

ABSTRACT
Inflammation is a hallmark of HIV infection. Among the multiple stimuli that can induce inflammation in untreated infection, ongoing viral replication is a primary driver. After initiation of effective combined antiretroviral therapy (cART), HIV replication is drastically reduced or halted. However, even virologically controlled patients may continue to have abnormal levels of inflammation. A number of factors have been proposed to cause inflammation in HIV infection: among others, residual (low-level) HIV replication, production of HIV protein or RNA in the absence of replication, microbial translocation from the gut to the circulation, co-infections, and loss of immunoregulatory responses. Importantly, chronic inflammation in HIV-infected individuals increases the risk for a number of non-infectious co-morbidities, including cancer and cardiovascular disease. Thus, achieving a better understanding of the underlying mechanisms of HIV-associated inflammation in the presence of cART is of utmost importance. Extracellular vesicles have emerged as novel actors in intercellular communication, involved in a myriad of physiological and pathological processes, including inflammation. In this review, we will discuss the role of extracellular vesicles in the pathogenesis of HIV infection, with particular emphasis on their role as inducers of chronic inflammation.
KEYWORDS: Extracellular vesicles, inflammation, HIV, pro-inflammatory cytokines, exosomes, HIF-1α
Introduction
Inflammation and HIV infection
Human Immunodeficiency Virus type 1 (HIV) continues to threaten global health, with 1.8 million new infections diagnosed in 2017 [1]. Disruption of the HIV viral replication cycle by a combination of drugs (combined antiretroviral therapy, or cART) reduces plasma HIV RNA levels to below the limit of detection of commercial assays (<50 copies/mL) [2,3]. Moreover, in most patients, cART can also restore CD4 + T cell counts in blood, thus preventing manifestation of acquired immunodeficiency syndrome (AIDS). However, immune dysfunction and chronic inflammation are not completely resolved during cART.
Inflammation is a defence response to invading pathogens or endogenous danger signals that contains and helps to eradicate threats to the organism. However, if inflammation goes unchecked or persists in time, it can result in pathology. During HIV infection, even in cART-treated (virally suppressed) individuals, parameters of inflammation such as C-reactive protein [4] and the pro-inflammatory cytokines interleukin (IL)-6 [4], TNF-α and BAFF [5] remain elevated for long periods of time. These parameters are predictors of a series of clinical conditions collectively known as serious non-AIDS events (SNAEs) [6], including cardiovascular disease [7], cancer [8], neurological disease [9], liver disease [10] and overall mortality [11]. Therefore, understanding the mechanisms underlying persistent inflammation could lead to the identification of new targets to reduce the increased risk of SNAEs, and thus is a priority in HIV research.
Several mechanisms have been proposed to explain persistent inflammation during HIV infection. A widely accepted mechanism is the increase in intestinal permeability due to the loss of mucosal CD4 + T cells during early infection with HIV. This results in the translocation of microbial products, such as lipopolysaccharides (LPS), to the circulation, which then stimulates monocytes/macrophages to secrete pro-inflammatory molecules [12]. Ongoing HIV replication and exposure to other microorganisms, like human cytomegalovirus, may also contribute to persistent immune activation and inflammation [13,14]. Furthermore, loss of immunoregulatory responses [15] and lymphoid fibrosis [16] have also been associated with markers of inflammation and morbidity. Evidence collected during the last few years suggests that Extracellular Vesicles play important functions in inflammation, and this will be the central topic of this Review.
Intercellular communication mediated by EVs
The term Extracellular Vesicle (EV) refers to structures limited by a lipid bilayer, released by cells into their extracellular environment, and that contain various components of the cell of origin. Different types of EVs have been described, based on their intracellular biogenesis location [17]. Exosomes are small EVs (approximately 50–150 nm in diameter) of endosomal origin that initially form as intraluminal vesicles inside late endosomal compartments. They are secreted following fusion of the endosomal limiting membrane with the plasma membrane. Other types of EVs, generally called ectosomes, microvesicles or microparticles, are secreted by direct budding from the cellular plasma membrane, which results in EVs of various diameters: as small as exosomes (<150 nm) or larger, up to a few micrometres [18]. Whatever their initial site of formation, plasma membrane or endosomes, EVs have the same topological orientation of their limiting membrane as the cell itself: they contain cytosolic components and they expose the extracellular domain of transmembrane proteins inserted into their lipid bilayer. This structure is identical to that of enveloped viruses such as HIV, which can thus be considered as a particular type of EV.
EVs are not a mere reflection of the cell from where they originate, but they have their own composition and, therefore, new functions. Indeed, EVs contain only a selection of the proteins of the secreting cell, mainly from the plasma membrane, endosomal compartments, and cytosolic components, although some large EVs may also include proteins from other intracellular compartments, such as endoplasmic reticulum or nucleus [19,20]. Moreover, EVs may contain specific repertoires of mRNAs and non-coding RNAs, suggesting the existence of mechanisms that control the sorting of these molecules, although these mechanisms are still unclear [21].
It is generally accepted that following secretion, EVs mediate the delivery to a target cell of nucleic acids, lipids and proteins that are expressed on the surface or contained within the EV lumen, although the actual mechanisms of such delivery are still poorly understood [18]. Irrespective of the mechanism, it is clear that EVs can modulate the function of recipient cells, inducing or suppressing different pathways in physiological and pathological processes [22].
Role of EVs in immunity
EVs mediate communication between cells of the immune system, participating in the regulation of immune responses both in normal physiology and pathological conditions [23,24]. Indeed, EVs can modulate different aspects and steps of adaptive immune responses, including antigen presentation [25,26], T cell polarization [27] and T cell function [28,29]. In pathological conditions, it has been shown that EVs can participate in the pathogenesis of autoimmune diseases [30]. In addition, EVs participate in innate immune responses, and in particular in inflammation. It has been shown that EVs secreted both by pathogens [31–34] and infected cells [35,36] carry pathogen-associated molecular patterns (PAMPs) and, therefore, trigger inflammation upon interaction with innate immune cells. Moreover, EVs containing danger signals released from stressed or injured cells contribute to inflammation induction and persistence [37,38]. Additionally, EVs containing cytokines [39–42], enzymes involved in the biosynthesis of lipid mediators [43,44] and other chemotactic signals [45] can contribute to the propagation of inflammation.
Role of EVs in viral infections
EVs play diverse and important roles during viral infections, leading to both pro- and anti-viral effects. On the one hand, EVs have been shown to contribute to viral transmission and spread from infected to uninfected cells, mainly by masking viral antigens and PAMPs, thus avoiding immune recognition [46–49]. Remarkably, viruses usually considered as “naked” or non-enveloped, such as hepatitis A virus (HAV), some members of the Enterovirus genus, rotavirus, norovirus and picornavirus were shown to be released from cells cloaked in host-derived membranes [50–53], facilitating the delivery of a high level of inoculum to the receiving host.
In addition to their role in the promotion of viral transmission, EVs released during viral infections could contribute to viral replication by dampening innate antiviral mechanisms, such as natural killer (NK)-cell functions [47,54,55] and type I interferon responses [56]. Moreover, EVs can impair adaptive antiviral immune responses by suppressing Th1 responses [57] or by promoting Treg cell expansion [58].
In contrast to the pro-viral effects mentioned above, EVs produced by infected cells or by bystander cells have also been shown to be able to restrict viral replication by triggering or amplifying host antiviral responses. Along these lines, it has been shown that EVs containing HAV [59] and hepatitis C virus RNA [60] activate type I interferon responses following uptake by uninfected plasmacytoid dendritic cells. Likewise, EVs released by latent Epstein Barr Virus-infected B cells activated non-infected dendritic cells (DCs) [61]. In addition, intercellular transmission of cellular antiviral factors by EVs can also constitute a mechanism for generating an interferon-induced antiviral response in non-infected cells, as was observed for hepatitis B virus [62] and herpes simplex virus-1 [63] infections. Interestingly, EVs from uninfected cells can also restrict viral replication. For instance, it has been shown that uninfected primary human trophoblasts, which are highly resistant to viral infection, can confer EV-mediated antiviral resistance to different target cells by inducing the autophagy pathway [64].
In sum, in the complex biology of viral infections, EVs constitute an intercellular communication mechanism used both by virus and host, intermingling antiviral and inflammatory responses with viral spread. In this sense, understanding the role played by EVs in HIV infection remains a crucial research goal that will allow us to gain knowledge on HIV pathogenesis and to contribute to patient care and treatment strategies.
Considerations regarding limitations in techniques for separating EVs from virions and for EV isolation from body fluids
HIV is released from infected cells by a process that in many aspects resembles the formation of exosomes and other EVs in non-infected cells. It was thus proposed already 16 years ago that HIV hijacks the exosome biogenesis pathway, to hide from the host immune system by adopting a structure and composition similar to innocuous endogenous EVs [65]. Similarity of structure and biophysical properties of small EVs and HIV (similar diameters of 50–150 nm, similar densities by flotation in sucrose) makes their separation difficult, and it is now clearly recognized that they are co-purified by standard virus or exosomal purification protocols (differential ultracentrifugation and/or sucrose cushions) [66,67]. Two techniques have been used so far to separate endogenous EVs from HIV: immuno-depletion of the former by anti-CD45 antibodies, since CD45 was described as excluded from the virus but present in EVs [68], or separation by pelleting into iodixanol gradients (commercially known as OptiPrepTM), where the virus reaches the bottom, dense fractions faster than endogenous EVs [69]. However, both approaches have limitations. Indeed, we recently observed that, in the T lymphoid cell line Jurkat, CD45 is mainly released in large EVs, and hardly present in small EVs. Thus, the utility of CD45 to separate small EVs and/or exosomes from HIV is limited (LMJ, CT, unpublished results). As for the velocity gradient, even though HIV is indeed mainly recovered in the bottom dense fractions, we recently observed that endogenous EVs are only slightly “lighter” than the virus, and are recovered in overlapping fractions of such gradient. In addition, by many different separation approaches, we could show that acetylcholinesterase (AChE), which was previously proposed to identify EVs specifically [69], is in fact mainly present in small non-EV structures recovered in the light fractions of a velocity gradient, separated from the overlapping EVs and virions [70]. Thus, some of the previous attempts at analysing the functions of EVs in the HIV life cycle (described below in Sections 2 and 3), may have instead established functions of either co-isolated large EVs (CD45+) or non-vesicular structures (AChE+), leaving open the question of a role of small EVs and/or exosomes.
In addition, isolation of vesicles from complex biological samples, such as blood, poses an extra challenge due to the presence of other materials that could diminish EVs preparations purity. That is the case of plasma samples, which contain large amounts of lipoproteins that could be co-isolated with EVs. Moreover, large proteins and protein aggregates formed in the plasma also co-precipitate with EVs following ultracentrifugation. EV precipitation by polymers (for instance, by using the commercial kit ExoQuickTM) is not specific for EVs, and precipitated material certainly includes non-vesicular components [71]. It is generally assumed that size exclusion chromatography (SEC) (probably complemented with density gradient fractionation) is one of the best options available to obtain highly pure EVs from plasma [72,73]. Therefore, caution is required when interpreting data obtained for plasma-purified EVs, since without rigorous demonstration of EV purity and assessment of non-EV fractions as controls, it is difficult to argue that a detected activity is EV-borne and not related to soluble or non-EV components [71].
Finally, it is important to stress that given the difficulty of analysing the in vivo effect of EVs, together with the lack of experimental models of HIV infection other than the primate models, most studies published on EVs and HIV infection were performed either in vitro or ex vivo. Thus, evidence supporting in vivo relevance is scarce.
In the following paragraphs we review the current knowledge of the functions reported for EVs in the context of HIV infection. Many of the studies used separation methods that we now know are not specific for EVs. We will nonetheless describe such studies as they were presented at the time of their publication. We will focus especially on the immunomodulatory effects of EVs and introduce a possible link between immune activation and cellular metabolism.
Implications of EVs for HIV infection
Effect of EVs on HIV replication in T cells and macrophages
Using the available, imperfect EV and HIV separation approaches, some groups have reported different effects of EVs and the co-isolated HIV particles on either infectivity or immune responses. In this section, we will briefly discuss the inhibitory and stimulatory roles of EVs on HIV infection. A comprehensive review of these mechanisms has been recently published elsewhere [74]. It is striking that EVs can play both promoting and inhibitory roles in HIV replication. Although the reasons for these discrepancies are not clear, they likely reflect the fact that EVs of different size, biogenesis and composition are used in the different studies. The existence of these incongruities further highlights the need to clearly characterize the EVs used in each study and the purification method used to obtain them.
Inhibitory role of EVs during HIV infection
Different mechanisms have been proposed to explain the observation that EVs produced by HIV-infected cells are able to inhibit HIV infection of target cells. For instance, it has been shown that CD4+ EVs produced by a CD4 + T cell line can inhibit the infectivity of HIV particles by binding to the viral envelope protein (gp120), which would thus not be available to bind to the CD4 receptor expressed on target cells [75]. In addition, EVs can transfer antiviral effector molecules to the recipient cells, thus blocking viral infection or restricting viral propagation. Along these lines, it has been postulated that infected cells can transfer family members of the host restriction factor APOBEC3, thus conferring resistance to HIV infection to bystander uninfected T cells [76]. However, in this study, the presence of other factors in EVs that contribute to HIV inhibition cannot be ruled out. Finally, Haque et al studied the role of EVs produced by monocytic cells exposed to cigarette smoke condensate (CSC) on HIV-1 replication, showing that CSC-treated monocytes produce EVs with increased anti-HIV activity, likely due to modified incorporation of antioxidant molecules into EVs [77].
EVs have also been implicated as effectors of the adaptive anti-viral immune response. For instance, it has been shown that EVs secreted by CD8 + T cells display potent non-cytotoxic antiretroviral activity by inhibiting HIV transcription [78].
Concerning the effect of EVs purified from body fluids, it has been shown that EVs isolated from semen, breast milk and vaginal fluid have an inhibitory effect on HIV infection [79–84]. Interestingly, blood serum-derived exosomes, although internalized by target cells, were not able to block HIV replication [80,83].
Promotion of HIV infection/replication by EVs
Another set of studies reported the opposite effect of EVs on HIV infection, showing that they can act as facilitators of viral transmission and replication. A distinctive mechanism by which EVs positively regulate HIV infection is by mediating the transfer of the HIV co-receptors CXCR4 and CCR5 to target cells that do not express these molecules or that express them at low levels, thus expanding the number of susceptible target cells [85,86]. Another mechanism that facilitates HIV transmission is the EV-mediated induction of maturation of both HIV-infected and bystander non-infected DCs. Maturation of DCs is accompanied by an increase of the adhesion molecule ICAM-1, which in turn facilitates interaction of DCs with CD4 + T cells. Consequently, lymphocyte activation is promoted, and transfer of the virus from DCs to CD4 + T cells is enhanced [87]. Another pro-viral effect has been described for macrophage-derived EVs, which have been shown to capture viral particles in clusters, thus engaging virus-cell interaction and facilitating the transfer of HIV to neighbouring uninfected cells [88]. Finally, an indirect mechanism through which EVs promote HIV replication is by mediating the transfer of HIV-derived proteins or nucleic acids that promote CD4 + T cell activation, thus rendering them more permissive to viral infection [89–92].
Effect of EVs on HIV latency/reactivation
While cART typically leads to the reduction of viral load to undetectable levels, a relatively small but stable reservoir of latently infected cells persists life-long in HIV-infected individuals [93]. This reservoir is fully capable of producing infectious virus when the host cell is reactivated by antigens, cytokines or when cART is discontinued. A series of studies have shown that EVs either from uninfected [94] or HIV-infected [95,96] cells can reactivate latent HIV infection through different mechanisms. In contrast, recent evidence from our group has demonstrated an alternative possibility of reversing HIV latency by withdrawing EVs from latently infected U1 and ACH2 cell lines [97]. Under EV-depleted conditions, these two cell lines underwent viral reactivation, and further functional analyses indicated that alterations in cellular lipid production could be a potential underlying mechanism for this reactivation. In the same study, we demonstrated that HIV virions produced in the EV-depleted condition were more infectious, and cells cultured in this depleted condition were more susceptible to infection [97]. We speculate that the discrepancy between our observation and the studies that analysed the role of EVs produced by infected cells on HIV latency reactivation could be due to the presence of viral factors, especially Tat, in the EV preparations. Altogether, these studies suggest that manipulating EVs in the cellular environment, or modulating cellular recognition of EVs, could be used to affect HIV latency.
In summary, these studies using different experimental systems and EVs purified by different methodologies show that vesicles can both inhibit and promote HIV replication. The use of physiologically-relevant models and of EVs obtained by standardized methodologies will help to clarify the role of EVs in HIV infection in vivo.
EVs and inflammation during HIV infection
As in other chronic inflammatory conditions, such as rheumatoid arthritis [30,98] and cardiovascular disease [99], there is an increasing amount of evidence suggesting that EVs constitute potential contributors to chronic inflammation during HIV infection.
Circulating EVs in HIV patients
EVs have been reported to be more abundant in the plasma of HIV patients as compared to non-infected individuals [100–102]. However, since different isolation and quantification methods – not all of them specific for EVs or properly validated – were used in these reports, these data should be interpreted with caution.
In the work by Lee et al., plasma from viraemic and non-viraemic HIV infected patients as well as from healthy donors was pooled, and EVs were purified by differential centrifugation followed by sucrose gradient. Comparison of EVs purified from the plasma of HIV-infected subjects with those from uninfected controls revealed that infected individuals, irrespective of virological status, had an 18- to 22-fold increase in total protein content. Nanoparticle Tracking Analysis (NTA) revealed a similar increase (median 17.9-fold) in the number of particles for non-viraemic HIV-infected patients versus healthy controls [100]. Preliminary data from our laboratory (PSP, AR, MO, unpublished results) also showed higher total protein content (1.6 fold) in plasma EVs from HIV-infected individuals purified by SEC followed by centrifugation, as compared with uninfected controls. In addition, in longitudinal samples from a well-established simian immunodeficiency virus (SIV)/macaque model [103], SEC-separated EVs increased by around 50% from before infection to acute-phase infection, declined back to baseline during antiretroviral treatment, and experienced a transient and slight (<50%) increase during treatment interruption (YH and KWW, unpublished data). These longitudinal results suggest the possibility that at least some of the reported increases in circulating EVs, particles, or protein are cohort-specific.
Abundance of EVs in HIV patients was also observed in other studies where plasma EVs were isolated by EV precipitation [101,102]. These studies also arrived at the conclusion that HIV-infected individuals have higher numbers of circulating EVs. However, due to the methodology used in these studies, it is likely that non-vesicular material was also present in the preparations analysed.
Cellular origin of circulating EVs
Cellular origin of EVs circulating in plasma of HIV-infected individuals is uncertain. A mass spectrometry analysis of highly purified EVs (isolated from healthy donor plasma with a combination of iodixanol density cushion and SEC), detected mostly cell markers for erythrocytes (CD235a) and platelets (CD41, CD61, CD62p) [73], thus suggesting that erythrocyte- and platelet-derived EVs would account for the majority of plasma vesicles. Nevertheless, contribution of other cell types such as endothelial cells or lymphocytes to the mixture of circulating EVs is also likely, as evidenced by other analysis methods such as multiplexed bead-based flow cytometry [104]. Additionally, circulating EVs could be generated from a broad spectrum of cells, including those of the immune system, upon activation, stress or apoptotic death.
It has been proposed that HIV-infected T cells release increased amounts of EVs, an effect mainly due to the accessory protein Nef. Indeed, Nef expression was shown to stimulate the release of microvesicle clusters by HIV-infected cell lines and primary CD4 + T cells from HIV-infected individuals [105] as well as from in vitro infected CD4 + T cells [89]. However, the percentage of infected CD4 + T cells in HIV+ individuals is too small to account for any increase in plasma EVs, even if EV secretion is promoted by Nef.
As mentioned above, a hallmark of HIV infection is chronic immune activation and inflammation, which are maintained by increased levels of circulating LPS, among other mechanisms. Remarkably, pro-inflammatory cytokines and LPS promote EV release from dendritic cells [106,107] and endothelial cells [108]. We thus speculate that in HIV-infected patients, any increase in the number of circulating EVs comes from a broad population of persistently activated immune cells, instead of HIV-infected cells only. Our unpublished data from the SIV/macaque model mentioned above are supportive of this notion, showing that during treatment interruption, when only a small number of cells are infected, the absolute increase in EVs is much greater than the number of released viruses. Also supporting this idea, the afore-mentioned study by Lee et al. suggested a myeloid/macrophage-like cell population origin of plasma EVs during HIV infection, according to the cytokine cargo pattern (e.g. IL-12p40, GRO, NAP-2) [100]. Despite this indirect evidence, whether activated immune cells contribute to the increase in circulating EVs observed in HIV patients in vivo is an interesting question that needs further investigation.
Pro-inflammatory effects of HIV-related EVs
Several studies suggest that EVs produced during HIV infection are capable of stimulating inflammation. For instance, EVs purified by immunoaffinity from the plasma of HIV-positive subjects incubated with the monocytic cell line THP-1 exerted transcriptional activation of genes implicated in interferon responses, cytokines/chemokines, immune activation, innate immune responses, and inflammation [101]. Another study also reported that EVs isolated with ExoQuick from conditioned medium of HIV-infected U1 cell line or from serum from HIV+ infected individuals induced TNFα release by THP-1 cells [109].
TNFα secretion induced by EVs produced during HIV infection has also been reported by Lee et al. who, proposed that Nef expression in HEK-293T cells activated the TNF-converting enzyme ADAM17 and shuttled it into EVs, which, following uptake by recipient cells, induced pro-TNF cleavage in endosomal compartments [110]. The same group later described that ADAM17+ EVs obtained by differential centrifugation from the plasma of cART-treated patients induced the cleavage of endosomal pro-TNF and its subsequent secretion in PBMCs and macrophages [111].
Our group has recently demonstrated that EVs released by HIV-infected primary CD4 + T cells promote pro-inflammatory cytokine production by recipient CD4 + T cells and monocyte-derived macrophages [112]. In addition, plasma EVs from HIV-infected individuals isolated by SEC followed by centrifugation triggered the release of pro-inflammatory cytokines (IL-6, IL1-β and TNF-α) by primary monocyte-derived macrophages [112]. Our results indicate that EV-mediated inflammation during HIV infection is coordinated with the activity of the transcription factor HIF-1α (see below).
Pro-inflammatory cargoes in HIV-EVs
The repertoire of molecules that can be loaded into EVs is vast, and both viral and host elements could contribute to the development of inflammation.
Virus-derived pro-inflammatory cargos
The convergence of HIV and EV biogenesis suggests that HIV products, including RNA and proteins may be packed within EVs. HIV elements present in EVs could then contribute to stimulation of immune cells.
As stated above, the viral protein Nef from both human and simian immunodeficiency virus has been reported to be incorporated into EVs [89,105,113]. Moreover, almost all Nef in the plasma of HIV patients was found to be associated with CD45+ EVs [114]. Interestingly, Nef-containing EVs were found even in the plasma of HIV-infected individuals under antiretroviral therapy and with undetectable viral loads [100,114]. The main effect reported for Nef+ EVs on recipient cells has been the promotion of apoptosis in primary CD4 + T cells [89] and cell lines [114], which could contribute to CD4 + T cell depletion. Moreover, as described above, Nef+ EVs promote TNF-α release by recipient cells [111]. It should be noted, though, that some have questioned EV incorporation of Nef [115], indicating that further studies may be required.
Regarding viral RNA cargo, unspliced HIV genomic sequences have been identified in EVs obtained by iodixanol gradient from chronically infected U937 cells [116]. Moreover, viral small RNAs have been described in EVs released by HIV-infected cells, also obtained by iodixanol gradient. Trans-activation response (TAR) RNA was found in EVs derived from infected Jurkat and primary T cells, predominantly in a stem-loop form [92]. The same group later demonstrated that exosomal TAR RNA from HIV-infected cells was able to modulate the gene expression of pro-inflammatory cytokines, such as IL-6 and TNF-β, in human primary macrophages, through a Toll-like receptor (TLR) 3-dependent pathway [117]. TAR and other small RNAs (called by the authors viral miRNAs: vmiR88 and vmiR99) were also detected in EV-containing releasate of U1 and infected primary alveolar macrophages, and in sera from HIV+ individuals precipitated by ExoQuick [109]. Interestingly, in this work, vmiR88 and vmiR99, but not TAR, stimulated human macrophage TNFα release in a TLR8-dependent way. (See, however, discussion of these mechanisms below.)
Host-derived pro-inflammatory cargoes
EVs produced during HIV infection have been reported to contain multiple host-derived inflammatory mediators. Konadu et al. described that levels of 21 cytokines and chemokines, such as IL-1α, TNF-α and CXCL10, were significantly higher in plasma EVs isolated by iodixanol gradient from HIV-infected patients (n = 15) than in those from non-infected subjects (n = 10) [118]. Another study analysed plasma EVs isolated by differential centrifugation plus sucrose gradient, showing that whereas EVs from healthy donors contained only 3 cytokines/chemokines/soluble factors from a 120-protein array, those from HIV-infected individuals harboured as many as 23 [100]. Moreover, although EVs from both non-viraemic and viraemic patients exhibited a similar pattern of protein expression, viraemic patients with low CD4 counts harboured more and higher levels of pro-inflammatory factors, including e.g. TNFα, IL-12p40, sIL-6R, sTNF-RI, GRO [100]. Finally, packaging of cytokines and chemokines has also been shown in EVs derived from the plasma of HIV-infected alcohol drinkers and cigarette smokers [119]. Nevertheless, whether these EVs harbouring inflammatory cytokines exert an effect on recipient cells has not been explored.
HIV infection and cellular activation may also induce changes in cell miRNA profiles [120,121], which could be reflected in EV cargo. Indeed, EVs from HIV-infected macrophages obtained by iodixanol gradient were reported to contain 38 miRNAs that were not present in EVs released by uninfected cells and 13 upregulated miRNAs [122], amongst which miR-29a has been described to enhance LPS induced inflammatory response in macrophages [123]. In addition, the pro-inflammatory miRNAs miR-155 and miR-223 were found in EV-containing material precipitated by ExoQuick from the plasma of ART-naive HIV patients [102].
Altogether, data presented in this section reveal EVs under the spotlight of possible mechanisms related to the sustained inflammation in people living with HIV. A working model depicting the role of EVs in HIV pathogenesis is presented in Figure 1. Whether EVs are implicated in the development of SNAES is an open question that needs further investigation.
Figure 1.
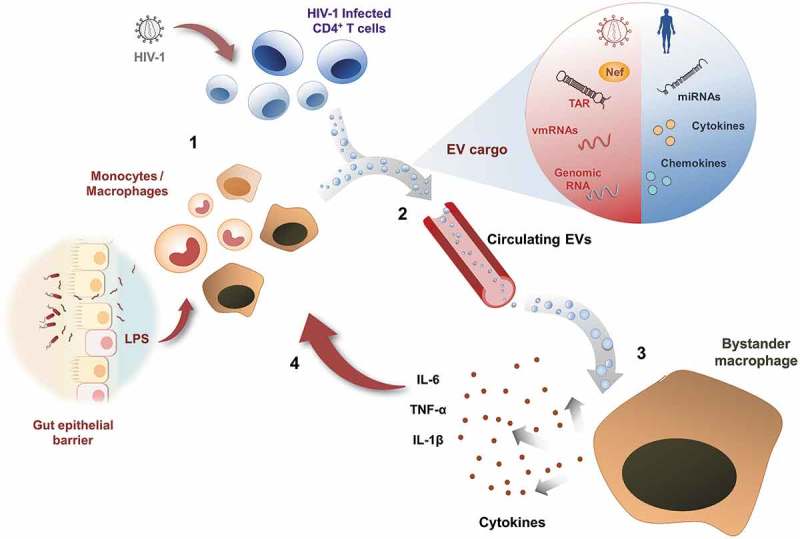
Model of chronic inflammation enhanced by extracellular vesicles.
HIV replication or expression of viral components in latently infected cells, in conjunction with bacterial PAMPs released into circulation as a consequence of microbial translocation in the gut, constitute a persistent activating stimuli for immune cells such as T lymphocytes, monocytes and macrophages [1]. These activated cells release into circulation EVs containing pro-inflammatory molecules from the host or HIV-derived PAMPs (as detailed in the enlarged vesicle) [2]. Macrophages exposed to circulating EVs become activated and release inflammatory cytokines [3], which in turn contribute to a positive loop of systemic chronic inflammation [4].
HIF-1α and EV production and function
HIF-1α is a transcription factor that was originally described for its function in controlling the cellular response to hypoxia [124]. However, further investigation on the functions of HIF-1α revealed that it is also associated with development of inflammatory responses. Indeed, PAMP recognition by myeloid cells activates the HIF-1α pathway, which plays a key role in regulating cell aggregation, motility, invasiveness, and bacterial killing [125]. Furthermore, it has been reported that HIF-1α promotes the transcription of IL-1β [126], thus directly contributing to the development of inflammation. Moreover, HIF-1α was shown to play a critical role in the induction of trained immunity (the acquisition of enhanced capability to respond to certain stimuli by cells of the innate immune system) by fungal components [127]. Thus, HIF-1α is intimately associated with a pro-inflammatory activity in macrophages.
Results recently published by our group show that HIV infection triggers HIF-1α activity in CD4 + T cells, enhancing viral replication [112]. Remarkably, induction of HIF-1α activity is not limited to infected CD4 + T cells because EVs released by these cells, as well as EVs isolated from the plasma of HIV-infected individuals, in turn promote HIF-1α activity in bystander lymphocytes and macrophages. Interestingly, pharmacological inhibition of the EV-mediated induction of HIF-1α activity in bystander macrophages reduces the secretion of the pro-inflammatory cytokines IL-6, IL-1β and TNF-α. Thus, we postulate that during HIV infection, the induction of HIF-1α activity, mediated both by the virus and by EVs, contributes to inflammation.
The coordinated actions of EVs and HIF-1α that we observe during HIV infection are in line with previous studies showing the cross-talk between EVs and HIF-1α activity. For instance, HIF-1α promotes the transcription of two Rab proteins (Rab22a and Rab27a) that control intracellular trafficking routes required for EV secretion [29,128]. Moreover, it has been previously shown that EVs can promote HIF-1α activity in target cells by delivering HIF-1α protein [129,130] or a long non-coding RNA that stabilizes HIF-1α mRNA by preventing its degradation [131]. Based on our results and those from other groups, we propose a model where HIF-1α and EVs coordinately interact to sustain inflammation during HIV infection (Figure 2).
Figure 2.
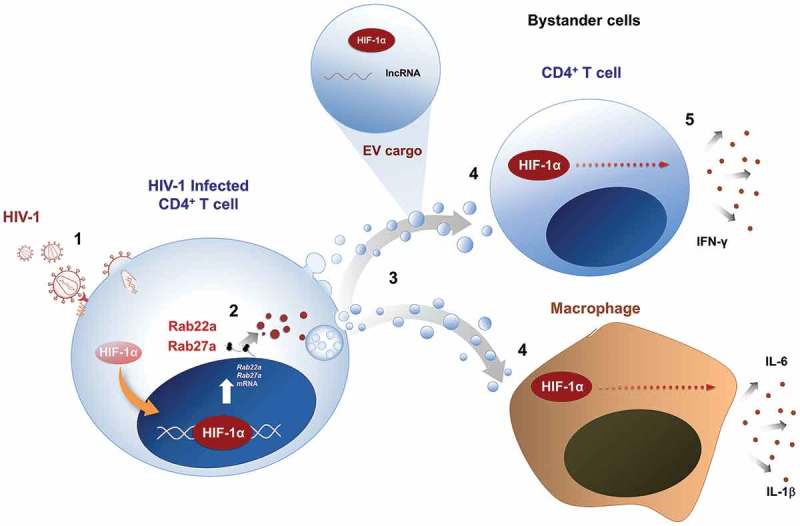
Proposed role of HIF-1α in the inflammatory effects in HIV infection.
HIV-1 infection of CD4 + T cells [1] triggers HIF-1α activity, which promotes the transcription of Rab22a and Rab27a [2], thus favouring intracellular trafficking routes involved in EV secretion [3]. Released EVs, in turn, promote HIF-1α activity in target cells [4], probably by delivering HIF-1α protein, or a lncRNA that stabilizes HIF-1α mRNA by preventing its degradation. Induction of HIF-1α in EV-recipient CD4 + T cells and macrophages is required for secretion of pro-inflammatory cytokines [5]. Thus, HIF-1α and EVs coordinately promote inflammation during HIV infection.
In conclusion, HIF-1α appears to be a key player linking metabolic conditions with inflammatory effects in HIV infection, and it could be a potential pharmacological target to reduce chronic inflammation in HIV patients. However, further studies are necessary to understand the mechanisms by which HIF-1α regulates the production of pro-inflammatory EVs by HIV-infected CD4 + T cells, and the effect provoked by enhanced glycolysis in this process.
Role of EVs in HIV-associated neuroinflammation
Chronic inflammation during HIV infection occurs not only in the periphery, but also in the central nervous system [132]. Entering the brain shortly after infection, usually via infiltrating macrophages [133–136], HIV productively infects microglia and macrophages [137–140]. Astrocytes may also be infected, albeit non-productively [141,142]. The presence of HIV and HIV proteins provokes release of cytokines and reactive oxygen species (ROS) from activated cells, whether infected or not [143–145]. In turn, inflammation contributes to the HIV-associated neurocognitive disorders (HAND): HIV-associated dementia (HAD or HIV-D), mild neurocognitive/motor disorders (MND), and asymptomatic neurocognitive impairment (ANI) [132,146,147]. While HAD, an AIDS-defining condition, has become relatively rare as a result of cART-mediated virologic control, other forms of HAND persist in effectively treated individuals.
Several roles for EVs have been proposed in HIV-associated neuroinflammation. Prominently, glial cells such as astrocytes and microglia have important modulatory functions in neuroinflammation and neurodegenerative diseases [148,149] and communicate with other cells through mechanisms that include EVs. Both harmful and protective effects of glial EVs have been postulated [150,151]. Interestingly, EVs from both glia and neurons also reach the periphery, where they may guide immune responses to brain lesions [152] and serve as biomarkers for CNS disease [153,154]. We will now examine several specific mechanisms attributed to EVs in HIV-associated neuroinflammation.
Dysregulation of glutamine metabolism could lead to up-regulation of EV biogenesis and excitotoxicity
One contribution of EVs to HAND may involve altered glutamate metabolism. Glutamate is the most abundant neurotransmitter in the brain but in excess can induce excitotoxicity [155]. Proinflammatory cytokines such as IL-1β and TNF-α promote EV release from mouse astrocytes [156] and from macrophages and microglia [157], apparently through a glutaminase pathway [158]. Released EVs may also carry and transfer glutaminase [157]. Dysregulation and translocation of glutaminase is associated with elevated glutamate levels within the recipient cell and in the extracellular milieu, driving excitotoxicity and neuronal cell death [155,159]. These interesting findings raise several questions for future studies, such as whether release of glutaminase into the extracellular space is primarily mediated by EVs and what the other potentially pro-inflammatory cargoes of these EVs might be.
EV-associated host and viral small RNAs
Another mechanism for EV effects in the CNS is the transfer of miRNAs. In retroviral infection, Yang et al examined the effects of the HIV Tat protein on astrocytes and discovered that astrocyte-released EVs promoted microglial migration [149], a hallmark of CNS innate inflammatory responses [160]. This effect was mediated by EV transfer of miR-9 both in vitro and in vivo, through post-transcriptional regulation of PTEN [149]. Hu and colleagues reported that a different host miRNA, miR-29a, was transferred by EVs and regulated both HIV Tat and morphine-associated neuronal dysfunction [161]. Finally, Yelamanchili et al found that EVs induced neurotoxicity by shuttling miR-21 to neurons [162]. Specifically, EV miR-21 exerted its effects by activating TLR7 [162].
EVs may also transfer small RNA fragments derived from viral transcripts [92,163]. As discussed above, HIV-infected primary alveolar macrophages were shown to produce EVs containing small viral RNAs that elicited TNF-α release by uninfected human macrophages through TLR8 activation in vitro [109]. Possibly, this could also occur in microglia or macrophages of the CNS. Peripheral EVs bearing these small RNAs might also gain entry into the CNS.
The miRNA and small RNA findings reviewed here raise the possibility of developing small RNA drugs to inhibit or enhance identified small RNAs. However, some of the findings are also puzzling and require additional investigations. For example, in the case of the signalling studies, it is difficult to understand how small RNAs carried inside an EV would engage a toll-like receptor, since the small RNA and the receptor are separated by the EV membrane bilayer. Thus, the EV-associated small RNA should be somehow transported across the membrane, or instead, the miRNA and receptor should remain functional as the EV is degraded by endosome acidification. Alternatively, the effects attributed to EV RNA could actually be due to extra-EV RNA, perhaps enhanced by association with the surface of the EV.
EVs as carriers of HIV proteins and mediators of neurotoxicity
Various HIV proteins, even if not incorporated into virions, can exert pro-inflammatory and cytotoxic effects. Both microglia [164] and astrocytes [165,166] have been reported to release the HIV accessory protein Nef in EVs. EV-associated Nef may then induce neurotoxicity [166], compromise the blood-brain barrier [164], or stimulate release of amyloid beta peptide in vitro [167]. HIV Tat may also be released in EVs and induce neurotoxicity [168]. Importantly, these studies suggest that HIV proteins, produced at low levels during replicative latency, can contribute to chronic inflammation.
In conclusion, EVs appear to contribute to neuroinflammation and neurotoxicity in HIV infection in numerous ways: by influencing metabolism, carrying miRNAs, and transferring HIV proteins even during latent infection.
Conclusions and perspectives
The sustained inflammatory state in HIV-infected individuals is associated with increased morbidity and mortality. It is generally assumed that chronic inflammation underlies a series of pathologies, including neurocognitive disorders, vascular dysfunction and alterations in coagulation state that leads to multimorbidity. Understanding the mechanisms responsible for persistent inflammation, even in patients undertaking cART, is a priority in the field that could lead to the development of new therapeutic interventions to reduce immunopathogenesis.
Increasing evidence suggests that EVs are implicated in the chronic inflammation observed during HIV infection. EVs are more abundant in the plasma of HIV infected individuals. Although it has been proposed that HIV-infected T cells release increased amounts of EVs, it is possible to speculate that a broader population of persistently activated immune cells, such as macrophages, dendritic cells and CD4 + T lymphocytes, could be responsible for any increase in the number of circulating EVs. Further research is needed to corroborate this hypothesis.
EVs isolated from the plasma of HIV-infected patients present pro-inflammatory effects ex vivo, inducing activation of monocytes/macrophages and stimulation of inflammatory cytokines secretion. EVs produced during HIV infection were shown to contain different pro-inflammatory cargoes, both virus- and host- derived, including Nef, viral RNAs, cytokines, chemokines and inflammatory miRNAs. Unfortunately, widespread use of imperfect EV separation techniques and the lack of controls to determine EV purity suggest that caution should be applied when interpreting many of these studies. Moreover, as the EV field evolves and markers of different kinds of vesicles are defined and re-defined, roles originally assigned to small EVs and/or exosomes should be reassessed. Despite these limitations, we consider that there is ample evidence for the notion that circulating EVs have a relevant role in enhancing and maintaining immune activation and inflammation in HIV patients. In addition, EVs appear to contribute to neuroinflammation and neurotoxicity in HIV infection by influencing metabolism, carrying miRNAs, and transferring HIV proteins to the CNS even during latent infection. Nevertheless, whether EVs are also implicated in the development of SNAEs requires further research.
Interestingly, the transcription factor HIF-1α has emerged as a possible key player linking EV-secretion and EV-inflammatory effects in HIV infection. Further studies addressing the mechanisms underlying the association between HIV infection, increase in HIF-1α activity and EV-mediated inflammation will undoubtedly contribute to our understanding of HIV immunopathogenesis, opening a door to the discovery of novel pharmacological targets to reduce chronic inflammation in HIV patients.
Funding Statement
This publication was supported by the USA National Institutes of Health, National Institute on Drug Abuse Grant No. [R01DA040385] (to KWW, CT and MO) and Grant No. [R01DA047807] (to KWW); National Institute of Allergy and Infectious Diseases Grant No. [R01AI144997] (to KWW); [UG3CA241694] (KWW), supported by the NIH Common Fund, through the Office of Strategic Coordination/Office of the NIH Director; by the Argentinean National Agency for Science and Technology Promotion (ANPCYT) under Grant No. [PICT 2015-0658] to MO; by Fondation ARC pour la recherche sur le Cancer [PGA1 RF20180208962], ANRS (2015-1), French National Research Agency [ANR-10-IDEX-0001-02 PSL* and ANR-11-LABX-0043] and SIDACTION [17-1-AAE- 1138] to CT.
Acknowledgments
We thank members of our research groups for technical advice and support, and for critical reading of the manuscript.
Disclosure of interest
The authors report no conflicts of interest. As stated above, this publication was supported by NIH grants R01DA040385, R01DA047807, R01AI144997, and UG3CA241694.
References
- [1].World Health Organization HIV/AIDS [Internet]; 2018. [cited 2019 July22]. Available from: https://www.who.int/en/news-room/fact-sheets/detail/hiv-aids
- [2].Riddler SA, Aga E, Bosch RJ, et al. Continued slow decay of the residual plasma viremia level in HIV-1 – infected adults receiving long-term antiretroviral therapy. J Infect Dis. 2016;213(4):556–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Maldarelli F, Palmer S, King MS, et al. ART suppresses plasma HIV-1 RNA to a stable set point predicted by pretherapy viremia. PLoS Pathog. 2007;3(4):e46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Neuhaus J, Jacobs, Jr DR Jr, Baker JV, et al. Markers of inflammation, coagulation, and renal function are elevated in adults with HIV infection. J Infect Dis. 2012;42(12):3334–3345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Wada NI, Jacobson LP, Margolick JB, et al. The effect of HAART-induced HIV suppression on circulating markers of inflammation and immune activation. AIDS. 2015;29(4):463–471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Tenorio AR, Zheng Y, Bosch RJ, et al. Soluble markers of inflammation and coagulation but not T-cell activation predict non-AIDS-defining morbid events during suppressive antiretroviral treatment. J Infect Dis. 2014;210(8):1248–1259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Hsue PY, Deeks SG, Hunt PW.. Immunologic basis of cardiovascular disease in HIV-infected adults. J Infect Dis. 2012;205(SUPPL. 3):375–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Marks MA, Rabkin CS, Engels EA, et al. Markers of microbial translocation and risk of AIDS-related lymphoma. AIDS. 2013. January;27(3):469–474. [DOI] [PubMed] [Google Scholar]
- [9].Ancuta P, Kamat A, Kunstman KJ, et al. Microbial translocation is associated with increased monocyte activation and dementia in AIDS patients. PLoS One. 2008;3(6):e2516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Andrade BB, Hullsiek KH, Boulware DR, et al. Biomarkers of inflammation and coagulation are associated with mortality and hepatitis flares in persons coinfected with HIV and hepatitis viruses. J Infect Dis. 2013;207(9):1379–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Kuller LH, Lundgren J, Neaton JD, et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008;5(10):e203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Brenchley JM, Price DA, Schacker TW, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006. December;12(12):1365–1371. [DOI] [PubMed] [Google Scholar]
- [13].Martinez-Picado J, Deeks SG. Persistent HIV-1 replication during antiretroviral therapy. Curr Opin HIV AIDS. 2016;11(4):417–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Gianella S, Letendre S. Cytomegalovirus and HIV: a dangerous Pas de Deux. J Infect Dis. 2016;214(Suppl 2):S67–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Hunt PW, Landay AL, Sinclair E, et al. A low T regulatory cell response may contribute to both viral control and generalized immune activation in HIV controllers. PLoS One. 2011. January;6(1):e15924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Estes JD, Haase AT, Schacker TW. The role of collagen deposition in depleting CD4+ T cells and limiting reconstitution in HIV-1 and SIV infections through damage to the secondary lymphoid organ niche. Semin Immunol. 2008;20(3):181–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Witwer KW, Théry C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J Extracell Vesicles [Internet] 2019;8(1):1648167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Mathieu M, Martin-Jaular L, Lavieu G, et al. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication [Internet] Nat Cell Biol. 2019;21(1):9–17. Available from: http://www.nature.com/articles/s41556-018-0250-9. [DOI] [PubMed] [Google Scholar]
- [19].Minciacchi VR, You S, Spinelli C, et al. Large oncosomes contain distinct protein cargo and represent a separate functional class of tumor-derived extracellular vesicles. Oncotarget. 2015. May;6(13):11327–11341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Keerthikumar S, Gangoda L, Liem M, et al. Proteogenomic analysis reveals exosomes are more oncogenic than ectosomes. Oncotarget. 2015. June;6(17):15375–15396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Mateescu B, Kowal EJK, van Balkom BWM, et al. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA - an ISEV position paper. J Extracell Vesicles. 2017;6(1):1286095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Yanez-Mo M, Siljander PR-M, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–593. [DOI] [PubMed] [Google Scholar]
- [24].Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol [Internet] 2014;14(3):195–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Théry C, Duban L, Segura E, et al. Indirect activation of naïve CD4+T cells by dendritic cell-derived exosomes. Nat Immunol. 2002;3(12):1156–1162. [DOI] [PubMed] [Google Scholar]
- [26].Zitvogel L, Regnault A, Lozier A, et al. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med. 1998. May;4(5):594–600. [DOI] [PubMed] [Google Scholar]
- [27].Tkach M, Kowal J, Zucchetti AE, et al. Qualitative differences in T-cell activation by dendritic cell-derived extracellular vesicle subtypes. EMBO J. 2017;36(20):3012–3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Okoye IS, Coomes SM, Pelly VS, et al. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity. 2014. July;41(1):89–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Zhang F, Li R, Yang Y, et al. Specific decrease in B-cell-derived extracellular vesicles enhances post-chemotherapeutic CD8+ T cell responses. Immunity. [Internet] 2019. February 12: pii: S1074 [cited 2019 March8]. Available from: https://www.sciencedirect.com/science/article/pii/S1074761319300330?via%3Dihub [DOI] [PubMed] [Google Scholar]
- [30].Turpin D, Truchetet ME, Faustin B, et al. Role of extracellular vesicles in autoimmune diseases. Autoimmun Rev [Internet] 2016;15(2):174–183. [DOI] [PubMed] [Google Scholar]
- [31].Kim M-R, Hong S-W, Choi E-B, et al. Staphylococcus aureus-derived extracellular vesicles induce neutrophilic pulmonary inflammation via both Th1 and Th17 cell responses. Allergy. 2012. October;67(10):1271–1281. [DOI] [PubMed] [Google Scholar]
- [32].Hong S-W, Kim M-R, Lee E-Y, et al. Extracellular vesicles derived from staphylococcus aureus induce atopic dermatitis-like skin inflammation. Allergy. 2011. March;66(3):351–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Nakao R, Hasegawa H, Ochiai K, et al. Outer membrane vesicles of porphyromonas gingivalis elicit a mucosal immune response. PLoS One. 2011;6(10):e26163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Prados-Rosales R, Baena A, Martinez LR, et al. Mycobacteria release active membrane vesicles that modulate immune responses in a TLR2-dependent manner in mice. J Clin Invest. 2011. April;121(4):1471–1483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Bhatnagar S, Shinagawa K, Castellino FJ, et al. Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and i. Blood. 2007;110(9):3234–3244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Singh PP, LeMaire C, Tan JC, et al. Exosomes released from M. tuberculosis infected cells can suppress IFN-gamma mediated activation of naive macrophages. PLoS One. 2011. April;6(4):e18564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Ayna G, Krysko DV, Kaczmarek A, et al. ATP release from dying autophagic cells and their phagocytosis are crucial for inflammasome activation in macrophages. PLoS One. 2012;7(6):e40069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Goh FG, Midwood KS. Intrinsic danger: activation of toll-like receptors in rheumatoid arthritis. Rheumatology (Oxford). 2012. January;51(1):7–23. [DOI] [PubMed] [Google Scholar]
- [39].Gulinelli S, Salaro E, Vuerich M, et al. IL-18 associates to microvesicles shed from human macrophages by a LPS/TLR-4 independent mechanism in response to P2X receptor stimulation. Eur J Immunol. 2012. December;42(12):3334–3345. [DOI] [PubMed] [Google Scholar]
- [40].Nickel W, Rabouille C. Mechanisms of regulated unconventional protein secretion. Nat Rev Mol Cell Biol. 2009. February;10(2):148–155. [DOI] [PubMed] [Google Scholar]
- [41].Pizzirani C, Ferrari D, Chiozzi P, et al. Stimulation of P2 receptors causes release of IL-1 β − loaded microvesicles from human dendritic cells. Blood. 2012;109(9):3856–3864. [DOI] [PubMed] [Google Scholar]
- [42].Cossetti C, Iraci N, Mercer TR, et al. Extracellular vesicles from neural stem cells transfer IFN-gamma via Ifngr1 to activate Stat1 signaling in target cells. Mol Cell. 2014. October;56(2):193–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Esser J, Gehrmann U, D’Alexandri FL, et al. Exosomes from human macrophages and dendritic cells contain enzymes for leukotriene biosynthesis and promote granulocyte migration. J Allergy Clin Immunol. 2010. November;126(5):1032–1040. 1040.e1–4. [DOI] [PubMed] [Google Scholar]
- [44].Majumdar R, Tavakoli Tameh A, Parent CA. Exosomes mediate LTB4 release during neutrophil chemotaxis. PLoS Biol. 2016. January;14(1):e1002336. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- [45].Kriebel PW, Majumdar R, Jenkins LM, et al. Extracellular vesicles direct migration by synthesizing and releasing chemotactic signals. J Cell Biol. 2018. August;217(8):2891–2910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Bello-Morales R, Praena B, de la Nuez C, et al. Role of microvesicles in the spread of herpes simplex virus 1 in oligodendrocytic cells. J Virol. 2018;92(10):1–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Yang Y, Han Q, Hou Z, et al. Exosomes mediate hepatitis B virus (HBV) transmission and NK-cell dysfunction. Cell Mol Immunol [Internet] 2017;14(5):465–475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Bukong TN, Momen-Heravi F, Kodys K, et al. Exosomes from hepatitis C infected patients transmit HCV infection and contain replication competent viral RNA in complex with Ago2-miR122-HSP90. PLoS Pathog. 2014;10(10):e1004424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Ramakrishnaiah V, Thumann C, Fofana I, et al. Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc Natl Acad Sci. 2013;110(32):13109–13113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Feng Z, Hensley L, McKnight KL, et al. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature [Internet] 2013;496(7445):367–371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Chen Y-H, Du W, Hagemeijer MC, et al. Phosphatidylserine vesicles enable efficient en bloc transmission of enteroviruses. Cell. 2015. February;160(4):619–630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Santiana M, Ghosh S, Ho BA, et al. Vesicle-cloaked virus clusters are optimal units for inter-organismal viral transmission. Cell Host Microbe. 2018;24(2):208–220.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].van der Grein SG, Defourny KAY, Rabouw HH, et al. Picornavirus infection induces temporal release of multiple extracellular vesicle subsets that differ in molecular composition and infectious potential. PLOS Pathog [Internet] 2019;15(2):e1007594 Available from: http://dx.plos.org/10.1371/journal.ppat.1007594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Dukers DF, Meij P, Vervoort MBHJ, et al. Direct immunosuppressive effects of EBV-encoded latent membrane protein 1. J Immunol. 2000;165(2):663–670. [DOI] [PubMed] [Google Scholar]
- [55].Keryer-Bibens C, Pioche-Durieu C, Villemant C, et al. Exosomes released by EBV-infected nasopharyngeal carcinoma cells convey the viral latent membrane protein 1 and the immunomodulatory protein galectin 9. BMC Cancer. 2006;6:1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Fu Y, Zhang L, Zhang F, et al. Exosome-mediated miR-146a transfer suppresses type I interferon response and facilitates EV71 infection. PLoS Pathog. 2017;13(9):1–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Klibi J, Niki T, Riedel A, et al. Blood diffusion and Th1-suppressive effects of galectin-9-containing exosomes released by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Blood. 2009;113(9):1957–1967. [DOI] [PubMed] [Google Scholar]
- [58].Cobb DA, Kim OK, Golden-Mason L, et al. Hepatocyte-derived exosomes promote T follicular regulatory cell expansion during hepatitis C virus infection. Hepatology. 2018;67(1):71–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Feng Z, Walker CM, Lemon SM, et al. Human pDCs preferentially sense enveloped hepatitis A virions. J Clin Invest. 2015;125(1):169–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Dreux M, Garaigorta U, Boyd B, et al. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe. 2012;12(4):558–570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Baglio SR, van Eijndhoven MAJ, Koppers-Lalic D, et al. Sensing of latent EBV infection through exosomal transfer of 5′pppRNA. Proc Natl Acad Sci. 2016;113(5):E587–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Li J, Liu K, Liu Y, et al. Exosomes mediate the cell-to-cell transmission of IFN-α-induced antiviral activity. Nat Immunol. 2013;14(8):793–803. [DOI] [PubMed] [Google Scholar]
- [63].Deschamps T, Kalamvoki M. Extracellular vesicles released by herpes simplex virus 1-infected cells block virus replication in recipient cells in a STING-dependent manner. J Virol. 2018. September;92(18):e01102-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Delorme-Axford E, Donker RB, Mouillet JF, et al. Human placental trophoblasts confer viral resistance to recipient cells. Proc Natl Acad Sci U S A. 2013;110(29):12048–12053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Gould SJ, Booth AM, Hildreth JEK. The Trojan exosome hypothesis. Proc Natl Acad Sci U S A. 2003. September;100(19):10592–10597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Bess JWJ, Gorelick RJ, Bosche WJ, et al. Microvesicles are a source of contaminating cellular proteins found in purified HIV-1 preparations. Virology. 1997. March;230(1):134–144. [DOI] [PubMed] [Google Scholar]
- [67].Nolte-’t Hoen E, Cremer T, Gallo RC, et al. Extracellular vesicles and viruses: are they close relatives? Proc Natl Acad Sci U S A. 2016. August;113(33):9155–9161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Ott DE. Purification of HIV-1 virions by subtilisin digestion or CD45 immunoaffinity depletion for biochemical studies. Methods Mol Biol. 2009;485:15–25. [DOI] [PubMed] [Google Scholar]
- [69].Cantin R, Diou J, Bélanger D, et al. Discrimination between exosomes and HIV-1: purification of both vesicles from cell-free supernatants. J Immunol Methods. 2008;338(1–2):21–30. [DOI] [PubMed] [Google Scholar]
- [70].Liao Z, Martin-Jaular L, Soueidi E, et al. Acetylcholinesterase is not a generic marker of extracellular vesicles. J Extracell Vesicles. 2019;8(1):1628592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the international society for extracellular vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles Internet]. 2018;8(1):1535750 Available from: https://www.tandfonline.com/doi/full/10.1080/20013078.2018.1535750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Böing AN, Van Der Pol E, Grootemaat AE, et al. Single-step isolation of extracellular vesicles from plasma by size-exclusion chromatography. Int Meet ISEV Rotterdam. 2014;3:118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Karimi N, Cvjetkovic A, Jang SC, et al. Detailed analysis of the plasma extracellular vesicle proteome after separation from lipoproteins. Cell Mol Life Sci [Internet] 2018;75(15):2873–2886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Dias MVS, Costa CS, Da Silva LLP. The ambiguous roles of extracellular vesicles in HIV replication and pathogenesis. Front Microbiol. 2018;9(OCT):1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].de Carvalho JV, de Castro RO, da Silva EZM, et al. Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLoS One. 2014;9(11):e113691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Khatua AK, Taylor HE, Hildreth JEK, et al. Exosomes packaging APOBEC3G confer human immunodeficiency virus resistance to recipient cells. J Virol. 2009. January;83(2):512–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Haque S, Sinha N, Ranjit S, et al. Monocyte-derived exosomes upon exposure to cigarette smoke condensate alter their characteristics and show protective effect against cytotoxicity and HIV-1 replication. Sci Rep [Internet] 2017;7(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Tumne A, Prasad VS, Chen Y, et al. Noncytotoxic suppression of human immunodeficiency virus type 1 transcription by exosomes secreted from CD8+ T cells. J Virol. 2009. May;83(9):4354–4364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Madison MN, Jones PH, Okeoma CM. Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology. 2015. August;482:189–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Madison MN, Roller RJ, Okeoma CM. Human semen contains exosomes with potent anti-HIV-1 activity. Retrovirology. 2014. November;11:102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Verhoef K, Tijms M, Berkhout B. Optimal Tat-mediated activation of the HIV-1 LTR promoter requires a full-length TAR RNA hairpin. Nucleic Acids Res. 1997. February;25(3):496–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Welch JL, Kaddour H, Schlievert PM, et al. Semen exosomes promote transcriptional silencing of HIV-1 by disrupting NF-kappaB/Sp1/Tat circuitry. J Virol. 2018. November;92(21):e00731-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Naslund TI, Paquin-Proulx D, Paredes PT, et al. Exosomes from breast milk inhibit HIV-1 infection of dendritic cells and subsequent viral transfer to CD4+ T cells. AIDS. 2014. January;28(2):171–180. [DOI] [PubMed] [Google Scholar]
- [84].Smith JA, Daniel R. Human vaginal fluid contains exosomes that have an inhibitory effect on an early step of the HIV-1 life cycle. AIDS. 2016. November;30(17):2611–2616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Rozmyslowicz T, Majka M, Kijowski J, et al. Platelet- and megakaryocyte-derived microparticles transfer CXCR4 receptor to CXCR4-null cells and make them susceptible to infection by X4-HIV. AIDS. 2003. January;17(1):33–42. [DOI] [PubMed] [Google Scholar]
- [86].Mack M, Kleinschmidt A, Bruhl H, et al. Transfer of the chemokine receptor CCR5 between cells by membrane-derived microparticles: a mechanism for cellular human immunodeficiency virus 1 infection. Nat Med. 2000. July;6(7):769–775. [DOI] [PubMed] [Google Scholar]
- [87].Mercier SK, Donaghy H, Botting RA, et al. The microvesicle component of HIV-1 inocula modulates dendritic cell infection and maturation and enhances adhesion to and activation of T lymphocytes. PLoS Pathog. 2013. October;9(10):e1003700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Kadiu I, Narayanasamy P, Dash PK, et al. Biochemical and biologic characterization of exosomes and microvesicles as facilitators of HIV-1 infection in macrophages. J Immunol [Internet] 2012;189(2):744–754. Available from: http://www.jimmunol.org/cgi/doi/10.4049/jimmunol.1102244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Lenassi M, Cagney G, Liao M, et al. HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic. 2010;11(1):110–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Arenaccio C, Chiozzini C, Columba-Cabezas S, et al. Exosomes from human immunodeficiency virus type 1 (HIV-1)-infected cells license quiescent CD4+ T lymphocytes to replicate HIV-1 through a Nef- and ADAM17-dependent mechanism. J Virol. 2014. October;88(19):11529–11539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Arakelyan A, Fitzgerald W, Zicari S, et al. Extracellular vesicles carry hiv env and facilitate HIV infection of human lymphoid tissue. Sci Rep. 2017. May;7(1):1695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Narayanan A, Iordanskiy S, Das R, et al. Exosomes derived from HIV-1-infected cells contain trans-activation response element RNA. J Biol Chem. 2013;288(27):20014–20033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Siliciano JD, Kajdas J, Finzi D, et al. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat Med. 2003. June;9(6):727–728. [DOI] [PubMed] [Google Scholar]
- [94].Barclay RA, Schwab A, DeMarino C, et al. Exosomes from uninfected cells activate transcription of latent HIV-1. J Biol Chem. 2017. July;292(28):11682–11701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Arenaccio C, Anticoli S, Manfredi F, et al. Latent HIV-1 is activated by exosomes from cells infected with either replication-competent or defective HIV-1. Retrovirology. 2015;12(1):1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Tang X, Lu H, Dooner M, et al. Exosomal Tat protein activates latent HIV-1 in primary, resting CD4+ T lymphocytes. JCI Insight. 2018. April;3(7):95676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [97].Liao Z, Muth DC, Eitan E, et al. Serum extracellular vesicle depletion processes affect release and infectivity of HIV-1 in culture. Sci Rep. 2017. May;7(1):2558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Buzas EI, György B, Nagy G, et al. Emerging role of extracellular vesicles in inflammatory diseases. Nat Rev Rheumatol [Internet] 2014;10(6):356–364. [DOI] [PubMed] [Google Scholar]
- [99].Boulanger CM, Loyer X, Rautou PE, et al. Extracellular vesicles in coronary artery disease. Nat Rev Cardiol [Internet] 2017;14(5):259–272. [DOI] [PubMed] [Google Scholar]
- [100].Lee JH, Schierer S, Blume K, et al. HIV-Nef and ADAM17-containing plasma extracellular vesicles induce and correlate with immune pathogenesis in chronic HIV infection. EBioMedicine [Internet] 2016;6(1):03–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Chettimada S, Lorenz DR, Misra V, et al. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci Rep. 2018;8:7227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Hubert A, Subra C, Jenabian MA, et al. Elevated abundance, size, and MicroRNA content of plasma extracellular vesicles in viremic HIV-1+ patients: correlations with known markers of disease progression. J Acquir Immune Defic Syndr. 2015;70(3):219–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Beck SE, Queen SE, Metcalf Pate KA, et al. An SIV/macaque model targeted to study HIV-associated neurocognitive disorders. J Neurovirol. 2018. April;24(2):204–212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Koliha N, Wiencek Y, Heider U, et al. A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J Extracell Vesicles. 2016;5:29975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Muratori C, Cavallin LE, Krätzel K, et al. Massive secretion by T cells is caused by HIV Nef in infected cells and by Nef transfer to bystander cells. Cell Host Microbe. 2009;6(3):218–230. [DOI] [PubMed] [Google Scholar]
- [106].Subra C, Simard S, Mercier S, et al. Dendritic cells pulsed with HIV-1 release exosomes that promote apoptosis in CD4 + T lymphocytes. J Clin Cell Immunol [Internet] 2011;7(1). [Google Scholar]
- [107].Mfunyi CM, Vaillancourt M, Vitry J, et al. Exosome release following activation of the dendritic cell immunoreceptor: a potential role in HIV-1 pathogenesis. Virology [Internet] 2015;484:103–112. [DOI] [PubMed] [Google Scholar]
- [108].Hromada C, Mühleder S, Grillari J, et al. Endothelial extracellular vesicles-promises and challenges. Front Physiol. 2017 May;8:275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Bernard MA, Zhao H, Yue SC, et al. Novel HIV-1 MiRNAs stimulate TNFa release in human macrophages via TLR8 signaling pathway. PLoS One. 2014;9(9):e106006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Lee JH, Wittki S, Bräu T, et al. HIV Nef, paxillin, and Pak1/2 regulate activation and secretion of TACE/ADAM10 proteases. Mol Cell. 2013;49(4):668–679. [DOI] [PubMed] [Google Scholar]
- [111].Ostalecki C, Wittki S, Lee JH, et al. HIV Nef- and Notch1-dependent endocytosis of ADAM17 induces vesicular TNF secretion in chronic HIV infection. EBioMedicine [Internet] 2016;13:294–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Duette G, Pereyra Gerber P, Rubione J, et al. Induction of HIF-1α by HIV-1 infection in CD4+ T cells promotes viral replication and drives extracellular vesicle-mediated inflammation. MBio [Internet] 2018;9(5):e00757–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [113].McNamara RP, Costantini LM, Myers TA, et al. Nef secretion into extracellular vesicles or exosomes is conserved across human and simian immunodeficiency viruses. MBio. 2018;9(1):1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Raymond AD, Campbell-Sims TC, Khan M, et al. HIV type 1 Nef is released from infected cells in CD45+ microvesicles and is present in the plasma of HIV-infected individuals. AIDS Res Hum Retroviruses [Internet] 2011;27(2):167–178. Available from: http://www.liebertonline.com/doi/abs/10.1089/aid.2009.0170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Luo X, Fan Y, Park IW, et al. Exosomes are unlikely involved in intercellular Nef transfer. PLoS One. 2015;10(4):1–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Columba Cabezas S, Federico M. Sequences within RNA coding for HIV-1 Gag p17 are efficiently targeted to exosomes. Cell Microbiol. 2013;15(3):412–429. [DOI] [PubMed] [Google Scholar]
- [117].Sampey GC, Saifuddin M, Schwab A, et al. Exosomes from HIV-1-infected cells stimulate production of pro-inflammatory cytokines through trans-activating response (TAR) RNA. J Biol Chem. 2016;291(3):1251–1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [118].Konadu KA, Huang MB, Roth W, et al. Isolation of exosomes from the plasma of HIV-1 positive individuals. J Vis Exp [Internet] 2016;(107):1–9. Available from: http://www.jove.com/video/53495/isolation-of-exosomes-from-the-plasma-of-hiv-1-positive-individuals [DOI] [PMC free article] [PubMed] [Google Scholar]
- [119].Kodidela S, Ranjit S, Sinha N, et al. Cytokine profiling of exosomes derived from the plasma of HIV-infected alcohol drinkers and cigarette smokers. PLoS One. 2018;13(7):e0201144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Swaminathan G, Navas-Martin S, Martin-Garcia J. MicroRNAs and HIV-1 infection: antiviral activities and beyond. J Mol Biol. 2014. March;426(6):1178–1197. [DOI] [PubMed] [Google Scholar]
- [121].Huang J, Wang F, Argyris E, et al. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat Med. 2007. October;13(10):1241–1247. [DOI] [PubMed] [Google Scholar]
- [122].Roth WW, Huang MB, Konadu KA, et al. Micro RNA in exosomes from HIV-infected macrophages. Int J Environ Res Public Health. 2015;13(1):ijerph13010032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [123].Tang B, Li X, Ren Y, et al. MicroRNA-29a regulates lipopolysaccharide (LPS)-induced inflammatory responses in murine macrophages through the Akt1/NF-κB pathway. Exp Cell Res. 2017;360(2):74–80. [DOI] [PubMed] [Google Scholar]
- [124].Semenza GL, Wang GL. A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation. Mol Cell Biol. 1992. December;12(12):5447–5454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [125].Cramer T, Yamanishi Y, Clausen BE, et al. HIF-1alpha is essential for myeloid cell-mediated inflammation. Cell. 2003. March;112(5):645–657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [126].Zhang W, Petrovic J-M, Callaghan D, et al. Evidence that hypoxia-inducible factor-1 (HIF-1) mediates transcriptional activation of interleukin-1beta (IL-1beta) in astrocyte cultures. J Neuroimmunol. 2006. May;174(1–2):63–73. [DOI] [PubMed] [Google Scholar]
- [127].Cheng S-C, Quintin J, Cramer RA, et al. mTOR- and HIF-1alpha-mediated aerobic glycolysis as metabolic basis for trained immunity. Science. 2014. September;345(6204):1250684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [128].Wang T, Gilkes DM, Takano N, et al. Hypoxia-inducible factors and RAB22A mediate formation of microvesicles that stimulate breast cancer invasion and metastasis. Proc Natl Acad Sci. 2014;111(31):E3234–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Aga M, Bentz GL, Raffa S, et al. Exosomal HIF1alpha supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene. 2014. September;33(37):4613–4622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [130].Gonzalez-King H, Garcia NA, Ontoria-Oviedo I, et al. Hypoxia inducible factor-1alpha potentiates jagged 1-mediated angiogenesis by mesenchymal stem cell-derived exosomes. Stem Cells. 2017. July;35(7):1747–1759. [DOI] [PubMed] [Google Scholar]
- [131].Chen F, Chen J, Yang L, et al. Extracellular vesicle-packaged HIF-1alpha-stabilizing lncRNA from tumour-associated macrophages regulates aerobic glycolysis of breast cancer cells. Nat Cell Biol. 2019. April;21(4):498–510. [DOI] [PubMed] [Google Scholar]
- [132].Saylor D, Dickens AM, Sacktor N, et al. HIV-associated neurocognitive disorder — pathogenesis and prospects for treatment. Nat Rev Neurol. 2016. April;12(4):234–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Shaw G, Harper M, Hahn B, et al. HTLV-III infection in brains of children and adults with AIDS encephalopathy. Science. 1985. January;227(4683):177–182. [DOI] [PubMed] [Google Scholar]
- [134].Hong S, Banks WA. Role of the immune system in HIV-associated neuroinflammation and neurocognitive implications. Brain Behav Immun. 2015. March;45:1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Sillman B, Woldstad C, Mcmillan J, et al. Neuropathogenesis of human immunodeficiency virus infection. Handb Clin Neurol. 2018;152:21–40. [DOI] [PubMed] [Google Scholar]
- [136].Witwer KW, Gama L, Li M, et al. Coordinated regulation of SIV replication and immune responses in the CNS. PLoS One. 2009. December 19;4(12):e8129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Bissel SJ, Wiley CA. Human immunodeficiency virus infection of the brain: pitfalls in evaluating infected/affected cell populations. Brain Pathol. 2004. January;14(1):97–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Fischer-Smith T, Bell C, Croul S, et al. Monocyte/macrophage trafficking in acquired immunodeficiency syndrome encephalitis: lessons from human and nonhuman primate studies. J Neurovirol. 2008. January;14(4):318–326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [139].Williams KC, Corey S, Westmoreland SV, et al. Perivascular macrophages are the primary cell type productively infected by simian immunodeficiency virus in the brains of macaques: implications for the neuropathogenesis of AIDS. J Exp Med. 2001. April 17;193(8):905–915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [140].Cosenza MA, Zhao M-L, Si Q, et al. Human brain parenchymal microglia express CD14 and CD45 and are productively infected by HIV-1 in HIV-1 encephalitis. Brain Pathol. 2006. April;12(4):442–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [141].Borjabad A, Brooks AI, Volsky DJ. Gene expression profiles of HIV-1-infected glia and brain: toward better understanding of the role of astrocytes in HIV-1-associated neurocognitive disorders. J Neuroimmune Pharmacol. 2010. March;5(1):44–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [142].Ivey NS, MacLean AG, Lackner AA. Acquired immunodeficiency syndrome and the blood-brain barrier. J Neurovirol. 2009. April;15(2):111–122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Tyor WR, Glass JD, Griffin JW, et al. Cytokine expression in the brain during the acquired immunodeficiency syndrome. Ann Neurol. 1992. April;31(4):349–360. [DOI] [PubMed] [Google Scholar]
- [144].Lipton SA, Yeh M, Dreyer EB. Update on current models of HIV-related neuronal injury: platelet-activating factor, arachidonic acid and nitric oxide. Adv Neuroimmunol. 1994;4(3):181–188. [DOI] [PubMed] [Google Scholar]
- [145].Alonso K, Pontiggia P, Medenica R, et al. Cytokine patterns in adults with AIDS. Immunol Invest. 1997. April;26(3):341–350. [DOI] [PubMed] [Google Scholar]
- [146].Tavazzi E, Morrison D, Sullivan P, et al. Brain inflammation is a common feature of HIV-infected patients without HIV encephalitis or productive brain infection. Curr HIV Res. 2014;12(2):97–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [147].Antinori A, Arendt G, Becker JT, et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology. 2007. October 05;69(18):1789–1799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [148].Wang T, Gong N, Liu J, et al. HIV-1-infected astrocytes and the microglial proteome. J Neuroimmune Pharmacol. 2008. September;3(3):173–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [149].Yang L, Niu F, Yao H, et al. Exosomal miR-9 released from HIV Tat stimulated astrocytes mediates microglial migration. J Neuroimmune Pharmacol. 2018;13(3):330–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [150].Ranjit S, Patters BJ, Gerth KA, et al. Potential neuroprotective role of astroglial exosomes against smoking-induced oxidative stress and HIV-1 replication in the central nervous system. Expert Opin Ther Targets. 2018. August;22(8):703–714. [DOI] [PubMed] [Google Scholar]
- [151].Chaudhuri AD, Dastgheyb RM, Yoo S-W, et al. TNFα and IL-1β modify the miRNA cargo of astrocyte shed extracellular vesicles to regulate neurotrophic signaling in neurons article. Cell Death Dis. 2018;9(3):363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Dickens AM, Tovar-y-Romo LB, Yoo S-W, et al. Astrocyte-shed extracellular vesicles regulate the peripheral leukocyte response to inflammatory brain lesions. Sci Signal. 2017. Apr;10(473:Eaai7696). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [153].Goetzl EJ, Schwartz JB, Abner EL, et al. High complement levels in astrocyte-derived exosomes of Alzheimer disease. Ann Neurol. 2018. Mar;83(3):544–552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [154].Pulliam L, Sun B, Mustapic M, et al. Plasma neuronal exosomes serve as biomarkers of cognitive impairment in HIV infection and Alzheimer’s disease. J Neurovirol. 2019. January. doi: 10.1007/s13365-018-0695-4. [Epub ahead of print]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [155].Zhao J, Lopez AL, Erichsen D, et al. Mitochondrial glutaminase enhances extracellular glutamate production in HIV-1-infected macrophages: linkage to HIV-1 associated dementia. J Neurochem. 2003. November;88(1):169–180. [DOI] [PubMed] [Google Scholar]
- [156].Wang K, Ye L, Lu H, et al. TNF-α promotes extracellular vesicle release in mouse astrocytes through glutaminase. J Neuroinflammation. 2017. December;14(1):87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [157].Wu B, Huang Y, Braun AL, et al. Glutaminase-containing microvesicles from HIV-1-infected macrophages and immune-activated microglia induce neurotoxicity. Mol Neurodegener. 2015. December;10(1):61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [158].Wu B, Liu J, Zhao R, et al. Glutaminase 1 regulates the release of extracellular vesicles during neuroinflammation through key metabolic intermediate alpha-ketoglutarate. J Neuroinflammation. 2018;15(1):1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [159].Ye L, Huang Y, Zhao L, et al. IL-1β and TNF-α induce neurotoxicity through glutamate production: a potential role for neuronal glutaminase. J Neurochem. 2013. June;125(6):897–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [160].Rivest S. Regulation of innate immune responses in the brain. Nat Rev Immunol. 2009. June;9(6):429–439. [DOI] [PubMed] [Google Scholar]
- [161].Hu G, Yao H, Chaudhuri AD, et al. Exosome-mediated shuttling of microRNA-29 regulates HIV Tat and morphine-mediated neuronal dysfunction. Cell Death Dis. 2012. August 31;3:e381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Yelamanchili SV, Lamberty BG, Rennard DA, et al. MiR-21 in extracellular vesicles leads to neurotoxicity via TLR7 signaling in SIV neurological disease. Douek DC, editor. PLoS Pathog. 2015. July;11(7):e1005032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Klase Z, Kale P, Winograd R, et al. HIV-1 TAR element is processed by Dicer to yield a viral micro-RNA involved in chromatin remodeling of the viral LTR. BMC Mol Biol. 2007. August 01;8:63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].Raymond AD, Diaz P, Chevelon S, et al. Microglia-derived HIV Nef+ exosome impairment of the blood-brain barrier is treatable by nanomedicine-based delivery of Nef peptides. J Neurovirol. 2016. April;22(2):129–139. [DOI] [PubMed] [Google Scholar]
- [165].Dominkuš PP, Ferdin J, Plemenitaš A, et al. Nef is secreted in exosomes from Nef.GFP-expressing and HIV-1-infected human astrocytes. J Neurovirol. 2017. October;23(5):713–724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [166].Sami Saribas A, Cicalese S, Ahooyi TM, et al. HIV-1 Nef is released in extracellular vesicles derived from astrocytes: evidence for Nef-mediated neurotoxicity. Cell Death Dis. 2017. January;8(1):e2542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Khan MB, Lang MJ, Huang MB, et al. Nef exosomes isolated from the plasma of individuals with HIV-associated dementia (HAD) can induce Aβ1–42secretion in SH-SY5Y neural cells. J Neurovirol. 2016;22(2):179–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [168].Rahimian P, He JJ. Exosome-associated release, uptake, and neurotoxicity of HIV-1 Tat protein. J Neurovirol. 2016. December;22(6):774–788. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- World Health Organization HIV/AIDS [Internet]; 2018. [cited 2019 July22]. Available from: https://www.who.int/en/news-room/fact-sheets/detail/hiv-aids