

Abstract
Echinococcosis represents a major public health problem worldwide and is considered a neglected disease by the World Health Organization. The etiological agents are Echinococcus tapeworms, which display elaborate developmental traits that imply a complex control of gene expression. MicroRNAs (miRNAs), a class of small regulatory RNAs, are involved in the regulation of many biological processes such as development and metabolism. They act through the repression of messenger RNAs (mRNAs) usually by binding to the 3’ untranslated region (3’UTR). Previously, we described the miRNome of several Echinococcus species and found that miRNAs are highly expressed in all life cycle stages, suggesting an important role in gene expression regulation. However, studying the role of miRNAs in helminth biology remains a challenge. To develop methodology for functional analysis of miRNAs in tapeworms, we performed miRNA knockdown experiments in primary cell cultures of Echinococcus multilocularis, which mimic the development of metacestode vesicles from parasite stem cells in vitro. First, we analysed the miRNA repertoire of E. multilocularis primary cells by small RNA-seq and found that miR-71, a bilaterian miRNA absent in vertebrate hosts, is one of the top five most expressed miRNAs. Using genomic information and bioinformatic algorithms for miRNA binding prediction, we found a high number of potential miR-71 targets in E. multilocularis. Inhibition of miRNAs can be achieved by transfection of antisense oligonucleotides (anti-miRs) that block miRNA function. To this end, we evaluated a variety of chemically modified anti-miRs for miR-71 knockdown. Electroporation of primary cells with 2’-O-methyl modified anti-miR-71 led to significantly reduced miR-71 levels. Transcriptomic analyses showed that several predicted miR-71 targets were up-regulated in anti-miR-treated primary cells, including genes potentially involved in parasite development, host parasite interaction, and several genes of as yet unknown function. Notably, miR-71-silenced primary cell cultures showed a strikingly different phenotype from control cells and did not develop into fully mature metacestodes. These findings indicate an important function of miR-71 in Echinococcus development and provide, for the first time, methodology to functionally study miRNAs in a tapeworm.
Author summary
Echinococcosis, caused by the larval stages of tapeworms of the genus Echinococcus, is a neglected disease that affects millions of people world-wide. These parasites show elaborate developmental features that rely on a complex control of gene expression. microRNAs are small molecules which have been discovered in the last decades and control gene expression in animals, plants and viruses. microRNAs are highly expressed in several tapeworms but their biological function in these parasites is unknown. Assuming that microRNAs will be important for parasite development, we analysed the function of these molecules in Echinococcus multilocularis, employing an in vitro model that mimics the first developmental transitions which occur in the human host. By artificially decreasing the concentration of the highest expressed microRNA, we observed phenotypic alterations and inhibition of development. In addition, we identified possible mRNA molecules targeted by microRNAs and found that some of these are known for being involved in developmental processes in other organisms. This work provides novel methodology to study microRNA function in tapeworms. Furthermore, highly expressed parasite microRNAs that are absent in the host but fulfil an important role in parasite developmental processes can serve as selective drug targets against the underlying diseases.
Introduction
Echinococcosis represents a major public health and economic issue in many countries and is considered a neglected tropical disease by the World Health Organization (WHO) [1,2]. This zoonotic disease is caused by cestodes of the genus Echinococcus. The two major forms of the disease in humans are cystic and alveolar echinococcosis caused by E. granulosus sensu lato (s. l.) and E. multilocularis, respectively. To date, the primary focus for echinococcosis treatment relies on surgery of the metacestode or chemotherapy using benzimidazoles [3]. Alveolar echinococcosis is a life-threatening zoonosis prevalent in the Northern Hemisphere where foxes act as definitive hosts [4]. Infection of the mammalian intermediate host (rodents and humans) is initiated by oral uptake of infectious eggs, which contain the oncosphere larva. Upon hatching from the egg in the host intestine, the oncosphere penetrates the intestinal epithelium and gains access to the host organs. Typically within the liver the parasite then undergoes a developmental transition towards the metacestode stage which is entirely driven by parasite stem cells (germinative cells) that have been carried to the host by the oncosphere [5]. As an asexual multiplication stage the metacestode grows multivesicularly and infiltrates the surrounding host tissue, eventually leading to organ failure. In natural rodent infections, head regions of the future adult worm (protoscoleces) are formed from the cellular germinal layer of the metacestode, and are subsequently taken up when the definitive host takes its prey [6]. A crucial role of germinative cells in parasite development and metacestode formation has previously been demonstrated. In particular, the germinative cells were shown to be the only mitotic cell type of Echinococcus which gives rise to all differentiated cells [5,7]. The E. multilocularis primary cell cultivation system [8], which initially contains around 80% germinative cells [5], is routinely used in developmental and immunological studies to mimic, in vitro, the transition of oncosphere-derived stem cells into metacestode vesicles [9–12]. Furthermore, due to their decisive role in parasite proliferation, the germinative cells are one of the most important cell types for the development of novel chemotherapeutics against echinococcosis [13]. The particular characteristics of the indirect and complex Echinococcus life cycle suggest that all these development processes require tightly controlled regulation of gene expression. MicroRNAs (miRNAs), a class of small regulatory RNAs, are involved in the regulation of many biological processes, primarily through the repression of messenger RNAs (mRNAs) by typically binding to the 3’ untranslated region (3’UTR) of target genes [14]. Upon binding of miRNAs to the 3’UTR, target mRNAs are either cleaved or, more common in metazoans, destabilized or translationally repressed [14]. The main determinant of this interaction is the seed region, an evolutionarily conserved sequence located in the 5′ end of the miRNA (nucleotides 2–8). miRNAs have already been identified in several cestode species including E. multilocularis [15] and E. granulosus s. l. [15–17]. In previous work, we found that miRNAs are highly expressed in all life cycle stages analysed, suggesting an important function in gene regulation of these zoonotic parasites. In addition, we found miRNAs differentially expressed between life cycle stages and species, and identified both parasite-specific miRNAs as well as several which are divergent from miRNAs of the host. We also previously predicted miRNA targets in E. granulosus, E. multilocularis and Taenia solium, using computational tools [18,19]. The studies on miRNAs in cestodes so far mainly focused on miRNA identification and computational predictions of target genes which inevitably leads to high false positive rates [20]. For this reason, there is still a lack of comprehensive functional and experimental validation of these putative miRNA targets. miR-71-5p (miR-71 from now on), a bilaterian miRNA absent in vertebrate hosts, is one of the top five most expressed miRNAs in the Echinococcus germinative layer and protoscoleces [15–17], suggesting that miR-71 may play stage-independent roles relevant to parasite homeostasis [15]. In addition, it was the miRNA with the highest number of predicted targets, suggesting that miR-71 controls several cellular processes in Echinococcus [18]. Studying the role of miRNAs in parasite biology and host interaction still remains a challenge. To this end, bioinformatic strategies adapted to each phylum as well as experimental strategies such as miRNA knockdown have to be developed [21]. Up to now, successful knockdown of platyhelminth parasite miRNAs has only been demonstrated for Schistosoma japonicum [22].
In the present study, we aimed to determine the role of miR-71 in Echinococcus metacestode development. To this end, we performed miR-71 knockdown experiments in E. multilocularis primary cell cultures followed by analysis of metacestode ontogenesis at the morphological level. In addition, using whole transcriptome and bioinformatic analyses, we identified potential target genes of miR-71 in E. multilocularis.
Methods
Ethics statement
All experiments in animals were carried out in accordance with European and German regulations on the protection of animals (Tierschutzgesetz, Section 6). Ethical approval of the study was obtained by the local ethics committee of the government of Lower Franconia (permit no. 55.2–2531.01-61/13).
Animals and parasites
All experiments were performed with the E. multilocularis isolate Ingrid which was propagated in Mongolian jirds (Meriones unguiculatus) as previously described [8]. For miRNA identification and expression profiling of E. multilocularis primary cell culture isolate H95 was used. The isolation of parasite larvae from infected jirds was performed as previously described [7].
Echinococcus multilocularis primary cell culture
Primary cell culture was performed as described in the previous publication [8], with some modifications. Briefly, metacestode vesicles in co-culture with RH-cells (rat hepatoma cells) were sieved with a tea strainer, washed twice with sterile PBS and incubated twice for 30 seconds in sterile distilled water. Then, metacestode vesicles were disrupted with a 10 ml pipette and centrifuged for 10 min at 2,000g at room temperature. Vesicles were washed again with PBS and centrifuged for 5 min at 250 g.
In order to achieve more standardized samples, the yield of the primary cell isolation process was quantified by measuring the cell density photometrically [8]. Using polystyrene cuvettes of 10 x 4 mm, (Sarstedt, Nümbrecht, Germany) 12.5 μl of the initial cell suspension were diluted with 987.5 μl 1xPBS. The optical density of this dilution was measured at a wavelength of 600 nm employing a Hitachi U-2000 spectrophotometer (New York, USA). One cell unit was defined as the cell suspension with a DO600 of 0.01. The in vitro cultivation of primary cells was carried out in 6-well plates (Nunc, Roskilde, DK) using 1000 cell units as starting material and 5.0 ml of cDMEM-A6/B4, a conditioned Dulbecco's Modified Eagle Medium used for fast primary cell growth as previously described [8]. The primary cells were placed in a closed Ziploc bag gassed with N2 and incubated at 37ºC overnight.
Small RNA isolation, small RNA library construction and sequencing
Small RNA isolation, library construction and sequencing of three biological replicates from 48 h primary cell cultures of E. multilocularis (Fig 1) were carried out as described in Macchiaroli et al. [23]. Briefly, cells were mechanically homogenized in Trizol (Invitrogen) for 10 s. Then, 200 μl of chloroform:isoamyl alcohol (24:1) were added and mixed thoroughly. Phase separation was carried out by centrifugation at maximum speed at 4°C. Then, 0.5× isopropanol and 4 μl of glycogen (5 mg/ml) were added to the aqueous phase and the RNA was pelleted by centrifugation at maximum speed at 4°C for 30 min. The resulting pellet was washed with 70% ethanol, air dried, and resuspended in nuclease-free water. The amount and integrity of total RNA was determined using a 2100 BioAnalyzer (Agilent, USA). RNA was concentrated by ethanol precipitation at -20°C overnight after elimination of polyadenylated mRNA using oligo-dT dynabead. Small RNA libraries were prepared using the NEBNext Multiplex Small RNA Library Prep Set for Illumina. Libraries were paired-end sequenced using an Illumina sequencing platform (HiSeq 2500) for 100 cycles.
Fig 1. Workflow for identification of miR-71 potential target genes in Echinococcus multilocularis primary cell culture.

The oligonucleotides complementary to miR-71 used were: anti-miR-71-2´OMe: 2’-O-methyl oligonucleotides in the complete sequence, anti-miR-71-LNA: locked nucleic acid and anti-miR-71-partial-2´OMe: 2´-O-methyl in some nucleotides. Three biological replicates were used for each experiment.
miRNA identification and expression profiling of Echinococcus multilocularis primary cell culture
In order to identify miRNAs from the small RNA libraries, the miRDeep2 software package [24] was used. Unique sequences were mapped to the E. multilocularis genome and used as input for miRNA prediction as previously described [15,16]. The initial miRDeep2 output list of candidate miRNA precursors of each library were manually curated to generate a final high confidence set of miRNAs retaining only candidate precursors with i) miRDeep2 score ≥ 4 ii) mature reads in more than one biological sample iii) star reads and/or seed conservation iv) no match to rRNA, tRNA or mRNA. The secondary structures and the minimum free energy of putative precursors and clusters were predicted using the mfold web server [25] and RNAfold software [26], respectively. To analyze miRNA expression, read counts of each individual miRNA in a sample (biological replicate) were normalized to the total number of mature miRNA read counts in that sample as previously described [16]. Then, normalized miRNA reads counts were averaged between the three biological replicates. Correlation analyses between pairs of independent biological replicates were performed (Pearson’s correlation coefficient).
miR-71 knockdown
In order to search for the best knockdown strategy for miR-71 in primary cell culture of E. multilocularis we evaluated 3 chemically modified anti-miRs (Fig 1), all containing substitutions of one of the no-bridging phosphate oxygen atoms with a sulphur atom (phosphorthioate): (1) 2’-O-methyl oligonucleotides in the complete sequence (anti-miR-71-2´OMe), (2) locked nucleic acid (anti-miR-71-LNA) and (3) 2´-O-methyl oligonucleotides modification in some nucleotides (anti-miR-71-partial-2´OMe). As controls, we used scrambled oligonucleotides for the anti-miR-71-2´OMe and the anti-miR-71-partial-2´OMe conditions and a negative control oligonucleotide designed by EXIQON for the anti-miR-71-LNA. The anti-miR sequences and their corresponding scrambled and negative control are shown in S1 Table. Also, primary cell culture was electroporated without any oligonucleotide (mock control). In the primary cell cultivation system, cell aggregates that give rise to metacestode vesicles are formed from parasite stem cells within 2–3 weeks [7]. For the electroporation procedure, anti-miR was added to a final concentration of 3 μM to 100 μl of primary cell aggregate suspension in selfmade siPORT electroporation buffer [27]. Electroporation procedure was used previously in E. multilocularis primary cell culture [27]. Samples were placed into a 1 mm gap electroporation cuvette (Electroporation Cuvettes Plus-BXT) and pulsed once (200 V, 13Ω and 50 μF, time constant ∼0.6 msec). After a 12-min incubation step at 37ºC, the electroporated aggregates were washed with cDMEM-A6/B4 and carefully transferred into appropriate culture wells with cDMEM-A6/B4. The cells were cultured for 24 h prior to RNA extraction or for 30 days by observation every 2 days under an inverted light microscope (Leica DM IRB). Vitality, phenotype and normal development of the aggregates was evaluated comparing the cells treated with each type of anti-miR to those treated with scrambled or negative controls, with mock electroporated aggregates and with aggregates without electroporation. This last control was used to check the normal development of the metacestodes vesicles used in the experiment.
Total RNA isolation after knockdown experiments
Total RNA was extracted from E. multilocularis primary cell samples after 24 h in culture (Fig 1) using Direct-zolTM RNA MiniPrep (ZYMO RESEARCH) according to the manufacturer's protocol. To maximize precipitation of small RNA, the eluted RNA was incubated overnight at -20°C with the addition of 0.1 volumes of 3 M sodium acetate (pH 5.2), 2.5 volumes of ethanol and 2 μl of glycogen (10mg/ml). RNA was centrifuged at 14,000 g for 30 min at 4°C, washed in 75% ethanol, air dried at room temperature and resuspended in 20 μl of nuclease-free water. Samples were stored at -80°C until use. RNA concentration was determined using a Nanodrop 1000 spectrophotometer (Thermo Scientific) and RNA integrity was assessed with an Agilent 2100 Bioanalyzer (Agilent Technologies) using a RNA 6000 Nano chip.
RT-qPCR analysis of miR-71 in Echinococcus multilocularis primary cell culture after knockdown experiments
Knockdown of miR-71 and RT-qPCR (retrotranscription followed by real time polymerase chain reaction) was performed. For this, miRNA cDNA synthesis was performed as in Macchiaroli et al. [16] using 5 ng of input RNA. PCR was performed in a StepOne Plus cycler (Applied Biosystems, U.S.A.). The PCR mix consisted of 5 x HOT FIREPol EvaGreen qPCR Mix Plus 2 μl (1x), 0.4 μl (200nM) of each primer, 5.2 μl of water DNAse free and 2 μl of diluted cDNA. The cycling conditions were: 15 min at 95°C, followed by 40 x (15 s at 95°C, 20 s at 60°C) and a final step of 30 s at 72°C. Data collection was performed at 72°C. For primer design, the mature miRNA sequences identified in this work were used (S2 Table). In order to find a reference gene for normalization of the qPCR data, those miRNAs that showed high expression in primary cells of E. multilocularis were evaluated. The data were analysed by two programs: NormFinder [28] and BestKeeper [29]. The miR-4989 was chosen as reference gene since it fulfilled the stability requirements: i) low stability index according to NormFinder analysis ii) Cp standard deviation ≤0,5 with coefficient of correlation ~1 according to BestKeeper analysis (S1 Fig). Primer sequences, amplicon size and PCR efficiency values for all the primers are described in S2 Table.
Source of genomic and annotation data
The genomic, annotation and CDS transcript data of E. multilocularis [30] are based on the EMULTI002 assembly deposited in the NCBI (National Center for Biotechnology Information) database and was downloaded from the WormBase Parasite database WBPS11-WS265 (http://parasite.wormbase.org/).
Library construction, sequencing and bioinformatics analyses of RNA-seq data after knockdown experiments
To determine the effects of miR-71 knock-down on protein coding genes, mRNA libraries (paired-end reads) were prepared using TruSeq Stranded mRNA library prep (Illumina), according to the manufacturer´s instructions except that the provided PCR mastermix was substituted for Kapa HiFi PCR mastermix. In total, 18 libraries were prepared from three biological replicates of primary cell culture of E. multilocularis (Ingrid isolate) treated with anti-miR-71-2´OMe or anti-miR-71-LNA, their respective controls (scrambled or negative control), mock and cells without electroporation. Samples treated with anti-miR-71-partial-2´OMe were not sequenced (see methods). Samples were sequenced with a read length of 75bp and library depth of 2–3 Gb at the Wellcome Trust Sanger Institute, United Kingdom using an Illumina HiSeq v4 sequencer. The RNA-seq data are available in European Nucleotide Archive (ENA) accession number ERP 106379.
The quality of the Illumina raw sequences produced by deep sequencing and the sequences produced after removing the adaptors, were analysed with FastQC and MultiQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc). To remove the adaptors, Trimmomatic tool [31] was used and the parameters used were as follows: LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 MINLEN:45. The processed reads were first mapped to the E. multilocularis genome with HISAT2 [32] with the options—fr—dta. The assemble transcript tool StringTie [32] was used for assembling the reads in transcripts to explain the data and in this case the parameters were: -G -A -C -B -e -M 0.45. A file containing the reference gene models of E. multilocularis was used to guide the genome alignment and the StringTie assembly. This network flow algorithm can reconstruct transcripts more accurately than some previous methods [32,33] because it computes abundance and exon–intron structure at the same time. For differential expression analysis, a gene read-count data matrix was produced with the script prepDE.py provided by StringTie authors.
The DESeq2 Bioconductor package [34], a gene expression analysis tool based on the negative binomial distribution, was used for the exploratory data analysis of the gene-count matrix of the anti-miR-treated and control samples. An initial pre-filtering of the expression matrix was used to remove those genes with less than 10 reads in total, keeping 9,703 genes for the following steps of size factors and dispersion estimations. Normalized and variance-stabilized (using option “blind = TRUE” within DESeq2 Bioconductor package) transformed data was used for a principal component analysis of the top 500 highest-transcribed genes. This analysis shows the samples in the 2D plane spanned by the first two principal components, allowing exploring the effect of experimental covariates, batch effects and helping to visualize sample-to-sample distances. For differential expression, a single factor analysis was performed per anti-miR library, with log2 fold changes and Wald test p values calculated for the comparison anti-miR versus the suitable reference control level. P values were adjusted by FDR correction and shrinkage of log2 fold change estimations were performed following a normal prior distribution [34]. The code and data for differential expression analysis are available at https://github.com/natinreg/PNTD_miR-71.
Bioinformatic prediction of miR-71 target genes in Echinococcus multilocularis
To identify potential targets of miR-71 in E. multilocularis, three hundred nucleotides were extracted downstream from the stop codon of each E. multilocularis coding gene using customs scripts as previously performed for T. solium [19]. The miRanda algorithm [35] was used to perform the prediction of miR-71 target sites using the predicted set of E. multilocularis 3′UTRs. The parameters used were: i) strict seed pairing; ii) score threshold: 140; iii) energy threshold: -17 kcal/mol, iv) gap open penalty: -9; v) gap extend penalty: -4; vi) scaling parameter: 4.
RT-qPCR of selected miR-71 target genes of Echinococcus multilocularis primary cell culture
To validate the differential expression of three selected potential targets of miR-71: serine:threonine protein kinase (EmuJ_000250800), T cell immunomodulatory protein (EmuJ_000440000) and frizzled (EmuJ_000085700) between treated cells with the anti-miR-71-2´OMe and scrambled-2´OMe, RT-qPCR for the same three biological replicates analysed in the RNA-seq was performed (see methods). For this, cDNA synthesis was performed as previously described [5], using 80 ng of total RNA as input. Quantitative PCR (qPCR) was performed in a StepOne Plus cycler (Applied Biosystems, U.S.A.). The PCR mix consisted of 5 x HOT FIREPol EvaGreen qPCR Mix plus 2.4 μl (1x), 0.72 μl (300nM) of each primer, 6.96 μl of DNAse free water and cDNA. The cycling conditions were: 15 min at 95°C, followed by 40 x (15 s at 95°C, 20 s at 60°C) and a final step of 20 s at 72°C. Data collection was performed at 72°C. For normalization, RT-qPCR with the constitutive gene em-elp (EmuJ_000485800) was performed [36]. Primer sequences, amplicon size and PCR efficiency values for the primers are described in S3 Table.
Results
miR-71 is the highest expressed miRNA in Echinococcus multilocularis primary cell cultures
We previously showed that the E. multilocularis miRNA repertoire comprises 39 mature members, all of which are expressed in the metacestode stage [15]. In the present work we determined the miRNA profile in primary cell cultures, freshly established from metacestode vesicles (S1 Table). Correlation analyses between pairs of biological replicates indicated high technical reproducibility and low biological variation (r > 0.95). We found that several miRNAs were highly expressed in primary cell culture. In particular, miR-71, miR-9, miR-10, let-7 and miR-4989 (a member of miR-277 family) represented the top five most abundant miRNAs and accounted for about 70% of total miRNA expression (Fig 2). miR-71 displayed the highest expression level in E. multilocularis primary cell cultures.
Fig 2. Top five most abundant miRNAs in Echinococcus multilocularis primary cell culture at 48 h.

The average proportion of top five most abundant miRNA reads relative to the total number of mature miRNAs is shown.
miR-71 is critical for normal development of Echinococcus multilocularis in vitro
RNA knockdown approaches against mRNAs have previously been carried out in E. multilocularis [27] and were strongly dependent on electroporation of primary cells. In this work, we evaluated the best strategy for miR-71 knockdown in primary cell culture of E. multilocularis using three chemically modified anti-miRs (Fig 1). At 24 h post transfection (48 h cell culture), cells treated with anti-miR-71-2’OMe showed a 78.4% (± 3.4) reduction of miR-71 expression (Fig 3). Likewise, cells treated with anti-miR-71-LNA and anti-miR-71-partial-2´OMe showed a reduction of miR-71 expression (S2 Fig). After 30 days of cultivation, clear phenotypic alterations were observed (Fig 1). In control and mock cultures, parasite cells developed as previously described [7] and first formed cellular aggregates with small, red cavities (S3A Fig) which probably result from an accumulation of phenol red provided by the culture medium. After around one week of incubation, the cellular aggregates enlarged due to germinative cell proliferation [5,7] and most of the cavities lost red staining, most probably due to the formation of a tegument that excludes phenol red. At the end of development, after around 2–3 weeks, mature metacestode vesicles were formed in culture (S3B–S3D Fig). On the other hand, the development in cell cultures transfected with anti-miR-71-2´OMe was blocked at the first stage and displayed clear morphological differences when compared to scrambled and mock controls (Fig 4). Cultures transfected with anti-miR-71-2´OMe consistently contained red cavities which did not increase in size and remained stained throughout incubation. Also, we did not observe the formation of mature metacestode vesicles in these cultures, at least until three weeks of incubation. In transfections performed with the other chemically modified anti-miRs, no change at the phenotype level was observed with respect to controls (S4 Fig). Taking into account that partial 2´OMe was the less effective modified oligonucleotide in miR-71 knockdown and that phenotypic effects were not observed we did not continue the analysis with this modified anti-miR.
Fig 3. Knockdown of Echinococcus multilocularis miR-71 in primary cell culture with 2’-O-methyl oligonucleotides (anti-miR-71-2´OMe).
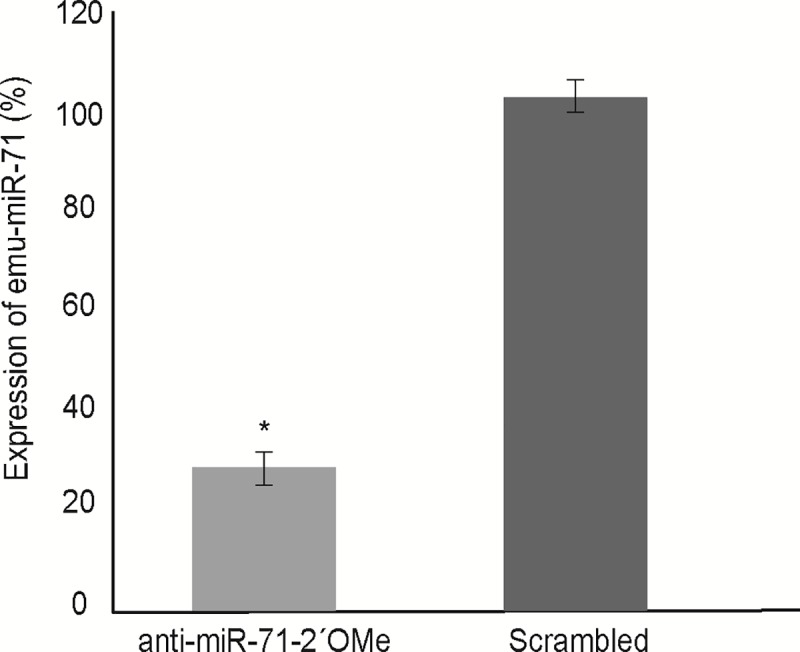
The primary cell culture was electroporated with anti-miR and the respective scrambled control. The effects on the levels of endogenous miR-71 was determined by RT-qPCR at 24 h post-electroporation (48 h culture). Data illustrate representative results with the mean and standard error derived from three technical replicates for each of the three independent biological replicates. The results were calibrated with the average (normalized ct) of the cells treated with the scrambled oligonucleotide. * P ≤ 0.05.
Fig 4. Light microscope images of Echinococcus multilocularis primary cell culture 4 weeks post electroporation.
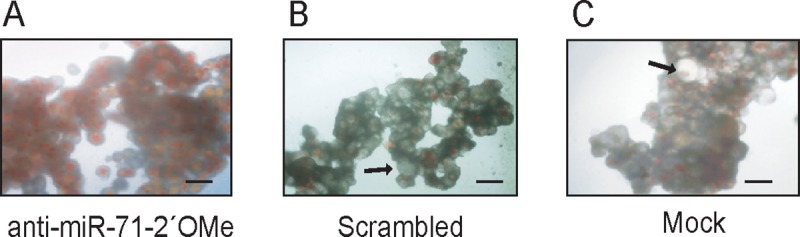
(A) Cells transfected with anti-miR-71-2’OMe by electroporation. The red cavities have been maintained and metacestodes vesicles were not observed. (B) Cells transfected by electroporation with scrambled-2´OMe control in which young cysts (arrow) developed. (C) Mock electroporated cells in which young cysts (arrow) developed. Bars indicate 50 μm.
miR-71 regulates relevant targets at the beginning of the development of Echinococcus multilocularis metacestodes in vitro
According to the canonical role of miRNAs in gene regulation [14], we expected an up-regulation of miR-71 target gene expression upon miR-71 knockdown. We thus explored by RNA-Seq the expression profiles of predicted miR-71 targets. To this end, RNA was analyzed from controls and from miR-71 knockdown samples 24 h after electroporation (Fig 1). We obtained between 11 and 15 million high quality paired-end reads per sample, with 98–99% overall mapping rate to the E. multilocularis genome (S5 Table).
After DESeq2 normalization and variance stabilizing transformation of the gene-count expression matrix, a principal component analysis of all samples was performed (S5 Fig). Firstly, these data indicated a clear effect of electroporation on gene expression since the first component splits the not electroporated samples from all the remaining conditions. Second, the treatment with anti-miR-71-2´OMe was the only knockdown procedure showing a consistent separation of anti-miR-71 and scrambled treated samples as can be seen along the second component (anti-miR-71-2´OMe and scrambled-2´OMe sample). Our results showed that 1,292 genes significantly changed transcription levels (adjusted p-value < 0.1) upon miR-71 knockdown (S6 Table). Of these genes, 744 (57.6%) were significantly up-regulated (log2FC_shrinkage>0, adjusted p-value < 0.1) and 372 of them displayed increased expression of more than 40%. (Fig 5, red points in the right of the volcano plot).
Fig 5. Volcano plot shows the gene statistical significance (on the y-axis) over the log2 fold change after normal shrinkage (on the x-axis) for all the samples from differential expression analysis between anti-miR-71-2´OMe vs scramble-2´OMe control.

Red points correspond to 416 differentially expressed transcripts (372 up-regulated, in the right of the plot) with an adjusted p-value less than 0.1 and an absolute log2 fold change above 0.48.
Interestingly, although miR-71 expression was apparently downregulated upon anti-miR-71 LNA electroporation (see above) when compared to control LNA, we did not see differential gene expression between these samples (S6 Fig) in accordance with the lack of differential phenotype (S4 Fig).
To identify potential miR-71 target genes, we matched it's seed sequence to 3'UTRs of E. multilocularis transcripts and predicted overall 252 candidate target sites (249 genes) (S7 Table). The up-regulated mRNAs and the predicted targets were integrated (Fig 1). After combining the list of up-regulated mRNAs (see above) and the list of candidate target genes, 43 mRNAs fulfilling both criteria were identified (Fig 6 and S8 Table). Finally, we selected from this intersection those genes with shrinkage_FC greater than 1.4 (log2 FoldChange_Shrinkage >0.48) and p adj< 0.05 (16 genes). These genes are represented in Table 1, where 17 target sites are depicted since one of the genes has two miR-71 target sites. Since evolutionary conservation of target sites provides more reliability to target predictions [37] we determined the presence of miR-71 recognition sites in the orthologous 3’UTRs from Echinococcus canadensis, E. granulosus and T. solium for this group of genes. We observed that 13 of the 16 selected genes showed miR-71 conserved target sites in E. granulosus, E. canadensis and/or T. solium orthologs (Table 1), adding confidence to these results. We also determined the energy of miRNA-target interaction [35], target site localization in the 3’UTR [38] and type site since these features are related to target site efficacy [14]. Eight out of the 17 target sites (47%), were located near the stop codon (at least 15 nt from the stop codon) and can thus be considered effective sites [14] (Table 1). The type of regulatory site was also analysed. We observed that all the target sites were canonical sites and belonged to the three site types with higher efficacy according to Bartel [14]: 7mer-A1, 7mer-m8 and 8mer (Table 1 and S9 Table). Among the 16 selected targets (Table 1), we found genes involved in development such as a member of the frizzled-family of GPCRs (EmuJ_000085700) for Wnt ligands that participate in canonical and non-canonical signalling [39,40]; genes related to the establishment of multicellularity such as protocadherin (EmuJ_000848200) [41]; transcription factors (EmuJ_000191100, EmuJ_001186600) [42]; serine:threonine protein kinase encoding genes that could participate in multiple cellular processes (EmuJ_000250800) [43] and genes involved in mRNA stability such as PAB dependent poly(A) specific ribonuclease [44], a protein involved in miRNA function (EmuJ_000610700) [45]. Another interesting target that participates in parasite-host communication is the T-cell immunomodulatory protein (EmTIP) (EmuJ_000440000) [12]. Additionally, three of the miR-71 targets code for proteins without annotation that are tapeworm specific: EmuJ_000085500 (also targeted by miR-281-3p) [18], EmuJ_000865800 and EmuJ_001140200. EmuJ_000865800 codes for a protein with at least one transmembrane domain and the EmuJ_000085500 encoded protein has a coiled-coil and RecF/RecN/SMC N terminal domain (S10 Table). We investigated by RT-qPCR three potential targets that were selected among the 16 genes due to their predicted role in development or host-parasite communication and found indeed increased expression of all three mRNAs, thus verifying the RNA-Seq data (Fig 7).
Fig 6. Venn diagram showing the integration of the target genes of Echinococcus multilocularis predicted by miRanda using genomic information, and genes over-expressed in miR-71 knocked down primary cells (P-value adjustment < 0.1).
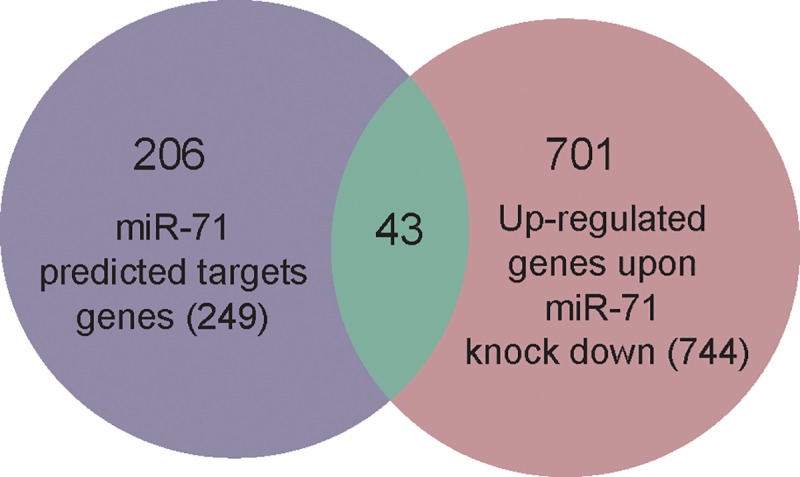
Table 1. Sixteen potential target genes of miR-71 in Echinocuccus multilocularis primary cell culture up-regulated after transfection with anti-miR-71-2´OMe.
(log2FoldChange_Shrinkage > 0.48 and P-value adjustment < 0.05). Position of miR-71 target site in the 3’ UTR of each target gene, conservation of miR-71 targeting in Echinococcus canadensis, Echinococcus granulosus and Taenia solium orthologs and type of site according to Bartel [14] are also shown.
| Gene ID | Annotation | FoldChange | UTR_start | UTR_end | Conservation | Type of site |
|---|---|---|---|---|---|---|
| EmuJ_000085500 | Expressed conserved protein | 1.4 | 14 | 33 | T. solium, E. canadensis and E. granulosus | 7mer-A1 |
| EmuJ_000085700 | Frizzled | 1.7 | 156 | 180 | T. solium and E. granulosus | 7mer-m8 |
| EmuJ_000124300 | CTR1, high affinity copper uptake protein 1 | 1.5 | 27 | 50 | T. solium, E. canadensis and E. granulosus | 8mer |
| EmuJ_000145900 | Glutamate receptor interacting protein 1 | 1.6 | 5 | 26 | T. solium | 8mer |
| EmuJ_000154000 | E3 ubiquitin protein ligase MARCH8 | 1.5 | 33 | 54 | T. solium, E. canadensis and E. granulosus | 7mer-m8 |
| EmuJ_000165500 | USP6 N terminal protein | 1.4 | 23 | 45 | T. solium, E. canadensis and E. granulosus | 8mer |
| EmuJ_000191100 | Transcription factor SOX 14 | 1.6 | 54 | 74 | T. solium, E. canadensis and E. granulosus | 8mer |
| EmuJ_000228700 | Leucine rich repeat, typical subtype | 1.6 | 186 | 206 | T. solium, E. canadensis and E. granulosus | 7mer-m8 |
| EmuJ_000250800 | Serine:threonine protein kinase | 1.4 | 218 | 239 | E. granulosus | 7mer-m8 |
| EmuJ_000440000 | T cell immunomodulatory protein | 1.5 | 26 | 47 | T. solium, E. canadensis and E. granulosus | 8mer |
| EmuJ_000440000 | T cell immunomodulatory protein | 1.5 | 84 | 106 | E. canadensis and E. granulosus | 7mer-m8 |
| EmuJ_000473600 | Sodium dependent phosphate transport protein 1 | 1.4 | 154 | 174 | T. solium and E. granulosus | 7mer-m8 |
| EmuJ_000610700 | PAB dependent poly(A) specific ribonuclease | 1.5 | 25 | 49 | n/c 2 | 8mer |
| EmuJ_000848200 | Protocadherin alpha 7 | 1.7 | 9 | 32 | n/c | 7mer-m8 |
| EmuJ_000865800 | n/a1 | 2.1 | 12 | 33 | n/c | 8mer |
| EmuJ_001140200 | Expressed conserved protein | 2.8 | 169 | 191 | n/c | 8mer |
| EmuJ_001186600 | Ets variant gene 4 | 1.6 | 25 | 46 | T. solium | 7mer-m8 |
1 Without annotation in Wormbase parasite
2 Without conservation in T. solium, E. canadensis and E. granulosus
Fig 7.

Effect of miR-71 suppression in miR-71 level (A), and in the mRNA level of the potential target genes EmuJ_000085700 (frizzled) (B), EmuJ_000440000 (T cell immunomodulatory protein) (C) and EmuJ_000250800 (serine:threonine protein kinase) (D). Primary cells were obtained from Echinococcus multilocularis metacestodes (Ingrid isolate). The cells were electroporated with anti-miR-71-2´OMe or scrambled-2´OMe and their effects on the levels of endogenous miRNA (A) and predicted target mRNAs (B,C,D) determined by RT-qPCR at 24 h post-electroporation (48 h cell culture). Data illustrate representative results with the mean and standard error derived from triplicate experiments. * mean P ≤ 0.05.
Discussion
In this study, the role of miRNAs in the early development of E. multilocularis was addressed by performing miRNA knockdown experiments. To our knowledge, this is the first report of successful miRNA knockdown in tapeworms. miR-71 was selected for knockdown experiments since, among other features, miR-71 displayed high expression levels in E. multilocularis primary cells as well as in the larval stages of several cestodes [19]. High abundance of an individual miRNA is considered functionally important since several miRNA complementary sequences usually compete for actual miRNA target sites [14]. The results of this work suggest that E. multilocularis miR-71 is necessary for proper early development of metacestode vesicles in vitro.
Since there were no previous reports on successful miRNA knockdown in cestodes, we herein assayed different types of chemically modified anti-miRs to optimize protocols for effective functional genomics in these organisms. We found that transfection by electroporation with 2’-O-methyl fully modified oligonucleotide antisense to E. multilocularis miR-71, significantly reduced miR-71 levels. This is probably due to miR-71 degradation since it is known that this type of modified oligonucleotides promote miRNA degradation [46,47]. Notably, cells transfected with this oligonucleotide showed a different phenotype from control oligonucleotide-transfected cells and did not develop metacestode vesicles. Since primary cell culture is enriched in stem cells which are able to produce metacestode vesicles [5], this result suggests that miR-71 regulates early events that are important for parasite development, probably involving stem cells. Furthermore, significant changes in mRNA levels were observed in ~16% of protein coding genes, suggesting that miR-71 knockdown altered the primary cell culture transcriptome. It is interesting to note that antisense oligonucleotides with 2'OMe modification were also already used to successfully alter miRNA expression in S. japonicum [22], suggesting that this type of modification is important for establishing functional genomics methodology in flatworms. The level of target mRNA upregulation observed upon miRNA knockdown was similar to our results, since we observed that most miR-71 mRNA targets were up-regulated between 1.4 and 1.6 times. This modest level of repression is typical for miRNA regulation and has also already been described in other studies [14]. We also attempted knockdown of miR-71 using LNA anti-miR but neither phenotypic alterations nor changes in mRNA levels were observed although miR-71 levels were decreased as measured by RT-qPCR. One possible explanation for these results is that most LNA molecules were retained in endosomes [48] and were not able to interact with cytosolic miR-71. The LNA anti-miR retained in endosomes could have been released after cell lysis to form strong heteroduplex with the miRNA [49], which is characteristic of this type of oligonucleotides and often causes false results in RT-qPCR.
The combined approach employed in this work allowed identifying possible miR-71 target genes. Importantly, the transcriptomic studies performed allowed to determine that miR-71 and the putative target genes are co-expressed in E. multilocularis primary cell culture, which is a necessary condition for target validation. Among these genes, some are presumably involved in parasite development such as a frizzled-family member of GPCRs which presumably acts in the Wnt signaling pathway. This evolutionarily conserved pathway regulates crucial aspects of cell fate determination, cell migration, cell polarity, neural patterning and organogenesis in metazoan embryonic development [50]. E. multilocularis and other flatworms possess genes associated with each of the three known Wnt pathways [51,52], and recently Koziol et al. [53] showed that the Wnt pathway is involved in anterior-posterior specification in E. multilocularis larvae. In concordance with our results, this signalling pathway has recently also been shown to be regulated by miRNAs in the platyhleminth S. japonicum [22]. We also found an ortholog of Serine:threonine protein kinases among the Echinococcus miR-71 targets. Protein kinase function is evolutionarily conserved from Escherichia coli to human and plays a role in a multitude of cellular processes, including division, proliferation, apoptosis, and differentiation [43]. Another miR-71 possible target, bearing 2 recognition sites in its 3’ UTR, is EmTIP. This gene is an ortholog of the human T-cell immunomodulatory protein, TIP [54]. The putative target site for miR-71 in this gene is conserved in E. canadensis and T. solium. This protein was shown to be necessary for E. multilocularis primary stem cell proliferation and metacestode development and it was suggested that it could be associated with cell-cell/extracellular matrix interactions [12]. Also, it stimulates IFN ɣ secretion from human CD4+ T-cells, suggesting that this protein could be important for host-parasite communication. Our results suggest that, by controlling EmTIP expression, miR-71 could be involved in the regulation of establishment of multicellularity and regulation of host immune response. Other interesting possible miR-71 targets are protocadherin, a cell adhesion protein linked with multicellularity establishment [41] and two transcription factors orthologs that play important roles in various biological processes [55]. Interestingly, miR-71 could also target PAB dependent poly(A) specific ribonuclease, a deadenylation protein required for miRNA-mediated silencing in several model organisms [45]. Thus, miR-71 seems to downregulate the expression of a protein necessary for miRNA function, suggesting that it can constitute a negative feedback loop.
Bioinformatically predicted miRNA targets that did not show upregulation upon miRNA knockdown can be false-positive. However, we cannot discard all of them as miR-71 targets since under the conditions assayed, the miRNA recognition site could be bound to other (regulatory) molecules and/or the 3’UTR could adopt a structure that does not allow miRNA binding. In other conditions or life cycle stages these genes might nevertheless be targeted by miR-71. On the other hand, variation in expression levels of genes that were not predicted miR-71 targets can be explained as indirect effects of miR-71 knockdown. Another possibility is that some of these genes are real targets but were not predicted as a consequence of the stringency of the parameters used for target prediction.
Due to their involvement in gene regulation, miRNAs have been identified as high-value targets for therapy. Several miRNA-targeting drugs are now in clinical trials or even close to market launch [56], including miRNA-targeting drugs against viral pathogens [57]. Inhibition of an endogenous miRNA can be achieved by the delivery of antisense oligonucleotides that bind and inhibit its interaction with the mRNA target. This approach has challenges such as the delivery into the cell of oligonucleotides that are poorly cell-permeable, and consequently, delivery strategies such as encapsulation into nanoliposomes are needed [58]. However, miRNA targeting has advantages over small molecule drugs directed against proteins. Since one miRNA can regulate the expression of several protein coding genes, by inhibiting one miRNA, it is theoretically possible to target several proteins. Interestingly, the use of small molecule modulators to target specific miRNAs is being addressed as an alternative strategy of miRNA-based therapy [56]. Parasitic helminth miRNAs have been proposed as potential targets of intervention strategies [59–61]. Furthermore, targeting parasite miRNAs that are protostome-specific, or bilaterian-specific but absent or divergent from host orthologs, could lead to selective drugs. Focusing on E. multilocularis, available drugs, i.e. benzimidazoles, are only parasitostatic probably due to their inability to target the key parasite stem cell population [13]. By inhibiting miR-71, as shown in this study, which is probably highly expressed in parasite stem cells, these problems could possibly be circumvented.
Taken together, the high miR-71 expression in stem-cell enriched primary culture of E. multilocularis, the inhibition of parasite early development by interfering miR-71 expression, the target genes related with parasite development and the high proportion of genes with altered expression after miRNA knockdown, suggest that this miRNA is a master regulator of gene expression in E. multilocularis. Furthermore, miR-71 is absent in the vertebrate host, suggesting that this miRNA is a potential selective drug target. The findings of the present study unveil miR-71 function in Echinococcus development and provide a methodology for miRNA functional analysis in this parasite that could be applied to related tapeworms.
Supporting information
miR-4989 was the gene that kept low stability index according to NormFinder analysis and CP standard deviation ~0,5 with coefficient of correlation ~1 according to BestKeeper analysis. (A) Primary cell culture transfected with anti-miR-71-2’-O-methyl chemical modification in the complete sequence (anti-miR-71-2´OMe), (B) Primary cell culture transfected with anti-miR-71-locked nucleic (anti-miR-71-LNA).
(TIF)
Knockdown of Echinococcus multilocularis miR-71 in primary cell culture with (A) locked nucleic acid (LNA) and (B) 2’-O-methyl oligonucleotides in some nucleotides (Partial-2´OMe). The primary cell culture was electroporated with anti-miR and the respective scrambled control. Their effects on the levels of endogenous miR-71 was determined by RT-qPCR at 24 h post-electroporation. Data illustrate representative results with the mean and standard error derived from triplicate experiments. The results were calibrated with the average (ct) of the mock. * mean P ≤ 0.05
(TIF)
(A) Cells after 10 days of cultivation. Arrow indicates the first red cavities. (B) 15–20 days later the aggregates started to create bigger cavities (arrow). (C) 20–30 days from the first day of cultivation only few rad cavities are observed. (D) Mature metacestode without red staining (arraow) are observed.
(TIFF)
Transfected cells with (A) anti-miR-71-LNA (B) Negative-control-LNA (C) anti-miR-71-partial-2´OMe (D) Scrambled-partial-2´OMe control. Transfected cells by electroporation with the anti-miRs and respective controls showed the development of metacestode vesicles (arrows) for all conditions. Bars indicate 50 μm.
(XLS)
2OME with yellow label (1,2,3): Biological replicates of primary cell culture treated with anti-miR-71-2´OMe. 2OMES with red label (1,2,3): Biological replicates of primary cell culture treated with Scrambled-2´OMe. LNA with pink label (1,2,3): Biological replicates of primary cell culture treated with anti-miR-71-LNA. LNAS with blue label (1,2,3): Biological replicates of primary cell culture treated with Negative-Control-anti-miR-71-LNA. CE with green label (1,2,3): Biological replicates of primary cell culture electroporated (Mock). CSE with light blue label (1,2,3): Biological replicates of primary cell culture without any treatment.
(TIF)
No differential expression is observed (p value less than 0.1).
(TIF)
(XLS)
(XLS)
(XLS)
(XLS)
Raw paired-end reads were 75 bp long. Quality and adapter trimming was performed with Trimmomatic, for a minimum final length of 45 bp. Genome alignment and transcript assembly and quantification was performed with HISAT2 plus StringTie. Expression values are given as transcripts per million (TPM).
(XLS)
(XLS)
(XLSX)
(XLS)
(XLSX)
(XLS)
Acknowledgments
We would like to thank Monika Bergmann from Consultant Laboratory for Echinococcosis, Institute for Hygiene and Microbiology, University of Würzburg, for excellent technical assistance. We would also like to thank Dr Hugo Naya, from Bioinformatics Unit, Institut Pasteur de Montevideo, for receiving MP for bioinformatic analyses.
Data Availability
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ebi.ac.uk/ena/data/view/PRJEB24548.
Funding Statement
This work was supported by ERANET-LAC Project ELAC2015/T080544 (to MR and KB); the Wellcome Trust (https://wellcome.ac.uk/), grant 107475/Z/15/Z (to KB; FUGI), and grant WT 098051; Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), Argentina, Project PICT 2017-2966 (to MR); Consultant Laboratory for Echinococcosis of the Robert Koch Institute; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Projects: PIP 2015 (to MR), CAPGBA-CAPES 070/30 and SNCAD ID 924 (to L.K.). MP is a recipient of a Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Argentina, doctoral fellowship. MC was recipient of Boehringer Ingelheim Fonds Travel Grant and DAAD-UBA fellowship. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Brunetti E, Heller T, Richter J, Kaminstein D, Youkee D, Giordani MT, et al. Application of Ultrasonography in the Diagnosis of Infectious Diseases in Resource-Limited Settings. Curr Infect Dis Rep. 2016. February 18;18(2):6 10.1007/s11908-015-0512-7 [DOI] [PubMed] [Google Scholar]
- 2.Moro P, Schantz PM. Echinococcosis: a review. Int J Infect Dis. 2009. March;13(2):125–33. 10.1016/j.ijid.2008.03.037 [DOI] [PubMed] [Google Scholar]
- 3.Brunetti E, Kern P, Vuitton DA, Writing Panel for the WHO-IWGE. Expert consensus for the diagnosis and treatment of cystic and alveolar echinococcosis in humans. Acta Trop. 2010. April;114(1):1–16. 10.1016/j.actatropica.2009.11.001 [DOI] [PubMed] [Google Scholar]
- 4.Kern P. Clinical features and treatment of alveolar echinococcosis. Curr Opin Infect Dis. 2010. October;23(5):505–12. 10.1097/QCO.0b013e32833d7516 [DOI] [PubMed] [Google Scholar]
- 5.Koziol U, Rauschendorfer T, Zanon Rodríguez L, Krohne G, Brehm K. The unique stem cell sysem of the immortal larva of the human parasite Echinococcus multilocularis. Evodevo. 2014. March 6;5(1):10 10.1186/2041-9139-5-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Eckert J, Deplazes P. Biological, Epidemiological, and Clinical Aspects of Echinococcosis, a Zoonosis of Increasing Concern. Vol. 17, Clinical Microbiology Reviews. 2004. p. 107–35. 10.1128/CMR.17.1.107-135.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Spiliotis M, Lechner S, Tappe D, Scheller C, Krohne G, Brehm K. Transient transfection of Echinococcus multilocularis primary cells and complete in vitro regeneration of metacestode vesicles. Int J Parasitol. 2008. July;38(8–9):1025–39. 10.1016/j.ijpara.2007.11.002 [DOI] [PubMed] [Google Scholar]
- 8.Spiliotis M, Brehm K. Axenic in vitro cultivation of Echinococcus multilocularis metacestode vesicles and the generation of primary cell cultures. Methods Mol Biol. 2009;470:245–62. 10.1007/978-1-59745-204-5_17 [DOI] [PubMed] [Google Scholar]
- 9.Hemer S, Konrad C, Spiliotis M, Koziol U, Schaack D, Förster S, et al. Host insulin stimulates Echinococcus multilocularis insulin signalling pathways and larval development. BMC Biol. 2014. January 27;12(1):5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Förster S, Koziol U, Schäfer T, Duvoisin R, Cailliau K, Vanderstraete M, et al. The role of fibroblast growth factor signalling in Echinococcus multilocularis development and host-parasite interaction. Siles-Lucas M, editor. PLoS Negl Trop Dis. 2019. March 8;13(3):e0006959 10.1371/journal.pntd.0006959 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Koziol U. Evolutionary developmental biology (evo-devo) of cestodes. Exp Parasitol. 2017. September;180:84–100. 10.1016/j.exppara.2016.12.004 [DOI] [PubMed] [Google Scholar]
- 12.Nono JK, Lutz MB, Brehm K. EmTIP, a T-Cell immunomodulatory protein secreted by the tapeworm Echinococcus multilocularis is important for early metacestode development. Dalton JP, editor. PLoS Negl Trop Dis. 2014. January 2;8(1):e2632 10.1371/journal.pntd.0002632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brehm K, Koziol U. On the importance of targeting parasite stem cells in anti-echinococcosis drug development. Parasite. 2014. December 22;21:72 10.1051/parasite/2014070 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bartel DP. Metazoan MicroRNAs. Cell. 2018. March;173(1):20–51. 10.1016/j.cell.2018.03.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cucher M, Macchiaroli N, Kamenetzky L, Maldonado L, Brehm K, Rosenzvit MC. High-throughput characterization of Echinococcus spp. metacestode miRNomes. Int J Parasitol. 2015. March;45(4):253–67. 10.1016/j.ijpara.2014.12.003 [DOI] [PubMed] [Google Scholar]
- 16.Macchiaroli N, Cucher M, Zarowiecki M, Maldonado L, Kamenetzky L, Rosenzvit M. microRNA profiling in the zoonotic parasite Echinococcus canadensis using a high-throughput approach. Parasit Vectors. 2015. February 6;8(1):83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bai Y, Zhang Z, Jin L, Kang H, Zhu Y, Zhang L, et al. Genome-wide sequencing of small RNAs reveals a tissue-specific loss of conserved microRNA families in Echinococcus granulosus. BMC Genomics. 2014. August 29;15(1):736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Macchiaroli N, Maldonado LL, Zarowiecki M, Cucher M, Gismondi MI, Kamenetzky L, et al. Genome-wide identification of microRNA targets in the neglected disease pathogens of the genus Echinococcus. Mol Biochem Parasitol. 2017. June;214:91–100. 10.1016/j.molbiopara.2017.04.001 [DOI] [PubMed] [Google Scholar]
- 19.Pérez MG, Macchiaroli N, Lichtenstein G, Conti G, Asurmendi S, Milone DH, et al. microRNA analysis of Taenia crassiceps cysticerci under praziquantel treatment and genome-wide identification of Taenia solium miRNAs. Int J Parasitol. 2017. September;47(10–11):643–53. 10.1016/j.ijpara.2017.04.002 [DOI] [PubMed] [Google Scholar]
- 20.Pinzón N, Li B, Martinez L, Sergeeva A, Presumey J, Apparailly F, et al. microRNA target prediction programs predict many false positives. Genome Res. 2017. February;27(2):234–45. 10.1101/gr.205146.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rosenzvit M, Cucher M, Kamenetzky L, Macchiaroli N, Prada L, Camicia F. MicroRNAs in Endoparasites In: Johnson James C., editor. MicroRNA and Non-Coding RNA: Technology, Developments and Applications. New York: Nova Science Publishers; 2013. p. 65–92. [Google Scholar]
- 22.Zhu L, Zhao J, Wang J, Hu C, Peng J, Luo R, et al. MicroRNAs Are Involved in the Regulation of Ovary Development in the Pathogenic Blood Fluke Schistosoma japonicum. Collins JJ, editor. PLoS Pathog. 2016. February 12;12(2):e1005423 10.1371/journal.ppat.1005423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Macchiaroli N, Cucher M, Kamenetzky L, Yones C, Bugnon L, Berriman M, et al. Identification and expression profiling of microRNAs in Hymenolepis. Int J Parasitol. 2019. March;49(3–4):211–23. 10.1016/j.ijpara.2018.07.005 [DOI] [PubMed] [Google Scholar]
- 24.Friedländer MR, Mackowiak SD, Li N, Chen W, Rajewsky N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012. January;40(1):37–52. 10.1093/nar/gkr688 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zuker M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003. July 1;31(13):3406–15. 10.1093/nar/gkg595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gruber AR, Lorenz R, Bernhart SH, Neuböck R, Hofacker IL. The Vienna RNA websuite. Nucleic Acids Res. 2008. July 1;36(Web Server issue):W70–4. 10.1093/nar/gkn188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Spiliotis M, Mizukami C, Oku Y, Kiss F, Brehm K, Gottstein B. Echinococcus multilocularis primary cells: Improved isolation, small-scale cultivation and RNA interference. Mol Biochem Parasitol. 2010. November;174(1):83–7. 10.1016/j.molbiopara.2010.07.001 [DOI] [PubMed] [Google Scholar]
- 28.Andersen CL, Jensen JL, Ørntoft TF. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004. August 1;64(15):5245–50. 10.1158/0008-5472.CAN-04-0496 [DOI] [PubMed] [Google Scholar]
- 29.Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol Lett. 2004. March;26(6):509–15. 10.1023/b:bile.0000019559.84305.47 [DOI] [PubMed] [Google Scholar]
- 30.Tsai IJ, Zarowiecki M, Holroyd N, Garciarrubio A, Sánchez-Flores A, Brooks KL, et al. The genomes of four tapeworm species reveal adaptations to parasitism. Nature. 2013. April 4;496(7443):57–63. 10.1038/nature12031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014. August 1;30(15):2114–20. 10.1093/bioinformatics/btu170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pertea M, Kim D, Pertea GM, Leek JT, Salzberg SL. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat Protoc. 2016. August 11;11(9):1650–67. 10.1038/nprot.2016.095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat Protoc. 2012. March 1;7(3):562–78. 10.1038/nprot.2012.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014. December 5;15(12):550 10.1186/s13059-014-0550-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Enright AJ, John B, Gaul U, Tuschl T, Sander C, Marks DS. MicroRNA targets in Drosophila. Genome Biol. 2003;5(1):R1 10.1186/gb-2003-5-1-r1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brehm K, Wolf M, Beland H, Kroner A, Frosch M. Analysis of differential gene expression in Echinococcus multilocularis larval stages by means of spliced leader differential display. Int J Parasitol. 2003. September 30;33(11):1145–59. 10.1016/s0020-7519(03)00169-3 [DOI] [PubMed] [Google Scholar]
- 37.Bartel DP. MicroRNA Target Recognition and Regulatory Functions. Cell. 2009. January 23;136(2):215 10.1016/j.cell.2009.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet. 2008. February;9(2):102–14. 10.1038/nrg2290 [DOI] [PubMed] [Google Scholar]
- 39.MacDonald BT, He X. Frizzled and LRP5/6 Receptors for Wnt/ -Catenin Signaling. Cold Spring Harb Perspect Biol. 2012. December 1;4(12):a007880–a007880. 10.1101/cshperspect.a007880 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.van Amerongen R, Nawijn MC, Lambooij J-P, Proost N, Jonkers J, Berns A. Frat oncoproteins act at the crossroad of canonical and noncanonical Wnt-signaling pathways. Oncogene. 2010. January 5;29(1):93–104. 10.1038/onc.2009.310 [DOI] [PubMed] [Google Scholar]
- 41.Maître J-L, Heisenberg C-P. Three Functions of Cadherins in Cell Adhesion. Curr Biol. 2013. July 22;23(14):R626–33. 10.1016/j.cub.2013.06.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zeitlinger J, Stark A. Developmental gene regulation in the era of genomics. Dev Biol. 2010. March 15;339(2):230–9. 10.1016/j.ydbio.2009.12.039 [DOI] [PubMed] [Google Scholar]
- 43.Manning G, Plowman GD, Hunter T, Sudarsanam S. Evolution of protein kinase signaling from yeast to man. Trends Biochem Sci. 2002. October;27(10):514–20. 10.1016/s0968-0004(02)02179-5 [DOI] [PubMed] [Google Scholar]
- 44.Parker R, Song H. The enzymes and control of eukaryotic mRNA turnover. Nat Struct Mol Biol. 2004. February;11(2):121–7. 10.1038/nsmb724 [DOI] [PubMed] [Google Scholar]
- 45.Flamand MN, Wu E, Vashisht A, Jannot G, Keiper BD, Simard MJ, et al. Poly(A)-binding proteins are required for microRNA-mediated silencing and to promote target deadenylation in C. elegans. Nucleic Acids Res. 2016. July 8;44(12):5924–35. 10.1093/nar/gkw276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Krützfeldt J, Rajewsky N, Braich R, Rajeev KG, Tuschl T, Manoharan M, et al. Silencing of microRNAs in vivo with “antagomirs.” Nature. 2005. December 30;438(7068):685–9. 10.1038/nature04303 [DOI] [PubMed] [Google Scholar]
- 47.Torres AG, Fabani MM, Vigorito E, Gait MJ. MicroRNA fate upon targeting with anti-miRNA oligonucleotides as revealed by an improved Northern-blot-based method for miRNA detection. RNA. 2011. May 1;17(5):933–43. 10.1261/rna.2533811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Buntz A, Killian T, Schmid D, Seul H, Brinkmann U, Ravn J, et al. Quantitative fluorescence imaging determines the absolute number of locked nucleic acid oligonucleotides needed for suppression of target gene expression. Nucleic Acids Res. 2019. January 25;47(2):953–69. 10.1093/nar/gky1158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stenvang J, Petri A, Lindow M, Obad S, Kauppinen S. Inhibition of microRNA function by antimiR oligonucleotides. Silence. 2012. January 9;3(1):1 10.1186/1758-907X-3-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Komiya Y, Habas R. Wnt signal transduction pathways. Organogenesis. 2008. April;4(2):68–75. 10.4161/org.4.2.5851 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Olson PD, Zarowiecki M, Kiss F, Brehm K. Cestode genomics—progress and prospects for advancing basic and applied aspects of flatworm biology. Parasite Immunol. 2012. February;34(2–3):130–50. 10.1111/j.1365-3024.2011.01319.x [DOI] [PubMed] [Google Scholar]
- 52.Riddiford N, Olson PD. Wnt gene loss in flatworms. Dev Genes Evol. 2011. October 3;221(4):187–97. 10.1007/s00427-011-0370-8 [DOI] [PubMed] [Google Scholar]
- 53.Koziol U, Jarero F, Olson PD, Brehm K. Comparative analysis of Wnt expression identifies a highly conserved developmental transition in flatworms. BMC Biol. 2016. December 4;14(1):10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fiscella M, Perry JW, Teng B, Bloom M, Zhang C, Leung K, et al. TIP, a T-cell factor identified using high-throughput screening increases survival in a graft-versus-host disease model. Nat Biotechnol. 2003. March 24;21(3):302–7. 10.1038/nbt797 [DOI] [PubMed] [Google Scholar]
- 55.Shaikhibrahim Z, Wernert N. ETS transcription factors and prostate cancer: The role of the family prototype ETS-1 (Review). Int J Oncol. 2012. February 21;40(6):1748–54. 10.3892/ijo.2012.1380 [DOI] [PubMed] [Google Scholar]
- 56.Schmidt MF. Drug target miRNAs: chances and challenges. Trends Biotechnol. 2014. November;32(11):578–85. 10.1016/j.tibtech.2014.09.002 [DOI] [PubMed] [Google Scholar]
- 57.van der Ree MH, de Vree JM, Stelma F, Willemse S, van der Valk M, Rietdijk S, et al. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: a phase 1B, double-blind, randomised controlled trial. Lancet. 2017. February;389(10070):709–17. 10.1016/S0140-6736(16)31715-9 [DOI] [PubMed] [Google Scholar]
- 58.Broderick JA, Zamore PD. MicroRNA therapeutics. Gene Ther. 2011. December 28;18(12):1104–10. 10.1038/gt.2011.50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Britton C, Winter AD, Gillan V, Devaney E. microRNAs of parasitic helminths–Identification, characterization and potential as drug targets. Int J Parasitol Drugs Drug Resist. 2014. August;4(2):85–94. 10.1016/j.ijpddr.2014.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Manzano-Román R, Siles-Lucas M. MicroRNAs in parasitic diseases: potential for diagnosis and targeting. Mol Biochem Parasitol. 2012. December;186(2):81–6. 10.1016/j.molbiopara.2012.10.001 [DOI] [PubMed] [Google Scholar]
- 61.Cai P, Gobert GN, McManus DP. MicroRNAs in Parasitic Helminthiases: Current Status and Future Perspectives. Trends Parasitol. 2016. January;32(1):71–86. 10.1016/j.pt.2015.09.003 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
miR-4989 was the gene that kept low stability index according to NormFinder analysis and CP standard deviation ~0,5 with coefficient of correlation ~1 according to BestKeeper analysis. (A) Primary cell culture transfected with anti-miR-71-2’-O-methyl chemical modification in the complete sequence (anti-miR-71-2´OMe), (B) Primary cell culture transfected with anti-miR-71-locked nucleic (anti-miR-71-LNA).
(TIF)
Knockdown of Echinococcus multilocularis miR-71 in primary cell culture with (A) locked nucleic acid (LNA) and (B) 2’-O-methyl oligonucleotides in some nucleotides (Partial-2´OMe). The primary cell culture was electroporated with anti-miR and the respective scrambled control. Their effects on the levels of endogenous miR-71 was determined by RT-qPCR at 24 h post-electroporation. Data illustrate representative results with the mean and standard error derived from triplicate experiments. The results were calibrated with the average (ct) of the mock. * mean P ≤ 0.05
(TIF)
(A) Cells after 10 days of cultivation. Arrow indicates the first red cavities. (B) 15–20 days later the aggregates started to create bigger cavities (arrow). (C) 20–30 days from the first day of cultivation only few rad cavities are observed. (D) Mature metacestode without red staining (arraow) are observed.
(TIFF)
Transfected cells with (A) anti-miR-71-LNA (B) Negative-control-LNA (C) anti-miR-71-partial-2´OMe (D) Scrambled-partial-2´OMe control. Transfected cells by electroporation with the anti-miRs and respective controls showed the development of metacestode vesicles (arrows) for all conditions. Bars indicate 50 μm.
(XLS)
2OME with yellow label (1,2,3): Biological replicates of primary cell culture treated with anti-miR-71-2´OMe. 2OMES with red label (1,2,3): Biological replicates of primary cell culture treated with Scrambled-2´OMe. LNA with pink label (1,2,3): Biological replicates of primary cell culture treated with anti-miR-71-LNA. LNAS with blue label (1,2,3): Biological replicates of primary cell culture treated with Negative-Control-anti-miR-71-LNA. CE with green label (1,2,3): Biological replicates of primary cell culture electroporated (Mock). CSE with light blue label (1,2,3): Biological replicates of primary cell culture without any treatment.
(TIF)
No differential expression is observed (p value less than 0.1).
(TIF)
(XLS)
(XLS)
(XLS)
(XLS)
Raw paired-end reads were 75 bp long. Quality and adapter trimming was performed with Trimmomatic, for a minimum final length of 45 bp. Genome alignment and transcript assembly and quantification was performed with HISAT2 plus StringTie. Expression values are given as transcripts per million (TPM).
(XLS)
(XLS)
(XLSX)
(XLS)
(XLSX)
(XLS)
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ebi.ac.uk/ena/data/view/PRJEB24548.