

Abstract
Edible marine algae, or seaweeds, are a rich source of several bioactive compounds including phytosterols, carotenoids, and polysaccharides. Over the last decades, seaweed-derived constituents turned out to not only reside in the systemic circulation, but are able to cross the blood-brain barrier to exert neuro-active functions both in homeostatic and pathological conditions. Therefore, seaweed-derived constituents have gained increasing interest for their neuro-immunomodulatory and neuroprotective properties, rendering them interesting candidates for the management of several neurodegenerative disorders. In particular seaweed-derived phytosterols gained interest for the treatment of neurodegenerative disorders as they potentiate neuroplasticity, enhance phagocytic clearance of neurotoxic peptides and have anti-inflammatory properties. Though, the anti-inflammatory and anti-oxidative properties of other constituents including carotenoids, phenols and polysaccharides have recently gained more interest. In this review, we provide an overview of a selection of the described neuro-active properties of seaweed-derived constituents with a focus on phytosterols.
Keywords: algae, carotenoids, neuro-active, phenols, phytosterols, polysaccharides, seaweed
Introduction
The marine ecosystem covers more than 70% of the world’s surface and hosts a wide variety of macro- and micro-organisms. Among these organisms, marine algae are of particular interest as they have been attributed medicinal properties due to their distinct nutritional composition. Marine algae are a rich source of bioactive compounds and secondary metabolites, such as peptides, lectins, carotenoids, polysaccharides, fatty acids, flavonoids, and phytosterols, distinguishing them from terrestrial plants (Ngo et al., 2012; Yende et al., 2014). Edible marine algae, also referred to as seaweed, can be classified into three main evolutionary and phylogenetically distinguishable classes that differ in nutritional and chemical composition: brown (Phaeophyceae), red (Rhodophyta) and green (Chlorophyta) algae (Yende et al., 2014). Since seaweeds require 0.1% photosynthetic light, they can be found in subtidal as well as intertidal waters. Potential medicinal properties of seaweeds have been explored for ages in traditional east Asian medicine, rendering China and Indonesia the two major cultivators and consumers of seaweed nowadays (Dhargalkar and Pereira, 2005; Yende et al., 2014). Interestingly, over the past decades, scientific interest in unravelling the exact pharmacological properties of marine algae and their constituents increased tremendously. Beneficial effects of seaweeds have been studied in the context of anti-inflammatory and anti-oxidant functions in divergent pathological conditions such as cancer, atherosclerosis, skin abnormalities, and neurodegeneration (Liu et al., 2012). In this review we discuss the neuroprotective properties of seaweed and its constituents, focussing on phytosterols in particular (Table 1 and Figure 1).
Table 1.
The neuro-active effects of seaweed derived constituents and their biological effects for the management of neurodegenerative disorders
| Constituent family | Individual constituent | Biological impact | Reference |
|---|---|---|---|
| Phytosterol | Fucosterol | Reduces Aβ plaque formation | Cárdenas-Aguayo Mdel et al., 2014 |
| Increases acetylcholine in brain | Andrade et al., 2013; Oh et al., 2018 | ||
| Counteracts memory deficits | Jun et al., 2016 | ||
| 24(S)-Saringosterol | Reduces Aβ plaque formation | Cárdenas-Aguayo Mdel et al., 2014 | |
| Increases Aβ clearance | Cárdenas-Aguayo Mdel et al., 2014 | ||
| Counteracts memory deficits | Jung et al., 2016; Alghazwi et al., 2019 | ||
| Sitosterol | Reduces Aβ plaque formation | Volkman et al., 1994; Burg et al., 2013; Wang et al., 2013 | |
| Anti-inflammatory | Liu et al., 2019 | ||
| Stigmasterol | Reduces Aβ plaque formation | Suganthy et al., 2010 | |
| Caretonoids | Beta-carotene | Anti-oxidant | Parajuli et al., 2013 |
| Fucoxanthin | Reduces Aβ plaque formation | Jung et al., 2016; Xiang et al., 2017; Jang et al., 2018 | |
| Anti-oxidant | Parajuli et al., 2013; Koivisto et al., 2014; Ruszkiewicz and Albrecht, 2015 | ||
| Counteracts memory deficits | Jung et al., 2016; Alghazwi et al., 2019 | ||
| Apo-9′- fucoxanthinone | Anti-inflammatory | Zhao et al., 2015 | |
| Phenols | Phloroglucinol | Anti-oxidant | Zhao et al., 2015 |
| Reduces Aβ-induced dendritic spine reduction | Zhao et al., 2015 | ||
| Counteracts memory deficits | Zhao et al., 2015 | ||
| Polysaccharides | Fucoidan | Counteracts memory deficits | Gao et al., 2012; Jung et al., 2016; Alghazwi et al., 2019 |
|
Sargassum fusiforme polysaccharide 65 |
Counteracts memory deficits | Gao et al., 2012 |
Aβ: Amyloid beta.
Figure 1.
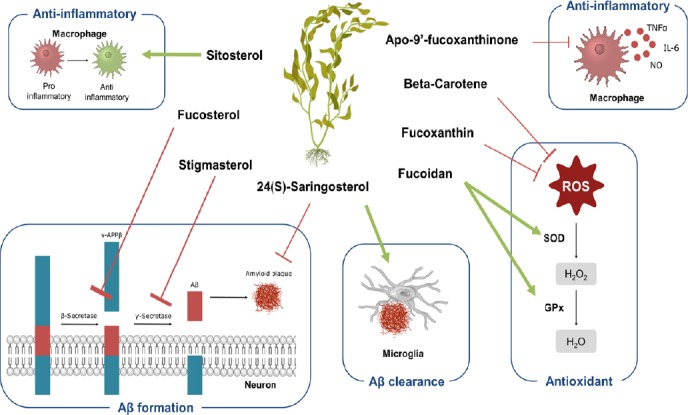
An overview of the key mechanism underlying bioactive properties of seaweed-derived constituents.
Phytosterols present in seaweed have mainly been described to be anti-inflammatory (sitosterol) and to interfere in neurotoxic peptide formation and clearance (fucosterol, stigmasterol, 24(S)-saringosterol). Especially anti-oxidative (beta-carotene, fucoxanthin) and anti-inflammatory (apo-9′-fucoxanthinone) properties have been attributed to carotenoids although their anti-inflammatory features. The polysaccharides in seaweed called fucoidan have been shown to be able to normalize ROS and GPx levels, essential for preventing ROS accumulation. Images were modified from Reactome icon library and Servier Medical Art, licensed under a Creative Common Attribution 3.0 Generic License (Sidiropoulos et al., 2017). Aβ: Amyloid beta; GPx: glutathione peroxidase; ROS: reactive oxygen species; SOD: superoxide dismutase.
Search Strategy and Selation Criteria
The approached search strategy was based on a focussed literature review. Databases used for the search included PubMed and Google Scholar. Search terms were combined in different manners and included: “seaweed constituents”, “phytosterols”, “neuromodulation”, “fucoidans”, “polyssaccharides”, “phenols”, “caretenoids”, “neuroinflammation”, “neurodegeneration”, “macroalgae”, “memory”, “cognition”.
Phytosterols as Neuro-Active Constituents
To date, phytosterols (e.g., plant sterols and plant stanols) have been enriched in functional foods to lower circulating levels of cholesterol and thereby reduce the risk of cardiovascular diseases. Yet, up to now, no trials have been conducted (with clinical endpoints) that evaluate the health effects of plant sterols or stanols in cardiovascular disorders. However, given that phytosterols would be capable of lowering cholesterol levels, they would not only reduce the risk of cardiovascular events, but could also be beneficial in central nervous system (CNS) disorders. While phytosterols can solely be derived from the diet, in the CNS, cholesterol is almost entirely synthesized locally (Hijmans et al., 2015; Vanmierlo et al., 2015). We and others have previously shown that phytosterols are able to cross the blood-brain barrier (BBB) and accumulate in the CNS where they can be incorporated in lipid rafts of CNS-resident cells and subsequently modify protein-protein interactions (Jansen et al., 2006; Vanmierlo et al., 2012). Phytosterols may therefore exert neuromodulatory properties. However, most studies thus far focussed on phytosterols present in terrestrial plants such as sitosterol, campesterol, and stigmasterol. Interestingly, seaweed is enriched in other specific phytosterols such as fucosterol and saringosterol which have been shown to exert neuromodulatory effects on synapse integrity and cognition, in health and disease. Importantly, mainly Phaeophyceae seaweed species could be considered a developable resource for phytosterols as all Rhodophyceae species are featured by cholesterol as dominant sterol (Al Easa et al., 1995). Chlorophyceae differs from the other classes because the dominant sterol seems to vary within the order or, for the same order, within the family (Al Easa et al., 1995). Phaeophyceae enriched with phytosterols may therefore offer a promising therapeutic strategy in the prevention or treatment of neurodegenerative disorders.
Neurodegenerative disorders are often featured by a wide range of diverse and intertwined neuro-inflammatory processes, leading to primary or secondary CNS damage. Neuro-inflammation is not only a central mediator of neurodegeneration, it also plays a pivotal role in permitting, facilitating, and orchestrating CNS repair. Therefore, modulating neuro-inflammatory processes can hold the key for treating multiple neurodegenerative disorders such as Alzheimer’s disease (AD) and multiple sclerosis (MS). Recently, it has been shown that the phytosterol sitosterol is able to modulate macrophage functions by augmenting the polarization of macrophages towards an anti-inflammatory phenotype, suggesting an interesting strategy for neurodegenerative disease management (Liu et al., 2019). Furthermore, emerging evidence indicates that phytosterols exert anti-inflammatory functions by activating nuclear liver X receptors (LXRs). LXRs are key transcription factors essential for modulating the sterol metabolism and homeostasis, and regulating immunomodulatory processes. The anti-inflammatory properties of phytosterols via LXRs can mainly be attributed to a mechanism of transrepression of toll-like receptor activation (Al Easa et al., 1995). Although phytosterols such as stigmasterol, fucosterol, brassicasterol, and sitosterol have been reported to activate LXRs, it has recently been shown that these phytosterols were unable to activate LXRs at physiological concentrations that can be reached through dietary supplementation (Bogie et al., 2019). Interestingly, the seaweed-derived phytosterol 24(S)-saringosterol was able to activate LXRs at these physiological concentrations (Chen et al., 2014; Bogie et al., 2019). In contrast with full synthetic LXR agonists, both pure 24(S)-saringosterol and a 24(S)-saringosterol-containing seaweed extract of Sargassum fusiforme did not induce typical LXR-dependent side effects (e.g. hypertriglyceridemia and hepatic steatosis) likely because 24(S)-saringosterol predominantly activates LXRβ (Plat et al., 2005; Bogie et al., 2019). For that reason, seaweed-derived 24(S)-saringosterol may provide an attractive strategy for ceasing and modulating neuro-inflammation in neurodegenerative disorders.
Synaptic loss is an early characteristic of aging and multiple neurodegenerative disorders (e.g., AD, schizophrenia, amyotrophic lateral sclerosis). During disease progression, synaptic loss is prominent and coincides with cognitive decline, sensory disturbances, and motor impairments. Newly formed synapses are often formed as a compensatory mechanism for coping with the pathological consequences (Jansen et al., 2012). In order to maintain synaptic integrity and to support synapse remodelling, distal axons require additional sterols that cannot be supplied sufficiently from the distant nerve cell body. LXRs act as cholesterol metabolite sensors that induce LXR-responsive genes required for maintaining cellular cholesterol turnover from astrocytes to neurons. Activation of LXRs may therefore enhance synaptic integrity and cognitive function. The LXR agonist T0901317 has been shown to stimulate the formation of new synapses in aged AD mice by enhancing cerebral cholesterol turnover (Vanmierlo et al., 2011). Seaweed-derived phytosterols may consequently also offer an interesting therapeutic strategy for enhancing synapse remodelling due to the activation of LXRs.
AD is featured by the accumulation of the toxic amyloid-β (Aβ) peptide. Although physiological levels of Aβ may be crucial for synaptic plasticity and neuronal survival, high concentrations eventually causes neurotoxicity and cell death (Cárdenas-Aguayo Mdel et al., 2014). The seaweed-derived fucosterol has been shown to be a non-competitive inhibitor of the β-secretase, an enzyme crucial in the formation of the toxic Aβ monomers (Jung et al., 2016). Furthermore, 24(S)-saringosterol has been found to reduce neuronal Aβ secretion while stimulating microglia-mediated clearance of Aβ. In line, dietary supplementation with either the brown seaweed Sargassum fusiforme, which contains high levels of 24(S)-saringosterol, or its lipid extract reduced Aβ plaque load and improved cognition in a mouse model for AD (Jung et al., 2016). However, besides 24(S)-saringosterol, also fucosterol, fucoidan, and fucoxanthin present in seaweed have been reported to counteract memory deficits and can therefore at least partially contribute to the therapeutic effects seen upon Sargassum fusiforme and its extract supplementation (Oh et al., 2018; Alghazwi et al., 2019). For instance, fucosterol showed to be not only a selective inhibitor of cholinesterases and therefore subsequently increased the levels of the neurotransmitter acetylcholine in the brain, but also reduced and prevented the formation of Aβ peptides (Suganthy et al., 2010; Andrade et al., 2013). Furthermore, fucosterol alleviates Aβ-induced endoplasmic reticulum (ER) stress and cognitive impairment suggesting that fucosterol can attenuate ER stress-induced ageing-associated cognitive decline (Oh et al., 2018). Since 24(S)-saringosterol is an auto-oxidation product of fucosterol, a contribution of 24(S)-saringosterol to these described health benefits of fucosterol cannot be ruled out yet ( Volkman et al., 1994).
Not only seaweed-derived phytosterols, but also phytosterols highly present in terrestrial plants have been reported to modulate AD pathology. Substitution of membrane cholesterol with sitosterol modulates the non-amyloidogenic processing of the amyloid precursor protein and may therefore reduce Aβ formation (Wang et al., 2013). In accordance with this, in vitro experiments showed that stigmasterol suppresses the activity of the lipid-raft associated γ-secretase thereby reducing Aβ generation and modulating AD pathogenesis (Burg et al., 2013; Koivisto et al., 2014).
Seaweed-derived phytosterols have been shown to be neuro-active modulators in the CNS. By ameliorating neuro-inflammation, stimulating synapse remodelling, diminishing neurotoxic protein accumulation and improving cognitive functioning in the CNS, seaweed-enriched phytosterols can be considered an interesting therapeutic strategy for the treatment of neurodegenerative disorders.
Non-Sterol Constituents as Neuro-Active Compounds
Besides phytosterols, numerous other seaweed-derived constituents have been reported to possess neuromodulating effects, rendering them interesting therapeutic molecules in strategies to modulate neuro-inflammatory and neuro-degenerative processes.
Carotenoids have often been investigated for their anti-oxidative properties. Although beta-carotene has been described as the most important anti-oxidant, fucoxanthin, highly present in Phaeophyceae, has gained interest for modulating CNS-related processes. In AD, accumulation of reactive oxygen species (ROS) leads to oxidative stress which negatively affects synaptic plasticity and causes neuronal death, leading to memory deficits and cognitive impairments. Upon arachidonic acid-induced oxidative damage, both beta-carotene and fucoxanthin were able to prevent ROS formation (Jang et al., 2018). Reducing ROS accumulation may therefore ameliorate the AD-associated deficiencies in memory and cognitive functions. Additionally, in cerebral cortical neurons, SH-SY5Y-, and PC12 cells, fucoxanthin inhibited the formation and aggregation of Aβ peptides, indicating a potential role for fucoxanthin in the treatment of AD (Zhao et al., 2015; Xiang et al., 2017; Alghazwi et al., 2019). Besides anti-oxidative properties, carotenoids have been shown to exhibit anti-inflammatory features such as inhibiting the inflammatory response in macrophages. Apo-9′-fucoxanthinone isolated from brown alga Sargassum muticum suppressed NO and prostaglandin E2 production in LPS-stimulated RAW 264.7 macrophages. Furthermore, in bone marrow-derived phagocytes, apo-9′-fucoxanthinone attenuated nuclear factor-kB-induced inducible nitric oxide synthase and cyclooxigensase-2 expression and suppresses CpG-induced production of pro-inflammatory cytokines interleukin-12 p40, interleukin-6 and tumor necrosis factor-α (Yang et al., 2015).
Next, seaweed-derived phenols have been described to possess neuroprotective properties. The polyphenol phloroglucinol, which is plentiful in the brown alga specie Ecklonia cava, was found to attenuate Aβ-induced ROS accumulation in the hippocampal neuronal cell line HT-22 (Yang et al., 2015). Potentially by acting as an anti-oxidant, phloroglucinol ameliorated the Aβ-induced reduction in dendritic spine density in primary rat hippocampal neurons and attenuated cognitive impairment in vivo in an animal model of AD (Yang et al., 2015).
The family of homo- and heteropolysaccharides called fucoidans, are widely distributed in brown algae. Biological effects of fucoidans have been extensively described and include mainly antitumor, antiviral and anticoagulant activities. Recently the neuro-active properties of fucoidans have been investigated in the context of AD. Fucoidans have been shown to normalize the levels of superoxide dismutase, glutathione peroxidase, choline acetyltransferase and acetylcholinesterase in the hippocampus of rats injected with Aβ (Gao et al., 2012). Cognitive impairments observed upon the Aβ infusion in these animals were ameliorated upon fucoidan treatment, suggesting a therapeutic potential of fucoidans in the context of AD (Gao et al., 2012). Additionally, Sargassum fusiforme polysaccharide 65 (SFPS65), a fucoidan, has been shown to ameliorate spatial learning and memory deficits in Swiss Albino mice (Hu et al., 2016). Further studies are however required to elucidate the underlying mechanisms of SFPS65-induced cognitive improvement.
Besides individual non-sterol constituents, the combination of multiple constituents to modulate cognitive processes, such as depression, anxiety and memory have been extensively investigated. A cross-sectional study conducted in Japan, where seaweed is largely being consumed, reported that depressive-like symptoms during pregnancy were reduced upon increased seaweed consumption, independent of the seaweed taxonomy (Miyake et al., 2014). Interestingly, a hydrophilic extract made from the green seaweed Ulva sp. reduced depressive-like behaviour in rodents. Although the exact underlying mechanisms remain unknown, the neuro-active compound is thought to be a family of sulphated polysaccharides, the ulvanse, which are highly present in the extract (Violle et al., 2018). The hypothesis of polysaccharides being involved in the observed anti-depressive actions is further strengthened by the finding that a water-soluble extract derived from the green seaweed Ulva Lactuca improved depressive symptoms in humans who experience a reduced feeling of motivation and pleasure (Allaert et al., 2018). A hexane extract of brown algae, containing a high amount of hydrocarbons, made from Sargassum plagyophylum acted as an antidepressant in a mouse model of despair (Haskell-Ramsay et al., 2018; Mesripour et al., 2019). However, not only hydrocarbons of brown seaweed but also plant phenols present in the methanolic extracts of Sargassum swartzii, Sargassum plagyophylum, Stoechospermum marginatum and Nizamuddinia zanardinii exhibited an antidepressant-like activity potentially by acting and modulating the mono-aminergic system in rodents (Siddiqui et al., 2017; Mesripour et al., 2019).
Since the glucose stores in the brain are limited, a steady supply of glucose is required to maintain optimal cognitive function (Haskell-Ramsay et al., 2018). Several studies have shown that consumption of foods low on the glycemic index (GI) scale, which tend to release glucose slowly and steadily, have beneficial effects on cognition compared to high GI foods (Philippou and Constantinou, 2014). The drawback of a low GI diet is the limiting types of food that can be consumed. An alternative approach to slow the carbohydrate absorption is inhibiting carbohydrate digestion (Haskell-Ramsay et al., 2018). Although the anti-diabetic drug acarbose inhibits the key enzymes of carbohydrate digestion and absorption, α-amylase and α-glucosidase (Roth et al., 2003), it induces side effects including gas accumulation and abdominal distention (Bischoff, 1994). A natural alternative to this anti-diabetic drug could be seaweed. The brown algae Ascophyllum nodosum (Apostolidis and Lee, 2010; Lordan et al., 2013; Pantidos et al., 2014) and Fucus vesiculosus (Lordan et al., 2013) have been shown to inhibit α-glucosidase to a greater extent than acarbose and α-amylase to a lesser extent (Haskell-Ramsay et al., 2018), an effect that has been shown to correlate with the seaweed phenol content (Apostolidis and Lee, 2010). Importantly, no side effects have been observed yet (Paradis et al., 2011). Seaweeds may therefore improve glucoregulation without side effects, which is suggested to have cognitive benefits. Nevertheless, it needs to be kept in mind that several studies showed the presence of toxic elements, such as radioactive isotopes and toxic metals (e.g., As, Cd, Pb and Al) in multiple seaweed species potentially entailing health risks (Rubio et al., 2017). Similar to the composition of bioactive constituents, the concentrations of toxic metals and traces differs upon seaweed taxonomy and cultivation conditions (Rubio et al., 2017). Although there are exceptions, toxic metals found in seaweed have not been reported to exceed the reference value (< 5%) upon consumption of 4 g seaweed per day, the average amount of seaweed the Japanese consume, indicating a negligible safety hazard (Rubio et al., 2017).
Concluding Remarks
The role of seaweed and its constituents as neuro-active compounds have gained tremendous interest in the last decade. Moreover, not only seaweed but also the multiple individual constituents can modify several CNS-related processes for modulating neuro-inflammation and neurodegeneration. Especially phytosterols have been extensively studied for their neuroprotective properties. Phytosterols can be found both in terrestrial plants and in multiple seaweed species with the most promising class being the Phaeophyceae algae. Due to the variety of biological actions, phytosterols offer a promising therapeutic strategy for ceasing the intertwined neuro-inflammatory and neurodegenerative processes featuring multiple CNS disorders. Sitosterol enhances the polarization of bone marrow-derived macrophages towards a more anti-inflammatory phenotype. Both fucosterol and stigmasterol have been described to reduce Aβ plaque formation by inhibiting β- and γ-secretases respectively. Interestingly, 24(S)-saringosterol not only reduces Aβ production but additionally increased microglial clearance of Aβ peptides, providing an attractive strategy for enhancing the clearance of neurotoxic peptides in general. Furthermore, in contrast to phytosterols common in the Western diet, 24(S)-saringosterol recently has been shown to be an LXRβ agonist at physiological concentrations. Due to the subsequent transrepression of toll-like receptors upon LXR activation, 24(S)-saringosterol possess a potential anti-inflammatory benefit. Furthermore, carotenoids, phenols and polysaccharides present in seaweed recently gained interest for their potential to treat CNS disorders. The carotenoids beta-carotene and fucoxanthin gained tremendous interest for their anti-oxidative properties, while apo-9′-fucoxanthinone reduced pro-inflammatory cytokine secretion by activated macrophages. Moreover, the seaweed-derived phenol, phloroglucinol, has been shown to harbour neuroprotective properties since it preserves dendritic spine density in vitro in primary hippocampal neurons cultured with Aβ. Finally, fucoidans, the polysaccharides widely distributed in algae, protect mice against cognitive decline, albeit the exact underlying mechanism remains to be elucidated.
By unravelling the neuro-active seaweed-derived constituents, the emerging field of seaweed constituent-based pharma gains more interest. For instance, semi-synthetic phytosterols are being developed and characterised for their potential to activate LXRs (Castro Navas et al., 2018). The semi-synthetic phytosterols have so far been described as positive LXR modulators rendering them interesting therapeutic strategies for modulating neuro-inflammation and enhancing neuroprotection (Castro Navas et al., 2018). As mentioned before, not only separate constituents but seaweed as a whole can be used for their neuroprotective properties. The usage of seaweed supplementation in the food industry combines the beneficial properties of multiple nutraceuticals for the application of disease prevention and disease management. Yet, the composition of bioactive constituents in the seaweed is highly variable and dependent on seasonality of sampling, taxonomic entity, location and related growth conditions. By identifying the most promising constituents, seaweed production and harvest can be optimised by utilizing more controlled systems before harvesting the seaweed. Nevertheless, the required amount of seaweed intake to reach neuro-active concentrations of the constituent of interest exceeds the dietary feasibility. Use of crude plant extracts with increased concentrations of its constituents may bypass this limitation and are easily implemented in the diet or as therapeutic intervention. Taken together, further identifying and unravelling the key neuro-active compounds in multiple seaweed species may lead to the development of nutraceutical treatment options for managing several neurodegenerative disorders.
Acknowledgments:
The authors sincerely thank Chuck Ungerman for editing the paper.
Footnotes
Conflicts of interest: We declare no conflicts of interest.
Financial support: NWO-TTW (Netherlands Organisation for Scientific Research), No. 16437; Alzheimer Nederland, No. WE.03-2018-06.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: NWO-TTW (Netherlands Organisation for Scientific Research), No. 16437; Alzheimer Nederland, No. WE.03-2018-06.
C-Editors: Zhao M, Li JY; T-Editor: Jia Y
References
- 1.Al Easa HS, Kornprobst JM, Rizk AM. Major sterol composition of some algae from Qatar. Phytochemistry. 1995;39:373–374. [Google Scholar]
- 2.Alghazwi M, Smid S, Musgrave I, Zhang W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta (Aβ1-42) toxicity and aggregation. Neurochem Int. 2019;124:215–224. doi: 10.1016/j.neuint.2019.01.010. [DOI] [PubMed] [Google Scholar]
- 3.Allaert FA, Demais H, Collén PN. A randomized controlled double-blind clinical trial comparing versus placebo the effect of an edible algal extract (Ulva Lactuca) on the component of depression in healthy volunteers with anhedonia. BMC Psychiatry. 2018;18:215–225. doi: 10.1186/s12888-018-1784-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Andrade PB, Barbosa M, Matos RP, Lopes G, Vinholes J, Mouga T, Valentão P. Valuable compounds in macroalgae extracts. Food Chem. 2013;138:1819–1828. doi: 10.1016/j.foodchem.2012.11.081. [DOI] [PubMed] [Google Scholar]
- 5.Apostolidis E, Lee CM. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J Food Sci. 2010;75:H97–102. doi: 10.1111/j.1750-3841.2010.01544.x. [DOI] [PubMed] [Google Scholar]
- 6.Bischoff H. Pharmacology of alpha-glucosidase inhibition. Eur J Clin Invest. 1994;24:3–10. [PubMed] [Google Scholar]
- 7.Bogie J, Hoeks C, Schepers M, Tiane A, Cuypers A, Leijten F, Chintapakorn Y, Suttiyut T, Pornpakakul S, Struik D, Kerksiek A, Liu HB, Hellings N, Martinez-Martinez P, Jonker JW, Dewachter I, Sijbrands E, Walter J, Hendriks J, Groen A, et al. Dietary sargassum fusiforme improves memory and reduces amyloid plaque load in an Alzheimer’s disease mouse model. Sci Rep. 2019;9:4908. doi: 10.1038/s41598-019-41399-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Burg VK, Grimm HS, Rothhaar TL, Grösgen S, Hundsdörfer B, Haupenthal VJ, Zimmer VC, Mett J, Weingärtner O, Laufs U, Broersen LM, Tanila H, Vanmierlo T, Lütjohann D, Hartmann T, Grimm MO. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J Neurosci. 2013;33:16072–16087. doi: 10.1523/JNEUROSCI.1506-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cárdenas-Aguayo Mdel C, Silva-Lucero Mdel C, Cortes-Ortiz M, Jiménez-Ramos B, Gómez-Virgilio L, Ramírez-Rodríguez G, Vera- Arroyo E, Fiorentino-Pérez R, García U, Luna-Muñoz J, Meraz-Ríos MA. Physiological role of amyloid beta in neural cells: the cellular trophic activity. Neurochemistry. 2014 doi: 10.5772/57398. [Google Scholar]
- 10.Castro Navas FF, Giorgi G, Maggioni D, Pacciarini M, Russo V, Marinozzi M. C24-hydroxylated stigmastane derivatives as Liver X Receptor agonists. Chem Phys Lipids. 2018;212:44–50. doi: 10.1016/j.chemphyslip.2018.01.005. [DOI] [PubMed] [Google Scholar]
- 11.Chen Z, Liu J, Fu Z, Ye C, Zhang R, Song Y, Zhang Y, Li H, Ying H, Liu H. 24(S)-Saringosterol from edible marine seaweed Sargassum fusiforme is a novel selective LXRβ agonist. J Agric Food Chem. 2014;62:6130–6137. doi: 10.1021/jf500083r. [DOI] [PubMed] [Google Scholar]
- 12.Dhargalkar VK, Pereira N. Seaweed: promising plant of the millennium. Sci Cult. 2005;71:60–66. [Google Scholar]
- 13.Gao Y, Li C, Yin J, Shen J, Wang H, Wu Y, Jin H. Fucoidan, a sulfated polysaccharide from brown algae, improves cognitive impairment induced by infusion of Aβ peptide in rats. Environ Toxicol Pharmacol. 2012;33:304–311. doi: 10.1016/j.etap.2011.12.022. [DOI] [PubMed] [Google Scholar]
- 14.Haskell-Ramsay CF, Jackson PA, Dodd FL, Forster JS, Bérubé J, Levinton C, Kennedy DO. Acute post-prandial cognitive effects of brown seaweed extract in humans. Nutrients. 2018;10:85–99. doi: 10.3390/nu10010085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hijmans BS, Tiemann CA, Grefhorst A, Boesjes M, van Dijk TH, Tietge UJ, Kuipers F, van Riel NA, Groen AK, Oosterveer MH. A systems biology approach reveals the physiological origin of hepatic steatosis induced by liver X receptor activation. FASEB J. 2015;29:1153–1164. doi: 10.1096/fj.14-254656. [DOI] [PubMed] [Google Scholar]
- 16.Hu P, Li Z, Chen M, Sun Z, Ling Y, Jiang J, Huang C. Structural elucidation and protective role of a polysaccharide from Sargassum fusiforme on ameliorating learning and memory deficiencies in mice. Carbohydr Polym. 2016;139:150–158. doi: 10.1016/j.carbpol.2015.12.019. [DOI] [PubMed] [Google Scholar]
- 17.Jang EJ, Kim SC, Lee JH, Lee JR, Kim IK, Baek SY, Kim YW. Fucoxanthin the constituent of Laminaria japonica, triggers AMPK-mediated cytoprotection and autophagy in hepatocytes under oxidative stress. BMC Complement Altern Med. 2018;18:97. doi: 10.1186/s12906-018-2164-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jansen D, Janssen CI, Vanmierlo T, Dederen PJ, van Rooij D, Zinnhardt B, Nobelen CL, Janssen AL, Hafkemeijer A, Mutsaers MP, Doedée AM, Kuipers AA, Broersen LM, Mulder M, Kiliaan AJ. Cholesterol and synaptic compensatory mechanisms in Alzheimer’s disease mice brain during aging. J Alzheimers Dis. 2012;31:813–826. doi: 10.3233/JAD-2012-120298. [DOI] [PubMed] [Google Scholar]
- 19.Jansen PJ, Lütjohann D, Abildayeva K, Vanmierlo T, Plösch T, Plat J, von Bergmann K, Groen AK, Ramaekers FC, Kuipers F, Mulder M. Dietary plant sterols accumulate in the brain. Biochim Biophys Acta. 2006;1761:445–453. doi: 10.1016/j.bbalip.2006.03.015. [DOI] [PubMed] [Google Scholar]
- 20.Jung HA, Ali MY, Choi RJ, Jeong HO, Chung HY, Choi JS. Kinetics and molecular docking studies of fucosterol and fucoxanthin, BACE1 inhibitors from brown algae Undaria pinnatifida and Ecklonia stolonifera. Food Chem Toxicol. 2016;89:104–111. doi: 10.1016/j.fct.2016.01.014. [DOI] [PubMed] [Google Scholar]
- 21.Koivisto H, Grimm MO, Rothhaar TL, Berkecz R, Lütjohann D D, Giniatullina R, Takalo M, Miettinen PO, Lahtinen HM, Giniatullin R, Penke B, Janáky T, Broersen LM, Hartmann T, Tanila H. Special lipid-based diets alleviate cognitive deficits in the APPswe/PS1dE9 transgenic mouse model of Alzheimer’s disease independent of brain amyloid deposition. J Nutr Biochem. 2014;25:157–169. doi: 10.1016/j.jnutbio.2013.09.015. [DOI] [PubMed] [Google Scholar]
- 22.Liu L, Heinrich M, Myers S, Dworjanyn SA. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in Traditional Chinese Medicine: a phytochemical and pharmacological review. J Ethnopharmacol. 2012;142:591–619. doi: 10.1016/j.jep.2012.05.046. [DOI] [PubMed] [Google Scholar]
- 23.Liu R, Hao D, Xu W, Li J, Li X, Shen D, Sheng K, Zhao L, Xu W, Gao Z, Zhao X, Liu Q, Zhang Y. β-Sitosterol modulates macrophage polarization and attenuates rheumatoid inflammation in mice. Pharm Biol. 2019;57:161–168. doi: 10.1080/13880209.2019.1577461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lordan S, Smyth TJ, Soler-Vila A, Stanton C, Ross RP. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013;141:2170–2176. doi: 10.1016/j.foodchem.2013.04.123. [DOI] [PubMed] [Google Scholar]
- 25.Mesripour A, Rabian N, Yegdaneh A. The effect of different partitions of seaweed Sargassum plagyophylum on depression behavior in mice model of despair. J Complement Integr Med. 2019 doi: 10.1515/jcim-2018-0207. doi: 10.1515/jcim-2018-0207. [DOI] [PubMed] [Google Scholar]
- 26.Miyake Y, Tanaka K, Okubo H, Sasaki S, Arakawa M. Seaweed consumption and prevalence of depressive symptoms during pregnancy in Japan: Baseline data from the Kyushu Okinawa Maternal and Child Health Study. BMC Pregnancy Childbirth. 2014;14:301. doi: 10.1186/1471-2393-14-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ngo DH, Vo T, Ngo DN, Wijesekara I, Kim SK. Biological activities and potential health benefits of bioactive peptides derived from marine organisms. Int J Biol Macromol. 2012;51:378–383. doi: 10.1016/j.ijbiomac.2012.06.001. [DOI] [PubMed] [Google Scholar]
- 28.Oh JH, Choi JS, Nam TJ. Fucosterol from an Edible Brown Alga Ecklonia stolonifera prevents soluble amyloid beta-induced cognitive dysfunction in aging rats. Mar Drugs. 2018 doi: 10.3390/md16100368. doi: 10.3390/md16100368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pantidos N, Boath A, Lund V, Conner S, McDougall GL. Phenolic-rich extracts from the edible seaweed, ascophyllum nodosum, inhibit α-amylase and α-glucosidase: Potential anti-hyperglycemic effects. J Funct Foods. 2014;10:201–209. [Google Scholar]
- 30.Paradis ME, Couture P, Lamarche B. A randomised crossover placebo-controlled trial investigating the effect of brown seaweed (Ascophyllum nodosum and Fucus vesiculosus) on postchallenge plasma glucose and insulin levels in men and women. Appl Physiol Nutr Metab. 2011;36:913–919. doi: 10.1139/h11-115. [DOI] [PubMed] [Google Scholar]
- 31.Parajuli B, Sonobe Y, Horiuchi H, Takeuchi H, Mizuno T, Suzumura A. Oligomeric amyloid β induces IL-1β processing via production of ROS: implication in Alzheimer’s disease. Cell Death Dis. 2013;4:e975. doi: 10.1038/cddis.2013.503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Philippou E, Constantinou M. The influence of glycemic index on cognitive functioning: a systematic review of the evidence. Adv Nutr. 2014;5:119–130. doi: 10.3945/an.113.004960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Plat J, Nichols JA, Mensink RP. Plant sterols and stanols: effects on mixed micellar composition and LXR (target gene) activation. J Lipid Res. 2005;46:2468–2476. doi: 10.1194/jlr.M500272-JLR200. [DOI] [PubMed] [Google Scholar]
- 34.Roth J, Ziak M, Zuber C. The role of glucosidase II and endomannosidase in glucose trimming of asparagine-linked oligosaccharides. Biochimie. 2003;85:287–294. doi: 10.1016/s0300-9084(03)00049-x. [DOI] [PubMed] [Google Scholar]
- 35.Rubio C, Napoleone G, Luis-González G, Gutiérrez AJ, González-Weller D, Hardisson A, Revert C. Metals in edible seaweed. Chemosphere. 2017;173:572–579. doi: 10.1016/j.chemosphere.2017.01.064. [DOI] [PubMed] [Google Scholar]
- 36.Ruszkiewicz J, Albrecht J. Changes in the mitochondrial antioxidant systems in neurodegenerative diseases and acute brain disorders. Neurochem Int. 2015;88:66–72. doi: 10.1016/j.neuint.2014.12.012. [DOI] [PubMed] [Google Scholar]
- 37.Siddiqui PJA, Khan A, Uddin N, Khaliq S, Rasheed M, Nawaz S, Hanif M, Dar A. Antidepressant-like deliverables from the sea: evidence on the efficacy of three different brown seaweeds via involvement of monoaminergic system. Biosci Biotechnol Biochem. 2017;81:1369–1378. doi: 10.1080/09168451.2017.1313697. [DOI] [PubMed] [Google Scholar]
- 38.Sidiropoulos K, Viteri G, Sevilla C, Jupe S, Webber M, Orlic-Milacic M, Jassal B, May B, Shamovsky V, Duenas C, Rothfels K, Matthews L, Song H, Stein L, Haw R, D’Eustachio P, Ping P, Hermjakob H, Fabregat A. Reactome enhanced pathway visualization. Bioinformatics. 2017;33:3461–3467. doi: 10.1093/bioinformatics/btx441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Suganthy N, Karutha Pandian S, Pandima Devi K. Neuroprotective effect of seaweeds inhabiting South Indian coastal area (Hare Island, Gulf of Mannar Marine Biosphere Reserve): Cholinesterase inhibitory effect of Hypnea valentiae and Ulva reticulata. Neurosci Lett. 2010;468:216–219. doi: 10.1016/j.neulet.2009.11.001. [DOI] [PubMed] [Google Scholar]
- 40.Vanmierlo T, Bogie JF, Mailleux J, Vanmol J, Lütjohann D, Mulder M, Hendriks JJ. Plant sterols: Friend or foe in CNS disorders? Prog Lipid Res. 2015;58:26–39. doi: 10.1016/j.plipres.2015.01.003. [DOI] [PubMed] [Google Scholar]
- 41.Vanmierlo T, Rutten K, Dederen J, Bloks VW, van Vark-van der Zee LC, Kuipers F, Kiliaan A, Blokland A, Sijbrands EJ, Steinbusch H, Prickaerts J, Lütjohann D, Mulder M. Liver X receptor activation restores memory in aged AD mice without reducing amyloid. Neurobiol Aging. 2011;32:1262–1272. doi: 10.1016/j.neurobiolaging.2009.07.005. [DOI] [PubMed] [Google Scholar]
- 42.Vanmierlo T, Weingärtner O, van der Pol S, Husche C, Kerksiek A, Friedrichs S, Sijbrands E, Steinbusch H, Grimm M, Hartmann T, Laufs U, Böhm M, de Vries HE, Mulder M, Lütjohann D. Dietary intake of plant sterols stably increases plant sterol levels in the murine brain. J Lipid Res. 2012;53:726–735. doi: 10.1194/jlr.M017244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Violle N, Rozan P, Demais H, Nyvall Collen P, Bisson JF. Evaluation of the antidepressant- and anxiolytic-like effects of a hydrophilic extract from the green seaweed Ulva sp. in rats. Nutr Neurosci. 2018;21:248–256. doi: 10.1080/1028415X.2016.1276704. [DOI] [PubMed] [Google Scholar]
- 44.Volkman JK, Barrett SM, Dunstan GA, Jeffrey SM. Sterol biomarkers for microalgae from the green algal class Prasinophyceae. Org Geochem. 1994;21:1211–1218. [Google Scholar]
- 45.Wang J, Wu F, Shi C. Substitution of membrane cholesterol with β-sitosterol promotes nonamyloidogenic cleavage of endogenous amyloid precursor protein. Neuroscience. 2013;247:227–233. doi: 10.1016/j.neuroscience.2013.05.022. [DOI] [PubMed] [Google Scholar]
- 46.Xiang S, Liu F, Lin J, Chen H, Huang C, Chen L, Zhou Y, Ye L, Zhang K, Jin J, Zhen J, Wang C, He S, Wang Q, Cui W, Zhang J. Fucoxanthin inhibits β-amyloid assembly and attenuates β-amyloid oligomer-induced cognitive impairments. J Agric Food Chem. 2017;65:4092–4102. doi: 10.1021/acs.jafc.7b00805. [DOI] [PubMed] [Google Scholar]
- 47.Yang EJ, Ahn S, Ryu J, Choi MS, Choi S, Chong YH, Hyun JW, Chang MJ, Kim HS. Phloroglucinol attenuates the cognitive deficits of the 5XFAD mouse model of Alzheimer’s disease. PLoS One. 2015;10:e0135686. doi: 10.1371/journal.pone.0135686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yende SR, Harle UN, Chaugule BB. Chaugule, Therapeutic potential and health benefits of Sargassum species. Pharmacogn Rev. 2014;8:1–7. doi: 10.4103/0973-7847.125514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhao X, Zhang SP, An CN, Zhang HN, Sun Y, Li YM, Pu XP. Neuroprotective effect of fucoxanthin on β-amyloid-induced cell death. Zhonguo Yaoxue Zazhi. 2015;24:467–474. [Google Scholar]