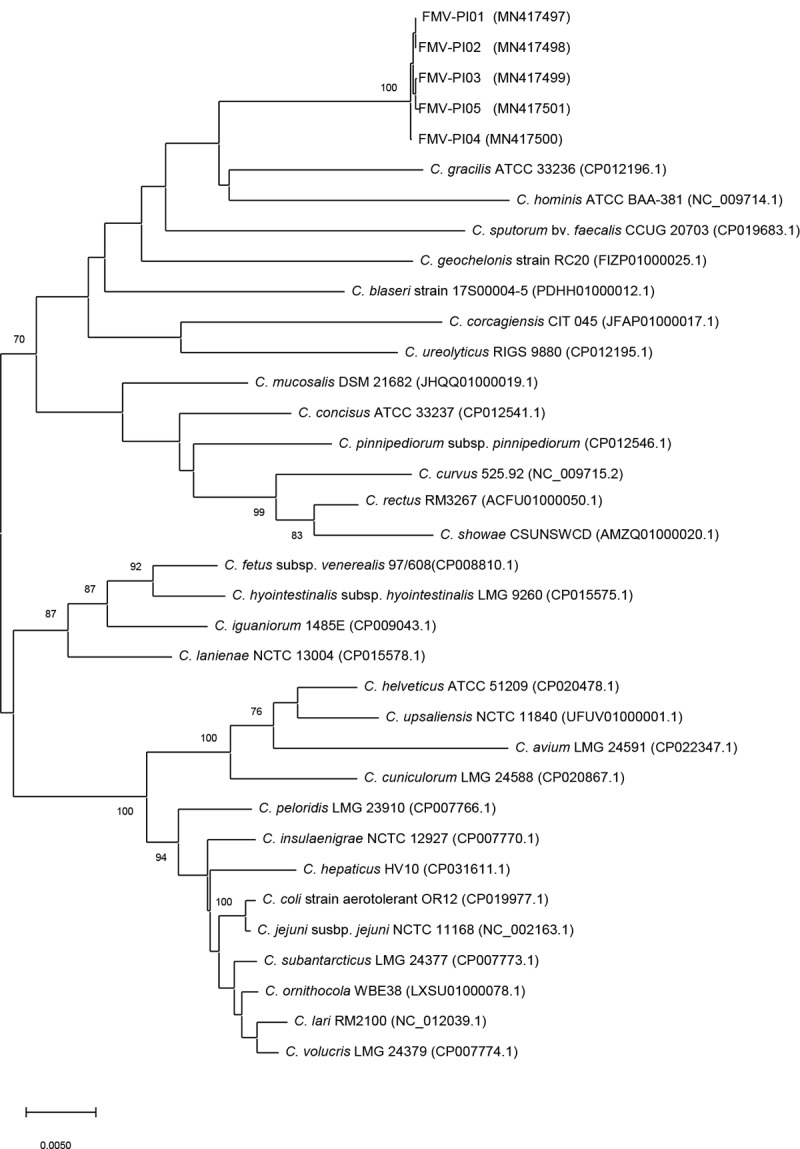
Bootstrap values (%) obtained from 1000 simulations are indicated at the nodes. Bootstrap values lower than 70% are not shown. Bar: 0.0050 substitutions per site.
Fig 3. Phylogenetic tree based on hsp60 gene sequences of Campylobacter species, reconstructed by the neighbour-joining method.
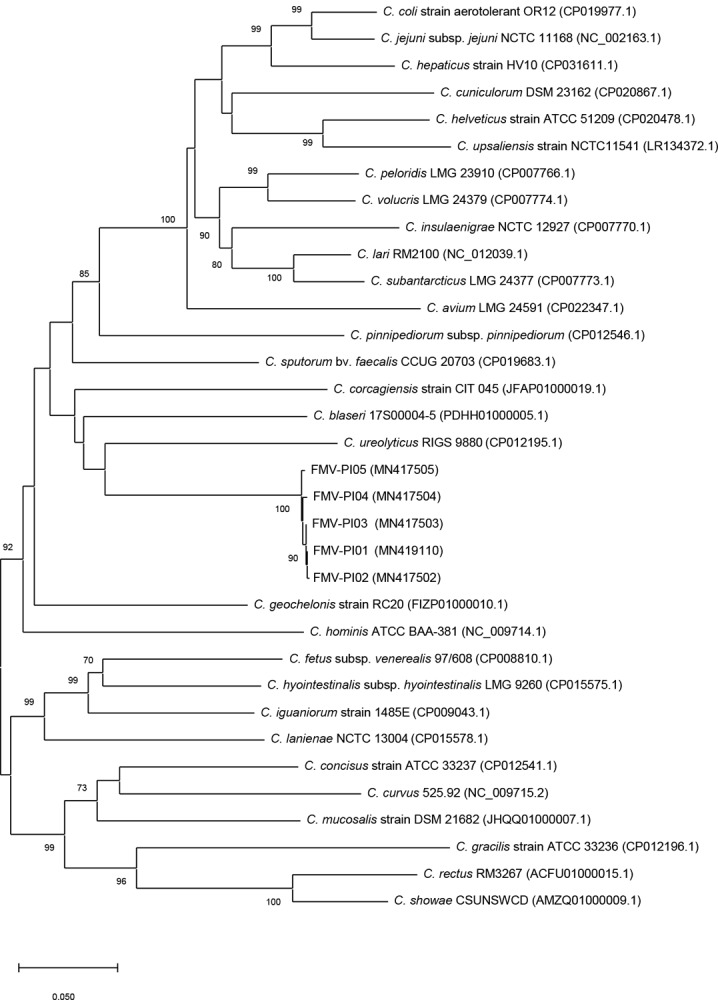
Bootstrap values (%) obtained from 1000 simulations are indicated at the nodes. Bootstrap values lower than 70% are not shown. Bar: 0.050 substitutions per site.
The 16S rRNA phylogenetic analysis identified C. gracilis, C. hominis, C. sputorum, C. geochelonis, C. blaseri, C. corcagiensis and C. ureolyticus as the most related taxa. This was also observed in the phylogenetic analysis based on hsp60 gene, except for C. gracilis, which was not grouped with FMV-PI isolates. The hsp60 gene analysis provided a better phylogenetic resolution, with a higher number of branches with high bootstrap values. Although the 16S rRNA gene is the most commonly accepted for use in taxonomic studies, phylogenetic analysis based on hsp60 gene is more discriminative for some bacterial taxa as the family Campylobacteraceae [33,49,50]. A greater interspecies variation in the nucleotide sequence of hsp60 gene than in 16S rRNA gene may explain these findings [50].
Genomic characterization
The Whole Genome Shotgun project was deposited at DDBJ/ENA/GenBank under the accession VWSJ00000000. The version described in this paper is version VWSJ01000000. The genome of isolate FMV-PI01 is 1 767 933 bp long, composed of 98 contigs, with 28.3% G+C content. This G+C content is one of the lowest reported for species of the genus Campylobacter. The G+C content of a bacterial species may reflect adaptation to environmental niches and lifestyles, since non-free-living bacteria generally have shorter genomes with lower G+C content, promoting energy conservation in environments with scarcity of nutrients [51,52]. Interestingly, C. sputorum, also with one of the lowest G+C contents within the genus Campylobacter (29%) [53], colonizes the bull’s preputial mucosa, exhibiting niche preferences similar to herein described isolates.
The ANI analysis revealed that homology with other Campylobacter species was less than 75%, which is far below the 95–96% cut-off for isolates of the same species [54]. These nucleotide similarities are in accordance with the hsp60 and 16S rRNA genes phylogenetic analysis results, showing that C. ureolyticus, C. corcagiensis and C. blaseri are the most closely related taxa with ANI values of 74.3%, 73.3% and 73.0%, respectively, followed by C. geochelonis (72.3%), C. sputorum (72.2%), C. hominis (70.1%) and C. gracilis (65.9%). The ANI values of these two latter species (C. gracilis and C. hominis) support the results of the analysis based on hsp60 gene rather than the 16S rRNA gene. Overall, these findings support that the herein described isolates belong to the Campylobacter genus, representing a novel species, for which the designation Campylobacter portucalensis sp. nov. is proposed. The ANI analysis homology between Campylobacter portucalensis sp. nov. and its most related Campylobacter species is shown in Table 3.
Table 3. Average nucleotide identity (ANI) values (%) based on BLAST for C. portucalensis sp. nov. and the most related Campylobacter species.
Strains: 1 –C. portucalensis sp. nov. FMV-PI01; 2 –C. hominis ATCC BAA-381; 3 –C. fetus subsp. fetus 82–40; 4 –C. lari RM2100; 5 –C. insulaenigrae NCTC 12927; 6 –C. hyointestinalis subsp. hyointestinalis LMG 9260; 7 –C. hepaticus HV10; 8 –C. jejuni subsp. jejuni LMG 9872; 9 –C. corcagiensis CIT 045; 10 –C. blaseri 17500004–5; 11 –C. ureolyticus DSM 20703; 12 –C. sputorum bv. faecalis CCUG 20703; 13 –C. gracilis ATCC 33236; 14 –C. concisus ATCC 33237; 15 –C. geochelonis RC20.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | * | 70.1 | 69.8 | 69.7 | 69.6 | 69.6 | 69.3 | 69.3 | 73.3 | 73.0 | 74.7 | 72.2 | 65.9 | 68.4 | 72.3 |
| 2 | 70.2 | * | 69.2 | 68.3 | 68.2 | 69.1 | 68.2 | 68.3 | 69.8 | 69.7 | 71.6 | 70.2 | 68.2 | 68.3 | 70.2 |
| 3 | 69.4 | 68.5 | * | 68.2 | 67.9 | 78.5 | 67.6 | 67.8 | 68.8 | 69.4 | 69.3 | 69.5 | 66.6 | 68.2 | 69.7 |
| 4 | 70.0 | 68.5 | 68.9 | * | 81.9 | 68.8 | 74.3 | 74.5 | 69.0 | 69.8 | 69.8 | 69.8 | 65.5 | 68.0 | 69.0 |
| 5 | 69.7 | 68.3 | 68.4 | 81.9 | * | 68.4 | 74.1 | 74.0 | 68.6 | 69.5 | 69.6 | 69.3 | 65.4 | 67.5 | 68.5 |
| 6 | 69.1 | 68.4 | 78.6 | 67.9 | 67.7 | * | 67.4 | 67.8 | 68.7 | 69.0 | 69.2 | 69.5 | 66.7 | 68.6 | 69.7 |
| 7 | 69.4 | 68.2 | 68.5 | 74.5 | 74.5 | 68.2 | * | 84.4 | 68.6 | 69.1 | 69.3 | 69.2 | 65.1 | 67.0 | 68.1 |
| 8 | 69.1 | 68.1 | 68.1 | 74.4 | 73.9 | 68.3 | 84.4 | * | 68.5 | 68.8 | 69.1 | 68.9 | 65.6 | 67.6 | 68.4 |
| 9 | 72.3 | 68.9 | 68.6 | 67.8 | 67.8 | 68.5 | 67.5 | 67.7 | * | 71.2 | 73.0 | 70.3 | 65.4 | 67.7 | 71.1 |
| 10 | 72.5 | 69.3 | 69.2 | 68.9 | 68.6 | 69.2 | 68.3 | 68.3 | 71.8 | * | 72.8 | 71.2 | 65.4 | 67.9 | 72.6 |
| 11 | 74.8 | 71.6 | 69.8 | 69.7 | 69.4 | 69.8 | 69.1 | 69.2 | 73.9 | 73.1 | * | 72.0 | 65.8 | 68.6 | 72.3 |
| 12 | 71.9 | 70.2 | 69.8 | 69.4 | 69.1 | 69.8 | 68.8 | 68.8 | 71.2 | 71.5 | 71.8 | * | 66.2 | 68.8 | 70.9 |
| 13 | 65.5 | 67.2 | 66.2 | 64.6 | 64.4 | 66.4 | 64.4 | 64.8 | 65.2 | 64.7 | 64.8 | 65.4 | * | 67.7 | 67.0 |
| 14 | 67.7 | 67.4 | 68.3 | 67.0 | 66.6 | 68.6 | 66.5 | 66.9 | 67.7 | 67.8 | 67.81 | 68.2 | 68.0 | * | 69.0 |
| 15 | 71.4 | 69.2 | 69.3 | 67.9 | 67.2 | 69.3 | 67.1 | 67.5 | 71.0 | 72.1 | 71.33 | 70.2 | 67.3 | 68.8 | * |
A total of 1877 coding sequences (CDS) and 41 RNAs (36 transfer RNA and 5 ribosomal RNA genes) were identified in FMV-PI01’s genome. Of the identified CDS, 497 were assigned to 191 subsystems. Subsystems with higher number of genes are related to metabolic processes, and include “protein metabolism” (n = 126), “amino acids and derivatives” (n = 136) and “cofactors, vitamins, prosthetic groups and pigments” (n = 65) (Fig 4). Nineteen genes involved in “Virulence, disease and defence” and 16 genes related to “stress response” were identified (Fig 4).
Fig 4. Subsystem category distribution in the genome of isolate FMV-PI01, based on the RAST server.

The CRISPRCasFinder identified one CRISPR-Cas system in FMV-PI01’s genome, which contains 15 CRISPR repeats with 13 CRISPR spacers and a type III-D Cas system. Despite CRISPR-Cas systems have been associated with phage defence mechanisms, these systems may also play a role in bacterial virulence and host immune evasion [55]. The diversity of CRISPR-Cas systems within the Campylobacter genus is wide, being the Cas systems type I and II the most common among Campylobacter species [55]. However, the type III-D, which was found in the genome of C. portucalensis sp. nov., was also identified in C. fetus. The mechanisms behind the selection of each system in different bacterial species [56], as well as the role of Cas system type III-D in C. fetus, remain unclear.
A more detailed analysis using BLASTP algorithm allowed the identification of genes potentially involved in adhesion and invasion to host cells. These genes encode homologous of the fibronectin/fibrinogen binding protein (98% query cover, 62% identity), collagenase like peptidase of U32 family (100% query cover, 81% identity) and Campylobacter invasion antigen B, ciaB (100% query cover, 64% identity). These virulence factor coding genes are present in several Campylobacter species, namely in C. fetus and C. jejuni [57]. Adhesion of C. jejuni to host cells is mediated by the fibronectin binding protein [58–60], and the mutational inactivation of the ciaB gene reduced the invasion in INT 407 cells, revealing that ciaB is involved in the internalization of C. jejuni [61,62]. Therefore, the presence of these genes in the genome of C. portucalensis is potentially related to host cell adhesion and invasion in the bovine reproductive tract. In addition, genes encoding multidrug efflux pumps of the resistance-nodulation-cell division family were found, namely a CmeABC efflux pump. These transporters are present in several species of Campylobacter (e.g. C. jejuni, C. coli, C. fetus and C. lari) and contribute to multidrug resistance [63]. For instance, this efflux pump in C. jejuni is involved in resistance to bile salts [64], macrolides, tetracycline [65], ciprofloxacin and other antimicrobials [66].
The C. portucalensis FMV-PI01’s genome encodes a type IV secretion system (T4SS), containing virB2-virB11/virD4 genes, highly homologous to the C. fetus subsp. venerealis T4SS (99% coverage and 97.2% identity). This T4SS was confirmed to contribute to C. fetus subsp. venerealis virulence properties, namely invasive and cytotoxic potential [67,68]. The fact that C. portucalensis sp. nov. and C. fetus subsp. venerealis, the causative agent of Bovine Genital Campylobacteriosis, share virulence factor coding genes and are both inhabitants of the bull preputial mucosa, may suggest that this novel Campylobacter species has the potential to cause disease in cattle. The herd from which the isolate FMV-PI01 was obtained presented signs of reproductive failure compatible with Bovine Genital Campylobacteriosis, the reason for the disease investigation. As in the above disease, where the bull acts as an asymptomatic carrier and signs of disease are only reflected on the female (embryonic and fetal mortalities) and herd (fertility rate, calving pattern and calving interval) sides [14], the bull from which the samples were taken was clinically sound. However, since several interacting factors may contribute to beef cattle herd’s infertility, one cannot conclude that the observed reproductive failure was the result of the infection with C. portucalensis sp. nov. To further investigate the pathogenic potential of C. portucalensis sp. nov in cattle fertility, research in the female reproductive tract needs to be addressed.
The evaluation of the pathogenic potential based on the PathogenFinder analysis showed that the probability of isolate FMV-PI01 being a human pathogen was 82.6%, indicating that this isolate may have the potential to cause disease in humans.
Conclusion
The distinct phenotypic and genotypic characteristics of the bacterial isolates confirm the identification of a novel species within the Campylobacter genus, for which the name Campylobacter portucalensis sp. nov. is proposed. C. portucalensis sp. nov. is an inhabitant of bulls’ preputial mucosa with unknown pathogenic potential.
Description of Campylobacter portucalensis sp. nov
Campylobacter portucalensis sp. nov. (por.tu.cal.en’sis. N.L. masc. adj. portucalensis referring to Portugal, from where the type strain was originally isolated).
In Columbia agar (supplemented with 5% sheep blood), after 48 hours in microaerobic atmosphere at 37°C, colonies are punctiform (1 mm in diameter), convex, circular with smooth margins and without any pigments. Colonies are non-haemolytic. Cells are Gram-negative, straight rods (length 1.68±0.07 μm and width 0.44±0.01 μm). A flagellum is absent and cells are non-motile. Occasional longer filamentous cells are observable.
Growth is observed on blood agar at 37°C under microaerobic and anaerobic (weak growth), but not aerobic conditions. Does not require H2 supplementation to grow. Isolates grow in microaerobic conditions at 37°C and 42°C but not at 25°C. Strains may differ in their ability to grow on blood agar medium supplemented with 1% glycine. Growth is not observed on blood agar medium supplemented with 2% and 3.5% NaCl. Unable to grow on Macconkey agar or Mueller-Hinton agar.
Phenotypically, C. portucalensis sp. nov. is oxidase positive and catalase negative. Negative for urease activity. Unable to hydrolyse hippurate and indoxyl acetate. Does not reduce nitrate. Does not produce hydrogen sulfide in TSI medium.
The genomic G+C content of the type strain is 28.3%.
The type strain FMV-PI01 (= LMG 31504, = CCUG 73856) was isolated from the reproductive tract of a bull (Bos taurus) sampled in the Alentejo province of Portugal in 2018.
Data Availability
All relevant data are within the manuscript.
Funding Statement
This study was supported by Fundação para a Ciência e a Tecnologia (FCT - www.fct.pt), under the 02/SAICT/2017, project no. 030145, co-funded by Fundo Europeu de Desenvolvimento Regional (FEDER). In addition, the study was funded by Centro de Investigação Interdisciplinar em Sanidade Animal (http://ciisa.fmv.ulisboa.pt/en/) (Project UID/CVT/00276/2019, funded by FCT). Marta Silva and Gonçalo Pereira are PhD students supported by grants from FCT, SFRH/BD/125657/2016 and SFRH/BD/130923/2017, respectively. Andrew Hemphill is funded by the Swiss National Science Foundation grant No. 310030_184662. Elisabete Silva is funded by FCT (DL 57/2016/CP1438/CT0001). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Vandamme P, Dewhirst FE, Paster BJ, On SLW. Genus I Campylobacter In: Garrity GM, Brenner DJ, Krieg NR, Staley JT, editors. Bergey’s Manual of Systematic Bacteriology. New York: Springer-Verlag: 2005. p. 1147–1160. [Google Scholar]
- 2.Gölz G, Rosner B, Hofreuter D, Josenhans C, Kreienbrock L, Löwenstein A, et al. Relevance of Campylobacter to public health—the need for a one health approach. Int J Med Microbiol. 2014;304(7):817–823. 10.1016/j.ijmm.2014.08.015 . [DOI] [PubMed] [Google Scholar]
- 3.Sahin O, Yaeger M, Wu Z, Zhang Q. Campylobacter-associated diseases in animals. Annu Rev Anim Biosci. 2017;8(5):21–42. 10.1146/annurev-animal-022516-022826 . [DOI] [PubMed] [Google Scholar]
- 4.Lastovica AJ, On SLW, Zhang L. The Family Campylobacteraceae In: Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F, editors. The Prokaryotes. Heidelberg: Springer; 2014. p. 307–335. [Google Scholar]
- 5.bacterio.net [Internet]. List of prokariotic names with standing in nomenclature (LPSN) [cited 2019 Jun 5]. Available from: http://www.bacterio.net/campylobacter.html
- 6.Shinha T. Fatal bacteremia caused by Campylobacter gracilis, United States. Emerg Infect Dis. 2015;21(6):1084–1085. 10.3201/eid2106.142043 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lawson AJ, On SLW, Logan JM, Stanley J. Campylobacter hominis sp. nov., from the human gastrointestinal tract. Int J Syst Evol Microbiol. 2001;51:651–660. 10.1099/00207713-51-2-651 . [DOI] [PubMed] [Google Scholar]
- 8.Vandamme P, Debruyne L, De Brandt E, Falsen E. Reclassification of Bacteroides ureolyticus as Campylobacter ureolyticus comb. nov., and emended description of the genus Campylobacter. Int J Syst Evol Microbiol. 2010;60: 2016–2022. 10.1099/ijs.0.017152-0 . [DOI] [PubMed] [Google Scholar]
- 9.Gilbert MJ, Zomer AL, Timmerman AJ, Spaninks MP, Rubio-García A, Rossen JW, et al. Campylobacter blaseri sp. nov., isolated from common seals (Phoca vitulina). Int J Syst Evol Microbiol. 2018;68(5):1787–1794. 10.1099/ijsem.0.002742 . [DOI] [PubMed] [Google Scholar]
- 10.Hakkinen M, Heiska H, Hänninen ML. Prevalence of Campylobacter spp. in cattle in Finland and antimicrobial susceptibilities of bovine Campylobacter jejuni strains. Appl Environ Microbiol. 2007;73(10):3232–3238. 10.1128/AEM.02579-06 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Inglis GD, Kalischuk LD, Busz HW, Kastelic JP. Colonization of cattle intestines by Campylobacter jejuni and Campylobacter lanienae. Appl Environ Microbiol. 2005;71(9):5145–5153. 10.1128/AEM.71.9.5145-5153.2005 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Koziel M, Lucey B, Bullman S, Corcoran GD, Sleator RD. Molecular-based detection of the gastrointestinal pathogen Campylobacter ureolyticus in unpasteurized milk samples from two cattle farms in Ireland. Gut Pathog. 2012;4:14 10.1186/1757-4749-4-14 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Quinn PJ, Markey BK, Leonard FC, FitzPatrick ES, Fanning S, Hartigan PJ editors. Veterinary Microbiology and Microbial Disease. 2nd ed. Chichester: Wiley-Blackwell; 2011. [Google Scholar]
- 14.Michi NA, Favetto PH, Kastelic J, Cobo ER. A review of sexually transmitted bovine trichomoniasis and campylobacteriosis affecting cattle reproductive health. Theriogenology. 2016;85(5):781–791. 10.1016/j.theriogenology.2015.10.037 . [DOI] [PubMed] [Google Scholar]
- 15.Monke HJ, Love BC, Wittum TE, Monke DR, Byrum BA. Effect of transport enrichment medium, transport time, and growth medium on the detection of Campylobacter fetus subsp. venerealis. J Vet Diagn Invest. 2002:14(1):35–39. 10.1177/104063870201400107 . [DOI] [PubMed] [Google Scholar]
- 16.Chaban B, Guerra AG, Hendrick SH, Waldner CL, Hill JE. Isolation rates of Campylobacter fetus subsp. venerealis from bovine preputial samples via passive filtration on nonselective medium versus selective medium, with and without transport medium. Am J Vet Res. 2013;74(8):1066–1069. 10.2460/ajvr.74.8.1066 . [DOI] [PubMed] [Google Scholar]
- 17.Ye J, Coulouris G, Zaretskaya I, Cutcutache I, Rozen S, Madden TL. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinformatics. 2012;13:134 10.1186/1471-2105-13-134 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35:1547–1549. 10.1093/molbev/msy096 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–4680. 10.1093/nar/22.22.4673 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Weisburg WG, Barns SM, Pelletier DA, Lane DJ. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol. 1991; 173 (2):697–703. 10.1128/jb.173.2.697-703.1991 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gorkiewicz G, Feierl G, Schober C, Dieber F, Köfer J, Zechner R, et al. Species-specific identification of campylobacters by partial 16S rRNA gene sequencing. J Clin Microbiol. 2003; 41(6):2537–2546. 10.1128/JCM.41.6.2537-2546.2003 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.On SLW, Holmes B. Effect of inoculum size on the phenotypic characterization of Campylobacter species. J Clin Microbiol. 1991;29(5):923–926 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.On SLW, Holmes B. Reproducibility of tolerance tests that are useful in the identification of campylobacteria. J Clin Microbiol. 1991;29:1785– 1788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Public Health England [Internet]. UK standards for microbiology investigations: Motility test. [cited 2019 Nov 18]. Available from: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/762926/TP_21i4.pdf
- 25.Hemphill A, Croft SL. Electron microscopy in parasitology In: Rogan MM, editor. Analytical parasitology. Heidelberg: Springer-Verlag: 1997. p. 227–268. [Google Scholar]
- 26.Hemphill A, Vonlaufen N, Naguleswaran A, Keller N, Riesen M, Guetg N, et al. Tissue culture and explant approaches to studying and visualizing Neospora caninum and its interactions with the host cell. Microsc Microanal. 2004;10(5):602–620. 10.1017/S1431927604040930 . [DOI] [PubMed] [Google Scholar]
- 27.Basto AP, Müller J, Rubbiani R, Stibal D, Giannini F, Süss-Fink G, et al. Characterization of the activities of dinuclear thiolato-bridged arene ruthenium complexes against Toxoplasma gondii. Antimicrob Agents Chemother. 2017;24;61(9). 10.1128/AAC.01031-17 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75 10.1186/1471-2164-9-75 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Overbeek R, Olson R, Pusch GD, Olsen GJ, Davis JJ, Disz T, et al. The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acid Res. 2014;42:206–214. 10.1093/nar/gkt1226 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Richter M, Rosselló-Móra R, Glöckner FO, Peplies J. JSpeciesWS: a web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics. 2016; 32(6):929–931. 10.1093/bioinformatics/btv681 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Couvin D, Bernheim A, Toffano-Nioche C, Touchon M, Michalik J, Néron B, et al. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res, 46:246–251. 10.1093/nar/gky425 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cosentino S, Voldby Larsen M, Møller Aarestrup F, Lund O. PathogenFinder—Distinguishing friend from foe using bacterial whole genome sequence data. PLoS One. 2013;8(10):e77302 10.1371/journal.pone.0077302 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.On SLW, Miller WG, Houf K, Fox JG, Vandamme P. Minimal standards for describing new species belonging to the families Campylobacteraceae and Helicobacteraceae: Campylobacter, Arcobacter, Helicobacter and Wolinella spp. Int J Syst Evol Microbiol. 2017;67:5296–5311. 10.1099/ijsem.0.002255 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kaakoush NO, Castaño-Rodríguez N, Mitchell HM, Man SM. Global epidemiology of Campylobacter infection. Clin Microbiol Reviews. 2015;28(3):687–720. 10.1128/CMR.00006-15 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Koziel M, O’Doherty P, Vandamme P, Corcoran GD, Sleator RD, Lucey B. Campylobacter corcagiensis sp. nov., isolated from faeces of captive lion-tailed macaques (Macaca silenus). Int J Syst Evol Microbiol. 2014;64:2878–2883. 10.1099/ijs.0.063867-0 . [DOI] [PubMed] [Google Scholar]
- 36.Gilbert MJ, Miller WG, Leger JS, Chapman MH, Timmerman AJ, Duim B, et al. Campylobacter pinnipediorum sp. nov. isolated from pinnipeds, comprising Campylobacter pinnipediorum subsp. pinnipediorum subsp. nov. and Campylobacter pinnipediorum subsp. caledonicus susbp. nov. Int J Syst Evol Microbiol. 2017;67:1961–1968. 10.1099/ijsem.0.001894 . [DOI] [PubMed] [Google Scholar]
- 37.Logan JM, Burnens A, Linton D, Lawson AJ, Stanley J. Campylobacter lanienae sp. nov., a new species isolated from workers in an abattoir. Int J Syst Evol Microbiol. 2000;50:865–872. 10.1099/00207713-50-2-865 . [DOI] [PubMed] [Google Scholar]
- 38.Van TT, Elshagmani E, Gor MC, Scott PC, Moore RJ. Campylobacter hepaticus sp. nov., isolated from chickens with spotty liver disease. Int J Syst Evol Microbiol. 2016;66:4518–4524. 10.1099/ijsem.0.001383 . [DOI] [PubMed] [Google Scholar]
- 39.Rossi M, Debruyne L, Zanoni RG, Manfreda G, Revez J, Vandamme P. Campylobacter avium sp. nov., a hippurate-positive species isolated from poultry. Int J Syst Evol Microbiol. 2009; 59:2364–2369. 10.1099/ijs.0.007419-0 . [DOI] [PubMed] [Google Scholar]
- 40.Inglis GD, Hoar BM, Whiteside DP, Morck DW. Campylobacter canadensis sp. nov., from captive whooping cranes in Canada. Int J Syst Evol Microbiol. 2007;57:2636–2644. 10.1099/ijs.0.65061-0 . [DOI] [PubMed] [Google Scholar]
- 41.Zanoni RG, Debruyne L, Rossi M, Revez J, Vandamme P. Campylobacter cuniculorum sp. nov., from rabbits. Int J Syst Evol Microbiol. 2009;59:1666–1671. 10.1099/ijs.0.007286-0 . [DOI] [PubMed] [Google Scholar]
- 42.Piccirillo A, Niero G, Calleros L, Pérez R, Naya H, Iraola G. Campylobacter geochelonis sp. nov. isolated from the western hermann’s tortoise (Testudo hermanni hermanni). Int J Syst Evol Microbiol. 2016;66:3468–3476. 10.1099/ijsem.0.001219 . [DOI] [PubMed] [Google Scholar]
- 43.Foster G, Holmes B, Steigerwalt AG, Lawson PA, Thorne P, Byrer DE, et al. Campylobacter insulaenigrae sp. nov., isolated from marine mammals. Int J Syst Evol Microbiol. 2004;54:2369–2373. 10.1099/ijs.0.63147-0 . [DOI] [PubMed] [Google Scholar]
- 44.Debruyne L, Broman T, Bergström S, Olsen B, On SL, Vandamme P. Campylobacter subantarcticus sp. nov., isolated from birds in the sub-Antarctic region. Int J Syst Evol Microbiol. 2010;60:815–819. 10.1099/ijs.0.011056-0 . [DOI] [PubMed] [Google Scholar]
- 45.Debruyne L, Broman T, Bergström S, Olsen B, On SL, Vandamme P. Campylobacter volucris sp. nov., isolated from black-headed gulls (Larus ridibundus). Int J Syst Evol Microbiol. 2010;60:1870–1875. 10.1099/ijs.0.013748-0 . [DOI] [PubMed] [Google Scholar]
- 46.Gilbert MJ, Kik M, Miller WG, Duim B, Wagenaar JA. Campylobacter iguaniorum sp. nov., isolated from reptiles. Int J Syst Evol Microbiol. 2015;65:975–982. 10.1099/ijs.0.000048 . [DOI] [PubMed] [Google Scholar]
- 47.Cáceres A, Muñoz I, Iraola G, Díaz-Viraqué F, Collado L. Campylobacter ornithocola sp. nov., a novel member of the Campylobacter lari group isolated from wild bird faecal samples. Int J Syst Evol Microbiol. 2017;67;1643–1649. 10.1099/ijsem.0.001822 . [DOI] [PubMed] [Google Scholar]
- 48.Stackebrandt E, Goebel BM. A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int J Syst Bacteriol. 1994;44:846–49. [Google Scholar]
- 49.Sakamoto M, Ohkuma M. Usefulness of the hsp60 gene for the identification and classification of Gram-negative anaerobic rods. J Med Microbiol. 2010;59:1293–1302. 10.1099/jmm.0.020420-0 . [DOI] [PubMed] [Google Scholar]
- 50.Kärenlampi RI, Tolvanen TP, Hänninen ML. Phylogenetic analysis and PCR-restriction fragment length polymorphism identification of Campylobacter species based on partial groEL gene sequences. J Clin Microbiol. 2004; 42 (12): 5731–5738. 10.1128/JCM.42.12.5731-5738.2004 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Rocha EP, Danchin A. Base composition bias might result from competition for metabolic resources. Trends Genet. 2002;18:291–294. 10.1016/S0168-9525(02)02690-2 . [DOI] [PubMed] [Google Scholar]
- 52.Mann S, Chen YP. Bacterial genomic G+C composition-eliciting environmental adaptation. Genomics. 2010;95:7–15. 10.1016/j.ygeno.2009.09.002 . [DOI] [PubMed] [Google Scholar]
- 53.Iraola G, Pérez R, Naya H, Paolicchi F, Pastor E, Valenzueça S, et al. Genomic evidence for emergence and evolution of pathogenicity and niche preferences in the genus Campylobacter. Genome Biol Evol. 2014;6(9):2392–2405. 10.1093/gbe/evu195 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Richter M, Rosselló-Móra R. Shifting the genomic gold standard for the prokaryotic species definition. Proc Natl Acad Sci U S A. 2009;106(45):19126–19131. 10.1073/pnas.0906412106 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Louwen R, Staals RHJ, Endtz HP, van Baarlen P, van der Oost J. The role of CRISPR-Cas systems in virulence of pathogenic bacteria. Microbiol Mol Biol Rev. 2014;78:74–88. 10.1128/MMBR.00039-13 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Westra ER, van Houte S, Gandon S, Whitaker R. The ecology and evolution of microbial CRISPR-Cas adaptive immune systems. Philos Trans R Soc Lond B Biol Sci. 2019;374(1772):20190101 10.1098/rstb.2019.0101 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ali A, Soares SC, Santos AR, Guimarães LC, Barbosa E, Almeida SS, et al. Campylobacter fetus subspecies: comparative genomics and prediction of potential virulence targets. Gene. 2012;508(2):145–56. 10.1016/j.gene.2012.07.070 . [DOI] [PubMed] [Google Scholar]
- 58.Krause-Gruszczynska M, van Alphen LB, Oyarzabal OA, Alter T, Hänel I, Schliephake A, et al. Expression patterns and role of CadF protein in Campylobacter jejuni and Campylobacter coli. FEMS Microbiol Lett. 2007;274:9–16. 10.1111/j.1574-6968.2007.00802.x . [DOI] [PubMed] [Google Scholar]
- 59.Ziprin RL, Young CR, Stanker LH, Hume ME, Konkel ME. The absence of cecal colonization of chicks by a mutant of Campylobacter jejuni not expressing bacterial fibronectin-binding protein. Avian Dis. 1999;43(3):686–589 . [PubMed] [Google Scholar]
- 60.Monteville MR, Yoon JE, Konkel ME. Maximal adherence and invasion of INT 407 cells by Campylobacter jejuni requires the CadF outer-membrane protein and microfilament reorganization. Microbiology. 2003;149:153–165. 10.1099/mic.0.25820-0 . [DOI] [PubMed] [Google Scholar]
- 61.Konkel ME, Kim BJ, Rivera-Amill V, Garvis SG. Identification of proteins required for the internalization of Campylobacter jejuni into cultured mammalian cells. Adv Exp Med Biol. 1999;473:215–224. 10.1007/978-1-4615-4143-1_22 . [DOI] [PubMed] [Google Scholar]
- 62.Konkel ME, Kim BJ, Rivera-Amill V, Garvis SG. Bacterial secreted proteins are required for the internalization of Campylobacter jejuni into cultured mammalian cells. Mol Microbiol. 1999;32(4):691–701. 10.1046/j.1365-2958.1999.01376.x . [DOI] [PubMed] [Google Scholar]
- 63.Guo B, Lin J, Reynolds DL, Zhang Q. Contribution of the multidrug efflux transporter CmeABC to antibiotic resistance in different Campylobacter species. Foodborne Pathog Dis. 2010;7(1):77–83. 10.1089/fpd.2009.0354 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lin J, Sahin O, Michel LO, Zhang Q. Critical role of multidrug efflux pump CmeABC in bile resistance and in vivo colonization of Campylobacter jejuni. Infect Immun. 2003;71(8):4250–4259. 10.1128/IAI.71.8.4250-4259.2003 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Gibreel A, Wetsch NM, Taylor DE. Contribution of the CmeABC efflux pump to macrolide and tetracycline resistance in Campylobacter jejuni. Antimicrob Agents Chemother. 2007;51(9):3212–3216. 10.1128/AAC.01592-06 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lin J, Michel LO, Zhang Q. CmeABC functions as a multidrug efflux system in Campylobacter jejuni. Antimicrob Agents Chemother. 2002;46(7):2124–2131. 10.1128/AAC.46.7.2124-2131.2002 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gorkiewicz G, Kienesberger S, Schober C, Scheicher SR, Gülly C, Zechner R, et al. A genomic island defines susbspecies-specific virulence features of the host-adapted pathogen Campylobacter fetus subsp. venerealis. J Bacteriol. 2010;192(2):502–517. 10.1128/JB.00803-09 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kienesberger S, Trummler CS, Fauster A, Lang S, Sprenger H, Gorkiewicz G, et al. Interbacterial macromolecular transfer by the Campylobacter fetus subsp. venerealis type IV secretion system. J Bacteriol. 2011;193(3):744–758. 10.1128/JB.00798-10 . [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the manuscript.