

Abstract
Although several studies have focused on the dynamics of bacterial food community, little is known about the variability of batch production and microbial changes that occur during storage. The aim of the study was to characterize the microbial spoilage community of minced pork meat samples, among different food production and storage, using both 16S rRNA gene sequencing and classical microbiology. Three batches of samples were obtained from four local Belgian facilities (A–D) and stored until shelf life under food wrap (FW) and modified atmosphere packaging (MAP, CO2 30%/O2 70%), at constant and dynamic temperature. Analysis of 288 samples were performed by 16S rRNA gene sequencing in combination with counts of psychrotrophic and lactic acid bacteria at 22°C. At the first day of storage, different psychrotrophic counts were observed between the four food companies (Kruskal-Wallist test, p-value < 0.05). Results shown that lowest microbial counts were observed at the first day for industries D and A (4.2 ± 0.4 and 5.6 ± 0.1 log CFU/g, respectively), whereas industries B and C showed the highest results (7.5 ± 0.4 and 7.2 ± 0.4 log CFU/g). At the end of the shelf life, psychrotrophic counts for all food companies was over 7.0 log CFU/g. With metagenetics, 48 OTUs were assigned. At the first day, the genus Photobacterium (86.7 and 19.9% for food industries A and C, respectively) and Pseudomonas (38.7 and 25.7% for food companies B and D, respectively) were dominant. During the storage, a total of 12 dominant genera (>5% in relative abundance) were identified in MAP and 7 in FW. Pseudomonas was more present in FW and this genus was potentially replaced by Brochothrix in MAP (two-sided Welch’s t-test, p-value < 0.05). Also, a high Bray-Curtis dissimilarity in genus relative abundance was observed between food companies and batches. Although the bacteria consistently dominated the microbiota in our samples are known, results indicated that bacterial diversity needs to be addressed on the level of food companies, batches variation and food storage conditions. Present data illustrate that the combined approach provides complementary results on microbial dynamics in minced pork meat samples, considering batches and packaging variations.
Keywords: minced meat, metagenetics, spoilage bacteria, modified atmosphere packaging, food wrap packaging
Introduction
Meat and meat products are highly perishable, with colonization and development of a variety of microorganisms, especially bacteria. This is due to complex nutrient-rich environment with chemical and physical conditions favorable to bacterial development (Nychas et al., 2008; Pennacchia et al., 2009; Chaillou et al., 2015; Garnier et al., 2017). Moreover, minced meat can be contaminated by different types of microorganisms from several sources, such as raw materials, equipment, environment and handling involved in the production process. Abiotic factors (temperature, gaseous atmosphere, pH, NaCl levels, etc.) can also select certain bacteria (Mann et al., 2016; Stellato et al., 2016; Rouger et al., 2018). However, it is well known that richness and abundance of microbiota present in food products, and especially meats, play an important role in the microbial safety and the shelf life of the products (Zhao et al., 2015; Pinu, 2016). Microbial growth on meat to unacceptable levels and the various metabolic activities contribute to its deterioration by altering the structure, color and flavor of the meat (Mann et al., 2016). This leading to a reduction in food quality to the point of not being edible for human consumption (Holm et al., 2013; Silbande et al., 2016; Stellato et al., 2016), with alterations in the sensorial qualities of the product, particularly the aspect, with discoloration and gas production, and the presence of an off-odors and off-flavors (Stoops et al., 2015). Thus, food spoilage is problematic for two main reasons: first, it renders food unfit for human consumption and, secondly, it results in significant economic losses (Dalcanton et al., 2013; Pinter et al., 2014; Den Besten et al., 2017).
As mentioned by Benson et al. (2014), the microbial population that colonizes and ultimately spoils minced pork meat is highly variable, depending on which groups of microbial taxa the product has been exposed to and perhaps even the order in which they are encountered. Using traditional cultivation methods, the microbial composition and diversity in fresh meat have been widely investigated (Zhao et al., 2015), but it is well known that traditional identification and culture-based methods for pathogens or food spoilage microbes are time-consuming (Pinu, 2016). Moreover, ecological studies at the genus-species level are required because the same storage conditions may affect differently the species in the same groups of bacteria (Pennacchia et al., 2011; Stoops et al., 2015), and because not all the members of this microbiota contribute to food spoilage. Several studies in meat microbiology have established that spoilage is caused only by a dominated fraction of the initial microbial association (Nychas et al., 2008). These spoilage microorganisms have been designated as Ephemeral/Specific Spoilage Organisms (E(S)SOs) (Benson et al., 2014; Zotta et al., 2019). Therefore, as discussed by De Filippis et al. (2013), the concept of succession of spoilage-related microbial groups is very important, and many studies have been performed to investigate the dynamics and changes of the meat microbiota during storage.
Developed during the last decades, the next generation sequencing methodologies provide a powerful tool to study microbial community structure and composition shifts at different stages of ripening, allowing the detection of minor bacterial populations (Riquelme et al., 2015), at variable taxonomic depth (Pothakos et al., 2014; Chaillou et al., 2015; Parente et al., 2016). The introduction of molecular methods, especially culture-independent approaches, have contributed to the exploration of various food microbiota (Galimberti et al., 2015; Pinu, 2016; Garofalo et al., 2017; Parlapani et al., 2018), as for beverages (Elizaquivel et al., 2015), vegetables (Lee et al., 2017; Gu et al., 2018; Liu et al., 2019), and for dairy (Nalbantoglu et al., 2014; Riquelme et al., 2015; Ceugniez et al., 2017; Porcellato et al., 2018), seafood (Li et al., 2018; Parlapani et al., 2018; Silbande et al., 2018), and meat products (Cocolin et al., 2004; Pennacchia et al., 2011; Nieminen et al., 2012; Benson et al., 2014; Greppi et al., 2015; Polka et al., 2015; Stoops et al., 2015; Zhao et al., 2015; Delhalle et al., 2016; Mann et al., 2016; Carrizosa et al., 2017; Cauchie et al., 2017; Kaur et al., 2017; Korsak et al., 2017; Peruzy et al., 2019; Vester Lauritsen et al., 2019), in order to assess the microbial levels and diversity of food and food products (Nieminen et al., 2012; Pothakos et al., 2014; Lee et al., 2017; Rouger et al., 2018). The interest of this method to characterize the dominant spoilage bacteria in pork meat and meat products was also described (Andritsos et al., 2012; Mann et al., 2016; Raimondi et al., 2018; Li et al., 2019; Peruzy et al., 2019).
In this context, the aim of the present study was to assess the microbial spoilage community and dynamics of minced pork meat samples, among different conditions of production and food storage, using both 16S rRNA gene sequencing and classical microbiology.
Materials and Methods
Sampling
Fresh minced pork meat (MPM) samples packed with a food wrap film were obtained from four local small and medium-sized Belgian manufacturers (food companies A, B, C, and D) at the day of the production, corresponding to the day of slaughtering. Three batches for each manufacturer were used, with a 1-week interval between sampling (Supplementary Figure S1).
According to the recipe MPM is composed of 100% minced pork meat (70% lean, 30% fat), no salt, no spices, no additives, no eggs and no sugar are added. At the day of the production, the water activity of this product was 0.98 ± 0.02 and the pH value was 5.80 ± 0.05 (n = 12). pH of the homogenized samples (5 g in 45 ml of KCl) was measured with a pH meter (Knick 765 Calimatic, Allemagne). The water activity was measured for homogenized samples on the basis of the relative humidity measurement of the air balance in the micro enclosure at 25 ± 0.4°C (Thermoconstanter TH200, Novasina, Switzerland).
Minced pork meat samples were packed (100 g), in triplicate, in two different types of non-sterile packaging.
The first packaging concerns a tray (187 × 137 × 36, polyester 10 μm, homo-polymer polypropylene 50 μm, NutriPack, France) under modified atmosphere (MAP, CO2 30%/O2 70% ± 0.1%) (Olympia V/G, Technovac, Italy) using packaging wrap (PP/EVOH/PP) with random gas measurements (CheckMate 3, Dansensor, France).
The second packaging concerns a tray (175 × 135 × 22, polystyrene) under food wrap packing (FW) using cling film (Clinofilm).
Food Storage
According to the requirements for implementing microbiological tests of chilled perishable and highly perishable foodstuffs (AFNOR, 2010, NF V01-003), MPM samples were stored during 3 days of shelf life under FW, and during 6 days under MAP packaging, at constant and dynamic temperature: at (i) 2°C (± 1°C), (ii) 8°C (± 1°C), (iii) 12°C (±1°C), and (iv) for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C (2/8°C ± 1°C), in climatic chambers (Sanyo MIR 254).
Samples were analyzed at the first day of inoculation (day 0) and at the last day of storage (day 3 in FW and day 6 in MAP, n = 288) (Supplementary Figure S1).
Plate Count Enumeration
Twenty-five grams of product were randomly collected from the trays at the surface and at depth, without homogenization, and put into a Stomacher bag with a mesh screen liner (80 μm pore size) (Biomérieux, Basingstoke, England, ref 80015) under aseptic conditions. Buffered peptone water (BPW, 10 g/L peptone, 5 g/L sodium chloride, #3564684, Bio-Rad, Marnes-la-Coquette, France) (225 mL) was automatically added to each bag (Dilumat, Biomérieux, Belgium) and the samples were homogenized for 2 min in a Stomacher (Bagmixer, Interscience, France). From this primary suspension, decimal dilutions in maximum recovery diluent (10 g/L peptone, 8.5 g/L sodium chloride, #CM0733, Oxoid, Hampshire, England) were prepared for microbiological analysis, and 0.1 mL aliquots of the appropriate dilutions were plated onto media for each analysis (Spiral plater, DW Scientific, England). Total viable counts (TVC) for the aerobic psychrotrophic flora were performed on plate count agar (PCA agar, #3544475, Bio-Rad, Marnes-la-Coquette, France), and for the lactic acid bacteria (LAB) on de Man, Rogosa and Sharpe (MRS agar, #CM0361, Oxoid, Hampshire, England), after incubation at 22°C (Pothakos et al., 2014) for 72 h (model 1535 incubator, Shel Lab, Sheldon Manufacturing Inc., United States).
DNA Extraction and 16S rDNA Amplicon Sequencing
Bacterial DNA was extracted from each primary suspension, previously stored at −80°C, using the DNEasy Blood and Tissue kit (QIAGEN Benelux BV, Antwerp, Belgium) following the manufacturer’s recommendations. The resulting DNA extracts were eluted in DNAse/RNAse free water and their concentration and purity were evaluated by means of optical density using the NanoDrop ND-1000 spectrophotometer (Isogen, Sint-Pieters-Leeuw, Belgium). DNA samples were stored at −20°C until used for 16S rDNA amplicon sequencing.
PCR-amplification of the V1-V3 region of the 16S rDNA library preparation were performed with the following primers (with Illumina overhand adapters), forward (5′-GAGAGTTTGATYMTGGCTCAG-3′) and reverse (5′-ACCGCGGCTGCTGGCAC-3′). Each PCR product was purified with the Agencourt AMPure XP beads kit (Beckman Coulter; Pasadena, CA, United States) and submitted to a second PCR round for indexing, using the Nextera XT index primers 1 and 2. Thermocycling conditions consisted of a denaturation step of 4 min at 94°C, followed by 25 cycles of denaturation (15 s at 94°C), annealing (45 s at 56°C) and extension (60 s at 72°C), with a final elongation step (8 min at 72°C). These amplifications were performed on an EP Mastercycler Gradient System device (Eppendorf, Hamburg, Germany). The PCR products of approximately 650 nucleotides were run on 1% agarose gel electrophoresis and the DNA fragments were plugged out and purified using a Wizard SV PCR purification kit (Promega Benelux, Leiden, Netherlands). After purification, PCR products were quantified using the Quanti-IT PicoGreen (ThermoFisher Scientific, Waltham, MA, United States) and diluted to 10 ng/μL. A final quantification, by quantitative (q)PCR, of each sample in the library was performed using the KAPA SYBR® FAST qPCR Kit (KapaBiosystems, Wilmington, MA, United States) before normalization, pooling and sequencing on a MiSeq sequencer using V3 reagents (Illumina, San Diego, CA, United States).
Bioinformatics Analysis
The 16S rRNA gene sequence reads were processed with MOTHUR (Schloss et al., 2009). The quality of all sequence reads was denoised using the Pyronoise algorithm implemented in MOTHUR. The sequences were checked for the presence of chimeric amplification using ChimeraSlayer (developed by the Broad Institute)1. The obtained read sets were compared to a reference data-set of aligned sequences of the corresponding region derived from the SILVA database of full-length rRNA gene sequences (version v1.2.11)2 implemented in MOTHUR (Pruesse et al., 2012; Pothakos et al., 2014; Cauchie et al., 2017). The final reads were clustered into operational taxonomic units (OTUs), using the nearest neighbor algorithm using MOTHUR with a 0.03 distance unit cut off. A taxonomic identity was attributed to each OTU by comparison to the SILVA database, using an 80% homogeneity cut off. As MOTHUR is not dedicated to the taxonomic assignment beyond the genus level, all unique sequences for each OTU were compared to the SILVA data-set 111, using a BLASTN algorithm. For each OTU, a consensus detailed taxonomic identification was given based upon the identity (<1% mismatch with the aligned sequence) and the metadata associated with the best hit (validated bacterial species or not) (Delcenserie et al., 2014; Cauchie et al., 2017).
16S rDNA Data Analysis
A correcting factor for 16S rDNA gene copy numbers was applied for any taxon i (Eq. 1) (Kembel et al., 2012; Louca et al., 2018).
| (Eq. 1) |
Where Ai is the real abundance of 16S genes from the taxon in the sample, Nk is the number of reads for the taxon in the sample k, and Ci is determined by the genomic 16S copy number of that taxon. To obtain each gene copy number, Ribosomal RNA Database (rrnDB) (Stoddard et al., 2015) and EzBioCloud database (Yoon et al., 2017) were used.
Then, to compare the relative abundance of OTUs, the number of reads of each taxon were normalized (Nri) as described by Chaillou et al. (2015). Reads counts of each taxon i in the sample k were divided by a sample-specific scaling factor (Si) (Eq. 2) (Fougy et al., 2016; Rouger et al., 2018):
| (Eq. 2) |
Where Ai is the real abundance of 16S genes from that taxon obtained with a correcting factor for 16S rDNA gene copy numbers, Sk is the normalization factor associated with sample k.
The sample-specific scaling factor was calculated by (Eq. 3):
| (Eq. 3) |
Where Sk is the sample-specific scaling factor associated with sample k, Tk is the number of total reads in the sample k, me is the median value of total reads for all the samples of the dataset. Reads counts of all samples were then transformed into a percentage of each OTUs.
All biosample raw reads were deposited at the National Center for Biotechnology Information (NCBI) and are available under de BioProject ID PRJNA551357. The raw data supporting the conclusions of this manuscript will be made available by EC to any qualified researcher.
Statistical Analysis
Statistical Analysis on Microbiological Results
Non-parametric statistical tests were used to compare the classical microbiology result between samples taken on the day of production and at the end of shelf life for a same temperature. With the help of R software (R Core Team, 2016), Kruskal-Wallis test was performed to make a comparison between the food industries on a certain day (i.e., day 0 or day 3) (stats package, kruskal.test function). An Analysis of Covariance (ANCOVA) was also performed to evaluate the interactions between the storage conditions and the food origin on psychrotrophic counts (FactoMineR package, AovSum function). All tests were considered as significant for a p-value of < 0.05.
Statistical Analysis on 16S rDNA Results
Alpha diversity for each sample was evaluated by richness estimation (Chao1 estimator), microbial biodiversity (inverse of the Simpson index, coverage), and the population evenness (Simpson evenness) using MOTHUR (version 1.40.5)3 (Riquelme et al., 2015; Zhao et al., 2015). Rarefaction curves were calculated for all samples to ensure that sequencing depth was sufficient: OTUs identified were plotted as a function of sequences obtained per sample. High diversity coverage was achieved with all curves reaching asymptotes from 3000 reads (Supplementary Figure S2). Using Explicet, alpha and beta diversity indices were also calculated with bootstrapped sequencing data4 (Robertson et al., 2013; Mann et al., 2016).
Beta-diversity was assessed with Explicet using the Bray-Curtis index on a 0-1 scale.
Using STAMP (v2+) software5, a 2-sided Welch’s t-test was performed on metagenetic results and confidence intervals were calculated according to the Newcombe-Wilson method. A Principal Component Analysis (PCoA) was also applied to classify and cluster samples according to the identified OTUs for the two packaging (Tukey-Kramer test in conjunction with an ANOVA) (Parlapani et al., 2018). The differences were considered significant for a corrected p-value of less than 0.05 (Parks et al., 2014).
Results
Microbiological Analysis
As expected, psychrotrophic and lactic aerobic counts increased during the shelf life with increasing the temperature (Tables 1, 2).
TABLE 1.
Results of psychrotrophic aerobic counts in minced pork meat samples according to the origin, the food packaging and the temperature of storage.
| Industries/packaging | Day 0 | End of the shelf life (day 3) | |||
| 2°C | 8°C | 12°C | 2/8°C | ||
| FW | |||||
| A | 5.6 ± 0.1 | 6.5 ± 0.6 | 8.3 ± 0.4∗ | 8.3 ± 0.5∗ | 8.3 ± 0.3∗ |
| B | 7.5 ± 0.4 | 7.5 ± 0.4 | 8.3 ± 0.0∗ | 8.3 ± 0.2∗ | 8.3 ± 0.9∗ |
| C | 7.2 ± 0.4 | 7.3 ± 0.5 | 7.8 ± 0.0 | 7.8 ± 0.2 | 7.6 ± 1.3 |
| D | 4.2 ± 0.4 | 4.6 ± 0.2 | 7.2 ± 0.2∗ | 8.3 ± 0.0∗ | 6.6 ± 0.2∗ |
| Kruskal-Wallis test | 9.43 (0.02)° | 8.74 (0.03)° | 9.02 (0.03)° | 5.71 (0.13) | 9.68 (0.02)° |
| MAP | |||||
| A | 5.6 ± 0.1 | 6.5 ± 0.1∗ | 7.9 ± 0.1∗ | 8.3 ± 0.3∗ | 7.9 ± 0.2∗ |
| B | 7.5 ± 0.4 | 7.9 ± 0.1 | 8.3 ± 0.0∗ | 8.3 ± 0.0∗ | 8.3 ± 0.0∗ |
| C | 7.2 ± 0.4 | 7.5 ± 0.2 | 7.6 ± 0.1 | 8.3 ± 0.1∗ | 7.8 ± 0.6 |
| D | 4.2 ± 0.4 | 5.2 ± 0.3∗ | 7.9 ± 0.1∗ | 8.1 ± 0.1∗ | 7.2 ± 0.1∗ |
| Kruskal-Wallis test | 9.43 (0.02)° | 10.39 (0.02)° | 9.68 (0.02)° | 3.45 (0.33) | 8.94 (0.03)° |
Values given as log CFU/g (mean ± SD, n = 3) at 2, 8, 12, and 2/8°C. FW (food wrap packaging), MAP (modified atmosphere packaging),° significant Kruskal-Wallis value (p < 0.05) with p-value between bracket, ∗significant Wilcoxon value (p < 0.05).
TABLE 2.
Results of lactic aerobic counts in minced pork meat samples according to the origin, the food packaging and the temperature of storage.
| Industries/packaging | Day 0 | End of the shelf life (day 6) | |||
| 2°C | 8°C | 12°C | 2/8°C | ||
| FW | |||||
| A | 5.2 ± 0.2 | 6.4 ± 0.4 | 7.8 ± 0.1∗ | 7.8 ± 0.2∗ | 7.4 ± 0.2∗ |
| B | 5.5 ± 0.6 | 5.5 ± 0.5 | 7.1 ± 0.3∗ | 7.9 ± 0.2∗ | 6.8 ± 0.4∗ |
| C | 5.2 ± 0.7 | 6.7 ± 0.2∗ | 7.4 ± 0.1∗ | 7.6 ± 0.1∗ | 7.0 ± 0.2∗ |
| D | 3.5 ± 0.2 | 4.4 ± 0.3∗ | 5.9 ± 0.4∗ | 7.5 ± 0.1∗ | 5.1 ± 0.3∗ |
| Kruskal-Wallis test | 8.90 (0.04)° | 9.15 (0.03)° | 9.67 (0.02)° | 7.62 (0.05) | 8.44 (0.04)° |
| MAP | |||||
| A | 5.2 ± 0.2 | 7.1 ± 0.2∗ | 8.0 ± 0.18∗ | 8.2 ± 0.09∗ | 8.2 ± 0.09∗ |
| B | 5.5 ± 0.6 | 6.6 ± 0.6∗ | 7.8 ± 0.21∗ | 7.7 ± 0.16∗ | 7.8 ± 0.15∗ |
| C | 5.2 ± 0.7 | 7.3 ± 0.2∗ | 7.6 ± 0.06∗ | 7.9 ± 0.09∗ | 7.5 ± 0.07∗ |
| D | 3.5 ± 0.2 | 5.2 ± 0.4∗ | 7.5 ± 0.07∗ | 7.8 ± 0.03∗ | 6.8 ± 0.24∗ |
| Kruskal-Wallis test | 8.90 (0.04)° | 8.44 (0.04)° | 9.05 (0.03)° | 8.27 (0.04)° | 9.45 (0.02)° |
Values given as log CFU/g (mean ± SD, n = 3) at 2, 8, 12, and 2/8°C. FW (food wrap packaging), MAP (modified atmosphere packaging),° Significant Kruskal-Wallis value (p < 0.05) with p-value between bracket, ∗significant t-student value (p < 0.05).
Compared to the TVC values, LAB counts showed highest results for food industries A and D.
At day 0, different microbiological counts were observed between food companies for TVC (Kruskal-Wallis test, H = 9.43, p-value = 0.02) and for LAB (Kruskal-Wallis tests, H = 8.90, p-value = 0.04). The lowest psychrotrophic populations were observed for food industries D (4.2 ± 0.4 log CFU/g) and A (5.6 ± 0.1 log CFU/g), whereas minced pork meat samples from B to C showed the highest results (7.5 ± 0.4 and 7.2 ± 0.4 log CFU/g, respectively).
At the end of the shelf life, the natural logarithm of the TVC for all food companies was over 7.0 log CFU/g. At this time, the Analysis of Covariance revealed also a significant effect of the food companies (p-value = 0.00000998) and the temperature of storage (p-value = 0.00000095) on microbial total counts. Psychrotrophic counts seems also to be influenced by the interaction of the food industry and the temperature (p-value = 000442), but not by others interactions terms (p-value > 0.05).
Carbon Dioxygen Production
As shown in Figure 1, carbon dioxygen values increased with highest temperatures, except for the food companies C and D which shown relatively stable measurements. Results at 2/8°C are not shown in this paper.
FIGURE 1.

Box plots show the carbon dioxide measurements at the end of the shelf life, for the four food companies (A–D) at (A) 2°C, (B) 8°C, and (C) 12°C. The boxes represent the interquartile range between the first (Q1) and the third (Q3) quartiles; the vertical black line insides the box is the median obtained from the three batches analyzed by food industries; the two dotted line is the difference of 25% below the Q1 or above the Q3. The presence of stars indicated that samples deviated significantly from the carbon dioxide value at day 0 (30.0 ± 0.1%).
Alpha Diversity of Bacteria With 16S rDNA Amplicon Sequencing
Over 4,200 reads per sample were generated with pyrosequencing. In total, 48 mains OTUs were assigned. The number of OTUs, the bacterial diversity, richness estimators and coverage are presented in Supplementary Tables S1–S3). The highest number of identified species was encountered for the food industries C and D.
Bacterial Communities at the Family and Genus Levels
The relative abundance results obtained by metagenetics analysis (expressed in%) in FW and MAP packaging at Family (Figure 2) and Genus (Figure 3) levels (>5%) are represented in cumulated histograms for all samples. These data including the relative abundance of sequences are also summarized in Supplementary Tables S4–S6). The taxa representing <5% in relative abundance were merged in the category of “Others.” “Others” in FW are mainly composed by the genera Bacillus, Carnobacterium, Enterococcus, Hafnia, Myroides, Rahnella, Staphylococcus, Serratia, Streptococcus, Weissella and Xanthomonas in FW. While it concerns Bacillus, Carnobacterium, Enterococcus, Hafnia, Rahnella, Staphylococcus, Streptococcus and Xanthomonas in MAP. Full data on taxa found in high (>5%) and low (<5%) frequencies will be made available by EC to any qualified researcher.
FIGURE 2.
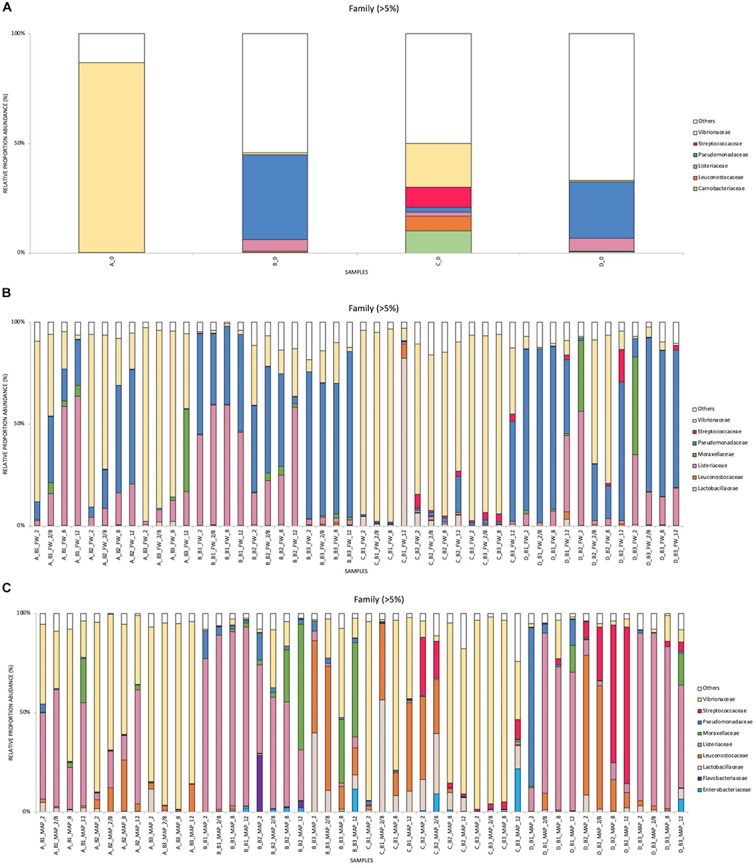
Cumulated histograms of the relative abundance (%) of taxa and the dynamics of the bacterial community identified by metagenetics at Family levels, during cold storage of minced pork meat in relation to the food packaging and the origin of samples (food companies and batches). (A) food samples analyzed at day 0 for the four companies (A–D), (B) storage in FW (food wrap) packaging, (C) storage in MAP (modified atmosphere) packaging. At Family levels, the taxa representing <5% in relative abundance were merged in the category of “Others”. Legend: batch 1 (B1), batch 2 (B2), batch 3 (B3), at 2°C (2), at 8°C (8), at 12°C (12), and for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C (2/8).
FIGURE 3.
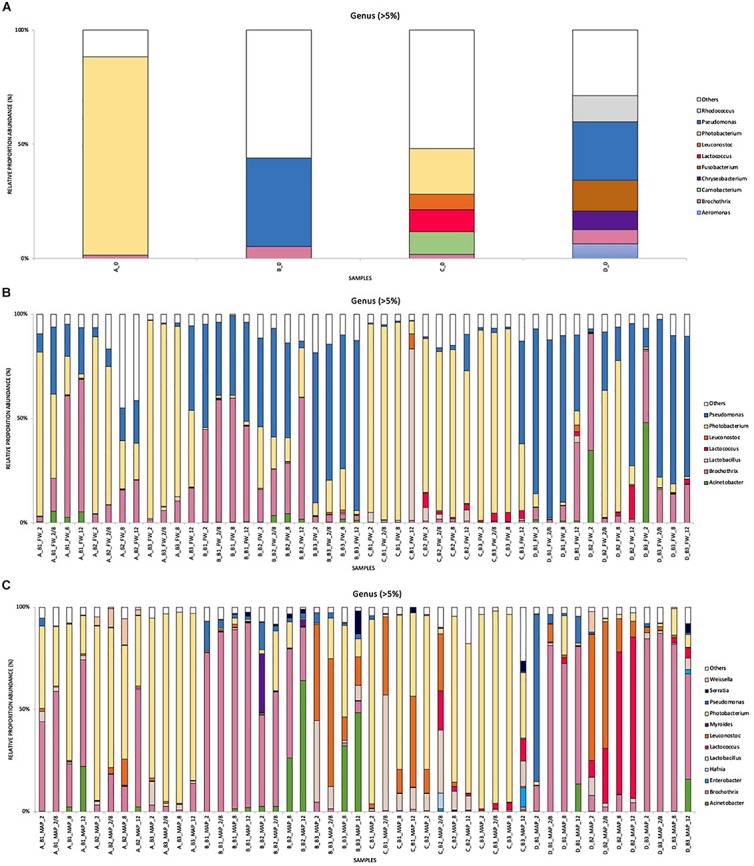
Cumulated histograms of the relative abundance (%) of taxa and the dynamics of the bacterial community identified by metagenetics at Genus levels, during cold storage of minced pork meat in relation to the food packaging and the origin of samples (food companies and batches). (A) food samples analyzed at day 0 for the four companies (A–D), (B) storage in FW (food wrap) packaging, (C) storage in MAP (modified atmosphere) packaging. At Genus levels, the taxa representing <5% in relative abundance were merged in the category of “Others”. These data including the relative abundance of sequences are also summarized in Supplementary Tables S4–S6. Legend: batch 1 (B1), batch 2 (B2), batch 3 (B3), at 2°C (2), at 8°C (8), at 12°C (12), and for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C (2/8).
According to Figures 2, 3, the food companies show a high variability in the distribution of read percentages at day 0. At this time, the genus Photobacterium is the most represented for A and C (86.7 and 19.9%, respectively), while it concerns the genus Pseudomonas for the industries B and D (38.7 and 25.7%, respectively).
At the end of the shelf life, a total of 12 genera were identified as dominant (taxa representing more than 5% in relative abundance) in MAP and only seven genera in FW. These seven genera are all identical to those found in MAP.
For all samples, the percentage of “unassigned” reads was relatively low (7.1 ± 3.7).
Effect of the Food Packaging on the Bacterial Communities
However, although dominant genera were identified across all samples, the two different types of packaging were characterized by different microbiota, with only some genera in common (Supplementary Figure S3). At the end of the shelf life, Pseudomonas was more present in FW and this genus was potentially replaced by Brochothrix in the MAP packaging (Welch’s t-test, p-value < 0.05) (Figure 4).
FIGURE 4.

Extended bar plot showing the bacterial populations whose mean relative abundance differed between food wrap (FW) and modified atmosphere (MAP) packaging at genus scale. The relative abundance and the difference in mean proportions are illustrated for the statistically different taxa (p < 0.05).
At this time, the major OTUs groups (Figure 5) are therefore different according to the food packaging: Brochothrix thermosphacta, Lactobacillus algidus, Photobacterium kishitanii, Photobacterium phosphoreum, Pseudomonas psychrophila, and Pseudomonas sp. are dominant in FW. While it concerns Acinetobacter sp., Brochothrix thermosphacta, Lactobacillus algidus, Lactococcus piscium, Leuconostoc inhae, Leuconostoc gelidum, Leuconostoc sp., Photobacterium kishitanii, Photobacterium phosphoreum, and Pseudomonas sp. in MAP.
FIGURE 5.

Heatmap of relative read abundance at species level for all samples (expressed in %) among the different storage conditions. Only the most abundant OTUs obtained in this study are specially indicated (>1%). Others OTUs are gathered in “Others OTUs.” Legend: food companies (A–D), with three batches each (B1, B2, B3), analyzed at the first (0) and the last day of storage, in food wrap (FW) and modified atmosphere (MAP) packaging. Temperature of storage: 2°C (2), 8°C (8), 12°C (12), and for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C (2/8).
Variability of the Minced Pork Meat Ecosystem Between Samples
Genus relative abundance shows a high Bray-Curtis dissimilarity during the storage, and between the food companies and batches (Figure 6).
FIGURE 6.

Global microbial dissimilarity obtained by metagenetics between samples for different conditions of storage. The heatmap shows the Bray-Curtis dissimilarity measure based on relative abundance of OTUS (genus scale). Values are given in dissimilarity counts (1 = 100% dissimilar, 0 = 0% dissimilar). Legend: analysis at day 0 (0), at day 3 (3) and at day 6 (6); food companies (A–D); three batches each (1, 2, 3); in food wrap (FW) and in modified atmosphere (MAP) packaging. Temperature of storage: 2°C in FW (A), for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C in FW (B), 8°C in FW (C), 12°C in FW (D), 2°C in MAP (E), for a third of the shelf life at 2°C and for the rest of the shelf life at 8°C in MAP (F), 8°C in MAP (G), 12°C in MAP (H).
At day 0, samples showed a high dissimilarity (>70%) with the metadata groupings at the end of the shelf life. At this time, the food company A seems not to shared OTUs in common with the three others food industries.
At the end of the shelf life, Bray-Curtis index seems indicating that a relative similarity exists for OTUs contained within food companies A and C, and within B and D. This index also indicates a relative similarity concerning the temperature of storage, except for the industry D.
A synthetic view about the Bray-Curtis index between samples according to the food origin and storage condition is summarized in Table 3.
TABLE 3.
Dominant bacteria represented in minced pork meat samples according to storage conditions.
| Food companies | First day of storage | Last day of storage | ||
| Batch | FW | MAP | ||
| A |
Photobacterium sp., Ph. phosphoreum |
1 |
Brochothrix thermosphacta, Photobacterium kishitanii, Pseudomonas sp. |
B. thermosphacta, Ph. kishitanii |
| 2 |
B. thermosphacta, Ph. kishitanii, Pseudomonas sp. |
B. thermosphacta, Ph. kishitanii, Weissella sp. |
||
| 3 |
Ph. phosphoreum, Pseudomonas sp. |
Ph. phosphoreum | ||
| B |
Pseudomonas sp., Ps. psychrophila |
1 |
B. thermosphacta, Ps. psychrophila |
B. thermosphacta, Ps. psychrophila |
| 2 |
B. thermosphacta, Photobacterium sp., Pseudomonas sp. |
Acinetobacter sp., B. thermosphacta, Photobacterium sp. |
||
| 3 |
Ph. kishitanii, Ph. phosphoreum, Pseudomonas sp. |
Acinetobacter sp., Lactobacillus sp., Leuconostoc sp., Ln. gelidum, Photobacterium sp., Ph. kishitanii |
||
| C |
Photobacterium sp., Ph. kishitanii |
1 |
Lactobacillus algidus, Ph. kishitanii |
Lb. algidus, Ln. carnosum, Ln. inhae, Ph. kishitanii |
| 2 |
Photobacterium sp., Ph. kishitanii, Pseudomonas sp., Ps. phychrophila |
Lb. algidus, Lactococcus piscium, Ln. inhae, Ph. kishitanii |
||
| 3 |
Ph. kishitanii, Pseudomonas sp. |
Ph. kishitanii | ||
| D |
Pseudomonas sp., Ps. psychrophila, Ps. syncyanea |
1 |
B. thermosphacta, Pseudomonas sp., |
B. thermosphacta, Photobacterium sp., Pseudomonas sp. |
| 2 |
Acinetobacter sp., B. thermosphacta, Photobacterium sp., Ps. psychrophila |
B. thermosphacta, Lc. piscium, Ln. gelidum, Ln. inhae |
||
| 3 |
Acinetobacter sp., Brochothrix sp., B. thermosphacta, Pseudomonas sp., |
B. thermosphacta, Ph. kishitanii |
||
At species level, the taxa representing <20% in relative abundance were not considered as dominant in this table. FW (food wrap packaging), MAP (modified atmosphere packaging).
Discussion
In this study, we investigated the microbial spoilage community and dynamics of minced pork meat samples, among different conditions of production and food storage, using both 16S rRNA gene sequencing and classical microbiology. Indeed, whereas the dynamics of the bacterial community of meat and meat products have been studied before, Stoops et al. (2015) reported that little is known about differences in microbial changes during storage, and among the variability of batches production. Meat and meat products are highly perishable, with colonization and development of a great variety of microorganisms (Nychas et al., 2008; Pennacchia et al., 2009; Chaillou et al., 2015; Stellato et al., 2016; Garnier et al., 2017). The product composition (low/high pH, low/high concentration of glucose, water activity, …) and the storage conditions (temperature of storage and packaging conditions for example) may favor growth of microorganisms, that are responsible for the formation of spoilage (Argyri et al., 2015; Reid et al., 2017). This can lead to visible growth (slime, colonies), as textural changes, off-odors or off-flavors (Casaburi et al., 2014; Chaillou et al., 2015; Stoops et al., 2015; Del Blanco et al., 2017). In this context, minced meat is a potentially hazardous food product, vulnerable to bacterial spoilage, with a very short shelf life (Geeraerts et al., 2017) due to abundant and diverse substrate for bacterial growth and favorable growth conditions (Benson et al., 2014). In our study, the minced pork meat samples present a high water activity and a near-neutral pH which are in accordance with previous studies on this food matrix (Blixt and Borch, 2002; Andritsos et al., 2012).
The initial contamination of products, and also the initial level of lactic acid bacteria, is also a key factor that can influence the spoilage dynamic during storage (De Filippis et al., 2013). In our results, the microbial counts of the four manufacturers were quite different and psychrotrophic counts were higher for two food industries (Tables 1, 2). High levels of initial contamination in minced pork meat samples were also observed by Peruzy et al. (2019). This difference of the initial bacterial contamination is not in relation with the size of the company. These results can be explained by the fact that multiple sources of contamination can contribute to the initial composition of the meat microbiota (De Filippis et al., 2013), such as at the farm (hygiene practices, the conditions of animal transport, etc.) and at the slaughterhouse (automatic level of the process, cleaning practices, etc.). Initial carcass contamination can be also environmental, with contamination by tools, machines, and surfaces of slaughter equipment (Mann et al., 2016; Moretro et al., 2016). In addition, subsequent handling of meat in the operations of slicing, sectioning, portioning, and transferring in packages can determine further contamination in the handling points (Del Blanco et al., 2017).
The bacterial count at the end of the shelf life was over 7.0 log CFU/g, indicating that meat had probably begun to be deteriorated and would not be suitable for human consumption (Zhao et al., 2015). Indeed, it is generally recognized that microbial spoilage of meat occurs when counts reach arbitrary level between 7.0 log CFU/g (Nychas et al., 2008; Pothakos et al., 2014; Stoops et al., 2015; Reid et al., 2017; Spanu et al., 2018) and 8.00 log CFU/g (Nychas et al., 2008; Fall et al., 2012; Pothakos et al., 2014; Chaillou et al., 2015; Reid et al., 2017). However, these values are only indicative and refer here to the total viable count. Food spoilage needs to be assessed to the genus-species level, because potentially protective bacteria can also occur in food products.
As discussed by Del Blanco et al. (2017), common approaches for delaying meat spoilage and improving meat shelf life are available, including good hygienic practices and all the storage conditions. Among these, low storage temperatures and adequate packaging are considered as the most important factors (Koutsoumanis et al., 2006; Andritsos et al., 2012; Kaur et al., 2017). During the storage at 2°C, the arbitrary level of 7.0 log CFU/g was sometimes not reached. In addition, it can be observed that the microbial kinetics from 2 to 8°C were quite similar to those at 8°C, as described by Cauchie et al. (2017).
In relation with the food packaging, the most common used in meat and meat products are vacuum packaging and modified atmosphere packaging (MAP) (Caryé et al., 2005; Koutsoumanis et al., 2008; Dalcanton et al., 2013; Chaix et al., 2015; Silbande et al., 2016). In this study, a food wrap (FW) and a MAP (30% CO2 – 70% O2) packaging are used. The composition of modified atmosphere systems can be an effective way to reduce the growth rate of spoilage aerobic organisms and modify the microbial ecology of the product. But their effectiveness strongly depends on the initial microbial contamination of raw materials, storage temperature, film permeability and the carbon dioxide concentration used (20–40% is commonly used to suppress microbial growth) (Simpson and Carevic, 2004; Rotabakk et al., 2006; Stoops et al., 2015; Guillard et al., 2016; Saraiva et al., 2016; Couvert et al., 2017). The carbon dioxide concentration was here theoretically sufficient to limit the microbial growth. However, the higher percentage of oxygen can also enhance the growth of aerobic microbial communities in our samples. Moreover, some bacteria are able to grow in variable food packaging, as Photobacterium which is CO2-tolerant (Dalgaard, 1995; Fuertez-Perez et al., 2019). Also, in accordance with Stoops et al. (2015), it can be observed a significant production of carbon dioxide. This production may be the reflect of the development of bacterial groups belonging to lactic acid bacteria, Brochothrix or Enterobacteriaceae (Caryé et al., 2005). As environment of slaughtering and processing steps (Stellato et al., 2016), packaging materials can also be a source of contamination because they are not sterile in study. Further studies based on microbial contamination of food trays would also be interesting.
According to this, and based on the study by Stoops et al. (2015), viable counts are not suitable to characterize the microbial diversity of food products and to investigate thoroughly shifts in the bacterial communities during storage. Indeed, culture-dependent techniques largely underestimated the species richness and abundance. For a more detailed characterization of microbial communities in samples, originating from different ecological niches, a sequence-based approach was used, allowing identification of OTUs at various taxonomic levels (species, genus or family levels) (Stoops et al., 2015). However, without extensive studies involving a large number of samples under different storage conditions it will not be possible to determine exactly the bacterial ecosystem and the role of individual spoilage species (Pennacchia et al., 2011; Rouger et al., 2018). According to this, we analyzed minced meat samples from four different food companies, with three different batches per industries. In addition to previous studies based on the microbial description of minced meat samples (Stoops et al., 2015; Peruzy et al., 2019), our study aims to understand and monitor microbial dynamics and variability between food companies and food batches, according to different storage conditions.
In our results, the observed microbial diversity was relatively high, and the most abundant bacteria differ among samples. As observed by Stoops et al. (2015) in minced meat samples, an increase of microbial counts is coinciding with a decrease in bacterial diversity during storage. At the end of the storage period, the major genus taxa are represented by Pseudomonas in FW and Brochothrix in MAP. But it can also be observed a high diversity between food companies and batches (Table 3). Our results are in accordance with Peruzy et al. (2019), which also observed a dominance of the genus Pseudomonas, Brochothrix, and Carnobacterium in minced pork meat samples. Moreover, these results are not surprising because the microbial populations of refrigerated meat and pork-meat products are mainly composed by Pseudomonas spp., cold tolerant Enterobaceriaceae, lactic acid bacteria (such as Lactobacillus spp., Lactococcus spp., Leuconostoc spp., Carnobacterium spp., etc.), Brochothrix thermosphacta, Clostridium spp. (Koort et al., 2005; Liu et al., 2006; Nychas et al., 2008; Pennacchia et al., 2009, 2011; Casaburi et al., 2014; Stellato et al., 2016; Del Blanco et al., 2017; Geeraerts et al., 2017) and Weissella spp. (Pothakos et al., 2014; Stellato et al., 2016). Other genera isolated frequently from fresh pork meats are Acinetobacter spp., Aeromonas spp., Enterococcus spp., and Moraxella spp. (Zhao et al., 2015; Mann et al., 2016). However, these results are not completely in accordance with Stoops et al. (2015) because this study mentioned that Lactobacillus algidus and Leuconostoc sp. became the dominant bacteria in minced meat samples stored at 5°C under modified atmosphere (66% O2, 25% CO2, and 9% N2). These differences can be explained by different meat compositions (beef in the study by Stoops et al. (2015) and pork in our study), the initial contamination of samples, and the gas mixture used.
The results also showed the interest of using culture-independent method to better understand the changes of food microbiota over time, and in each food companies, according to the storage conditions. Indeed, metagenetics approach produce a large amount of data in a very short time (Cocolin et al., 2018; Den Besten et al., 2018), allowing to interpret and use these data to help agri-food companies in their decisions regarding food safety and quality decisions. Moreover, all the OTUs-species described as potentially spoilers in our study are well described in the literature (Table 4), and in minced pork meat samples (Stoops et al., 2015; Peruzy et al., 2019). The bacterial species present in our samples are also able to grow in meat matrices, and they are potentially responsible of spoilage effects, which can affect color, flavor, visual aspect, etc. (Pothakos et al., 2015). Sensory analyses would be interesting in this context, but were not performed in this study. Moreover, the enzymatic decarboxylation of amino acids, or the transamination of aldehydes and ketones, by bacteria results in the formation and accumulation of biogenic amines (BAs) (Jastrzębska et al., 2016). Biogenic amines (e.g., b-phenylethylamine, cadaverine, histamine, putrescrine, spermidine, spermine, tyramine and tryptamine) are reported in various foods including meat, fish, cheese, and wine (Papageorgiou et al., 2018). They can have health implications, such as allergic reactions, but also contribute to spoilage due to their putrid aroma (Stanborough et al., 2017). Therefore, as proposed by Cheng et al. (2016), the sum of BAs could be used as an indicator of pork meat quality and freshness during storage. Li et al. (2014) also showed that some BAs could be used as spoilage indicators of chilled pork.
TABLE 4.
Examples of some microbial species occurring during chilled storage of meat and their potential spoilage effects.
| Bacteria | Growth conditions | Spoilage effects | References |
| Actinetobacter spp. | Especially present in dairy and seafood products. | Low spoilage potential but can enhanced the growth of other spoilage bacteria by means of quorum sensing. | Pinu, 2016; Ghasemi-Varnamkhasti et al., 2018; Odeyemi et al., 2018; Hahne et al., 2019 |
| Brochothrix spp. | In different gas composition, such as under air, modified atmosphere and vacuum-packaging. More tolerant in oxygen-depleted and CO2-enriched environments. | Sour, acid and cheesy odor. | Koutsoumanis et al., 2008; Nychas et al., 2008; Ercolini et al., 2011; Doulgeraki et al., 2012; Zhao et al., 2015; Mann et al., 2016; Del Blanco et al., 2017; Reid et al., 2017; Mansur et al., 2019 |
| Carnobacterium spp. | In all types of packaging conditions. Predominance in low O2 packaging. | Spoilage effect can vary, producing volatile molecules with low sensory impacts (fruity or fermented odors, …) | Casaburi et al., 2011; Doulgeraki et al., 2012; Pothakos et al., 2015 |
| Lactobacillus spp. (Lb. sakei, Lb. fuchuensis, Lb. plantarum, Lb. curvatus, Lb. algidus, Lb. oligofermentans, …) | In all types of packaging conditions. Predominance with high concentration of CO2. | Severe acidification, emission of off-odor compounds and ropy slime. However, lactic acid bacteria may produce lactic acid, which inhibits the growth of other families of bacteria. And some species can produce bacteriocins. | Kato et al., 2000; Fadda et al., 2010; Doulgeraki et al., 2012; Dalcanton et al., 2013; Nieminen et al., 2015; Pothakos et al., 2015; Zhao et al., 2015; Alvarez-Sieiro et al., 2016; Mann et al., 2016; Woraprayote et al., 2016; Stefanovic et al., 2017; Mansur et al., 2019 |
| Lactococcus spp. | In various types of packaging. | Traditionally they have not been considered as spoilage microorganisms, but the spoilage potential of these bacteria is still scarcely known. | Kato et al., 2000; Doulgeraki et al., 2012; Rahkila et al., 2012; Dalcanton et al., 2013; Pothakos et al., 2014; Zhao et al., 2015; Mann et al., 2016; Mansur et al., 2019 |
| Leuconostoc spp. (Ln. gelidum, Ln. carnosum, Ln. mesenteroides, …) | Under aerobic, vacuum and modified atmosphere packaging. Predominance with high concentration of O2. | Buttery aroma, formation of slime, blowing of packages, green discoloration. | Kato et al., 2000; Doulgeraki et al., 2012; Dalcanton et al., 2013; Nieminen et al., 2015; Pothakos et al., 2015; Zhao et al., 2015; Mann et al., 2016; Mansur et al., 2019 |
| Photobacterium spp. | Under air, vacuum and modified atmosphere packaging. More frequently present in seafood products. | Typically not associated with spoilage of meat. Responsible for reducing TMAO to TMA, off-odor (produce volatile organic compounds) and biogenic amine formation. The mechanism underlying spoilage has not been clarified. | Nieminen et al., 2016; Li et al., 2019 |
| Pseudomonas spp. | In different gas composition, such as under air, modified atmosphere and vacuum-packaging. Predominance under aerobic low temperature. Limitation in the bacterial flora by the presence of CO2 and/or the limitation of O2 in MAP packaging. | Slime, discoloration, off-odor producing. | Koutsoumanis et al., 2008; Nychas et al., 2008; Ercolini et al., 2011; Andritsos et al., 2012; Doulgeraki et al., 2012; Zhao et al., 2015; Mann et al., 2016; Del Blanco et al., 2017; Reid et al., 2017; Liu et al., 2018; Spanu et al., 2018; Mansur et al., 2019 |
| Weissella spp. | Some can be found in salted and fermented foods. Present in vacuum packaging. | Greenish appearance. Can plays an important role in the fermentation process. Some species can produce bacteriocins. | Pothakos et al., 2015; Martins et al., 2016; Kim et al., 2017; Kariyawasam et al., 2019 |
However, it is important to add that some bacteria can be considered as protective, such as some lactic acid bacteria. As mentioned by Singh (2018), the presence of high LAB communities does not necessarily result in quality defect, and their intra-species variation to cause spoilage has already been recognized (Pothakos et al., 2015).
In the present study, we designed a method to collect MPM samples in order to explore the bacterial communities and diversity among different food origin and storage conditions. Indeed, the modification of the composition of the spoilage flora during storage is an important factor in assessing food quality (Holm et al., 2013). Although the bacteria consistently dominated the microbiota of MPM samples are known, results indicated that bacterial diversity needs to be addressed on the level of food companies and batches variations. As discussed by Rouger et al. (2017), it is important to overcome variability to better understand the factors underlying the diversity of spoilage bacterial communities, by (i) defining reproducible and reliable experimental conditions to lead to biological interpretation, or (ii) to multiplying sampling or experiments to obtain statistical significance of the results (Chaillou et al., 2015; Rouger et al., 2017). A seasonal effect on the microbial quality of minced meat has also been reported by Andritsos et al. (2012). In this paper, no conclusions about bacterial ecosystems for others food companies, or for different times of the year, should be dawn. Further data are so needed to determine diversity of spoilage microbiota in minced pork meat samples, according to others food industries, sampling periods and storage conditions. Also, a comparative evaluation of spoilage-related bacterial species and metabolic profiles, with growth parameters of these potentially spoilage bacteria in samples, will be studied in another study.
In conclusion, the combination of both culture-dependent and culture-independent analyses enabled us to explore the microbial communities of minced pork meat samples under different food origin and storage conditions, as previously described by Stoops et al. (2015). In our study, microbial changes during storage were monitored, according to a sampling in four food companies and for several batches. In accordance with previous studies we found that Pseudomonas and Brochothrix dominate the community at the end of the shelf life in FW and MAP, respectively, together with Photobacterium. The major OTUs groups are also often associated with pork meat spoilage in the scientific literature. And these results are also in accordance with studies conducted on the microbiota of minced meat by Stoops et al. (2015) and Peruzy et al. (2019). Psychrophilic spoilers dominated the microbiota of our samples, but each sample harbored a unique pork meat microbiota, depending on the manufacturing batch and the packaging used. The gas mixture and the temperature condition used in this study are probably the most important factors implied to the dynamics of the bacterial community. Further researches on the main contamination during slaughter production process, such as importance of processing environment, procedures and storage conditions, are desirable to provide a complete assessment of the microbiome of minced meat and to limit incidents of unexpected spoilage.
Data Availability Statement
All biosample raw reads were deposited at the National Center for Biotechnology Information (NCBI) and are available under the BioProject ID PRJNA551357.
Author Contributions
EC did the experiments, interpreted the results and wrote the manuscript. LD performed the experiments, supervised analyses and revised the manuscript. BT, PF, FF, GB, and GD were involved in the design of the study and provided help for interpretation of the results. AT and SB participated to the experiments. NK participated to the design of the study, interpretation of the results and writing of the manuscript. All authors read and approved the final manuscript.
Conflict of Interest
PF and SB (Quality Partner sa, Liège, Belgium) were employed by the Department of Food Sciences (Faculty of Veterinary Medicine, University of Liège, Liège, Belgium) to perform 16S rRNA gene amplicon sequencing. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the producers who provided the minced pork meat samples.
Footnotes
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.03074/full#supplementary-material
References
- AFNOR, (2010). Hygiène Des Aliments, Lignes Directrices pour la Réalisation de Tests de Vieillissement Microbiologique, Aliments Périssables et très Périssables Réfrigérés, NF V01-003. France: Association française de normalisation. [Google Scholar]
- Alvarez-Sieiro P., Montalban-Lopez M., Mu D., Kuipers O. P. (2016). Bacteriocins of lactic acid bacteria: extending the family. Appl. Microbiol. Biotechnol. 100 2939–2951. 10.1007/s00253-016-7343-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andritsos N. D., Mataragas M., Mavrou E., Stamatiou A., Drosinos E. H. (2012). The microbiological condition of minced pork prepared at retail stores in Athens. Greece. Meat Sci. 91 486–489. 10.1016/j.meatsci.2012.02.036 [DOI] [PubMed] [Google Scholar]
- Argyri A. A., Mallouchos A., Panagou E. S., Nychas G.-J. E. (2015). The dynamics of the HS/SPME-GC-MS as a tool to assess the spoilage of minced beef stored under different packaging and temperature conditions. Int. J. Food Microbiol. 193 51–58. 10.1016/j.ijfoodmicro.2014.09.020 [DOI] [PubMed] [Google Scholar]
- Benson A. K., David J. R. D., Gilbreth S. E., Smith G., Nietfeldt J., Legge R., et al. (2014). Microbial successions are associated with changes in chemical profiles of a model refrigerated fresh pork sausage during an 80-day shelf life study. Appl. Environ. Microbiol. 80 5178–5194. 10.1128/AEM.00774-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blixt Y., Borch E. (2002). Comparison of shelf life of vacuum-packed pork and beef. Meat Sci. 60 371–378. 10.1016/S0309-1740(01)00145-0 [DOI] [PubMed] [Google Scholar]
- Carrizosa E., Benito M. J., Ruiz-Moyano S., Hernandez A., del Carmen Villalobos M., Martin A., et al. (2017). Bacterial communities of fresh goat meat packaged in modified atmosphere. Food Microbiol. 65 57–63. 10.1016/j.fm.2017.01.023 [DOI] [PubMed] [Google Scholar]
- Caryé M. E., Garro O., Vignolo G. (2005). Effect of storage temperature and gas permeability of packaging film on the growth of lactic acid bacteria and Brochothrix thermosphacta in cooked meat emulsions. Food Microbiol. 22 505–512. 10.1016/j.fm.2005.01.003 [DOI] [Google Scholar]
- Casaburi A., Nasi A., Ferrocino I., Di Monaco R., Mauriello G., Villani F., et al. (2011). Spoilage-related activity of Carnobacterium maltaromaticum strains in air-storedand vacuum-packed meat. Appl. Environ. Microbiol. 77 7382–7393. 10.1128/AEM.05304-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casaburi A., Piombino P., Nychas G.-J., Villani F., Ercolini D. (2014). Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 45 83–102. 10.1016/j.fm.2014.02.002 [DOI] [PubMed] [Google Scholar]
- Cauchie E., Gand M., Kergourlay G., Taminiau B., Delhalle L., Korsak N., et al. (2017). The use of the 16S rRNA gene metagenetic monitoring of refrigerated food products for understanding the kinetics of microbial subpopulations at different storage temperatures: the example of white pudding. Int. J. Food Microbiol. 247 70–78. 10.1016/j.ijfoodmicro.2016.10.012 [DOI] [PubMed] [Google Scholar]
- Ceugniez A., Taminiau B., Coucheney F., Jacques P., Delcenserie V., Daube G., et al. (2017). Use of a metagenetic approach to monitor the bacterial microbiota of ≪ Tomme d’Orchies ≫ cheese during the ripening process. Int. J. Food Microbiol. 247 65–69. 10.1016/j.ijfoodmicro.2016.10.034 [DOI] [PubMed] [Google Scholar]
- Chaillou S., Chaulot-Talmon A., Caekebeke H., Cardinal M., Christieans S., Denis C., et al. (2015). Origin and ecological selection of core and food-specific bacterial communities associated with meat and seafood spoilage. Int. Soc. Microb. Ecol. J. 9 1105–1118. 10.1038/ismej.2014.202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaix E., Guillaume C., Gontard N., Guillard V. (2015). Diffusivity and solubility of CO2 in dense solid food products. J. Food Eng. 166 1–9. 10.1016/j.jfoodeng.2015.05.023 [DOI] [Google Scholar]
- Cheng W., Sun D.-W., Cheng J.-H. (2016). Pork biogenic amine index (BAI) determination based on chemometric analysis of hyperspectral imaging data. LWT Food Sci. Technol. 73 13–19. 10.1016/j.lwt.2016.05.031 [DOI] [Google Scholar]
- Cocolin L., Mataragas M., Bourdichon F., Doulgeraki A., Pilet M.-F., Jagadeesan B., et al. (2018). Next generation microbiological risk assessment meta-omics: the next need for integration. Int. J. Food Microbiol. 287 10–17. 10.1016/j.ijfoodmicro.2017.11.008 [DOI] [PubMed] [Google Scholar]
- Cocolin L., Rantsiou K., Iacumin L., Urso R., Cantoni C., Comi G. (2004). Study of the ecology of fresh sausages and characterization of populations of lactic acid bacteria by molecular methods. Appl. Environ. Microbiol. 70 1883–1894. 10.1128/AEM.70.4.1883-1894.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couvert O., Guégan S., Hézard B., Huchet V., Lintz A., Thuault D., et al. (2017). Modeling carbon dioxide effect in a controlled atmosphere and its interactions with temperature and pH on the growth of L. monocytogenes and P. fluorescens. Food Microbiol. 68 89–96. 10.1016/j.fm.2017.07.003 [DOI] [PubMed] [Google Scholar]
- Dalcanton F., Pérez-Rodriguez F., Posada-Izquierdo D., de Arageo G. M.-F., Garcia-Gimeno R. M. (2013). Modelling growth of Lactobacillus plantarum and shelf life of vacuum-packaged cooked chopped pork at different temperatures. Int. J. Food Sci. Technol. 48 2580–2587. 10.1111/ijfs.12252 [DOI] [Google Scholar]
- Dalgaard P. (1995). Qualitative and quantitative characterization of spoilage bacteria from packed fish. Int. J. Food Microbiol. 26 319–333. 10.1016/0168-1605(94)00137-U [DOI] [PubMed] [Google Scholar]
- De Filippis F., La Storia A., Villani F., Ercolini D. (2013). Exploring the sources of bacterial spoilers in beefsteaks by culture-independent high-throughput sequencing. PLoS One 8:e70222. 10.1371/journal.pone.0070222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Del Blanco A., Caro I., Quinto E. J., Mateo J. (2017). Quality changes in refrigerated stored minced pork wrapped with plastic cling film and the effect of glucose supplementation. Meat Sci. 126 55–62. 10.1016/j.meatsci.2016.12.007 [DOI] [PubMed] [Google Scholar]
- Delcenserie V., Taminiau B., Delhalle L., Nezer C., Doyen P., Crevecoeur S., et al. (2014). Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 97 6046–6056. 10.3168/jds.2014-8225 [DOI] [PubMed] [Google Scholar]
- Delhalle L., Korsak N., Taminiau B., Nezer C., Burteau S., Delcenserie V., et al. (2016). Exploring the bacterial diversity of Belgian steak tartare using metagenetics and quantitative real-time PCR analysis. J. Food Prot. 79 220–229. 10.4315/0362-028X.JFP-15-185 [DOI] [PubMed] [Google Scholar]
- Den Besten H. M. W., Amézquita A., Bover-Cid S., Dagnas S., Ellouze M., Guillou S., et al. (2018). Next generation of microbiological risk assessment: potential of omics data for exposure assessment. Int. J. Food Microbio. 287 18–27. 10.1016/j.ijfoodmicro.2017.10.006 [DOI] [PubMed] [Google Scholar]
- Den Besten H. M. W., Aryani D. C., Metselaar K. I., Zwietering M. H. (2017). Microbial variability in growth and heat resistance of a pathogen and a spoiler: all variabilities are equal but some are more equal than others. Int. J. Food Microbiol. 240 24–31. 10.1016/j.ijfoodmicro.2016.04.025 [DOI] [PubMed] [Google Scholar]
- Doulgeraki A. I., Ercolini D., Villani F., Nychas G.-J. E. (2012). Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 157 130–141. 10.1016/j.ijfoodmicro.2012.05.020 [DOI] [PubMed] [Google Scholar]
- Elizaquivel P., Pérez-Cataluna A., Yépez A., Aristimuno C., Jiménez E., Cocconcelli P. S., et al. (2015). Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chica, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 198 9–18. 10.1016/j.ijfoodmicro.2014.12.027 [DOI] [PubMed] [Google Scholar]
- Ercolini D., Ferrocino I., Nasi A., Ndagijimana M., Vernocchi P., La Storia A., et al. (2011). Monitoring of microbial metabolites and bacterial diversity in beef stored under different packaging conditions. Appl. Environ. Microbiol. 77 7372–7381. 10.1128/AEM.05521-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fadda S., Lopez C., Vignolo G. (2010). Role of lactic acid bacteria during meat conditioning and fermentation: peptides generated as sensorial and hygienic biomarkers. Meat Sci. 86 66–79. 10.1016/j.meatsci.2010.04.023 [DOI] [PubMed] [Google Scholar]
- Fall P. A., Pilet M. F., Leduc F., Cardinal M., Duflos G., Guérin C., et al. (2012). Sensory and physicochemical evolution of tropical cooked peeled shrimp inoculated by Brochothrix thermosphacta and Lactococcus piscium CNCM I-4031 during storage at 8°C. Int. J. Food Microbiol. 152 82–90. 10.1016/j.ijfoodmicro.2011.07.015 [DOI] [PubMed] [Google Scholar]
- Fougy L., Desmonts M.-H., Coeuret G., Fassel C., Hamon E., Hézard B., et al. (2016). Reducing salt in raw pork sausage increases spoilage and correlated with reduced bacterial diversity. Appl. Environ. Microbiol. 82 3928–3939. 10.1128/AEM.00323-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuertez-Perez S., Hauschild P., Hilgarth M., Vogel R. F. (2019). Biodiversity of Photobacterium spp. isolated from meats. Front. Microbiol. 10:2399. 10.3389/fmicb.2019.02399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galimberti A., Bruno A., Mezzasalma V., De Mattia F., Bruni I., Labra M. (2015). Emerging DNA-based technologies to characterize food ecosystems. Food Res. Int. 69 424–433. 10.1016/j.foodres.2015.01.017 [DOI] [Google Scholar]
- Garnier L., Valence F., Pawtowski A., Auhustsinava-galerne L., Frotté N., Baroncelli R., et al. (2017). Diversity of spoilage fungi associated with various French dairy products. Int. J. Food Microbiol. 241 191–197. 10.1016/j.ijfoodmicro.2016.10.026 [DOI] [PubMed] [Google Scholar]
- Garofalo C., Bancalari E., Milanovic V., Cardinali F., Osimani A., Sardaro M. L. S., et al. (2017). Study of the bacterial diversity of foods: PCR-DGGE versus LH-PCR. Int. J. Food Microbiol. 242 24–36. 10.1016/j.ijfoodmicro.2016.11.008 [DOI] [PubMed] [Google Scholar]
- Geeraerts W., Pothakos V., De Vuyst L., Leroy F. (2017). Diversity of the dominant bacterial species on sliced cooked pork products at expiration date in the Belgian retail. Food Microbiol. 65 236–243. 10.1016/j.fm.2017.03.007 [DOI] [PubMed] [Google Scholar]
- Ghasemi-Varnamkhasti M., Apetrei C., Lozano J., Anyogu A. (2018). Potential use of electronic noses, electronic tongues and biosensors as multisensory systems for spoilage examination in foods. Trends Food Sci. Technol. 80 71–92. 10.1016/j.tifs.2018.07.018 [DOI] [Google Scholar]
- Greppi A., Ferrocino I., La Storia A., Rantsiou K., Ercolini D., Cocolin L. (2015). Monitoring of the microbiota of fermented sausages by culture independent rRNA-based approaches. Int. J. Food Microbiol. 212 67–75. 10.1016/j.ijfoodmicro.2015.01.016 [DOI] [PubMed] [Google Scholar]
- Gu G., Ottesen A., Bolten S., Ramachandran P., Reed E., Rideout S., et al. (2018). Shifts in spinach microbial communities after chlorine washing and storage at compliant and abusive temperatures. Food Microbiol. 73 73–84. 10.1016/j.fm.2018.01.002 [DOI] [PubMed] [Google Scholar]
- Guillard V., Couvert O., Stahl V., Hanin A., Denis C., Huchet V., et al. (2016). Validation of a predictive model coupling gas transfer and microbial growth in fresh food packed under modified atmosphere. Food Microbiol. 58 43–55. 10.1016/j.fm.2016.03.011 [DOI] [PubMed] [Google Scholar]
- Hahne J., Isele D., Berning J., Lipski A. (2019). The contribution of fast growing, psychrotrophic microorganisms on biodiversity of refrigerated raw cow’s milk with high bacterial counts and their food spoilage potential. Food Microbiol. 79 11–19. 10.1016/j.fm.2018.10.019 [DOI] [PubMed] [Google Scholar]
- Holm E. S., Schäfer A., Koch A. G., Petersen M. A. (2013). Investigation of spoilage in saveloy samples inoculated with four potential spoilage bacteria. Meat Sci. 93 687–695. 10.1016/j.meatsci.2012.11.016 [DOI] [PubMed] [Google Scholar]
- Jastrzębska A., Kowalska S., Szłyk E. (2016). Studies of levels of biogenic amines in meat samples in relation to the content of additives. Food Addit. Contam. Part A hem. Anal. Control Expo Risk Assess 33 27–40. 10.1080/19440049.2015.1111525 [DOI] [PubMed] [Google Scholar]
- Kariyawasam K. M. G. M. M., Jeewanthi R. K. C., Lee N.-K., Paik H.-D. (2019). Characterization of cottage cheese unsing Weissella cibaria D30: physicochemical, antioxidant, and antilisterial properties. J. Dairy Sci. 102 3887–3893. 10.3168/jds.2018-15360 [DOI] [PubMed] [Google Scholar]
- Kato Y., Sakala R. M., Hayashidani H., Kiuchi A., Kaneuchi C., Ogawa M. (2000). Lactobacillus algidus sp. nov., a psychrophilic lactic acid bacterium isolated from vacuum-packaged refrigerated beef. Int. J. Syst. Envol. Microbiol. 50 1143–1149. 10.1099/00207713-50-3-1143 [DOI] [PubMed] [Google Scholar]
- Kaur M., Shang H., Tamplin M., Ross T., Bowman J. P. (2017). Culture-dependent and culture-independent assessment of spoilage community growth on VP lamb meat from packaging to past end of shelf-life. Food Microbiol. 68 71–80. 10.1016/j.fm.2017.06.015 [DOI] [PubMed] [Google Scholar]
- Kembel S. W., Wu M., Eisen J. A., Green J. L. (2012). Incorporating 16S gene copy number information improves estimates of microbial diversity and abundance. PLOS Comput. Biol. 8 1–11. 10.1371/journal.pcbi.1002743.g001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim E., Cho Y., Lee Y., Han S.-K., Kim C.-G., Choo D.-W., et al. (2017). A proteomic approach for rapid identification of Weissella species isolated from Korean fermented foods on MALDI-TOF MS supplemented with an in-house database. Int. J. Food Microbiol. 243 9–15. 10.1016/j.ijfoodmicro.2016.11.027 [DOI] [PubMed] [Google Scholar]
- Koort J., Murros A., Coenye T., Eerola S., Vandamme P., Sukura A., et al. (2005). Lactobacillus oligofermentans sp. nov., associated with spoilage of modified-atmosphere-packaged poultry products. Appl. Environ. Microbiol. 71 4400–4406. 10.1128/AEM.71.8.4400-4406.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korsak N., Taminiau B., Hupperts C., Delhalle L., Nezer C., Delcenserie V., et al. (2017). Assessment of bacterial superficial contamination in classical or ritually slaughtered cattle using metagenetics and microbial analysis. Int. J. Food Microbiol. 247 79–86. 10.1016/j.ijfoodmicro.2016.10.013 [DOI] [PubMed] [Google Scholar]
- Koutsoumanis K., Stamatiou A., Skandamis P., Nychas G.-J. E. (2006). Development of a microbial model for the combined effect of temperature and pH on spoilage of ground meat, and validation of the model under dynamic temperature conditions. Appl. Environ. Microbiol. 72 124–134. 10.1128/AEM.72.1.124-134.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koutsoumanis K. P., Stamatiou A. P., Drosinos E. H., Nychas G.-J. E. (2008). Control of spoilage microorganisms in minced pork by a self-developed modified atmosphere induced by the respiratory activity of meat microflora. Food Microbiol. 25 915–921. 10.1016/j.fm.2008.05.006 [DOI] [PubMed] [Google Scholar]
- Lee M., Song J. H., Jung M. Y., Lee S. H., Chang J. Y. (2017). Large-scale targeted metagenomics analysis of bacterial ecological changes in 88 kimchi samples during fermentation. Food Microbiol. 66 173–183. 10.1016/j.fm.2017.05.002 [DOI] [PubMed] [Google Scholar]
- Li M., Tian L., Zhao G., Zhang Q., Gao X., Huang X., et al. (2014). Formation of biogenic amines and growth of spoilage-related microorganisms in pork stored under different packaging conditions applying PCA. Meat Sci. 96 843–848. 10.1016/j.meatsci.2013.09.023 [DOI] [PubMed] [Google Scholar]
- Li N., Zhang Y., Wu Q., Gu Q., Chen M., Zhang Y., et al. (2019). High-throughput sequencing analysis of bacterial community composition and quality characteristics in refrigerated pork during storage. Food Microbiol. 83 86–94. 10.1016/j.fm.2019.04.013 [DOI] [PubMed] [Google Scholar]
- Li Q., Zhang L., Luo Y. (2018). Changes in microbial communities and quality attributes of white muscle and dark muscle from common carp (Cyprinus carpio) during chilled and freeze-chilled storage. Food Microbiol. 73 237–244. 10.1016/j.fm.2018.01.011 [DOI] [PubMed] [Google Scholar]
- Liu F., Yang R.-Q., Li Y.-F. (2006). Correlations between growth parameters of spoilage micro-organisms and shelf-life of pork stored under air and modified atmosphere at -2, 4 and 10°C. Food Microbiol. 23 578–583. 10.1016/j.fm.2005.10.002 [DOI] [PubMed] [Google Scholar]
- Liu X., Ji L., Wang X., Li J., Shu J., Sun A. (2018). Role of RpoS in stress resistance, quorum sensing and spoilage potential of Pseudomonas fluorescens. Int. J. Food Microbiol. 270 31–38. 10.1016/j.ijfoodmicro.2018.02.011 [DOI] [PubMed] [Google Scholar]
- Liu Z., Li J., Wei B., Huang T., Xiao Y., Peng Z., et al. (2019). Bacterial community and composition in Jiang-shui and Suan-cai revealed by high-throughput sequencing of 16S rRNA. Int J Food Microbiol. 306:108271. 10.1016/j.ijfoodmicro.2019.108271 [DOI] [PubMed] [Google Scholar]
- Louca S., Doebeli M., Parfrey L. W. (2018). Correcting for 16S rRNA gene copy numbers in microbiome surveys remains an unsolved problem. Microbiome 6 1–12. 10.1186/s40168-018-0420-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann E., Wetzels S. U., Pinior B., Metzler-Zebeli B. U., Wagner M., Schmitz-Esser S. (2016). Psychrophile spoilers dominate the bacterial microbiome in musculature samples of slaughter pigs. Meat Sci. 117 36–40. 10.1016/j.meatsci.2016.02.034 [DOI] [PubMed] [Google Scholar]
- Mansur A. R., Song E.-J., Cho Y.-S., Nam Y.-D., Choi Y.-S., Kim D.-O., et al. (2019). Comparative evaluation of spoilage-related bacterial diversity and metabolite profiles in chilled beef stored under air and vacuum packaging. Food Microbiol. 77 166–172. 10.1016/j.fm.2018.09.006 [DOI] [PubMed] [Google Scholar]
- Martins W. F., Longhi D. A., Costa Menezes N. M., da Silva Camargo A. P., Borges Laurindo J., Falcao, et al. (2016). Predicting growth of Weissella viridescens in culture medium under dynamic temperature conditions. Procedia Food Sci. 7 37–40. 10.1016/j.profoo.2016.02.082 [DOI] [Google Scholar]
- Moretro T., Moen B., Heir E., Hansen A. A., Langsrud S. (2016). Contamination of salmon fillets and processing plants with spoilage bacteria. Int. J. Food Microbiol. 237 98–108. 10.1016/j.ijfoodmicro.2016.08.016 [DOI] [PubMed] [Google Scholar]
- Nalbantoglu U., Cakar A., Dogan H., Abaci N., Ustek D., Sayood K., et al. (2014). Metagenomic analysis of the microbial community in kefir grains. Food Microbiol. 41 42–51. 10.1016/j.fm.2014.01.014 [DOI] [PubMed] [Google Scholar]
- Nieminen T. T., Dalgaard P., Björkroth J. (2016). Volatile organic compounds and Photobacterium phosphoreum associated with spoilage of modified-atmosphere-packaged raw pork. Int. J. Food Microbiol. 218 86–95. 10.1016/j.ijfoodmicro.2015.11.003 [DOI] [PubMed] [Google Scholar]
- Nieminen T. T., Koskinen K., Laine P., Hultman J., Säde E., Paulin L., et al. (2012). Comparison of microbial communities in marinated and unmarinated broiler meat by metagenomics. Int. J. Food Microbiol. 157 142–149. 10.1016/j.ijfoodmicro.2012.04.016 [DOI] [PubMed] [Google Scholar]
- Nieminen T. T., Nummela M., Björkroth J. (2015). Packaging gas selects lactic acid bacterial communities on raw pork. J. Appl. Microbiol. 119 1310–1316. 10.1111/jam.12890 [DOI] [PubMed] [Google Scholar]
- Nychas G. J. E., Skandamis P. N., Tassou C. C., Koutsoumanis K. P. (2008). Meat spoilage during distribution. Meat Sci. 78 77–89. 10.1016/j.meatsci.2007.06.020 [DOI] [PubMed] [Google Scholar]
- Odeyemi O. A., Burke C. M., Bolch C. C. J., Stanley R. (2018). Spoilage microbial community profiling by 16S rRNA amplicon sequencing of modified atmospohere packaged live mussels stored ar 4°C. Food Res. Int. 121 568–576. 10.1016/j.foodres.2018.12.017 [DOI] [PubMed] [Google Scholar]
- Papageorgiou M., Lambropoulou D., Morrison C., Klodzinska E., Namiesnik J., Plotka-Wasylka J. (2018). Literature update of analytical methods for biogenic amines determination in food and beverages. Trends Analyt. Chem. 98 128–142. 10.1016/j.trac.2017.11.001 [DOI] [Google Scholar]
- Parente E., Cocolin L., De Filippis F., Zotta T., Ferrocino I., O’Sullivan O., et al. (2016). FoodMicrobionet: a database for the visualization and exploration of food bacterial communities based on network analysis. Int. J. Food Microbiol. 219 28–37. 10.1016/j.ijfoodmicro.2015.12.001 [DOI] [PubMed] [Google Scholar]
- Parks D. H., Tyson G. W., Hugenholtz P., Beiko R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinform. 30 3123–3124. 10.1093/bioinformatics/btu494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parlapani F. F., Michailidou S., Pasentsis K., Argiriou A., Krey G., Boziaris I. S. (2018). A meta-barcoding approach to assess and compare the storage temperature-dependent bacterial diversity of gilt-head sea bream (Sparus aurata) originating from fish farms from two geographically distinct areas of Greece. Int. J. Food Microbiol. 278 36–43. 10.1016/j.ijfoodmicro.2018.04.027 [DOI] [PubMed] [Google Scholar]
- Pennacchia C., Ercolini D., Villani F. (2009). Development of a real-time PCA assay for the specific detection of Brochothrix thermosphacta in fresh and spoiled raw meat. Int. J. Food Microbiol. 134 230–236. 10.1016/j.ijfoodmicro.2009.07.005 [DOI] [PubMed] [Google Scholar]
- Pennacchia C., Ercolini D., Villani F. (2011). Spoilage-related microbiota associated with chilled beef stored in air or vacuum pack. Food Microbiol. 28 84–93. 10.1016/j.fm.2010.08.010 [DOI] [PubMed] [Google Scholar]
- Peruzy M. F., Murru N., Yu Z., Cnockaert M., Joossens M., Proroga Y. T. R., et al. (2019). Determination of the microbiological contamination in minced pork by culture dependent and 16S amplicon sequencing analysis. Int. J. Food Microbiol. 290 27–35. 10.1016/j.ijfoodmicro.2018.09.025 [DOI] [PubMed] [Google Scholar]
- Pinter M. D., Harter T., McCarthy M. J., Augustine M. P. (2014). Toward using NMR to screen for spoiled tomatoes stored in 1,000 L, aseptically sealed, metal-lined totes. Sensors 14 4167–4176. 10.3390/s140304167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinu F. R. (2016). Early detection of food pathogens and food spoilage microorganisms: application of metabolomics. Trends Food Sci. Technol. 54 213–215. 10.1016/j.tifs.2016.05.018 [DOI] [Google Scholar]
- Polka J., Rebecchi A., Pisacana V., Morelli L., Puglisi E. (2015). Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 46 342–356. 10.1016/j.fm.2014.08.023 [DOI] [PubMed] [Google Scholar]
- Porcellato D., Aspholm M., Skeie S. B., Monshaugen M., Brendehaug J., Mellegard H. (2018). Microbial diversity of consumption milk during processing and storage. Int. J. Food Microbiol. 266 21–30. 10.1016/j.ijfoodmicro.2017.11.004 [DOI] [PubMed] [Google Scholar]
- Pothakos V., Devlieghere F., Villani F., Björkroth J., Ercolini D. (2015). Lactic acid bacteria and their controversial role in fresh meat spoilage. Meat Sci. 109 66–74. 10.1016/j.meatsci.2015.04.014 [DOI] [PubMed] [Google Scholar]
- Pothakos V., Taminiau B., Huys G., Nezer C., Daube G., Devlieghere F. (2014). Psychrotrophic lactic acid bacteria associated with production batch recalls and sporadic cases of early spoilage in Belgium between 2010 and 2014. Int. J. Food Microbiol. 191 157–163. 10.1016/j.ijfoodmicro.2014.09.013 [DOI] [PubMed] [Google Scholar]
- Pruesse E., Peplies J., Glïckner F. O. (2012). SINA: accurate high-throughput multiple sequence aligment of ribosomal RNA genes. Bioinformatics 28 1823–1829. 10.1093/bioinformatics/bts252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: https://www.R-project.org/ [Google Scholar]
- Rahkila R., Nieminen T., Johansson P., Säde E., Björkroth J. (2012). Characterization and evaluation of the spoilage potential of Lactococcus piscium isolates from modified atmosphere packaged meat. Int. J. Food Microbiol. 156 50–59. 10.1016/j.ijfoodmicro.2012.02.022 [DOI] [PubMed] [Google Scholar]
- Raimondi S., Nappi M. R., Sirangelo T. M., Leonardi A., Amaretti A., Ulrici A., et al. (2018). Bacterial community of industrial raw sausage packaged in modified atmosphere throughout the shelf life. Int. J. Food Microbiol. 280 78–86. 10.1016/j.ijfoodmicro.2018.04.041 [DOI] [PubMed] [Google Scholar]
- Reid R., Fanning S., Whyte P., Kerry J., Lindqvist R., Yu Z., et al. (2017). The microbiology of beef carcasses and primals during chilling and commercial storage. Food Microbiol. 61 50–57. 10.1016/j.fm.2016.08.003 [DOI] [PubMed] [Google Scholar]
- Riquelme C., Câmara S., Enes Dapkevicius M., de L. N., Vinuesa P., da Silva C. C. G., et al. (2015). Characterization of the bacterial biodiversity in Pico cheese (an artisanal Azorean food). Int. J. Food Microbiol. 192 86–94. 10.1016/j.ijfoodmicro.2014.09.031 [DOI] [PubMed] [Google Scholar]
- Robertson C. E., Harris J. K., Wagner B. D., Granger D., Browne K., Tatem B., et al. (2013). Explicet: graphical used interface software for metadata-driven management, analysis and visualization of microbiome data. Bioinformatics 29 3100–3101. 10.1093/bioinformatics/btt526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rotabakk B. T., Birkeland S., Jeksrud W. K., Sivertsvik M. (2006). Effect of modified atmosphere packaging and soluble gas stabilization on the shelf life of skinless chicken breast fillets. J. Food Sci. 71 124–131. 10.1111/j.1365-2621.2006.tb08915.x [DOI] [Google Scholar]
- Rouger A., Moriceau N., Prévost H., Remenant B., Zagorec M. (2018). Diversity of bacterial communities in French chicken cuts stored under modified atmosphere packaging. Food Microbiol. 70 7–16. 10.1016/j.fm.2017.08.013 [DOI] [PubMed] [Google Scholar]
- Rouger A., Remenant B., Prévost H., Zagorec M. (2017). A method to isolate bacterial communities and characterize ecosystems from food products: validation and utilization in as a reproductible chicken meat model. Int. J. Food Microbiol. 247 38–47. 10.1016/j.ijfoodmicro.2016.04.028 [DOI] [PubMed] [Google Scholar]
- Saraiva C., Fontes M. C., Patarata L., Martins C., Cadavez V., Gonzalas-Barron U. (2016). Modelling the kinetics of Listeria monocytogenes in refrigerated fresh beef under different packaging atmospheres. Food Sci. Technol. 66 664–671. 10.1016/j.lwt.2015.11.026 [DOI] [Google Scholar]
- Schloss P. D., Westcott S. L., Ryabin T., Hall J. R., Hartmann M., Hollister E. B., et al. (2009). Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75 7537–7541. 10.1128/AEM.01541-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silbande A., Adenet S., Chopin C., Cornet J., Smith-Ravin J., Rochefort K., et al. (2018). Effect of vacuum and modified atmosphere packaging on the microbiological, chemical and sensory properties of tropical red drum (Sciaenops acellatus) fillets stored at 4°C. Int. J. Food Microbiol. 266 31–41. 10.1016/j.ijfoodmicro.2017.10.015 [DOI] [PubMed] [Google Scholar]
- Silbande A., Adenet S., Smith-Ravin J., Joffraud J.-J., Rochefort K., Leroi F. (2016). Quality assessment of ice-stored tropical yellowfin tuna (Thunnu albacares) and influence of vacuum and modified atmosphere packaging. Food Microbiol. 60 62–72. 10.1016/j.fm.2016.06.016 [DOI] [PubMed] [Google Scholar]
- Simpson R., Carevic E. (2004). Designing a modified atmosphere packaging system for foodservice portions on nonrespiring foods: optimal gas mixture and food/headspace ratio. Foodserv. Res. Int. 14 257–272. 10.1111/j.1745-4506.2004.tb00194.x [DOI] [Google Scholar]
- Singh V. P. (2018). Recent approaches in food bio-preservation – a review. Open Vet. J. 8 104–111. 10.4314/ovj.v8i1.16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spanu C., Piras F., Mocci A. M., Nieddu G., De Santis E. P. L., Scarano C. (2018). Use of Carnobacterium spp. protective culture in MAP packed Ricotta fresca cheese to control Pseudomonas spp. Food Microbiol. 74 50–56. 10.1016/j.fm.2018.02.020 [DOI] [PubMed] [Google Scholar]
- Stanborough T., Fegan N., Powell S. M., Tamplin M., Chandry P. S. (2017). Insight into the genome of Brochothrix thermosphacta, a problematic meat spoilage bacterium. Appl. Environ. Microbiol. 83 1–20. 10.1128/AEM.02786-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanovic E., Fitzgerald G., McAulliffe O. (2017). Advances in the genomics and metabolomics of dairy lactobacilli: a review. Food Microbiol. 61 33–49. 10.1016/j.fm.2016.08.009 [DOI] [PubMed] [Google Scholar]
- Stellato G., La Storia A., De Filippis F., Borriello G., Villani F., Ercoloni D. (2016). Overlap of spoilage-associated microbiota between meat and the meat processing environment in small-scale and large-scale retail distributions. Appl. Environ. Microbiol. 82 4045–4054. 10.1128/AEM.00793-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoddard S. F., Smith B. J., Hein R., Roller B. R. K., Schmidt T. M. (2015). rrnDB: improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic Acids Res. 43 593–598. 10.1093/nar/gku1201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoops J., Ruyters S., Busschaert P., Spaepen R., Verreth C., Claes J., et al. (2015). Bacterial community dynamics during colt storage of minced meat packaged under modified atmosphere and supplemented with different preservatives. Food Microbiol. 48 192–199. 10.1016/j.fm.2014.12.012 [DOI] [PubMed] [Google Scholar]
- Vester Lauritsen C., Kjeldgaard J., Ingmer H., Bisgaard M., Christensen H. (2019). Microbiota emcompassing putative spoilage bacteria in retail packaged broiler meat and commercial broiler abattoir. Int. J. Food Microbiol. 300 14–21. 10.1016/j.ijfoodmicro.2019.04.003 [DOI] [PubMed] [Google Scholar]
- Woraprayote W., Malila Y., Sorapukdee S., Swetwiwathana A., Benjakul S., Visessanguan W. (2016). Bacteriocins from lactic acid bacteria and their applications in meat and meat products. Meat Sci. 120 118–132. 10.1016/j.meatsci.2016.04.004 [DOI] [PubMed] [Google Scholar]
- Yoon S. H., Ha S. M., Kwon S., Lim J., Kim Y., Seo H., et al. (2017). Introducing EzBioCloud: a taxonomically united database of 16S rRNA and whole genome assemblies. Int. J. Syst. Evol. Microbiol. 67 1613–1617. 10.1099/ijsem.0.001755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao F., Zhou G., Ye K., Wang S., Xu X., Li C. (2015). Microbial changes in vacuum-packed chilled pork during storage. Meat Sci. 100 145–149. 10.1016/j.meatsci.2014.10.004 [DOI] [PubMed] [Google Scholar]
- Zotta T., Parente E., Ianniello R. G., De Filippis F., Ricciardi A. (2019). Dynamics of bacterial communities and interaction networks in thawed fish fillets during chilled storage in air. Int. J. Food Microbiol. 293 102–113. 10.1016/j.ijfoodmicro.2019.01.008 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All biosample raw reads were deposited at the National Center for Biotechnology Information (NCBI) and are available under the BioProject ID PRJNA551357.