

Highlights
* Self-development incorporates both personal and social/group identity. * Self-development involves both cognitive and socioemotional processes. * An extended network of brain regions supports self-development, broadly conceived. * Autism, depression associated with hypoactivation, hyperactivation of the network. * Integrating cognitive, socioemotional, neuroimaging approaches will advance the field.
Keywords: Self-development, fMRI, Medial prefrontal cortex, Perspective-taking, Social exclusion, Identity
Abstract
This review integrates cognitive, socioemotional, and neuroimaging perspectives on self-development. Neural correlates of key processes implicated in personal and social identity are reported from studies of children, adolescents, and adults, including autobiographical memory, direct and reflected self-appraisals, and social exclusion. While cortical midline structures of medial prefrontal cortex and medial posterior parietal cortex are consistently identified in neuroimaging studies considering personal identity from a primarily cognitive perspective (“who am I?”), additional regions are implicated by studies considering personal and social identity from a more socioemotional perspective (“what do others think about me, where do I fit in?”), especially in child or adolescent samples. The involvement of these additional regions (including tempo–parietal junction and posterior superior temporal sulcus, temporal poles, anterior insula, ventral striatum, anterior cingulate cortex, middle cingulate cortex, and ventrolateral prefrontal cortex) suggests mentalizing, emotion, and emotion regulation are central to self-development. In addition, these regions appear to function atypically during personal and social identity tasks in autism and depression, exhibiting a broad pattern of hypoactivation and hyperactivation, respectively.
Self-development results in an intricate and somewhat paradoxical product. We grow to see ourselves as unique, coherent, and bounded entities with personal preferences, abilities, and characteristics (Harter, 1999), but we also acquire a sense of being intimately connected with others in relationships and groups (Ruble et al., 2004). Most research attempting to understand this complex phenomenon has been philosophical or behavioral in nature, but for over a decade and a half, neuroimaging technologies have been employed in attempts to uncover how the brain supports the endpoint of this prolonged path to a rich and multifaceted self, and in the last several years developmental populations have been folded into the scope of these inquiries (Pfeifer et al., 2007, Sebastian et al., 2008). In this manuscript, we review evidence identifying the neural correlates of the self and its development. As an organizing principle, we argue that the most successful and complete approach to understanding how the brain supports self-development will integrate cognitive, socioemotional, and neuroimaging perspectives.
The self in psychological science is an umbrella term encompassing such diverse constructs as self-awareness, agency, self-recognition, self-esteem, self-concept, the social self and self-enhancement (Leary and Tangney, 2003, Sedikides and Spencer, 2007). Likewise, evolutionary approaches such as that of Robins and colleagues (Robins et al., 2008) have identified several adaptive functions of the self: self-regulation, identity formation, and its utility as an information-processing filter and basis for understanding others. In the current paper we take an ontogenetic perspective on just one of these many facets of the self – namely that of identity and its formation. One classic developmental approach (Erikson, 1968) divides identity into personal aspects that mark the self as a distinctive individual, and social aspects that highlight group affiliations, roles, and relationships (as well as into the ego which is aware of the self and its continuity over time). This approach includes the familiar depiction of the adolescent conflict over fidelity (identity vs. role confusion), which highlights three predominant themes of self-development emphasized in this manuscript: who am I, what do others think about me, and where do I fit in?
Much of the answer to “who am I?” is embodied in the self-concept, which contains evaluative self-knowledge that is organized by domains (social, academic, athletic, family, etc.) and contributes to overall global self-worth (Harter, 1999, Marsh, 1990). In adults, the self is considered an exceptional structure for organizing information; it conveys mnemonic advantages (the self-reference effect; Rogers et al., 1977, Symons and Johnson, 1997), and facilitates behavioral responses owing to implicit associations between attributes and the self (Greenwald and Banaji, 1989, Markus, 1977). A large body of research, primarily in the field of social psychology, therefore takes this and other related cognitive perspectives on the self's functions (Greenwald, 1981, Kihlstrom et al., 2002, Kihlstrom and Klein, 1994, Klein, 2001, Klein et al., 1996).
Various markers of self-referential cognition that contribute to personal identity development undergo considerable change during childhood and adolescence; this is an extended period of time during which the changes are many and varied. In toddlers, mirror self-recognition and personal pronoun use provide early indicators of self-development (Lewis and Carmody, 2008). Self-appraisals featuring an evaluative component emerge in early childhood, but these are both more concrete and more positive than self-appraisals of children in the later elementary school years (Damon and Hart, 1988, Harter, 2006). The self-reference effect first emerges around the transition to middle childhood (5–6 years of age) and then increases to adult levels by about age 10 (Ray et al., 2009). In parallel, during this same timeframe children move from using global information to predict others’ behaviors to using traits to make more accurate inferences, suggesting that younger children do not understand the association between psychological traits and consistent patterns of behavior in others or themselves (Alvarez et al., 2001). Finally, evidence suggests children begin using social comparisons to inform lasting self-evaluations by approximately 7–9 years of age (Harter, 2006, Ruble and Frey, 1991, Stipek and MacIver, 1989), which leads to an overall decline in the positivity of self-evaluations, although in some circumstances even preschool children use social comparisons, especially to connect or contrast the self with others (Butler, 1998, Rhodes and Brickman, 2008).
Just after these important cognitive skills are acquired in late childhood, the social landscape begins to undergo massive changes in adolescence, particularly relating to the questions “what do others think about me” and “where do I fit in?” Another classic developmental approach (Elkind, 1967, Elkind, 1985) suggests that cognitive advances related to perspective-taking abilities are associated with attributes people consider to be quintessentially adolescent, like self-consciousness or belief in one's own invulnerability and individuality. Although this perspective has been critiqued empirically and theoretically (Vartanian, 2000), it remains likely that the development of these perspective-taking skills probably increases the ability, if not also the tendency, to reason about others’ opinions of the self (Pfeifer et al., 2009). In addition, the relative importance of different perspectives is shifting during this stage, dictating which “others” are relevant. Adolescents must balance conflicting needs to individuate from their parents and to feel connected (Casey et al., 2010, Steinberg and Silverberg, 1986), fueling the necessity of developing intimate, supportive friendships and identifying with social groups to which they belong (Rubin et al., 2008), including division into cliques and crowds that differ according to behaviors and values (Brown, 2004). Peers therefore become much more important (particularly in the social domain, while parents retain some influence in academics; Bouchey and Harter, 2005, Pfeifer et al., 2009, Steinberg and Silverberg, 1986), as indicated by significant increases in time spent together without adult supervision (Brown, 2004, Csikszentmihalyi and Larson, 1984); acceptance by peers is critical, and rejection is costly (Coie, 1990).
Importantly, personal and social identity development can be seen as mutually informative. For example, both recent theorists and pioneers in psychology (Baldwin, 1895, James, 1890, Harter, 1999, Mead, 1934) persuasively argue that the evaluative self-knowledge supporting personal identity should be recognized as both as a cognitive and social construction. The consequences of this dual nature are that the self is significantly affected by the cognitive capabilities of a given individual, as well as by family members, peers, and other close relationship partners (although these social inputs are likely still mediated by cognitive processes). Meanwhile, developing a sense of being accepted within relevant peer groups can be viewed as deriving in part from a growing understanding of one's own abilities, values, and preferences, which either complement or conflict with possible social identities. For example, being praised by a coach for game-winning performances but chided by a parent for failures on exams may be associated with representing the self as athletic but not smart, and identifying as part of the “jocks” instead of the “brains” in high school. In other words, what I believe others think about me can affect who I think I am, which can influence where I believe I fit in. Finally, the driving forces behind these cognitive and social developments are assumed to be not only each other, but also neural changes. Specifically, a more developed brain (across a variety of indices, including structure, function, and connectivity) cultivates more advanced cognitive capacities (e.g., Paus, 2005). At the same time, the onset of puberty and adolescence marks a turning point in relationships with peers and increased responsiveness in socioemotional contexts (Blakemore et al., 2010, Casey et al., 2010, Nelson et al., 2005, Pfeifer et al., 2011, Spear, 2004, Steinberg, 2008). Indeed, developmental psychologists recognize the combined importance of cognitive, socioemotional, and physical changes that contribute to all aspects of maturation (Dahl, 2004), and the case of self-development is no exception.
In this review, we hope to demonstrate the value of a developmental social cognitive neuroscience approach to the self, as depicted in Fig. 1: both cognitive and socioemotional processes influence self-development (as well as bidirectionally influencing each other), and both are supported by (and likely impact) brain structure, connectivity, and function. First, neuroimaging studies taking a cognitive or a social perspective on the self will be reviewed, in that order. We emphasize the socioemotional perspective – which includes cultural, affective, and motivational effects – because it may be particularly important during self-development. Next, we describe the corresponding neuroimaging findings in children and adolescents, as well as summarize patterns of structural development in the brain regions identified as relevant. To conclude, we propose that by integrating cognitive, socioemotional, and neuroimaging perspectives it is possible to achieve real advances in understanding both typical and atypical self-development, and we briefly discuss how this approach may be relevant to the study of personal and social identity in various disorders, illustrated by the cases of autism and depression.
Fig. 1.

Model of cognitive, socioemotional, and neural influences on self-development. In this model, three factors interact to influence self-development: cognition, socioemotional functioning, and brain development. Bidirectional influences are noted between each of these factors, and there is noted change over time in both neural and psychological components of the model.
1. Cognitive perspectives
Many aspects of cognition related to self-development have been explored in adults using neuroimaging techniques, such as agency (Powell et al., 2010) and self-recognition (Kaplan et al., 2008). However, in this section we focus on neuroimaging evidence regarding two key phenomena that contribute to development of personal identity from a cognitive viewpoint: autobiographical memory and direct self-appraisals (specifically, retrieving or evaluating personal attributes or preferences). Autobiographical memories differ from direct self-appraisals because the former are event-specific, whereas the latter are generalized across many instances.
Autobiographical memories are intertwined with personal identity in many ways (Fivush, 2010). They comprise a knowledge base of general events and event-specific knowledge spanning multiple lifetime periods, which index different parts of this knowledge base, and are typically very rich in imagery and sensory–perceptual detail (Conway and Pleydell-Pearce, 2000). In adults, autobiographical memory can be distinguished from non-self-referential memory by activation in both medial posterior parietal cortex (mPPC; includes precuneus, posterior cingulate, and retrosplenial cortex) and medial prefrontal cortex (mPFC; Svoboda et al., 2006, Summerfield et al., 2009), two regions collectively referred to as cortical midline structures (CMS; Northoff and Bermpohl, 2004).
In a sense, autobiographical memory can be seen as a cornerstone of personal identity development – a vast source of information about the self that is compiled and integrated to produce self-evaluations. Neuroimaging studies of direct self-appraisals target these byproducts, typically asking participants to reflect on characteristics of the self, as well as varied familiar and personally known others, and report whether evaluative traits are descriptive of a given target (self or other; for examples, see Johnson et al., 2002, Kelley et al., 2002). There has been much evidence amassed to indicate that in adults, self-appraisals rely on CMS, similar to those found in autobiographical memory: mPFC (spanning its ventral, anterior rostral, and dorsal aspects) and adjacent rostral anterior cingulate cortex (ACC), as well as mPPC (Mitchell, 2009, Northoff and Bermpohl, 2004, Northoff et al., 2006, Northoff et al., 2010).
2. Cortical midline structures
The above summary suggests CMS are prime candidates for supporting cognitive processes associated with self-development. However, an ongoing challenge in the field has been to clearly specify the functional subdivisions of mPFC and mPPC with respect to their roles in self-appraisals and autobiographical memory. These regions are extensive: for example, mPFC is composed of the anterior cingulate gyrus and medial aspects of superior and middle frontal gyri, which extends over multiple putative Brodmann Areas (10/9/8/32/24) associated with distinct cytoarchitectural features. Complicating matters, diverse terminology has been used by the many different research groups investigating mPFC function. Some groups simply divide mPFC into two subregions (dorsal and ventral), usually according to the superior–inferior axis (approximately at z = 0). Others reserve the ventral distinction for cortex located approximately 0.5–1.0 cm lower, and separate mPFC into three subregions (dorsal, anterior rostral, and ventral). Groups also vary as to whether they distinguish between anterior rostral mPFC and adjacent rostral ACC. In contrast, the terminology has been relatively more consistent with respect to mPPC. The specific terminology used for the remainder of this review is depicted in Fig. 2.
Fig. 2.
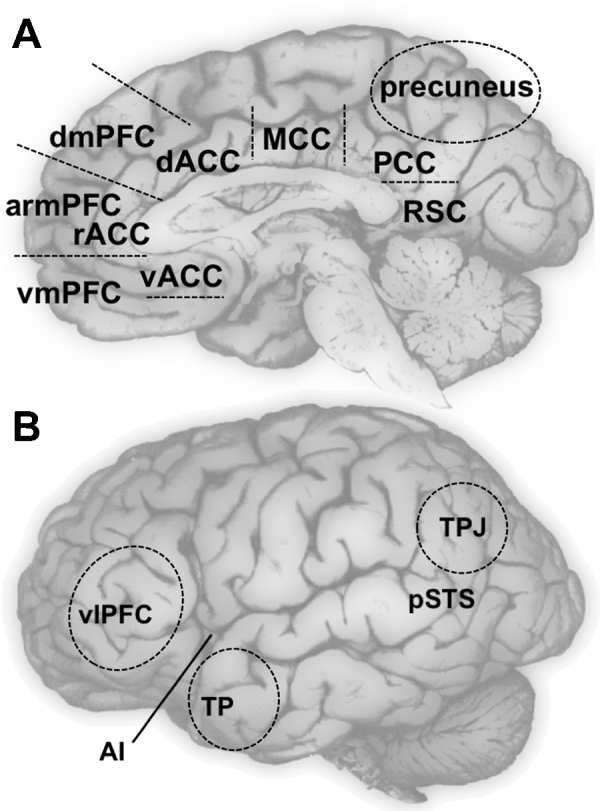
Brain regions of interest for self-development. Panel A displays a medial view of the brain with approximate locations of dorsal, anterior rostral, and ventral medial prefrontal cortex (dmPFC, armPFC, and vmPFC, respectively); dorsal, rostral, and ventral anterior cingulate cortex (dACC, rACC, and vACC, respectively); middle cingulate cortex (MCC); and medial posterior parietal cortex (mPPC) which includes precuneus, posterior cingulate cortex (PCC) and retrosplenial cortex (RSC). Panel B displays a lateral view of the brain with approximate locations of tempo–parietal junction (TPJ), posterior superior temporal sulcus (pSTS), temporal poles (TP), anterior insula (AI, which is underneath the lateral surface), and ventral lateral prefrontal cortex (vlPFC). Ventral striatum (VS), a subcortical structure, is not pictured.
In our opinion, increased specificity will be key to advancing the field, both in terms of spatial location of activations and psychological constructs (for a similar argument, see Johnson et al., 2009, Johnson et al., 2006). Readers should take care to not rely on terminology alone before a consensus has been reached in the field, but rather note reported locations in coordinate space and examine rendered images carefully. The rationale behind this viewpoint is that the adult literature seems to indicate each of the regions in Fig. 2 may carry out different aspects of self-appraisals, autobiographical memory, and other identity-related phenomena.
There is an apparent gradient of specialization in mPFC, such that dorsal aspects are engaged in more cognitive aspects of self-appraisals, while ventral and anterior rostral aspects seem sensitive to affective and motivational components (Moran et al., 2006, Ochsner et al., 2005). Anterior rostral mPFC may be strongly engaged by positive, approach-oriented, self-relevant thought such as reflection on hopes and aspirations (Johnson et al., 2006, Packer and Cunningham, 2009), envisioning positive future events happening to the self (D’Argembeau et al., 2008), and prospection more generally (Addis et al., 2007). Anterior rostral mPFC is also often referred to in the literature as rostral ACC, and for the purposes of this review we consider these regions as a group. Together, they may be involved specifically in first-person perspective-taking (Ames et al., 2008), shifting between or integrating external, exogenous stimulus-oriented attention and internally-generated, endogenous stimulus-independent information (Burgess et al., 2007, Gilbert et al., 2006, Gusnard et al., 2001, Mason et al., 2007), or distinguishing self and other by responding preferentially to the self and close or preferred others (Benoit et al., 2010, Chen et al., 2010, Chiao et al., 2009, Heatherton et al., 2006, Krienen et al., 2010, Northoff et al., 2010, Ray et al., 2009). In contrast, dorsal mPFC activations are associated with decreasing similarity between self and other evaluated targets (Benoit et al., 2010, Jenkins et al., 2008, Mitchell et al., 2005, Mitchell et al., 2006, Ochsner et al., 2005, Tamir and Mitchell, 2010). To sum up, mPFC appears to support not only cognitive but also affective and motivational aspects of self-appraisals, including those that are focused on the present/future as well as those that are abstracted and integrated from individual experiences of the past self.
Subregions in mPPC likewise exhibit varied sensitivity to diverse aspects of self-referential processing, although investigations of mPFC functioning are not yet as abundant as studies of mPFC. In general, research suggests that mPPC may serve as the basis for autonoetic consciousness – that is, recalling past experiences and having the feeling of “remembering” them (Andreasen et al., 1995, Lou et al., 2004, Moran et al., 2009). Imagery appears to be supported by more anterior aspects of the precuneus, while posterior regions of the precuneus subserve episodic memory retrieval (Cavanna and Trimble, 2006). In addition to this anterior/posterior distinction in the precuneus, a dorsal/ventral distinction may be present in the posterior cingulate, namely that ventral posterior cingulate and retrosplenial cortex may be more involved in affective processes than dorsal aspects of mPPC (Maddock, 1999, Vogt and Laureys, 2005). Together, the posterior cingulate and retrosplenial cortex have also been referred to as “evaluative,” that is, assigning significance to events and assessing the relevance of different environmental stimuli (Vogt et al., 1992). Posterior cingulate may be focused more externally than internally, on others in addition to the self, as it is implicated in both perspective-taking (Benoit et al., 2010, Jackson et al., 2006, Pfeifer et al., 2009) and reflecting on personal duties and obligations (Johnson et al., 2006). Retrosplenial cortex appears to be involved in both autobiographical memory and visuo-spatial processing (Svoboda et al., 2006), and may be involved in switching between allocentric and egocentric frames of reference during memory recall, imagery and navigation (Vann et al., 2009). Therefore, mPPC is broadly involved in episodic or autobiographical memory and autonoetic consciousness, but also in maintaining an external focus that extends self-perceptions beyond the individual, and engages perspective-taking. As such, mPPC may not be uniquely involved in self-referential processing (Qin and Northoff, 2011, Qin et al., 2011), and in some studies is observed to be more active when appraisals are made about others than the self, perhaps as a result of the specific retrieval processes that vary across studies (Lombardo et al., 2010a). Future research should attempt to directly manipulate personal familiarity and other memory retrieval conditions that may modulate activity in mPPC, to test this hypothesis more fully and help clarify the precise role of mPPC in self-appraisals.
3. Socioemotional perspectives
The above summary reveals that even when focusing on the self from a cognitive perspective, neuroimaging studies of adult direct self-appraisals and autobiographical memory appear to elicit social, contextual, affective, and motivational processes to some extent. In this section, we now turn to considering three phenomena that demonstrate the relevance of a socioemotional perspective on neural correlates of the self: reflected self-appraisals, cultural influences on self-construals, and social exclusion from peer groups.
The psychological construct of reflected appraisals is expressed well by the question, “what do others think about me?,” which represents perhaps the most straightforward example of socioemotional influences on personal identity development. Indeed, the actual or perceived opinions of others contained in reflected self-appraisals have been hypothesized to inform self-development for decades (Baldwin, 1895, Cooley, 1902, Harter, 1999, Mead, 1934, Pfeifer et al., 2009). Unlike the accumulation of neuroimaging data addressing autobiographical memory and direct self-appraisals, there have been just a few studies exploring the neural bases of reflected self-appraisals. These studies find that reflected self-appraisals engage not only CMS, but also regions involved in perspective-taking and theory of mind, including TPJ and adjacent posterior superior temporal sulcus (pSTS), as well as structures supporting affective processing, such as temporal poles and the insula (D’Argembeau et al., 2007, Ochsner et al., 2005; Pfeifer et al., invited revision under review). These additional regions have not historically been highlighted in studies of adult direct self-appraisals (Northoff et al., 2006, Northoff et al., 2010).
However, regions like TPJ, temporal poles, and the insula have all been receiving increasing attention for their role in self-referential processing (Benoit et al., 2010, Enzi et al., 2009, Legrand and Ruby, 2009, Northoff et al., 2010, van der Meer et al., 2010). Our own work has demonstrated that adults may engage these regions during direct as well as reflected self-appraisals, in certain contexts. In a study of Chinese young adults, the TPJ in particular was implicated alongside CMS in direct self-appraisals – but only in the social, not academic, domain (Pfeifer et al., invited revision under review). Eastern cultures are generally considered to be more interdependent than Western cultures (Markus and Kitayama, 1991), such as in the social domain in China, yet both historical and current empirical evidence suggests that in the academic domain Chinese individuals exhibit very independent self-construals and goals, even exceeding their Western counterparts on average (Li, 2006). Furthermore, to the extent participants believed others’ reflected self-appraisals differed from their own academic self-evaluations, they elicited activity in ventrolateral PFC (vlPFC), a region strongly associated with response inhibition (Aron et al., 2004). These neuroimaging findings suggest that direct self-appraisals may rely on perceived opinions of others more in some contexts than others, such as in relatively more interdependent than independent domains, and that reflected self-appraisals may require one to inhibit personally-held beliefs about the self when the two differ.
Other studies conducting cross-cultural comparisons of direct self-appraisals illustrate additional contextual effects on the neural correlates of the self. For example, although Western (British, American, Canadian, and Australian) and Chinese participants showed similar levels of anterior rostral mPFC activation during self- vs. other-referential processing, Chinese participants showed larger anterior rostral mPFC activation for mother- vs. other-referential processing, equal to that elicited by self-referential processing (Zhu et al., 2007). The authors concluded that Chinese participants employed anterior rostral mPFC for representations of both self and mother, reflecting a more interdependent social structure, whereas Western participants only represent the self in anterior rostral mPFC, reflecting a more independent social structure. Another study found that participants with an individualistic self-construal exhibited greater anterior rostral mPFC activation to general self-descriptions, like being shy in general, while participants with a collectivist self-construal showed greater activation in this region to contextual self-descriptions, like being sociable around one's family (Chiao et al., 2009).
Taken together, this line of neuroimaging research on direct and reflected self-appraisals suggests that not only do specific individuals impact the functioning of neural mechanisms supporting self-evaluations, so does one's cultural milieu (Han and Northoff, 2008, Kitayama and Park, 2010). Furthermore, dynamic cultural influences on behavioral and neural responses during self-appraisals suggest that these representations remain flexible even in adulthood, but static cultural influences are also apparent, and may reflect cultural effects on self-appraisals that may have been acquired early in development (Chiao et al., 2010). These effects may be determined to some degree by desires (implicit or explicit) on the part of an individual to “fit in,” to demonstrate similarity or cohesion within cultural or peer groups and cultivate a sense of belonging.
This leads directly to the final socioemotional perspective we wish to consider, which is tied to social identity via threats to group belonging, when the question “where do I fit in?” is answered with exclusion from a peer group. This approach is particularly likely to be affectively or motivationally relevant during adolescence, as children who are socially rejected face pervasive challenges to adjustment in the short and long-term (Brown and Huang, 1995, Coie, 1990, Coie and Cillessen, 1993, Prinstein and Aikins, 2004, Rubin et al., 2006). Interestingly, however, social exclusion and peer rejection research is rarely connected explicitly to the self in the neurosciences. In adults, social exclusion typically elicits activity in dorsal ACC and anterior insula, which often increases to the extent participants feel distressed about being rejected, report greater rejection sensitivity, or possess histories of rejection (Burklund et al., 2007, Eisenberger and Lieberman, 2004, Eisenberger et al., 2003). In these studies, vlPFC activity is often observed, and inversely correlated with dorsal ACC activity – possibly regulating feelings of distress. Other studies suggest that the dorsal ACC activity is due to the violation of an expectation or implicit social rule of inclusion and fair play, finding instead that ventral ACC is associated more specifically with social inclusion (Somerville et al., 2006) or exclusion (Bolling et al., 2010) than expectancy violations (see also Onoda et al., 2009). Another related negative outcome resulting from social identity is stereotype threat, which occurs when a member of a group that is stereotyped as performing poorly in a given domain subsequently performs poorly in that domain after being reminded of his or her relevant group membership (Steele and Aronson, 1995). A recent neuroimaging study investigating this phenomenon found that when the stereotype of poor math performance among females was primed, participants activated more ventral ACC, and engaged regions involved in mathematics less (Krendl et al., 2008; see also Derks et al., 2008). These lines of research suggest that social exclusion is painful, making it a very salient motivator of self-oriented behavior and cognition related to such experiences, and that dorsal and ventral ACC together contribute to detecting and processing of such social evaluative threats (Heatherton, 2011).
4. Beyond CMS
As reviewed in the prior section, the influence of social, cultural, affective, and motivational processes on neural responses linked to personal and social identity can be observed at multiple levels of interpersonal experience. From considering what others may think about oneself to being included or excluded by peers, it is clear that CMS are not the only brain regions implicated in important self-related functions. In this section, we very briefly summarize the functions of these additional regions noted, which extend the network involved in maintaining personal and social identity: TPJ and adjacent pSTS, temporal poles, anterior insula, dorsal and ventral ACC, and vlPFC.
Reasoning about other people's beliefs or other mental states via third-person perspective-taking is thought to rely on TPJ, a region well-known for its association with theory-of-mind (D’Argembeau et al., 2007, Ruby and Decety, 2003, Samson et al., 2004, Saxe and Kanwisher, 2003, Saxe and Powell, 2006), although attentional processes also effectively recruit this region (Mitchell, 2008). A primary function of the adjacent pSTS is thought to be extracting information about goals and intentions from biological motion within a social context, which would support mentalizing (Pelphrey et al., 2004). Like TPJ and pSTS, the temporal poles have also been associated with theory-of-mind and mentalizing (Frith and Frith, 2003), but seem more specifically to be involved with linking perceptual memories of social interactions with emotions, creating a store of social semantic memory (Olson et al., 2007). If correct, this would give the temporal poles a critical role in supporting social components of autobiographical memory. The insula (particularly its anterior aspect) is recognized for its involvement in various aspects of interoceptive awareness, especially in subjective feelings associated with internal states (Craig, 2009), and has been suggested to mediate interactions with other brain networks based on the personal salience of an event (Menon and Uddin, 2010). Activation in anterior insula is most likely to occur in conjunction with mPFC in self-referential tasks with an emotional component, and has been shown to increase as the degree of self-relatedness of the task or emotion increases (Enzi et al., 2009, Modinos et al., 2009, Phan et al., 2004). Classically, dorsal and ventral ACC have been depicted as preferentially responsive to cognitive control and affect, respectively (e.g., Bush et al., 2000). However, more current understandings suggest dorsal ACC is involved in detecting and evaluating conflict, while ventral ACC supports conflict regulation, in both emotional and unemotional circumstances (Etkin et al., 2011). Dorsal ACC activity may also reflect the distressing aspects of physical and social pain (Eisenberger and Lieberman, 2004). Finally, vlPFC (particularly in the right hemisphere) appears directly responsible for exerting cognitive control to inhibit prepotent responses, or engage in tasks where inhibition is required such as set switching or memory retrieval (Aron et al., 2004). Together, these regions highlight the important contributions of mentalizing, emotion, and emotion regulation to social identity, and socioemotional aspects of the self.
5. Developmental perspectives
In this subsection, we discuss the growing neuroimaging evidence addressing the progression of self-development from various socioemotional and cognitive perspectives. On the one hand, there are many gaps to be filled. To our knowledge there are no fMRI studies of autobiographical memory in children. This is an area ripe for further inquiry given its extended developmental trajectory, and relation to parenting and discourse (Fivush, 2010). In addition, there are currently no developmental fMRI studies of how self-referential processing varies across cultures, or according to closeness or similarity to targets of comparison. On the other hand, evidence is beginning to accumulate about development of brain function related to self-appraisals, as well as social rejection.
In the first such study, children (ages 9–11 years) and young adults (ages 23–32 years) evaluated themselves and a familiar, fictional other (Harry Potter) with reference to social and academic competencies (Pfeifer et al., 2007). In both groups, a region of anterior rostral mPFC (extending into dorsal mPFC and rostral ACC in children) was more active during direct appraisals of self than other. Only adults engaged dorsal mPFC more when evaluating the other social target than themselves. During self-appraisals, children showed more activation in anterior rostral mPFC than adults, even engaging it above a resting baseline (a pattern that is rarely observed in adults during self-appraisals). A similar pattern was seen in a subsequent study (Pfeifer et al., 2009) of early adolescents (ages 11–14 years) and young adults (ages 23–30 years). Rather than contrasting evaluations of self and other, direct and reflected self-appraisals were compared. Yet the same condition as in the prior study (i.e., the process of making direct self-evaluations in social and academic domains) engaged anterior rostral and dorsal mPFC and ACC more in adolescents than adults, and activated anterior rostral mPFC above a resting baseline in adolescents but not adults. A third fMRI study found that for 7–13-year-old boys, recall of psychological trait words increased with age when the words were referenced to the self, but not when the words were referenced to the subjects’ mother (Ray et al., 2009). The neural basis of this developing self-reference effect was traced to activation in regions involved in the processing of emotion and memory, including rostral and ventral ACC, inferior frontal gyrus (IFG), and the caudate head.
Together these studies suggest anterior rostral and dorsal mPFC, as well as dorsal, rostral, and ventral ACC, are all more active in child or adolescent direct self-appraisals, in comparison with young adults. Based on the prior information in this review, there are several interpretations of these findings that can be proposed. One possibility is that children and adolescents are engaging in more active processes of integration and abstraction of self-knowledge – constructing self-appraisals rather than efficiently retrieving them (Pfeifer et al., 2007). The dorsal and ventral ACC activations suggest that perhaps direct self-appraisals elicited conflict detection and regulation processes (Pfeifer et al., 2009), such as that which might be expected when self-evaluations are influenced by multiple, sometimes conflicting perspectives (see Harter, 1999). In addition, there may be more overlap between self and others during childhood (Pfeifer et al., 2007, Ray et al., 2009). Together, these proposals lead to the prediction that longitudinal changes in responses from these regions during direct self-appraisals may be related to the individuation process across the transition to adolescence, as teenagers develop stable personal identities, although self-concepts continue to evolve over the lifespan.
During this identity development process in adolescence, however, what others think about the self may be intertwined with who they think they are – even on a neural level. In an fMRI study comparing direct self-appraisals with reflected self-appraisals from various viewpoints (Pfeifer et al., 2009), adolescents engaged CMS (anterior rostral and dorsal mPFC, mPPC) as well as TPJ, pSTS, and temporal poles during all forms of self-appraisals – both direct and reflected, unlike adults. CMS were most responsive when adolescents took their best friend's perspective on their social attributes, and their mom's perspective on their academic abilities. These findings are consistent with the idea that vulnerability of self-perceptions to social influences may be amplified in adolescence, as inclusion and affiliation with peers becomes increasingly important, and developing personal and social identities that foster a sense of continuity, coherence, and belonging emerges as a major developmental task (Brown, 2004, Erikson, 1968, Marcia, 1980, Steinberg and Silverberg, 1986). It would be interesting for future studies to explore whether individual differences in engagement of perspective-taking regions during direct self-appraisals in adolescence predicts outcomes like social anxiety, or susceptibility to peer influence over behavior and perceptions (for a similar line of inquiry with respect to emotion processing, see Guyer et al., 2008). Perhaps excessive recruitment of a region like TPJ during adolescent direct self-appraisals is associated with greater risk. Clearly, a variety of implications for health, academics, and general well-being can be derived from these hypotheses, making them important targets for future research.
As personal and social identity development proceed in tandem, and peers become increasingly important, the costs of rejection may become increasingly salient. Interestingly, the adolescent pattern of neural response to social exclusion is both similar to and different from that of adults. In one of the initial fMRI studies of adolescent peer rejection, the insula (but not the dorsal ACC) was relatively more activated during social exclusion than inclusion (Masten et al., 2009). Meanwhile, vlPFC, ventral striatum, and dorsal mPFC were negatively correlated with activity in regions that may have indexed distress responses, including dorsal ACC, insula, amygdala, and ventral ACC. In another study, anticipations of peer rejection or acceptance were combined with a reflected self-appraisal process, in which brain activity was assessed while adolescents reported whether they thought a peer was interested in interacting with them. Interestingly, both ventral striatum and insula activity increased with age for female but not male adolescents, when attempting to judge the interest of peers with whom participants previously reported they were more interested in interacting (Guyer et al., 2009). This suggests the threat of rejection from appealing peers becomes increasingly distressing to adolescent girls, and they may have to engage greater regulatory resources to cope. Yet another study found that ventral mPFC/ACC and the striatum were engaged when children, adolescents, and adults received social acceptance feedback, or expected to receive positive social evaluations, and the activity of these regions increased with age while expecting positive feedback (Gunther Moor et al., 2010). Meanwhile, lateral PFC among other regions was increasingly involved in processing social rejection feedback with age. One may hypothesize that, following social exclusion, failures to regulate (using vlPFC, ventral striatum, dorsal mPFC, or some combination thereof) may be associated with greater distress and probability that the peer rejection will be marked as self-relevant. Future studies should continue to build upon methods of studying neural responses to actual or anticipated peer rejection in a way that allows for the assessment of its impact on personal and social identity, such as self-evaluations of interpersonal competence or desire to affiliate with various peer groups.
6. Brain development
The previous subsections have highlighted a set of core brain regions implicated in personal and social identity development: CMS including mPFC (ventral, dorsal, and anterior rostral aspects, as well as adjacent rostral ACC) and mPPC (precuneus, posterior cingulate, and retrosplenial cortex). Other regions have also been implicated in a more extended network relevant to self-development, particularly when viewed from a social perspective: TPJ, pSTS, temporal poles, dorsal and ventral ACC, vlPFC, ventral striatum, and anterior insula. Understanding their contributions to self-development may be buoyed by understanding neuroanatomical development in these regions, which includes changes in the structural composition of the brain and in the dynamic aspects of how different brain regions function alone and interactively. In other words, the trajectories of both structural and functional development may directly impact social and cognitive functioning in a manner that holds meaningful implications for understanding self-development. For example, a recent study showed that structural maturation of the left TPJ was related to mirror self-recognition, personal pronoun use, and pretend play in toddlers (Lewis and Carmody, 2008), suggesting brain development may influence self-development even at a very early age. In this subsection, we summarize the available information about the development of these regions from a biological perspective.
Different brain regions involved in self-referential processing have different structural developmental trajectories. While there are many metrics of structural development (e.g., volume, gray and white matter density), we choose to focus here on cortical thickness.1 Gray matter exhibits an early period of rapid growth in childhood followed by a gradual thinning that extends into adulthood (Gogtay et al., 2004, Groeschel et al., 2010, Huttenlocher, 1979). Within this overall growth trend, however, the trajectories of reaching maximum cortical thickness vary for individual CMS components and other regions supporting cognitive functions of the self. A recent cohort-sequential study examined cortical development in a large sample of typically developing children, adolescents, and young adults (N = 375, ages 3.5–33.0 years; Shaw et al., 2008). The trajectory of cortical thickness in different regions frequently corresponded with cytoarchitecture, such that the most primitive cortex (i.e., allocortex) displayed linear decreases in cortical thickness, transitional cortex displayed quadratic functions over the age range sampled (increases followed by decreases, but lacking stabilization by the third decade of life), and homotypical isocortex displayed cubic functions (increases followed by decreases and then stabilization in the third decade of life). In addition to distinguishing regional patterns of cortical maturation,2 the study also provided animated representations of changes in cortical thickness between ages 5.0–15.0, which allow for the estimation of potentially meaningful structural milestones during a period that is critical for self-development (see http://neuro.cjb.net/content/vol28/issue14/images/data/3586/DC1/Movie2_medial.mov). We have used these representations to estimate the approximate age range of maximum cortical thickness in regions supporting personal and social identity development. Since peak cortical thickness can only be estimated if a region shows a non-linear (i.e., quadratic or cubic) growth function, some regions of interest in this review could not be estimated due to linear trajectories. Areas with linear growth functions, or those that were not reported specifically in the tables or depicted in the supplementary movies, included anterior insula, temporal poles, ventral striatum, and much of ventral ACC and retrosplenial cortex.
Cortical thickness in frontal regions of the CMS peaked first in ventral mPFC at around age 8.0–8.5 years. Beginning slightly later, anterior rostral mPFC reached maximum cortical thickness between 9.0 and 9.5 years, and rostral ACC between 9.0 and 10.5 years. The progression of peak thickness continued upward to reach dorsal mPFC at 10.0–10.5 years. The trend of cortical development in frontal CMS thus followed an inferior to superior progression with a simultaneous anterior to posterior progression. This corresponds to a general sequence in which affective or motivational processes (associated with ventral and anterior rostral mPFC) may be primary, and cognitive processes (associated with dorsal mPFC) may be secondary. The progression also suggests there may be more overlap between representations of self and others early in development, with differentiation emerging concurrently with the individuation process during the transition to adolescence. Posterior regions of the CMS followed a different developmental sequence, progressing from superior-posterior regions to anterior-inferior regions. Maximum cortical thickness was reached first in the precuneus between 8.0 and 10.0 years, continuing in anterior and inferior directions, to gradually encompass the posterior cingulate between 9.0 and 11.5 years. Several of the regions identified previously in addition to CMS were estimable. vlPFC reached peak cortical thickness around age 9.0–9.5 years bilaterally. While peak cortical thickness in TPJ was more difficult to estimate, it may be reached as early as 8.0–9.0 years in the right hemisphere, and in the left hemisphere around age 8.5–9.5 years, although regions of adjacent superior temporal gyrus (which may include pSTS) did not peak in cortical thickness until approximately 15.0 years. Most of dorsal (and some aspects of ventral) ACC peaked between 11.0 and 13.5 years. Finally, although anterior insula exhibited a linear trajectory, middle and posterior insula peaked in cortical thickness quite late, around 18.0 years of age. Together, these developmental trajectories suggest mentalizing, emotion, and emotion regulation processes might become increasingly important as children move into adolescence. Future studies should explore the relative contributions of biological and other contextual factors to these patterns, ranging from the onset of puberty to shifts toward greater peer orientation within a changing social landscape.
The participation of both frontal and parietal regions in personal and social identity processes points to another important factor: the development of structural and functional connectivity that supports communication between regions, which is associated with the maturation of white matter. In contrast to gray matter, white matter development shows a steady increase into adulthood (Giedd et al., 1999, Tamnes et al., 2010) that may influence neural connectivity both within CMS regions and between CMS and other regions, like TPJ. Studies show that in middle childhood, these regions exhibit stronger local, short-range connections, but over time (through adolescence into young adulthood), long-range connections are strengthened (Dosenbach et al., 2010, Fair et al., 2008, Supekar et al., 2010). Using TPJ involvement in reflected self-appraisals as an example, this may mean that in childhood, TPJ outputs (my classmates think I’m bad at sports) may not be conveyed as well to frontal regions such as mPFC as to posterior regions such as mPPC. Reflected self-appraisals at this early stage may draw primarily on autobiographical memories (the time I let the other team score the winning goal and everyone was mad at me) in short-range, medial parietal regions. Perhaps to the extent these reflected self-appraisals and autobiographical memories are emotionally salient, such content will be integrated and incorporated into self-evaluations, after connectivity between parietal and frontal regions matures. An alternative prediction is that, if the entire network is maturing, other processes that recruit this network should show similar developmental patterns (see Burnett and Blakemore, 2009). This derives from viewing the maturational changes in this network more broadly, perhaps as exhibiting a combination of patterns: an anterior-to-posterior shift (Sebastian et al., 2008), complemented by increased specialization across development in posterior cortical regions (Saxe et al., 2009).
Researchers should ultimately aspire to examine aspects of personal and social identity development concurrently with both structural and functional neuroimaging during the transition from childhood to adolescence, ideally in a longitudinal sample, to work towards disentangling cognitive, socioemotional, and biological effects. It is important to note that the brain represents just one of many possible biological contributions. Although brain function and structure are the biological factors that have received the most empirical attention thus far, widening the research base to include greater study of neuroendocrine, psychophysiological, genetic, and electrophysiological factors that contribute to self-development will be critical in the future.
7. Atypical self-development
This review demonstrates not only that neural foundations of self and identity can be considered from a cognitive perspective, but also that there are important socioemotional influences to take into account, as well as bidirectional influences among these two factors and brain development. Taken together, the neuroimaging evidence from child and adolescent samples points to a critical role for mentalizing, emotion, and emotion regulation in typical self-development – more so than perhaps would be anticipated from adult studies alone, although that work has nicely highlighted the importance of social context in investigations that compare self-evaluations across cultures, or with evaluations of similar or close others. Brain regions associated with affective and motivational processes, such as anterior rostral mPFC, ventral posterior cingulate, ventral striatum, insula, and ventral ACC (Guyer et al., 2009, Masten et al., 2009, Pfeifer et al., 2007, Pfeifer et al., 2009, Ray et al., 2009), have been found to be more engaged in children and adolescents compared to adults, suggesting a significant contribution of emotion to personal and social identity development. Given this greater affective content, it is little wonder that regulatory demands may be greater across various self-relevant tasks: deciding who I am, considering what others think about me, and making efforts to fit in that are sometimes rebuffed. Perhaps areas involved in inhibition and the detection or regulation of conflict, like vlPFC or dorsal and ventral ACC, are not only involved in direct self-appraisals and responses to social exclusion during adolescence (Masten et al., 2009, Pfeifer et al., 2009), but will also be observed in adolescent reflected self-appraisals and autobiographical memories. Finally, the neuroimaging evidence also suggests self-development not only surges in tandem with emotions, but also draws deeply on mentalizing resources, and hinges on experiences and connections with peers, family members, and others. This is illustrated by greater recruitment of brain regions involved in perspective-taking, like TPJ and mPPC, during both direct and reflected self-appraisals in adolescence than adulthood (Pfeifer et al., 2009).
These conclusions imply self-development is likely to be perturbed in disorders that affect mentalizing abilities or are associated with atypical emotion processing and regulation, such as autism or depression (although many other disorders exhibit disrupted self-development, such as social anxiety, borderline personality disorder, and schizophrenia, but addressing each is beyond the scope of this review). There is a growing body of evidence that brain function during self-appraisals and social exclusion differs from adult norms in these and other populations. For example, recent studies of adults with autism have begun to demonstrate that mentalizing capabilities are positively associated with various indicators of personal identity (such as self-consciousness and the self-reference effect; Lombardo et al., 2007), which again suggests that these capabilities rely on each other for typical development and functioning to some meaningful extent. Similar patterns have been observed in children with autism, who frequently demonstrate deficits in autobiographical memory, prospection, the self-reference effect, and many other gauges of self-development (for reviews, see Hobson et al., 2006, Lind and Bowler, 2008, Lombardo and Baron-Cohen, 2010).
In one fMRI study, ventral and anterior rostral mPFC were hypoactive in adults with autism during direct self-appraisals and unlike neurotypical controls, did not distinguish between self and other on average (Lombardo et al., 2010b; for similar findings, see Chiu et al., 2008, Kennedy and Courchesne, 2008). Interestingly, middle cingulate cortex (MCC; another structure located on the cortical midline) was also differentially recruited by the two groups of participants. Specifically, when making mentalizing judgments MCC was more active for self than other in neurotypical adults, but it was more active for other than self in adults with autism. In other words, MCC preferentially responded to other-appraisals in autism, rather than the normative pattern of activating more during self-appraisals. Finally, greater engagement of ventral mPFC for self than other was observed in those individuals who evidenced less impairment on a retrospective diagnostic autism assessment providing a clinical measure of reciprocal social impairment. Together these results may imply that in the case of autism, one core challenge is that self-appraisals are not tagged with the same affective and motivational content as in typical development. Similar arguments have been made with respect to deficits in self-awareness and theory-of-mind in schizophrenia and alexithymia, namely that difficulties in comprehending one's own affective states and impaired capabilities for affective mentalizing are interrelated at behavioral and neural levels (Moriguchi et al., 2006, Shamay-Tsoory et al., 2007). Another core challenge in these disorders may be that mentalizing deficits impair the ability to make reflected self-appraisals with the ease and frequency of neurotypical adolescents. As of yet, there are no published fMRI studies of children or adolescents with autism or these other disorders engaging in direct or reflected self-appraisals, but this is a promising avenue for future research.
It is worth noting that in neurotypical adults, excessive self-referential behavior was associated with poor mentalizing, unlike in adults with autism (Lombardo et al., 2007). In other words, there may be a non-linear function characterizing the involvement of mentalizing regions in personal and social identity development, such that too much may be as harmful as too little. Recent fMRI studies of depression provide support for this hypothesis, finding that both depressed and neurotypical adults engage anterior rostral mPFC during self-appraisals, but that depressed adults engage more dorsal and/or anterior rostral mPFC (including rostral ACC), dorsal ACC, and dorsolateral PFC (dlPFC; Lemogne et al., 2009, Yoshimura et al., 2010). These activations have been interpreted to reflect excessive self-focus, negative affect, and resultant need for greater cognitive control. In one of these studies, greater depressive symptoms were correlated with anterior rostral mPFC activity (including rostral ACC) during self-appraisals, and responses in these regions were furthermore positively correlated with amygdala responses (Yoshimura et al., 2010; see also Lemogne et al., 2010). To our knowledge, there are no published fMRI studies of depressed children or adolescents engaging in direct or reflected self-appraisals. However, enhanced responses to reward in anterior rostral mPFC, and diminished responses to reward in ventral striatum, have been associated with greater depressive symptoms and less daily positive affect in an fMRI study of neurotypical adolescents (Forbes et al., 2010), and depressed adolescents similarly engaged less ventral striatum and more anterior rostral mPFC, dorsal ACC, and dlPFC (Forbes et al., 2009). A recent behavioral study also found that overly general autobiographical memories were predictive of concurrent and subsequent depressive symptomatology among adolescent girls (Hipwell et al., 2011; see also Drummond et al., 2006).
Finally, a common problem faced by children and adolescents with depression or autism is peer rejection (Attwood, 2008, Boivin and Hymel, 1997, Boivin et al., 1994, Little, 2001, Sebastian et al., 2009), which in the developmental psychology literature is often explicitly connected with negative social self-appraisals. Two published studies relating neural responses to social exclusion with depressive symptomatology have produced results consistent with the idea that exaggerated affective responses (and associated regulatory responses) to rejection are present in this context as well. Neurotypical young adults with low self-esteem demonstrated greater dorsal and ventral ACC responses during social exclusion, and reported greater distress afterwards (Onoda et al., 2010). In adolescence, greater ventral ACC responses to social exclusion were associated with longitudinal increases in parent-reported depressive symptoms 1 year after the fMRI scan (Masten et al., 2011b). Similar to the patterns demonstrated with respect to self-appraisals, wherein autism was associated with hypoactivation and depression with hyperactivation of relevant neural systems, one may hypothesize that in children, adolescents, or adults with autism, less affective or mentalizing responses to social exclusion will be observed. This was confirmed in recent studies of high-functioning adolescents with autism, who displayed significantly less activity than neurotypical adolescents in regions including anterior insula, ventral ACC, vlPFC, and ventral striatum during social exclusion relative to inclusion (Bolling et al., 2011, Masten et al., 2011a, McPartland et al., 2011; for discussion, see Sebastian and Blakemore, 2011).
Comparisons between these two populations (children and adolescents with autism and depression) should pay heed to their different patterns of onset and prevalence by gender. Autism is more common among males, while depression is more common among females. Autism is also typically diagnosed early in life (by 3 years of age), while rates of depression significantly increase in adolescence, particularly among girls. Clearly, variations in the onset and nature of these disorders will be associated with different stages of cognitive, socioemotional, and neurological maturation. Furthermore, challenges in one aspect of development are likely to affect the others, creating a potential cascade of effects. Factors such as these may be intrinsically related to the patterns of functioning observed in each population.
8. Conclusion
In summary, fMRI data from neurotypical children and adolescents as well as youth and adults with autism, depression, and other disorders confirm how critical the socioemotional perspective on self-development may be to understanding how our complex sense of the self – as unique and yet connected with others – is supported by the brain. The trajectories of cortical development in CMS (mPFC and mPPC) and the more extended group of regions identified in this review (including TPJ, pSTS, temporal poles, insula, dorsal and ventral ACC, ventral striatum, and vlPFC) suggest that mentalizing, emotion, and emotion regulation processes elicited by reflected self-appraisals and social exclusion may play very important roles in personal and social identity formation around the transition to adolescence. As constrained by structural brain development, this may be a period when self-development is most sensitive to affective, motivational, interpersonal, or cultural influences (see Casey et al., 2010). Conversely, the accumulation of experiences within a specific relationship, peer group, or cultural context may ultimately affect underlying biology, to facilitate diminished, enhanced, or even excessive responses in brain regions that support the socioemotional and cognitive construction of the self. We are only just beginning to understand the neural systems involved in typical and atypical self-development, but each step towards identifying component processes provides the opportunity to intervene in ways that will help all individuals develop full and healthy personal and social identities.
Footnotes
It should be noted that there are many ways of interpreting what these metrics of structural brain development mean at a cellular level. For example, changes in cortical thickness may represent proliferation of myelin into the cortical neuropil (Sowell et al., 2004), synaptic pruning, and so on.
Although this study focused on cortical maturation, given that the VS also emerged as a region of interest in this review, readers are referred to a recent study that examined trajectories of subcortical maturation (Ostby et al., 2009). In this study, the basal ganglia exhibited a linear decrease in gray matter volume (corrected for total brain volume) from ages 8–31 years, although the relationship between volume and age was weaker in the caudate than the putamen, pallidum, or accumbens.
References
- Addis D.R., Wong A.T., Schacter D.L. Remembering the past and imagining the future: common and distinct neural substrates during event construction and elaboration. Neuropsychologia. 2007;45(7):1363–1377. doi: 10.1016/j.neuropsychologia.2006.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez J.M., Ruble D.N., Bolger N. Trait understanding or evaluative reasoning? An analysis of children's behavioral predictions. Child Development. 2001;72(5):1409–1425. doi: 10.1111/1467-8624.00356. [DOI] [PubMed] [Google Scholar]
- Ames D.L., Jenkins A.C., Banaji M.R., Mitchell J.P. Taking another person's perspective increases self-referential neural processing. Psychological Science: A Journal of the American Psychological Society/APS. 2008;19(7):642–644. doi: 10.1111/j.1467-9280.2008.02135.x. [DOI] [PubMed] [Google Scholar]
- Andreasen N.C., O’Leary D.S., Cizadlo T., Arndt S., Rezai K., Watkins G.L., Hichwa R.D. Remembering the past: two facets of episodic memory explored with positron emission tomography. The American Journal of Psychiatry. 1995;152(11):1576–1585. doi: 10.1176/ajp.152.11.1576. [DOI] [PubMed] [Google Scholar]
- Aron A.R., Robbins T.W., Poldrack R.A. Inhibition and the right inferior frontal cortex. Trends in Cognitive Sciences. 2004;8(4):170–177. doi: 10.1016/j.tics.2004.02.010. [DOI] [PubMed] [Google Scholar]
- Attwood T. Jessica Kingsley Pub; 2008. The Complete Guide to Asperger's Syndrome. [Google Scholar]
- Baldwin J.M. University of Chicago Press; 1895. Mental Development in the Child and the Race. [Google Scholar]
- Benoit R.G., Gilbert S.J., Volle E., Burgess P.W. When I think about me and simulate you: medial rostral prefrontal cortex and self-referential processes. Neuroimage. 2010;50(3):1340–1349. doi: 10.1016/j.neuroimage.2009.12.091. [DOI] [PubMed] [Google Scholar]
- Blakemore S.J., Burnett S., Dahl R.E. The role of puberty in the developing adolescent brain. Human Brain Mapping. 2010;31(6):926–933. doi: 10.1002/hbm.21052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boivin M., Hymel S. Peer experiences and social self-perceptions: a sequential model. Developmental Psychology. 1997;33(1):135–145. doi: 10.1037//0012-1649.33.1.135. [DOI] [PubMed] [Google Scholar]
- Boivin M., Poulin F., Vitaro F. Depressed mood and peer rejection in childhood. Development and Psychopathology. 1994;6(03):483–498. [Google Scholar]
- Bolling D.Z, Pitskel N.B., Deen B., Crowley M.J., McPartland J.C., Kaiser M.D., Vander Wyk B.C., Wua J., Mayes L.C., Pelphrey K.A. Enhanced neural responses to rule violation in children with autism: A comparison to social exclusion. Developmental Cognitive Neuroscience. 2011;1(3):280–294. doi: 10.1016/j.dcn.2011.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolling D.Z., Pitskel N.B., Deen B., Crowley M.J., McPartland J.C., Mayes L.C., Pelphrey K.A. Dissociable brain mechanisms for processing social exclusion and rule violation. Neuroimage. 2010;54:2462–2471. doi: 10.1016/j.neuroimage.2010.10.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouchey H.A., Harter S. Reflected appraisals, academic self-perceptions, and math/science performance during early adolescence. Journal of Educational Psychology. 2005;97:673–686. [Google Scholar]
- Brown B.B. Adolescents’ relationships with peers. In: Lerner R.M., Steinberg L.D., editors. vol. 2. John Wiley & Sons Inc.; Hoboken, NJ: 2004. pp. 363–394. (Handbook of Adolescent Psychology). [Google Scholar]
- Brown B.B., Huang B. Examining parenting practices in different peer contexts: implications for adolescent trajectories. In: Crockett L.J., Crouter A.C., editors. Pathways Through Adolescence: Individual Development in Relation to Social Contexts. Lawrence Erlbaum Associates Inc.; Mahwah, NJ: 1995. pp. 151–174. [Google Scholar]
- Burgess P.W., Dumontheil I., Gilbert S.J. The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Sciences. 2007;11(7):290–298. doi: 10.1016/j.tics.2007.05.004. [DOI] [PubMed] [Google Scholar]
- Burklund L.J., Eisenberger N.I., Lieberman M.D. The face of rejection: rejection sensitivity moderates dorsal anterior cingulate activity to disapproving facial expressions. Social Neuroscience. 2007;2(3):238–253. doi: 10.1080/17470910701391711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnett S., Blakemore S.J. Functional connectivity during a social emotion task in adolescents and in adults. European Journal of Neuroscience. 2009;29:1294–1301. doi: 10.1111/j.1460-9568.2009.06674.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bush G., Luu P., Posner M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences. 2000;4(6):215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Butler R. Age trends in the use of social and temporal comparison for self-evaluation: examination of a novel developmental hypothesis. Child Development. 1998;69(4):1054–1073. [PubMed] [Google Scholar]
- Casey B.J., Duhoux S., Cohen M.M. Adolescence: what do transmission, transition, and translation have to do with it? Neuron. 2010;67(5):749–760. doi: 10.1016/j.neuron.2010.08.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanna A.E., Trimble M.R. The precuneus: a review of its functional anatomy and behavioural correlates. Brain: A Journal of Neurology. 2006;129(Pt 3):564–583. doi: 10.1093/brain/awl004. [DOI] [PubMed] [Google Scholar]
- Chen A.C., Welsh R.C., Liberzon I., Taylor S.F. ‘Do I like this person?’ A network analysis of midline cortex during a social preference task. Neuroimage. 2010;51(2):930–939. doi: 10.1016/j.neuroimage.2010.02.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiao J., Harada T., Komeda H., Li Z., Mano Y., Saito D. Dynamic cultural influences on neural representations of the self. Journal of Cognitive Neuroscience. 2010;22:1–11. doi: 10.1162/jocn.2009.21192. [DOI] [PubMed] [Google Scholar]
- Chiao J.Y., Harada T., Komeda H., Li Z., Mano Y., Saito D., Parrish T.B., Sadato N., Iidaka T. Neural basis of individualistic and collectivistic views of self. Human Brain Mapping. 2009;30(9):2813–2820. doi: 10.1002/hbm.20707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu P.H., Kayali M.A., Kishida K.T., Tomlin D., Klinger L.G., Klinger M.R., Montague P.R. Self responses along cingulate cortex reveal quantitative neural phenotype for high-functioning autism. Neuron. 2008;57(3):463–473. doi: 10.1016/j.neuron.2007.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coie J.D. Toward a theory of peer rejection. Peer Rejection in Childhood. 1990:365–401. [Google Scholar]
- Coie J.D., Cillessen A.H.N. Peer rejection: origins and effects on children's development. Current Directions in Psychological Science. 1993;2(3):89–93. [Google Scholar]
- Conway M.A., Pleydell-Pearce C.W. The construction of autobiographical memories in the self-memory system. Psychological Review. 2000;107(2):261–288. doi: 10.1037/0033-295x.107.2.261. [DOI] [PubMed] [Google Scholar]
- Cooley C.H. Charles Scribner's Sons; New York: 1902. Human Nature and the Social Order. [Google Scholar]
- Craig A.D. How do you feel—now? The anterior insula and human awareness. Nature Reviews Neuroscience. 2009;10(1):59–70. doi: 10.1038/nrn2555. [DOI] [PubMed] [Google Scholar]
- Csikszentmihalyi M., Larson R. Basic Books; New York: 1984. Being Adolescent. [Google Scholar]
- Dahl R.E. Adolescent brain development: a period of vulnerabilities and opportunities Keynote address. Annals of the New York Academy of Sciences. 2004;1021:1–22. doi: 10.1196/annals.1308.001. [DOI] [PubMed] [Google Scholar]
- Damon W., Hart D. Cambridge University Press; New York: 1988. Self-understanding in Childhood and Adolescence. [Google Scholar]
- D’Argembeau A., Ruby P., Collette F., Degueldre C., Balteau E., Luxen A., Salmon E. Distinct regions of the medial prefrontal cortex are associated with self-referential processing and perspective taking. Journal of Cognitive Neuroscience. 2007;19(6):935–944. doi: 10.1162/jocn.2007.19.6.935. [DOI] [PubMed] [Google Scholar]
- D’Argembeau A., Xue G., Lu Z.L., Van der Linden M., Bechara A. Neural correlates of envisioning emotional events in the near and far future. Neuroimage. 2008;40(1):398–407. doi: 10.1016/j.neuroimage.2007.11.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derks B., Inzlicht M., Kang S. The neuroscience of stigma and stereotype threat. Group Processes and Intergroup Relations. 2008;11:163–181. [Google Scholar]
- Dosenbach N.U.F., Nardos B., Cohen A.L., Fair D.A., Power J.D., Church J.A., Lessov-Schlaggar C.N. Prediction of individual brain maturity using fMRI. Science. 2010;329(5997):1358–1361. doi: 10.1126/science.1194144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drummond L.E., Dritschel B., Astell A., O’Carroll R.E., Dalgleish T. Effects of age, dysphoria, and emotion focusing on autobiographical memory specificity in children. Cognition & Emotion. 2006;20:488–505. doi: 10.1080/02699930500341342. [DOI] [PubMed] [Google Scholar]
- Eisenberger N.I., Lieberman M.D. Why rejection hurts: a common neural alarm system for physical and social pain. Trends in Cognitive Sciences. 2004;8(7):294–300. doi: 10.1016/j.tics.2004.05.010. [DOI] [PubMed] [Google Scholar]
- Eisenberger N.I., Lieberman M.D., Williams K.D. Does rejection hurt? An FMRI study of social exclusion. Science. 2003;302(5643):290–292. doi: 10.1126/science.1089134. [DOI] [PubMed] [Google Scholar]
- Elkind D. Egocentrism in adolescence. Child Development. 1967;38:1025–1034. [PubMed] [Google Scholar]
- Elkind D. Egocentrism redux. Developmental Review. 1985;5(3):218–226. [Google Scholar]
- Enzi B., de Greck M., Prösch U., Tempelmann C., Northoff G. Is our self nothing but reward? Neuronal overlap and distinction between reward and personal relevance and its relation to human personality. PLoS One. 2009;4(12):e8429. doi: 10.1371/journal.pone.0008429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erikson E.H. W.W. Norton & Company; 1968. Identity: Youth and Crisis. [Google Scholar]
- Etkin A., Egner T., Kalisch R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends in Cognitive Sciences. 2011;15(2):85–93. doi: 10.1016/j.tics.2010.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fair D.A., Cohen A.L., Dosenbach N.U.F., Church J.A., Miezin F.M., Barch D.M., Schlaggar B.L. The maturing architecture of the brain's default network. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(10):4028–4032. doi: 10.1073/pnas.0800376105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fivush R. The development of autobiographical memory. Annual Review of Psychology. 2010;62:559–582. doi: 10.1146/annurev.psych.121208.131702. [DOI] [PubMed] [Google Scholar]
- Forbes E.E., Hariri A.R., Martin S.L., Silk J.S., Moyles D.L., Fisher P.M., Dahl R.E. Altered striatal activation predicting real-world positive affect in adolescent major depressive disorder. The American Journal of Psychiatry. 2009;166(1):64–73. doi: 10.1176/appi.ajp.2008.07081336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forbes E.E., Ryan N.D., Phillips M.L., Manuck S.B., Worthman C.M., Moyles D.L., Dahl R.E. Healthy adolescents’ neural response to reward: associations with puberty, positive affect, and depressive symptoms. Journal of the American Academy of Child and Adolescent Psychiatry. 2010;49(2):162–172. doi: 10.1097/00004583-201002000-00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frith U., Frith C.D. Development and neurophysiology of mentalizing. Philosophical Transactions of the Royal Society of London B. 2003;358(1431):459–473. doi: 10.1098/rstb.2002.1218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giedd J.N., Blumenthal J., Jeffries N.O., Castellanos F.X., Liu H., Zijdenbos A., Rapoport J.L. Brain development during childhood and adolescence: a longitudinal MRI study. Nature Neuroscience. 1999;2:861–862. doi: 10.1038/13158. [DOI] [PubMed] [Google Scholar]
- Gilbert S.J., Spengler S., Simons J.S., Frith C.D., Burgess P.W. Differential functions of lateral and medial rostral prefrontal cortex (area 10) revealed by brain-behavior associations. Cerebral Cortex. 2006;16(12):1783–1789. doi: 10.1093/cercor/bhj113. [DOI] [PubMed] [Google Scholar]
- Gogtay N., Giedd J.N., Lusk L., Hayashi K.M., Greenstein D., Vaituzis A.C., Thompson P.M. Dynamic mapping of human cortical development during childhood through early adulthood. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(21):8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenwald A.G. Self and memory. Psychology of Learning and Motivation. 1981;15:201–236. [Google Scholar]
- Greenwald A.G., Banaji M.R. The self as a memory system: powerful, but ordinary. Journal of Personality and Social Psychology. 1989;57(1):41–54. [Google Scholar]
- Groeschel S., Vollmer B., King M.D., Connelly A. Developmental changes in cerebral grey and white matter volume from infancy to adulthood. International Journal of Developmental Neuroscience. 2010;28(6):481–489. doi: 10.1016/j.ijdevneu.2010.06.004. [DOI] [PubMed] [Google Scholar]
- Gunther Moor B., van Leijenhorst L., Rombouts S.A.R.B., Crone E.A., Van der Molen M.W. Do you like me? Neural correlates of social evaluation and developmental trajectories. Social Neuroscience. 2010;5(5–6):461–482. doi: 10.1080/17470910903526155. [DOI] [PubMed] [Google Scholar]
- Gusnard D.A., Akbudak E., Shulman G.L., Raichle M.E. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(7):4259–4264. doi: 10.1073/pnas.071043098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., Lau J.Y., McClure-Tone E.B., Parrish J., Shiffrin N.D., Reynolds R.C., Nelson E.E. Amygdala and ventrolateral prefrontal cortex function during anticipated peer evaluation in pediatric social anxiety. Archives of General Psychiatry. 2008;65(11):1303–1312. doi: 10.1001/archpsyc.65.11.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., McClure-Tone E.B., Shiffrin N.D., Pine D.S., Nelson E.E. Probing the neural correlates of anticipated peer evaluation in adolescence. Child Development. 2009;80(4):1000–1015. doi: 10.1111/j.1467-8624.2009.01313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han S., Northoff G. Culture-sensitive neural substrates of human cognition: a transcultural neuroimaging approach. Nature Reviews Neuroscience. 2008;9(8):646–654. doi: 10.1038/nrn2456. [DOI] [PubMed] [Google Scholar]
- Harter S. The Guilford Press; New York, NY: 1999. The Construction of the Self: A Developmental Perspective. [Google Scholar]
- Harter S. 6th ed. vol. 3. Wiley; 2006. The self. (Handbook of Child Psychology). pp. 505–568. [Google Scholar]
- Heatherton T.F. Neuroscience of self and self-regulation. Annual Review of Psychology. 2011;62:363–390. doi: 10.1146/annurev.psych.121208.131616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heatherton T.F., Wyland C.L., Macrae C.N., Demos K.E., Denny B.T., Kelley W.M. Medial prefrontal activity differentiates self from close others. Social Cognitive and Affective Neuroscience. 2006;1(1):18–25. doi: 10.1093/scan/nsl001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hipwell A.E., Sapotichne B., Klostermann S., Battista D., Keenan K. Autobiographical memory as a predictor of depression vulnerability in girls. Journal of Clinical Child and Adolescent Psychology. 2011;40(2):254–265. doi: 10.1080/15374416.2011.546037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobson P.R., Chidambi G., Lee A., Meyer J. Self awareness in developmental perspective. Monographs of the Society for Research in Child Development. 2006;71(2):1–28. doi: 10.1111/j.1540-5834.2006.00387.x. [DOI] [PubMed] [Google Scholar]
- Huttenlocher P.R. Synaptic density in human frontal cortex—developmental changes and effects of aging. Brain Research. 1979;163:195–205. doi: 10.1016/0006-8993(79)90349-4. [DOI] [PubMed] [Google Scholar]
- Jackson P.L., Brunet E., Meltzoff A.N., Decety J. Empathy examined through the neural mechanisms involved in imagining how I feel versus how you feel pain. Neuropsychologia. 2006;44(5):752–761. doi: 10.1016/j.neuropsychologia.2005.07.015. [DOI] [PubMed] [Google Scholar]
- James W. Holt; New York: 1890. Principles of Psychology. [Google Scholar]
- Jenkins A.C., Macrae C.N., Mitchell J.P. Repetition suppression of ventromedial prefrontal activity during judgments of self and others. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(11):4507–4512. doi: 10.1073/pnas.0708785105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson M.K., Nolen-Hoeksema S., Mitchell K.J., Levin Y. Medial cortex activity, self-reflection and depression. Social Cognitive and Affective Neuroscience. 2009;4(4):313–327. doi: 10.1093/scan/nsp022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson M.K., Raye C.L., Mitchell K.J., Touryan S.R., Greene E.J., Nolen-Hoeksema S. Dissociating medial frontal and posterior cingulate activity during self-reflection. Social Cognitive and Affective Neuroscience. 2006;1(1):56–64. doi: 10.1093/scan/nsl004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson S.C., Baxter L.C., Wilder L.S., Pipe J.G., Heiserman J.E., Prigatano G.P. Neural correlates of self-reflection. Brain. 2002;125(8):1808. doi: 10.1093/brain/awf181. [DOI] [PubMed] [Google Scholar]
- Kaplan J.T., Aziz-Zadeh L., Uddin L.Q., Iacoboni M. The self across the senses: an fMRI study of self-face and self-voice recognition. Social Cognitive and Affective Neuroscience. 2008;3(3):218–223. doi: 10.1093/scan/nsn014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley W.M., Macrae C.N., Wyland C.L., Caglar S., Inati S., Heatherton T.F. Finding the self? An event-related fMRI study. Journal of Cognitive Neuroscience. 2002;14(5):785–794. doi: 10.1162/08989290260138672. [DOI] [PubMed] [Google Scholar]
- Kennedy D.P., Courchesne E. Functional abnormalities of the default network during self- and other-reflection in autism. Social Cognitive and Affective Neuroscience. 2008;3(2):177–190. doi: 10.1093/scan/nsn011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kihlstrom J.F., Beer J.S., Klein S.B. Self and identity as memory. In: Leary M.R., Tangney J., editors. Handbook of Self and Identity. Guilford Press; New York: 2002. pp. 68–90. [Google Scholar]
- Kihlstrom J.F., Klein S.B. Handbook of Social Cognition. Lawrence Erlbaum Associates, Inc.; Hillsdale, NJ, US: 1994. The self as a knowledge structure. pp. 153–208. [Google Scholar]
- Kitayama S., Park J. Cultural neuroscience of the self: understanding the social grounding of the brain. Social Cognitive and Affective Neuroscience. 2010;5(2–3):111. doi: 10.1093/scan/nsq052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein S.B. Individual Self, Relational Self, Collective Self. Psychology Press/Taylor & Francis; Philadelphia, PA US: 2001. A self to remember: a cognitive neuropsychological perspective on how self creates memory and memory creates self. pp. 25–46. [Google Scholar]
- Klein S.B., Sherman R.W., Loftus J. The role of episodic and semantic memory in the development of trait self-knowledge. Social Cognition. 1996;14:277–291. [Google Scholar]
- Krendl A.C., Richeson J.A., Kelley W.M., Heatherton T.F. The negative consequences of threat: an fMRI investigation of the neural mechanisms underlying women's underperformance in math. Psychological Science. 2008;19:168–175. doi: 10.1111/j.1467-9280.2008.02063.x. [DOI] [PubMed] [Google Scholar]
- Krienen F.M., Tu P., Buckner R.L. Clan mentality: evidence that the medial prefrontal cortex responds to close others. The Journal of Neuroscience. 2010;30(41):13906–13915. doi: 10.1523/JNEUROSCI.2180-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leary M.R., Tangney J.P. The self as an organizing construct in the behavioral and social sciences. Handbook of Self and Identity. 2003:3–14. [Google Scholar]
- Legrand D., Ruby P. What is self-specific? Theoretical investigation and critical review of neuroimaging results. Psychological Review. 2009;116(1):252–282. doi: 10.1037/a0014172. [DOI] [PubMed] [Google Scholar]
- Lemogne C., Gorwood P., Bergouignan L., Pélissolo A., Lehéricy S., Fossati P. Negative affectivity, self-referential processing and the cortical midline structures. Social Cognitive and Affective Neuroscience. 2010 doi: 10.1093/scan/nsq049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemogne C., Le Bastard G., Mayberg H., Volle E., Bergouignan L., Lehéricy S., Fossati P. In search of the depressive self: extended medial prefrontal network during self-referential processing in major depression. Social Cognitive and Affective Neuroscience. 2009;4:305–312. doi: 10.1093/scan/nsp008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis M., Carmody D. Self-representation and brain development. Developmental Psychology. 2008;44:1329–1334. doi: 10.1037/a0012681. [DOI] [PubMed] [Google Scholar]
- Li J. Self in learning: Chinese adolescents’ goals and sense of agency. Child Development. 2006;77:482–501. doi: 10.1111/j.1467-8624.2006.00883.x. [DOI] [PubMed] [Google Scholar]
- Lind S.E., Bowler D. Episodic memory and autonoetic consciousness in autistic spectrum disorders: the roles of self-awareness, representational abilities and temporal cognition. Memory in Autism: Theory and Evidence. 2008:166–187. [Google Scholar]
- Little L. Peer victimization of children with Asperger spectrum disorders. Journal of the American Academy of Child and Adolescent Psychiatry. 2001;40:995–996. doi: 10.1097/00004583-200109000-00007. [DOI] [PubMed] [Google Scholar]
- Lombardo M.V., Barnes J.L., Wheelwright S.J., Baron-Cohen S. Self-referential cognition and empathy in autism. PLoS One. 2007;2(9):e883. doi: 10.1371/journal.pone.0000883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombardo M.V., Baron-Cohen S. Unraveling the paradox of the autistic self. WIREs Cognitive Science. 2010;1:393–403. doi: 10.1002/wcs.45. [DOI] [PubMed] [Google Scholar]
- Lombardo M.V., Chakrabarti B., Bullmore E.T., Wheelwright S.J., Sadek S.A., Suckling J., Baron-Cohen S. Shared neural circuits for mentalizing about the self and others. Journal of Cognitive Neuroscience. 2010;22:1623–1635. doi: 10.1162/jocn.2009.21287. [DOI] [PubMed] [Google Scholar]
- Lombardo M.V., Chakrabarti B., Bullmore E.T., Sadek S.A., Pasco G., Wheelwright S.J., Baron-Cohen S. Atypical neural self-representation in autism. Brain. 2010;133:611–624. doi: 10.1093/brain/awp306. [DOI] [PubMed] [Google Scholar]
- Lou H.C., Luber B., Crupain M., Keenan J.P., Nowak M., Kjaer T.W., Lisanby S.H. Parietal cortex and representation of the mental self. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(17):6827–6832. doi: 10.1073/pnas.0400049101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maddock R.J. The retrosplenial cortex and emotion: new insights from functional neuroimaging of the human brain. Trends in Neurosciences. 1999;22(7):310–316. doi: 10.1016/s0166-2236(98)01374-5. [DOI] [PubMed] [Google Scholar]
- Marcia J.E. Identity in adolescence. In: Adelson J., editor. vol. 5. Wiley; New York: 1980. pp. 145–160. (Handbook of Adolescent Psychology). [Google Scholar]
- Markus H. Self-schemata and processing information about the self. Journal of Personality and Social Psychology. 1977;35(2):63–78. [Google Scholar]
- Markus H.R., Kitayama S. Culture and the self: implications for cognition, emotion, and motivation. Psychological Review. 1991;98(2):224–253. [Google Scholar]
- Marsh, H.W., 1990. Self Description Questionnaire (SDQ) I: a theoretical and empirical basis for the measurement of multiple dimensions of preadolescent self-concept: a test manual and research monograph. Faculty of Education, University of Western Sydney, Macarthur, NSW Australia.
- Mason M.F., Norton M.I., Van Horn J.D., Wegner D.M., Grafton S.T., Macrae C.N. Wandering minds: the default network and stimulus-independent thought. Science. 2007;315(5810):393–395. doi: 10.1126/science.1131295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masten C.L., Colich N.L., Rudie J.D., Bookheimer S.Y., Eisenberger N.I., Dapretto M. An fMRI investigation of responses to peer rejection in adolescents with autism spectrum disorders. Developmental Cognitive Neuroscience. 2011;1(3):260–270. doi: 10.1016/j.dcn.2011.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masten C.L., Eisenberger N.I., Borofsky L.A., McNealy K., Pfeifer J.H., Dapretto M. Subgenual anterior cingulate responses to peer rejection: a marker of adolescents’ risk for depression. Development and Psychopathology. 2011;23(01):283–292. doi: 10.1017/S0954579410000799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masten C.L., Eisenberger N.I., Borofsky L.A., Pfeifer J.H., McNealy K., Mazziotta J.C., Dapretto M. Neural correlates of social exclusion during adolescence: understanding the distress of peer rejection. Social Cognitive and Affective Neuroscience. 2009;4(2):143–157. doi: 10.1093/scan/nsp007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPartland J.C., Crowley M.J., Perszyka D.R., Naplesa A.J., Mukerjia C.E., Wua J., Molfesea P., Bollinga D.Z., Pelphrey K.A., Mayes L.C. Temporal dynamics reveal atypical brain response to social exclusion in autism. Developmental Cognitive Neuroscience. 2011;1(3):271–279. doi: 10.1016/j.dcn.2011.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mead G.H. University of Chicago Press; Chicago, IL: 1934. Mind, Self and Society from the Standpoint of a Social Behaviorist. [Google Scholar]
- Menon V., Uddin L.Q. Saliency, switching, attention and control: a network model of insula function. Brain Structure & Function. 2010;214(5–6):655–667. doi: 10.1007/s00429-010-0262-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell J.P. Inferences about other minds. Philosophical Transactions of the Royal Society B. 2009;364:1309–1316. doi: 10.1098/rstb.2008.0318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell J.P. Activity in right temporo-parietal junction is not selective for theory-of-mind. Cerebral Cortex. 2008;18(2):262–271. doi: 10.1093/cercor/bhm051. [DOI] [PubMed] [Google Scholar]
- Mitchell J.P., Macrae C.N., Banaji M.R. Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron. 2006;50(4):655–663. doi: 10.1016/j.neuron.2006.03.040. [DOI] [PubMed] [Google Scholar]
- Mitchell J.P., Banaji M.R., Macrae C.N. The link between social cognition and self-referential thought in the medial prefrontal cortex. Journal of Cognitive Neuroscience. 2005;17(8):1306–1315. doi: 10.1162/0898929055002418. [DOI] [PubMed] [Google Scholar]
- Modinos G., Ormel J., Aleman A. Activation of anterior insula during self-reflection. PLoS One. 2009;4(2):e4618. doi: 10.1371/journal.pone.0004618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran J.M., Heatherton T.F., Kelley W.M. Modulation of cortical midline structures by implicit and explicit self-relevance evaluation. Social Neuroscience. 2009;4(3):197–211. doi: 10.1080/17470910802250519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran J.M., Macrae C.N., Heatherton T.F., Wyland C.L., Kelley W.M. Neuroanatomical evidence for distinct cognitive and affective components of self. Journal of Cognitive Neuroscience. 2006;18(9):1586–1594. doi: 10.1162/jocn.2006.18.9.1586. [DOI] [PubMed] [Google Scholar]
- Moriguchi Y., Ohnishi T., Lane R.D., Maeda M., Mori T., Nemoto K., Komaki G. Impaired self-awareness and theory of mind: an fMRI study of mentalizing in alexithymia. Neuroimage. 2006;32(3):1472–1482. doi: 10.1016/j.neuroimage.2006.04.186. [DOI] [PubMed] [Google Scholar]
- Nelson E.E., Leibenluft E., McClure E.B., Pine D.S. The social re-orientation of adolescence: a neuroscience perspective on the process and its relation to psychopathology. Psychological Medicine. 2005;35(02):163–174. doi: 10.1017/s0033291704003915. [DOI] [PubMed] [Google Scholar]
- Northoff G., Bermpohl F. Cortical midline structures and the self. Trends in Cognitive Sciences. 2004;8(3):102–107. doi: 10.1016/j.tics.2004.01.004. [DOI] [PubMed] [Google Scholar]
- Northoff G., Heinzel A., de Greck M., Bermpohl F., Dobrowolny H., Panksepp J. Self-referential processing in our brain—a meta-analysis of imaging studies on the self. Neuroimage. 2006;31(1):440–457. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Northoff G., Qin P., Feinberg T.E. Brain imaging of the self-conceptual, anatomical and methodological issues. Consciousness and Cognition. 2011;20:52–63. doi: 10.1016/j.concog.2010.09.011. [DOI] [PubMed] [Google Scholar]
- Ochsner K.N., Beer J.S., Robertson E.R., Cooper J.C., Gabrieli J.D., Kihsltrom J.F., D’Esposito M. The neural correlates of direct and reflected self-knowledge. Neuroimage. 2005;28(4):797–814. doi: 10.1016/j.neuroimage.2005.06.069. [DOI] [PubMed] [Google Scholar]
- Olson I.R., Plotzker A., Ezzyat Y. The enigmatic temporal pole: a review of findings on social and emotional processing. Brain. 2007;130:1718–1731. doi: 10.1093/brain/awm052. [DOI] [PubMed] [Google Scholar]
- Onoda K., Okamoto Y., Nakashima K., Nittono H., Ura M., Yamawaki S. Decreased ventral anterior cingulate cortex activity is associated with reduced social pain during emotional support. Social Neuroscience. 2009;4(5):443–454. doi: 10.1080/17470910902955884. [DOI] [PubMed] [Google Scholar]
- Onoda K., Okamoto Y., Nakashima K., Nittono H., Yoshimura S., Yamawaki S., Ura M. Effects of trait self-esteem on social pain by ostracism and activation of anterior cingulate cortex. Clinical Neurophysiology. 2010;121(7):e34. doi: 10.1093/scan/nsq002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostby Y., Tamnes C., Fjell A.M., Westlye L.T., Due-Tonnessen P., Walhovd K.B. Heterogeneity in subcortical brain development: a structural magnetic resonance imaging study of brain maturation from 8 to 30 years. Journal of Neuroscience. 2009;29:11772–11782. doi: 10.1523/JNEUROSCI.1242-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Packer D.J., Cunningham W.A. Neural correlates of reflection on goal states: the role of regulatory focus and temporal distance. Social Neuroscience. 2009;4(5):412–425. doi: 10.1080/17470910902750186. [DOI] [PubMed] [Google Scholar]
- Paus T. Mapping brain maturation and cognitive development during adolescence. Trends in Cognitive Sciences. 2005;9(2):60–68. doi: 10.1016/j.tics.2004.12.008. [DOI] [PubMed] [Google Scholar]
- Pelphrey K.A., Morris J.P., Mccarthy G. Grasping the intentions of others: the perceived intentionality of an action influences activity in the superior temporal sulcus during social perception. Journal of Cognitive Neuroscience. 2004;16(10):1706–1716. doi: 10.1162/0898929042947900. [DOI] [PubMed] [Google Scholar]
- Pfeifer J.H., Masten C.L., Moore W.E., III, Oswald T.M., Mazziotta J.C., Iacoboni M., Dapretto M. Entering adolescence: resistance to peer influence, risky behavior, and neural changes in emotion reactivity. Neuron. 2011;69(5):1029–1036. doi: 10.1016/j.neuron.2011.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeifer J.H., Lieberman M.D., Dapretto M. I know you are but what am I?!”: neural bases of self- and social knowledge retrieval in children and adults. Journal of Cognitive Neuroscience. 2007;19(8):1323–1337. doi: 10.1162/jocn.2007.19.8.1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeifer J.H., Masten C.L., Borofsky L.A., Dapretto M., Fuligni A.J., Lieberman M.D. Neural correlates of direct and reflected self-appraisals in adolescents and adults: when social perspective-taking informs self-perception. Child Development. 2009;80(4):1016–1038. doi: 10.1111/j.1467-8624.2009.01314.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan K.L., Wager T.D., Taylor S.F., Liberzon I. Functional neuroimaging studies of human emotions. CNS Spectrums. 2004;9:258–266. doi: 10.1017/s1092852900009196. [DOI] [PubMed] [Google Scholar]
- Powell L.J., Macrae C.N., Cloutier J., Metcalfe J., Mitchell J.P. Dissociable neural substrates for agentic versus conceptual representations of self. Journal of Cognitive Neuroscience. 2010;22(10):2186–2197. doi: 10.1162/jocn.2009.21368. [DOI] [PubMed] [Google Scholar]
- Prinstein M.J., Aikins J.W. Cognitive moderators of the longitudinal association between peer rejection and adolescent depressive symptoms. Journal of Abnormal Child Psychology. 2004;32(2):147–158. doi: 10.1023/b:jacp.0000019767.55592.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin P., Liu Y., Shi J., Wang Y., Duncan N., Gong Q., Weng X., Northoff G. Dissociation between anterior and posterior cortical regions during self-specificity and familiarity: a combined fMRI-meta-analytic study. Human Brain Mapping. 2011 doi: 10.1002/hbm.21201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin P., Northoff G. How is our self related to midline regions and the default-mode network? Neuroimage. 2011 doi: 10.1016/j.neuroimage.2011.05.028. [DOI] [PubMed] [Google Scholar]
- Ray R.D., Shelton A.L., Hollon N.G., Michel B.D., Frankel C.B., Gross J.J., Gabrieli J.D. Cognitive and neural development of individuated self-representation in children. Child Development. 2009;80(4):1232–1242. doi: 10.1111/j.1467-8624.2009.01327.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhodes M., Brickman D. Preschoolers’ responses to social comparisons involving relative failure. Psychological Science. 2008;19(10):969–972. doi: 10.1111/j.1467-9280.2008.02184.x. [DOI] [PubMed] [Google Scholar]
- Robins R.W., Tracy J.L., Trzesniewski K.H. Naturalizing the self. In: John O.P., Robins R.W., Pervin L.A., editors. Handbook of Personality: Theory and Research. 3rd ed. New York, NY; Guilford: 2008. pp. 375–398. [Google Scholar]
- Rogers T.B., Kuiper N.A., Kirker W.S. Self-reference and the encoding of personal information. Journal of Personality and Social Psychology. 1977;35(9):677–688. doi: 10.1037//0022-3514.35.9.677. [DOI] [PubMed] [Google Scholar]
- Rubin K., Fredstrom B., Bowker J. Future directions in … Friendship in childhood and early adolescence. Social Development. 2008;17(4):1085–1096. doi: 10.1111/j.1467-9507.2007.00445.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubin K.H., Bukowski W.M., Parker J.G. Peer interactions, relationships, and groups. In: Eisenberg N., Damon W., Lerner R.M., editors. Handbook of Child Psychology, vol. 3: Social, Emotional, and Personality Development. 6th ed. John Wiley & Sons; Hoboken, NJ: 2006. pp. 571–645. [Google Scholar]
- Ruble D.N., Alvarez J., Bachman M., Cameron J., Fuligni A., Coll C.G., Rhee E. The development of a sense of “we”: the emergence and implications of children's collective identity. In: Bennett M., Sani F., editors. The development of the social self. Psychology Press; Hove, UK: 2004. pp. 29–76. [Google Scholar]
- Ruble D.N., Frey K.S. Changing patterns of comparative behavior as skills are acquired: a functional model of self-evaluation. In: Suls J., Wills T., editors. Social Comparison: Contemporary Theory and Research. Erlbaum; Hillsdale, NJ: 1991. pp. 79–113. [Google Scholar]
- Ruby P., Decety J. What you believe versus what you think they believe: a neuroimaging study of conceptual perspective-taking. The European Journal of Neuroscience. 2003;17(11):2475–2480. doi: 10.1046/j.1460-9568.2003.02673.x. [DOI] [PubMed] [Google Scholar]
- Samson D., Apperly I.A., Chiavarino C., Humphreys G.W. Left temporoparietal junction is necessary for representing someone else's belief. Nature Neuroscience. 2004;7(5):499–500. doi: 10.1038/nn1223. [DOI] [PubMed] [Google Scholar]
- Saxe R., Kanwisher N. People thinking about thinking people: the role of the temporo-parietal junction in “theory of mind”. NeuroImage. 2003;19:1835–1842. doi: 10.1016/s1053-8119(03)00230-1. [DOI] [PubMed] [Google Scholar]
- Saxe R., Powell L.J. It's the thought that counts: specific brain regions for one component of theory of mind. Psychological Science. 2006;17(8):692–699. doi: 10.1111/j.1467-9280.2006.01768.x. [DOI] [PubMed] [Google Scholar]
- Saxe R.R., Whitfield-Gabrieli S., Scholz J., Pelphrey K.A. Brain regions for perceiving and reasoning about other people in school-aged children. Child Development. 2009;80:1197–1209. doi: 10.1111/j.1467-8624.2009.01325.x. [DOI] [PubMed] [Google Scholar]
- Sebastian C.L., Blakemore S.J. Understanding the neural response to social rejection in adolescents with autism spectrum disorders: a commentary on Masten et al., McPartland et al. and Bolling et al. Developmental Cognitive Neuroscience. 2011;1(3):256–259. doi: 10.1016/j.dcn.2011.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebastian C., Blakemore S.J., Charman T. Reactions to ostracism in adolescents with autism spectrum conditions. Journal of Autism and Developmental Disorders. 2009;39(8):1122–1130. doi: 10.1007/s10803-009-0725-4. [DOI] [PubMed] [Google Scholar]
- Sebastian C., Burnett S., Blakemore S.J. Development of the self-concept during adolescence. Trends in Cognitive Sciences. 2008;12(11):441–446. doi: 10.1016/j.tics.2008.07.008. [DOI] [PubMed] [Google Scholar]
- Sedikides C., Spencer S. Psychology Press; New York, NY: 2007. The Self: Frontiers in Social Psychology. [Google Scholar]
- Shamay-Tsoory S.G., Shur S., Barcai-Goodman L., Medlovich S., Harari H., Levkovitz Y. Dissociation of cognitive from affective components of theory of mind in schizophrenia. Psychiatry Research. 2007;149(1–3):11–23. doi: 10.1016/j.psychres.2005.10.018. [DOI] [PubMed] [Google Scholar]
- Shaw P., Kabani N.J., Lerch J.P., Eckstrand K., Lenroot R., Gogtay N., Greenstein D., Clasen L., Evans A., Rapoport J.L., Giedd J.N., Wise S.P. Neurodevelopmental trajectories of the human cerebral cortex. Journal of Neuroscience. 2008;28(14):3586–3594. doi: 10.1523/JNEUROSCI.5309-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville L.H., Heatherton T.F., Kelley W.M. Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Nature Neuroscience. 2006;9(8):1007–1008. doi: 10.1038/nn1728. [DOI] [PubMed] [Google Scholar]
- Sowell E.R., Thompson P.M., Leonard C.M., Welcome S.E., Kan E., Toga A.W. Longitudinal mapping of cortical thickness and brain growth in normal children. Journal of Neuroscience. 2004;24(38):8223–8231. doi: 10.1523/JNEUROSCI.1798-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spear L.P. Adolescent brain development and animal models. Annals of the New York Academy of Sciences. 2004;1021:23–26. doi: 10.1196/annals.1308.002. [DOI] [PubMed] [Google Scholar]
- Steele C.M., Aronson J. Stereotype threat and the intellectual test performance of African Americans. Journal of Personality and Social Psychology. 1995;69:797–811. doi: 10.1037//0022-3514.69.5.797. [DOI] [PubMed] [Google Scholar]
- Steinberg L. A social neuroscience perspective on adolescent risk-taking. Developmental Review. 2008;28(1):78–106. doi: 10.1016/j.dr.2007.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg L., Silverberg S.B. The vicissitudes of autonomy in early adolescence. Child Development. 1986;57:841–851. doi: 10.1111/j.1467-8624.1986.tb00250.x. [DOI] [PubMed] [Google Scholar]
- Stipek D., MacIver D. Developmental changes in children's assessment of intellectual competence. Child Development. 1989;60:521–538. [Google Scholar]
- Summerfield J.J., Hassabis D., Maguire E.A. Cortical midline involvement in autobiographical memory. Neuroimage. 2009;44(3):1188–1200. doi: 10.1016/j.neuroimage.2008.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Supekar K., Uddin L.Q., Prater K., Amin H., Greicius M.D., Menon V. Development of functional and structural connectivity within the default mode network in young children. Neuroimage. 2010;52(1):290–301. doi: 10.1016/j.neuroimage.2010.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svoboda E., McKinnon M.C., Levine B. The functional neuroanatomy of autobiographical memory: a meta-analysis. Neuropsychologia. 2006;44(12):2189–2208. doi: 10.1016/j.neuropsychologia.2006.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Symons C.S., Johnson B.T. The self-reference effect in memory: a meta-analysis. Psychological Bulletin. 1997;121(3):371–394. doi: 10.1037/0033-2909.121.3.371. [DOI] [PubMed] [Google Scholar]
- Tamir D.I., Mitchell J.P. Neural correlates of anchoring-and-adjustment during mentalizing. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(24):10827–10832. doi: 10.1073/pnas.1003242107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamnes C.K., Ostby Y., Fjell A.M., Westlye L.T., Due-Tønnessen P., Walhovd K.B. Brain maturation in adolescence and young adulthood: regional age-related changes in cortical thickness and white matter volume and microstructure. Cerebral Cortex. 2010;20(3):534–548. doi: 10.1093/cercor/bhp118. [DOI] [PubMed] [Google Scholar]
- van der Meer L., Costafreda S., Aleman A., David A.S. Self-Reflection and the brain: a theoretical review and meta-analysis of neuroimaging studies with implications for schizophrenia. Neuroscience and Biobehavioral Reviews. 2010;34(6):935–946. doi: 10.1016/j.neubiorev.2009.12.004. [DOI] [PubMed] [Google Scholar]
- Vann S.D., Aggleton J.P., Maguire E.A. What does the retrosplenial cortex do? Nature Reviews. Neuroscience. 2009;10:792–802. doi: 10.1038/nrn2733. [DOI] [PubMed] [Google Scholar]
- Vartanian L.R. Revisiting the imaginary audience and personal fable constructs of adolescent egocentrism: a conceptual review. Adolescence. 2000;35:639–662. [PubMed] [Google Scholar]
- Vogt B.A., Laureys S. Posterior cingulate, precuneal and retrosplenial cortices: cytology and components of the neural network correlates of consciousness. Progress in Brain Research. 2005;150:205–217. doi: 10.1016/S0079-6123(05)50015-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt B.A., Finch D.M., Olson C.R. Functional heterogeneity in cingulate cortex: the anterior executive and posterior evaluative regions. Cerebral Cortex. 1992;2(6):435–443. doi: 10.1093/cercor/2.6.435-a. [DOI] [PubMed] [Google Scholar]
- Yoshimura S., Okamoto Y., Onoda K., Matsunaga M., Ueda K., Suzuki S. Rostral anterior cingulate cortex activity mediates the relationship between the depressive symptoms and the medial prefrontal cortex activity. Journal of Affective Disorders. 2010;122(1–2):76–85. doi: 10.1016/j.jad.2009.06.017. [DOI] [PubMed] [Google Scholar]
- Zhu Y., Zhang L., Fan J., Han S. Neural basis of cultural influence on self-representation. Neuroimage. 2007;34(3):1310–1316. doi: 10.1016/j.neuroimage.2006.08.047. [DOI] [PubMed] [Google Scholar]