

Summary
Differentiation of human embryonic stem cells into pancreatic β cells holds great promise for the treatment of diabetes. Recent advances have led to the production of glucose-responsive insulin-secreting cells in vitro, but resulting cells remain less mature than their adult primary β cell counterparts. The barrier(s) to in vitro β cell maturation are unclear. Here, we evaluated a potential role for microRNAs. MicroRNA profiling showed high expression of let-7 family microRNAs in vivo, but not in in vitro differentiated β cells. Reduced levels of let-7 in vitro were associated with increased levels of the RNA binding protein LIN28B, a negative regulator of let-7 biogenesis. Ablation of LIN28B during human embryonic stem cell (hESC) differentiation toward β cells led to a more mature glucose-stimulated insulin secretion profile and the suppression of juvenile-specific genes. However, let-7 overexpression had little effect. These results uncover LIN28B as a modulator of β cell maturation in vitro.
Keywords: hESC-derived human pancreatic beta cells, LIN28, let-7, microRNAs, beta cell maturation
Graphical Abstract
Highlights
-
•
Let-7 is low in hESC-derived relative to in vivo matured human β cells
-
•
LIN28 is elevated in hESC-derived relative to in vivo matured human islet β cells
-
•
Depletion of LIN28, but not gain of let-7, promotes hESC-derived β cell maturation
-
•
Depletion of LIN28 promotes silencing of juvenile-specific transcriptional program
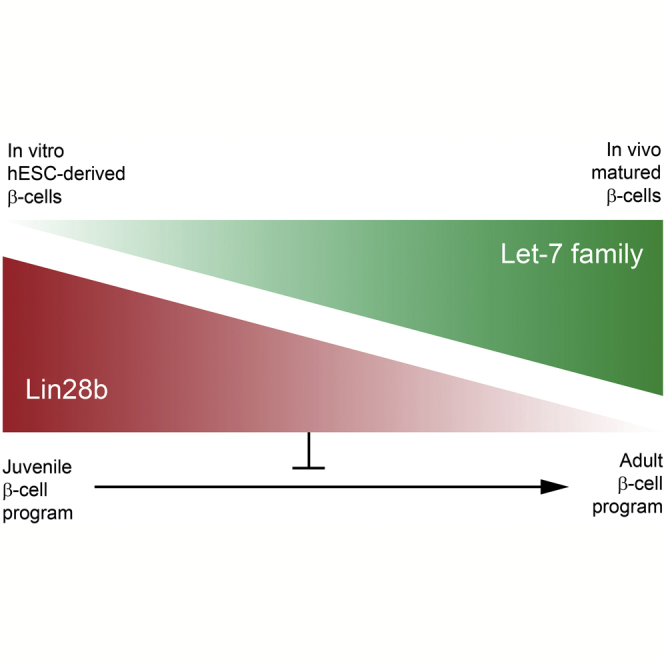
In this article, Blelloch, Hebrok, and colleagues show that in vitro hESC-derived islet β cells express elevated LIN28 and reduced let-7 levels relative to their in vivo matured counterparts. Reduction of LIN28, but not increase in let-7, promotes suppression of a juvenile-specific β cell transcriptional program, implicating LIN28 as a barrier to β cell maturation in vitro.
Introduction
A growing number of people are suffering from diabetes worldwide (Roglic and World Health Organization, 2016). Diabetes is a disease of imbalance between blood insulin and glucose levels secondary to pancreatic islet β cell loss or impaired function (Cerf, 2013). At present, type 1 diabetic (T1D) and end-stage type 2 diabetic (T2D) patients rely on exogenous injection of insulin to control blood glucose. While life sustaining, this therapy is arduous and prone to complications as it is virtually impossible to mimic the dynamic changes in insulin production and secretion performed by endogenous β cells. Transplantation of cadaveric islet cells provides an alternative option resulting in effective glycemic control, but these cells are in limited supply making it unfeasible for broad implementation (Sneddon et al., 2018). β cells produced by the differentiation of pluripotent stem cells, both human embryonic stem cells (hESCs) and induced pluripotent stem cells (iPSCs) hold great promise in filling this gap. Recent advancements have greatly improved the production of these cells in vitro (Nair et al., 2019, Velazco-Cruz et al., 2019, Veres et al., 2019). However, there remain differences between in vitro produced cells and endogenous adult β cells in their gene expression profile and secretory capacity. Therefore, it is important both conceptually and practically to understand the barriers to in vitro differentiation toward mature adult β cells. Since euglycemia can be restored in diabetic mice by transplantation of stem cell-derived pancreatic progenitors or β cell populations, it is speculated that the in vivo environment supports further maturation of generated β cells, although the changes that occur in β cells upon transplantation have not been elucidated.
Much of the progress in β cell differentiation has been achieved by optimizing combinations of signaling peptides and chemicals that recapitulate events that occur during normal development in vivo (Liew, 2010, Nair and Hebrok, 2015). MicroRNAs (miRNAs) represent another type of small molecule. They exist endogenously, function by coordinating the regulation of many targets, and can have profound effects on developmental cell fate decisions (Friedman et al., 2009, Shenoy and Blelloch, 2014). The let-7 family comprises one of the evolutionarily most conserved families of miRNAs (Friedman et al., 2009). Let-7 exists in a negative feedback loop with the RNA binding proteins LIN28A and LIN28B (Shyh-Chang and Daley, 2013). Let-7 inhibits production of the LIN28 proteins, while the LIN28 proteins suppress biogenesis of Let-7. This loop forms a bistable regulatory switch in a number of cell fate decisions (Thornton and Gregory, 2012). Of note, both let-7 and LIN28 have many other targets. Let-7 miRNAs act through their many targets to generally promote differentiation and suppress growth (Kumar et al., 2008, Roush and Slack, 2008), whereas LIN28 has the opposite effect both by inhibiting let-7 and through let-7 independent mechanisms, such as increasing translation of cell-cycle mRNAs (Tsialikas and Romer-Seibert, 2015). Here, we report an increase in let-7 and decrease in LIN28B during β cell maturation. The manipulation of LIN28B, but not let-7 levels, promoted a switch to a more mature adult-like β cell phenotype in vitro, uncovering a let-7 independent role for LIN28B in inhibiting β cell maturation.
Results
Let-7 Expression Increases with β Cell Maturation
To determine a potential role for miRNAs in the maturation of pancreatic β cells, we performed small RNA sequencing (RNA-seq) of in vitro stem cell-derived, in vivo matured, and human cadaveric islet cells. Human in vitro derived β-like cells were produced from hESCs using an INS-GFP reporter hESC line (Micallef et al., 2012), where GFP expression is under the control of the endogenous insulin promoter (Figure 1A, hESC immature β-like cells) (Faleo et al., 2017, Russ et al., 2015). Typically, 39.26% ± 4.09% INS-GFP+ cells were generated (Figures S1A and S1B). The β-like cells were also transplanted under the kidney capsule of immunodeficient mice to allow for further in vivo maturation for 4–5 weeks (referred to as in vivo matured hESC β cells). As the differentiation protocol produces a heterogeneous mixture of cells, the insulin-producing cells in both in vitro derived cultures and in vivo matured grafts were isolated by their GFP expression using fluorescence-activated cell sorting before transcriptome analysis. Cadaveric human islets were used as a proxy for pancreas-derived human β cells, although these islets contain a mix of cell types (approximately 50% β cells) (Cabrera et al., 2006).
Figure 1.

Let-7 Is Upregulated at Late-Stage β Cell Maturation
(A) Schematic outlining the differentiation protocol employed. In vivo matured β cells: β cells isolated from grafts post transplant. eBCs, enriched β clusters generated after inducing further maturation in vitro. Adapted from Nair et al. (2019).
(B) Volcano plot of differentially expressed miRNAs in hESC β-like cells (n = 3, independent samples) and human islets (n = 3, independent samples). Significant hits are shown in blue (p < 0.05). let-7 family hits are shown in red.
(C) Volcano plot of differentially expressed miRNAs in hESC β-like cells (n = 3, independent samples) and in vivo matured β cells (n = 3, independent samples). Significant hits are shown in blue (p < 0.05). let-7 family hits are shown in red.
(D) qRT-PCR verification of representative let-7 family member expression in human islets (n = 3, independent samples), hESC β-like cells (n = 4, independent samples), hESC eBCs (n = 3, independent samples), and in vivo matured β cells (n = 2, independent samples). Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
Analysis of small RNA-seq data for human islet cells versus in vitro derived hESC β-like cells uncovered 554 significantly differentially expressed miRNAs (adjusted p value < 0.05) (Figure 1B; Table S1). Of note, this contained a large number of let-7 family members that were highly upregulated in the islet cells relative to the in vitro derived cells (Figure 1B, red dots). The increase in let-7 could have been contributed by non-β cells within the cadaveric human islets. Therefore, we next analyzed small RNA-seq data from in vivo matured INS-GFP+ hESC β cells versus INS-GFP+ hESC β-like cells (Figures 1C; Table S2). This comparison of isogenic purified INS-GFP+ cell populations controls for genetic variation and cellular heterogeneity. Still, similar to the human islets, the in vivo matured β cells showed elevated expression of multiple let-7 family members relative to the β-like cells (Figure 1C, red dots).
Next, we validated the association between let-7 expression and β cell maturation using a differentiation protocol that allows for further maturation of β-like cells in vitro, by reaggregation and culture of purified insulin-expressing cells as organoids (Nair et al., 2019). We call the resulting in vitro matured cells as hESC-enriched β cell clusters (eBCs) (Figure 1A). We performed qRT-PCR for representative let-7 family members in hESC β-like cells, in vitro matured hESC eBCs, in vivo matured hESC β cells, and human islets (Figure 1D). Consistent with the sequencing data, both human islet cells and in vivo matured hESC β cells showed dramatically increased levels of all the let-7 family members. hESC eBCs also showed elevated levels of let-7 relative to hESC β-like cells, but the levels were below human islets and in vivo matured hESC β cells (Figure 1D). Together, these data show a positive association between let-7 levels and the maturation of β cells.
LIN28B Expression Is Downregulated during β Cell Maturation
Let-7 expression is regulated both transcriptionally and post-transcriptionally (Roush and Slack, 2008). Post-transcriptionally, the RNA binding proteins LIN28A&B suppress the biogenesis of mature let-7 family members (Nam et al., 2011, Piskounova et al., 2011, Viswanathan et al., 2008). In turn, let-7 itself functions to suppress hundreds of downstream mRNA targets including LIN28A&B (Rybak et al., 2008, Yang et al., 2010). To evaluate the impact of β cell maturation on the let-7 regulatory network of genes, we performed mRNA-seq in hESC β-like cells, in vivo matured hESC β cells, and human islet cells. Differential expression between human islets and hESC β-like cells showed differential expression of many transcripts (Figure 2A, blue dots, adj p < 0.05, Table S3), including elevated levels of LIN28B (but not LIN28A) in the in vitro cells (LIN28B, green dot, adj p = 4 × 10−5). Surprisingly, however, analysis of high-scoring Targetscan predicted targets of let-7 showed roughly equal distribution between up- and downregulated genes (Figure 2B, red dots). Furthermore, cumulative density analysis on the fold change of let-7 targets versus all genes, showed no shift in the curve (Figure 2C). We hypothesized this may be due to a confounding effect due to presence of other islet cell types besides β cells in human islets. Therefore, we next compared the GFP-sorted populations from hESC β-like cells and in vivo matured hESC β cells (Figure 2D, Table S4). Fewer genes were differentially expressed between these cell types, consistent with the common origin, and hence reduced heterogeneity between the two cell populations (Figure 2D, blue dots, adj p < 0.05). Again, LIN28B (but not LIN28A) was up in hESC β-like cells (LIN28B, green dot, adj p = 0.044). Also, predicted targets of let-7 were distributed equally among up- and downregulated genes (Figure 2E, red dots) and were not shifted in the cumulative density plot (Figure 2F). We validated the LIN28B findings by qRT-PCR and extended them to the hESC eBCs (Figure 2G). There was a progressive reduction in LIN28 levels when starting with hESC β-like cells, followed by hESC eBCs, in vivo matured hESC β cells, and finally human islet cells. The negative correlation between let-7 levels and LIN28B levels is consistent with their known negative feedback on each other (Rybak et al., 2008, Thornton and Gregory, 2012). However, the lack of enrichment of let-7 targets among the upregulated genes in hESC β-like cells suggests that let-7 downstream function may not play a major role in β cell maturation.
Figure 2.

LIN28 Is Downregulated at Late-Stage β Cell Maturation
(A) Volcano plot of differentially expressed mRNAs in hESC β-like cells (n = 3, independent samples) and human islets (n = 3, independent samples). Significant hits are shown in blue (p < 0.05). LIN28A and LIN28B are highlighted in yellow and green, respectively.
(B) Same as (A), except let-7 family target genes (predicted by TargetScan) are highlighted in red.
(C) Cumulative distribution of differential expression of all expressed mRNAs and let-7 targets, predicted by TargetScan, in the human islets versus hESC β-like cells from (A).
(D) Volcano plot of differentially expressed mRNAs in hESC β-like cells (n = 3, independent samples) and in vivo matured β cells (n = 3, independent samples). Significant hits are shown in blue (p < 0.05). LIN28A and LIN28B are highlighted in yellow and green, respectively.
(E) Same as (D), except let-7 family target genes (predicted by TargetScan) are highlighted in red.
(F) Cumulative distribution of differential expression of all expressed mRNAs and let-7 targets, predicted by TargetScan, in the in vivo matured β cells versus hESC β-like cells from (D).
(G) qRT-PCR verification of LIN28B expression in human islets (n = 3, independent samples), hESC β-like cells (n = 4, independent samples), hESC eBCs (n = 3, independent samples), and in vivo matured β cells (n = 3, independent samples). Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
LIN28B Downregulation Promotes hESC β Cell Maturation
As LIN28B was up and let-7 was down in the in vitro derived cells (both β-like cells and eBCs) relative to in vivo matured hESC β cells and human islet cells, we asked if suppression of LIN28B could promote further maturation of β cells. To suppress LIN28B, we initially attempted knocking down LIN28 using a virally transduced shRNA against the Lin28b mRNA. However, transduction even of a control vector led to poor differentiation (data not shown). Therefore, we implemented a doxycycline-inducible CRISPR knockout strategy with guide RNAs on either side of exon 3 (iCrLIN28B) (Gonzalez et al., 2014) (Figure 3A, also see Experimental Procedures). Both gRNAs are constitutively expressed under the control of U6 promoters, while Cas9 protein expression depends on doxycycline exposure. Doxycycline was introduced at different times during the differentiation process and the cells were treated until the end of the differentiation protocol (Figure S2A). Addition of doxycycline at day 0 resulted in 100% indel formation, but caused poor differentiation of INS-GFP+ cells (Figures S2B and S2C). In contrast, addition at day 3 resulted in 78% indel formation with only a marginal decrease in the number of INS-GFP+ cells at day 20 of differentiation (Figure S2D). Addition of doxycycline at later time points including days 6, 8, 10, and 14 resulted in lower levels of indel formation (Figure S2B), indicating a relative resistance to the inducible CRISPR knockout at later stages of differentiation. Furthermore, cells assessed at the β-like and eBC stages that were treated with doxycyline from day 3, consistently showed 70%–80% indel formation (Figure S2E). This result was corroborated with western blot analysis showing a 50% reduction in LIN28B protein levels at β-like stage of differentiation (Figure 3B). Because addition of doxycycline starting at day 3 resulted in the greatest loss of LIN28B while retaining near normal INS-GFP+ cell numbers, further experiments were performed using this treatment regime.
Figure 3.

LIN28B Downregulation Promotes hESC-β Cell Maturation
(A) Generation of iCrLIN28B. SA, splice acceptor; 2A, self-cleaving 2A peptide; Puro, puromycin resistance gene; TRE, tetracycline response element; Cas9, Cas9 protein; U6, U6 promoter; Neo, neomycin resistance gene; CAG, constitutive synthetic promoter; M2rtTA, reverse tetracycline trans-activator sequence and protein. gRNA1 and gRNA2 were designed to target in LIN28B exon 3.
(B) Western blot analysis of LIN28B in d20 (hESC β-like) clusters generated from iCrLIN28B line plus/minus Cas9 induction with doxycycline during differentiation. Dox–, no doxycycline treatment. Dox+, doxycycline treatment from d3 to d20. Left panel: representative western blot. Right panel: quantification of four independent western blots (n = 4, independent samples, two LIN28 isoforms relative to GAPDH). Values are average ± SEM. Statistical significance was calculated using paired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
(C) qRT-PCR analysis of representative let-7 family members in iCrLIN28B hESC eBCs plus/minus induction of Cas9 as in (B). n = 5 independent samples for Dox–, n = 7 independent samples for Dox+. Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
(D) qRT-PCR analysis of selected gene expression in hESC eBCs plus/minus induction of Cas9 as in (B). Number of samples and statistics as in (C).
(E) Static glucose-stimulated insulin secretion (GSIS) of iCrLIN28B eBCs. Doxycycline treatment as in (B). n = 17 independent samples for Dox–, n = 22 independent samples, for Dox+. Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
(F) Dynamic GSIS of iCrLIN28B hESC eBCs in perifusion assays. Dox treatments as in (B). n = 4 independent samples for Dox–, n = 4 independent samples for Dox+. Values are average ± SEM. Area under the curve (AUC) was calculated for the secretion profiles. Error bars represent the standard error. Statistical significance was calculated using unpaired two-tailed t test.
(G) GSEA of differentially expressed genes between Dox+ versus Dox– iCrLIN28B hESC eBCs on previously identified gene set found upregulated in juvenile versus adult primary human β cells (Arda et al., 2016).
See also Figures S2, S3, and S4 and Tables S5 and S6.
In particular, we evaluated the effect of iCrLIN28B on let-7 production and β cell maturation on hESC eBCs. iCrLIN28B led to a small, albeit significant, increase in let-7 levels as determined by qRT-PCR for four representative let-7 family members (Figure 3C). To determine the impact of LIN28B depletion on β cell maturation, we first analyzed the expression of key β cell markers. qRT-PCR showed enhanced expression of a number of markers of β cells including PDX1, NKX6.1, NKX2.2, MAFA, NEUROD1, and ISL1 (Figure 3D). PDX1, NKX6.1, NKX2.2, and NEUROD1 are important in the maintenance of β cell identity and function (Chao et al., 2007, Gao et al., 2014, Gu et al., 2010, Schaffer et al., 2013). MAFA regulates insulin expression and promotes the functional maturation of β cells (Artner et al., 2010, Hang and Stein, 2011). ISL1 maintains the terminal differentiation program of β cells (Ediger et al., 2017). We also conducted a protein level characterization of these β cell markers by flow cytometry (Figure S3). Percentage of cells expressing PDX1, NKX6.1, PAX6, and ISL1 trended higher in LIN28B-deleted cells than controls, although the statistical tests did not reach a p value of 0.05.
Next, we tested the impact of LIN28 depletion on β cell function using both static and dynamic glucose stimulation insulin secretion (GSIS) studies. In the static GSIS assay, the fold increase in insulin release into the media was measured following an increase in the glucose concentration from 2.8 to 16.7 mM (stimulation index). The assay showed a significant, roughly 2-fold greater increase in the stimulation index in the doxycycline-treated versus untreated iCrLIN28B eBCs (Figure 3E; Table S5). For the dynamic assay, iCrLIN28B doxycycline-treated or untreated eBCs were perfused with low- and high-glucose buffers and insulin release was measured over time. In contrast to the static assay, the dynamic perifusion assay provides a comprehensive view of β cell function including the basal, first, and second phases of insulin secretion. The perifusion assay showed higher levels of insulin secretion in both the low- and high-glucose treatments for the doxycycline-treated iCrLIN28B eBCs relative to their untreated counterparts (Figure 3F). These perifusion results are highly reminiscent of differences in the secretion profiles previously reported for adult versus juvenile primary human islet samples (Arda et al., 2016).
The Arda et al. study also measured gene expression differences in adult versus juvenile primary human islet samples by RNA-seq. We therefore performed a similar RNA-seq comparison between our doxycycline-treated and untreated iCrLIN28B hESC eBCs (Figure S4A; Table S6). To compare the gene expression changes in our experiments with theirs we performed gene set enrichment analysis (GSEA) for genes upregulated in either adult or juvenile primary human islets. Although there was no enrichment for adult upregulated genes in our doxycycline-treated samples there was a highly significant depletion of the juvenile upregulated genes (Figures 3G and S4B). Together, these data show that the reduction of LIN28B during differentiation of hESCs to β cells in vitro promotes their maturation with a switch from a more juvenile to a more adult-like primary human β cell phenotype.
Let-7 Upregulation Alone Is Insufficient to Drive hESC-β Cell Maturation
Next we asked whether let-7 is acting downstream of LIN28B depletion. Induction of iCrLIN28 led to a small increase in let-7 levels (Figure 3C). Analysis of the differential expression between uninduced and induced iCrLIN28 showed no enrichment of let-7 targets among the downregulated set of genes (Figures S4C and S4D). To directly test the impact of let-7 on β cell maturation, we generated a cell line where a doxycycline-inducible let-7a/f/b transgene was targeted to the hAAVS1 locus (iLET-7) (Figure 4A). Doxycycline was added from day 14 to day 27 to induce let-7 at the later stages of differentiation (Figure 4B). Doxycycline treatment did not affect the percent of INS-GFP+ cells measured at day 20 (Figure S4E). qRT-PCR showed a 2- to 6-fold increase in let-7 in doxycycline-treated cells relative to no doxycycline controls, significantly higher than seen in the iCrLIN28B cells (Figure 4C, compare with 3C). These levels did not reduce LIN28B expression, but did suppress another well-known let-7 target, HMGA2 (Figure 4D). Static GSIS assays on the resulting day 27 eBCs did not show an improvement in the let-7-induced cells relative to uninduced controls, unlike Lin28B-deleted cells (Figure 4E, compare with Figures 3E; Table S5). Transcriptional markers of maturation were also mostly unchanged (Figure 4F). These data suggest that, while LIN28 acts a barrier to β cell maturation, it is likely acting independent of its role as an inhibitor of let-7 biogenesis.
Figure 4.

Let-7 Upregulation Alone Is Insufficient to Drive hESC-β Cell Maturation
(A) Generation of iLET-7 cell line. Pre-let-7a/f (m) and pre-let-7b (m) are genomic sequences for let-7a/f and let-7b precursors with LIN28 binding region mutated (see Experimental Procedures). OE, overexpression.
(B) Schematic outlining let-7 induction protocol. Doxycycline treatment was from d14 to d27. Fluorescence-activated cell sorting was performed on d20. GSIS and qRT-PCR were performed on d27.
(C) qRT-PCR analysis of representative let-7 family members in iLET-7 hESC eBCs. Doxycycline treatment as in (B). n = 6 independent samples for Dox–, n = 5 independent samples for Dox+. Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001; n.s., not significant.
(D) qRT-PCR analysis of LIN28B and HMGA2 expression in iLET-7 hESC eBCs. Doxycycline treatment, number of replicates and statistics as in (C).
(E) Static GSIS of iLET-7. Doxycycline treatment as in (B). n = 11 independent samples for Dox–, n = 10 independent samples for Dox+. Values are average ± SEM. Statistical significance was calculated using unpaired two-tailed t test. n.s., not significant. Compare with iCrLIN28B in Figure S4E.
(F) qRT-PCR analysis of selected gene expression in iLET-7 hESC eBCs. Dox treatment, number of replicates, and statistics as in (C).
See also Figure S4.
Discussion
Our findings uncover an important function for LIN28B during the course of hESC β cell maturation. The levels of let-7 and LIN28B correlate with the maturation status of the β cells. Let-7 family miRNAs were upregulated, while LIN28B was downregulated as the cells matured; human islets contain the highest levels of let-7 and lowest levels of LIN28B, followed by in vivo matured transplanted β cells, in vitro matured eBCs, and finally β-like cells. Furthermore, using an inducible CRISPR-Cas9 system we found that the deletion of LIN28B during the course of differentiation to eBCs improved GSIS. Interestingly, the change in the GSIS profile is reminiscent of the change previously reported when comparing adult versus juvenile primary human islets (Arda et al., 2016). Also consistent with this previous report is the observation that inducible deletion of LIN28B led to the downregulation of genes characteristic of juvenile primary human islets. In addition, the expression of a number of markers of pancreatic β cell function and maturation, including PDX1, NKX6.1, MAFA, PAX6, and ISL1, were higher upon LIN28B deletion. Together, these findings show that LIN28B suppresses maturation of hESC-derived β cells.
How LIN28B suppresses maturation is unclear. A major role of LIN28B is to inhibit the biogenesis of let-7 (Piskounova et al., 2011). However, in the context of hESC differentiation toward β cells, the reduction of LIN28B led to a very modest increase in the levels of let-7 family members. Furthermore, there was no enrichment among downregulated genes for let-7 targets and the overexpression of let-7 to levels higher than seen in LIN28B-deleted cells did not promote maturation. Therefore, LIN28 appears to be acting through let-7-independent mechanisms to enhance β cell maturation. Let-7-independent roles for LIN28 have been previously reported in different contexts (Peng et al., 2011, Tsialikas and Romer-Seibert, 2015, Xu et al., 2009, Zhang et al., 2016, Zhu et al., 2011). Importantly, our results do not rule out a role for the very high levels of let-7 seen in in vivo matured β cells and adult primary islets promote β cell maturation. We were not able to achieve those levels in our experiments.
A connection between LIN28 and glucose metabolism has been reported in mice. Whole-body LIN28A- and LIN28B-overexpressing transgenic animals are more sensitive to insulin and have reduced peripheral glucose levels (Zhu et al., 2011). These results compared with ours suggest opposite effects of LIN28 in the cells that produce insulin versus cells that receive the insulin signal. However, caution should be taken when comparing mouse and human β cell maturation. For example, in mouse, increased basal insulin secretion has been associated with immaturity (Blum et al., 2012, Puri et al., 2018), while in human, both basal and stimulated insulin secretion is higher in adult versus juvenile β cells (Arda et al., 2016).
Poor glucose management is associated with long-term diabetic consequences including diabetic retinopathy, nephropathy, and neuropathy (Cade, 2008). Transplantation of hESC-derived β cells holds great promise for improving glucose management and thus minimizing the negative consequences. Understanding barriers to β cell maturation has the potential to improve the functionality of transplanted cells. Our work provides insight into one such barrier whose activity could be targeted by ongoing efforts to find small-molecule inhibitors of LIN28 and its partners (Roos et al., 2016).
Experimental Procedures
Cell Culture and hESC-β Cell Differentiation
Undifferentiated INS-GFP hES cells (Micallef et al., 2012) were maintained and differentiated into hESC-β like cells and eBCs as described previously (Nair et al., 2019, Russ et al., 2015). Human islets were from the UCSF Islets and Cellular Production Facility. The procurement and use of human islets used in the study was approved by the institutional biosafety committee at UCSF. The study is compliant with all relevant ethical regulations regarding research involving human participants, and informed consent was obtained by all participants at the isolation facility.
Mice
NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ mice (NSG) were obtained from Jackson Laboratories. Mice used in this study were maintained according to protocols approved by the University of California, San Francisco Committee on Laboratory Animal Resource Center. Mouse kidney capsule grafts have been described previously (Russ et al., 2015).
Flow Cytometry
Stained cells were run on LSRFortessa X20 DualData and analyzed with FlowJo software. Detailed staining methods including antibodies in Supplemental Information.
iCRISPR LIN28 and iLet7
Construction of the iCRISPR line was built as described previously (Gonzalez et al., 2014). The iLET-7 strategy is shown in Figure 4A. Details in Supplemental Information.
Small RNA-Seq and RNA-Seq
Small RNA-seq libraries were made as described previously (Hafner et al., 2012). RNA-seq libraries were made by using SMART-Seq v4 Ultra Low Input RNA Kit for Sequencing (Takara) and Nextera XT DNA Library Preparation kit (Illumina) thereafter.
Small RNA-Seq and RNA-Seq Data Analysis
For RNA-seq analysis, the data were preprocessed using Kallisto (Bray et al., 2016) and index to Gencode v.24. For the miRNA-seq analysis, reads were aligned using Bowtie v1.1.2 (-n 0 -L 18 -best) to a hairpin genome downloaded from miRbase (Langmead et al., 2009). Differential expression analysis was performed using in R using the Limma-Voom analysis (Liu et al., 2015, Ritchie et al., 2015). Let-7 targets were obtained from the TargetScan Release 7.1:June 2016 let-7-5p/98-5p list. GSEA analysis was performed using the current release (July 16, 2018) from http://www.gsea-msigdb.org/gsea/index.jsp (Mootha et al., 2003, Subramanian et al., 2005).
qRT-PCR
Total RNA was extracted with RNeasy Micro Kit (QIAGEN), treated with DNase I Kit (QIAGEN), and reverse transcribed using SuperScript III Kit (Invitrogen) as per the manufacturer's instructions.
miRNA qRT-PCR has been described previously (Moltzahn et al., 2011). Primers and probes can be found in Supplemental Information.
Western Blots
Antibodies and concentrations used can be found in Supplemental Information. Imaging was performed using an Odyssey LICOR scanner and quantified using ImageJ.
T7 Endonuclease I Assay
Genomic regions flanking the CRSIPR target sites were PCR amplified. Purified PCR products were denatured and reannealed and then treated with the T7 Endonuclease I Assay (New England Biolabs). Indel percentage was determined by the formula: %gene modification = 100 × (1–(1–fraction cleaved)1/2).
GSIS Assays
For static insulin secretion assays, cells were treated at the indicated glucose concentrations, and supernatant was collected. For dynamic insulin secretion assays, eBCs were assayed using the perifusion system from Biorep Technologies. Flow-through was collected over the course of the experiment. C-peptide levels were measured using the STELLUX Chemi Human C-peptide ELISA kit (Alpco).
Author Contributions
X.Z., M.H., and R.B. conceived of the experimental study. X.Z. performed the experiments presented in the figures except as noted below. G.G.N. developed the protocol to produce β-like cells and eBCs and conducted the experiments with end-stage cells, including perifusion assays. M.-L.L. differentiated the cells. H.A.R. developed the iCrLIN28B and iLet-7 cell lines, and performed in vivo transplants. M.S. and H.A.R. performed original experiments leading to premise for project. M.S. also produced sequencing libraries for Figures 1B, 1C, 2A, and 2C. C.D.B. performed all genomic analyses. X.Z., R.B., and G.G.N. wrote the paper with help from M.H.
Acknowledgments
This work was funded by a grant from the Leona M. and Harry B. Helmsley Charitable Trust; United States, to R.B. Stem cell work in M.H. laboratory was supported by grants from the NIH (DK105831, DK108666); United States. G.G.N. and H.A.R. were supported by fellowship grants from the JDRF; United States.
Published: December 26, 2019
Footnotes
Supplemental Information can be found online at https://doi.org/10.1016/j.stemcr.2019.11.009.
Contributor Information
Matthias Hebrok, Email: matthias.hebrok@ucsf.edu.
Robert Blelloch, Email: robert.blelloch@ucsf.edu.
Accession Numbers
The GEO accession number for the genomic data presented is GSE108654.
Supplemental Information
The sequence of let-7 overexpression cassette in iLET-7. miRNAs that are excised by Dicer are in red, pre-miR-21 loop in blue.
References
- Arda H.E., Li L., Tsai J., Torre E.A., Rosli Y., Peiris H., Spitale R.C., Dai C., Gu X., Qu K. Age-dependent pancreatic gene regulation reveals mechanisms governing human beta cell function. Cell Metab. 2016;23:909–920. doi: 10.1016/j.cmet.2016.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artner I., Hang Y., Mazur M., Yamamoto T., Guo M., Lindner J., Magnuson M.A., Stein R. MafA and MafB regulate genes critical to beta-cells in a unique temporal manner. Diabetes. 2010;59:2530–2539. doi: 10.2337/db10-0190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blum B., Hrvatin S., Schuetz C., Bonal C., Rezania A., Melton D.A. Functional beta-cell maturation is marked by an increased glucose threshold and by expression of urocortin 3. Nat. Biotechnol. 2012;30:261–264. doi: 10.1038/nbt.2141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray N.L., Pimentel H., Melsted P., Pachter L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016;34:525–527. doi: 10.1038/nbt.3519. [DOI] [PubMed] [Google Scholar]
- Cabrera O., Berman D.M., Kenyon N.S., Ricordi C., Berggren P.O., Caicedo A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. U S A. 2006;103:2334–2339. doi: 10.1073/pnas.0510790103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cade W.T. Diabetes-related microvascular and macrovascular diseases in the physical therapy setting. Phys. Ther. 2008;88:1322–1335. doi: 10.2522/ptj.20080008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cerf M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol. (Lausanne) 2013;4:37. doi: 10.3389/fendo.2013.00037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao C.S., Loomis Z.L., Lee J.E., Sussel L. Genetic identification of a novel NeuroD1 function in the early differentiation of islet alpha, PP and epsilon cells. Dev. Biol. 2007;312:523–532. doi: 10.1016/j.ydbio.2007.09.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ediger B.N., Lim H.W., Juliana C., Groff D.N., Williams L.T., Dominguez G., Liu J.H., Taylor B.L., Walp E.R., Kameswaran V. LIM domain-binding 1 maintains the terminally differentiated state of pancreatic beta cells. J. Clin. Invest. 2017;127:215–229. doi: 10.1172/JCI88016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faleo G., Russ H.A., Wisel S., Parent A.V., Nguyen V., Nair G.G., Freise J.E., Villanueva K.E., Szot G.L., Hebrok M. Mitigating ischemic injury of stem cell-derived insulin-producing cells after transplant. Stem Cell Reports. 2017;9:807–819. doi: 10.1016/j.stemcr.2017.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman R.C., Farh K.K., Burge C.B., Bartel D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19:92–105. doi: 10.1101/gr.082701.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao T., McKenna B., Li C., Reichert M., Nguyen J., Singh T., Yang C., Pannikar A., Doliba N., Zhang T. Pdx1 maintains beta cell identity and function by repressing an alpha cell program. Cell Metab. 2014;19:259–271. doi: 10.1016/j.cmet.2013.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez F., Zhu Z., Shi Z.D., Lelli K., Verma N., Li Q.V., Huangfu D. An iCRISPR platform for rapid, multiplexable, and inducible genome editing in human pluripotent stem cells. Cell Stem Cell. 2014;15:215–226. doi: 10.1016/j.stem.2014.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu C., Stein G.H., Pan N., Goebbels S., Hornberg H., Nave K.A., Herrera P., White P., Kaestner K.H., Sussel L. Pancreatic beta cells require NeuroD to achieve and maintain functional maturity. Cell Metab. 2010;11:298–310. doi: 10.1016/j.cmet.2010.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hafner M., Renwick N., Farazi T.A., Mihailovic A., Pena J.T., Tuschl T. Barcoded cDNA library preparation for small RNA profiling by next-generation sequencing. Methods. 2012;58:164–170. doi: 10.1016/j.ymeth.2012.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hang Y., Stein R. MafA and MafB activity in pancreatic beta cells. Trends Endocrinol. Metab. 2011;22:364–373. doi: 10.1016/j.tem.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar M.S., Erkeland S.J., Pester R.E., Chen C.Y., Ebert M.S., Sharp P.A., Jacks T. Suppression of non-small cell lung tumor development by the let-7 microRNA family. Proc. Natl. Acad. Sci. U S A. 2008;105:3903–3908. doi: 10.1073/pnas.0712321105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead B., Trapnell C., Pop M., Salzberg S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009;10:R25. doi: 10.1186/gb-2009-10-3-r25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liew C.G. Generation of insulin-producing cells from pluripotent stem cells: from the selection of cell sources to the optimization of protocols. Rev. Diabet. Stud. 2010;7:82–92. doi: 10.1900/RDS.2010.7.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu R., Holik A.Z., Su S., Jansz N., Chen K., Leong H.S., Blewitt M.E., Asselin-Labat M.L., Smyth G.K., Ritchie M.E. Why weight? Modelling sample and observational level variability improves power in RNA-seq analyses. Nucleic Acids Res. 2015;43:e97. doi: 10.1093/nar/gkv412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Micallef S.J., Li X., Schiesser J.V., Hirst C.E., Yu Q.C., Lim S.M., Nostro M.C., Elliott D.A., Sarangi F., Harrison L.C. INS(GFP/w) human embryonic stem cells facilitate isolation of in vitro derived insulin-producing cells. Diabetologia. 2012;55:694–706. doi: 10.1007/s00125-011-2379-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moltzahn F., Hunkapiller N., Mir A.A., Imbar T., Blelloch R. High throughput microRNA profiling: optimized multiplex qRT-PCR at nanoliter scale on the fluidigm dynamic arrayTM IFCs. J. Vis. Exp. 2011;54 doi: 10.3791/2552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mootha V.K., Lindgren C.M., Eriksson K.F., Subramanian A., Sihag S., Lehar J., Puigserver P., Carlsson E., Ridderstrale M., Laurila E. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003;34:267–273. doi: 10.1038/ng1180. [DOI] [PubMed] [Google Scholar]
- Nair G., Hebrok M. Islet formation in mice and men: lessons for the generation of functional insulin-producing β-cells from human pluripotent stem cells. Curr. Opin. Genet. Dev. 2015;32:171–180. doi: 10.1016/j.gde.2015.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nair G.G., Liu J.S., Russ H.A., Tran S., Saxton M.S., Chen R., Juang C., Li M.L., Nguyen V.Q., Giacometti S. Recapitulating endocrine cell clustering in culture promotes maturation of human stem-cell-derived beta cells. Nat. Cell Biol. 2019;21:263–274. doi: 10.1038/s41556-018-0271-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nam Y., Chen C., Gregory R.I., Chou J.J., Sliz P. Molecular basis for interaction of let-7 microRNAs with Lin28. Cell. 2011;147:1080–1091. doi: 10.1016/j.cell.2011.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng S., Chen L.L., Lei X.X., Yang L., Lin H., Carmichael G.G., Huang Y. Genome-wide studies reveal that Lin28 enhances the translation of genes important for growth and survival of human embryonic stem cells. Stem Cells. 2011;29:496–504. doi: 10.1002/stem.591. [DOI] [PubMed] [Google Scholar]
- Piskounova E., Polytarchou C., Thornton J.E., LaPierre R.J., Pothoulakis C., Hagan J.P., Iliopoulos D., Gregory R.I. Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms. Cell. 2011;147:1066–1079. doi: 10.1016/j.cell.2011.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puri S., Roy N., Russ H.A., Leonhardt L., French E.K., Roy R., Bengtsson H., Scott D.K., Stewart A.F., Hebrok M. Replication confers beta cell immaturity. Nat. Commun. 2018;9:485. doi: 10.1038/s41467-018-02939-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie M.E., Phipson B., Wu D., Hu Y., Law C.W., Shi W., Smyth G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015;43:e47. doi: 10.1093/nar/gkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roglic G., World Health Organization . World Health Organization; 2016. Global Report on Diabetes. [Google Scholar]
- Roos M., Pradere U., Ngondo R.P., Behera A., Allegrini S., Civenni G., Zagalak J.A., Marchand J.R., Menzi M., Towbin H. A small-molecule inhibitor of Lin28. ACS Chem. Biol. 2016;11:2773–2781. doi: 10.1021/acschembio.6b00232. [DOI] [PubMed] [Google Scholar]
- Roush S., Slack F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008;18:505–516. doi: 10.1016/j.tcb.2008.07.007. [DOI] [PubMed] [Google Scholar]
- Russ H.A., Parent A.V., Ringler J.J., Hennings T.G., Nair G.G., Shveygert M., Guo T., Puri S., Haataja L., Cirulli V. Controlled induction of human pancreatic progenitors produces functional beta-like cells in vitro. EMBO J. 2015;34:1759–1772. doi: 10.15252/embj.201591058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rybak A., Fuchs H., Smirnova L., Brandt C., Pohl E.E., Nitsch R., Wulczyn F.G. A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat. Cell Biol. 2008;10:987–993. doi: 10.1038/ncb1759. [DOI] [PubMed] [Google Scholar]
- Schaffer A.E., Taylor B.L., Benthuysen J.R., Liu J., Thorel F., Yuan W., Jiao Y., Kaestner K.H., Herrera P.L., Magnuson M.A. Nkx6.1 controls a gene regulatory network required for establishing and maintaining pancreatic beta cell identity. PLoS Genet. 2013;9:e1003274. doi: 10.1371/journal.pgen.1003274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shenoy A., Blelloch R.H. Regulation of microRNA function in somatic stem cell proliferation and differentiation. Nat. Rev. Mol. Cell Biol. 2014;15:565–576. doi: 10.1038/nrm3854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shyh-Chang N., Daley G.Q. Lin28: primal regulator of growth and metabolism in stem cells. Cell Stem Cell. 2013;12:395–406. doi: 10.1016/j.stem.2013.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sneddon J.B., Tang Q., Stock P., Bluestone J.A., Roy S., Desai T., Hebrok M. Stem cell therapies for treating diabetes: progress and remaining challenges. Cell Stem Cell. 2018;22:810–823. doi: 10.1016/j.stem.2018.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian A., Tamayo P., Mootha V.K., Mukherjee S., Ebert B.L., Gillette M.A., Paulovich A., Pomeroy S.L., Golub T.R., Lander E.S. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U S A. 2005;102:15545–15550. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thornton J.E., Gregory R.I. How does Lin28 let-7 control development and disease? Trends Cell Biol. 2012;22:474–482. doi: 10.1016/j.tcb.2012.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsialikas J., Romer-Seibert J. LIN28: roles and regulation in development and beyond. Development. 2015;142:2397–2404. doi: 10.1242/dev.117580. [DOI] [PubMed] [Google Scholar]
- Velazco-Cruz L., Song J., Maxwell K.G., Goedegebuure M.M., Augsornworawat P., Hogrebe N.J., Millman J.R. Acquisition of dynamic function in human stem cell-derived beta cells. Stem Cell Reports. 2019;12:351–365. doi: 10.1016/j.stemcr.2018.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veres A., Faust A.L., Bushnell H.L., Engquist E.N., Kenty J.H., Harb G., Poh Y.C., Sintov E., Gurtler M., Pagliuca F.W. Charting cellular identity during human in vitro beta-cell differentiation. Nature. 2019;569:368–373. doi: 10.1038/s41586-019-1168-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viswanathan S.R., Daley G.Q., Gregory R.I. Selective blockade of microRNA processing by Lin28. Science. 2008;320:97–100. doi: 10.1126/science.1154040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu B., Zhang K., Huang Y. Lin28 modulates cell growth and associates with a subset of cell cycle regulator mRNAs in mouse embryonic stem cells. RNA. 2009;15:357–361. doi: 10.1261/rna.1368009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X., Lin X., Zhong X., Kaur S., Li N., Liang S., Lassus H., Wang L., Katsaros D., Montone K. Double-negative feedback loop between reprogramming factor LIN28 and microRNA let-7 regulates aldehyde dehydrogenase 1-positive cancer stem cells. Cancer Res. 2010;70:9463–9472. doi: 10.1158/0008-5472.CAN-10-2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J., Ratanasirintrawoot S., Chandrasekaran S., Wu Z., Ficarro S.B., Yu C., Ross C.A., Cacchiarelli D., Xia Q., Seligson M. LIN28 regulates stem cell metabolism and conversion to primed pluripotency. Cell Stem Cell. 2016;19:66–80. doi: 10.1016/j.stem.2016.05.009. [DOI] [PubMed] [Google Scholar]
- Zhu H., Shyh-Chang N., Segre A.V., Shinoda G., Shah S.P., Einhorn W.S., Takeuchi A., Engreitz J.M., Hagan J.P., Kharas M.G. The Lin28/let-7 axis regulates glucose metabolism. Cell. 2011;147:81–94. doi: 10.1016/j.cell.2011.08.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The sequence of let-7 overexpression cassette in iLET-7. miRNAs that are excised by Dicer are in red, pre-miR-21 loop in blue.