

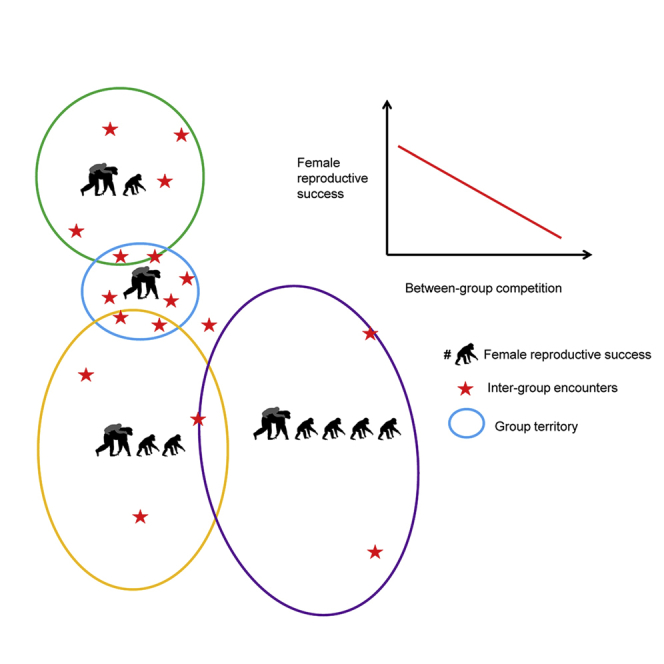
Summary
Between-group competition in social animals appears to be a prominent selective pressure shaping the evolution of territoriality and cooperation [1, 2, 3, 4]. Evidence for an effect of between-group competition on fitness in territorial species, however, is mostly lacking because of difficulty in measuring between-group competition and its long-term impact [5]. Between-group competition corresponds to a complex set of interactions between neighboring groups, and its intensity seems to depend on the competitive abilities of each interacting group [6, 7]. We tested whether the competitive ability of groups and the pressure exerted by neighboring groups affected the reproductive success of wild female chimpanzees (Pan troglodytes verus). Using long-term data on four neighboring groups in the Taï National Park, Côte d’Ivoire, collected over the course of 54 observation years, we measured the competitive ability of habituated groups using the number of mature males and the pressure exerted by non-habituated neighbors with an index of neighbor pressure that combined the frequency of neighboring encounters and related spatial information. Importantly, we found that experiencing low neighbor pressure provides fitness benefits through increased offspring survival and shorter inter-birth intervals. Also, many males in a group are associated with shorter inter-birth intervals. We conclude that high between-group competition hampers fast reproduction and offspring survival when exposure is during the prenatal period. Our findings suggest that having many males in a group results in fitness benefits and that between-group competition should be considered as a potential selective pressure that shaped key social adaptations in the hominoid lineage.
Keywords: neighbor pressure, territoriality, inter-birth interval, survival, within-group competition
Graphical Abstract
Highlights
-
•
High neighbor pressure during pregnancy affects chimpanzees’ offspring survival
-
•
Female chimpanzees reproduce slower when neighbor pressure is high
-
•
Reproductive rates are shorter when many males are present in a group
-
•
Within-group competition makes female reproduction slower
Lemoine et al. use long-term data on four neighboring groups of wild chimpanzees and show that between-group competition, expressed by a novel index of neighbor pressure and a group’s competitive ability, reduces female reproductive success. This suggests that between-group competition represents a relevant selective pressure in territorial species.
Results
We investigated the effects of chimpanzee between-group competition on female reproductive success within the conceptual framework of the inter-group dominance hypothesis [8], which postulates that between-group competition results in a between-group hierarchy with more dominant groups enjoying better reproductive success than less dominant ones. A group’s dominance depends on its competitive capacity relative to other groups, measured by group size or number of individuals of the dominant sex [6]. The dominance of neighboring groups can be estimated by the intensity of the pressure they exert on a particular group [9]. Thus, to investigate the impact of between-group competition on reproductive success, one needs to consider both the group’s competitive ability and the level of neighbor pressure experienced. We define neighbor pressure as a component of between-group competition corresponding to a threat caused by presence of neighbors in one’s territory. Reproductive success should improve when individuals live in a group with high competitive ability and low neighbor pressure. We tested these predictions using 54 observation years of ranging and demographic data on four neighboring groups (north, middle, south, and east) of wild chimpanzees (Pan troglodytes verus) in the Taï National Park, Ivory Coast [10, 11]. Chimpanzees are a suitable species to study the effects of between-group competition as they are predominantly hostile toward neighbors and inter-group encounters can be lethal [12, 13, 14]. Lethal aggression between neighbors depends on the number of males [13], so we used the number of mature males >12 years old (range 0−7 across groups) [15, 16] to approximate the competitive ability of habituated groups. The pressure exerted by neighbors is often measured by using rates of inter-group encounters [17, 18, 19, 20, 21], home-range overlap [22], or population density [23, 24]. However, low rates of inter-group encounters may not reflect low neighbor pressure but rather inter-group encounters avoidance, especially when power asymmetry exists [25]. Population density, although applicable for comparative studies of different populations [13], is not suitable for within-population comparisons where local variation in population density is usually low.
As locations of inter-group encounters can have an effect on the outcome of such contests [9, 19], we measured the neighbor pressure by a novel index combining information on occurrence and location of inter-group encounters (n = 384). This measure, which approximates the theoretical take-over pressure [17], assumes that the threat is higher if neighbor intrusion occurs close to the center of the residents’ territory [19] or in frequently used locations [9, 17, 26]. We then propose the following composite neighbor pressure index (NPI) = μ [(I) × (K)]j × Fj, where “I” represents the relative distance of the inter-group encounter to the territory center [26] and “K” the value of the inter-group encounters locations as a function of usage by the group over the past year [19]. Over a time period of interest j (i.e., during the 8.5 months of pregnancy), the mean (μ) value of the product of these two spatial measures is multiplied by F as the mean frequency of inter-group encounter occurrence [17, 18, 19]. We assume that the neighbor pressure index is high in a given group when inter-group encounters are more frequent and occur in locations that are close to the territory center and frequently used by the territory owners.
To assess the effect of between-group competition on chimpanzee female reproductive success, we analyzed how a group’s competitive ability, neighbor pressure, and within-group competition affected offspring survival and inter-birth interval lengths. Because the importance of inter-group encounters’ locations for chimpanzees’ dynamics of between-group competition has not previously been tested, and to assess whether our index is an adequate measure of costs due to between-group competition, we performed ad hoc model comparisons using an information-theoretic approach [27] between the models, including the neighbor pressure index, and subsequent models testing the effect of each component of the neighbor pressure index separately while keeping the other variables in all models constant. We included vocal and physical inter-group encounters when computing the neighbor pressure index, because both types of inter-group encounters are known to trigger a stress response [28, 29]. Although they differ in the risk of injury, they are unlikely to differ in their long-term effects on territory usage, such as location avoidance. However, because physical inter-group encounters trigger a stronger cortisol response than vocal inter-group encounters [29], we computed further survival models testing the effect of the number of physical and vocal inter-group encounters or the number of physical inter-group encounters only, which we compared to the original model that tested the effect of the neighbor pressure index. We did not consider patrolling behavior, as we concentrated on evidence of neighbor detection.
Effects of Neighbor Pressure on Survival Probability
We first modeled the survival probability of offspring (age range: 0–234 months; n = 81) using a Cox proportional hazards mixed-effects model, in which we averaged each predictor across the months of pregnancy for each mother (n = 44), thus reflecting potential prenatal effects on survival. Test predictors were the number of mature males (range: 0.5–6.25) and the neighbor pressure index (range: 0.0001–0.617) as measures of between-group competition and the number of within-group weaned individuals (range: 4.12–40.25) as a measure of within-group competition. Because territory sizes differ between groups, we considered food availability scores (range: 0.66–10.31) based on the monthly fruit availability index (FAI) [30] multiplied by the yearly territory sizes to represent both between-group and within-group competition. We controlled for the mother’s dominance rank, her age at offspring birth, and the sex of the offspring. We included mother and group identity as random effects. We monitored offspring survival continuously from birth to the end of the study period, during which 37 offspring died. Mortality rates (number of deaths) did not differ significantly between groups (chi-square test χ 2 = 6.77, df = 3, p = 0.08; n(North) = 9, n(Middle) = 4, n(South) = 17, n(East) = 7; Table S1).
The test predictors significantly affected the probability of survival (likelihood ratio test [LRT]: χ 2 = 9.50; df = 4; p = 0.04). Specifically, high neighbor pressure (z = 2.62; p = 0.008; Table 1; Figure 1A) during pregnancy decreased offspring survival. Neither the number of mature males (z = −0.21; p = 0.84) nor the number of weaned individuals (z = −0.27; p = 0.79) affected survival. A subsequent analysis testing survival up to 3 years of age (to restrict offspring mortality to the dependency period) almost reached significance (LRT: χ 2 = 9.99, df = 4, p = 0.04; specific effect of the neighbor pressure index: z = 1.95, p = 0.051; Table S2; Figure 1B). A weaker effect here than when examining survival across all ages may be attributed to the fact that survival before weaning (≤3 years) mostly depends on mother’s survival [10, 31], with the exception of few adoption cases [32, 33]. Also, to determine whether the negative impact of neighbor pressure on survival was limited to the prenatal period or also acts similarly during lactation, we conducted three similar survival analyses in which the test and control variables were averaged across the first year, the first 2 years, and the first 3 years of lactation. The probability of survival over lifetime was not significantly affected by the test predictors in all three models (first year—LRT: χ 2 = 2.47, df = 4, p = 0.64; Data S1A; 2 years—LRT: χ 2 = 1.73, df = 4, p = 0.78; Data S1B; 3 years—LRT: χ 2 = 1.62, df = 4, p = 0.80; Data S1C), indicating more pronounced effects of neighbor pressure during the prenatal than the lactation period.
Table 1.
Offspring Survival Is Negatively Associated with Neighbor Pressure during Pregnancy
| Terms | b | SE | p | 95% CI |
|---|---|---|---|---|
| Food availabilitya,b | 0.294 | 0.214 | 0.170 | 0.95, 1.87 |
| Number of individualsa,b | −0.058 | 0.216 | 0.790 | 0.67, 1.33 |
| Number of malesa,b | −0.049 | 0.240 | 0.840 | 0.61, 1.47 |
| Neighbor pressurea,b | 1.025 | 0.391 | 0.008c | 1.51, 4.85c |
| Sex_infant_maled | −0.777 | 0.398 | 0.051 | 0.19, 1.13 |
| Age of motherb,d | 0.596 | 0.231 | 0.010c | 1.15, 2.87c |
| Rank of motherb,d | −0.117 | 0.197 | 0.550 | 0.66, 1.19 |
Cox proportional hazards model on offspring survival based on the pregnancy period (N = 81 offspring for 44 mothers; 37 death events). A positive regression coefficient b indicates an increased likelihood for the hazard to occur (e.g., death). Means and SD of z-transformed variables and test statistics are given in Data S2. Food availability is multiplied by each group territory size. See also Table S3 and Data S2.
Test predictors.
z-transformed.
95% confidence intervals (CIs) and p values indicating a statistically significant effect; 95% CIs are calculated using the R function “coxph” and are based on the hazard ratios (the exponent of the coefficients b).
Control predictors.
Figure 1.

Effects of Between-Group Competition and Within-Group Competition on Female Reproductive Success
(A) Effect of neighbor pressure values during pregnancy on offspring survival, up to 150 months.
(B) Effect of neighbor pressure values on offspring survival up to 3 years old (see also Table S2); red curve for neighbor pressure below the mean (range: 0–0.061; N = 56 pregnancies) and blue curve for neighbor pressure above the mean (range: 0.062–0.617; N = 25 pregnancies).
(C) Neighbor pressure effect on inter-birth interval length.
(D) Effect of group size (measured by the number of weaned individuals) on inter-birth interval length.
(E) Effect of the number of mature males on inter-birth interval length. Dashed lines show the fitted models and the gray areas their 95% confidence interval. Letters and associated colors correspond to the different communities: E, east group; M, middle group; N, north group; S, south group.
To compare the effect of the neighbor pressure index with the effect of the individual components of the index on survival during the pregnancy period, we conducted a model comparison based on Akaike information criterion (AIC) [27]. This indicated that the neighbor pressure index had the strongest effect on survival, although the individual components showed a lesser impact on survival (Table 3; Data S2). Additional comparisons investigating the effects of the number of physical encounters only, or physical and vocal encounters, both showed less impact on survival than the effects of the neighbor pressure index (Table S3; Data S2).
Table 3.
Model Comparisons between a Model Using the Composite Index for Neighbor Pressure (Model NPI) and Subsequent Models Replacing the Composite Index by the Frequency F of Inter-group Encounters (Model Frequency F), the Degree of Intrusion (Model Intrusion I), and the Kernel of the Inter-group Encounters Locations (Model Kernel K)
| Analysis | Model | df | logLik | AIC | Delta AIC | AIC Weight |
|---|---|---|---|---|---|---|
| Survival | model NPI | 7 | −122.261 | 259.5 | 0 | 0.509 |
| Survival | model I | 7 | −123.018 | 260.7 | 1.25 | 0.272 |
| Survival | model K | 7 | −123.886 | 261.8 | 2.32 | 0.160 |
| Survival | model F | 7 | −124.870 | 263.7 | 4.29 | 0.060 |
| Inter-birth interval | model NPI | 18 | −493.188 | 1,022.4 | 0 | 0.666 |
| Inter-birth interval | model I | 18 | −494.619 | 1,025.2 | 2.86 | 0.159 |
| Inter-birth interval | model K | 18 | −495.126 | 1,026.3 | 3.88 | 0.096 |
| Inter-birth interval | model F | 18 | −495.322 | 1,026.6 | 4.27 | 0.079 |
See also Data S2.
Effects of Between-Group Competition on Inter-birth Interval Length
Inter-birth interval is a life history trait commonly used to assess female reproductive success [34]. High-ranking females show higher reproductive rates compared to low-ranking females, creating shorter inter-birth intervals in most mammals [34, 35, 36, 37, 38, 39, 40, 41]. In some chimpanzee populations, females have shorter inter-birth intervals with increasing group territory size [42]. Also, high-ranking females showed faster reproductive rates due to access to more preferred food in their core area [43, 44]. These studies suggest that better access to resources improves reproductive conditions and allows females to reproduce faster [45]. It has also been shown, in various species [46] and in Taï chimpanzees [47], that inter-birth interval lengths differ depending on the sex of the previous infant. In sum, we expect higher reproductive rates (shorter inter-birth intervals) for better compared to poorer reproductive conditions, which we expect to be negatively affected by higher between-group and within-group competitions. We fitted a linear mixed model (LMM) [48] investigating whether variation in inter-birth interval lengths was influenced by mean values of the number of mature males (range: 0.8–7) and of the neighbor pressure index (range: 0.0035–0.2577) as between-group competition measures and by the number of within-group weaned individuals (range: 8–38.07) as within-group competition measure. We included food availability multiplied by territory sizes (range: 0.76–9.58), where low values are expected to be associated with high between-group and within-group competition. We controlled for the sex of the previous infant [47] and for the mother’s mean dominance rank and her age at conception. The covariates were averaged for each female across the time period (range: 1–4 years) when females should resume reproduction (3 years after the last birth until next conception). We only included females with a previous offspring that survived at least 3 years (estimated period for post-partum amenorrhea [49]; n = 71 inter-birth intervals for 44 mothers; inter-birth intervals mean ± SD = 1,800.8 ± 343.3 days; range: 1,218–2,709 days). The original model included the interactions of dominance rank with all other variables, as dominance is known to influence the length of inter-birth intervals [39, 43, 44], and random effects comprised the identity of groups and mothers.
The test predictors significantly affected inter-birth interval length (LRT: χ 2 = 18.89; df = 8; p = 0.01), but none of the interactions were significant. Therefore, we fitted a reduced model without the interactions (Tables 2 and S4). Both between-group and within-group competitions affected inter-birth intervals’ length as predicted. Female inter-birth intervals were longer when neighbor pressure was high (Figure 1C: estimate ± SE = 86.96 ± 34.15; df = 1; p = 0.037) and when the number of weaned individuals was high (Figure 1D: estimate ± SE = 124.62 ± 34.07; df = 1; p = 0.018) and shorter when the number of mature males was high (Figure 1E: estimate ± SE = −125.37 ± 39.39; df = 1; p = 0.015). Like for the survival model, the ad hoc comparison of AIC for models differing only in the inter-group encounter parameters revealed that the variance of inter-birth intervals is better explained by the model including the neighbor pressure index, followed by the intrusion and the past usage models (Table 3).
Table 2.
Inter-birth Intervals Are Affected by Neighbor Pressure, Number of Mature Males, and Group Size
| Terms | χ2 | p | 95% CI |
|---|---|---|---|
| Intercept | 1,584, 1,783 | ||
| Food availabilitya,b | 0.492 | 0.482 | −52.67, 104.32 |
| Number of individualsa,b | 5.590 | 0.018c | 56.17, 196.50c |
| Number of malesa,b | 5.860 | 0.015c | −209.04, −43.68c |
| Neighbor pressurea,b | 4.344 | 0.037c | 20.87, 153.85c |
| Sex infant maled | 5.598 | 0.017c | 72.63, 376.36c |
| Age of motherb,d | 0.282 | 0.595 | −46.06, 99.15 |
| Rank of motherb,d | 0.200 | 0.654 | −86.70, 56.36 |
The results correspond to a reduced LMM not including the interactions (see text). Means and SD of z-transformed variables and variance components of the random effects and residuals are given in Table S4. Food availability is multiplied by each group territory size. See also Table S4.
Test predictors.
z-transformed.
95% CIs and p values indicating a statistically significant effect.
Control predictors.
Discussion
This study provides evidence for effects of between-group competition on female reproductive success in a highly territorial social species under natural conditions. First, high neighbor pressure is associated with reduced offspring survival and longer inter-birth intervals. Second, high competitive ability is associated with short female inter-birth intervals. Our results support the notion that between-group competition can act as a selective pressure shaping social behavior, because females in groups with high neighbor pressure and few males experience reduced reproductive success. The role of between-group competition in shaping specific adaptations, so far, has been difficult to test due to the need for long-term data on several groups within a population (but see [50] for experimental evidence) and due to the difficulty in measuring levels of between-group competition. Here, we investigate the potential costs of between-group competition, although the question of what chimpanzees compete over will need further investigation relating neighbor pressure and socio-environmental conditions.
Using the neighbor pressure index and each group’s competitive ability both as measures of between-group competition, we can tease apart the effects due to the neighbor pressure from those due to a given group’s offensive capacity. The neighbor pressure index not only impacts significantly on reproductive success measures but also explains their variance better than the rate and the number of inter-group encounters (all types and physical only). Importantly, the models including the location parameters are more supported than the models with the rate of inter-group encounters. These results reinforce the idea that inter-group encounters’ locations influence the costs of between-group competition. High neighbor pressure during pregnancy, but not during lactation, reduced offspring survival, indicating a prenatal effect of neighbor pressure on offspring survival. Possible mechanisms explaining this effect could involve repeated high maternal hypothalamic pituitary adrenal activity caused by nutritional stress related to limited access to nutritious food resources, such as nut tree patches, in contested areas of the territory when neighbor pressure is high [51] or caused by mothers’ repeated exposure to inter-group encounters, a known stressor in chimpanzees [28, 29, 52]. In chimpanzees, mothers’ stress levels during gestation are associated with that of their offspring during development [53]. Like in humans, prenatal stress in chimpanzees may increase the susceptibility to offspring disease and thus mortality [54] or trigger epigenetic changes as seen in primates experiencing strong environmental pressures [55]. It should be noted that high neighbor pressure can be precipitated by higher mortality in one group relative to another, with weakened groups experiencing more neighbor pressure, and death is caused by a number of factors. In Taï chimpanzees, for example, these include poaching, leopard predation, and disease [10, 11, 56, 57]. Nonetheless, our results suggest that those who experienced high neighbor pressure prenatally are more vulnerable to mortality in general, even if causes of mortality are likely not directly linked to between-group competition.
Our results indicate that, under favorable conditions, such as periods of low neighbor pressure, chimpanzee females produce faster, which results in shorter inter-birth intervals. Causal factors promoting this result could be tested in future studies. Likely candidates include improved access to patchy resources under low neighbor pressure conditions and an increase of territory size for more competitive groups [42, 58]. Chimpanzees rely on ephemeral ripe fruits and may experience nutritional stress in low fruit seasons [59], and access to key food patches could also be obstructed by neighbors’ presence. As for the effect on survival, repeated exposure to inter-group encounters and a higher neighbor pressure could also act as a repeated stressor, resulting in negative effects on reproductive success. This may be particularly the case when high neighbor pressure is sustained over months or several years, such as during the inter-birth intervals (range: 1–4 years). High glucocorticoids during inter-group encounters are likely linked to the elevated risk of aggression resulting in injury or death, particularly of inter-group infanticide [28, 29], even though these rates are lower in Taï than in other chimpanzee populations [13, 60].
Reproductive rates were also shorter when more males were present in a group. Large numbers of males are associated with larger territories in Taï [10, 18]. These results replicate and confirm previous findings in chimpanzees [42] and other primate species [61], in which females reproduce faster when territory size is increased. More males also indicate a strong competitive ability at the between-group level (better fighting potential and larger territories) [18, 60], which should reduce neighbor pressure [8, 17]. Having many males may reduce incursions of neighbors and thus reduce the neighbor pressure experienced and potential loss of territory, which would benefit females, although high neighbor pressure could increase the loss of territory and thus increase within-group competition [62]. However, we cannot rule out that many males increase sexual within-group competition, in which case females might counteract increased sexual coercion by males [63] by reproducing faster. Inter-birth intervals were longer with high numbers of weaned individuals, suggesting costs of within-group feeding competition, despite no significant effect of food availability. In sum, increased reproductive rates seem a beneficial strategy for females when reproductive conditions are favorable.
Between-group competition is considered a strong driver of group territoriality and within-group cooperation [1, 2, 3, 4, 14]. Our study demonstrates the potential of between-group competition to drive selection for particular social traits in wild chimpanzees, a highly territorial species. Similar effects are expected in other territorial social species with hostile inter-group encounters associated with high potential costs [21]. Territoriality can be an evolutionary stable strategy when intrusions by neighbors are costly for territory owners [64]. Thus, the potential costs incurred by between-group competition and the advantage of increased number of males likely shaped the intense territoriality of chimpanzees. Similarly, the capacity of chimpanzees to cooperate as a group could have been selected for in a context of intense between-group competition [14, 65]. It would be adaptive to reduce the potential costs of neighbor incursions by cooperative border patrols, a strategy used by chimpanzees [14, 15], but this remains to be tested. In humans, intense levels of between-group competition have been proposed as a major selective pressure shaping the evolution of certain social adaptations until recently considered particularly specialized in humans, such as group-level cooperation with non-kin [66] and strong in-group cooperation in the face of out-group threat underpinned by facilitative neuroendocrine mechanisms [14, 67]. This study in chimpanzees offers a demonstration that between-group competition holds the potential to drive selection of particular traits and thus adds strong support to other chimpanzee studies [14, 68] indicating that such between-group competition-driven social adaptations have a much older evolutionary history within hominoids than previously thought. Intense levels of between-group competition, in conjunction with effects of group dominance [8], likely could have led to group extinctions in the hominin lineage [69, 70]; thus, our study adds information to conditions in which extinction of groups could have occurred and which specific adaptations facilitating reduction of risk could have been under selection.
STAR★Methods
Key Resources Table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Experimental Models: Organisms/Strains | ||
| Pan troglodytes verus | Taï Chimpanzee Project, Taï National Park, Ivory Coast | N/A |
| Software and Algorithms | ||
| R 3.4.0 | https://r-project.org | |
| survival v2.44-1 | [71] | CRAN (https://cran.r-project.org) |
| coxme v2.2-10 | [72] | CRAN (https://cran.r-project.org) |
| lme4 v1.1-14 | [73] | CRAN (https://cran.r-project.org) |
| car v.3.0-2 | [74] | CRAN (https://cran.r-project.org) |
| MuMin v.1.42-1 | [75] | CRAN (https://cran.r-project.org) |
| AICcmodavg v.2.2-1 | [76] | CRAN (https://cran.r-project.org) |
| Other | ||
| Variable measurements and analysis scripts | This paper | N/A |
Lead Contact and Materials Availability
Further information and requests for resources and reagents should be directed to and will be fulfilled by the Lead Contact, Sylvain Lemoine (sylvain_lemoine@eva.mpg.de). This study did not generate new unique reagents.
Experimental Model and Subject Details
We collected non-invasive observational data on four habituated wild chimpanzee groups from the Taï National Park, Ivory Coast: North (0-4 adult males, 6-12 adult females), South (2-7 adult males, 6-21 adult females), Middle (1-3 adult males, 2-4 adult females) and East (3-7 adult males, 9-18 adult females). Data collection extended from 1997 to 2016, cumulating in 54 years across groups [10, 11]. The length of the study periods varied between communities (North: 19 years, South: 17 years, Middle: 5 years; East: 9 years – Table S1). We chose 12 years old as the cut-off age for males, based on evidence for social dominance, participation to border patrol and paternity [77, 78, 79], similarly to previous studies on chimpanzees where adolescent males were also considered [15, 68]. We considered females as adult as soon as they presented exaggerated sexual swellings (minimum age: 9.5 years old). We used the number of weaned individuals, which is equivalent to the number of individuals that can feed independently from their mothers and travel without assistance, as a proxy for within-group feeding competition. We expressed the number of males and weaned individuals in the analyses as mean monthly values. Dominance ranks of females were calculated daily using the Elo rating method [80], in which we used submissive unidirectional pant grunt vocalizations to establish hierarchies [81]. Within each group and for each female, we used the mean rank over the particular period of time considered. Age and rank were poorly correlated (Pearson R = 0.43) so both variables could be included in the analyses. All behavioral data collections follow the ethical guidelines of the Max Planck Society, of the Ministère de l’Enseignement Supérieur et de la Recherche Scientifique and the Ministère de Eaux et Forêts in Côte d’Ivoire, and of the Office Ivoirien des Parcs et Réserves.
Method Details
Food availability and neighbor pressure index
We calculated food availability on a monthly basis following a standard food availability index (FAI) [30], that combines chimpanzee food fruiting phenology scores (absence/presence of mature fruits), density of tree species and mean basal areas of each tree species. Densities and basal areas were measured separately for each chimpanzee community home-range, and phenology trails are established on each territory, so that differences between groups in terms of FAI not only consider differences in fruit productivity but also local density differences. Overall, there were no differences in the monthly FAI scores between the four study groups (one-way ANOVA: F3,526 = 1.323, p = 0.266). Since each group lives in territories of different sizes, we multiplied the FAI scores by the yearly territory sizes to take into account these between-group differences: ([FAI x territory size] / 10). We calculated yearly territory sizes based on 95% utilization distribution of all locations cumulated from 1st January to 31st December of each year. Locations were established during focal follows by using hand maps in which a grid of 500 m x 500 m cells was allocated.
To calculate the neighbor pressure index (NPI), we considered all types of inter-group encounters (physical: n = 103 – 55 involving physical aggression, 48 only visual contact; and vocal: n = 281, across all groups; Table S1), as both types are known to trigger a behavioral and physiological response [14, 28, 29]. Physical inter-group encounters correspond to those in close proximity where visual contact between opponents occurred even in the absence of fight or physical aggression (n = 48). For each inter-group encounter we extracted two metrics related to their location: (i) I: the extent of incursion of neighbors, measured as the distance between the location of the encounter and the center of the territory (measured for the last 12 months), relative to the distance to the border delimited by the kernel distribution of 75% of the locations (used as a cut-off point to delimit core areas [18]); (ii) K: the past usage by the target group during the last 12 months of the location of the inter-group encounter, measure based on the kernel distribution of the ranging locations expressed every 10%. We computed the product of these two metrics and used the mean product value across each specific time frame considered in the different models (8.5 months of pregnancy for survival analysis, female-specific for inter-birth interval analysis), which we multiplied by the mean frequency of occurrence of the inter-group encounter (F) based on the number of observation days between consecutive encounters, in order to control for variation in the observation effort. The three components vary in the same direction, resulting into large values of the index associated with a stronger neighbor pressure. Correlations coefficients (Pearson) between the three components, based on the all inter-group encounters, are: I – K (0.53), I – F (0.004), K – F (−0.02).
Offspring survival and inter-birth interval
For each group, we selected the offspring born during the study period for which we could access the variables of interest during the 8.5 months of their mother’s pregnancy (North n = 24 offspring; Middle n = 4 offspring; South n = 33 offspring; East n = 20 offspring). We did not include offspring that died the same month than their mother died. Orphans were included in the dataset (n = 13, among which 3 died). Emigrant individuals were considered as alive at the age of departure (n = 2). Results of the survival analysis based on pregnancy periods are provided in Tables 1 and S2 and Data S2.
For the survival analysis based on the lactation period (Data S1), we included for the first year 80 offspring (North n = 23; Middle n = 3; South n = 34; East n = 20), for the two first years 70 offspring (North n = 21; Middle n = 3; South n = 28; East n = 18) and for the three first years 58 offspring (North n = 19; Middle n = 1; South n = 24; East n = 14). The events considered were deaths. All individuals were followed during their lifespan and continuously censored. Inter-birth intervals were defined as the number of days between two consecutive births for the same mother, under the condition that the former offspring reached at least three years old. During each inter-birth interval, the period during which a female could potentially become receptive again and conceive was chosen as the specific period for each female between the 3rd year after the last birth to the predicted month of the next conception (last birth + 3 years, to next conception). This period varied between 1 and 4 years across groups. The choice of three years after the last birth makes our study comparable to studies conducted in Gombe [42]. We obtained 71 inter-birth intervals in total (22 for 10 females in North, 35 for 21 females in South, 2 for 2 females in Middle and 12 for 10 females in East).
Quantification and Statistical Analysis
To analyze the effect of between-group competition and within-group competition on offspring survival, we conducted a mixed-effects Cox proportional hazards model, using the R packages “survival” (version 2.44-1) [71], and “coxme” (version 2.2-10) [72]. Across the pregnancy period of the mother and across the three time windows related to the lactation period, we computed the mean values of the different covariates. Test predictors included the number of mature males in the group, neighbor pressure, the number of weaned individuals (proxy for within-group competition) and food availability multiplied by territory size. We controlled for the rank of the mother, her age at offspring birth and the sex of the infant. For the lactation periods, we took the age of the mother at the end of the considered period (after 1, 2 and 3 years). We included mother and group identity as random effects. Estimates and p values are provided in the main text; coefficients, standard-error and 95% confidence intervals in Table 1, while hazard ratios, means and standard-deviation of the original values are provided in the Supplemental Information. Confidence intervals of the hazard ratios (always positive) were obtained using the function coxph from the “survival” R package. Results of the survival analyses during lactation are provided in Data S1A, S1B, and S1C.
We fitted a linear mixed model (LMM) with Gaussian structure using the function lmer of the R-package “lme4” (version 1.1-14) [73] to establish the influence of key predictors on the length of inter-birth intervals. For each female-specific period, we averaged the values of the covariates across the given periods. The test and control predictors in this analysis were identical to the survival analysis, however, as females of different ranks may respond differently, we included the interactions of dominance rank with all other predictors. None of the interactions was significant, so to investigate each covariate as main effects, we ran a reduced model without the interactions. Estimates and p values are provided in the main text; Chisq value and 95% confidence intervals in Table 2, while random slopes variances, mean and standard-deviation of original values and effect sizes are provided in Table S4. Significance threshold was set for p values at 0.05.
To keep type I error rates at the nominal level of 5%, we included in each model the random slopes of test predictors within the random effect of “mother identity” and “group identity,” but not the correlation parameters between random intercepts and random slopes terms, as in each model these correlations were not identifiable [82, 83]. Continuous predictors were z-transformed to a mean of zero and a standard deviation of one. Models were fitted using R version 3.4.0. The assumptions of normally distributed and homogeneous residuals were assessed by visually inspecting a qqplot and a plot of residuals against fitted values. None of these plots indicated obvious deviation from these assumptions. We checked for model stability by excluding each level of the random effect one at a time from the data and comparing the model estimates derived from these subsets of the data with those derived from the full dataset. This revealed that our models are stable. Variance Inflation Factors (VIF) [84] were derived using the function vif from the R-package ”car” (version 3.0-2) [74] applied to a standard linear model excluding the random effects. Collinearity was not problematic in either model. In order to establish the significance of the full as compared to the null models (comprising only control predictors, random effects and slopes), we used likelihood ratio test (R function anova with argument test set to “Chisq” [85]). To allow for a likelihood ratio test, we fitted the models using Maximum Likelihood rather than Restricted Maximum Likelihood [85]. P values for the individual effects were based on likelihood ratio tests comparing the full with respective reduced models ([48] R function drop1). Ad hoc model comparisons were carried on using the R-package “MuMin” (version 1.42-1) [75] with the function model.sel and the package “AICcmodavg” (version 2.2-1) [76] with the function aictab. We processed model comparisons in the survival analysis based on pregnancy and in the inter-birth interval model, by comparing models which include the neighbor pressure index with subsequent models replacing the index by each of its components (Model I: intrusion degree; Model K: kernel of location; and Model F: rate of inter-group encounters). Not all pregnant females were present during all inter-group encounters. However, the nature of chimpanzee fission-fusion society and the loudness of the vocal exchanges make it problematic to exclude the possibility that those not present nonetheless heard the encounter. Also, those not present would nonetheless be restricted by any subsequent territory use limitations. Thus, we used a conservative approach using all inter-group encounters that occurred during pregnancy. To test this rationale, we also provide a complementary model comparison in the survival analysis based on the pregnancy period, in which we compare the model including the neighbor pressure index to subsequent models replacing the index by the total number of inter-group encounters (Model N), the total number of physical encounters (Model P), the number of all types of encounters in which specific pregnant females were present (Model SN) and the number of physical encounters in which specific pregnant females were present (Model SP). Specific effects of each variable for these different models can be found in Data S2, while results of this model comparison are provided in Table S3.
Data and Code Availability
The datasets and codes supporting the current study have not been deposited in a public repository because these are part of further investigation, but they are available from the corresponding author on request.
Acknowledgments
We are grateful to the Ministère de l’Enseignement Supérieur et de la Recherche Scientifique of Côte d’Ivoire and the Office Ivorien des Parcs et Réserves for permissions to conduct the research and also to the Centre Suisse de Recherches Scientifiques and the staff members of the Taï Chimpanzee Project for their long-term support. This study was funded by the Max Planck Society and the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement no. 679787).
Author Contributions
Conceptualization, S.L. and R.M.W.; Methodology, S.L., C.C., and R.M.W.; Investigation, S.L.; Writing – Original Draft, S.L.; Writing – Review & Editing, S.L., L.S., A.P., C.B., C.C., and R.M.W.; Funding Acquisition, C.C. and R.M.W.; Resources, S.L., L.S., A.P., C.B., C.C., and R.M.W.; Supervision, C.C. and R.M.W.
Declaration of Interests
The authors declare no competing interests.
Published: January 2, 2020
Footnotes
Supplemental Information can be found online at https://doi.org/10.1016/j.cub.2019.11.039.
Contributor Information
Sylvain Lemoine, Email: sylvain_lemoine@eva.mpg.de.
Roman M. Wittig, Email: wittig@eva.mpg.de.
Supplemental Information
(A) During the first year of lactation. (B) During the two first years of lactation. (C) During the three first years of lactation.
Each model differs only by the component related to inter-group encounters.
References
- 1.Gaston A.J. The evolution of group territorial behavior and cooperative breeding. Am. Nat. 1978;112:1091–1100. [Google Scholar]
- 2.Radford A.N., Majolo B., Aureli F. Within-group behavioural consequences of between-group conflict: a prospective review. Proc. Biol. Sci. 2016;283:20161567. doi: 10.1098/rspb.2016.1567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Choi J.K., Bowles S. The coevolution of parochial altruism and war. Science. 2007;318:636–640. doi: 10.1126/science.1144237. [DOI] [PubMed] [Google Scholar]
- 4.Bowles S. Did warfare among ancestral hunter-gatherers affect the evolution of human social behaviors? Science. 2009;324:1293–1298. doi: 10.1126/science.1168112. [DOI] [PubMed] [Google Scholar]
- 5.Clutton-Brock T., Sheldon B.C. Individuals and populations: the role of long-term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol. Evol. 2010;25:562–573. doi: 10.1016/j.tree.2010.08.002. [DOI] [PubMed] [Google Scholar]
- 6.Parker G.A. Assessment strategy and the evolution of fighting behaviour. J. Theor. Biol. 1974;47:223–243. doi: 10.1016/0022-5193(74)90111-8. [DOI] [PubMed] [Google Scholar]
- 7.Hammerstein P. The role of asymmetries in animal contests. Anim. Behav. 1981;29:193–205. [Google Scholar]
- 8.Crofoot M.C., Wrangham R.W. Intergroup aggression in primates and humans: the case for a unified theory. In: Kappeler P.M., Silk J., editors. Mind the Gap. Springer Berlin Heidelberg; 2010. pp. 171–195. [Google Scholar]
- 9.Markham A.C., Alberts S.C., Altmann J. Intergroup conflict: ecological predictors of winning and consequences of defeat in a wild primate population. Anim. Behav. 2012;82:399–403. doi: 10.1016/j.anbehav.2012.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Boesch C., Boesch-Achermann H. Oxford Univ. Press; 2000. The Chimpanzees of the Taï Forest: Behavioural Ecology and Evolution. [Google Scholar]
- 11.Wittig R.M. Taï chimpanzees. In: Vonk J., Shackelford T., editors. Encyclopedia of Animal Cognition and Behavior. Springer International Publishing; 2018. pp. 1–7. [Google Scholar]
- 12.Wilson M.L., Wrangham R.W. Intergroup relations in chimpanzees. Annu. Rev. Anthropol. 2003;32:363–392. [Google Scholar]
- 13.Wilson M.L., Boesch C., Fruth B., Furuichi T., Gilby I.C., Hashimoto C., Hobaiter C.L., Hohmann G., Itoh N., Koops K. Lethal aggression in Pan is better explained by adaptive strategies than human impacts. Nature. 2014;513:414–417. doi: 10.1038/nature13727. [DOI] [PubMed] [Google Scholar]
- 14.Samuni L., Preis A., Mundry R., Deschner T., Crockford C., Wittig R.M. Oxytocin reactivity during intergroup conflict in wild chimpanzees. Proc. Natl. Acad. Sci. USA. 2017;114:268–273. doi: 10.1073/pnas.1616812114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mitani J.C., Watts D.P. Correlates of territorial boundary patrol behaviour in wild chimpanzees. Anim. Behav. 2005;70:1079–1086. [Google Scholar]
- 16.Hasegawa M., Kutsukake N. Bayesian competitiveness estimation predicts dominance turnover among wild male chimpanzees. Behav. Ecol. Sociobiol. 2015;69:89–99. [Google Scholar]
- 17.Port M., Kappeler P.M., Johnstone R.A. Communal defense of territories and the evolution of sociality. Am. Nat. 2011;178:787–800. doi: 10.1086/662672. [DOI] [PubMed] [Google Scholar]
- 18.Lehmann J., Boesch C. Social influences on ranging patterns among chimpanzees (Pan troglodytes verus) in the Taï National Park, Côte d’Ivoire. Behav. Ecol. 2003;14:642–649. [Google Scholar]
- 19.Crofoot M.C., Gilby I.C., Wikelski M.C., Kays R.W. Interaction location outweighs the competitive advantage of numerical superiority in Cebus capucinus intergroup contests. Proc. Natl. Acad. Sci. USA. 2008;105:577–581. doi: 10.1073/pnas.0707749105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cheney D.L. Intragroup cohesion and intergroup hostility: the relation between grooming distributions and intergroup competition among female primates. Behav. Ecol. 1992;3:334–345. [Google Scholar]
- 21.Mosser A., Packer C. Group territoriality and the benefits of sociality in the African lion, Panthera leo. Anim. Behav. 2009;78:359–370. [Google Scholar]
- 22.Grueter C.C. Home range overlap as a driver of intelligence in primates. Am. J. Primatol. 2015;77:418–424. doi: 10.1002/ajp.22357. [DOI] [PubMed] [Google Scholar]
- 23.Hyman J., Hughes M., Searcy W.A., Nowicki S. Individual variation in the strength of territory defense in male song sparrows: correlates of age, territory tenure, and neighbor aggressiveness. Behaviour. 2004;141:15–27. [Google Scholar]
- 24.Bailey D.H., Geary D.C. Hominid brain evolution: testing climatic, ecological, and social competition models. Hum. Nat. 2009;20:67–79. [Google Scholar]
- 25.Smith J.M., Parker G.A. The logic of asymmetric contests. Anim. Behav. 1976;24:159–175. [Google Scholar]
- 26.Kokko H., López-Sepulcre A., Morrell L.J. From hawks and doves to self-consistent games of territorial behavior. Am. Nat. 2006;167:901–912. doi: 10.1086/504604. [DOI] [PubMed] [Google Scholar]
- 27.Burnham K.P., Anderson D.R., Huyvaert K.P. AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011;65:23–35. [Google Scholar]
- 28.Wittig R.M., Crockford C., Weltring A., Langergraber K.E., Deschner T., Zuberbühler K. Social support reduces stress hormone levels in wild chimpanzees across stressful events and everyday affiliations. Nat. Commun. 2016;7:13361. doi: 10.1038/ncomms13361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Samuni L., Preis A., Deschner T., Wittig R.M., Crockford C. Cortisol and oxytocin show independent activity during chimpanzee intergroup conflict. Psychoneuroendocrinology. 2019;104:165–173. doi: 10.1016/j.psyneuen.2019.02.007. [DOI] [PubMed] [Google Scholar]
- 30.Anderson D.P., Nordheim E.V., Moermond T.C., Gone Bi Z.B., Boesch C. Factors influencing tree phenology in Taï National Park, Côte d’Ivoire. Biotropica. 2005;37:631–640. [Google Scholar]
- 31.Nakamura M., Hayaki H., Hosaka K., Itoh N., Zamma K. Brief communication: orphaned male chimpanzees die young even after weaning. Am. J. Phys. Anthropol. 2014;153:139–143. doi: 10.1002/ajpa.22411. [DOI] [PubMed] [Google Scholar]
- 32.Boesch C., Bolé C., Eckhardt N., Boesch H. Altruism in forest chimpanzees: the case of adoption. PLoS ONE. 2010;5:e8901. doi: 10.1371/journal.pone.0008901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Samuni L., Wittig R.M., Crockford C. Adoption in the Tai chimpanzees: costs, benefits, and strong social relationships. In: Boesch C., Wittig R.M., editors. The Chimpanzees of the Taï Forest: 40 Years of Research. Cambridge University; 2019. pp. 141–158. [Google Scholar]
- 34.Clutton-Brock T.H., Albon S.D., Guinness F.E. Maternal dominance, breeding success and birth sex ratios in red deer. Nature. 1984;308:358–360. [Google Scholar]
- 35.Cassinello J., Alados C.L. Female reproductive success in captive Ammotragus lervia (Bovidae, Artiodactyla). Study of its components and effects of hierarchy and inbreeding. J. Zool. 1996;239:141–153. [Google Scholar]
- 36.Pusey A., Williams J., Goodall J. The influence of dominance rank on the reproductive success of female chimpanzees. Science. 1997;277:828–831. doi: 10.1126/science.277.5327.828. [DOI] [PubMed] [Google Scholar]
- 37.Altmann J., Alberts S.C. Variability in reproductive success viewed from a life-history perspective in baboons. Am. J. Hum. Biol. 2003;15:401–409. doi: 10.1002/ajhb.10157. [DOI] [PubMed] [Google Scholar]
- 38.Pluháček J., Bartoš L., Čulík L. High-ranking mares of captive plains zebra Equus burchelli have greater reproductive success than low-ranking mares. Appl. Anim. Behav. Sci. 2006;99:315–329. [Google Scholar]
- 39.Stockley P., Bro-Jørgensen J. Female competition and its evolutionary consequences in mammals. Biol. Rev. Camb. Philos. Soc. 2011;86:341–366. doi: 10.1111/j.1469-185X.2010.00149.x. [DOI] [PubMed] [Google Scholar]
- 40.Majolo B., Lehmann J., de Bortoli Vizioli A., Schino G. Fitness-related benefits of dominance in primates. Am. J. Phys. Anthropol. 2012;147:652–660. doi: 10.1002/ajpa.22031. [DOI] [PubMed] [Google Scholar]
- 41.Gélin U., Wilson M.E., Coulson G., Festa-Bianchet M. Experimental manipulation of female reproduction demonstrates its fitness costs in kangaroos. J. Anim. Ecol. 2015;84:239–248. doi: 10.1111/1365-2656.12266. [DOI] [PubMed] [Google Scholar]
- 42.Williams J.M., Oehlert G.W., Carlis J.V., Pusey A.E. Why do male chimpanzees defend a group range? Anim. Behav. 2004;68:523–532. [Google Scholar]
- 43.Thompson M.E., Kahlenberg S.M., Gilby I.C., Wrangham R.W. Core area quality is associated with variance in reproductive success among female chimpanzees at Kibale National Park. Anim. Behav. 2007;73:501–512. [Google Scholar]
- 44.Murray C.M., Eberly L.E., Pusey A.E. Foraging strategies as a function of season and rank among wild female chimpanzees (Pan troglodytes) Behav. Ecol. 2006;17:1020–1028. [Google Scholar]
- 45.Setchell J.M. Alternative reproductive tactics in primates. In: Oliveira R.F., Taborsky M., Brockmann H.J., editors. Alternative Reproductive Tactics. Cambridge University Press; 2008. pp. 373–398. [Google Scholar]
- 46.Trivers R.L., Willard D.E. Natural selection of parental ability to vary the sex ratio of offspring. Science. 1973;179:90–92. doi: 10.1126/science.179.4068.90. [DOI] [PubMed] [Google Scholar]
- 47.Boesch C. Evidence for dominant wild female chimpanzees investing more in sons. Anim. Behav. 1997;54:811–815. doi: 10.1006/anbe.1996.0510. [DOI] [PubMed] [Google Scholar]
- 48.Baayen R.H. Cambridge University Press; 2011. Analyzing Linguistic Data: a Practical Introduction to Statistics Using R. [Google Scholar]
- 49.Deschner T., Boesch C. Can the patterns of sexual swelling cycles in female Taï chimpanzees be explained by the cost-of-sexual-attraction hypothesis? Int. J. Primatol. 2007;28:389–406. [Google Scholar]
- 50.Sillett T.S., Rodenhouse N.L., Holmes R.T. Experimentally reducing neighbor density affects reproduction and behavior of a migratory songbird. Ecology. 2004;85:2467–2477. [Google Scholar]
- 51.Lupien S.J., McEwen B.S., Gunnar M.R., Heim C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009;10:434–445. doi: 10.1038/nrn2639. [DOI] [PubMed] [Google Scholar]
- 52.Sobolewski M.E., Brown J.L., Mitani J.C. Territoriality, tolerance and testosterone in wild chimpanzees. Anim. Behav. 2012;84:1469–1474. [Google Scholar]
- 53.Murray C.M., Stanton M.A., Wellens K.R., Santymire R.M., Heintz M.R., Lonsdorf E.V. Maternal effects on offspring stress physiology in wild chimpanzees. Am. J. Primatol. 2018;80:e22525. doi: 10.1002/ajp.22525. [DOI] [PubMed] [Google Scholar]
- 54.Entringer S., Buss C., Wadhwa P.D. Prenatal stress, development, health and disease risk: a psychobiological perspective-2015 Curt Richter Award Paper. Psychoneuroendocrinology. 2015;62:366–375. doi: 10.1016/j.psyneuen.2015.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lea A.J., Altmann J., Alberts S.C., Tung J. Resource base influences genome-wide DNA methylation levels in wild baboons (Papio cynocephalus) Mol. Ecol. 2016;25:1681–1696. doi: 10.1111/mec.13436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Köndgen S., Kühl H., N’Goran P.K., Walsh P.D., Schenk S., Ernst N., Biek R., Formenty P., Mätz-Rensing K., Schweiger B. Pandemic human viruses cause decline of endangered great apes. Curr. Biol. 2008;18:260–264. doi: 10.1016/j.cub.2008.01.012. [DOI] [PubMed] [Google Scholar]
- 57.Hoffmann C., Zimmermann F., Biek R., Kuehl H., Nowak K., Mundry R., Agbor A., Angedakin S., Arandjelovic M., Blankenburg A. Persistent anthrax as a major driver of wildlife mortality in a tropical rainforest. Nature. 2017;548:82–86. doi: 10.1038/nature23309. [DOI] [PubMed] [Google Scholar]
- 58.Mitani J.C., Watts D.P., Amsler S.J. Lethal intergroup aggression leads to territorial expansion in wild chimpanzees. Curr. Biol. 2010;20:R507–R508. doi: 10.1016/j.cub.2010.04.021. [DOI] [PubMed] [Google Scholar]
- 59.Wessling E.G., Deschner T., Mundry R., Pruetz J.D., Wittig R.M., Kühl H.S. Seasonal variation in physiology challenges the notion of chimpanzees (Pan troglodytes verus) as a forest-adapted species. Front. Ecol. Evol. 2018;6:60. [Google Scholar]
- 60.Boesch C., Crockford C., Herbinger I., Wittig R., Moebius Y., Normand E. Intergroup conflicts among chimpanzees in Taï National Park: lethal violence and the female perspective. Am. J. Primatol. 2008;70:519–532. doi: 10.1002/ajp.20524. [DOI] [PubMed] [Google Scholar]
- 61.Koenig A. Competition for resources and its behavioral consequences among female primates. Int. J. Primatol. 2002;23:759–783. [Google Scholar]
- 62.Cheney D.L., Seyfarth R.M. The influence of intergroup competition on the survival and reproduction of female vervet monkeys. Behav. Ecol. Sociobiol. 1987;21:375–386. [Google Scholar]
- 63.Clutton-Brock T.H., Parker G.A. Sexual coercion in animal societies. Anim. Behav. 1995;49:1345–1365. [Google Scholar]
- 64.Hinsch M., Komdeur J. Defence, intrusion and the evolutionary stability of territoriality. J. Theor. Biol. 2010;266:606–613. doi: 10.1016/j.jtbi.2010.07.033. [DOI] [PubMed] [Google Scholar]
- 65.Puurtinen M., Heap S., Mappes T. The joint emergence of group competition and within-group cooperation. Evol. Hum. Behav. 2015;36:211–217. [Google Scholar]
- 66.Flinn M.V., Geary D.C., Ward C.V. Ecological dominance, social competition, and coalitionary arms races: why humans evolved extraordinary intelligence. Evol. Hum. Behav. 2005;26:10–46. [Google Scholar]
- 67.De Dreu C.K.W. Oxytocin modulates cooperation within and competition between groups: an integrative review and research agenda. Horm. Behav. 2012;61:419–428. doi: 10.1016/j.yhbeh.2011.12.009. [DOI] [PubMed] [Google Scholar]
- 68.Langergraber K.E., Watts D.P., Vigilant L., Mitani J.C. Group augmentation, collective action, and territorial boundary patrols by male chimpanzees. Proc. Natl. Acad. Sci. USA. 2017;114:7337–7342. doi: 10.1073/pnas.1701582114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Eller E., Hawks J., Relethford J.H. Local extinction and recolonization, species effective population size, and modern human origins. 2004. Hum. Biol. 2009;81:805–824. doi: 10.3378/027.081.0623. [DOI] [PubMed] [Google Scholar]
- 70.Boyd R., Richerson P.J. Culture and the evolution of human cooperation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009;364:3281–3288. doi: 10.1098/rstb.2009.0134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Therneau T.M., Lumley T. Package ‘survival’: survival analysis. 2014. https://tbrieder.org/epidata/course_reading/e_therneau.pdf
- 72.Therneau T.M. Package ‘coxme’: mixed effects Cox models. 2019. https://cran.r-project.org/web/packages/coxme/coxme.pdf
- 73.Bates D., Mächler M., Bolker B., Walker S. Fitting linear mixed-effects models using lme4. arXiv. 2014 https://arxiv.org/abs/1406.5823 arXiv:1406.5823v1. [Google Scholar]
- 74.Fox J., Weisberg S. Second Edition. Sage; 2011. An R Companion to Applied Regression. [Google Scholar]
- 75.Bartoń K. Package ‘MuMin’: multi-model inference. 2019. https://cran.r-project.org/web/packages/MuMIn/index.html
- 76.Mazerolle M.J. Package ‘AICcmodavg’: Model selection and multimodel inference based on (Q)AIC(c) 2019. https://cran.r-project.org/web/packages/AICcmodavg/index.html
- 77.Goodall J. Belknap Press of Harvard University; 1986. The Chimpanzees of Gombe: Patterns of Behavior. [Google Scholar]
- 78.Nishida T. Tokyo University Press; 1990. The Chimpanzees of the Mahale Mountains: Sexual and Life History Strategies. [Google Scholar]
- 79.Boesch C., Kohou G., Néné H., Vigilant L. Male competition and paternity in wild chimpanzees of the Taï forest. Am. J. Phys. Anthropol. 2006;130:103–115. doi: 10.1002/ajpa.20341. [DOI] [PubMed] [Google Scholar]
- 80.Neumann C., Duboscq J., Dubuc C., Ginting A., Maulana A., Agil M., Widdig A., Engelhardt A. Assessing dominance hierarchies: validation and advantages of progressive evaluation with Elo-rating. Anim. Behav. 2011;82:911–921. [Google Scholar]
- 81.Wittig R.M., Boesch C. Food competition and linear dominance hierarchy among female chimpanzees of the Taï National Park. Int. J. Primatol. 2003;24:847–867. [Google Scholar]
- 82.Schielzeth H., Forstmeier W. Conclusions beyond support: overconfident estimates in mixed models. Behav. Ecol. 2009;20:416–420. doi: 10.1093/beheco/arn145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Barr D.J., Levy R., Scheepers C., Tily H.J. Random effects structure for confirmatory hypothesis testing: keep it maximal. J. Mem. Lang. 2013;68:255–278. doi: 10.1016/j.jml.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Field A. Sage; 2009. Discovering Statistics Using SPSS, Third Edition. [Google Scholar]
- 85.Dobson A.J., Barnett A.G. Third Edition. CRC; 2008. An Introduction to Generalized Linear Models. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) During the first year of lactation. (B) During the two first years of lactation. (C) During the three first years of lactation.
Each model differs only by the component related to inter-group encounters.
Data Availability Statement
The datasets and codes supporting the current study have not been deposited in a public repository because these are part of further investigation, but they are available from the corresponding author on request.