

Abstract
Anthropogenic climate change induces unprecedented variability in a broad range of environmental parameters. These changes will impact material properties and animal biomechanics, thereby affecting animal performance and persistence of populations. Climate change implies warming at the global level, and it may be accompanied by altered wind speeds, wave action, ocean circulation, acidification as well as increased frequency of hypoxic events. Together, these environmental drivers affect muscle function and neural control and thereby movement of animals such as bird migration and schooling behaviour of fish. Altered environmental conditions will also modify material properties of animals. For example, ocean acidification, particularly when coupled with increased temperatures, compromises calcified shells and skeletons of marine invertebrates and byssal threads of mussels. These biomechanical consequences can lead to population declines and disintegration of habitats. Integrating biomechanical research with ecology is instrumental in predicting the future responses of natural systems to climate change and the consequences for ecosystem services such as fisheries and ecotourism.
Keywords: Climate change, Biomechanics, Locomotion, Animals
Here, we summarise how the dominant environmental changes associated with climate change can impact the biomechanics and movement of organisms.
Introduction
Anthropogenic climate change will dominate ecosystem flux for the next decades if not centuries. The recent literature has given a much clearer picture of the expected impacts of global warming on physiological performance and thermal tolerance (Gunderson and Stillman, 2015; Sinclair et al., 2016). However, the impacts of climate change are much broader, including changes in ocean circulation and acidification, increased frequency of hypoxic events, increased storm activity, altered rainfall patterns and flow regimes of freshwater streams and rivers (Aarnes et al., 2017; Baynes et al., 2018; Doney, 2010; Doney et al., 2009). These changes will challenge the biomechanical performance of organisms.
Biomechanics is concerned with the function of biological systems and materials that underlie animal structure and movement, and environmentally induced changes in biomechanical properties will have far-reaching consequences for ecosystems (Higham et al., 2016). For example, increased storm activity and wave action can have pronounced impacts on intertidal communities. The biomechanical stress of increased wave action can damage and remove macroalgae and invertebrates from rocky shores (Jensen and Denny, 2016; Jonsson et al., 2006). Some organisms have biomaterial properties that increase resilience to high physical impacts (Blamires et al., 2017; O'Donnell et al., 2013). Nonetheless, environmental changes such as in temperature, physical impacts stemming from wave of wind action, and ocean acidification, interact and thereby increase the total impact on individuals and on interactions between species (Domenici et al., 2017b; Kroeker et al., 2016). Anthropogenic climate change represents a dominant selection pressure and one that is novel to most ecosystems. It is timely now to highlight the importance of biomechanics for climate change responses: the field has much to offer to increase understanding of the effects of climate change, but it has not yet made a concerted effort to address the new problems arising with the Anthropocene. The challenge lies in integrating large-scale biogeographic features of climate with small-scale physiological and biomechanical properties of individuals (Torossian et al., 2016). Biomechanics can answer how environmental stressors affect the mechanical properties and the motion of organisms and can develop predictive mechanistic models (Gaylord et al., 2001) that can significantly advance understanding of the ecological consequences of climate change. Below, we summarise how the dominant environmental changes associated with climate change can impact the biomechanics and movement of organisms.
Musculoskeletal function and movement
Animal movement depends on muscle and neural function, the cardiovascular system and metabolism, and the performance of these physiological systems is sensitive to environmental perturbations such as changes in temperature, pH, oxygen level, water flow and wind speed (Fig. 1). Climate change influences these parameters both by causing steady increases in temperature and decreases in oceanic pH, for example, and by increasing extreme climate events (van de Pol et al., 2017). Environmental impacts on animal movement can compromise ecological networks such as food web structures and energy transfer between trophic levels by modulating predator–prey interactions (Gibert et al., 2016).
Figure 1.
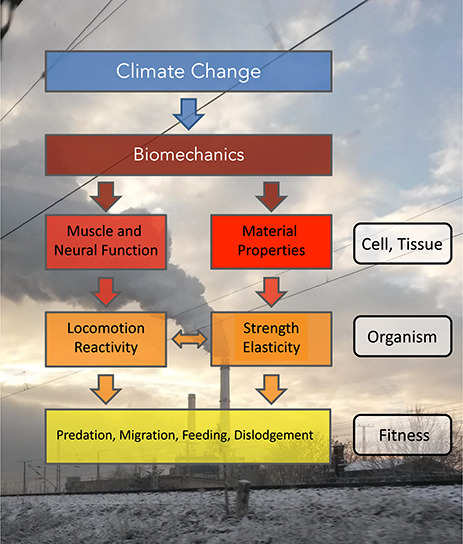
The effects of climate change on biomechanics and their potential consequences on the fitness of organisms. Conceptual diagram showing the effects of climate change at various biological levels. Climate change can affect: (i) muscle/neural function and (ii) material properties. In turn, these can affect the locomotor performance and reactivity of organism, as well as their strength and elasticity. These changes in organismal performance can lead to changes at the fitness level, in terms of vulnerability to predators and dislodgment, as well as feeding.
The movement capacity of individuals and species will determine the relevant scale (from local to global) of environmental variability that affects performance and ultimately reproductive success. Migrating birds, for example, are exposed to local variation at their breeding and overwintering grounds and to global variation as a result of migration, and together these will determine reproductive success (Kentie et al., 2018; Shaffer et al., 2006). Altered wind and air currents modulate the biomechanics and energetics of bird flight and thereby affect survivorship and demographics (McCabe et al., 2017; Sapir et al., 2014; Weisshaupt et al., 2017). In contrast, sessile organisms (e.g. intertidal mussels) are exposed only to local, but nonetheless complex, environmental variation (Choi et al., 2019). Species with greater range of movement, such as migrating fish and birds, essentially undergo extensive bouts of endurance exercise while exposed to relatively large-scale climate variation. Endurance exercise is sensitive to muscle fatigue mediated by biomechanical constraints of intrinsic muscle function and metabolic energy supply (Allen et al., 2008; Biewener, 2016). The combined effects of environmental parameters such as altered temperatures and flows (e.g. air and ocean currents, streams) can increase the cost of transport (Halsey, 2016; Seebacher et al., 2016) and decrease muscle power output (James and Tallis, 2019), thereby compromising the movement objective (Martin et al., 2015). Migrating salmon, for example, face reduced reproductive success because warmer water and altered flow conditions affect migration success (Eliason et al., 2011; Fenkes et al., 2016).
Climate change has already led to increased temperatures and increased wave heights in oceans (Aarnes et al., 2017; Hemer et al., 2013; Young et al., 2011). Increased wave height results in greater impact force and may therefore lead to increased physiological demand on marine organisms (Bejarano et al., 2017; Forrester et al., 2016). At the same time, temperature changes alter muscle contractile function (James and Tallis, 2019), which can modify the impact of other drivers. Intertidal organisms are exposed to several environmental drivers concurrently (Choi et al., 2019), and wave action is of particular importance because it can dislodge individuals and effectively remove them from the population (Jensen and Denny, 2016). One of the most important traits for survival of intertidal organisms is the adhesion strength to the substrate (Trussell et al., 1993), and the foot muscle of snails, for example, has to produce greater force for the animal to stay in situ as wave action increases (Forrester et al., 2016). At the same time, force needs to be produced longer if the increase in wave action is chronic. Increased wave action elicited a beneficial training effect on muscle by increasing tenacity, but increasing water temperature decreased endurance in an intertidal snail (Nerita atramentosa) (Clayman and Seebacher, 2019). Ocean warming would therefore negate the beneficial increase in tenacity that could render snails more resistant to acute impacts of wave action.
Warmer waters are often associated with low oxygen concentrations (Nudds et al., 2020). Hypoxia and increasing temperature together influence metabolic scope and swimming energetics and kinematics (Claireaux and Lefrancois, 2007; Domenici et al., 2013, Nudds et al., 2020), which can lead to altered schooling behaviour in fish (Domenici et al., 2017a). Increasing ocean temperature is often also associated with decreasing pH. Both as single and multiple stressors, acidification, temperature, and hypoxia can have pronounced effects on escape kinematics of fish that alter predator–prey interactions (Domenici et al., 2019). Temperature often has a stronger effect than other environmental drivers (Domenici et al., 2019; Nudds et al., 2020). Overall, the climate-induced interaction between environmental drivers affects escape responses and swimming kinematics by decreasing muscle performance and interfering with brain and sensory function (Domenici et al., 2019). Animals may compensate for changes in their environment, and acclimation can render individuals more resilient to environmental variability (Le Roy et al., 2017). However, there is considerable variation between species in their capacity for plastic responses, which may determine spatial patterns of distribution of different species across climate gradients (Padilla et al., 2019). One of the main challenges associated with predictions about the effect of global warming at the ecological level is that body temperature affects individual performance and that temperature varies through time within an individual and through space between individuals in a population (Denny, 2019). Theoretical work suggests that for any given individual this variation decreases performance and the range of viable temperatures (i.e. the range of temperatures within which animals can reproduce successfully). However, variation among individuals increases the viable range of the population (Denny, 2019). Clearly, investigating the effects of thermal history (as well as that of other main stressors) and plastic responses to variation at different time scales needs to be a priority in order to improve predictions of the effects of climate change on organisms.
In terrestrial environments, climate change can alter wind speeds (Young et al., 2011), and altered wind speed patterns can influence migratory success in birds (Nourani et al., 2017). Wind speed patterns can also disrupt predator–prey interaction by physically disturbing and dislodging individuals and by disrupting locomotion (Cherry and Barton, 2017). The terrestrial environment, including wind speed and temperature, can influence the energetic cost of transport (Chappell et al., 2004; Halsey, 2016), which is a function of running kinematics (Kram and Taylor, 1990). Plastic responses to temperature can affect muscle and locomotor performance (James, 2013; James and Tallis, 2019; Padilla et al., 2019), but probably not cost of transport (Jahn and Seebacher, 2019).
Structural properties
Ocean acidification, in particular, has a strong negative effect on the structural properties of skeletons and shells of many marine invertebrates. Ocean acidification compromises calcification and growth twofold: the hypercapnia associated with increased CO2 causes an energetic constraint and allocation trade-off that curtails growth, and it reduces the calcium carbonate building blocks essential for calcification (Byrne and Fitzer, 2019). Increasing temperature can exacerbate this allocation trade-off. For example, shell strength of the mussel Mytilus edulis was negatively affected by increasing water temperatures which caused an allocation trade-off, where energy was directed away from shell formation to support temperature-induced increases in maintenance costs (Mackenzie et al., 2014).
The combined effects of ocean acidification and increasing temperature are similar in bryozoans. Bryozoans are a phylum of aquatic invertebrates, which play an important ecological role as bioconstructors in shallow marine habitats. Their calcareous skeleton is quite different in structure and composition from that of other marine calcifying organisms such as molluscs. However, current and predicted future decreases in ocean pH and increases in temperature are detrimental for calcification of their skeleton, as well as growth rate and reproduction (Taylor et al., 2014). In contrast, climate change-induced decreases in pH and warming had opposite effects in the tube built by the tube worm Hydroides elegans. Decreased pH weakened the tube and lowered resistance to predator attack, while increasing temperature increased mineral density and resistance to predator attack (Li et al., 2016)
Corals are particularly sensitive to the combined changes in storm activity and wave action and ocean acidification. Branching corals are more susceptible to the combination of increased wave action and storm activity combined with sea-level rise than massive corals (Baldock et al., 2014). Vulnerability of corals to wave stress is exacerbated particularly when corals lose condition and structural strength (Baldock et al., 2014). These individual responses scale up to populations and may render populations vulnerable to decline and extinction. Acidification rendered populations of Acropora hyacinthus vulnerable to collapse by altering calcification rates and thereby compromising growth and skeletal strength (Madin et al., 2012). These biomechanical impacts may be exacerbated by ocean warming, which can cause decreases in condition and coral bleaching (Hughes et al., 2017; Mellin et al., 2019).
Similar to corals, the interactions between environmental drivers have detrimental ecological impacts on marine mussels (Kroeker et al., 2016). Mussels are common on temperate rocky shores around the world, where they form dense aggregations that provide essential habitat for ecological communities, and mussels are important as human food sources (Seitz et al., 2013). Given their abundance and ecological and commercial importance, mussels have become a model system to study the interactions between individual responses to interacting environmental drivers and their consequences for population dynamics and persistence (Carrington et al., 2015). Mussels attach to their substrate via byssal threads, and the biomechanical properties of the threads, and their sensitivity to environmental impacts such as ocean warming and acidification determines the survival of individuals and the persistence of populations (Newcomb et al., 2019). Acidification and high temperature can affect mussel attachment strength and reduce mussel survival, though mussels may be more vulnerable to the negative effects of ocean warming than ocean acidification (Newcomb et al., 2019). However, single environmental drivers such as increased CO2 do not necessarily reduce attachment strength of the byssal thread (Dickey et al., 2018). In addition to using multiple stressors, recent studies have made considerable advances in understanding the ecological relevance of climate change studies by using realistic fluctuating environmental conditions such as those likely to be experienced by organisms in nature (Frieder et al., 2014; Gobler et al., 2017; Jarrold et al., 2017). Chan and Tong (2020) found that the responses of larval sea urchins (measured as growth of larval arms) were less pronounced when they were exposed to fluctuating low pH conditions, compared to a constant low pH. Therefore, particularly for larvae which are subject to varying environmental conditions throughout their dispersal, further work on the rate of physiological response to realistic (i.e. fluctuating) environmental conditions along the dispersive pathway is fundamental in order to predict the effects of climate change (Chan and Tong 2020).
Terrestrial ecosystems are of course also exposed to warming climates and storm activity. However, other environmental drivers such as rainfall patterns and drought and wind regimes have unique effects on non-aquatic habitats (Trenberth, 2011). The extended phenotype of terrestrial organisms may show plasticity in response to this environmental variation. Similar to the byssal thread of mussels, the strength and performance of other biological materials such as spider silk is influenced by environmental conditions (Blamires et al., 2017). Climate change can affect spider webs by altering environmental variables that impact the physiological processes responsible for the production of silk. Temperature and humidity, in particular, can alter web architecture and performance (Blamires and Sellers, 2019).
Conclusions
Climate change and its consequences are unavoidable now. A greater understanding of the impact of climate change on existing ecosystems will translate to better management and conservation of natural resources and biodiversity. The combined impacts of climate change on the biomechanics of animals are mainly due to effects on material properties and muscle/neural functions. These effects can have consequences for the fitness of the individual through their impacts on organismal strength and locomotor performance (Figure 1). Research in the area can be encapsulated by ecological biomechanics (ecomechanics), which provides a mechanistic framework linking environment-driven changes in the form and function of individuals, to community interactions and populations (Denny and Gaylord, 2010). This approach has been used successfully to model mussel biology with a focus on linking material properties to molecular and population responses (Carrington et al., 2015). Biomechanical research is of particular interest here because it can link organismal traits to species interactions and community structure in the face of unprecedented environmental variability resulting from anthropogenic climate change (Gaylord et al., 2019). It will be of particular importance that predictive models incorporate interactions between environmental drivers and natural fluctuations in those drivers.
In a broader context, the synthesis provided here can be applied to practical problems. The natural environment provides essential services to human societies. Ecological biomechanics can facilitate understanding of the mechanistic basis underlying many practical problems associated with climate change and can be a key element for developing relevant management and conservation strategies. For example, the sustainability of fisheries around the world depends on prediction of large-scale movements of commercial species. Movement of fish schools will depend on prevailing thermal conditions and sufficient food supply, which in turn will be affected by ocean acidification and ocean currents. These relationships, and others associated with aquaculture, farming, ecotourism, etc., will continue to change, and there is an urgent need for better mechanistic basis that can help predict the direction and magnitude of that change.
Funding
F.S. was supported by the Australian Research Council Discovery Grant DP180103036. P.D. was supported by European Union’s Horizon 2020 research and innovation program under the grant agreement No. 773713 (PANDORA).
Contributor Information
Paolo Domenici, IAS-CNR, Località Sa Mardini, Torregrande, Oristano, 09170 Italy.
Frank Seebacher, School of Life and Environmental Sciences A08, University of Sydney, Sydney, NSW 2006, Australia.
References
- Aarnes OJ, Reistad M, Breivik Ø, Bitner-Gregersen E, Ingolf Eide L, Gramstad O, Magnusson AK, Natvig B, Vanem E (2017) Projected changes in significant wave height toward the end of the 21st century: Northeast Atlantic. J Geophys Res 122: 3394–3403. [Google Scholar]
- Allen DG, Lamb GD, Westerblad H (2008) Skeletal muscle fatigue: cellular mechanisms. Physiol Rev 88: 287–332. [DOI] [PubMed] [Google Scholar]
- Baldock TE, Karampour H, Sleep R, Vyltla A, Albermani F, Golshani A, Callaghan DP, Roff G, Mumby PJ (2014) Resilience of branching and massive corals to wave loading under sea level rise – a coupled computational fluid dynamics-structural analysis. Mar Pollut Bull 86: 91–101. [DOI] [PubMed] [Google Scholar]
- Baynes ERC, Lageweg WI, McLelland SJ, Parsons DR, Aberle J, Dijkstra J, Henry P-Y, Rice SP, Thom M, Moulin F (2018) Beyond equilibrium: re-evaluating physical modelling of fluvial systems to represent climate changes. Earth-Sci Rev 181: 82–97. [Google Scholar]
- Bejarano S, Jouffray J-B, Chollett I, Allen R, Roff G, Marshell A, Steneck R, Ferse SCA, Mumby PJ (2017) The shape of success in a turbulent world: wave exposure filtering of coral reef herbivory. Funct Ecol 31: 1312–1324. [Google Scholar]
- Biewener AA. (2016) Locomotion as an emergent property of Muscle contractile dynamics. J Exp Biol 219: 285–294, 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blamires SJ, Blackledge TA, Tso I-M (2017) Physicochemical property variation in spider silk: ecology, evolution, and synthetic production. Annu Rev Entomol 62: 443–460. [DOI] [PubMed] [Google Scholar]
- Blamires SJ, Sellers W (2020) Temperature and humidity effects on orb web performance: implications for spider foraging in a changing climate. Conserv Physiol 7: coz083. doi: 10.1093/conphys/coz083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Byrne M, Fitzer S (2019) The impact of environmental acidification on the microstructure and mechanical integrity of marine invertebrate skeletons. Conserv Physiol 7: coz062. doi: 10.1093/conphys/coz062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrington E, Waite JH, Sará G, Sebens KP (2015) Mussels as a model system for integrative ecomechanics. Annu Rev Mar Sci 7: 443–469. [DOI] [PubMed] [Google Scholar]
- Chan KYK, Tong CSD (2019) Temporal variability modulates pH impact on larval sea urchin development. Conserv Physiol in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chappell MA, Garland T, Rezende EL, Gomes FR (2004) Voluntary running in deer mice: speed, distance, energy costs and temperature effects. J Exp Biol 207: 3839–3854. [DOI] [PubMed] [Google Scholar]
- Cherry MJ, Barton BT (2017) Effects of wind on predator-prey interactions. Food Webs 13: 92–97. [Google Scholar]
- Choi F, Gouhier T, Lima FP, Rilov G, Seabra R, Helmuth BS (2019) Mapping physiology: biophysical mechanisms define scales of climate change impacts. Conserv Physiol 7: coz028. doi: 10.1093/conphys/coz028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claireaux G, Lefrançois C (2007) Linking environmental variability and fish performance: integration through the concept of scope for activity. Philos Trans R Soc Lond B Biol Sci 362: 2031–2041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayman S, Seebacher F (2019) Increased wave action promotes muscle performance but increasing temperatures cause a tenacity–endurance trade-off in intertidal snails (Nerita atramentosa). Conserv Physiol 7: coz039. doi: 10.1093/conphys/coz039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny MW. (2019) Performance in a variable world: using Jensen's inequality to scale up from individuals to populations. Conserv Physiol 7: coz053. doi: 10.1093/conphys/coz053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denny MW, Gaylord B (2010) Marine ecomechanics. Annu Rev Mar Sci 2: 89–114. [DOI] [PubMed] [Google Scholar]
- Dickey G, Preziosi BM, Clark CT, Bowden TJ (2018) The impact of ocean acidification on the byssal threads of the blue mussel (Mytilus edulis). PLoS One 13: e0205908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domenici P, Allan BJM, Lefrançois C, McCormick MI (2019) The effect of climate change on the escape kinematics and performance of fishes: implications for future predator-prey interactions. Conserv Physiol 7: coz078. doi: 10.1093/conphys/coz078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domenici P, Herbert NA, Lefrancois C, Steffensen JF, McKenzie DJ (2013) The effects of hypoxia on fish swimming performance and behaviour In Palstra AP, Planas JV, eds, Swimming Physiology of Fish. Springer, Heidelberg. [Google Scholar]
- Domenici P, Steffensen JF, Marras S (2017a) The effect of hypoxia on fish schooling. Phil Trans R Soc B 372: 20160236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domenici P, Torres R, Manríquez PH (2017b) Effects of elevated carbon dioxide and temperature on locomotion and the repeatability of lateralization in a keystone marine mollusc. J Exp Biol 220: 667–676. [DOI] [PubMed] [Google Scholar]
- Doney SC. (2010) The growing human footprint on coastal and open-ocean biogeochemistry. Science 328: 1512–1516. [DOI] [PubMed] [Google Scholar]
- Doney SC, Fabry VJ, Feely RA, Kleypas JA (2009) Ocean acidification: the other CO2 problem. Annu Rev Mar Sci 1: 169–192. [DOI] [PubMed] [Google Scholar]
- Eliason EJ, Clark TD, Hague MJ, Hanson LM, Gallagher ZS, Jeffries KM, Gale MK, Patterson DA, Hinch SG, Farrell AP (2011) Differences in thermal tolerance among sockeye salmon populations. Science 332: 109–112. [DOI] [PubMed] [Google Scholar]
- Fenkes M, Shiels HA, Fitzpatrick JL, Nudds RL (2016) The potential impacts of migratory difficulty, including warmer waters and altered flow conditions, on the reproductive success of salmonid fishes. Comp Biochem Phys A 193: 11–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forrester GE, Macfarlan RJA, Holevoet AJ, Merolla S (2016) Dislodgement force and shell morphology vary according to wave exposure in a tropical gastropod (Cittarium pica). Mar Biol Res 12: 986–992. [Google Scholar]
- Frieder CA, Gonzalez JP, Bockmon EE, Navarro MO, Levin LA (2014) Can variable pH and low oxygen moderate ocean acidification outcomes for mussel larvae? Global Change Biology 20: 754–764. [DOI] [PubMed] [Google Scholar]
- Gaylord B, Barclay KM, Jellison BM, Jurgens LJ, Ninokawa AT, Rivest EB, Leighton R (2019) Ocean change within shoreline communities: from biomechanics to behaviour and beyond. Conserv Physiol 7: coz077. doi: 10.1093/conphys/coz077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaylord B, Hale BB, Denny MW (2001) Consequences of transient fluid forces for compliant benthic organisms. J Exp Biol 204: 1347–1360. [DOI] [PubMed] [Google Scholar]
- Gibert JP, Chelini M-C, Rosenthal MF, DeLong JP (2016) Crossing regimes of temperature dependence in animal movement. Glob Chan Biol 22: 1722–1736. [DOI] [PubMed] [Google Scholar]
- Gunderson AR, Stillman JH (2015) Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc R Soc B 282: 20150401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobler CJ, Clark HR, Griffith AW, Lusty MW (2017) Diurnal fluctuations in acidification and hypoxia reduce growth and survival of larval and juvenile bay scallops (Argopecten irradians) and hard clams (Mercenaria mercenaria). Front Mar Sci 3: 282. [Google Scholar]
- Halsey LG. (2016) Terrestrial movement energetics: current knowledge and its application to the optimising animal. J Exp Biol 219: 1424–1431. [DOI] [PubMed] [Google Scholar]
- Hemer MA, Fan Y, Mori N, Semedo A, Wang XL (2013) Projected changes in wave climate from a multi-model ensemble. Nat Clim Chang 3: 471–476. [Google Scholar]
- Higham TE, Rogers SM, Langerhans RB, Jamniczky HA, Lauder GV, Stewart WJ, Martin CH, Reznick DN (2016) Speciation through the lens of biomechanics: locomotion, prey capture and reproductive isolation. Proc R Soc B 283: 20161294–20161210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes TP et al. (2017) Coral reefs in the Anthropocene. Nature 546: 82–90. [DOI] [PubMed] [Google Scholar]
- Jahn M, Seebacher F (2019) Cost of transport is a repeatable trait but is not determined by mitochondrial efficiency in zebrafish (Danio rerio). J Exp Biol 222: jeb201400–jeb201407. [DOI] [PubMed] [Google Scholar]
- James RS. (2013) A review of the thermal sensitivity of the mechanics of vertebrate skeletal muscle. J Comp Physiol B 183: 723–733. [DOI] [PubMed] [Google Scholar]
- James RS, Tallis J (2019) The likely effects of thermal climate change on vertebrate skeletal muscle mechanics with possible consequences for animal movement and behaviour. Conserv Physiol 7: coz066. doi: 10.1093/conphys/coz066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarrold MD, Humphrey C, McCormick MI, Munday PL (2017) Diel CO2 cycles reduce severity of behavioural abnormalities in coral reef fish under ocean acidification. Sci Rep 7: 10153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen MM, Denny MW (2016) Life in an extreme environment: characterizing wave-imposed forces in the rocky intertidal zone using high temporal resolution hydrodynamic measurements. Limnol Oceanogr 61: 1750–1761. [Google Scholar]
- Jonsson PR, Granhag L, Moschella PS, Aberg P, Hawkins SJ, Thompson RC (2006) Interactions between wave action and grazing control the distribution of intertidal macroalgae. Ecology 87: 1169–1178. [DOI] [PubMed] [Google Scholar]
- Kentie R, Coulson T, Hooijmeijer JCEW, Howison RA, Loonstra AHJ, Verhoeven MA, Both C, Piersma T (2018) Warming springs and habitat alteration interact to impact timing of breeding and population dynamics in a migratory bird. Global Change Biol 24: 5292–5303. [DOI] [PubMed] [Google Scholar]
- Kram R, Taylor R (1990) Energetics of running: a new perspective. Nature 346: 265–267. [DOI] [PubMed] [Google Scholar]
- Kroeker KJ et al. (2016) Interacting environmental mosaics drive geographic variation in mussel performance and predation vulnerability. Ecol Lett 19: 771–779. [DOI] [PubMed] [Google Scholar]
- Le Roy A, Loughland I, Seebacher F (2017) Differential effects of developmental thermal plasticity across three generations of guppies (Poecilia reticulata): canalization and anticipatory matching. Sci Rep 7: 4313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C, Meng Y, He C, Chan VBS, Yao H, Thiyagarajan V (2016) Mechanical robustness of the calcareous tubeworm Hydroides elegans: warming mitigates the adverse effects of ocean acidification. Biofouling 32: 191–204. [DOI] [PubMed] [Google Scholar]
- Mackenzie CL, Ormondroyd GA, Curling SF, Ball RJ, Whiteley NM, Malham SK (2014) Ocean warming, more than acidification, reduces shell strength in a commercial shellfish species during food limitation. PLoS One 9: e86764–e86769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madin JS, Hughes TP, Connolly SR (2012) Calcification, storm damage and population resilience of tabular corals under climate change. PLoS One 7: e46637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin BT, Nisbet RM, Pike A, Michel CJ, Danner EM (2015) Sport science for salmon and other species: ecological consequences of metabolic power constraints. Ecol Lett 18: 535–544. [DOI] [PubMed] [Google Scholar]
- McCabe JD, Olsen BJ, Osti B, Koons PO (2017) The influence of wind selectivity on migratory behavioral strategies. Behav Ecol 29: 160–168. [Google Scholar]
- Mellin C et al. (2019) Spatial resilience of the great barrier reef under cumulative disturbance impacts. Glob Chan Biol 67: 345–315. [DOI] [PubMed] [Google Scholar]
- Newcomb LA, George MN, O'Donnell MJ, Carrington E (2019) Only as strong as the weakest link: structural analysis of the combined effects of elevated temperature and pCO2 on mussel attachment. Conserv Physiol 7: coz068. doi: 10.1093/conphys/coz068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nourani E, Yamaguchi NM, Higuchi H (2017) Climate change alters the optimal wind-dependent flight routes of an avian migrant. Proc R Soc B 284: 20170149–20170146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nudds RL, Ozolina K, Fenkes M, Wearing OH, Shiels HA (2020) Extreme temperature combined with hypoxia affects swimming performance in brown trout (Salmo trutta). Conserv Physiol in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Donnell MJ, George MN, Carrington E (2013) Mussel byssus attachment weakened by ocean acidification. Nat Clim Chang 3: 587–590. [Google Scholar]
- Padilla P, Ducret V, Bonneaud C, Courant J, Herrel A (2019) Acclimation temperature effects on locomotor traits differ in adult aquatic anurans (X. tropicalis and X. laevis) from different latitudes: implications for climate change. Conserv Physiol 7: coz019. doi: 10.1093/conphys/coz019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sapir N, Horvitz N, Wikelski M, Avissar R, Nathan R (2014) Compensation for lateral drift due to crosswind in migrating European bee-eaters. J Ornithol 155: 745–753. [Google Scholar]
- Seebacher F, Borg J, Schlotfeldt K, Yan Z (2016) Energetic cost determines voluntary movement speed only in familiar environments. J Exp Biol 219: 1625–1631. [DOI] [PubMed] [Google Scholar]
- Seitz RD, Wennhage H, Bergström U, Lipcius RN, Ysebaert T (2013) Ecological value of coastal habitats for commercially and ecologically important species. ICES J Mar Sci 71: 648–665. [Google Scholar]
- Shaffer SA et al. (2006) Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. Proc Natl Acad Sci U S A 103: 12799–12802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinclair BJ et al. (2016) Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures? Ecol Lett 19: 1372–1385. [DOI] [PubMed] [Google Scholar]
- Taylor PD, Lombardi C, Cocito S (2014) Biomineralization in bryozoans: present, past and future. Biol Rev 90: 1118–1150. [DOI] [PubMed] [Google Scholar]
- Torossian JL, Kordas RL, Helmuth B (2016) Cross-scale approaches to forecasting biogeographic responses to climate change. Adv Ecol Res 55: 371–433. [Google Scholar]
- Trenberth KE. (2011) Changes in precipitation with climate change. Clim Res 47: 123–138. [Google Scholar]
- Trussell GC, Johnson AS, Rudolph SG, Gilfillan ES (1993) Resistance to dislodgement: habitat and size-specific differences in morphology and tenacity in an intertidal snail. Mar Ecol Prog Ser 100: 135–144. [Google Scholar]
- van de Pol M, Jenouvrier S, Cornelissen JHC, Visser ME (2017) Behavioural, ecological and evolutionary responses to extreme climatic events: challenges and directions. Phil Trans R Soc B 372: 20160134–20160116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisshaupt N, Lehmann V, Arizaga J, Maruri M (2017) Radar wind profilers and avian migration: a qualitative and quantitative assessment verified by thermal imaging and moon watching. Methods Ecol Evol 8: 1133–1145. [Google Scholar]
- Young IR, Ziegler S, Barbanin AV (2011) Global trends in wind speed and wave height. Science 332: 451–455. [DOI] [PubMed] [Google Scholar]