

Abstract
Slit and its receptor Roundabout (Robo) are important for neuronal development and neo-angiogenesis in various neoplastic and non-neoplastic diseases. Angiogenesis is a key factor for tumor growth and other angiogenesis-dependent diseases including rheumatoid arthritis, and chronic inflammation Recently, over-expression of Slit/Robo1 family proteins has been reported in several types of malignancy. We explored the expression of Robo1 in neoplastic and non-neoplastic diseases with a focus on newly formed blood vessels. Three hundred and thirty four cases of malignancy and forty five cases of angiogenic diseases were recruited. Using the A7241A Robo1 monoclonal antibody, Robo1 expression was validated by immunohistochemistry. Among malignant cases, endothelial cells of newly formed blood vessels in 283 tumors (84.7%) exhibited positive staining with above antibody. In non-neoplastic diseases, newly formed blood vessels were positive in 70.6% (12/17) cases of chronic inflammation, 100% (18/18) cases of pyogenic granuloma and 83.3% (5/6) cases of rheumatoid arthritis. Recently, newly anti-angiogenesis therapy is drawing attention as effective therapy for angiogenesis-dependent diseases without regard to their neoplastic or non-neoplastic nature. Our results showed a large number of neoplastic and non-neoplastic diseases showed positive staining for ROBO1 by immunohistochemistry. Thus, Robo1 targeted therapy may create new strategies for the treatment of angiogenic-dependent diseases through the suppression of angiogenesis. Further, besides the majority of liver cell carcinomas (23/28, 82.1%), Robo1 was positive in 100% of the squamous cell carcinoma of the esophagus, uterine cervix, lung and skin. Thus, immunohistochemical evaluation of Robo1 may be useful as an additional diagnostic tool for liver cell carcinomas and squamous cell carcinomas.
Keywords: Angiogenesis, immunohistochemistry, inflammation, neoplasia, ROBO1
Introduction
The Slit family members and their receptor Roundabout (Robo) are important factors in neuronal development and leukocyte cell migration [1]. Recent work on mammals has indicated there are three Slit proteins and four forms of Robo (Robo1-4) [2,3]. ROBO1 is the human homolog of the Drosophila roundabout gene, which defines a novel subfamily of the immunoglobulin superfamily highly conserved from fruit flies to mammals [2]. The human Robo1 protein consists of five immunoglobulin-like (Ig) and three fibronectin type3-like (Fn3) domains in its extracellular region, and four cytoplasmic motifs termed CC0, CC1, CC2 and CC3 in its intracellular region [3].
Angiogenesis is an important process under a variety of physiological and pathological conditions such as hemostasis, ischemia and hypoxia and wound healing, as well as in cancer growth and metastasis [4-6]. It occurs not only in tumor formation, but also in a range of non-neoplastic diseases that may be classified together as either angiogenesis-dependent [5] or angiogenic [7]. Angiogenesis-dependent diseases include diabetic retinopathy, rheumatoid arthritis, and chronic inflammation [5]. Various growth factors and receptors such as VEGF are involved in angiogenesis [6], and the angiogenic properties of neuronal guidance factors, such as semaphorines, ephrins, slits and netrins, and their cognate receptors, have recently attracted a great deal of attention [8,9]. Involvement of most members of the Slit/Robo family has been reported, but their roles in the progression of angiogenesis-dependent disease including various cancers have not been clearly defined [10]. Robo1 is expressed in human primary lymphatic endothelial cells (hLECs) and Slit2/Robo1 signaling promotes the lymphangiogenic activity of cultured hLECs [11]. In addition, Slit-Robo1 signaling pathways play a causal role in tumor angiogenesis in chemical-induced hamster squamous cell carcinogenesis [12]. MicroRNA218 inhibits the metastasis of gastric cancer by acting on the receptor Robo1 through angiogenesis, and this Slit-miR218-Robo1 pathway plays a pivotal role in gastric cancer metastasis [13]. Increased immunoreactivity to Robo1 and Slit is an important factor for recurrence, clinical stage, and angiogenesis in endometrial cancer cases [14].
Slit/Robo family members may impact on the genesis of many solid tumors [15], and multiple Slit/Robo pathway genes are inactivated by hypermethylation of their promotors in cervical cancer progression [16]. Low Robo1 expression is associated with the development of cholangiocarcinomas [17], and homogenous deletions have been observed in lung and breast tumor cell lines [18]. Most mice with deletion of exon 2 of ROBO1 die because of delayed lung maturation, but survivors develop bronchial hyperplasia [19]. In mice, ROBO1 is a classic tumor suppressor gene requiring inactivation of both alleles for lung adenocarcinoma and lymphoma development [20]. Thus, in some tumors, it has been postulated as a tumor suppressor gene [21].
However, Ito et al showed contrasting overexpression of Robo1 in hepatocellular carcinomas [22]. Breast cancer cells and tissues derived from breast cancer patients express Robo1 and 2 receptors, and Slit treatment inhibits CXCR4-induced breast cancer cell chemotaxis, chemoinvasion and adhesion during metastasis [23]. Slit and Robo1 are both overexpressed in prostate tumors [24] while Robo1 and Robo4 were found to be overexpressed in colorectal cancer at the mRNA level and immunohistochemistry demonstrated respective locations in tumor cells, and endothelial cells of tumor vessels [25]. Thus, roles of the Slit/Robo pathway in human cancers appear complex and tissue-specific expression patterns of Robo1 clearly require further investigation.
We previously developed a monoclonal antibody for Robo1, A7241A, which may be utilized for immunohistochemistry [22]. In this study, we made an extensive survey of Robo1 expression in various human tissues and newly formed blood vessels in both neoplastic and non-neoplastic diseases and demonstrated broad distribution in endothelial cells of in both tumors and angiogenic diseases. These findings provide insight into physiological and pathological process. These may have implications for diagnosis as well as therapy.
Materials and methods
Specimen characteristics
This study was approved by the Ethics Committees of Niigata Medical and Dental University and The University of Tokyo. The tissue specimens were collected at Niigata Medical and Dental University Hospital from 2003 to 2010, and consisted of 438 normal samples (heart, aorta, oral mucosa, salivary gland, breast, larynx, lung, esophagus, stomach, small intestine, colon, liver, gallbladder, pancreas, spleen, kidney, ureter, urinary bladder, adrenal gland, testis, ovary, uterus, Fallopian tube, placenta, prostate, thyroid, parathyroid, tonsil, thymus, eye, brain, abdominal lymph node, skin, nasal cavity, tongue, pituitary gland, choroid plexus, and skeletal muscle (Table 1), 334 of various carcinomas and 45 angiogenic diseases (Table 2). The carcinomas comprised 28 cases of hepatocellular carcinoma, 13 of skin carcinoma (squamous cell carcinoma), 11 of cervical cancer (squamous cell carcinoma), 13 of esophageal squamous cell carcinoma, 4 cases of esophageal adenocarcinoma, 68 of lung cancer (squamous cell carcinoma, adenocarcinoma, small cell carcinoma, large cell carcinoma, and carcinoid tumor), 16 of renal cell carcinoma, 35 of pancreas cancer (adenocarcinoma), 35 cases of colon cancer (adenocarcinoma), 36 of ovarian cancer (adenocarcinoma) and 11 of gastric cancer (adenocarcinoma) (Table 3). As background hepatocellular carcinoma-associated diseases, there were 18 cases of liver cirrhosis, two of precirrhosis, 4 cases of liver fibrosis, and 2 cases of chronic active hepatitis (Table 4). As angiogenic diseases 17 cases of chronic inflammation (granulation), 16 of pyogenic granuloma, 6 of rheumatoid arthritis and 6 of bulla were included (Table 2). All tissues were fixed in 10% formaldehyde and embedded in paraffin.
Table 1.
Robo1 expression in human normal tissues
| Organ | Tssues | N | Results |
|---|---|---|---|
| Skin | Squamous epithelium | 11 | ± (basal cell layer) |
| Chroid plexus | 2 | - | |
| Heart | Striated muscle | 6 | - |
| Kidney | Glomeruli | 13 | ± (blood vessel) |
| Esophagus | Squamous epithelium | 10 | ± (basal cell layer) |
| Lung | 30 | - | |
| Liver | 22 | - | |
| Colon | 35 | - | |
| Stomach | 6 | - | |
| Small intestine | 6 | - | |
| Pancreas | 35 | - | |
| Spleen | 6 | ± (blood vessel) | |
| Breast | Duct epithelium | 44 | - |
| Uterine corpus | Endometrial stroma | 33 | + (blood vessel) |
| Ovary | 36 | - | |
| Uterine cervix | Squamous epithelium | 10 | ± (basal cell layer) |
| Abdominal lymph node | 6 | + (blood vessel) | |
| Gallbladder | 6 | - | |
| Parathyroid | 3 | - | |
| Thyroid | 6 | - | |
| Adrenal gland | 10 | - | |
| Salivary gland | 6 | - | |
| Pituitary gland | 3 | - | |
| Brain | 2 | - | |
| Cerebellum | 2 | - | |
| Thymus | 3 | - | |
| Tonsil | 6 | ± (blood vessel) | |
| Oral cavity | Squamous epithelium | 6 | - |
| Skeletal muscle | 6 | - | |
| Nasal cavity | Ciliated epithelium | 6 | - |
| Urinary bladder | Transitional epithelium | 6 | - |
| Tongue | Squamous epithelium | 6 | - |
| Aorta | Endotheium | 6 | - |
| Larynx | Squamous epithelium | 2 | - |
| Fallopian tube | 6 | - | |
| Plascenta | 6 | - | |
| Tastis | 12 | - | |
| Ureter | 6 | - | |
| Eye | Cornea | 6 | - |
| Prostate | 6 | - |
Table 2.
ROBO1 expression in new blood vessel in angiogenesis-dependent diseases
| New blood vessels | Type | N | Immunostaining | % |
|---|---|---|---|---|
| Malignant tumor | 334 | +++ 283/334 | 84.7 | |
| Chronic inflammation | Granulation | 17 | +++ 12/17 | 70.6 |
| Angiogenesis-dependent diseases | ||||
| Granuloma pyogenicum | 16 | +++ 16/16 | 100 | |
| Rheumatoid arthritis | 6 | +++ 5/6 | 83.3 | |
| Bulla | 6 | -~± 1/6 | 16.7 |
Table 3.
Robo1 expression in human cancers and related non-neoplastic diseases
| Organ | Histological type | Positivity | Staining | % |
|---|---|---|---|---|
| Skin | Squamous cell carcinoma | 13/13 | +++ | 100 |
| Uterine cervix | Squamous cell carcinoma | 11/11 | +++ | 100 |
| Esophagus | Sqaumous cell carcinoma | 13/13 | +++ | 100 |
| Adenocarcinoma | 0/4 | - | 0 | |
| Lung | Squamoua cell carcinoma | 27/39 | ++ | 70 |
| Adenocarcinoma | 3/16 | -~± | 17.8 | |
| Small cell carcinoma | 3/5 | -~+ | 60 | |
| Large cell carcinoma | 0/4 | - | 0 | |
| Carcinoid | 4/4 | ++ | 100 | |
| Liver | Hepatocellular carcinoma | 22/28 | -~+++ | 78.6 |
| Cirrhosis | 0/10 | - | 0 | |
| Hepatitis | 0/10 | - | 0 | |
| Breast | Ductal carcinoma | 11/44 | -~+ | 25 |
| Kidney | Renal cell carcinoma | 0/16 | - | 0 |
| Pancreas | Adenocarcinoma | 8/35 | -~± | 22.9 |
| Colon | Adenocarcinoma | 5/35 | -~± | 14.3 |
| Ovary | Adenocarcinoma | 3/36 | -~± | 8.3 |
| Stomach | Adenocarcinoma | 1/11 | -~± | 9 |
-: negative, ±: weakly positive, +: positive, ++: moderately positive, +++: strongly positive.
Table 4.
Immunohistochemical expression of Robo1 and glypican by HCC
| No. | Differentiation of HCC | Age | Sex | Background | Virus | Glypican staining | Robo1 staining |
|---|---|---|---|---|---|---|---|
| 1 | Poor | 74 | M | LC | C | + | +++ |
| 2 | Poor | 49 | M | LC | B | + | +++ |
| 3 | Poor | 76 | M | LF | C | - | +++ |
| 4 | Poor | 71 | F | AIH | - | ++ | +++ |
| 5 | Poor moderate | >59 | M | CAH | B | + | +++ |
| 6 | Poor moderate | >71 | F | LC | C | ++ | +++ |
| 7 | Moderate~poor | 62 | M | LC | B | - | ++ |
| 8 | Moderate~poor | 63 | M | LF | B | - | +++ |
| 9 | Moderate~poor | 55 | F | LC | B | ++ | ++ |
| 10 | Moderate~poor | 55 | M | LC | B | ±~+ | +++ |
| 11 | Moderate~poor | 69 | M | LC | NBNC | - | + |
| 12 | Moderate | 76 | F | LF | C | + | +++ |
| 13 | Moderate | 80 | M | Lc | C | ++ | +++ |
| 14 | Moderate | 60 | F | LC | C | + | +++ |
| 15 | Moderate | 70 | F | LC | C | + | +++ |
| 16 | Moderate | 70 | F | LC | C | - | +++ |
| 17 | Moderate | 46 | M | LC | C | ++ | + |
| 18 | Moderate | 73 | F | LC | C | ++ | + |
| 19 | Moderate | 65 | M | LC | C | -~± | +++ |
| 20 | Moderate | 70 | M | LF | NBNC | - | +++ |
| 21 | Well~moderate | 50 | M | CAH | B | - | -~+++ |
| 22 | Well | 71 | M | B | C | ++ | - |
| 23 | Well | 62 | M | C | C | - | - |
| 24 | Well | 81 | M | C | C | ND | - |
| 25 | Well | 71 | M | C | C | - | - |
| 26 | Well | 67 | M | C | C | - | - |
| 27 | Well | 71 | M | C | C | - | -~+++ |
| 28 | Well | 66 | M | C | C | - | -~+++ |
LC: Liver cirrhosis, LF: Liver fibrosis, CAH: Chronic active hepatitis, AIH: Autoimmune hepatitis, NBNC: non-B non-C, ND: not done, -: negative, ±: positive, +: positive, ++: moderately positive, +++: strongly positive.
Western blotting
Chinese hamster ovary cells (CHO) stably expressing human Robo1 (Robo1/CHO) or Robo4 (Robo4/CHO) were established using the Flp-In system (Life Technologies Japan, Tokyo) with insertion of cDNAs for Robo1 or Robo4 into a pcDNA5/FRT mammalian expression vector (Life Technologies Japan) followed by transfection of construct FLP-CHO cells (Life Technologies Japan) according to the manufacturer’s instructions. The human hepatocellular carcinoma cell line HepG2 and human small cell lung carcinoma cell line NCI-H69 were obtained from American Type Culture Collection (ATCC). Aliquots of 1×107 cells were homogenized with a radio immunoprecipitation assay (RIPA) buffer containing 50 mM Tris-HCl (pH 7.6), 150 mM NaCl, 1% Nonidet-P40, 0.5% Sodium Deoxycholate, 0.1% SDS and Protease Inhibitor Cocktail-Ethylenediaminetetraacetic acid free (CompleteTM, Roche Diagnostics KK), then subjected to SDS-PAGE followed by western blot analysis using the A7241A Robo1 monoclonal antibody at a final concentration of 1 ng/μl. The immuno-reacted band for Robo1 on membranes was detected by the ECL (Enhanced chemiluminescence) method using SuperSignalTM West Dura Extended Duration Substrate (Life Technologies Japan) and an LAS-4000 Image analyzer (GE Healthcare Japan, Tokyo).
Immunohistochemistry
For detection of Robo1 protein in tissue sections 3, 3’-diaminobenzidine (DAB) immunostaining method was applied. The anti-Robo1 monoclonal antibody A7241A was generated by immunizing with an N-terminal peptide of Robo1, as described [22]. Simple stain multi PO (Nichirei, Tokyo, Japan) was used for secondary antibody staining. The 1G12 glypican 3 antibody (Nichirei, Tokyo, Japan) specific for hepatocellular carcinoma was applied for a comparison. Briefly, 3 μm sections were deparaffinized, rehydrated, and treated with 3% hydrogen peroxide for 15 minutes to inhibit endogenous peroxidase activity. Following antigen epitope retrieval by autoclaving in Tris-HCL buffer pH9.0, the slides were incubated with Robo1 antibody with a dilution of ×500 overnight at room temperature. Glypican 3 antibody was used without dilution. After incubation with a rabbit anti-mouse secondary antibody, detection was with biotin-free horseradish peroxidase-labeled polymers obtained from DAKO Corporation (Tokyo, Japan). Staining was visualized using DAB substrate-chromogen solution with hematoxylin counterstaining.
Results
Reactivity of the A7241A anti-Robo1 antibody
To confirm the specific immunoreactivity of A7241A anti-human Robo1 monoclonal antibody, we performed western blot analysis. As shown in Figure 1, a band for Robo1 at 240 kDa was clearly detected in each lane for Robo1 transfected CHO cells (positive control), HepG2 cells and NCI-H69 cells, while no signals were detected in the lane for Robo4 transfected CHO cells (negative control). No other indispensable bands were observed.
Figure 1.
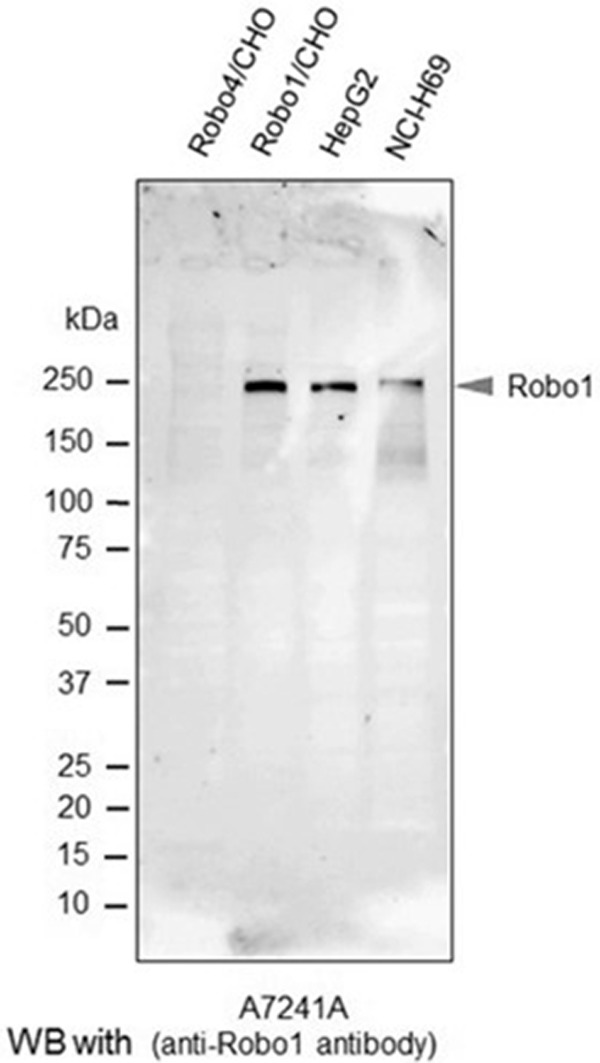
Western blot analysis with the anti-Robo1 antibody. Robo4/CHO (negative control), Robo1/CHO (positive control), HepG2 and NCI-H69 cells were homogenized in RIPA buffer, and subjected to SDS-PAGE followed by western blot analysis using the A7241A anti-human Robo1 monoclonal antibody. The arrowhead in the image by the ECL method indicates the Robo1 band.
Robo1 expression in various normal tissues
The expression pattern of Robo1 in various normal tissue is tabulated in Table 1. Robo1 was detected a membrane staining by immunohistochemistry. In normal tissues, weak staining was seen only in basal squamous cells in the skin, esophagus, tonsils, buccal mucosa, tongue, larynx, and uterine cervix. The other organs exhibited no staining for Robo1. Blood vessels of normal tissue showed weak staining in renal glomeruli, spleen, endometrium of uterine corpus, abdominal lymph nodes, and tonsil.
Robo1 expression in newly formed blood vessel in various cancer tissue
Results for expression patterns of Robo1 in newly formed blood vessels in various tumors and angiogenic diseases are summarized in Table 2. Among 334 malignancies, endothelial cells in 283 tumors (84.7%) exhibited positive staining with the Robo1 antibody, including examples of squamous cell carcinoma of the cervix, squamous cell carcinoma of the skin, adenocarcinoma of the stomach, and squamous cell carcinoma and adenocarcinoma of the lung (Figure 2A-C). It was striking that positive immunostaining for Robo1 was observed in vascular endothelial cells such as gastric adenocarcinoma and lung adenocarcinoma which were Robo-1 negative in epithelial elements (Figure 2B, 2C).
Figure 2.
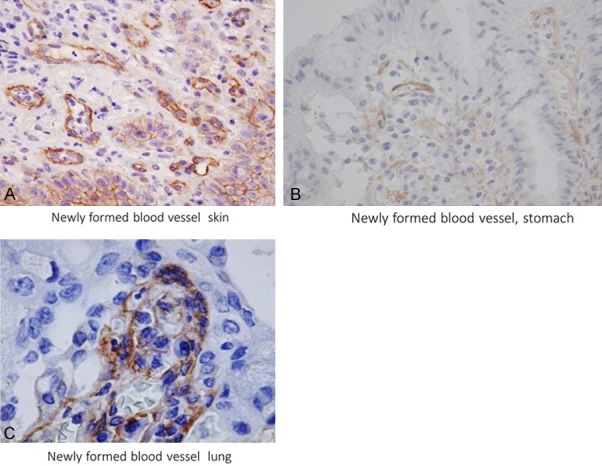
Robo1 expression in newly formed blood vessels of a malignant tumor. Skin squamous cell carcinoma (A) gastric adenocarcinoma (B), and lung adenocarcinoma (C). Robo1 immunostaining. H.E stain, (A, B) ×400; (C) ×1000.
Robo1 expression in newly formed blood vessels in non-cancerous tissues
Robo1 was also positive in newly organized vascular endothelial cells in various non-neoplastic angiogenic diseases (51/57, 89.5%) (Table 2). Newly formed blood vessels were positive for Robo1 immunostaining in 70.6% (12/17) cases of chronic inflammation, 100% (18/18) cases of pyogenic granuloma (16/16) (Figure 3A) and 83.3% (5/6) cases of rheumatoid arthritis (Figure 3B). In contrast, blood vessels of the bulla showed only weak staining in only one case (1/6). Vascular endothelial cells involved in the granulation in pyogenic granuloma stained more intensely than those far from the granulation sites (Figure 3A).
Figure 3.
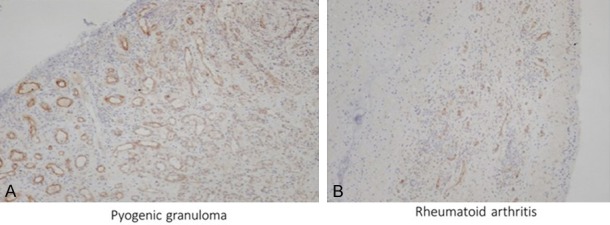
Robo1 expression in inflammatory diseases. Pyogenic granuloma (A) and rheumatoid arthritis (B) and exhibit positive Robo1 staining in their newly formed blood vessels. In pyogenic granuloma, Robo1 staining in the newly formed blood vessels is stronger in the surface region than near the normal tissue (A). Robo1 immunostaining. Robo1 immunostaining, (A, B) ×100.
Robo1 expression in epithelial cells of malignant tumors
Data for Robo1 expression in a variety of malignancies (334 cases) are shown in Table 3. In addition to the majority of liver cell carcinomas, Robo1 was positive in 100% of the squamous cell carcinoma cases of the esophagus, uterine cervix, and skin (Figure 4A). In the lung, 70% of the squamous cell carcinoma cases and 60% of the small cell carcinoma and 100% of the carcinoid tumor cases displayed Robo1 expression (Figure 4B, 4C). In breast carcinoma (ductal carcinoma) focal positive staining was observed. Adenocarcinomas tended to be less positive despite newly-formed blood vessels exhibiting strong expression (283 cases) (Figure 2C). Renal cell carcinomas exhibited no expression, along with large cell carcinomas of the lung.
Figure 4.
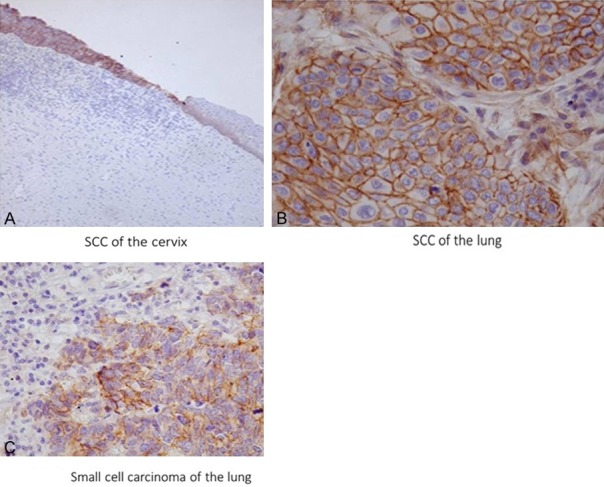
Positive immunostaining for epithelial cells of squamous cell carcinoma of the uterine cervix (A), squamous carcinoma of the lung (B), and small cell carcinoma of the lung (C). Note that strong membrane staining is apparent in squamous cell carcinoma of the uterus, but not in non-neoplastic squamous epithelium (right) except weakly in the basal layer. Robo1 immunostaining (A) ×100; (B, C) ×400.
Robo1 and glypican expression in hepatocellular carcinomas
Data for expression of Robo1 and glypican in individual hepatocellular carcinomas are tabulated in Table 4. In 28 cases, Robo1 was positive in 23 (82.1%) (Figure 5A-C; Table 4). Metastases also displayed positive binding of the antibody and detection of minute HCC foci was possible (Figure 5B). Vascular invasion by HCC was also detected by immunohistochemistry (Figure 5C). In agreement with the results of Ito et al. [22], no Robo1 expression was seen in background non-neoplastic hepatic diseases such as cirrhosis and hepatitis (Figure 5B, 5C). Glypican 3 proved positive in the cytoplasm of 15 of 28 cases (53.8%).
Figure 5.
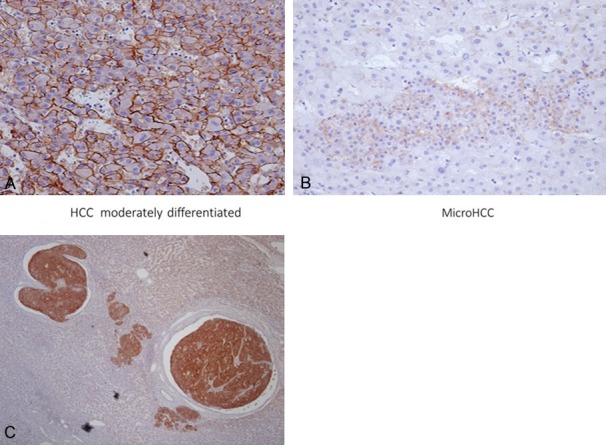
Robo1 staining in neoplastic and non-neoplastic liver tissues. Normal liver tissue (B, C) lacks staining, in contrast to HCC positivity. HCC exhibits strong membrane expression of Robo1 (A). A small focus of HCC can be seen surrounded by normal liver tissue (B). Tumor cells of HCC invading into vascular space are evident (C). Robo1 immunostaining (A, B) ×200; (C) ×40.
Discussion
The term angiogenesis is generally applied to the growth of primitive microvessels that are the size of capillary blood vessels, a process that is orchestrated by a range of angiogenic factors, such as VEGF and angiogenesis inhibitors [6,26,27]. Folkman originally suggested that tumor growth is dependent on angiogenesis [27]. In tumorigenesis, tumor cell-induced angiogenesis and tumor metastasis both involve cell migration. The Slit2 protein attracts endothelial cells in newly formed blood vessels and promotes tube formation through the Slit/Robo signaling cascade [3,28]. In this setting, Robo1, together with Robo4, may play important roles [3].
Growing evidence suggests that the induction of angiogenesis is a discrete component of the tumor phenotype, often activated during early, preneoplastic stages [4,29]. It is well-known that tumor growth depends on certain tumor angiogenesis factors, such as VEGF, α- and β-FGF, the inhibitor thrombospondin-1 and interferon α/β, and the balance of these factors [29]. However, their relations with Robo1, involved in endothelial cell migration, has remained unclear. Slit/Robo1 mediated human umbilical vein endothelial cell migration is suppressed by Robo4 [30]. In the present study, 90% of malignant tumors showed Robo1 positivity in their newly-formed blood vessels, even examples of otherwise Robo1-negative cancers such as gastric and lung adenocarcinomas (Figure 2B, 2C).
At present, a variety of anti-angiogenic drugs, such as Avastin and Endostatin, have been approved in many countries [31]. Anti-angiogenic therapy is widely applied [4,6,28,29,32]. For example, an anti VEGF agent was shown to be effective in certain cancers combined with chemotherapy [33]. MicroRNA218 inhibits the metastasis of gastric cancer by acting on the receptor Robo1, and this Slit-miR218-Robo1 pathway plays a pivotal role in gastric cancer metastasis [13]. Recently, it was verified in the mouse that inhibition of Robo1 by microRNA was effective for involution of gastric cancer through blocked angiogenesis [34]. These findings indicate that Slit/Robo signaling could be a novel therapeutic target of cancer therapy.
Previous studies on the expression of Robo1 in various tumors have generated contradictory results. In this study, squamous cell carcinoma of the skin, esophagus, and uterine cervix exhibited positive staining, whereas most adenocarcinomas of the esophagus, lung, and stomach did not. This result indicates a tissue specific expression pattern, with functions of Slit2/Robo1 signaling being cell-type dependent. In gastric cancer, activation of Slit/Robo1 signaling up-regulates the Wnt/β catenin pathway [35], and such activation of Wnt/β signaling also promotes genesis of colorectal cancer through Src signaling [36]. However, in our study, Robo1 was not found to be expressed in a large proportion of both gastric and colorectal cancers. The roles of Robo1 in carcinogenesis and neovascularization of cancer tissue clearly must be considered separately.
In addition to the angiogenesis evident in tumors, our results showed this as a key component of some inflammatory diseases. Rheumatoid arthritis is an inflammatory angiogenic disease which is mediated by pro-inflammatory and pro-angiogenic cytokines, characterized by VEGF decrease after the administration of certain effective drugs [37]. In our study, Robo1 was positive in newly organized blood vessels, not only in cancerous tissues, but also in granulation, pyogenic granuloma and rheumatoid arthritis. These findings point to new strategies of neutralizing Robo1 activity for the inhibition of both tumor growth and also amelioration of non-neoplastic angiogenic diseases.
The findings in this study suggest that Robo1 might be a useful marker in some types of cancer, especially in squamous cell carcinomas. This is the first study to specifically demonstrate utility of the Robo1 antibody for the diagnosis of squamous cell carcinoma. Robo1 also appears useful as a marker of hepatocellular carcinoma (HCC), in line with data of an RNA experiment by Ito et al [22] which indicated 70% of cases to be positive. Glypican 3 is well-known as the marker of hepatocellular carcinoma, but staining is focal [38]. Our study further showed that only 53.8% of the HCC cases were positive for Glypican 3 compared with 82.1% for Robo1, suggesting that the latter may be a more sensitive immunohistochemical marker. Furthermore, minute foci of HCC were detectable using Robo1.
Acknowledgements
This work was supported by the Program for Development of New Functional Antibody Technologies of the New Energy and Industrial Technology Development Organization (NEDO) of Japan, the Funding Program for World-Leading Innovative R&D on Science and Technology (FIRST program) of Japan Society for the Promotion of Science.
Disclosure of conflict of interest
None.
References
- 1.Rao Y, Wong K, Ward M, Jurgensen C, Wu JY. Neuronal migration and molecular conservation with leukocytechemotaxes. Genes Dev. 2002;16:2973–2984. doi: 10.1101/gad.1005802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kidd T, Brose K, Mitchell KJ, Fetter RD, Tessier-Levigne M, Goodman CS, Tear G. Roundabout controls axon crossing of the CNS midline and defines novel subfamily of evolutionary conserved guidance receptors. Cell. 1998;92:205–215. doi: 10.1016/s0092-8674(00)80915-0. [DOI] [PubMed] [Google Scholar]
- 3.Legg JA, Herbert JM, Clissold P, Bicknell R. Slits and roundabouts in cancer, tumor angiogenesis, and endothelial cell migration. Angiogenesis. 2008;11:13–21. doi: 10.1007/s10456-008-9100-x. [DOI] [PubMed] [Google Scholar]
- 4.Hanahan D, Weinberg R. The hallmarks of cancer. Cell. 2000;100:57–70. doi: 10.1016/s0092-8674(00)81683-9. [DOI] [PubMed] [Google Scholar]
- 5.Folkman J. Angiogenesis: an organizing principle for drug discovery. Drug Discovery. 2007;6:273–286. doi: 10.1038/nrd2115. [DOI] [PubMed] [Google Scholar]
- 6.Ferrara N. Role of vascular endothelial growth factor in physiologic and pathologic angiogenesis: therapeutic implications. Semin Oncol. 2002;29(Suppl 16):10–14. doi: 10.1053/sonc.2002.37264. [DOI] [PubMed] [Google Scholar]
- 7.Carmeliet P, Jain RK. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases. Nat Rev Drug Discov. 2011;10:417–427. doi: 10.1038/nrd3455. [DOI] [PubMed] [Google Scholar]
- 8.Klagsbrun M, Eichman A. A role for axon guidance receptors and ligands in blood vessel development and tumor angiogenesis. Cytokine Growth Factor Rev. 2005;16:535–548. doi: 10.1016/j.cytogfr.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 9.Shima D, Mailhos C. Vascular development biology: getting nervous. Curr Opin Gen Develop. 2000;10:536–542. doi: 10.1016/s0959-437x(00)00124-6. [DOI] [PubMed] [Google Scholar]
- 10.Fujiwara M, Ghazizadeh M, Kawanami O. Potential role of the Slit/Robo signal pathway in angiogenesis. Vasc Med. 2006;11:115–121. doi: 10.1191/1358863x06vm658ra. [DOI] [PubMed] [Google Scholar]
- 11.Yang XM, Han HX, Sui F, Dai YM, Chen M, Geng JG. Slit-Robo signaling mediates lymphangiogenesis and promotes tumor lymphatic metastasis. Biochem Biophysic Res Commun. 2010;396:571–577. doi: 10.1016/j.bbrc.2010.04.152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wang LJ, Zhao Y, Han B, Ma YG, Zhang J, Yang DM, Mao JW, Tang FT, Li WD, Yang Y, Wang R, Geng JG. Targeting Slit-Roundabout signaling inhibits tumor angiogenesis in chemical-induced squamous cell carcinogenesis. Cancer Sci. 2008;99:510–517. doi: 10.1111/j.1349-7006.2007.00721.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tie J, Pan Y, Zhao L, Wu K, Liu J, Sun S, Guo XG, Wang B, Gang Y, Zhang YG, Qiao TD, Zhao QC, Nie YZ, Fan DM. MIR-218 inhibits invasion and metastasis of gastric cancer by targeting the Robo1 receptor. PLoS Gen. 2010;6:e1000879. doi: 10.1371/journal.pgen.1000879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ma S, Liu X, Geng JG, Giuo SW. Increased SLIT immunoreactivity as a biomarker for recurrence in endometrial carcinoma. Am J Obst Gynecol. 2010;202:68.e1–68.e11. doi: 10.1016/j.ajog.2009.07.040. [DOI] [PubMed] [Google Scholar]
- 15.Mimmi S Ballard, Hink L. A roundabout way to cancer. Adv Cancer Res. 2012;114:187–235. doi: 10.1016/B978-0-12-386503-8.00005-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Narayan G, Goparaju G, Arias-Pulido H, Kaufman AN, Schneider A, Dürst M, Mansukhani M, Pothuri B, Murty VV. Promotor hypermethylation-mediated inactivation of multiple Slit-Robo pathway genes in cervical cancer progression. Mol Cancer. 2006;5:1–10. doi: 10.1186/1476-4598-5-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mano Y, Aishima S, Fukuhara T, Tanaka Y, Kubo Y, Motomura T, Toshima T, Iguchi T, Shirabe K, Maehara Y, Oda Y. Decreased roundabout 1 expression promotes development of intrahepatic cholangiocarcinoma. Human Pathol. 2013;44:2419–2426. doi: 10.1016/j.humpath.2013.03.022. [DOI] [PubMed] [Google Scholar]
- 18.Sundaresan V, Chung G, Heppell-Parton A, Xiong J, Grundy Y, Roberts Y, James L, Cahn A, Bench A, Douglas J, Minna J, Sekido Y, Lerman M, Latif L, Bergh J, Li H, Lower N, Ogilvie D, Rabbitts P. Homozygoud deletion at 3p12 in breast and lung cancer. Oncogene. 1998;17:1723–1729. doi: 10.1038/sj.onc.1202103. [DOI] [PubMed] [Google Scholar]
- 19.Xian J, Clark C, Fordam R, Pannell R, Rabbitts TH, Rabbitts PH. Inadequate lung development and bronchial hyperplasia in mice with a targeted deletion in the DUTT1/Robo1 gene. PNAS. 2001;98:15062–15066. doi: 10.1073/pnas.251407098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xian J, Aitchison A, Bobrow L, Corbett G, Pannel R, Rabbitts T, Rabbitts P. Targeted disruption of the 3p12 gene, DUTT1/Robo1, predisposes mice to lung adenocarcinomas and lymphomas with methylation of the gene promotor. Cancer Res. 2004;64:6432–6437. doi: 10.1158/0008-5472.CAN-04-2561. [DOI] [PubMed] [Google Scholar]
- 21.Dallol A, Forgacs E, Martinez A, Sekido Y, Walker R, Kishida T, Rabbitts P, Maher ER, Minna JD, Latif L. Tumour specific promotor region methylation of the human homologue of the Drosophila roundabout gene DUTT1 (ROBO1) in human cancers. Oncogene. 2002;21:3020–3028. doi: 10.1038/sj.onc.1205421. [DOI] [PubMed] [Google Scholar]
- 22.Ito H, Funahashi S, Yamauchi N, Shibahara J, Midorikawa Y, Kawai S, Kinoshita Y, Watanabe A, Hippo Y, Ohtomo T, Iwanari H, Nakajima A, Makuuchi M, Fukayama M, Hirata Y, Hamakubo T, Kodama T, tsuchiya M, Aburatani H. Identification of ROBO1 as a novel hepatocellular carcinoma antigen and a potential therapeutic and diagnostic target. Clin Cancer Res. 2006;12:3257–3264. doi: 10.1158/1078-0432.CCR-05-2787. [DOI] [PubMed] [Google Scholar]
- 23.Prasad A, Fernandez AZ, Rao Y, Ganju RK. Mechanisms of signal transduction: Slit-protein mediated inhibition of CXCR4-induced chemotactic and chemoinvasive signaling pathways in breast cancer cells. J Biol Chem. 2004;279:9115–9124. doi: 10.1074/jbc.M308083200. [DOI] [PubMed] [Google Scholar]
- 24.Latil A, Chene L, Cochant-Priollet B, Mangin P, Fournier G, Berthon P, Cussenot O. Quantification of expression of netrins, Slits, and their receptors in human prostate tumors. Int J Cancer. 2003;103:306–315. doi: 10.1002/ijc.10821. [DOI] [PubMed] [Google Scholar]
- 25.Gröne J, Doebler O, Loddenkemper C, Hotz B, Buhr HJ, Bhargava S. Robo1/Robo4: differential expression of angiogenic markers in colorectal cancer. Oncol Rep. 2006;15:1437–1443. [PubMed] [Google Scholar]
- 26.Folkman J. Angiogenesis. Ann Rev Med. 2006;57:1–18. doi: 10.1146/annurev.med.57.121304.131306. [DOI] [PubMed] [Google Scholar]
- 27.Folkman J. Tumor angiogenesis: therapeutic implications. New Eng J Med. 1971;18:1182–1186. doi: 10.1056/NEJM197111182852108. [DOI] [PubMed] [Google Scholar]
- 28.Wang B, Xiao Y, Ding BB, Zhang N, Yuan XB, Gui L, Qian KX, Duan S, Chen ZJ, Rao Y, Geng JG. Induction of tumor angiogenesis by Slit-Robo signaling and inhibition of cancer growth by blocking Robo activity. Cancer Cell. 2003;4:19–29. doi: 10.1016/s1535-6108(03)00164-8. [DOI] [PubMed] [Google Scholar]
- 29.Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996;86:353–364. doi: 10.1016/s0092-8674(00)80108-7. [DOI] [PubMed] [Google Scholar]
- 30.Enomoto S, Mitsui K, Kawamura T, Iwanari H, Daigo K, Horiuchi K, Minami T, Kodama T, Hamakubo T. Suppression of Slit2/Robo1 mediated HUVEC migration by Robo4. Biochem Biophys Res Comm. 2016;469:797–802. doi: 10.1016/j.bbrc.2015.12.075. [DOI] [PubMed] [Google Scholar]
- 31.Folkman J. Antiangiogenesis in cancer therapy-endostatin and its mechanisms of action. Experi Cell Res. 2006;312:594–607. doi: 10.1016/j.yexcr.2005.11.015. [DOI] [PubMed] [Google Scholar]
- 32.Holash J, Thurston G, Rudge JS, Yancopoulos GD, Adjei AA, Bergers J, Pytowski B, Pegram S, Gordon MS. Inhibitors of growth factor receptors, signaling pathways and angiogenesis as therapeutic molecular agents. Cancer Metastasis Rev. 2006;25:243–252. doi: 10.1007/s10555-006-8504-6. [DOI] [PubMed] [Google Scholar]
- 33.Jain RK, Duda DG, Clark JW, Loeffler JS. Lessons for phase III clinical trials on anti-VEGF therapy for cancer. Nature Clin Pract. 2006;3:24–39. doi: 10.1038/ncponc0403. [DOI] [PubMed] [Google Scholar]
- 34.Zhang XY, Dong JQ, He Y, Zhao M, Liu Z, Wang N, Jiang MZ, Zhe Z, Gang L, Liu HM, Nie YZ, Fan DM, Tie J. miR-218 inhibited tumor angiogenesis by targeting ROBO1 in gastric cancer. Gene. 2017;615:42–49. doi: 10.1016/j.gene.2017.03.022. [DOI] [PubMed] [Google Scholar]
- 35.Shi R, Liu W, Liu B, Xu Z, Chen L, Zhang Z. Slit2 expression and its correlation with subcellular localization of β-catenin in gastric cancer. Oncol Rep. 2013;30:1883–1889. doi: 10.3892/or.2013.2662. [DOI] [PubMed] [Google Scholar]
- 36.Zhang QQ, Zhou D, Lei Y, Zheng L, Chen SX, Gou HJ, Gu QL, He XD, Lan T, Qi CL, Li JC, Ding YQ, Qiao L, Wang LJ. Slit2/Robo1 signaling promote intestinal tumorigenesis through Src-mediated activation of the Wnt/β-catenin pathway. Oncotarget. 2014;6:3123–3135. doi: 10.18632/oncotarget.3060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nagashima M, Asano G, Yoshino S. Imbalance in production between vascular endothelial growth factor and endostatin in patients with rheumatoid arthritis. J Rheumatol. 2000;27:2339–2342. [PubMed] [Google Scholar]
- 38.Antelli F, Chuang ST, Yang XJ, Wang HL. Value of Glypican 3 immunostaining in the diagnosis of hepatocellular carcinoma on needle biopsy. Am J Clin Pathol. 2008;130:219–223. doi: 10.1309/WMB5PX57Y4P8QCTY. [DOI] [PubMed] [Google Scholar]