

Abstract
The nucleus accumbens (NAc) is a subcortical brain structure known primarily for its roles in pleasure, reward, and addiction. Despite less focus on the NAc in pain research, it also plays a large role in the mediation of pain and is effective as a source of analgesia. Evidence for this involvement lies in the NAc’s cortical connections, functions, pharmacology, and therapeutic targeting. The NAc projects to and receives information from notable pain structures, such as the prefrontal cortex, anterior cingulate cortex, periaqueductal gray, habenula, thalamus, etc. Additionally, the NAc and other pain-modulating structures share functions involving opioid regulation and motivational and emotional processing, which each work beyond simply the rewarding experience of pain offset. Pharmacologically speaking, the NAc responds heavily to painful stimuli, due to its high density of μ opioid receptors and the activation of several different neurotransmitter systems in the NAc, such as opioids, dopamine, calcitonin gene-related peptide, γ-aminobutyric acid, glutamate, and substance P, each of which have been shown to elicit analgesic effects. In both preclinical and clinical models, deep brain stimulation of the NAc has elicited successful analgesia. The multi-functional NAc is important in motivational behavior, and the motivation for avoiding pain is just as important to survival as the motivation for seeking pleasure. It is possible, then, that the NAc must be involved in both pleasure and pain in order to help determine the motivational salience of positive and negative events.
Keywords: analgesia, circuitry, deep brain stimulation, nociception, nucleus accumbens, pain, pain relief, pain signaling, reward, striatum
Introduction
The nucleus accumbens (NAc) is a subcortical brain structure located within the ventral striatum. It is often referred to as the pleasure center of the brain for its primary role in positive affect and feelings of reward, but this is an incomplete description. There is also evidence of a role in motivation, behavior, and a limbic-motor interface (Floresco, 2015; Salgado and Kaplitt, 2015). Another well-known role of the NAc is in addiction and reactivity to several psychoactive drugs. Because of these various functions, the NAc has been implicated in numerous other psychological disorders, such as addiction, obsessive-compulsive disorder, anxiety, depression, Alzheimer’s disease, Huntington’s disease, Parkinson’s disease, and obesity (Salgado and Kaplitt, 2015). Its wide range of neural connections has led to an increase in research prevalence in recent years, particularly as a target for potential treatments of both physiological and psychological disorders. However, an important aspect of NAc functioning that is frequently overlooked is pain.
Pain is a predominant issue in modern healthcare; it can be an aversive sensory experience and is highly comorbid with injury and disease. The evolutionary benefit of pain that allows the detection and prevention of dangerous encounters can quickly become maladaptive in cases of chronic pain, lasting beyond treatment and preventing a return to normal functioning. Current pain management methods are not well suited to handle long-term pain treatment, and many pain medications come with extreme adverse side effects even in short term use, such as addiction and overdose. New developments in pain treatment are difficult to research because of the particularly complex nature of pain neurophysiology. This review explores the neural connectivity of the NAc in attempt to ascertain its involvement in pain and pain treatment.
Search Strategy
Articles cited in this neurological review were found via electronic searches of the following databases: Academic Search Complete, Medline, Science Direct, and PubMed. Searches were conducted using the phrase nucleus accumbens combined with the following key words: pain, analgesia, nociception, antinociception, reward circuitry, pain circuitry, and pain signaling. Articles were included if they were determined to contribute to the understanding of the nucleus accumbens and the neuroscience of pain processing.
Afferents and Efferents
Initial evidence of the role of the NAc in pain processing comes from its connections to other brain structures known for their roles in pain modulation. The NAc receives dopaminergic projections from the ventral tegmental area and substantia nigra and glutamatergic projections from the prefrontal cortex, amygdala, thalamus, prelimbic cortex, hippocampus, and subiculum (Gupta and Young, 2018; Li et al., 2018). The anatomically distinct shell and core of the NAc have additional distinctions in their input regions. Afferents to the core include the perirhinal cortices, dorsal prelimbic cortex, the substantia nigra, and the anterior cingulate cortex (Salgado and Kaplitt, 2015; Li et al., 2018). Afferents to the shell include, the ventromedial prefrontal cortex, the basolateral amygdala, and the lateral hypothalamus (Salgado and Kaplitt, 2015; Li et al., 2018). The core and the shell of the NAc both receive afferent input from the ventral tegmental area (Li et al., 2018). However, the core receives projections primarily from the lateral region of the ventral tegmental area, whereas the shell receives projections primarily from the medial region of the ventral tegmental area (Li et al., 2018). Of note, researchers have found multitudes of neurons from the spinal cord that directly innervate the NAc via retrograde labeling, suggesting a strong link to incoming pain messages (Gear and Levine, 2011; Salgado and Kaplitt, 2015).
Efferents from the NAc tend to travel to areas within the diencephalon and pallidal complex, such as the nucleus mediodorsalis thalami, stria terminalis, lateral habenular nucleus, lateral hypothalamus, substantia nigra, the globus pallidus, and the subpallidal region (Bálint et al., 2011; Salgado and Kaplitt, 2015). The core is known to project to areas such as the dorsolateral region of the ventral pallidum, whereas the shell is known to project to the rostral-caudal regions of the lateral hypothalamus, extended amygdala, and ventromedial region of the ventral pallidum (Bálint et al., 2011; Salgado and Kaplitt, 2015). Both core and shell have been found to project to the entopeduncular nucleus, a region of the rodent brain considered to be equivalent to the human globus pallidus.
Links to areas known to be involved in pain processing, particularly the anterior cingulate cortex, prefrontal cortex, amygdala, somatosensory cortex, thalamus, and indeed the spinal cord itself as mentioned previously, suggest that the NAc receives nociceptive input (Bálint et al., 2011; Gear and Levine, 2011; Salgado and Kaplitt, 2015; Li et al., 2018). Lateral regions of the habenula have been shown to receive projections from both the medial frontal cortex and the NAc (Bianco and Wilson, 2009). Connections between these pain-related brain structures and the NAc suggest an NAc contribution to pain signaling. The NAc efferents to the amygdala and the ventral pallidum are noteworthy, as they have each been shown to be modulators of pain in previous and recent research (Kato et al., 2016).
Connectivity of both the NAc core and shell include feedback loops with areas involved in pain processing. This suggests that the NAc receives nociceptive input and projects nociceptive signals along the pain pathway (Seminowicz et al., 2019). From this, it can be deduced that the NAc plays an active and important role in multiple aspects of pain, and may even be a critical structure for the neural oscillations involved in pain signaling (Rauschecker et al., 2015). Connections with a wide variety of areas involved in sensation, emotion, and cognition could indicate that the NAc serves as a waystation for multiple components of pain processing, as pictured in Figure 1. The NAc and other structures of the reward pathway could be highly active in pain processing, due to the inherently rewarding aspect of relief from pain.
Figure 1.

Cortical connectivity of the nucleus accumbens.
Highlighted are afferent, efferent, and reciprocal connections of the nucleus accumbens and other pain-related brain structures, including excitatory (red) and inhibitory (blue) connections and neurotransmitter systems. ACC: Anterior cingulate cortex; AMY: amygdala; BG: basal ganglia; GABA: γ-aminobutyric acid; L Hyp: left hypothalamus; PAG: periaqueductal gray; PFC: prefrontal cortex; Thal: thalamus; SN: substantia nigra; VP: ventral pallidum; VTA: ventral tegmental area.
Functions
NAc functions also serve as particularly strong evidence for NAc involvement in pain processing. Specifically, the NAc and other pain-related structures engage in opioid regulation functions, which involve analgesia and reward. For instance, sites that have been identified as important in processing morphine-based antinociception include the nucleus raphe magnus, the locus coeruleus, the amygdala, the substantia nigra, the nucleus reticularis gigantocellularis, the periaqueductal gray (PAG), the habenula, and the NAc. It is logical to assert that these structures do not act independently in their mediation of analgesia. Evidence for this lies in morphine microinjection research, in which cannula injections of morphine increase the pain thresholds of rabbits and rats when targeted to either the PAG, the habenula, or the NAc (Ma et al., 1992; Khalilzadeh and Saiah, 2017). Similarly, naloxone, a known opioid antagonist, will negate the antinociceptive effects of morphine when its microinjection is targeted to any one of the above structures, including the NAc alone (Dill and Costa, 1977; Saigusa et al., 2017). Previous literature indicates a bidirectional loop of pain modulation between the PAG, the habenula, and the NAc (Ma et al., 1992; Borsook et al., 2016). Evidence for this statement comes from measurement of rabbits’ escape response latency, which increased after injections of morphine into the PAG, habenula, and NAc, and then significantly decreased again with the introduction of naloxone to each of those areas. The authors argued for the existence of an analgesia-related pathway between these structures due to effects of analgesia from morphine that had been injected into the NAc, for instance, could be negated by injecting naloxone into the habenula (Ma et al., 1992). With this research, the authors claimed to prove not only bidirectional pathways between the three structures, but also an ascending, analgesia-modulating pathway between them not previously known. Connections between these structures is illustrated in Figure 2.
Figure 2.
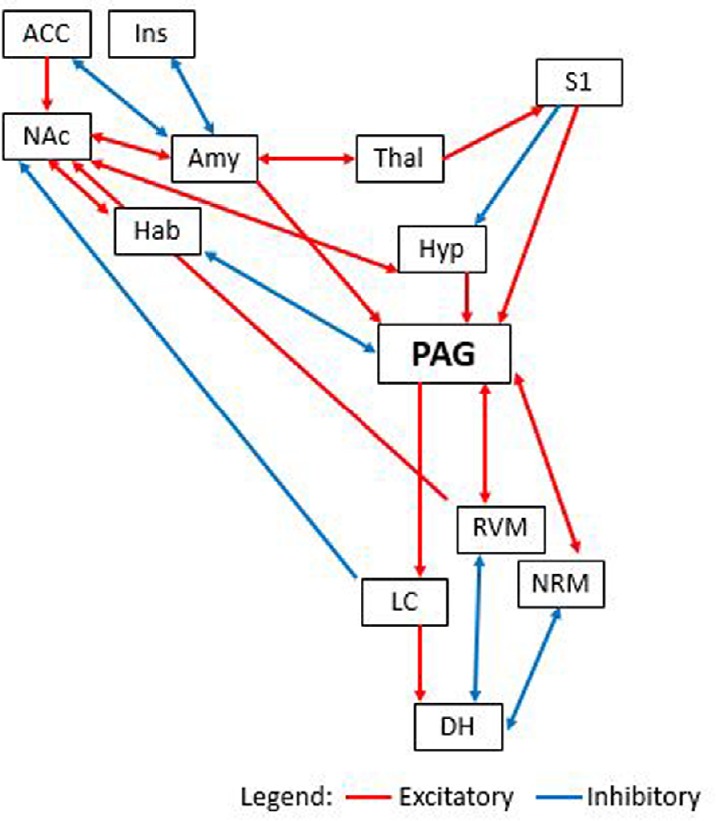
Cortical connections between the nucleus accumbens and periaqueductal gray.
This figure represents both ascending and descending pain pathways, highlighting the nucleus accumbens and periaqueductal gray involvement, including excitatory (red) and inhibitory (blue) connections. ACC: Anterior cingulate cortex; Amy: amygdala; DH: dorsal horn of the spinal cord; Hab: habenula; Hyp: hypothalamus; Ins: insular cortex; LC: locus coeruleus; NAc: nucleus accumbens; NRM: nucleus raphe magnus; PAG: periaqueductal gray; RVM: rostral ventral medulla; S1: primary somatosensory cortex; Thal: thalamus.
Research on synaptic activity during motivational and emotional processing reveals brain structures that are involved in motivational and emotional aspects of pain processing (Okun et al., 2016; Massaly et al., 2019). Behavioral patterns that arise from noxious or reward stimuli are also occurring in response to many different dimensions of information processing and internal regulation (Okun et al., 2016). Brain structures recently shown to be involved in these conflicts in motivational processing, such as the ventral tegmental area, the amygdala, the extended amygdala, and the NAc, are also involved in reward circuitry. Specifically, the aforementioned brain structures are considered vital for processing information and weighing conflicting values during emotional and motivational processing (Joseph et al., 2016). These motivational functions tie in closely with not only reward-seeking behavior, but also behaviors of avoiding aversive or dangerous stimuli (i.e., pain).
Structures in the reward pathway are most commonly considered to singularly involved in reward processing. However, researchers have postulated that aversive and rewarding stimuli might function as two sides of one behavioral spectrum (Shur and Ritvo, 1970; Hu, 2016). One study of reward and aversion involved exposure of human subjects to noxious and nonnoxious thermal stimuli during functional magnetic resonance imaging (Becerra et al., 2001). In a 46oC thermal application, the NAc was observed to have separate temporal phases of activation. The experimenters posit that differences between the noxious and nonnoxious thermal stimuli in timing of the activation could signify aversive versus noxious stimuli activations. The NAc and other reward regions primarily exhibited responses during the early phase activations, confirming previous suggestions of a primary function of stimulus evaluation regarding motivational dimensions (Breiter and Rosen, 1999). Increased signal in the early phase of activation was seen in the anterior portion of the NAc, whereas decreased signal was observed in the posterior portion during the later phase of the activation response. This signal decrease is in direct opposition to signal increases known to be associated with administration of morphine and cocaine in this region, which indicates additional processing of stimuli in the NAc, outside of reward evaluation, at the onset of pain stimuli.
Additional experiments have indicated endogenous opioid binding within the NAc of humans undergoing pain stimuli (Bodnar, 2018). These results corroborate results from studies of the NAc in various animals, implicating links to pain processing in the NAc (Kato et al., 2016). Studies involving blood-oxygen level dependency have revealed much more complex patterns in response to pain within the NAc (Becerra er al., 2001). The increased blood-oxygen level dependency signal during the early phases of pain stimulation and the decreased blood-oxygen level dependency signal during the later phases mentioned earlier has been seen in multiple studies, and has even been found in post hoc analyses of previous functional magnetic resonance imaging datasets (Becerra et al., 2001). The replicability of this result suggests that the positive or negative direction of the signal response in the NAc could be indicative of rewarding/aversive evaluation of the stimulus. In particular, the fact that this distinction in response times is also dispersed across locations within the structure of the NAc, with the early phase activating the anterior portion and the late phase activating the posterior portion, implies functional distinctions between these two sections (Becerra et al., 2001).
Additionally, it has been argued that the activation in the NAc in the early phase of pain activation is reflective of anticipation or evaluation functions (Kato et al., 2016; Shafiei et al., 2018). On the other hand, the activation in the late phase might work as an interpretation of stimulus valence and carry some analgesic functions (Shafiei et al., 2018). These results are also corroborated with animal studies (Kato et al., 2016). Evidence for these hypotheses include examples of healthy non-pain control groups receiving low doses of morphine, who were observed to have activation in the opposite direction (positive instead of negative) in the NAc (Becerra et al., 2001; Reakkamnuan et al., 2017). Additionally, studies that manipulate D1 and D2 dopamine agonists observe similar patterns of early and late phase activation (Shafiei et al., 2018). Finally, arguments that early phase activation in the NAc do not reflect simple valence or salience of motivational stimuli are supported with the theory that, mechanistically, the rewarding effects of various stimulants (amphetamine, morphine, cocaine, etc.) originate with the inhibition of inhibitory medium spiny neurons in the NAc, which lead to trans-synaptic, slow responses (Kohnomi et al., 2017).
As mentioned previously, the NAc is associated most strongly with activation of the reward system and addiction. In fact, even accumbal involvement in pain processing can be traced to reward and reinforcement. Survival behaviors that reduce the possibility of injury or damage are negatively reinforced (Navratilova et al., 2012). Unfortunately, NAc involvement in negative reinforcement is much less well researched than its involvement in positive reinforcement. What little research there is has shown definite negative reinforcement orientation in this region. A primary example is that pain offset and placebo-based analgesia can both activate overlapping brain regions, particularly those involved in reward and approach behavior, such as the ventral tegmental area and its dopamine projection to the NAc (Becerra and Borsook, 2008; Zubieta and Stohler, 2009). Previous literature implicates accumbal neurons in the signaling of reward salience and decision making in terms of reward-based approach behaviors (Saunders and Robinson, 2012). In light of these assertions, Navratilova and colleagues hypothesized that the experience of relief from persistent pain would result in activation of the mesolimbic dopaminergic pathway, a necessary step in the negative reinforcement of aforementioned survival behaviors (Navratilova et al., 2012). They tested their hypothesis by inducing surgical incision-based pain in rats, which was alleviated with peripheral nerve block. The researchers posit that negative reinforcement was demonstrated when the peripheral nerve block led to increased conditioned place preference. However, to definitively demonstrate that this mesolimbic dopaminergic pathway is truly necessary for peripheral nerve block-induced conditioned place preference, Navratilova and colleagues tested whether conditioned place preference could be blocked by inactivating the dopamine neurons in the ventral tegmental area. A microinjection of lidocaine, which inhibits passage fibers, injected into the ventral tegmental area at least 10 minutes before the peripheral nerve block did prevent the eventual conditioned place preference, suggesting a necessity of this dopamine transmission to the NAc in order for negative reinforcement to work and to affect behavior (Navratilova et al., 2012). These results confirm previous results in human subjects that demonstrate increasing accumbal activation at the relief of acute painful stimulus (Becerra and Borsook, 2008).
Pharmacological Mechanisms of Pain in the Nucleus Accumbens
Opioid
Opiate drugs help decrease perceptions of painful stimuli by mimicking the effect of endorphins, which serve as an endogenous opiate system. These drugs act prominently in specific areas of the midbrain, medulla, and pons to elicit antinociceptive effects, as identified by microinjection studies (Erfanparast et al., 2018). Whereas the first regions identified as strongly responsive to morphine were the PAG, locus coeruleus, and nucleus raphe magnus, the NAc was eventually shown to respond to similar antinociceptive methods, such as morphine microinjection and electrical stimulation (Hong et al., 2017; Yoshida et al., 2019). In fact, the first method used to determine the involvement of the locus coeruleus in analgesia was deep brain stimulation, a method later extended to the PAG and the NAc with equally successful results (Segal and Sandberg, 1977). It has also been determined that there exists a descending pathway involving noradrenaline that originates from the locus coeruleus and culminates in the spinal cord that is said to be responsible for modulation of spinal antinociception (Llorca-Torralba et al., 2016; Yoshida et al., 2019). In the nucleus raphe magnus, administering opiate drugs or electrically stimulating the area resulting in analgesic effect, and lesions to this area block the efficacy of analgesic relief of morphine (Campion et al., 2016; Yoshida et al., 2019). Additionally, studies on central microinjections of opioids suggest an involvement of the nucleus reticularis gigantocellularis in antinociception (Satoh et al., 1983).
Overall, opioid-based analgesia involves processes undergone by opioid receptor subtypes (Mizoguchi et al., 2014). There is strong evidence for delta and kappa opioid receptor-based analgesic processing in the spinal cord, whereas μ opioid receptors are most predominant in supraspinal analgesic processing. Two subtypes of μ receptors include the μ1 receptor, known to have high affinity binding to most opioids and enkephalins, and the μ2 receptor, which shows preferential affinity for binding to morphine (Mizoguchi et al., 2014). Sites of μ1 receptor binding have been shown to be involved in many opioid actions, supraspinal antinociception in particular (Mizoguchi et al., 2014). Increased levels of binding sites for μ1 have been identified in the PAG regions, and it has been posited that increased density of these binding sites results in increased possibility of sensitivity to opioid analgesics (Bodnar, 2018). The purpose of explaining the details of opioid processing in known pain processing regions is to demonstrate the synaptic similarities to the NAc. It is worth noting that the NAc also mediates opiate analgesia by way of μ-opioid receptors (Rosas et al., 2016). Research has shown that persistent pain increases the activity of opioids in the NAc (Bodnar, 2018). Morphine injection into the NAc has been shown to be sufficient to produce analgesia, suggesting an involvement of this structure in endogenous pain modulation (Kohnomi et al., 2017).
Dopamine
Dopamine is known to be highly involved in both reward and addiction. The substantia nigra and ventral tegmental area send dopamine projections to the NAc (Hikida et al., 2016). Projections from the latter part of the mesolimbic dopaminergic pathway appear to target the shell in particular, whereas the core appears to be innervated by the nigrostriatal system (Salgado and Kaplitt, 2015). Additionally, dopamine turnover is significantly higher in the NAc than the rest of the striatum (Schepisi et al., 2014).
The core has more dopaminergic transporters than the shell, and dopamine appears to have preferential binding to D2-type receptors in the core (Zhu et al., 2013; Salgado and Kaplitt, 2015). Conversely, the shell has a greater amount of dopamine receptors and opioids tend to cause more dopamine increase in the shell than they do in the core (Zhu et al., 2013; Salgado and Kaplitt, 2015). In pain processing, dopaminergic pathways are used in transmission between NAc, ventral tegmental area, and the medial prefrontal cortex.
Activation of or agonists for receptors of several different neurotransmitter systems in the NAc, such as calcitonin gene-related peptide (CGRP), neuropeptide Y, opioid, and acetylcholine, have all been shown to elicit analgesic effects (Desai et al., 2013; Yan and Yu, 2013). However, the neurotransmitter system most prevalent in this region is dopamine, the stimulation of which has been demonstrated to produce significant antinociception (Moradi et al., 2015). In fact, endogenous dopamine gets released in the NAc at the onset of noxious stimulation and modulates analgesia (Moradi et al., 2015). Additionally, injection of dopamine antagonists into the NAc are known to prevent the analgesia that would have been derived from the drug stimulation of the ventral tegmental area (Navratilova et al., 2012). Regardless of agonist specific to particular dopaminergic receptor subtypes, it appears that increased dopamine in the NAc results in analgesia. Studies on this topic have explored this effect from many different angles. For instance, one laboratory produced results demonstrating that bupivacaine, a local anesthetic, injected into the core of the NAc, actually enhances the nociceptive effect of formalin by inactivating the region, implicating accumbal involvement in ascending pain modulation (Magnusson and Martin, 2002). Interestingly, injection of another local anesthetic, lidocaine, into the NAc shell does not enhance the formalin effect, but alleviates pain (Magnusson and Martin, 2002; Chang et al., 2014). Magnusson and Martin (2002) suggested that this discrepancy might be due to the differential anatomy and connectivity of the core and shell, arguing that the core might be more involved in persistent behavior such as continuous pain, whereas the shell is more heavily involved in the onset of pain.
There is also strong evidence of two distinct dopaminergic innervations to the NAc, one rewarding and pleasurable, and the other aversive and negative (Lammel et al., 2011; Gentry et al., 2019). These networks are both responsible for evaluating affect and salience of stimuli, though they seem to respond to different types of stimuli (Hikida et al., 2010). There is one pathway associated with positive affect and reward that operates on direct spiny neurons. This direct pathway acts primarily on D1 dopamine receptors, and its activity is enhanced by increased dopamine at these receptors. The other pathway is associated with negative affect and aversion and operates on indirect spiny neurons. The activity of this indirect pathway is diminished when dopamine activates D2-type receptors (Hikida et al., 2010; Lammel et al., 2011; Ren et al., 2015).
Exogenous introduction of selective dopamine agonists into the NAc elicits analgesia. Similarly, injection of quinpirole, a selective D2 dopamine receptor agonist, blocked nociception during phase 2 of formalin stimulation in the same study (Taylor et al., 2003). The quinpirole might have accomplished this by causing interference with ascending pain messaging or modifying D2 neuron output (Escobar et al., 2015). Finally, the analgesic effect of the quinpirole could be reversed with induction of raclopride, a D2 dopamine receptor antagonist, solidifying the idea that stimulating D2 receptors in the NAc can block acute inflammatory pain (Taylor et al., 2003). Previous research suggests that this might be due to primary afferent C-fiber depletion (and to a lesser extent some Aδ-fibers), as shown in experiments with capsaicin in phase 2 specifically (Sheibani et al., 2010). Because Taylor and colleagues found that quinpirole worked specifically during phase 2 rather than phase 1 of formalin stimulation, it’s possible that accumbal D2 receptor agonism specifically targets noxious stimuli transmission via C-fibers (Taylor et al., 2003).
Additional literature corroborates these findings. For instance, one laboratory found that microinjections of raclopride into the NAc also blocked the analgesic effects of ventral tegmental area-injected morphine and NAc-injected amphetamine (Oswald et al., 2015). Additionally, another laboratory discovered that neuronal death of dopamine via 6-hydoxydopamine in the NAc prevents the analgesic effect of amphetamine (Clarke and Franklin, 1992). It has also been shown via microdialysis that various types of noxious stimuli, such as tail pinch and capsaicin in rats, elicit the release of dopamine in the NAc (Rougé-Pont et al., 1993; Schmidt et al., 2002; Dias et al., 2015).
Finally, there have been clinical studies of dopamine use in human subjects for the treatment of neuropathic pain. These studies include the use of levo-dopa, and have been demonstrated to be effective (Zeng et al., 2018). This suggests a likelihood of efficacy in persistent pain management with dopamine receptor agonists.
Calcitonin gene-related peptide
CGRP is a neuropeptide known widespread nervous system applications. CGRP has been shown to be important in certain aspects of pain modulation as well, such as the potentiation of excitatory signals induced by dorsal horn substance P (Bree and Levy, 2018). CGRP8–37, a selective CGRP receptor antagonist, can cause severe nociception in rats without previous painful stimulus, rats with inflammatory pain, and rats experiencing neuropathic pain (Yu et al., 2009). Opioid peptides and CGRP interactions have been observed during pain modulation in rat spinal cords. For instance, CGRP produced analgesic effects in the nucleus raphe magnus involving opioid receptors (Roeder et al., 2016). This is possibly indicative of some of the CGRP activity that occurs in the NAc, given its quantity of opioid receptors. Direct administration of CGRP into pain structures can alleviate pain, as demonstrated in experiments in which CGRP injections into the PAG increase hindpaw withdrawal latency (Roeder et al., 2016).
Morphological studies have indicated significantly high densities of receptor binding sites for CGRP in the NAc (Liu and Borgland, 2015). One study that involved the administration of CGRP into the NAc demonstrated significant increases in hindpaw withdrawal latency, proving a CGRP-mediated analgesic effect in the NAc, and linking pain processing in the NAc to similar pain processing in the PAG (Li et al., 2001). To further prove this point, the researchers administered the CGRP antagonist CGRP8–37 into the NAc and observed an antagonization of the analgesic effect. These results were also demonstrated earlier in a 1989 study, in which CGRP injected into the NAc resulted in significant analgesia, which was then attenuated by administration of CGRP antagonist (Liu and Borgland, 2015). Overall, these results suggests a significant role of the CGRP1 receptor in the NAc specifically in terms of pain modulation, as the selectivity of the CGRP8–37 antagonist is very high, and primarily binds with the CGRP1 receptor subtype (Bell and McDermott, 1996).
Studies have revealed innervations from the NAc to the habenula, which has been shown to innervate the PAG. Later research demonstrated the nociception-related pathway between the NAc and the PAG specifically as well (Yu et al., 1988; Yu and Han, 1990; Sardi et al., 2018). Arguably, the administration of CGRP into the NAc activates and facilitates the descending pain pathway from the NAc through the PAG to mediate antinociceptive response.
γ-Aminobutyric acid
γ-Aminobutyric acid (GABA)ergic pathways are used in transmission between the NAc, ventral pallidum, ventral tegmental area, and rostromedial tegmentum. Research has effectively demonstrated the use of GABA secretion in descending pain modulation from specific brain structures involved in pain processing, such that it might inhibit the transmission of pain signals in the appropriate pathways (Kanematsu et al., 2006). Recently, research has shown that GABA receptors and dopamine receptors are both involved in large amounts of activity regulation in the NAc specifically (Hjelmstad, 2004; Chen et al., 2006). It has previously been suggested that exogenous administration of GABA could attenuate responses of “pain-excited neurons” in the NAc, leading to antinociception modulated by GABAA receptors (Xu et al., 2005).
Introduction of noxious stimulation to morphine-dependent rats and measurement of GABA influences on synaptic activity in the NAc revealed prolonged latency of electrical activity in the NAc after GABA injection (Xu et al., 2008). The researchers hypothesize that GABA regulates pain processing in cases of morphine dependency, with the activation of the GABAA receptor. They also report that administering GABA would cause GABAergic neurons to antagonize the effects of morphine, additionally weakening excitability of dopaminergic neurons. Finally, during the duration of a pain stimulus, dopamine concentration in the NAc gradually elevates while GABA concentration in that region gradually diminishes. Thus, the excitability of GABA neurons are gradually inhibited, reducing pain threshold.
Glutamate
Additionally, glutamatergic innervation of NAc GABAergic inhibitory neurons from the ventral tegmental area has been shown to drive aversive responses, in contrast to the typical excitatory glutamatergic input from other structures. Photoactivation by the NAc on mesoaccumbic glutamate fibers has been shown to promote this aversive response (Qi et al., 2016). One group of researchers has examined vesicular glutamate transporters (VGLUTs) levels in the NAc in terms of a reward model, in which rats self-administer sucrose, and a pain model, in which rats undergo spared nerve injury (Tukey et al., 2013). Previously, this sucrose administration has been shown to result in increased psychomotor activity, which indicates reward-seeking behavior in rodents, and the spared nerve injury model has been shown to lead to sensory allodynia and other pain behaviors (Wang et al., 2011). What they found was decreases in VGLUT1 and VGLUT3 during the pain model and an increase in VGLUT3 during sucrose administration. It could be argued from these results that co-release of glutamate with acetylcholine and other neurotransmitters decrease under painful stimulation and increase in the presence of reward (Tukey et al., 2013). The decrease in VGLUT1 in the pain model could arise from a change in connectivity due to motivational learning.
Substance P
In the spinal cord specifically, substance P is active in pain signaling and mediates the autonomic reflexes that activate in response to noxious stimuli (O’Connor et al., 2004). Substance P is also one of the neuropeptides that are believed to affect functioning of dopamine in the NAc. The first bit of evidence for this claim is the locations substance P can be found in. The largest densities of substance P are found in the nigrostriatal dopamine system, and slightly smaller densities are found in the ventral tegmental area and the NAc-specifically in the terminals of dopamine neurons (Al-Hasani et al., 2015). Researchers have discovered that injecting substance P into the ventral tegmental area, which projects dopamine to the NAc, will lead to dopamine increase (Xia et al., 2010). Additionally, a specific interaction between dopamine and substance P in the NAc has been identified. In their research, it was discovered that substance P can prolong the effects of dopamine activation (Koob and Volkow, 2010). However, there remains much to investigate in terms of what the role is of substance P in the NAc, and what it means for pain processing.
Why would the nucleus accumbens be involved in pain processing?
Evolution, biology, and behavior answer ‘why’ questions with resourcefulness, intelligence, and natural selection. Keeping warm and fed through winter, providing for offspring, and reducing injury all promote survival, and each of these behaviors is inherently rewarding because of this. One prime example of this phenomenon is the rewarding aspects of relief from pain or avoidance of pain.
It seems obvious at first why organisms must experience pain: it is an adaptive, survival-promoting phenomenon that alerts to the possibility of tissue damage, which leads to behaviors such as tending to the wound or removing the noxious stimulus, increasing probability of recovery and survival. Thus, the ‘why’ of the existence of pain could be conveniently answered by promoting the organism’s likelihood to engage in survival behavior. However, this definition only applies to acute pain experienced in the face of actual danger. There are numerous instances in which pain lasts well beyond the treatment of a disease or the healing of an injury, and there are numerous other instances in which pain arises seemingly without a physiological cause.
Clearly, pain is a highly subjective individual experience with dimensions that include sensation, affect, and cognition. While the sensory aspect of pain such as stimulus source, location, and intensity is associated with nociceptor activation, the overall experience of the nociception can be altered by context and motivation. In fact, the activation of nociceptors does not always result in pain, and the sensation of pain can be experienced without nociceptive stimulus (Fields and Margolis, 2015). Thus, the affective, aversive aspect of pain is vital for promoting survival behaviors, such as withdrawal from the noxious stimulus. Additionally, it is critical that the aversion that results from pain experience is rewarding, so that it affects memory and promotes learning actions to avoid and safety behaviors to engage in in order to prevent future harm (Fields and Margolis, 2015). Therefore, neural circuitry involved in reward, motivation, and learning, such as in the NAc are essential aspects of pain processing.
An important point to note is that pharmacologically, the pain relief depends heavily on the reward system. For instance, opioids as analgesics do not work simply by blocking the transmission of pain messages; they mimic the effects of endogenous endorphins that get released in response to a complex situation. For instance, these endorphins can be released to help actively suppress pain when there is some painful stimulus present that it would be rewarding to ignore, such as when ignoring a broken ankle to outrun a bear. In this way, pain and pleasure must be analyzed within one circuit or structure, so that competing motivations even in the presence of pain can be weighed and assigned value. The NAc is a motivational behavior region of the brain, and it is one of the sources of conflicting motivations that are experienced during pain. Without its involvement in both pain and reward/motivation, humans and animals would not be able to make these decisions in complex situations.
Therapeutic Targeting: Deep Brain Stimulation in the Nucleus Accumbens for Analgesia
Recently, research into deep brain stimulation as a treatment for intractable pain has increased. This process has been used to successfully reduce chronic and treatment-resistant pain in humans, and acute pain in animal models, although the regions typically targeted for pain have included the thalamus, hypothalamus, motor cortex, anterior cingulate cortex, and the ventral tegmental area (Levy et al., 2010; Li et al., 2016). Because of the involvement of the NAc in pain processing, it is reasonable to suggest that deep brain stimulation of this region will also alleviate pain, particularly since the areas previously targeted in analgesic deep brain stimulation are so highly connected to the NAc, which forwards inhibitory projections to the medial thalamus, which projects forward to dorsal horn neurons, effectively closing the gate and producing analgesia. In fact, there have been a couple studies that have demonstrated effective alleviation of pain via deep brain stimulation of the NAc in humans (Levy et al., 2010; Mallory et al., 2012).
Conclusion
The NAc, often overlooked as nothing more than a pleasure center, has widespread cortical connections that imply its involvement in numerous areas of functioning. This region is particularly overlooked in pain research, where the sensation component of the pain neuromatrix is often the primary focus of study, leading to the exclusion of the affective and cognitive components to which the NAc is so highly linked. The NAc is in a prime location for pain signaling, as it receives innervation from both the prefrontal cortex and limbic regions (such as the hippocampus and amygdala), and projects forward to motor regions (Mallory et al., 2012). In this way, the NAc plays a direct role in the modulation of pain processing via cortical circuitry and chemical neurotransmission.
Footnotes
Conflicts of interest: Portions of this review are part of Haley N. Harris’ PhD dissertation. Dr. Yuan-Bo Peng is Haley N. Harris’ s PhD mentor.
Financial support: None.
Copyright license agreement: The Copyright License Agreement has been signed by both authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Ghulam Hussain, Government College University Faisalabad, Pakistan.
P-Reviewer: Hussain G; C-Editors: Zhao M, Yu J; T-Editor: Jia Y
References
- 1.Al-Hasani R, McCall JG, Shin G, Gomez AM, Schmitz GP, Bernardi JM, Pyo CO, Park SI, Marcinkiewcz CM, Crowley NA, Krashes MJ, Lowell BB, Kash TL, Rogers JA, Bruchas MR. Distinct subpopulations of nucleus accumbens dynorphin neurons drive aversion and reward. Neuron. 2015;87:1063–1077. doi: 10.1016/j.neuron.2015.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bálint E, Mezey S, Csillag A. Efferent connections of nucleus accumbens subdivisions of the domestic chicken (Gallus domesticus): an anterograde pathway tracing study. J Comp Neurol. 2011;519:2922–2953. doi: 10.1002/cne.22672. [DOI] [PubMed] [Google Scholar]
- 3.Becerra L, Borsook D. Signal valence in the nucleus accumbens to pain onset and offset. Eur J Pain. 2008;12:866–869. doi: 10.1016/j.ejpain.2007.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Becerra L, Breiter HC, Wise R, Gonzalez RG, Borsook D. Reward circuitry activation by noxious thermal stimuli. Neuron. 2001;32:927–946. doi: 10.1016/s0896-6273(01)00533-5. [DOI] [PubMed] [Google Scholar]
- 5.Bell D, McDermott B. Calcitonin gene-related peptide in the cardiovascular system: Characterization of receptor populations and their (patho) physiological significance. Pharmacol Rev. 1996;48:253–288. [PubMed] [Google Scholar]
- 6.Bianco IH, Wilson SW. The habenular nuclei: a conserved asymmetric relay station in the vertebrate brain. Philos Trans R Soc B Biol Sci. 2009;364:1005–1020. doi: 10.1098/rstb.2008.0213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bodnar RJ. Endogenous Opiates and Behavior: 2016. Peptides. 2018;101:167–212. doi: 10.1016/j.peptides.2018.01.011. [DOI] [PubMed] [Google Scholar]
- 8.Borsook D, Linnman C, Faria V, Strassman AM, Becerra L, Elman I. Reward deficiency and anti-reward in pain chronification. Neurosci Biobehav Rev. 2016;68:282–297. doi: 10.1016/j.neubiorev.2016.05.033. [DOI] [PubMed] [Google Scholar]
- 9.Bree D, Levy D. Development of CGRP-dependent pain and headache related behaviours in a rat model of concussion: Implications for mechanisms of post-traumatic headache. Cephalalgia. 2018;38:246–258. doi: 10.1177/0333102416681571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Breiter HC, Rosen BR. Functional magnetic resonance imaging of brain reward circuitry in the human. Ann N Y Acad Sci. 1999;877:523–547. doi: 10.1111/j.1749-6632.1999.tb09287.x. [DOI] [PubMed] [Google Scholar]
- 11.Campion KN, Saville KA, Morgan MM. Relative contribution of the dorsal raphe nucleus and ventrolateral periaqueductal gray to morphine antinociception and tolerance in the rat. Eur J Neurosci. 2016;44:2667–2672. doi: 10.1111/ejn.13378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chang PC, Pollema-Mays SL, Centeno MV, Procissi D, Contini M, Baria AT, Martina M, Apkarian AV. Role of nucleus accumbens in neuropathic pain: linked multi-scale evidence in the rat transitioning to neuropathic pain. Pain. 2014;155:1128–1139. doi: 10.1016/j.pain.2014.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen G, Kittler J, Moss S, Yan Z. Dopamine D3 receptors regulate GABAa receptor function through a phospho-dependent endocytosis mechanism in nucleus accumbens. J Neurosci. 2006;26:2513–2521. doi: 10.1523/JNEUROSCI.4712-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clarke PBS, Franklin KBJ. Infusions of 6-hydroxydopamine into the nucleus accumbens abolish the analgesic effect of amphetamine but not of morphine in the formalin test. Brain Res. 1992;580:106–110. doi: 10.1016/0006-8993(92)90932-y. [DOI] [PubMed] [Google Scholar]
- 15.Dias EV, Sartori CR, Marião PR, Vieira AS, Camargo LC, Athie MC, Pagliusi MO, Tambeli CH, Parada CA. Nucleus accumbens dopaminergic neurotransmission switches its modulatory action in chronification of inflammatory hyperalgesia. Eur J Neurosci. 2015;42:2380–2389. doi: 10.1111/ejn.13015. [DOI] [PubMed] [Google Scholar]
- 16.Dill RE, Costa E. Behavioural dissociation of the enkephalinergic systems of nucleus accumbens and nucleus caudatus. Neuropharmacology. 1977;16:323–326. doi: 10.1016/0028-3908(77)90068-5. [DOI] [PubMed] [Google Scholar]
- 17.Erfanparast A, Tamaddonfard E, Seyedin S. Involvement of central opiate receptors in modulation of centrally administered oxytocin-induced antinociception. Iran J Basic Med Sci. 2018;21:1275–1280. doi: 10.22038/ijbms.2018.26302.6449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Escobar AP, Cornejo FA, Olivares-Costa M, González M, Fuentealba JA, Gysling K, España RA, Andrés ME. Reduced dopamine and glutamate neurotransmission in the nucleus accumbens of quinpirole-sensitized rats hints at inhibitory D2 autoreceptor function. J Neurochem. 2015;134:1081–1090. doi: 10.1111/jnc.13209. [DOI] [PubMed] [Google Scholar]
- 19.Fields HL, Margolis EB. Understanding opioid reward. Trends Neurosci. 2015;38:217–225. doi: 10.1016/j.tins.2015.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Floresco SB. The nucleus accumbens: an interface between cognition, emotion, and action. Annu Rev Psychol. 2015;66:25–52. doi: 10.1146/annurev-psych-010213-115159. [DOI] [PubMed] [Google Scholar]
- 21.Gear RW, Levine JD. Nucleus accumbens facilitates nociception. Exp Neurol. 2011;229:502–506. doi: 10.1016/j.expneurol.2011.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gentry RN, Schuweiler DR, Roesch MR. Dopamine signals related to appetitive and aversive events in paradigms that manipulate reward and avoidability. Brain Res. 2019;1713:80–90. doi: 10.1016/j.brainres.2018.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gupta I, Young AMJ. Metabotropic glutamate receptor modulation of dopamine release in the nucleus accumbens shell is unaffected by phencyclidine pretreatment: In vitro assessment using fast-scan cyclic voltammetry rat brain slices. Brain Res. 2018;1687:155–161. doi: 10.1016/j.brainres.2018.03.007. [DOI] [PubMed] [Google Scholar]
- 24.Hikida T, Kimura K, Wada N, Funabiki K, Nakanishi Shigetada S. Distinct roles of synaptic transmission in direct and indirect striatal pathways to reward and aversive behavior. Neuron. 2010;66:896–907. doi: 10.1016/j.neuron.2010.05.011. [DOI] [PubMed] [Google Scholar]
- 25.Hikida T, Morita M, Macpherson T. Neural mechanisms of the nucleus accumbens circuit in reward and aversive learning. Neurosci Res. 2016;108:1–5. doi: 10.1016/j.neures.2016.01.004. [DOI] [PubMed] [Google Scholar]
- 26.Hjelmstad G. Dopamine excites nucleus accumbens neurons through the differential modulation of glutamate and GABA release. J Neurosci. 2004;24:8621–8628. doi: 10.1523/JNEUROSCI.3280-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hong SI, Nguyen TL, Ma SX, Kim HC, Lee SY, Jang CG. TRPV1 modulates morphine-induced conditioned place preference via p38 MAPK in the nucleus accumbens. Behav Brain Res. 2017;334:26–33. doi: 10.1016/j.bbr.2017.07.017. [DOI] [PubMed] [Google Scholar]
- 28.Hu H. Reward and aversion. Annu Rev Neurosci. 2016;39:297–324. doi: 10.1146/annurev-neuro-070815-014106. [DOI] [PubMed] [Google Scholar]
- 29.Joseph JE, Zhu X, Lynam D, Kelly TH. Modulation of meso-limbic reward processing by motivational tendencies in young adolescents and adults. Neuroimage. 2016;129:40–54. doi: 10.1016/j.neuroimage.2015.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kanematsu T, Yasunaga A, Mizoguchi Y, Kuratani A, Kittler J, Jovanovic J. Modulation of GABAa receptor phosphorylation and membrane trafficking by phospholipase C-related inactive protein/protein phosphatase 1 and 2A signaling complex underlying brain-derived neurotrophic factor-dependent regulation of GABAergic inhibition. J Biol Chem. 2006;281:22180–22189. doi: 10.1074/jbc.M603118200. [DOI] [PubMed] [Google Scholar]
- 31.Kato T, Ide S, Minami M. Pain relief induces dopamine release in the rat nucleus accumbens during the early but not late phase of neuropathic pain. Neurosci Lett. 2016;629:73–78. doi: 10.1016/j.neulet.2016.06.060. [DOI] [PubMed] [Google Scholar]
- 32.Khalilzadeh E, Saiah G. The possible mechanisms of analgesia produced by microinjection of morphine into the lateral habenula in the acute model of trigeminal pain in rats. Res Pharm Sci. 2017;12:241. doi: 10.4103/1735-5362.207205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kohnomi S, Ebihara K, Kobayashi M. Suppressive regulation of lateral inhibition between medium spiny neurons via dopamine D1 receptors in the rat nucleus accumbens shell. Neurosci Lett. 2017;636:58–63. doi: 10.1016/j.neulet.2016.10.049. [DOI] [PubMed] [Google Scholar]
- 34.Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology. 2010;35:217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lammel S, Ion DI, Roeper J, Malenka RC. Projection-specific modulation of dopamine neuron synapses by aversive and rewarding stimuli. Neuron. 2011;70:855–862. doi: 10.1016/j.neuron.2011.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Levy R, Deer TR, Henderson J. Intracranial neurostimulation for pain control: a review. Pain Physician. 2010;13:157–165. [PubMed] [Google Scholar]
- 37.Li A, Sibi J, Yang X, Chiao JC, Peng YB. Stimulation of the ventral tegmental area increased nociceptive thresholds and decreased spinal dorsal horn neuronal activity in rat. Exp Brain Res. 2016;234:1505–1514. doi: 10.1007/s00221-016-4558-z. [DOI] [PubMed] [Google Scholar]
- 38.Li N, Lundeberg T, Yu LC. Involvement of CGRP and CGRP1 receptor in nociception in the nucleus accumbens of rats. Brain Res. 2001;901:161–166. doi: 10.1016/s0006-8993(01)02341-1. [DOI] [PubMed] [Google Scholar]
- 39.Li Z, Chen Z, Fan G, Li A, Yuan J, Xu T. Cell-type-specific afferent innervation of the nucleus accumbens core and shell. Front Neuroanat. 2018;12:1–16. doi: 10.3389/fnana.2018.00084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liu S, Borgland SL. Regulation of the mesolimbic dopamine circuit by feeding peptides. Neuroscience. 2015;289:19–42. doi: 10.1016/j.neuroscience.2014.12.046. [DOI] [PubMed] [Google Scholar]
- 41.Llorca-Torralba M, Borges G, Neto F, Mico JA, Berrocoso E. Noradrenergic locus coeruleus pathways in pain modulation. Neuroscience. 2016;338:93–113. doi: 10.1016/j.neuroscience.2016.05.057. [DOI] [PubMed] [Google Scholar]
- 42.Ma QP, Shi YS, Han JS. Further studies on interactions between periaqueductal gray, nucleus accumbens and habenula in antinociception. Brain Res. 1992;583:292–295. doi: 10.1016/s0006-8993(10)80036-8. [DOI] [PubMed] [Google Scholar]
- 43.Magnusson JE, Martin RV. Additional evidence for the involvement of the basal ganglia in formalin-induced nociception: The role of the nucleus accumbens. Brain Res. 2002;942:128–132. doi: 10.1016/s0006-8993(02)02489-7. [DOI] [PubMed] [Google Scholar]
- 44.Mallory GW, Abulseoud O, Hwang SC, Gorman DA, Stead SM, Klassen BT, Sandroni P, Watson JC, Lee KH. The nucleus accumbens as a potential target for central poststroke pain. Mayo Clin Proc. 2012;87:1025–1031. doi: 10.1016/j.mayocp.2012.02.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Massaly N, Copits BA, Wilson-Poe AR, Hipólito L, Markovic T, Yoon HJ, Liu S, Walicki MC, Bhatti DL, Sirohi S, Klaas A, Walker BM, Neve R, Cahill CM, Shoghi KI, Gereau RW, 4th, McCall JG, Al-Hasani R, Bruchas MR, Morón JA. Pain-induced negative affect is mediated via recruitment of the nucleus accumbens kappa opioid system. Neuron. 2019;102:564–573e6. doi: 10.1016/j.neuron.2019.02.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mizoguchi H, Takagi H, Watanabe C, Yonezawa A, Sato T, Sakurada T, Sakurada S. Involvement of multiple μ-opioid receptor subtypes on the presynaptic or postsynaptic inhibition of spinal pain transmission. Peptides. 2014;51:15–25. doi: 10.1016/j.peptides.2013.10.012. [DOI] [PubMed] [Google Scholar]
- 47.Moradi M, Yazdanian M, Haghparast A. Role of dopamine D2-like receptors within the ventral tegmental area and nucleus accumbens in antinociception induced by lateral hypothalamus stimulation. Behav Brain Res. 2015;292:508–514. doi: 10.1016/j.bbr.2015.07.007. [DOI] [PubMed] [Google Scholar]
- 48.Navratilova E, Xie JY, Okun A, Qu C, Eyde N, Ci S, Ossipov MH, King T, Fields HL, Porreca F. Pain relief produces negative reinforcement through activation of mesolimbic reward-valuation circuitry. Proc Natl Acad Sci U S A. 2012;109:20709–20713. doi: 10.1073/pnas.1214605109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.O’Connor TM, O’Connell J, O’Brien DI, Goode T, Bredin CP, Shanahan F. The role of substance P in inflammatory disease. J Cell Physiol. 2004;201:167–180. doi: 10.1002/jcp.20061. [DOI] [PubMed] [Google Scholar]
- 50.Okun A, McKinzie DL, Witkin JM, Remeniuk B, Husein O, Gleason SD, Oyarzo J, Navratilova E, McElroy B, Cowen S, Kennedy JD, Porreca F. Hedonic and motivational responses to food reward are unchanged in rats with neuropathic pain. Pain. 2016;157:2731–2738. doi: 10.1097/j.pain.0000000000000695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Oswald LM, Wand GS, Wong DF, Brown CH, Kuwabara H, Brašić JR. Risky decision-making and ventral striatal dopamine responses to amphetamine: a positron emission tomography [(11)C]raclopride study in healthy adults. Neuroimage. 2015;113:26–36. doi: 10.1016/j.neuroimage.2015.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Qi J, Zhang S, Wang HL, Barker DJ, Miranda-Barrientos J, Morales M. VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons. Nat Neurosci. 2016;19:725–733. doi: 10.1038/nn.4281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rauschecker JP, May ES, Maudoux A, Ploner M. Frontostriatal gating of tinnitus and chronic pain. Trends Cogn Sci. 2015;19:567–578. doi: 10.1016/j.tics.2015.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Reakkamnuan C, Cheaha D, Kumarnsit E. Nucleus accumbens local field potential power spectrums, phase-amplitude couplings and coherences following morphine treatment. Acta Neurobiol Exp (Wars) 2017;77:214–224. [PubMed] [Google Scholar]
- 55.Ren W, Centeno MV, Berger S, Wu Y, Na X, Liu X, Kondapalli J, Apkarian AV, Martina M, Surmeier DJ. The indirect pathway of the nucleus accumbens shell amplifies neuropathic pain. Nat Neurosci. 2015;19:220–222. doi: 10.1038/nn.4199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Roeder Z, Chen Q, Davis S, Carlson JD, Tupone D, Heinricher MM. Parabrachial complex links pain transmission to descending pain modulation. Pain. 2016;157:2697–2708. doi: 10.1097/j.pain.0000000000000688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Rosas M, Porru S, Fenu S, Ruiu S, Peana AT, Papale A, Brambilla R, Di Chiara G, Acquas E. Role of nucleus accumbens μ opioid receptors in the effects of morphine on ERK1/2 phosphorylation. Psychopharmacology (Berl) 2016;233:2943–2954. doi: 10.1007/s00213-016-4340-8. [DOI] [PubMed] [Google Scholar]
- 58.Rougé-Pont F, Piazza PV, Kharouby M, Le Moal M, Simon H. Higher and longer stress-induced increase in dopamine concentrations in the nucleus accumbens of animals predisposed to amphetamine self-administration. A microdialysis study. Brain Res. 1993;602:169–174. doi: 10.1016/0006-8993(93)90260-t. [DOI] [PubMed] [Google Scholar]
- 59.Saigusa T, Aono Y, Waddington JL. Mechanisms underlying δ- and μ-opioid receptor agonist-induced increases in extracellular dopamine level in the nucleus accumbens of freely moving rats. J Oral Sci. 2017;59:195–200. doi: 10.2334/josnusd.16-0874. [DOI] [PubMed] [Google Scholar]
- 60.Salgado S, Kaplitt MG. The nucleus accumbens: a comprehensive review. Stereotact Funct Neurosurg. 2015;93:75–93. doi: 10.1159/000368279. [DOI] [PubMed] [Google Scholar]
- 61.Sardi NF, Lazzarim MK, Guilhen VA, Marcílio RS, Natume PS, Watanabe TC, Lima MMS, Tobaldini G, Fischer L. Chronic sleep restriction increases pain sensitivity over time in a periaqueductal gray and nucleus accumbens dependent manner. Neuropharmacology. 2018;139:52–60. doi: 10.1016/j.neuropharm.2018.06.022. [DOI] [PubMed] [Google Scholar]
- 62.Satoh M, Kubota A, Iwama T, Wada T, Yasui M, Fujibayashi K, Takagi H. Comparison of analgesic potencies of mu, delta and kappa agonists locally applied to various CNS regions relevant to analgesia in rats. Life Sci. 1983;37:689–692. doi: 10.1016/0024-3205(83)90596-9. [DOI] [PubMed] [Google Scholar]
- 63.Saunders BT, Robinson TE. The role of dopamine in the accumbens core in the expression of Pavlovian-conditioned responses. Eur J Neurosci. 2012;36:2521–2532. doi: 10.1111/j.1460-9568.2012.08217.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schepisi C, Cianci S, Bedse G, Fu J, Gaetani S, Nencini P. Differences in the structure of drinking, cart expression and dopamine turnover between polydipsic and non polydipsic rats in the quinpirole model of psychotic polydipsia. Psychopharmacology (Berl) 2014;231:3889–3897. doi: 10.1007/s00213-014-3527-0. [DOI] [PubMed] [Google Scholar]
- 65.Schmidt BL, Tambeli CH, Barletta J, Luo L, Green P, Levine JD, Gear RW. Altered nucleus accumbens circuitry mediates pain-induced antinociception in morphine-tolerant rats. J Neurosci. 2002;22:6773–6780. doi: 10.1523/JNEUROSCI.22-15-06773.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Schwabe K, Polikashvili N, Krauss JK. Deficient sensorimotor gating induced by selective breeding in rats is improved by entopeduncular nucleus lesions. Neurobiol Dis. 2009;34:351–356. doi: 10.1016/j.nbd.2009.02.004. [DOI] [PubMed] [Google Scholar]
- 67.Segal M, Sandberg D. Analgesia produced by electrical stimulation of catecholamine nuclei in the rat brain. Brain Res. 1977;123:369–372. doi: 10.1016/0006-8993(77)90488-7. [DOI] [PubMed] [Google Scholar]
- 68.Seminowicz DA, Remeniuk B, Krimmel SR, Smith MT, Barrett FS, Wulff AB, Furman AJ, Geuter S, Lindquist MA, Irwin MR, Finan PH. Pain-related nucleus accumbens function: modulation by reward and sleep disruption. Pain. 2019;160:1196–1207. doi: 10.1097/j.pain.0000000000001498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Shafiei I, Vatankhah M, Zarepour L, Ezzatpanah S, Haghparast A. Role of D1- and D2-like dopaminergic receptors in the nucleus accumbens in modulation of formalin-induced orofacial pain: Involvement of lateral hypothalamus. Physiol Behav. 2018;188:25–31. doi: 10.1016/j.physbeh.2018.01.020. [DOI] [PubMed] [Google Scholar]
- 70.Sheibani V, Shamsizadeh A, Afarinesh MR, Rezvani ME. Neonatal capsaicin treatment modulates experience-Dependent plasticity in the rat barrel cortex. J Comp Neurol. 2010;518:3427–3438. doi: 10.1002/cne.22384. [DOI] [PubMed] [Google Scholar]
- 71.Shur M, Ritvo L. A principle of evolutionary biology for psychoanalysis: Schneirla’s evolutionary and developmental theory of biphasic processes underlying approach and withdrawal and Freud’s unpleasure and pleasure principles. J Am Psychoanal Assoc. 1970;18:422–439. doi: 10.1177/000306517001800210. [DOI] [PubMed] [Google Scholar]
- 72.Taylor BK, Joshi C, Uppal H. Stimulation of dopamine D2 receptors in the nucleus accumbens inhibits inflammatory pain. Brain Res. 2003;987:135–143. doi: 10.1016/s0006-8993(03)03318-3. [DOI] [PubMed] [Google Scholar]
- 73.Tukey DS, Lee M, Xu D, Eberle SE, Goffer Y, Manders TR, Ziff EB, Wang J. Differential effects of natural rewards and pain on vesicular glutamate transporter expression in the nucleus accumbens. Mol Brain. 2013;6:32. doi: 10.1186/1756-6606-6-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang J, Li D, Li X, Liu FY, Xing GG, Cai J, Wan Y. Phase-amplitude coupling between theta and gamma oscillations during nociception in rat electroencephalography. Neurosci Lett. 2011;499:84–87. doi: 10.1016/j.neulet.2011.05.037. [DOI] [PubMed] [Google Scholar]
- 75.Xia YF, Margolis EB, Hjelmstad GO. Substance P inhibits GABAB receptor signalling in the ventral tegmental area. J Physiol. 2010;588:1541–1549. doi: 10.1113/jphysiol.2010.188367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Xu Y, Xu M, Yan B. Effects of GABA on the electric activities of pain-related neurons in nucleus accumbens of rats. Neurosci Bull. 2005;21:53–57. [Google Scholar]
- 77.Xu Y, Xu MY, Li X. Modulation of gamma-aminobutyric acid on painful sense in central nervous system of morphine-dependent rats? Neurosci Bull. 2008;24:278–282. doi: 10.1007/s12264-008-0227-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Yan H, Yu LC. Influences of calcitonin gene-related peptide on mu opioid receptors in nucleus accumbens neurons of rats. Neuropeptides. 2013;47:125–131. doi: 10.1016/j.npep.2012.10.008. [DOI] [PubMed] [Google Scholar]
- 79.Yoshida K, Nonaka T, Nakamura S, Araki M, Yamamoto T. Microinjection of 26RFa, an endogenous ligand for the glutamine RF-amide peptide receptor (QRFP receptor), into the rostral ventromedial medulla (RVM), locus coelureus (LC), and periaqueductal grey (PAG) produces an analgesic effect in rats. Peptides. 2019;115:1–7. doi: 10.1016/j.peptides.2019.02.003. [DOI] [PubMed] [Google Scholar]
- 80.Yu L, Shi Y, Han J. A descending pathway of analgesia from nucleus accumbens to habenula. Chin Sci Bull. 1988;33:337–341. [Google Scholar]
- 81.Yu LC, Han JS. Habenula as a relay in the descending pathway from nucleus accumbens to periaqueductal gray subserving antinociception. Int J Neurosci. 1990;54:245–251. doi: 10.3109/00207459008986640. [DOI] [PubMed] [Google Scholar]
- 82.Yu LC, Hou JF, Fu FH, Zhang YX. Roles of calcitonin gene-related peptide and its receptors in pain-related behavioral responses in the central nervous system. Neurosci Biobehav Rev. 2009;33:1185–1191. doi: 10.1016/j.neubiorev.2009.03.009. [DOI] [PubMed] [Google Scholar]
- 83.Zeng Z, Liang Y, Chen Y, Jing X, Peng S, Tao E. Chronic back pain cured by low-dose levodopa: Is it a variant of restless legs syndrome? J Pain Res. 2018;11:277–279. doi: 10.2147/JPR.S156894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhu J, Bardo MT, Dwoskin LP. Distinct effects of enriched environment on dopamine clearance in nucleus accumbens shell and core following systemic nicotine administration. Synapse. 2013;67:57–67. doi: 10.1002/syn.21615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zubieta JK, Stohler CS. Neurobiological mechanisms of placebo responses. Ann N Y Acad Sci. 2009;1156:198–210. doi: 10.1111/j.1749-6632.2009.04424.x. [DOI] [PMC free article] [PubMed] [Google Scholar]