

The physiological effect of neurotrophic factors and their role in Alzheimer’s disease (AD): Neurotrophins (NTs) are a family of homologues proteins that play an essential role in neuronal cells growth, survival and differentiation. These proteins include the nerve growth factor (NGF), the brain derived neurotrophic factor (BDNF), NT-3 and NT-3, also known as NT-4/5.
The biological functions of NTs are regulated by binding to two distinct classes of receptors, namely p75 neurotrophin receptor and tropomyosin receptor kinase (Trk). p75 neurotrophin receptor is member of the tumour necrosis factor receptor family, which binds with no selectivity all of the neurotrophins. On the other hand, to each NT there is a specific Trk receptor: NGF binds to TrkA, BDNF and NT-4 to TrkB, NT-3 interacts mainly with TrkC, respectively (Bothwell, 2014).
NGF is the first discovered and best-characterized member of NTs family. Similarly to all the other neurotrophic factors, NGF is synthesized in a precursor form (pro-NGF) and is cleaved by proteases (e.g., plasmin, furin, matrix metalloproteinase (MMP7 and MMP3) to produce the mature and active form, which typically exerts its biological action as non-covalent homodimer.
The cholinergic system of the human brain is considered as the master regulator of mnemonic functions and its loss is strictly associated with the cognitive decline (Cuello et al., 2019). NGF is secreted by post-synaptic target tissues and it is known to activate, by the binding to TrkA receptor, the signaling pathways at the axonal growth cones. Retrograde NGF transport maintains the phenotype of basal forebrain cholinergic neurons and control the long distance communication between the axon terminals and the neuronal cell bodies.
Specifically, NGF binding to TrkA activates the three following signaling pathways: i) the phosphatidylinositol 3-kinase/protein kinase B (Akt); ii) the Ras/mitogen-activated protein kinase (ERK) and iii) the phospholipase C. In particular, the ERK or Akt pathways trigger the phosphorylation of transcription factor cAMP response element binding protein (CREB). This, in turn, induces the expression of proteins, including BDNF, which are directly involved in memory formation and consolidation.
Noteworthy, NGF metabolism is hampered in AD, thus determining a widespread loss of cholinergic neurons in the basal forebrain with consequent loss in cognition and memory. NGF signalling pathways control also the post-translational modifications of the amyloid precursor protein (APP) and the amyloid beta (Aβ) production in neurons. Indeed, the deprivation in NGF determines Aβ aggregation and tau hyperphosphorylation (Canu et al., 2017). BDNF can reduce the production of toxic Aβ oligomers through a mechanism that increases α-secretase processing of APP. It has been often reported that different NTs perform similar activities in physiological as well as pathological conditions. However, the levels of NGF and BDNF follow different trend in hippocampus of AD animal models: at early stages of pathology, Aβ oligomers lead to significant reductions in BDNF expression whereas, at later stages, deficits in the NGF metabolic pathway occur.
Peptides as NT mimicking systems: Taking into account the essential activity in neurite outgrowth and maintenance, neurotrophins represent a significant drug target in neurodegenerative diseases, memory consolidation and neurogenesis. The most relevant challenges in the use of NTs in therapy include the reduction of uncontrolled side effects as well as the increase of their bioavailability and delivery to the central nervous system, since NTs, due to their molecular size, barely cross the blood-brain barrier (BBB). Small molecules able to mimic NTs activity may overcome such limitations. Experimental and computational evidences pointed to the critical role of NTs N-terminus domain in the interaction with their respective Trk receptors. To this aim, we synthesized peptides encompassing first 14 and 12 N-terminal residues of NGF and BDNF, respectively NGF(1–14) and BDNF(1–12), as new mimic systems of the corresponding whole protein (Travaglia et al., 2012). These peptides induced cell proliferation as much as the respective proteins. Noteworthy, copper addition increased NGF and NGF(1–14) peptide activity, whereas decreased that of BDNF and BDNF(1–12) peptide. These data suggested that metal ions could modulate the activity of NTs and drive their different specialization in neurons. Hence, the metal dyshomeostasis in pathological condition may have a diverse impact on NT expression and activity.
Copper ions and AD: According to the “metal hypothesis” of sporadic AD (Barnham and Bush, 2014), the dyshomeostasis of metal ions is an early event in the onset and in the progression of the pathology. Such a hypothesis interlaces with the other two main theories on AD pathogenesis, namely the amyloid cascade and the oxidative stress.
A local imbalance of metal ions between extra- and intra-cellular space may favour their binding to Aβ, thus determining amyloid aggregation, which may result in formation of oligomeric precursors, amyloid fibres or amorphous aggregates, depending on the metal ion/peptide molar ratio. Furthermore, the increase of intracellular copper modulates APP metabolism and then Aβ release. The direct binding of the redox-active metals as Cu2+ may also generate reactive oxygen species and then oxidative stress.
The accumulation of metal ions in the brain can also be related to BBB damages. Neuroimaging studies in individuals with mild cognitive impairment and early AD have shown BBB breakdown in the hippocampus. Accordingly, a significant elevation in the copper to iron molar ratio is reported in serum of mild cognitive impairment subjects (Mueller et al., 2012). Such an increase appears to be transient in nature, as patients with early AD were nearly identical to healthy controls. Actually, the enhancement of exchangeable copper levels correlates with tissue oxidative damage (James et al., 2012). These copper pools, referred as metal not bound to the ceruloplasmin, are able to pass BBB and result increased in individuals suffering of AD (Squitti, 2014). Such evidence suggests that copper handling is displaced in the brain of AD patients although no difference may be observed in the total metal content compared to that of healthy individuals.
Copper signaling and NGF: Copper is an indispensable metal as metabolic cofactors in many enzymes but mounting experimental evidences point to its role as messenger to modulate the function of neurotransmitter receptors, affecting neurite outgrowth, synaptic plasticity and memory formation.
Such a messenger activity requires the presence of kinetically accessible pools of copper ions and the involvement of molecules able to bind, transport and release metal to a specific intracellular target.
Since the main brain areas of copper release correspond to same regions where NGF performs its activity, we hypothesized that the molecules involved in the functional copper signaling process could be the NTs (Figure 1). Actually, in vitro studies in PC12 cells indicated that only copper and zinc, among bivalent metal ions, are the most effective at altering the NGF conformation and then potentially affecting its biological activity (Maitra et al., 2000). To note, copper binds to NGF with higher affinity than zinc.
Figure 1.
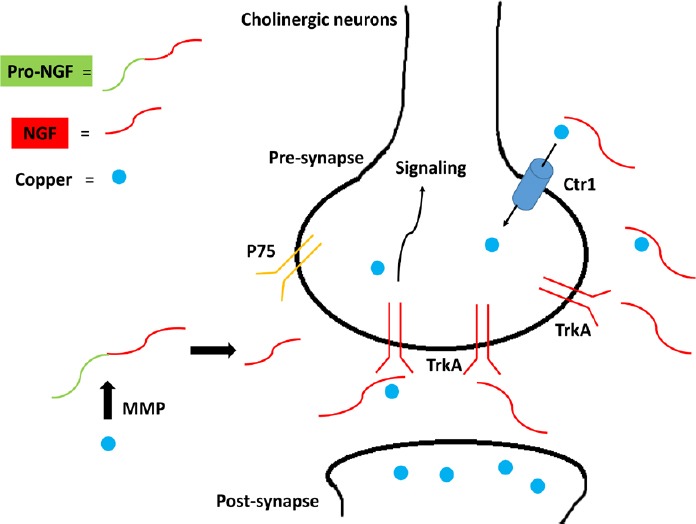
A graphic model showing how copper ions may affect NGF signaling.
Copper released in the synaptic space binds to nerve growth factor (NGF) so to influence interaction with tropomyosin receptor kinase A (TrkA) and p75 receptors. Copper may influence metalloprotease (MMP) activity and then NGF release. NGF may influence copper transport inside cells (copper transporter protein 1 (Ctr1)) and then control metal homeostasis.
The initial experimental findings on the interaction between NTs and metals suggested a potential toxic role of copper, analogous to that of heavy metals such as lead and mercury. Later, it has been reported that NGF increases cellular copper content and the metal ion deprivation by means of copper chelators decreases the neurite outgrowth induced by NGF (Birkaya and Aletta, 2005).
As the neurite outgrowth is a macro marker of neuronal differentiation, we determined that copper ion addition induced a two-fold increase in the NGF-driven expression of specific genes involved in neuronal development and differentiation, including Map2, Tubb3, Elavl4 and GAP- 43. These findings confirmed the influence of copper in the regulation role of NGF (Pandini et al., 2016).
To give insight on the molecular mechanism involved in the presence of copper, we synthesized other peptides mimicking the NGF and encompassing the first 14 residues, including the dimer d-NGF(1–15), a peptide with reverse sequence and another with a scrambled sequence (Naletova et al., 2019).
The specific interaction of NGF with its receptor is influenced by the presence of copper ions that lead to the decrease of TrKA/p75 internalization ratio, indicating that metal can modulate different signalling pathways of protein and then biological response. Instead the mimicking peptides NGF(1–14) and dNGF(1–15) induced exclusively TrkA phosphorylation, an effect strongly influenced by copper ions. It is to note that the medium in which cells grow contains metal ions. The addition of a copper extracellular chelator, as bathocuproine disulfonate (BCS), partially inhibited Tyr-490 phosphorylation of TrkA induced by both NGF and mimicking peptides, whereas a supplement of exogenous copper ions partially restored it.
Differently, the reverse and scrambled peptides both in absence or presence of metal ions have no activity, confirming that the N-terminus sequence of protein is specifically involved in receptor interaction and metal binding specifically increase recognition process.
The activation of TrkA by NGF or mimicking peptides triggered the signalling cascade of Akt and ERK. All these pathways converge to CREB phosphorylation (p-CREB) that is strongly enhanced by the presence of copper ions. Accordingly, BCS addition during the treatment significantly decreased the amount of p-CREB underlining a positive role of metal in regulating gene expression. Since the addition of BCS alone has no effect compared to the untreated controls, the copper chelation effect is strictly related to the contemporary presence of NGF or mimicking peptides.
According to the p-CREB level, NGF and mimicking peptides increased the transcription of BDNF gene, but differently from that observed for signaling activation, copper presence decreased the release of BDNF. These data confirm the hypothesis that copper ions may modulate differently NGF and BDNF expression and activity.
The copper binding with NGF and mimicking peptides enforces their cellular recognition and the incubation with an extracellular copper chelator may hinder this process. On the other hand, the observed effect of NGF and mimicking peptides on gene expression requires pathways that go beyond cellular recognition.
It is known that ionophore ligands increase intracellular metal bioavailability inducing activation of transcription factors and among these copper can up-regulate MMP involved in release of NGF from pro-NGF. It is to note that intracellular copper is mainly in the reduced form Cu+. Monovalent copper, as zinc, may bind to the cysteine thiolate of tyrosine phosphatases active sites, blocking its activity and maintaining a high level of phosphorylated tyrosine. Indeed, NGF and its mimicking peptides increased total tyrosine phosphorylation, an effect strengthened by copper ions and hampered by chelating agent, pointing to an effective role of intracellular metal content (Naletova et al., 2019).
Therefore, a pivotal question to explain the observed effects of metal on the NGF activity is: may NGF induce intracellular increase of copper contents?
Confocal microscopy data reported by us using fluorescent NGF mimicking peptides demonstrated how they perform also a ionophoric action which however required a specific recognition of the TrkA receptor (Naletova et al., 2019). These findings indicate that NGF/TrkA route regulates intracellular copper level and then could influence different factors involved in metal homeostasis as copper transporters system Ctr1, intracellular copper chaperones, glutathione and metal transcription factor.
It is to note that MMP expression and release due to the increase of intracellular copper may also control Aβ level through protease degradation processes so as to reduce the formation of Aβ neurotoxic oligomers increasing instead the amount of non toxic Aβ monomers. These latter are able to activate Akt and CREB, then crossing with NGF and copper ions signaling pathways. All these data reveal novel links between, NGF, copper ions and Aβ physiology, suggesting that AD onset can be risen by an altered balance between these three players.
Collectively, these findings highlight a relevant role of copper ions in the NGF activity under normal conditions promoting neurite outgrowth, CREB phosphorylation and then influencing cognitive processes and memory formation. The interaction of metals and neurotrophins provides a nexus between signalling and neuronal physiology, establishing a critical step in the study of neurodegenerative diseases.
Better understanding of copper driven NGF signal transduction and vice versa, the role of NGF in the copper homeostasis in neurons, could disclose new therapeutic interventions for the memory and cognitive processes maintenance as well as new pharmacological targets in AD.
Footnotes
Copyright license agreement: The Copyright License Agreement has been signed by the author before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
C-Editors: Zhao M, Li JY; T-Editor: Jia Y
References
- 1.Barnham KJ, Bush AI. Biological metals and metal-targeting compounds in major neurodegenerative diseases. Chem Soc Rev. 43:6727–6749. doi: 10.1039/c4cs00138a. [DOI] [PubMed] [Google Scholar]
- 2.Birkaya B, Aletta JM. NGF promotes copper accumulation required for optimum neurite outgrowth and protein methylation. J Neurobiol. 2005;63:49–61. doi: 10.1002/neu.20114. [DOI] [PubMed] [Google Scholar]
- 3.Bothwell M. NGF, BDNF, NT3, and NT4, neurotrophic factors. Handb Exp Pharmacol. 2014;220:3. doi: 10.1007/978-3-642-45106-5_1. [DOI] [PubMed] [Google Scholar]
- 4.Canu N, Amadoro G, Triaca V, Latina V, Sposato V, Corsetti V, Severini C, Ciotti MT, Calissano P. The intersection of NGF/TrkA signaling and amyloid precursor protein processing in Alzheimer’s disease neuropathology. Int J Mol Sci. 2017 doi: 10.3390/ijms18061319. doi: 103390/ijms18061319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cuello AC, Pentz R, Hall H. The brain NGF metabolic pathway in health and in Alzheimer’s pathology. Front Neurosci. 2019;13:62. doi: 10.3389/fnins.2019.00062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.James SA, Volitakis I, Adlard PA, Duce JA, Masters CL, Cherny RA, Bush AI. Elevated labile Cu is associated with oxidative pathology in Alzheimer disease. Free Radic Biol Med. 2012;52:298–302. doi: 10.1016/j.freeradbiomed.2011.10.446. [DOI] [PubMed] [Google Scholar]
- 7.Maitra R, Shamovsky IL, Wang W, Solc M, Lawrance G, Dostaler SM, Ross GM, Riopelle RJ. Differential effects of transition metal cations on the conformation and biological activities of nerve growth factor. Neurotox Res. 2000;2:321–341. [Google Scholar]
- 8.Mueller C, Schrag M, Crofton A, Stolte J, Muckenthaler MU, Magaki S, Kirsch W. Altered serum iron and copper homeostasis predicts cognitive decline in mild cognitive impairment. J Alzheimers Dis. 2012;29:341–350. doi: 10.3233/JAD-2011-111841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Naletova I, Satriano C, Pietropaolo A, Gianì F, Pandini G, Triaca V, Amadoro G, Latina V, Calissano P, Travaglia A, Nicoletti VG, La Mendola D, Rizzarelli E. The copper(II)-assisted connection between NGF and BDNF by means of nerve growth factor-mimicking short peptides. Cells. 2019 doi: 10.3390/cells8040301. doi: 103390/cells8040301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pandini G, Satriano C, Pietropaolo A, Gianì F, Travaglia A, La Mendola D, Nicoletti VG, Rizzarelli E. The inorganic side of NGF: copper(II) and zinc(II) affect the NGF mimicking signaling of the N-Terminus peptides encompassing the recognition domain of TrkA receptor. Front Neurosci. 2016;10:569. doi: 10.3389/fnins.2016.00569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Squitti R. Copper subtype of Alzheimer’s disease (AD): meta-analyses, genetic studies and predictive value of non-ceruloplasmim copper in mild cognitive impairment conversion to full AD. J Trace Elem Med Biol. 2014;28:482–485. doi: 10.1016/j.jtemb.2014.06.018. [DOI] [PubMed] [Google Scholar]
- 12.Travaglia A, Pietropaolo A, La Mendola D, Nicoletti VG, Rizzarelli E. The inorganic perspectives of neurotrophins and Alzheimer’s disease. J Inorg Biochem. 2012;111:130–137. doi: 10.1016/j.jinorgbio.2011.10.017. [DOI] [PubMed] [Google Scholar]