

Abstract
Objective:
The objective of this study was to investigate the effects of trimetazidine (TMZ) and coenzyme Q10 (CoQ10) on cisplatin-induced cardiotoxicity in rat cardiomyocytes.
Methods:
Rat cardiomyocytes were isolated and subjected to cisplatin (200 µM) treatment with and without TMZ (200 µM) and CoQ10 (200 mg/L) pretreatment. The cell viability, apoptosis, oxidant and antioxidant indicators, and mitochondrial dysfunction were examined.
Results:
TMZ or CoQ10 significantly attenuated cisplatin-induced cell viability inhibition (p<0.01) and apoptosis (p<0.001), and the combined use of TMZ and CoQ10 pretreatment exerted a pronounced effect compared to the effects of using each of these agents individually (p<0.05). TMZ or CoQ10 inhibited the levels of reactive oxidative species (ROS, p<0.01) and malondialdehyde (MDA, p<0.001 and p<0.01, respectively), elevated the activities of antioxidant enzymes superoxide dismutase (SOD, p<0.01) and catalase (CAT, p<0.01 and p<0.05, respectively), evidently enhanced nuclear translocation of nuclear factor erythroid 2-related factor 2 (Nrf2, p<0.05), alleviated mitochondrial membrane potential (ΔΨm) loss (p<0.05), and down-regulated the release of cytochrome c (cyto-c) into the cytosol (p<0.01) in cisplatin-treated cells. The combined use of TMZ and CoQ10 treatment was more effective than using either agent alone (p<0.01 for ROS, MDA, CAT, and cytosolic cyto-c; p<0.05 for SOD, nuclear Nrf2, and ΔΨm loss).
Conclusion:
TMZ and CoQ10 showed protective effects against cisplatin-induced cardiotoxicity via attenuating oxidative stress.
Keywords: cisplatin-induced cardiotoxicity, trimetazidine, coenzyme Q10, oxidative stress, mitochondrial dysfunction
Introduction
Cisplatin, a chemotherapeutic agent, is one of the most commonly used drugs for various types of malignancies; however, it has severe adverse effects (1). Cardiotoxicity events have been reported in many cases during or shortly after cisplatin treatment, including arrhythmias, ventricular hypertrophy, myocarditis, and angina pectoris (2, 3). Therefore, there is an absolute need to attenuate cisplatin-induced cardiotoxicity as clinical interventions have met little achievement (3, 4).
Oxidative stress is considered to be one of the major mechanisms involved in cisplatin-induced cardiotoxicity, as characterized by reactive oxidative species (ROS) production, lipid peroxidation, and inhibition of antioxidant enzymes such as glutathione (GSH), superoxide dismutase (SOD), and catalase (CAT) (5-7). On the other hand, excessive ROS can target mitochondria, resulting in mitochondrial dysfunction and release of cytochrome c (cyto-c) into the cytosol, which can subsequently induce caspase-dependent apoptosis (1, 8, 9). Hence, we hypothesize that timely intervention on oxidative stress may be an effective measure to prevent against cisplatin-induced cardiotoxicity.
The metabolic modulator trimetazidine (TMZ) is an antianginal drug, which reduces fatty acid oxidation via inhibition of 3-ketoacyl CoA thiolase and enhances myocardial glucose oxidation (10, 11). Coenzyme Q10 (CoQ10) is an essential component of the electron transport chain and participates in energy production. It is a fat-soluble antioxidant employed for the treatment of heart failure and other diseases (12). Evidence has shown that TMZ or CoQ10 can also be used as a supplement for cancer patients undergoing chemotherapy, and can effectively attenuate the side effects of the chemo-drugs, such as nephrotoxicity, hepatotoxicity, and ototoxicity because of their antioxidant properties (13-17). There is also a study showing the effective protection of CoQ10 against doxorubicin-induced cardiotoxicity (12). TMZ and CoQ have been reported to inhibit ROS accumulation and lipid peroxidation (15, 18), and alleviate ROS-triggered mitochondrial dysfunction (13, 19). Therefore, we speculate that TMZ and/or CoQ10 might protect against cisplatin-induced cardiotoxicity.
Nuclear factor erythroid 2-related factor 2 (Nrf2) has been considered a crucial modulator of antioxidant defense. Nrf2 is normally inactivated in the cytosol associated with its negative regulator Keap1. In response to stimuli, Nrf2 dissociates from Keap1, translocates to the nucleus, and up-regulates the expression of downstream antioxidant enzymes including SOD and CAT through binding to the antioxidant response element. Therefore, Nrf2 exerts a protective effect against ROS-induced injuries, including cardiotoxicity (20-22). Previous studies have indicated that Nrf2 is associated with cisplatin-induced toxicities (23, 24) and CoQ10-mediated antioxidant properties (23, 24); however, the possible involvement of Nrf2 in cisplatin-induced cardiotoxicity as well as TMZ and/or CoQ10-mediated antioxidant cardioprotection is largely unknown.
In this study, we aimed to demonstrate, for the first time, the possible protective effects of TMZ and/or CoQ10 on cisplatin-induced cardiotoxicity in vitro and illustrate the possible underlying mechanisms. These findings highlight the potential of TMZ and CoQ10 in protecting cancer patients from cisplatin-induced heart dysfunction.
Methods
Drugs and antibodies
Cisplatin and TMZ were purchased from MCE (Shanghai, China). CoQ10 was from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against Nrf2, Lamin A, and COX IV were from Abcam (Cambridge, MA, USA). Antibody against cyto-c was from Affinity Bioreagents (Golden, CO, USA). Antibody against GAPDH was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Isolation of rat cardiomyocytes
Ventricular myocytes were isolated from neonatal (2 days) Sprague Dawley (SD) rats as described in Ref. (25). The animals were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Briefly, hearts were excised from anesthetized rats, after which ventricles were separated, washed with D-Hanks solution three times, and minced into small fragments. Tissue fragments were further dissociated by repeated cycles of trypsinization using 0.0625% trypsin at 37 °C and centrifuged. Then, cell suspensions were preplated for 2 h into 100-mm dishes in Dulbecco’s modified Eagle medium/F-12 (Gibco, CA, USA) supplemented with 10% fetal bovine serum (Hyclone, UT, USA), 100 U/mL penicillin, and 100 µg/mL streptomycin (Gibco, CA, USA). The unattached cardiomyocytes were collected, adjusted to 5.0×105 cells/mL, and plated into 96-well plates (2×104 cells/well) or into 6-well plates (5×105 cells/well) until being used, up to five days. The experiments were approved by the Animal Care and Use Committee of Fudan University.
Cell viability
Cells were seeded into a 96-well culture plate and treated with cisplatin ranging from 0 to 800 µM for different periods of time (6 h, 12 h, 24 h, and 48 h). Cell viability was measured using Counting Kit-8 (CCK8) (Sigma-Aldrich, MO, USA). The CCK8 solution (10 µL) was added to each well and the plate was subsequently incubated at 37 °C for 2 h. The optical density of each well was measured using an enzyme linked immunosorbent assay (ELISA) reader at 450 nm.
To analyze the effect of TMZ and CoQ10 on cisplatin-treated cardiomyocytes, cells were divided into five groups according to the treatment, respectively, as follows: no treatment (control group), 200 µM of cisplatin treatment (cisplatin group), 200 µM of TMZ pretreatment (cisplatin+TMZ group), 200 mg/L of CoQ10 pretreatment (cisplatin+CoQ10 group), and a combination of TMZ and CoQ10 pretreatment (cisplatin+TMZ+CoQ10) prior to cisplatin application. Pretreatment with TMZ or/and CoQ10 was carried out 2 h prior to cisplatin treatment. CCK8 assay was performed as described above.
Cell apoptosis
Cells were seeded into 6-well plates and subjected to different treatments. The apoptotic cells were measured using an Annexin V-PE/7-AAD Apoptosis Detection Kit (Yeasen, Shanghai, China). Briefly, the cells were harvested, washed with ice-cold phosphate-buffered saline (PBS), and resuspended in 1× cold binding buffer. Next, cells were stained with 5 µL of Annexin V/PE and 10 µL of 7-AAD for 15 min at room temperature in darkness. Finally, the cells were subjected to flow cytometry (BD Biosciences, NJ, USA) and analyzed by CellQuest software (BD Biosciences, NJ, USA).
Mitochondrial membrane potential
JC-1 fluorochrome was employed to measure the mitochondrial membrane potential (ΔΨm) according to the manufacturer’s protocol (Cayman Chemicals, MI, USA). Cells were seeded into 6-well plates and subjected to different treatments. After treatment, the cells were incubated with 5 µM/L of JC-1 working solution for 15 min at 37 °C in darkness. Next, the cells were washed with PBS, resuspended in 1× assay buffer, and analyzed using a flow cytometer (BD Biosciences, NJ, USA).
Nuclear and cytoplasmic extraction
The lysates of nucleus and cytoplasm were prepared using a Nuclear/Cytosol Fractionation Kit (Biovision, CA, USA) following the manufacturer’s instructions. Briefly, cells were washed with ice-cold PBS and collected by centrifugation at 500×g for 3 min. The cell pellets were resuspended in ice-cold cytosol extraction buffer (CEB)-A and CEB-B containing protein inhibitors, incubated in ice for 10 min, and centrifuged at 12000×g for 5 min. The supernatant was obtained as a cytosol extract. The pellet was washed with PBS, incubated with a nuclear extraction buffer for 30 min, and then centrifuged at 16000×g for 10 min. The supernatant was saved as the nuclear fraction. All the steps were performed in ice or at 4 °C.
Mitochondrial and cytoplasmic extraction
Mitochondrial and cytoplasmic extracts were prepared using a Mitochondria/Cytosol Fractionation Kit (Biovision, CA, USA). Cells were collected and incubated with CEB, similarly as described above. Cells were homogenized in a Dounce tissue grinder (30 passes) and centrifuged at 700×g for 10 min. The supernatant was centrifuged at 10000×g for 30 min and the resulting supernatant was collected as the cytosolic fraction. The pellet was resuspended in mitochondrial extraction buffer, vortexed for 10 s as the mitochondrial fraction. All the steps were performed in ice or at 4 °C.
Western blot
Equal amounts of protein were denatured in a boiling water bath and then separated by 10% dodecyl sulfate polyacrylamide gels (SDS-PAGE), followed by being transferred to polyvinylidene difluoride (PVDF) membranes. Subsequently, the membranes were probed with 5% non-fat milk in tris-buffered saline and incubated with primary antibodies specific for Nrf2 and cyto-c. GAPDH, Lamin A, and COX IV served as the internal controls of cytosolic, nuclear, and mitochondrial fractions, respectively. After being washed five times, the membranes were incubated with Horseradish Peroxidase (HRP)-conjugated secondary antibodies at room temperature for 60 min and detected with a Novex® ECL Chemiluminescent Substrate Reagent Kit (Thermo Fisher Scientific, MA, USA) using a ChemiDoc™ XRS+ imaging system (Bio-Rad, California, USA). The signal intensities were quantified by ImageJ software.
Oxidative and antioxidant activity
To detect intracellular ROS production, cells were incubated with 50 µM of the DCFH-DA probe (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) for 30 min at 37 °C. The fluorescence intensities were measured using a fluorescence microplate reader at the excitation and emission wavelengths of 488 and 525 nm, respectively. SOD and CAT activities and malondialdehyde (MDA) content in cells were measured using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s protocols.
Statistical analysis
All data were analyzed using the GraphPad Prism 5 software (San Diego, CA, USA) and expressed as mean ± SD. The significance of differences between the groups was determined using the one-way analysis of variance (ANOVA) test followed by Tukey’s analysis. One-way repeated-measures ANOVA and post-hoc analysis (Bonferroni’s test) were performed for exploration of differences in cell viability upon treatment with 200 µM cisplatin for different periods of time. A p-value below 0.05 is generally considered statistically significant.
Results
TMZ and CoQ10 attenuated cisplatin-induced changes in cell viability and apoptosis
Cell viability was found to be significantly decreased in a dose- and time-dependent manner in cisplatin-treated cells than in control (p<0.001; Fig. 1a and 1b). Exposure to 200 µM of cisplatin for 24 h was chosen for the subsequent experiments. As expected, TMZ or CoQ10 pretreatment significantly attenuated the inhibitory effect of cisplatin on cell viability (p<0.01; Fig. 1c). The combined pretreatment with TMZ and CoQ10 significantly enhanced cell viability to a greater extent than pretreatment with either of TMZ or CoQ10 alone (p<0.05; Fig. 1c).
Figure 1.
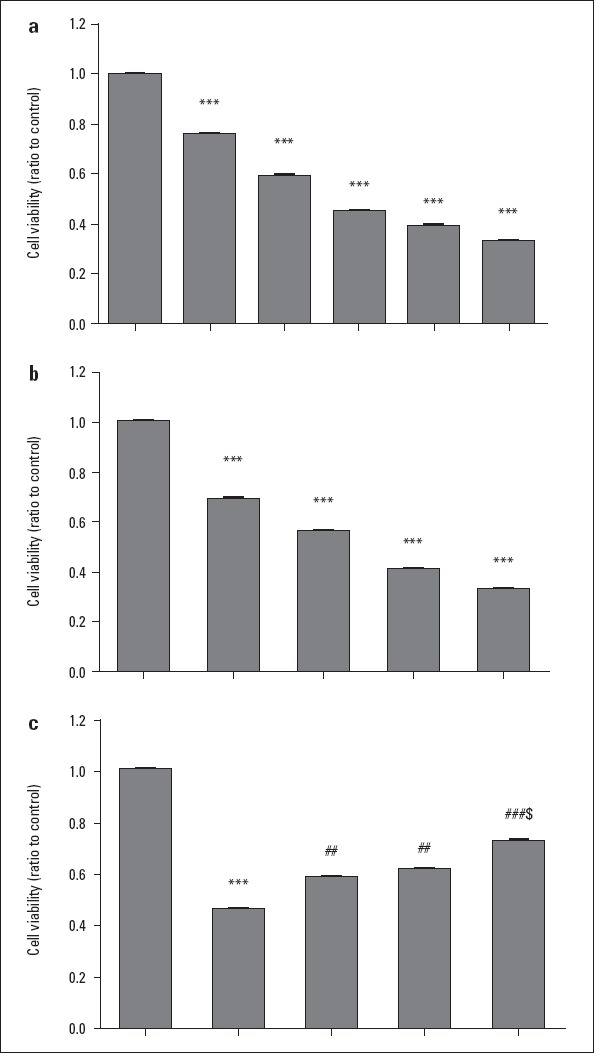
Toxic effect of cisplatin on the viability of rat cardiomyocytes and protective effects of TMZ and CoQ10. (a) Cell viability upon cisplatin (50 to 800 µM) treatment for 24 h. (b) Cell viability upon 200 µM cisplatin treatment for different periods of time (0, 6, 12, 24, and 48 h). (c) Cell viability in control, cisplatin, cisplatin+TMZ, cisplatin+CoQ10, and cisplatin+TMZ+CoQ10 groups. Data were normalized to that of control (assigned as a value of 1) and presented as mean±SD. ***P<0.001 as compared with control; ##P<0.01, and ###P<0.001 as compared with cisplatin-treated cells; and $P<0.05 as compared with TMZ or CoQ10-pretreated cells
TMZ - trimetazidine; CoQ10 - coenzyme Q10
On the other hand, cell apoptosis was analyzed by flow cytometry. As seen in Figure 2a and 2b, a significant induction of apoptosis was observed with 200 µM of cisplatin (p<0.001). However, TMZ or CoQ10 pretreatment markedly suppressed cisplatin-induced cell apoptosis (p<0.001), and the combined pretreatment with TMZ and CoQ10 exerted a more pronounced effect than pretreatment with each of these agents individually (p<0.05).
Figure 2.

TMZ and CoQ10 reduced cisplatin-induced cardiomyocyte apoptosis. (a) Representative scatter plot of cell apoptosis in control, cisplatin, cisplatin+TMZ, cisplatin+CoQ10, and cisplatin+TMZ+CoQ10 groups. (b) Apoptotic rates presented as mean±SD. ***P<0.001 as compared with control; ###P<0.001 as compared with cisplatin-treated cells; and $P<0.05 as compared with TMZ or CoQ10-pretreated cells
TMZ - trimetazidine; CoQ10 - coenzyme Q10
TMZ and CoQ10 inhibited ROS and MDA levels and elevated SOD and CAT activities in cisplatin-treated cells
As shown in Table 1, cisplatin caused significant increases in levels of ROS and MDA to about 2.8±0.06 (p<0.001) and 5.66±0.38 (p<0.001) fold, respectively, as compared to control, whereas these increases were effectively suppressed by the pretreatment with TMZ (p<0.01 for ROS; p<0.001 for MDA) or CoQ10 (p<0.01 for both). Furthermore, the combined pretreatment with TMZ and CoQ10 significantly inhibited ROS and MDA levels than pretreatment with TMZ or CoQ alone (p<0.01).
Table 1.
Trimetazidine and CoQ10 inhibited the levels of ROS and MDA and improved the activities of SOD and CAT in cisplatin-treated cells
| Factor | Control | Cisplatin | Cisplatin+TMZ | Cisplatin+CoQ10 | Cisplatin+TMZ+CoQ10 |
|---|---|---|---|---|---|
| ROS | 1±0.09 | 2.81±0.11*** | 2.36±0.08## | 2.21±0.1## | 1.77±0.08###$$ |
| MDA | 1±0.03 | 5.66±0.67*** | 3.75±0.17### | 4.1±0.31## | 2.77±0.22###$$ |
| SOD | 1±0.14 | 0.43±0.07*** | 0.58±0.05 | 0.55±0.01 | 0.67±0.02 |
| CAT | 1±0.03 | 0.25±0.05*** | 0.44±0.04## | 0.37±0.02# | 0.66±0.05###$$ |
P<0.001 as compared with control;
P<0.05,
P<0.01, and
P<0.001 as compared with cisplatin-treated cells; $P<0.05 and
P<0.01 as compared with TMZ or CoQ10-pretreated cells. TMZ – trimetazidine; CoQ10 - coenzyme Q10; ROS - reactive oxidative species; MDA – malondialdehyde; SOD - superoxide dismutase; CAT - catalase
In addition to elevating ROS and MDA production, antioxidant indicators such as SOD and CAT were analyzed. As indicated in Table 1, cisplatin was able to significantly decrease the activities of SOD (p<0.001) and CAT (p<0.001) compared to control. However, significant increases in SOD and CAT activities were observed upon pretreatment with TMZ (p<0.01 for both) or CoQ10 (p<0.01 for SOD; p<0.05 for CAT) than in cisplatin-treated cells. The combined pretreatment with TMZ and CoQ10 was more effective than using each of these drugs alone (p<0.05 for SOD; p<0.01 for CAT).
TMZ and CoQ10 enhanced nuclear Nrf2 expression in cisplatin-treated cells
Western blot data elucidated Nrf2 expression. As seen in Figure 3 and Table 2, nuclear Nrf2 expression was significantly decreased in cisplatin-treated cells than in the control (p<0.001). TMZ or CoQ10 pretreatment evidently enhanced Nrf2 expression in the nuclear fraction compared to cisplatin treatment alone (p<0.05). The combined pretreatment with TMZ and CoQ10 remarkably stimulated translocation of Nrf2 to the nucleus than pretreatment with each of these two drugs alone (p<0.05). However, the expression levels of Nrf2 in the cytosolic fractions among the groups did not differ significantly (p>0.05; Table 2).
Figure 3.
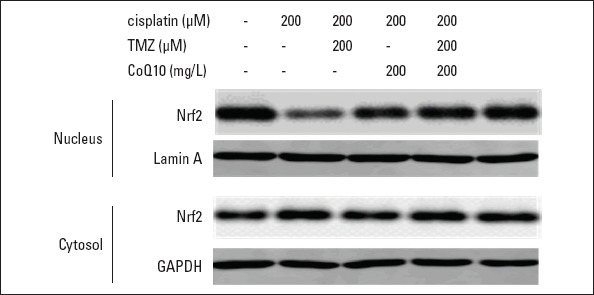
TMZ and CoQ10 enhanced nuclear Nrf2 expression in cisplatin-treated cells. Representative images of nuclear and cytosolic Nrf2 expression. GAPDH and Lamin A were used as the internal controls of cytosolic and nuclear fractions, respectively
TMZ - trimetazidine; CoQ10 - coenzyme Q10
Table 2.
Trimetazidine and CoQ10 enhanced nuclear Nrf2 expression and suppressed the release of cyto-c into cytosol in cisplatin-treated cells
| Factor | Control | Cisplatin | Cisplatin+TMZ | Cisplatin+CoQ10 | Cisplatin+TMZ+CoQ10 |
|---|---|---|---|---|---|
| Nrf2/Lamin A | 1±0.17 | 0.41±0.09*** | 0.6±0.07# | 0.63±0.1# | 0.87±0.06###$ |
| Cyto-c/GAPDH | 1±0.29 | 17.55±2.94*** | 10.76±1.35## | 9.65±1.78## | 6.71±1.02###$ |
| Nrf2/GAPDH | 1±0.15 | 1.23±0.12 | 1±0.16 | 1.06±0.07 | 0.91±0.19 |
| Cyto-c/COX IV | 1±0.22 | 0.24±0.07*** | 0.47±0.05# | 0.56±0.11## | 0.65±0.12###$ |
P<0.001 as compared with control;
P<0.05,
P<0.01, and
P<0.001 as compared with cisplatin-treated cells;
P<0.05 as compared with TMZ or CoQ10-pretreated cells.
TMZ - trimetazidine; CoQ10 - coenzyme Q10
TMZ and CoQ10 alleviated cisplatin-induced ΔΨm loss
JC-1 staining was used for the analysis of changes in ∆Ψm. As shown in Figure 4a and 4b, control cells emitted a red fluorescence, indicating the integrity of ∆Ψm. Cisplatin-treated cells showed a significant decrease in the red/green fluorescence intensity ratio (p<0.001), indicating loss of ∆Ψm. By contrast, TMZ or CoQ10 pretreatment markedly inhibited the loss of ∆Ψm (p<0.05). Furthermore, the combined pretreatment with TMZ and CoQ10 attenuated ∆Ψm loss to a greater extent than using either TMZ or CoQ10 alone (p<0.05).
Figure 4.

TMZ and CoQ10 attenuated cisplatin-induced ΔΨm dysfunction in cardiomyocytes. (a) Representative images of JC-1 staining. (b) Quantitative analysis of JC-1 staining presented as mean±SD. ***P<0.001 as compared with control; #P<0.05 and ###P<0.001 as compared with cisplatin-treated cells; and $P<0.05 as compared with TMZ or CoQ10-pretreated cells
TMZ - trimetazidine; CoQ10 - coenzyme Q10
TMZ and CoQ10 suppressed the release of cyto-c into cytosol caused by cisplatin
The expression of cyto-c was measured by Western blot. As displayed in Figure 5 and Table 2, cyto-c was mainly expressed in mitochondria in control cells. Cisplatin treatment led to a significantly increased expression of cytosolic cyto-c (p<0.001; Fig. 5 and Table 2) and a significantly decreased expression of mitochondrial cyto-c (p<0.001; Table 2) compared to control, indicating the release of cyto-c into cytosol. Pretreatment with TMZ or CoQ10 down-regulated the expression level of cytosolic cyto-c (p<0.01; Fig. 5 and Table 2) and up-regulated the level of mitochondrial cyto-c (p<0.05 and p<0.01, respectively; Fig. 5 and Table 2). Furthermore, the combined pretreatment with TMZ and CoQ10 showed a remarkable decrease in the expression of cytosolic cyto-c (p<0.05; Fig. 5 and Table 2) and a significant increase in the expression of mitochondrial cyto-c (p<0.05; Table 2).
Figure 5.
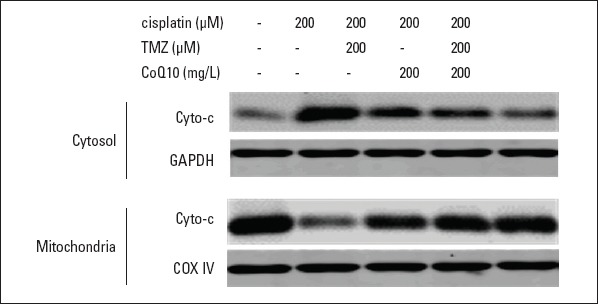
TMZ and CoQ10 suppressed the release of cyto-c into cytosol in cisplatin-treated cells. Representative images of mitochondrial and cytosolic cyto-c expression. GAPDH and COX IV served as the internal controls of cytosolic and mitochondrial fractions, respectively
TMZ - trimetazidine; CoQ10 - coenzyme Q10
Discussion
Evidence has shown that cisplatin chemotherapy can lead to cardiotoxicity (8, 26). This is in agreement with the present study where cisplatin significantly inhibited cell viability and induced apoptosis of rat cardiomyocytes. TMZ and CoQ10 are widely used for the management of heart failure diseases. Increasing evidence has shown that TMZ or CoQ10 can attenuate cisplatin-induced toxicities including nephrotoxicity, hepatotoxicity, and ototoxicity (13-17). Despite a lack of reports on the protective effect of TMZ and CoQ10 against cisplatin-induced cardiotoxicity, there is one study showing that CoQ10 can attenuate doxorubicin-induced cardiotoxicity (12). The results of the present study show that TMZ (200 µM) and CoQ10 (200 mg/L) significantly reduce apoptosis and increase cell viability in rat cardiomyocytes treated with cisplatin (200 µM), first demonstrating the cardioprotective role of TMZ and CoQ10 against cisplatin in vitro.
Oxidative stress is the main reason for cisplatin-induced cardiotoxicity (5-7). Experimental findings have suggested that up-regulated levels of ROS and MDA and down-regulated activities of antioxidant enzymes SOD and CAT are involved in cisplatin-induced cardiotoxicity, which agreed with the results of the present study (5-7). ROS overload can cause disruption or consumption of antioxidant defenses (2, 27, 28). In addition, excessive ROS may also induce membrane lipid peroxidation, which releases toxic products like MDA (29, 30), directly leading to cardiotoxicity (4). TMZ and CoQ10 have potent antioxidant properties that may ameliorate cisplatin-induced toxicities such as nephrotoxicity and ototoxicity (13-18). In the present study, TMZ and CoQ10 significantly prevented increases in ROS and MDA production and decreases in SOD and CAT activities caused by cisplatin. This is in accordance with some studies which suggest that TMZ and CoQ10 might elevate the activities of the antioxidant enzymes and act as free radical scavengers (13-18). In addition, MDA is directly associated with multiple tissue injuries. Other studies have suggested that TMZ and CoQ10 can ameliorate tissue toxicities through inhibition of MDA (15, 18).
Nrf2 is responsible for the activation of downstream antioxidant enzymes including SOD and CAT. Some investigations have shown suppressed Nrf2 activity associated with cisplatin-induced toxicities such as ototoxicity and nephrotoxicity (31, 32). This is similar to the present study where cisplatin significantly reduced the nuclear expression level of Nrf2 in rat cardiomyocytes. This result can partially explain why cisplatin attenuated the activities of SOD and CAT. Nrf2 has been proven to protect against ROS-induced cardiotoxicity (20-22) and is associated with TMZ and CoQ10-mediated antioxidant properties (23, 24, 33). Similarly, our data showed that TMZ or CoQ10 pretreatment could induce Nrf2 activation via promoting Nrf2 nuclear translocation, which was augmented by the combined use of TMZ and CoQ10. Furthermore, elevated nuclear expression of Nrf2 was paralleled with the raised activities of SOD and CAT caused by TMZ and CoQ10, indicating that TMZ and CoQ10-mediated enhanced antioxidant enzyme expression is attributable, in part, to increased Nrf2 activity.
Cardiomyocytes are mitochondria-rich cells. Cardiac mitochondrial dysfunction often occurs upon ROS accumulation and overproduction caused by cisplatin, which results in disruption of mitochondrial membrane potential and release of pro-apoptotic factor (8). In the present study, loss of ΔΨm and release of cyto-c into cytosol were observed in rat cardiomyocytes treated with cisplatin. Cyto-c translocation into the cytosol often precedes ΔΨm loss and is regarded as a sign of severe mitochondrial damage (34), which can subsequently induce caspase-dependent apoptosis of cardiomyocytes (8, 26). However, these disruptions caused by cisplatin can be improved by TMZ and CoQ10, suggesting that TMZ and CoQ10 may inhibit cell apoptosis partially through suppressing cisplatin-induced mitochondrial dysfunction. Other studies have also reported the positive modulation of TMZ and CoQ10 on mitochondrial function in damaged cardiomyocytes (35, 36), similar to the results of our study.
Conclusion
In conclusion, our study demonstrated that TMZ and CoQ10 significantly suppressed cisplatin-induced cardiotoxicity partially dependent on the Nrf2-related antioxidant defense system, which inhibits oxidative stress and mitochondrial dysfunction-mediated apoptosis. A combination of TMZ and CoQ10 might represent potential therapeutic agents for amelioration of cisplatin-induced cardiotoxicity.
Footnotes
Conflict of interest: None declared.
Peer-review: Externally peer-reviewed.
References
- 1.Dasari S, Tchounwou PB. Cisplatin in cancer therapy:molecular mechanisms of action. Eur J Pharmacol. 2014;740:364–78. doi: 10.1016/j.ejphar.2014.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.El-Awady el-SE, Moustafa YM, Abo-Elmatty DM, Radwan A. Cisplatin-induced cardiotoxicity:Mechanisms and cardioprotective strategies. Eur J Pharmacol. 2011;650:335–41. doi: 10.1016/j.ejphar.2010.09.085. [DOI] [PubMed] [Google Scholar]
- 3.Patane S. Cardiotoxicity:cisplatin and long-term cancer survivors. Int J Cardiol. 2014;175:201–2. doi: 10.1016/j.ijcard.2014.04.238. [DOI] [PubMed] [Google Scholar]
- 4.Kalam K, Marwick TH. Role of cardioprotective therapy for prevention of cardiotoxicity with chemotherapy:a systematic review and meta-analysis. Eur J Cancer. 2013;49:2900–9. doi: 10.1016/j.ejca.2013.04.030. [DOI] [PubMed] [Google Scholar]
- 5.Dugbartey GJ, Peppone LJ, de Graaf IA. An integrative view of cisplatin-induced renal and cardiac toxicities:Molecular mechanisms, current treatment challenges and potential protective measures. Toxicology. 2016;371:58–66. doi: 10.1016/j.tox.2016.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Topal I, Ozbek Bilgin A, Keskin Cimen F, Kurt N, Suleyman Z, Bilgin Y, et al. The effect of rutin on cisplatin-induced oxidative cardiac damage in rats. Anatol J Cardiol. 2018;20:136–42. doi: 10.14744/AnatolJCardiol.2018.32708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bahadır A, Ceyhan A, Öz Gergin Ö, Yalçın B, Ülger M, Özyazgan TM, et al. Protective effects of curcumin and beta-carotene on cisplatin-induced cardiotoxicity:An experimental rat model. Anatol J Cardiol. 2018;19:213–21. doi: 10.14744/AnatolJCardiol.2018.53059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Qian P, Yan LJ, Li YQ, Yang HT, Duan HY, Wu JT, et al. Cyanidin ameliorates cisplatin-induced cardiotoxicity via inhibition of ROS-mediated apoptosis. Exp Ther Med. 2018;15:1959–65. doi: 10.3892/etm.2017.5617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ozben T. Oxidative stress and apoptosis:impact on cancer therapy. J Pharm Sci. 2007;96:2181–96. doi: 10.1002/jps.20874. [DOI] [PubMed] [Google Scholar]
- 10.Onay-Besikci A, Ozkan SA. Trimetazidine revisited:a comprehensive review of the pharmacological effects and analytical techniques for the determination of trimetazidine. Cardiovasc Ther. 2008;26:147–65. doi: 10.1111/j.1527-3466.2008.00043.x. [DOI] [PubMed] [Google Scholar]
- 11.Molinari F, Pin F, Gorini S, Chiandotto S, Pontecorvo L, Penna F, et al. The mitochondrial metabolic reprogramming agent trimetazidine as an 'exercise mimetic'in cachectic C26-bearing mice. J Cachexia Sarcopenia Muscle. 2017;8:954–73. doi: 10.1002/jcsm.12226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chen PY, Hou CW, Shibu MA, Day CH, Pai P, Liu ZR, et al. Protective effect of Co-enzyme Q10 On doxorubicin-induced cardiomyopathy of rat hearts. Environ Toxicol. 2017;32:679–89. doi: 10.1002/tox.22270. [DOI] [PubMed] [Google Scholar]
- 13.Fatima S, Al-Mohaimeed N, Al-Shaikh Y, Tyagi P, Banu N, Hasan S, et al. Combined treatment of epigallocatechin gallate and Coenzyme Q10 attenuates cisplatin-induced nephrotoxicity via suppression of oxidative/nitrosative stress, inflammation and cellular damage. Food Chem Toxicol. 2016;94:213–20. doi: 10.1016/j.fct.2016.05.023. [DOI] [PubMed] [Google Scholar]
- 14.Scasso F, Sprio AE, Canobbio L, Scanarotti C, Manini G, Berta GN, et al. Dietary supplementation of coenzyme Q10 plus multivitamins to hamper the ROS mediated cisplatin ototoxicity in humans:A pilot study. Heliyon. 2017;3:e00251. doi: 10.1016/j.heliyon.2017.e00251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.El-Sherbeeny NA, Attia GM. The protective effect of trimetazidine against cisplatin-induced nephrotoxicity in rats. Can J Physiol Pharmacol. 2016;94:745–51. doi: 10.1139/cjpp-2015-0472. [DOI] [PubMed] [Google Scholar]
- 16.Ateyya H, Yosef H, Nader MA. Ameliorative effect of trimetazidine on cisplatin-induced hepatotoxicity in rats. Can J Physiol Pharmacol. 2016;94:225–30. doi: 10.1139/cjpp-2015-0304. [DOI] [PubMed] [Google Scholar]
- 17.Astolfi L, Simoni E, Valente F, Ghiselli S, Hatzopoulos S, Chicca M, et al. Coenzyme Q10 plus Multivitamin Treatment Prevents Cisplatin Ototoxicity in Rats. PLoS One. 2016;11:e0162106. doi: 10.1371/journal.pone.0162106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mirmalek SA, Gholamrezaei Boushehrinejad A, Yavari H, Kardeh B, Parsa Y, Salimi-Tabatabaee SA, et al. Antioxidant and Anti-Inflammatory Effects of Coenzyme Q10 on L-Arginine-Induced Acute Pancreatitis in Rat. Oxid Med Cell Longev. 2016;2016:5818479. doi: 10.1155/2016/5818479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Steggall A, Mordi IR, Lang CC. Targeting Metabolic Modulation and Mitochondrial Dysfunction in the Treatment of Heart Failure. Diseases. 2017;5:E14. doi: 10.3390/diseases5020014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.El-Agamy DS, El-Harbi KM, Khoshhal S, Ahmed N, Elkablawy MA, Shaaban AA, et al. Pristimerin protects against doxorubicin-induced cardiotoxicity and fibrosis through modulation of Nrf2 and MAPK/NF-kB signaling pathways. Cancer Manag Res. 2018;11:47–61. doi: 10.2147/CMAR.S186696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Shinjo T, Tanaka T, Okuda H, Kawaguchi AT, Oh-Hashi K, Terada Y, et al. Propofol induces nuclear localization of Nrf2 under conditions of oxidative stress in cardiac H9c2 cells. PLoS One. 2018;13:e0196191. doi: 10.1371/journal.pone.0196191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hao E, Lang F, Chen Y, Zhang H, Cong X, Shen X, et al. Resveratrol alleviates endotoxin-induced myocardial toxicity via the Nrf2 transcription factor. PLoS One. 2013;8:e69452. doi: 10.1371/journal.pone.0069452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li L, Du J, Lian Y, Zhang Y, Li X, Liu Y, et al. Protective Effects of Coenzyme Q10 Against Hydrogen Peroxide-Induced Oxidative Stress in PC12 Cell:The Role of Nrf2 and Antioxidant Enzymes. Cell Mol Neurobiol. 2016;36:103–11. doi: 10.1007/s10571-015-0224-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kabel AM, Elkhoely AA. Ameliorative Effect of Coenzyme Q10 and/or Candesartan on Carboplatin-Induced Nephrotoxicity:Roles of Apoptosis, Transforming Growth Factor-Beta1, Nuclear Factor Kappa-B And The Nrf2/HO-1 Pathway. Asian Pac J Cancer Prev. 2017;18:1629–36. doi: 10.22034/APJCP.2017.18.6.1629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liao XD, Wang XH, Jin HJ, Chen LY, Chen Q. Mechanical stretch induces mitochondria-dependent apoptosis in neonatal rat cardiomyocytes and G2/M accumulation in cardiac fibroblasts. Cell Res. 2004;14:16–26. doi: 10.1038/sj.cr.7290198. [DOI] [PubMed] [Google Scholar]
- 26.Zhang P, Yi LH, Meng GY, Zhang HY, Sun HH, Cui LQ. Apelin-13 attenuates cisplatin-induced cardiotoxicity through inhibition of ROS-mediated DNA damage and regulation of MAPKs and AKT pathways. Free Radic Res. 2017;51:449–59. doi: 10.1080/10715762.2017.1313414. [DOI] [PubMed] [Google Scholar]
- 27.Kisaoglu A, Borekci B, Yapca OE, Bilen H, Suleyman H. Tissue damage and oxidant/antioxidant balance. Eurasian J Med. 2013;45:47–9. doi: 10.5152/eajm.2013.08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Karthikeyan K, Bai BR, Devaraj SN. Cardioprotective effect of grape seed proanthocyanidins on isoproterenol-induced myocardial injury in rats. Int J Cardiol. 2007;115:326–33. doi: 10.1016/j.ijcard.2006.03.016. [DOI] [PubMed] [Google Scholar]
- 29.Yüce A, Ateşşahin A, Ceribaşi AO, Aksakal M. Ellagic acid prevents cisplatin-induced oxidative stress in liver and heart tissue of rats. Basic Clin Pharmacol Toxicol. 2007;101:345–9. doi: 10.1111/j.1742-7843.2007.00129.x. [DOI] [PubMed] [Google Scholar]
- 30.Yousef MI, Saad AA, El-Shennawy LK. Protective effect of grape seed proanthocyanidin extract against oxidative stress induced by cisplatin in rats. Food Chem Toxicol. 2009;47:1176–83. doi: 10.1016/j.fct.2009.02.007. [DOI] [PubMed] [Google Scholar]
- 31.Youn CK, Jo ER, Sim JH, Cho SI. Peanut sprout extract attenuates cisplatin-induced ototoxicity by induction of the Akt/Nrf2-mediated redox pathway. Int J Pediatr Otorhinolaryngol. 2017;92:61–6. doi: 10.1016/j.ijporl.2016.11.004. [DOI] [PubMed] [Google Scholar]
- 32.Wang SW, Xu Y, Weng YY, Fan XY, Bai YF, Zheng XY, et al. Astilbin ameliorates cisplatin-induced nephrotoxicity through reducing oxidative stress and inflammation. Food Chem Toxicol. 2018;114:227–36. doi: 10.1016/j.fct.2018.02.041. [DOI] [PubMed] [Google Scholar]
- 33.Zhang H, Liu M, Zhang Y, Li X. Trimetazidine Attenuates Exhaustive Exercise-Induced Myocardial Injury in Rats via Regulation of the Nrf2/NF-κB Signaling Pathway. Front Pharmacol. 2019;10:175. doi: 10.3389/fphar.2019.00175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Eleftheriadis T, Pissas G, Liakopoulos V, Stefanidis I. Cytochrome c as a Potentially Clinical Useful Marker of Mitochondrial and Cellular Damage. Front Immunol. 2016;7:279. doi: 10.3389/fimmu.2016.00279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kuzmicic J, Parra V, Verdejo HE, Lopez-Crisosto C, Chiong M, Garcia L, et al. Trimetazidine prevents palmitate-induced mitochondrial fission and dysfunction in cultured cardiomyocytes. Biochem Pharmacol. 2014;91:323–36. doi: 10.1016/j.bcp.2014.07.022. [DOI] [PubMed] [Google Scholar]
- 36.Kido K, Ito H, Yamamoto Y, Makita K, Uchida T. Cytotoxicity of propofol in human induced pluripotent stem cell-derived cardiomyocytes. J Anesth. 2018;32:120–31. doi: 10.1007/s00540-017-2441-0. [DOI] [PMC free article] [PubMed] [Google Scholar]