

Abstract
This review intends to rationalize the knowledge related to the aroma of grapes and to the aroma of wine with specific origin in molecules formed in grapes. The actual flavor of grapes is formed by the few free aroma molecules already found in the pulp and in the skin, plus by those aroma molecules quickly formed by enzymatic/catalytic reactions. The review covers key aroma components of aromatic grapes, raisins and raisinized grapes, and the aroma components responsible from green and vegetal notes. This knowledge is used to explain the flavor properties of neutral grapes. The aroma potential of grape is the consequence of five different systems/pools of specific aroma precursors that during fermentation and/or aging, release wine varietal aroma. In total, 27 relevant wine aroma compounds can be considered that proceed from grape specific precursors. Some of them are immediately formed during fermentation, while some others require long aging time to accumulate. Precursors are glycosides, glutathionyl and cysteinyl conjugates, and other non-volatile molecules.
Keywords: wine aging, glycosides, glutathione, mercaptans, terpenols, norisoprenoids, volatile phenols, vanillin
1. Introduction
Winemaking grapes are quite unique fruits because they are grown not to be immediately consumed, but to make wine with them. From this point of view, the study of grape aroma cannot be limited to the pool of molecules directly responsible for the odors and flavors of grape and grape juice but has also to include those other chemical structures that, more or less directly, are specific precursors of relevant wine aroma molecules. This task began more than 40 years ago when French and Australian researchers reported the existence of glycosides and other precursors of linalool [1,2]. The task, however, has proved to be extremely difficult due to many factors, such as the chemical and biochemical complexity of the precursor systems, the long times required to see aging effects in wine, or the analytical challenges associated to obtaining reliable representations of wine sensory properties from analytical data [3,4]. The truth is that nowadays, in spite of many significant advances, there are not accurate criteria or accepted methods able to provide a reliable assessment of the grape aroma potential, except perhaps for aromatic varietals such as Muscat or Gewürztraminer. This is a bit of a paradox; the grape genome was untangled more than 10 years ago [5], but yet, we do not have a clear understanding of all the grape metabolites which will ultimately contribute to the aromatic sensory properties of wine.
The reasons for this rather sluggish progress in linking grape molecular systems and wine odorants can be better understood with the help of the schema in Figure 1. The schema shows that grape contains at least seven different systems or pools of aroma precursors. Two out of the seven have relevance in grape but are not particularly important in wine aroma (the Strecker amino acid system and the fatty acid/peroxygenase system), while the other five play essential roles in the development of wine varietal aroma during wine aging, and/or in the development of wine flavor notes. If at the light of our present understanding, the different analytical strategies and concepts applied along the years for the study of grape aroma precursors are revisited, it will become evident that they provide information covering a rather limited fraction of wine varietal aroma. In fact, the general strategy followed to analyze grape glycosidic precursors deals with precursors belonging to just one or two out of the five pools. This is not to blame previous research, most of which was brilliantly carried out by pioneers, but to acknowledge the difficulties of the study, which with the limited analytical tools available in the 1980s, 1990s, and even the 2000s, hardly could have been done any better.
Figure 1.

Scheme showing the main systems/pools in grape of specific precursors of aroma molecules and their involvement in the development of wine varietal aroma and flavor.
The two first systematic approaches developed to study grape aroma precursors, which are yet the basis of the methods in use at present, were developed by Patrick Williams and coworkers in Australia [6] and by Ziya Gunata and coworkers in Montpellier [7]. In these approaches, grape glycosil aroma precursors are extracted from grape must or macerated grape solids with C18 or with XAD-2 polymeric sorbents, respectively. Much later, the use of more advanced polymeric sorbents providing a wider extraction of precursors was proposed [8], although as noted by Hampel et al., no sorbent was effective for all glycosides [9]. The glycosidic fractions are further hydrolyzed well by acid hydrolysis and enzymatic treatment [6], and well exclusively by enzymatic treatment [7].
The advantage of enzymatic treatment is that, in comparison to acid hydrolysis, it provides a relatively unbiased composition of the aglycones present in the extract, as far as the correct type of enzyme is used [9]. Under this approach the aroma of grape is divided into the free and the bound fractions [10,11]. Its major disadvantage is that, in many cases, the aglycone is not an odorant relevant for wine aroma, but an aroma-worthless volatile compound such as benzyl alcohol or an odorless precursor that only after a series of reactions, which can take a long time, will form the odorant. Attending to the scheme shown in Figure 1, enzymatic hydrolysis provides a useful estimate of wine aroma molecules derived from the pool of “glycosides of aroma molecules”, but not of those derived from the pool of “glycosides of precursors of aroma molecules” or from the other pools of precursors. Unfortunately, only some terpenols have direct glycosides, while important wine aroma molecules derived from norisoprenoids or grape phenols have not many direct glycosides. Consequently, enzymatic hydrolysis can assess the aroma potential of Muscat and other terpenol-related varietals, but not of “neutral varieties” [12].
For neutral varieties things are slightly more complicated, since the precursors of some relevant aroma molecules, such as norisoprenoids, require acid catalysis to undergo the chemical rearrangement processes through which the odorant is formed. Inevitably, this implies that labile aroma molecules, such as linalool and geraniol, will be degraded [9]. This problem is more evident in the many assays in which acid hydrolysis is carried out at high temperatures (100 °C). Under these conditions, as will be later detailed, there is a strong degradation of many relevant wine aroma molecules. Best results from the sensory point of view were obtained in the few studies in which acid hydrolysis was carried out at mild temperatures (45–50 °C). Only in these conditions the aroma hydrolysates obtained were able to induce significant sensory changes in wine [13,14]. However, some of the aroma descriptors developed during the hydrolysis, such as honey or tea [13], suggest that oxidation and thermal degradation processes are taking place under those conditions. These observations may question whether those hydrolysates are genuine representatives of wine varietal aroma and hence of grape potential aroma.
A recently presented strategy tries to sort out these limitations by using a most powerful extraction strategy, carrying out the hydrolysis in strict anoxia and in the presence of grape polyphenols [15]. Grape polyphenols and most specific aroma precursors, except those of dimethyl sulfide (DMS), are coextracted from dearomatized “mistellas” and reconstituted in synthetic wine. Under these conditions, hydrolysates obtained after 24 h display sensory profiles congruent with unoxidized wine odor nuances and specific for the grape variety (Alegre et al., in preparation). The approach is promising, yet requires proper validation.
In the present review we will make a distinction between the actual and the potential aromas of grapes, even if in many instances the boundaries between both categories are relatively blurred. Actual grape aroma integrates those aroma molecules and chemical systems responsible for the aromatic sensory properties (odor and flavor) of grapes and grape juices. On the other hand, potential grape aroma refers to the different grape molecules and grape chemical systems that are specific precursors of relevant wine odorants.
2. The Actual Aroma of Grapes and Musts
The concept of actual aroma includes not only the aroma molecules found as free forms in the grape or must, but also those others formed in the short time span in which grapes of grape juices are kept in the mouth during mastication and salivation. This can be better understood with the help of the scheme shown in Figure 2. In the figure, the precursor systems able to quickly release free aroma molecules are linked by discontinuous arrows to the “grape free aroma molecules pool”.
Figure 2.

Scheme showing the different aroma precursor systems/pools in grape and their relationship with the fraction of free aroma molecules which will ultimately be responsible for the odor and flavor of grapes and musts.
In common with many fruits, the actual aroma of grapes involves compounds in three related categories:
Free aroma, which refers to the aroma molecules found as such in the pulp and skin of the fruit, the grape in our case;
Aroma molecules formed by nearly instantaneous enzymatic/catalytical processes triggered during the disruption of fruit tissues [16,17];
Aroma molecules formed in the buccal cavity by the action of salivary or bacterial enzymes [18,19,20].
Compounds in the second category include a number of aldehydes, ketones and alcohols formed by peroxidation of fatty acids. Numerically the most abundant are compounds with six carbon atoms, so that compounds in this category are often named as C6-compounds [21,22]. It should be noted, however, that some powerful aroma compounds with a different number of carbon atoms can be also formed through this way, such as E-2-nonenal [23] or (E,Z)-2,6-nonadienal [24,25]. These powerful aroma compounds have much smaller odor thresholds, so that some of the green odors usually attributed to C6 aldehydes and alcohols could be in fact be caused by C9 aromas.
Compounds in the third category derive from two different types of precursors. It has been demonstrated that glutathionyl and cysteinyl precursors, which are odorless cysteine-S-conjugates, can release the aromatic thiol by the action of buccal microbiota [18]. The release takes 20–30 s and can induce a perception lasting for up to 3 min, which supports the idea that these precursors can have an outstanding role in the persistence of grape and wine aroma. In the case of glycosidic precursors of aroma molecules, it has been demonstrated that oral bacteria are able to hydrolyze glycosidic precursors, releasing an array of volatiles [19]. In the particular case of glycoconjugates of the volatile phenols derived from smoke exposure, it was demonstrated that enzymes in saliva are able to release enough volatiles to create a sensory perception [26]. In the case of glycoconjugates extracted from Gewürztraminer grapes, sensory effects in the mouth were only evident when tested at 5-times wine concentration and in the absence of wine volatiles, which may call into question the sensory relevance of the aroma volatiles released from those glycosides in the mouth [20]. However, all these in-mouth effects are highly variable between individuals, so that for some sensitive individuals they may have an effect. Additionally, a recent report [27] has revealed that glycosides extracted from the grape marc added to the must produce wines with longer aftertaste. This observation does not unequivocally demonstrate the role of glycosidic precursors in aftertaste but supports their importance on wine flavor.
In the case of grapes, the free aroma fraction is very small in most varieties, in agreement with the fact that most of them display weak and rather neutral odors and flavors. This should not be a surprise, since grapes are fruits extremely rich in water and do not contain special cellular or vacuolar structures in which nonpolar molecules such as aroma compounds can be safely stored. Hydrophobic molecules, including many aroma components, are stabilized in the pulp and skin by forming covalent bonds with polar molecules, such as sugars or amino acids, constituting fractions of specific aroma precursors which will be extensively discussed later on.
In the present section we will focus on the aroma molecules which can be found as free molecules in grapes or musts and which are likely contributors of sensory notes. The section will be divided into four subsections. The first one addresses the aroma molecules of those types of grapes showing clear and distinctive aromas, such as Muscat, Gewurztraminer and some hybrids between Vitis vinifera and labruscana. The second subsection summarizes the knowledge about the aroma molecules of raisins. The third subsection considers aroma molecules responsible for green, herbaceous, and vegetal aroma, many of which form a kind of common background in grapes of all types. The fourth and last subsection will briefly discuss about the aroma molecules responsible for the aroma characteristics of neutral grapes.
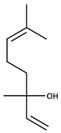
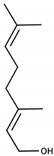
2.1. Key Aroma Compounds of Aromatic Grapes
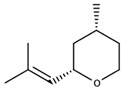
Among Vitis vinifera grape varieties, only those of the Muscat group have distinctive aroma and flavor [28]. These grapes contain important amounts of terpenols at levels above the odor threshold, as detailed in Table 1. The most important aroma compounds are linalool and geraniol, although those grapes also contain important levels of citral, citronellol, nerol, and α-terpineol. Another component, which attending to recent reports can be present at sensorily relevant levels, is geranic acid [29,30,31]. Muscat grapes can contain more than 5 mg/kg of these aroma compounds, in clear contrast to non-Muscat varieties which contain in general less than 0.5 mg/kg of these aroma compounds. Another relevant terpenic aroma compound is (Z)-rose oxide, which is responsible for the litchi-like or rose-like characteristic aroma of Gewürztraminer wines [32,33]. Rose oxide is a powerful aroma compound with an odor threshold one order of magnitude smaller than that of linalool [34]. It has been quantified in grapes from the Traminer family at 18 μg/L [35]. It has been recently found also in Muscat grapes [36] and a recent report even suggests that the intensity of Muscat aroma in grapes is strongly correlated to the presence of this molecule [37]. Its aromatic relevance in some aromatic grapes could have been underestimated simply because this molecule has been quantified in a reduced number of cases. Semiquantitative data provided by a recent report suggest that this aroma compound could be in fact relevant in the aroma profile not only of Gewurztraminer and Muscat, but also in Traminette and even in Riesling [38].
Table 1.
Structures, odor properties, and occurrence of the key odorants of aromatic grapes.
| Compound | Structure | Grape | Odor Description | Threshold | Range of Occurrence in Grapes |
|---|---|---|---|---|---|
| Linalool |
|
Muscat | Hyacinth, Muscat wine | 6 μg/L [48] | 0.06–1.5 mg/L [28] |
| Geraniol |
|
Muscat | Citrus, rose | 40 μg/L [49] | 0.09–1.1 mg/L [28] |
| (Z)-Rose oxide |
|
Traminer | Rose, litchi | 0.5 (l form) or 50 μg/L (d form) [34] | 7–29 μg/L [35] |
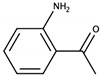
| o-Aminoacetophenone |
|
Concord | Sweet, caramel | 0.2 μg/L [50] | 10–20 μg/L [46] |
| Methyl anthranilate |
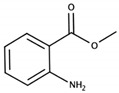
|
Concord | Orangine, sweet | 3 μg/L [51] | 0.8 mg/kg [39] 0.5–6 mg/kg [52] |
Among non Vitis vinifera cultivars there are some varieties known by their specific aroma. One of them is Vitis labruscana Bailey cv. Concord which contains at least four different aroma molecules at sensory-relevant levels. These are o-aminoacetophenone, methylfuraneol, methyl anthranilate, and furaneol [39,40]. Two of them, methyl anthranilate and o-aminoacetophenone, are involved in the characteristic “foxy” aroma of the variety (see Table 1). Remarkably, methyl anthranilate was identified as early as 1926 [41], while o-aminoacetophenone was identified in the 1980s [42]. Methyl anthranilate has been identified as one of the aroma components able to attract flies [43]. For its part, o-aminoacetophenone can eventually also develop in wines of Vitis vinifera varieties (mostly of German origin) where it causes a defect known as “untypical aging note” [44]. Furaneol (2,5-dimethyl-4-hydroxy-3(2H)-furanone) has also been identified as key odorant of muscadine (Vitis rotundifolia Michx), together with o-aminoacetophenone [45,46]. The potency and particular sensory characteristics of these aroma compounds make it so that those grapes are much appreciated as table grapes and also for making aromatic grape juice, but they are regarded as nonappropriate for making wine. In a recent paper, Wu et al. [29] study the aroma composition of 20 table grapes, 12 of which are hybrids between V. vinifera and V. labrusca. Interestingly, five of the hybrids showed strawberry aroma and four others foxy aroma, which suggests that the former contain large amounts of furaneol and of methylfuraneol, while the latter may contain methyl anthranilate and o-aminoacetophenone. Unfortunately, and this is a limitation of most recent studies carried out on grapes, all these polar and not very volatile aroma compounds cannot be easily determined by headspace solid phase microextraction (HS-SPME), which has become a kind of standard technique for the analysis of grape aroma. This explains the controversy about the implication of ethyl esters on the strawberry aroma of some of those grapes [29,47] and should warn about the risk of extracting conclusions about the sensory implications of analytical data when known essential aroma compounds have not been quantified: even if the profile of the volatiles quantified by HS-SPME is enough to obtain a highly satisfactory varietal differentiation, this does not mean that the varietal aroma profile is perfectly defined.
Some of these compounds can be also present, albeit at much smaller levels, in grapes from neutral varieties. For instance, furaneol was proposed time ago as a potential marker for the detection of forbidden hybrids (Vitis vinifera × non-vinifera) for making wine [53]. Furaneol can be present at levels above 1 mg/kg in non-viniferas, while it rarely will reach 0.05 mg/kg in vinifera wines [54].
Recent and quite extensive reports from Chinese researchers have confirmed that some table grapes contain a range of ethyl esters at concentrations above their thresholds [29,30,31,47]. These aroma compounds are found mainly as free compounds in the pulp and, in terms of odor activity values (OAVs), can amount to a relevant fraction of the odorants present in the grape. This fraction seems to be particularly high in “foxy” aroma grapes derived from V. labruscana [29] and also in some unfamiliar table-grapes [30]. For instance, in the cultivar “Honey Black”, these compounds account for more than 70% of the total OAV measured by the researchers. It is not clear, however, whether this aromatic power translates into specific aroma nuances. Ethyl esters are relatively ubiquitous aroma compounds and are normal constituents of the aroma of many fruits, so that they will likely contribute to generic fruity aroma nuances to grape flavor.
2.2. Key Aroma Compounds of Raisins and of “Raisinized” Grapes
Another type of grapes with intense and specific aroma and flavors are raisins, which are grapes naturally dried under the sun or by different artificial means. Some raisins are used to make dessert wines, such as Pedro Ximenez, and are, therefore, genuine winemaking grapes. Many other raisins are produced to be directly consumed as sweet grapes and confectionery ingredients. Their aroma composition is, however, of general interest for the wine industry, since winemaking grapes can undergo naturally spontaneous drying processes on the vine (raisining, as indicated in Figure 2) as the consequence of different maturation problems. As those problems become more frequent due to climate change, unwanted raisining will be an emerging problem in many vine growing areas [55]. In the event these raisinized grapes are fermented together with healthy grapes, the wine will eventually develop raisin and prune notes.
Raisins can contain different groups of key aroma compounds [23,56,57,58], which explains the high diversity of aroma nuances observed between different types of raisins and also supports the general complexity of raisin aroma. Leaving aside key terpenic odorants, such as linalool, geraniol, and rose oxide, which come directly from the fresh grape in the frequent case in which the raisins are made of aromatic grapes (Muscat and derivatives, Traminer and derivatives, Pedro Ximenez) [23], raisins can contain relevant odorants or groups of odorants produced or accumulated well during the own raisining process, during the last stages of grape maturation, and even during the storage of raisins.
The first aroma compound particularly relevant in raisins is β-damascenone, which seems to be a quite ubiquitous and key aroma component of many sun-dried grapes [23,57] and of the wines made with them [59]. β-Damascenone is a norisoprenoid derived from the degradation of carotenoids. It has a quite low odor threshold, close to the ng/L, and an odor reminding of prunes or overmatured plums. As will be later discussed, this molecule plays also a relevant role in the flavor of neutral grapes and in the sensory properties of wines. Its structure and odor properties, together with those of other important aroma compounds from the same family, can be seen in Table 2. Different studies confirm that β-damascenone tends to accumulate in grapes in the last periods of maturation [60,61,62,63], particularly in the case of late season berry dehydration (or raisining) [64,65], during the storage of the raisins [58], or even during the aging of wines made with raisins [66]. Its levels, however, have no clear relationship with sun exposure on the vine [67,68]. β-Damascenone plays an outstanding role in the fruity aroma characteristics of wine. At low concentrations it acts as aroma enhancer [69] but at levels above 2–3 μg/L it can induce the perception of overmatured fruit, particularly if methional is also present [70].
Table 2.
Structures, odor properties, and occurrence of norisoprenoids found above their threshold value in wine.
| Compound | Structure | Odor Descriptor | Threshold in Wine | Range of Occurrence in Wine |
|---|---|---|---|---|
| β-Damascenone |
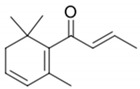
|
Plum, cooked apple | 50 ng/L [32] | n.d. to 10.5 μg/L [71] |
| β-Ionone |
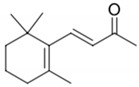
|
Violet, woody | 90 ng/L [72] | n.d. to 1.2 μg/L [71] |
| TDN |
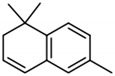
|
Kerosene-like | 2 μg/L [73] | n.d. to 255 μg/L [74] |
| TPB |
|
Green, cut-grass | 40 ng/L [75] | n.d. to 233 ng/L [76] |
n.d.: Not detected.
The second group of powerful aroma compounds likely formed during grape dehydration are Strecker aldehydes derived from the Strecker degradation of amino acids. The most relevant from the aromatic point of view are phenylacetaldehyde (honey odor) and methional (raw potato odor), which are important aroma constituents of Pedro Ximenez wines made with sun-dried grapes [59]. Phenylacetaldehyde has been also found at levels well above its threshold in raisins [23,57]. The formation of these compounds can be particularly intense in the frequent case in which dehydration occurs after or during the attack of the fungus Botrytis cinerea [77,78,79], which explains the high levels of both compounds in wines from Sauternes. These compounds arise by the reaction of the amino acid precursor with a quinone or other α-dicarbonyl. In grapes, the major source of dicarbonyls is the quinones from oxidizing polyphenols. The oxidation can begin by photoactivation (normal raisining) or by enzymatic action, which will be particularly intense in the presence of the powerful phenol-oxidase from Botrytis (laccase). Recent results suggest that the formation may take place after some time of the solar irradiation, since in a study of the effects of the storage on raisin aroma, phenylacetaldehyde strongly accumulated only after 12 months of storage of sun-dried raisins but not in air-dried raisins [58]. These compounds are relatively difficult to analyze because of their high activity towards many chromatographic phases and because of the adducts they form with SO2. This explains why many reports fail in their detection, particularly in the case of methional, so that their importance may be underestimated.
The third group of aroma components of raisins is formed by two odorous lactones derived from grape lipids, namely γ-nonalactone and massoia lactone. γ-Nonalactone is a well-known wine component [72] of coconut aroma whose levels in wine were first tentatively related to the development of prune character by Pons et al. [80]. The contribution to dry-fruit aroma has been recently shown to happen by perceptual interaction with furaneol and homofuraneol [81]. Its levels are increased in wines made from grapes affected by Botrytis [78,79], in late harvest wines [82], and in wines made from raisinized grapes [64]. Remarkably, γ-nonalactone is also a constituent of raisins [57]; its level and fate much depends on the type of grape, its pretreatment, time of storage and packaging material [58,83]. Massoia lactone (5,6-dihydro-6-pentyl-2H-pyran-2-one) has been recently identified as key aroma component in musts showing clear over-ripe characters of cooked plums and dried figs [84]. Both components, γ-nonalactone and massoia lactone, have been found at higher levels in wines made from partially dehydrated (raisinized) Shiraz grapes [65]. Massoia lactone has been also identified in the hydrolysates of phenolic and aromatic fractions (PAFs) extracted from grapes [15].
The fourth group of relevant aroma compounds formed during grape dehydration are some pyrazines with roasted aromas derived from Maillard reactions between sugars and amino acids. Wang et al. [23,57] identified at sensory-relevant levels 3-ethyl-2,5-dimethyl pyrazine and 2,6-diethylpyrazine. Both compounds were found to increase with storage of raisins [58].
Finally, and in common with any kind of grapes, raisins contain a relatively wide array of aldehydes and alcohols derived from the oxidation of grape fatty acids (FAOs). According to Wang et al. [23,57], pentanal, hexanal, heptanal, nonanal, decanal, (E)-2-hexenal, (E)-2-heptenal, (E)-2-octenal, (E)-2-nonenal, and 1-octen-3-ol can be found at levels above sensory thresholds.
The effects of dehydration on aroma composition are strongly dependent on many factors poorly controlled, such as the previous physiological state of the grape or the environmental conditions. Such variability has been observed for terpenols [85]. There are reports in which no changes in these compounds are observed during dehydration [86], others in which dramatic decreases were seen [87], and even others in which slight increases were measured [37,88]. A similar degree of diversity of patterns was also identified in the case of β-damascenone. Increased levels of this component, and also of γ-nonalactone [64] and of massoia lactone [65], have been observed and related to the prevalence of prune and fig character of the wines made with partially raisinized grapes [80]. In contrast, other studies have shown that shriveled grapes did not produce wines with higher β-damascenone content [89]. In the case of Strecker aldehydes, levels formed will be likely strongly related to the levels of the amino acid precursors (methionine and phenylalanine) present in the grape.
Regarding aldehydes and alcohols from FAOs, these compounds in general decrease during grape dehydration [56,86,88,90,91]. Such decreases may be attributed to a reduction in the lipoxygenase activity [86,90,91] of the raisinized grapes which cannot compensate for the general and continuous decrease of aldehydes by reaction with, among others, grape polyphenols.
2.3. Aroma Compounds Responsible for Vegetal and Green Aroma and Flavors
There are two families of aroma compounds which play a role in the vegetal, herbaceous, and green–unripe characteristics of grapes, musts and, eventually, wine: alkylmethoxypyrazines along with aldehydes and alcohols derived from the oxidation of fatty acids, or fatty acid oxidation-derived odorants (FAOs).
Alkylmethoxypyrazines are extremely powerful aroma molecules which accumulate in some grapes. They were first found in wines from Cabernet Sauvignon [92] and were further identified in Sauvignon Blanc juices and wines [93]. These compounds are 3-isobutyl-2-methoxypyrazine (IBMP), 3-secbutyl-2-methoxypyrazine (SBMP), and 3-isopropyl-2-methoxypyrazine (IPMP). Their properties are listed in Table 3. These compounds accumulate preferably in fruits grown under cool conditions and their levels decrease during ripening. They have been blamed for the specific green bell pepper character associated with Cabernet varieties, with a threshold for this character estimated to be just 15 ng/L [94]. Carmenere wines, which also belong to the Cabernet family, contain large amounts of these compounds too. Levels of IBMP were found to be strongly affected by climatic conditions and by vine genotype [95]. Temperatures during spring were found to be an important driver of green characters [96]. Levels of IBMP have been also positively related to altitude [97] and negatively related to light exposure, which limits accumulation but does not promote degradation [98]. Consequently, leaf removal significantly reduces accumulation of IBMP but only if it is carried out before veraison [99]. The relationship with nitrogen fertilization seems to be indirect, through the higher vigor [100]. Anecdotally, huge levels of IPMP can be induced by some foreign ladybeetles, causing great concern [101]. The levels of these compounds in wines from Spain and other southern countries are very low. It should be remarked, however, that strong negative correlations between the levels of these compounds—notably IBMP—and the different fruity and liquorice attributes of wines have been found in a recent work [102]. Such negative correlation would suggest that these compounds could be relevant suppressors at subthreshold level.
Table 3.
Structures, odor properties, and occurrence of alkylmethoxypyrazines.
| Compound | Structure | Odor Descriptor | Odor Threshold | Range of Occurrence in Grape Juice |
|---|---|---|---|---|
| 3-Isobutyl-2-methoxypyrazine |
|
Bell pepper, earthy | 2 ng/L (in water) [103]; 15 ng/L (in wine) [94] |
n.d. to 79 ng/L [93] |
| 3-Isopropyl-2-methoxypyrazine |
|
Green pea, earthy | 0.74–1.11 (hybrid grape juice) [104]; 2 ng/L in wine [105] |
n.d. to 6.8 ng/L [93] |
| 3-Secbutyl-2-methoxypyrazine |
|
Bell pepper | 1–2 ng/L (in water) [106] | n.d. to 1.3 ng/L [93] |
n.d.: Not detected.
The second family of compounds is formed by a relatively large number of aroma compounds, most of them aldehydes, derived from the oxidation of fatty acids or FAOs. Since quantitatively the most abundant were C6 alcohols and aldehydes, the family was first referred as the C6-family, however, some of the most powerful aroma compounds have nine carbon atoms, such as E-2-nonenal or (E,Z)-2,6-nonadienal. For instance, the most relevant aroma compound of Cabernet Sauvignon must, as assessed by aroma extract dilution analysis was (E,Z)-2,6-nonadienal [107]. Chemical structures and basic properties of these compounds are given in Table 4. This group of compounds derives from the enzymatic oxidation of fatty acids during must processing [22] and are well-known for the green odor of green leaves particularly evident in some teas [21]. The most powerful in aroma are the aldehydes, as usual, which have odor thresholds hundreds of times smaller than those of the corresponding alcohols. These aldehydes are surely responsible for the herbaceous note characteristics of some musts, particularly of those produced from unripe grapes. However, aldehydes are mostly eliminated during fermentation, in which they are enzymatically reduced to the corresponding alcohols. Consequently, the role of the family on the green and herbaceous (negative) aroma characteristics of wines has yet to be clearly demonstrated. FAO odorants decrease with maturity. Their levels are strongly related to grape variety [108] and also to the position in the bunch [109], being richer in the shoulder.
Table 4.
Structures, odor properties, and occurrence of FAO-related 1 family of compounds.
| Compound | Structure | Odor Descriptor | Threshold in Water | Ranges of Occurrence in Grape [23,25,57,61,110,111,112] |
|---|---|---|---|---|
| Hexanal |

|
Herbaceous | 5 μg/L [113] | 8–1300 μg/kg |
| (Z)-3-Hexenal |
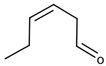
|
Grass | 0.25 μg/L [48] | 4–20 μg/kg |
| (E)-2-Hexenal |
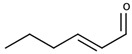
|
Grass | 17 μg/L [113] | 13–3800 μg/kg |
| (E,E)-2,4-Hexadienal |
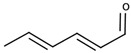
|
Grass | 60 μg/L [114] | 50–120 μg/kg |
| (Z)-3-Hexenol |
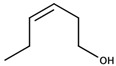
|
Grass | 70 μg/L [48] | 4–79 μg/kg |
| (E)-2-Hexenol |
|
Green | 400 μg/L [114] | |
| 1-Hexanol |
|
Green | 2500 μg/L [113] | 45–214 μg/kg |
| E-2-Nonenal |
|
Green, fatty | 0.17 μg/L [113] | |
| (E,Z)-2,6-Nonadienal |
|
Cucumber | 0.01 μg/L [115] | 113–482 μg/kg |
1 FAO: Fatty acid oxidation.
The vegetal aromas of Cabernet Sauvignon and other wines are, however, much more complex and cannot be completely explained just by analyzing IPMP and IBMP [116], or C6-alcohols. While some works from Allen’s group initially reported a high correlation between the sensory vegetative aroma notes of Cabernet Sauvignon grapes grown in five sites of Sonoma and IBMP levels, more recent reports have not been able to find any correlation [116]. In fact, a comprehensive understanding of the green and unripe characters of wines remains a major challenge for wine science today. Preliminary reports from our group suggest that (a) C6-alcohols together with IBMP can impart herbaceous notes to red wine [117]; (b) the concerted action of hexanol, the major C6 alcohol, with dimethyl sulfide and methanethiol, opposed to the action of acetaldehyde and linear fatty acids, could be related to the vegetal character of wine [70].
There is also strong evidence demonstrating the implication of 1,8-cineole, a terpineol of eucalyptus odor, in the green and minty characters of wine. In many instances, the origin of this molecule is exogenous, coming from leaves of Eucalyptus trees [118] or from invasive plants, such as Artemisia verlotiorum [119]. Highest levels are related to the presence of the Eucalyptus leaves or of small quantities of the plant in the fermentation tanks, but the molecule can accumulate in the berry skin at sensorily relevant levels [120]. Additionally, recent evidence has shown that the molecule can be found in unripe berries of Cabernet Sauvignon and Merlot [119], contributing to the green perception via perceptual interaction with IBMP. A third formation route of 1,8-cineole in wine as product of the reaction of limonene and other terpenols has been also reported [66,121].
2.4. Compounds Responsible for the Flavor of Neutral Grapes
The subtle flavor of neutral grapes is the consequence of the presence of very small amounts of a relatively large list of aroma compounds. The list includes nearly all the aroma compounds described in the three previous subsections, the difference being that neutral grapes do not contain any odorant at the concentrations at which it can be regarded to act as impact aroma compound. In fact, studies performed on the aroma composition of neutral varietals, such as Grenache, Monastrell, Tempranillo, Aglianico, or Uva di Troia, using direct liquid–liquid extraction or solid phase extraction only find at quantifiable levels C6 compounds together with minor levels of some hydrocarbons, alcohols, ketones, esters, and terpenes [122,123,124,125]. Methods using SPME can more easily find other nonpolar volatiles, because of its intrinsic higher concentration power [126], but at the expense of losing the most polar and less volatile ones, such as furaneol or vanillin derivatives.
Many neutral grapes contain low amounts of free furaneol, limonene, linalool, geraniol and other terpenols, β-damascenone, β-ionone and other norisoprenoids, and also of ethyl esters, such as ethyl butyrate, ethyl hexanoate, some volatile phenols, and vanillin derivatives. All these compounds, together with FAO derivatives, contribute concertedly to the subtle fruity flavor of neutral grapes. For instance, in one of the few works published about the gas chromatography-olfactometric (GCO) profiles of neutral grapes, the most relevant odorants were β-damascenone, β-ionone, ethyl hexanoate, ethyl octanoate, and different FAO derivatives (hexanal, decanal, and (E,Z)-2,6-nonadienal) [25]. With no impact aroma compound present, but with a relatively wide array of fruity–sweet–citric–flowery smelling aroma compounds present at low levels, there is a perceptual cooperation between all of them as described by Loscos et al. [127], whose outcome is a subtle sweet–fruity flavor.
There is also some evidence that neutral grapes of specific varieties contain eventually sensorily-relevant levels of rotundone. Rotundone is a sesquiterpene that is also present in grapes and can give a peppery aroma to grapes and wines [128]. In certain varieties, like Shiraz or Duras, and under favorable agronomical conditions [129,130], rotundone can accumulate in the berry exocarp in levels in the order of 600 ng/kg [128]. The synthesis pathway of rotundone in grape is not clear, but α-guaiene has been proposed as a potential precursor [131]. During the red wine winemaking maceration process, rotundone is extracted and can reach levels well above its perception threshold of 16 ng/L [128,132]. This characteristic peppery aroma is usually perceived positively among wine consumers [133].
Following the idea of aromatic series proposed by different authors [29,31,125], it can be stated that the aroma of neutral grapes is the consequence of the concerted action of 25–30 aroma compounds, with aroma nuances classifiable into seven odor categories:
Fruity: ethyl isobutyrate, ethyl butyrate, ethyl 3-methylbutyrate, ethyl hexanoate, ethyl octanoate, and eventually others;
Jammy, very sweet fruit: furaneol, homofuraneol, β-damascenone, γ-nonalactone, and massoia lactone;
Sweet–floral: vanillin, ethyl vanillate, β-ionone, β-phenylethyl acetate, and phenylacetaldehyde;
Floral–citric aroma compounds: linalool, geraniol, limonene, nonanal, and eventually others;
Herbaceous: hexanal, (Z)-3-hexenal, (E)-2-hexenal, (Z)-3-hexenol, (E)-2-nonenal, (E,Z)-2,6-nonadienal;
Peppery: rotundone;
Unspecific: 3-methylbutanal, ethyl acetate, diacetyl.
3. Grape Potential Aroma: Specific Aroma Precursors
3.1. Specific vs. Unspecific Precursors
Grape specific aroma precursors are non-volatile and hence odorless molecules which may rend a specific odoriferous molecule by the hydrolysis of a chemical bond, by spontaneous chemical rearrangement, or by a combination of both mechanisms. Many grape and grape-derived wine aroma molecules have specific aroma precursors. Remarkably, some of them have a relatively complex pool of different “specific precursors”. This is common in nature; for instance, apples contain more than eight different non-volatile molecules which by hydrolysis and further chemical rearrangement lead to β-damascenone [134]. A higher level of complexity regarding the number and type of precursor molecules is found in grapes. Such a pool of molecules is the pool of β-damascenone precursors. Similarly, there is a pool of precursors for linalool, for geraniol, for (Z)-rose oxide, for β-ionone, for furaneol, for TDN, for 3-mercaptohexanol, and for many other relevant grape-derived wine aroma compounds.
The word specific has an important meaning here. “Specific” means that the aroma compound will be formed by simple incubation of the pool of precursors extracted from grape at normal wine pH, or alternatively, by incubation in the presence of an enzyme. This definition deliberately excludes those precursor molecules which can be transformed into aroma compounds only by a complex metabolic action of yeast, bacteria or other micro-organisms. For instance, the amino acid isoleucine can be metabolized by Saccharomyces producing as byproducts isovaleric acid, isoamyl alcohol and isoamyl acetate. But isoleucine cannot be regarded as a specific precursor for these important aroma compounds, because their final levels are extraordinarily constrained by the metabolic requirements of yeast. In fact, yeast is able to produce all those compounds even if there is no isoleucine in the fermentation media. We rather should consider it as an unspecific precursor of the aroma molecule. This differentiation has a paramount importance for defining grape aroma potential. In general, wines made from grapes containing higher levels of specific precursors will develop higher levels of the aroma molecules derived from those precursors and/or will keep levels of those molecules for longer aging periods.
3.2. Grape Aroma vs. Grape-Derived Wine Aroma
As was schematized in Figure 1, grapes contain seven relatively well differentiated chemical/biochemical aroma precursor systems. As discussed previously, two of the systems—the fatty acid/enzymatic system and the Strecker amino acid system—have a major role in the development of the actual aroma of grapes, but to the best of our knowledge, they seem to have a rather limited role as wine aroma precursors. Both systems will influence wine aroma insofar as they form grape aroma molecules or precursors of aroma molecules, which will eventually pass to wine, but the systems as such do not survive fermentation. This explains why if the grape has not suffered raisination or over-ripening, the wine, generally, will not develop prune and overmatured character. On the contrary, the five other systems or molecular pools will be transferred to wine with different degrees of change induced by fermentation and will release or produce the specific aroma molecules at different moments of the winemaking process.
The wine odorants for which there is more or less strong evidence about the implication of grape specific precursors in their formation are summarized in Table 5, Table 6 and Table 7. The list includes 27 compounds: four norisoprenoids, five terpenes, six volatile phenols, four vanillin derivatives, ethyl cinnamate, two ethyl esters, two lactones, furaneol, DMS, and three polyfunctional mercaptans. Some compounds in the list, such as polyfunctional mercaptans, DMS, linalool, rose oxide, or TDN, can reach odor-impact levels. Some others, such as volatile phenols or vanillin derivatives, rather exert a cooperative effect on wine aroma. Mint lactones, recently identified at low levels in red wines from Bordeaux [135], limonene and 1,4- and 1,8-cineol, as well as some megastigmatrienones, may also play a role in minty, balsamic, and tobacco notes [66], but evidence about their implication is yet weak.
Table 5.
Wine norisoprenoid and terpene odorants coming from specific precursors.
| Aroma Molecule | Enzymatic Hydrolysis | Harsh Acid Hydrolysis | Mild/Long Term Acid Hydrolysis |
|---|---|---|---|
| Norisoprenoids | |||
| β-Damascenone | Not found; yes in raisins [23,57] and frozen grapes [136]; not in wines [137]; 0.17–0.5 ppb in frozen grapes [12] | 26 ppb [138]; detected by GCO [139]; 4–28 ppb depending varieties, unclear pulp/skin distribution [140]; 4–20 ppb depending location [140]; levels correlated to total norisoprenoids by enzymatic [141]; 2–4.5 ppb depending varieties [12] | Detected by GCO [142]; maxima (3.3 ppb) after short aging, then steady decrease [14]; steady increase all the aging in fermented samples [143]; maxima 7.1–7.3 ppb after medium aging in unfermented controls [143]; formed soon and stable, maxima 17 ppb [15]; idem, with maxima 7 ppb [66] |
| β-Ionone | Not found; yes in frozen grapes [136]; not in wines [137]; <0.11 ppb in frozen grapes [12] | Generally yes; not found in [12] | Maxima (1.9 ppb) after short aging, stable with time [14]; formed soon, stable for a while, maxima 7.7 ppb [15] |
| TDN | Not found; yes in frozen grapes [136]; not in wines [137]; 1–6 ppb (5–30% of levels found in harsh acid hydrolysis) in frozen grapes [12] | 8 ppb [138]; detected by GCO [139]; 1–35 ppb depending on varieties, unclear pulp/skin distribution [140]; n.d. to 26 ppb depending on place [140]; 8–89 ppb depending on varieties [12] | Linear increase with time, max 140 ppb [143]; idem, max at 61 ppb [15]; idem [66] |
| TPB | Not found; 0.2–3 ppb (2–22% of levels found in harsh acid hydrolysis) in frozen grapes [12] | 3 ppb [138]; 2–23 ppb depending varieties [12] | Continuously formed, maxima 9 ppb [66] |
| Terpenes | |||
| Linalool | Generally present; not found in Portuguese reds [140]; not found in Melon B [141]; not found in Shiraz [144]; found at low levels (less than 7% geraniol 1% total terpenes) [144] | 3% levels found in enzymatic [138]; 10–50% of levels found in enzymatic [12] | Found only in mild acid hydrolysis [141]; maxima after fermentation, sharp decrease in aging [14]; in Grenache, maxima after short aging [143]; formed very soon, sharp decrease [15,66] |
| Geraniol | Always found; up to 10% of total terpenes in Shiraz, 14% in Muscat [144] | No [138]; 3–30% of levels found in enzymatic [12] | Maxima in fermentation, sharp decrease in aging [14,143]; formed very soon, sharp decrease [15,66] |
| (Z)-Rose oxide | 11–29 ppb in Muscat, depending on maturity [145]; unrelated to free form in raisins [23] | 0.04 ppb in Muscat, 0.01 ppb in Grenache; not found in Verdejo, Tempranillo, Chardonnay, Cabernet Sauvignon, or Merlot [12] | |
| Geranic acid | Up to 2–3 ppm [146,147]; also found in raisins [23]; <4 ppb [145]; up to 7.5% total terpenes in Shiraz, 18% in Muscat [144] | Not found [138]; 0.5–50% of levels found in enzymatic [12] | 1.5 ppb in Chardonnay juices [148] |
| Piperitone | Derived from limonene, unknown accumulation pattern [149]; limonene accumulates in the first periods of aging, then slight decrease [66] | ||
n.d.: Not detected.
Table 6.
Wine benzenoid odorants coming from specific precursors.
| Aroma Molecule | Enzymatic Hydrolysis | Harsh Acid Hydrolysis | Mild/Long Term Acid Hydrolysis |
|---|---|---|---|
| Volatile Phenols | |||
| Guaiacol | Not found [146]: only in Brachetto, not in Aleatico, Malvasia, or Moscato [147]; <2 ppb [125]; up to 60 ppb in Rojal wine [137]; 0–41 ppb [150]; 10–76 ppb depending on vintage [151]; 15–44 ppb depending on vintage [152]; 17 ppb in Shiraz [144]; 0.4–2.3 ppb depending on varieties [12] | Detected by GCO [139]; <0.61 ppb, unrelated to enzymatic levels [12] | Detected by GCO [142]; Steady increase with time, maxima 4.3 ppb [14]; idem, maxima 6.3 ppb [143]; idem, maxima 14 ppb [15] |
| Eugenol | 1–8.3 ppb [146,147]; not found [125]; up to 33 ppb in Rojal wine [137]; present in less than half varieties, up to 16 ppb [150]; 84–216 ppb depending on vintage [151]; 12–20 ppb in Bobal depending on vintage [152]; n.d. to 9.4 ppb depending on variety [140]; 2.7–18 ppb depending on location [140]; 10 ppb in Shiraz [144]; 0.4–7 ppb depending on variety [12] | Detected by GCO [139]; <0.36 ppb, unrelated to enzymatic levels [12] | Steady increase, maxima 1.25 ppb [15] |
| Isoeugenol | Up to 14 ppb in Rojal wine [137]; 7.6–26 ppb depending on vintage [151]; 5–25 ppb depending on vintage [152]; 0.4–4.8 ppb depending on varieties [12] | <0.58 ppb, unrelated to enzymatic levels [12] | Detected by GCO [142] |
| 2,6-Dimethoxyphenol | 3–60 ppb [147]; n.d. to 13 ppb depending on varieties [12] | n.d. to 5.5 ppb depending on varieties [12] | Detected by GCO [142]; steady increase with time, maxima 33 ppb [14]; idem, maxima 142 ppb [15] |
| 4-Vinylguaiacol | 65–357 ppb [147]; <24 ppb [150]; 56–378 ppb depending on vintage [151]; 56–64 ppb depending on vintage in Bobal [152]; 2–114 ppb depending on varieties [140]; 2–178 ppb depending on location [140]; 21 ppb in Shiraz [144]; 39–162 ppb on depending varieties [12] | 40% of enzymatic [138]; detected by GCO [139]; 10–38 ppb depending on varieties, unrelated to enzymatic [12] | A maxima (21 ppb) after short aging, then decrease and steady increase [14]; continuous increase, maxima 5.5 ppm [143]; formed soon and stable, maxima at 1.3 ppm [15] |
| 4-Vinylphenol | 28–266 ppb [150]; 5–222 ppb depending on varieties [140]; 19–310 ppb depending on location [140]; 6 ppb in Shiraz [144]; 121–1739 ppb depending on varieties [12] | 9–21 ppb depending on varieties, unrelated to enzymatic [12] | A maxima after short aging (45 ppb), then decrease and steady increase, maxima 80 ppb [14]; continuous increase, maxima 4.4 ppm [143]; formed very soon, later steady decrease, maxima at 102 ppb [15] |
| Vanillin Derivatives | |||
| Vanillin | 27–42 ppb [147]; 361 ppb in skin of Uva di Troia [125]; 31–61 ppb [137]; <37 ppb [150]; 48–68 ppb depending on vintage [151]; 60–160 ppb depending on vintage in Bobal [152]; 31 ppb in Shiraz [144]; 40 ppb in Muscat [144]; <4.1 ppb [12] | 50% enzymatic [138]; detected by GCO [139]; <1.5 ppb [12] | Detected by GCO [142]; linear increase with time, maxima 45 ppb [14]; idem, maxima 91 ppb [143]; idem, maxima 123 ppb [15] |
| Methyl vanillate | 4–7 ppb [147]; <7 ppb [125]; up to 205 ppb in Rojal wine [137]; <42 ppb [150]; 12–147 ppb depending on vintage [151]; 9–143 depending on vintage in Bobal [152]; 25 ppb in Shiraz [144]; 154 ppb in Muscat [144]; <18 ppb [12] | <3.4 ppb [12] | 6 ppb in Chardonnay juices [148] |
| Ethyl vanillate | Up to 45 ppb in Rojal wine [137]; n.d. to 10 ppb depending on vintage in Bobal [152]; <12 ppb [12] | <3.1 ppb | |
| Acetovanillone | Up to 205 and 260 ppb in Rojal and Tortosí wines [137]; 1–12 ppb depending on vintage [151]; 42 ppb in Muscat, none in Shiraz [144]; 8–34 ppb depending on variety [12] | Detected by GCO [139]; <2.5 ppb, unrelated to enzymatic [12] | Unclear pattern [15]; 5 ppb in Chardonnay juices [148] |
| Cinnamic Acid Derivatives | |||
| Ethyl cinnamate | 7 ppb only in pulp from Uva di Troia [125]; <0.8 ppb [12]; its precursor, cinnamic acid has been found up to 7 ppb in fractions from wine, levels depending on vintage [137,151,152] | 12 ppb [138]; <0.11 ppb [12] | Detected by GCO [142] [15]; steady increase with time in some varietals, maxima 3.3 ppb [14]; maxima 3.3 ppb after short aging [143] |
n.d.: Not detected.
Table 7.
Wine miscellaneous odorants coming from specific precursors.
| Aroma Molecule | Enzymatic Hydrolysis | Harsh Acid Hydrolysis | Mild/Long Term Acid Hydrolysis |
|---|---|---|---|
| Ethyl cyclohexanoate | Its precursor, ethyl cyclohexanoic acid, found in unfermented mistellas [153] | ||
| Ethyl 4-methylpentanoate | Its precursor, ethyl 4-methylpentanoic acid, found in unfermented mistellas [153] | ||
| γ-Decalactone | No [125] | Identified [8] | Detected by GCO [15,142] |
| Massoia lactone | Detected by GCO [15] | ||
| Furaneol | Aglianico up to 2 ppm in pulp and 0.6 in skin, Uva di Troia 1,2 ppm in pulp, 90 ppb in skin [125]; 15–51 ppm in muscadine [46] | Detected by GCO [139] | Detected by GCO [15] |
| DMS | Only found in grape or grape mistellas not in precursor fractions [15] | ||
| Polyfunctional Mercaptans | |||
| 4-Methyl-4-mercaptopentan-2-one | Mostly released by yeast. | ||
| 3-Mercaptohexanol | Released by yeast. Detected by GCO in mild-acid hydrolyzates [15,142] | ||
| 3-Mercaptohexyl acetate | Formed by yeast from 3MH | ||
The tables summarize information relative to the presence of the odorants in hydrolysates obtained by enzymatic, harsh, or mild (long term) acid hydrolysis. This information is relevant to understand the genesis of the aroma compound and also to assess the relevance of the findings of the different reports. In some of the few studies using long term acid hydrolyses, there is additional information about the pattern of accumulation of the odorant with time. This information is crucial to understand the evolution of these aroma molecules during wine aging. As can be seen in Table 5, none of the four norisoprenoid odorants were present in enzymatic hydrolysates. Only in grapes kept frozen before the analysis, or in raisins, were these odorants found after enzymatic hydrolysis. In the case of terpenes (Table 5), volatile phenols, and vanillin derivatives (Table 6), enzymatic hydrolysis in general produced much higher levels than harsh acid hydrolysis. By contrast, most compounds are found at reasonable levels in hydrolysates obtained by long-term acid hydrolysis.
Large differences between compounds are also found regarding the pattern of accumulation during aging. Linalool and geraniol reach maximal levels immediately after fermentation or after a short aging time, and afterwards their levels decay dramatically. β-Damascenone and β-ionone reach maximal levels also after a relatively short aging period, but their levels decay slowly or stay stable (Table 5). By contrast, TDN, TPB, and most volatile phenols and vanillin derivatives steadily increase during aging (Table 5 and Table 6). 4-Vinylphenol and 4-vinylguaiacol follow more complex evolutions with at least two maxima, likely because of the number of precursors they have and their chemical reactivity. The evolution with time of some relevant odorants, such as (Z)-rose oxide, geranic acid, or piperitone is mostly unknown.
Data summarized in the tables also reveal the existence of huge variabilities in the levels of most compounds, regarding variety, vintage, location, or maturity. While some differences may be attributed just to the different analytical methodologies followed by the researchers, some others truly reflect a large diversity. Differences between Muscat grapes and “neutral” grapes regarding levels of terpenols are known, as well as those of furaneol between hybrids and Vitis vinifera varieties. However, data in Table 6 suggest that differences in the levels of some volatile phenols and vanillin derivatives are well above the order of magnitude.
Finally, Table 7 contains some odorants for which the existence of precursors can be expected but has not been demonstrated.
The following four sections deal with the different types of precursors responsible for all those odorants. The first section deals with glycosidic precursors, the second with other precursors, and the two last sections with glutathionyl and cysteinyl precursors and DMS precursors.
3.3. Glycoconjugates as Aroma Precursors
Some good reviews on these questions have been recently published [154,155,156]. Glycoconjugation is a clever way to solubilize and fix nonpolar and volatile aroma molecules and it is very common in nature [157]. Many secondary metabolites of plants are glycoconjugated, and in fact, glycoconjugation can be considered a relatively common last step of plant secondary metabolism and seems to be a primary sedative mechanism used by plants to maintain metabolic homeostasis [158] and to detoxify from potentially toxic (lipophilic and/or reactive nucleophiles) molecules [159]. Glycoconjugation takes place by reaction between a reactive functional group and an “activated” sugar. Activated sugars are UDP-glucose, UDP-rhamnose, UDP-galactose, UDP-xyloxe and glucuronic acid, where UDP stands for uracil-diphosphate glucose. The reactive functional groups are -COOH, -NH2, -SH, and -OH, among others.
In the case of grapes, little is known about the real activities and selectivities of glycosyltransferases, but at least 240 different types of these enzymes are coded in the grape genome [160]. Although glycosides may be more easily handled and transported by plant transport systems, recent evidences suggest that grape aroma glycosides are integrally formed in the grape.
Of course, major grape glycosides are those of flavonoids, phenolic acids, and anthocyanins, while aroma compounds represent quantitatively a quite modest fraction. In the case of aroma compounds, to date, all grape aroma-related derivatives have been found to be bound to a β-D-glucose, and such glucose can be further bound to malonic acid, arabinose, apiofuranose, or rhamnose to form the structures indicated in Figure 3.
Figure 3.

Sugar moieties of glycoside precursors. Adapted from [154].
Recently, two trisaccharides have been also tentatively identified in grape [161,162].
According to the type of aglycone, glycoconjugates in grapes can be broadly classified into the following categories:
Aliphatic alcohol derivatives;
Terpenes;
Norisoprenoids;
- Benzenoids, which can be further subdivided into:
- Benzyl and phenyl derivatives;
- Volatile phenols;
- Vanillins;
- Ethyl cinnamate.
Miscellaneous compounds.
Aliphatic alcohol derivatives can be quantitatively important, but they are quite unimportant from the aromatic point of view. Compounds in this group, among others, include isoamyl alcohol, hexanol, (Z)-3-hexenol, (E)-2-hexenol, 1-octen-3-ol, heptanol, and octanol [138].
Terpenes include a quite complex array of terpenes in different oxidation states. The list includes several terpenic diols which will be presented in the next section, together with linalool, α-terpineol, nerol, geraniol, and several of their oxides, including c-rose oxide. The most important from the sensory point of view are the same four as in the free fraction, namely linalool, geraniol, c-rose oxide, and geranic acid. Note that some of these compounds will suffer chemical rearrangements at acidic pHs. Different reports have estimated that between 77% to 83% of the total terpenic content in Riesling grapes are present as glycosides [163,164,165]. Some of them, such as different hydroxylated forms of the main terpenols or of geranic acid, seem to be majorly or even exclusively found as glycosides [147]. From the quantitative point of view, major aglycones of terpenes in neutral varietals are those of geraniol (Figure 4), (Z)-8-hydroxy-linalool (or (2Z)-2,6-dimethylocta-2,7-diene-1,6-diol), and p-menthene-7,8-diol with account to more than 60% of the peak area, eventually followed by those of linalool and geranic acid and those of the (E)- and (Z)-pyran linalool oxides [137,140,150,166]. A glycoside precursor of 1,8-cineole, namely 2-exo-hydroxy-1,8-cineole, has been also identified in Falanghinna grapes [167].
Figure 4.

Release of geraniol via acid-catalyzed hydrolysis of the geranyl-β-D-glucopyranoside.
There are a number of recent reports about the evolution of these precursors during grape maturation. Results show that the patterns of accumulation depend largely on the aroma compound [145], on the variety of grape [147], and on the vintage [146], which makes difficult to extract sound conclusions. In general, it can be said that glycosidic forms tend to increase with maturation following more regular accumulation patterns than free forms, which can show erratic patterns of evolution during maturation.
As summarized in Table 5, levels of linalool and geraniol are maximal in the wines immediately or shortly after fermentation, and levels decrease due to the poor stability of these molecules at wine pH. The pool of precursors which survived the fermentation seems to be essential for keeping the levels of these relevant aromas longer times [14,143].
Aglycones in the norisoprenoid family can be also extraordinarily complex and, not surprisingly, there are not aglycones representing the most relevant aroma compounds in this family, such as β-damascenone, β-ionone, TDN, or TPB. The major aglycones are 3-hydroxy-β-damascone, dihydro-β-ionone, and different ionols, particularly 3-oxo-a-ionol and vomifoliol [140,150]. This represent quite a nuisance, since the direct analysis of the aglycones (after careful enzymatic hydrolysis) or the direct HPLC-MS of the unaltered glycosidic precursors do not give clear information about the aroma potentiality of this important precursor fraction.
There is large difference between the four major nor-isoprenic odorants regarding the pattern of accumulation during aging. β-Damascenone and β-ionone reach maximal levels soon and then remain stable or steadily decrease with aging. By contrast, TDN and TPB are formed much more slowly during aging, with levels steadily increasing, as indicated in Table 5. A recent report has shown that fermented samples form TDN faster than unfermented controls, which suggests that some of the first chemical reactions in the sequence required to form TDN from 3,6-dihydroxy-β-ionone, its main precursor [168], are accelerated by yeast [143]. Such a report also demonstrates that levels of TDN formed during aging can be modulated by yeast.
Within the group of benzenoids (Table 6) there are several subgroups of volatile compounds usually identified in the hydrolysates of grape precursor fractions [8,12,14,142,147].
Benzyl and phenyl derivatives include benzaldehyde, benzoic acid, benzyl alcohol, and 2-phenylethanol. In many neutral grape varieties these compounds, particularly the latter two, are the major constituents of the glycosidic aroma fraction [137,140,150]. This has some practical relevance since the contribution of these glycosides to wine flavor can be considered negligible. One the one hand, the odor thresholds of both odorants are relatively high, and on the other hand, 2-phenylethanol is a main secondary product of yeast metabolism, so that levels derived from grape glycosides represent a quantitatively marginal fraction. The consequence is that indirect measures for the aromatic potential of neutral grapes [169] may be not related to the true aromatic potential but just to the general secondary metabolic activity of the grape.
Volatile phenols, such as guaiacol, eugenol, isoeugenol, 2,6-dimethyoxyphenol, 4-vinylguaiacol, and 4-vinylphenol, are relevant components of the hydrolysates obtained from fractions of precursors extracted from grapes or wines, as detailed in Table 6. All or some of them tend to score high in the different GCO studies carried out on hydrolysates [139,142,170]. Reported levels of all these compounds have ranges of variation depending on vintage and varieties close to two orders of magnitude, as summarized in Table 6. These compounds cannot be determined by harsh acid hydrolysis, even though most of them accumulate steadily during aging. The case of vinylguaiacol and vinylphenol is particularly interesting. Both can be considered detrimental for wine quality if present at high levels [171]. As recently documented, they can be formed via yeast phenolic acid decarboxylases from phenolic acids and also by enzymatic or acid hydrolyses of their glycosides [143].
Vanillin and other related compounds are also formed from different precursors. Although the levels of these important aroma compounds derived from the grape cannot rival with levels released by some types of oak wood, grapes contain a large number of precursors able to release significant levels of these compounds. Vanillin is one of the odorants of acid hydrolysates which always scores very high by GCO [15,155,158,159]. In the enzymatic hydrolysates obtained from some varieties, such as those of skins from Uva di Troia [125] vanillin can be found at high levels (more than 360 μg/kg). Additionally, vanillin can be also formed by oxidation of 4-vinylguaiacol [172].
Ethyl cinnamate has been also found at minor levels in the hydrolysates of precursor fractions extracted from grapes (see Table 6). Since cinnamic acid was also identified as aglycone after enzymatic hydrolysis, the precursor should be a glycoside. A glycoside of cinnamic acid has been recently identified in wine made from Korean black raspberries [173].
Within the miscellaneous section (Table 7), the most relevant odorant is furaneol. Furaneol glucopyranoside has been recently identified and quantified in the must of Muscat Bailey A (V. labrusca (Bailey) × V. vinifera (Muscat Hamburg)) [174]. The gene encoding the UDP-glucose: furaneol glucosyltransferase was also determined [175]. The same authors were also able to quantify this precursor in different grape varieties and in the parental concord. Concentrations of the precursor were much higher in the labrusca and in the hybrids, but normal grapes also contain low amounts of this precursor. This aroma compound has been systematically identified by GC olfactometry in the hydrolyzed precursor fractions extracted from Grenache [142], Aragonez [139], Pinot Noir [170], or Tempranillo [15], and it has been found as aglycone released by enzymatic hydrolysis of the precursor fraction from Aglianico and Uva di Troia [125].
Finally, it should be noted that several authors have reported the presence of glycosides of some fatty acids at relatively large levels in the enzymatic hydrolysates of precursor fractions extracted from wines. For instance, isovaleric acid was found at 109 μg/L, butyric acid at 412 μg/L, hexanoic at 336 μg/L, and octanoic acid at 295 μg/L [150,152]. These amounts are just slightly smaller than those formed by yeast.
3.4. Other Precursors: Molecules Which by Chemical Rearrangement or Esterification Form the Aroma Molecule
The first type of molecules includes a series of polyols discovered more than 30 years ago which by chemical rearrangements induced by the acid hydrolysis at wine pH produce different aroma active terpenols [2].
As shown in Figure 5, one of the diols (3,7-dimethyloct-1-ene-3,7-diol) rearranges to give linalool and α-terpineol. The other molecules are different terpenols of lesser olfactory importance such as myrcenol or ocimenol. The diols were also found to be present as glycosides [176]. Some C13-triols with a megastimagne structure were also further identified as potential precursors for some norisoprenoids such as vitispiranes and TDN [177]. At wine pH, these precursors can spontaneously form TDN, responsible for the kerosene–off odor developed by some wines during aging. Also a megastimagne structure, megastigm-5-en-7-yne-3,9-diol, was identified as precursor for β-damascenone [178]. This was later confirmed by synthesis of the pure molecule [179]. The dienyne derivative and the allenic diol, shown in Figure 6, were further identified in 2005 [180]. Both proceed from an allenic triol derived from the degradation of carotenoids such as neoxanthin [181].
Figure 5.

Polyol rearrangement at pH 3.2. Adapted from [2].
Figure 6.

Formation of β-damascenone from allenic triol. Adapted from [181].
Notably, Australian researchers have recently demonstrated that a ketone and a diketone derived from diol 5 can be transformed by the action of yeast in β-damascenone [182].
As previously mentioned, most of these molecules are also found as glycosides, which supposedly amount to a larger fraction of precursors.
Finally, in this section we should mention the two lactones and the two ethyl esters listed in Table 7: γ-decalactone and massoia lactone and ethyl cyclohexanoate and ethyl 4-methylpentanoate. The two lactones are primarily formed during grape dehydration, but since they accumulate in some wines or precursor fractions, it can be suggested that the corresponding γ-hydroxy or δ-hydroxy acids are present as precursors. As different glycosidic precursors of whisky lactones (γ-methyloctalactone) have been described in oak wood [183,184] the presence of some glycosides of the acids cannot be ruled out. In the case of the esters, the corresponding acids have been quantified in unfermented grape must [185].
3.5. S-Derivatives of Cysteine or Glutathione
Two recent reviews [186,187] have been published on cysteinyl or glutathionyl derivatives. Grapes contain some cysteinyl or glutathionyl derivatives which by hydrolysis of the S–C bond in the cysteine part can give some of the most powerful aroma molecules of wine and of nature in general. The aroma molecules are 4-methyl-4-mercaptopentan-2-one (4MMP), 3-mercaptohexanol (3MH), and 3-mercaptohexyl acetate (3MHA). The aroma properties of these relevant aroma compounds are summarized in the following Table 8 [187]:
Table 8.
Structures, odor properties, and occurrence of varietal thiols.
| Compound | Structure | Odor Descriptor | Threshold in Model Wine (ng/L) [188] | Range of Occurrence in Wine (ng/L) [189] |
|---|---|---|---|---|
| 4-Methyl-4-mercaptopentan-2-one |
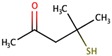
|
Box tree | 0.8 | n.d. to 90 |
| 3-Mercaptohexanol |
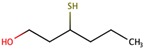
|
Grapefruit | 60 | n.d. to 7300 |
| 3-Mercaptohexyl acetate |
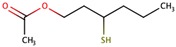
|
Box tree, passion fruit | 4 | n.d. to 440 |
n.d.: Not detected.
There are at least three or four more other varietal polyfunctional mercaptans in wine with far less aromatic importance.
All these aroma compounds are released by the action of β-lyase enzymes from yeasts from their specific precursors present in the grape must. The 3MHA requires, in addition, the acetylation of the alcohol 3MH by action of an acyltransferase also from yeast, as summarized in Figure 7.
Figure 7.

Biogenesis pathways of 4MMP, 3MH, and 3MHA. Adapted from [187].
Apart from the precursors described in Figure 7, very recent reports demonstrate also the existence of the glutathione precursor of 4-mercapto-4-methylpentan-2-ol [190] and of hexanal [191]. The first precursors discovered were the cysteinylated ones [192], and for over 10 years thiols were thought to be formed exclusively from cysteine conjugates. Glutathione precursors were identified much later and definitive evidence of their effective role as precursors of 3MH and 4MM4P was obtained only some years ago [193,194,195]. Recently, a glutamyl–cysteine dipeptide S-conjugate to 3MH has also been identified in must [196]. From the quantitative point of view, Glu–3MH precursor is the most concentrated, being present at levels between 8 and 35 times higher than those of the Cys–3MH precursor. In the case of MP, both can be at similar levels [197] (see Table 9).
Table 9.
Mean concentration of 4MMP and 3MH cysteinylated and gluthanionylated precursors in μg/L ± RSD% (n = 2) in eight grape varieties [197].
| Variety | CYS–MH | CYS–MMP | GLU–MH | GLU–MMP |
|---|---|---|---|---|
| Sauvignon Blanc | 174 ± 7 | 12.6 ± 1.4 | 1557 ± 86 | 7.7 ± 1.3 |
| Gewürztraminer | 89 ± 6 | 8.0 ± 1.5 | 1154 ± 56 | 6.6 ± 0.8 |
| Muscat | 157 ± 8 | n.d. | 1673 ± 71 | 8.3 ± 0.9 |
| Grenache | 172 ± 5 | 7.9 ± 1.2 | 1422 ± 63 | 9.4 ± 1.2 |
| Albariño | 158 ± 3 | 7.2 ± 0.7 | 1462 ± 80 | 8.4 ± 0.7 |
| Tempranillo | 205 ± 8 | 6.1 ± 1.8 | 1284 ± 76 | 10.3 ± 1.1 |
| Verdejo | 215 ± 9 | 7.3 ± 1.0 | 3397 ± 102 | n.d. |
| Chardonnay | 32 ± 4 | 0.4 ± 0.2 | 1405 ± 97 | n.d. |
n.d.: Not detected.
The conjugated thiol precursors are produced in the grape and concentrations are highest in the skin [198]. Little is known, however, about their biosynthesis and about the factors determining their accumulation during grape maturation. Levels are varietal-dependent, being highest in Sauvignon Blanc and Verdejo and close to null in Malvasia del Lazio, and increase during maturation [190]. Levels are also related to picking time [199], being maximum at early morning and later decreasing during the day. Interesting changes in amino acid levels during the day have been also identified as a consequence of leaf photosynthesis [200].
As it is also suggested in the previous figure, there is an additional prefermentative pathway leading to the in situ formation of 3MH precursors during grape processing before fermentation. According to this pathway, 3MH precursors form once the berry is damaged by reaction between E-2-hexenal formed via enzymatic oxidation of grape fatty acids and cysteine or glutathione present in the must. The existence of such pathway resulted as evident by the observed paradox that hand-picked grapes from Sauvignon Blanc produced wines much less aromatic than those harvested by machine [201]. The relative importance of the two different “kinds” of precursors, those already present in the grape and those formed in situ during early grape processing, is not clear. Subileau et al. showed that in their conditions (E)-2-hexenal was not a major contributor [194], while different studies confirm that machine-harvested grapes contain higher levels, with excessive oxidation being detrimental [201,202]. The effects of maceration time and pressing have been also studied by several authors, mostly concluding that prolonged maceration times leaded to higher levels of precursors [203,204]. More recently, Larcher et al. demonstrated that oxygen at harvest was essential for increased levels of precursors [205]. The apparent contradictory observations could be related to the existence of several concurrent factors not yet well controlled in the experiments such as the E-2-hexenal formation rate of the grape (dependent on grape lipoxygenases, oxygen, and grape fatty acids) and the cysteine and glutathione availability of the must.
Cysteinyl and glutathionyl precursors are poorly metabolized by most yeasts, so that levels of the precursors in the final wines can be high [206], particularly if the must contains high levels of glutathione [15]. It should be noted that there is evidence, some old [142,207] and some new [15], suggesting that the powerful polyfunctional mercaptans could be also formed by acid hydrolysis of the precursors. The role of this pool of compounds to keep longer levels of these powerful aroma compounds should not be ruled out.
3.6. S-Methylmethionine and Other DMS Precursors
Dimethyl sulfide is a quite remarkable wine aroma compound. It has been repeatedly identified as a powerful aroma enhancer [117,208] and, more specifically, as a contributor to blackberry and blackcurrant aroma nuances of red wines [209].
This compound can be formed by spontaneous hydrolysis of different precursors (very fast at alkaline medium) [210], of which S-methylmethionine (vitamin U) has been identified as the most important [211]. There are nearly no other reports about the occurrence and factors affecting the levels of this precursor in grapes, although its level has been found to be related to water deficit of vines [212]. Vines with moderate water deficit have higher potential for this compound and the concomitant higher levels of yeast assimilable nitrogen contained in the musts from those vines seem additionally to avoid the destruction (metabolization) of the precursor during fermentation.
3.7. The Action of Fungus and Other Exogenous Factors on Grape Actual and Potential Aroma
Finally, the aroma of the must or grapes reaching the cellar can be strongly affected by the presence of fungus or by some other exogenous factors. Wines made from grapes affected by noble rot have higher levels of 3MH, furaneol, sotolon, methional, and phenylacetaldehyde [59,77,78], while wines made from grapes affected by uncontrolled fungal attacks can develop fungal odors. Some of them, at smaller levels, are of course also present in noble rot wines, such as 1-octen-3-ol [78]. The infection with Botrytis cinerea also changes some must enzymes with effect on aroma (esterase and β-glucosidase).
Grapes affected by noble rot have also increased levels of cysteinyl precursors [213] and can have even an expanded number of this type of precursor [214], which helps explaining their particular aroma.
Regarding negative odors related to fungal attacks, 3-octanone, 1-octen-3-one, (E)-2-octenol, 1-octen-3-ol, 2-methyl isoborneol, TCA, geosmin, TBA, and pentachloroanisole are usually targeted as responsible for off-odors [215]. The type and levels are related to the strain of fungus; 50% of Botrytis cinerea strains induce geosmin, one strain induces anisol [216]
Following the exposure of vineyards to forest or bushfires, the occurrence of the smoke taint has been detected repeatedly; one review has been published recently about this off-flavor in wine [217]. Volatile phenols, like phenol, guaiacol, and their derivatives, that usually appear in wines as a consequence of barrel toasting or contamination with Brettanomyces yeasts, are present in greater quantities in wines produced with grapes exposed to smoke [218]. The evidence that free run juice of smoked grapes had trace levels of volatile phenols, while the same juice after several days of maceration showed levels in the range of hundreds of μg/L, proved that volatile phenols were stored in the skin rather that in the pulp [219]. Several studies have confirmed that the accumulation of volatile phenols takes place in the form of different glycoconjugates [220,221,222]. The release of volatile phenols from their precursor forms takes place not only during fermentation via enzymatic hydrolysis, but also via acid hydrolysis during post-bottle aging [223].
4. Final Conclusions
Both grape aroma and grape-derived wine aroma are formed by a relatively large group of odorants belonging to different chemical and biochemical families. Only in the specific cases of aromatic grapes are there clear impact compounds or families of compounds defining the aroma profile. In neutral varieties, grape aroma profiles are rather the consequence of the presence of more than 20 odorants imparting at least seven different types of aroma nuances. In the case of wine, up to 27 relevant wine odorants have specific origin in grape molecules or specific aroma precursors. Those odorants have, however, a much larger aromatic diversity than that observed between grape odorants, introducing or contributing to many different wine odor nuances such as fruity, jammy, floral, citrus, phenolic, spicy, empyreumatic, or green, and hence contributing decisively to wine quality.
Additionally, grape-derived wine aroma molecules accumulate in quite different time periods of winemaking; some of them are mostly released during fermentation, while some others accumulate only after long periods of aging. Within the first, remaining precursors in wine can have a crucial effect on keeping levels of odorants during aging, and therefore, in wine shelf-life. Within the latter, some of the odorants accumulating during aging, such as DMS, TDN, or TPB, may have controversial effects on wine quality, and may therefore have also a major influence on wine longevity.
For of all these reasons, the control of grape-derived wine aroma is an essential piece for controlling wine quality and wine shelf-life. Comprehensive analytical strategies for such a control have to face demanding challenges, which at present are not satisfactorily solved. On the one hand, aroma molecules with different chemophysical properties have to be simultaneously determined, which is nearly impossible using a single isolation strategy. On the other hand, the strategy has to sort out the difficult and non-obvious link between specific precursors and wine odorants. Surely this will require combining metabolomic approaches with new, comprehensive hydrolytical strategies. Both techniques are at hand but will require from researchers a clear awareness of all the dimensions of the analytical problem.
Funding
Funded by the Spanish Ministry of Economy and Competitiveness (MINECO) (project AGL2017-87373-C3-1-R). LAAE acknowledges the continuous support of Diputación General de Aragón (T29) and European Social Fund.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- 1.Cordonnier R., Bayonove C.L. Mise en evidence dans la baie de raisin, var. Muscat d’Alexandrie, de monoterpenes lies revelables par une ou plusieurs enzymes du fruit. Comptes Rendus de l’Académie des Sciences. 1974;278:3387–3390. [Google Scholar]
- 2.Williams P.J., Strauss C.R., Wilson B. Hydroxylated Linalool Derivatives as Precursors of Volatile Monoterpenes of Muscat Grapes. J. Agric. Food Chem. 1980;28:766–771. doi: 10.1021/jf60230a037. [DOI] [Google Scholar]
- 3.Cullere L., Lopez R., Ferreira V. Red Wine Technology. Elsevier; Amsterdam, The Netherlands: 2019. The Instrumental Analysis of Aroma-Active Compounds for Explaining the Flavor of Red Wines; pp. 283–307. [DOI] [Google Scholar]
- 4.Ferreira V., De-la-Fuente A., Sáenz-Navajas M.P. Wine aroma vectors and sensory attributes. In: Reynolds A., editor. Managing Wine Quality. 2nd ed. Woodhead Publishing (Elsevier); Amsterdam, The Netherlands: 2020. pp. 1–20. [Google Scholar]
- 5.Velasco R., Zharkikh A., Troggio M., Cartwright D.A., Cestaro A., Pruss D., Pindo M., FitzGerald L.M., Vezzulli S., Reid J., et al. A high quality draft consensus sequence of the genome of a heterozygous grapevine variety. PLoS ONE. 2007;2:e1326. doi: 10.1371/journal.pone.0001326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Williams P.J., Strauss C.R., Wilson B., Massy-Westropp R.A. Use of C18 reversed-phase liquid chromatography for the isolation of monoterpene glycosides and nor-isoprenoid precursors from grape juice and wines. J. Chromatogr. A. 1982;235:471–480. doi: 10.1016/S0021-9673(00)85911-7. [DOI] [Google Scholar]
- 7.Gunata Y.Z., Bayonove C.L., Baumes R.L., Cordonnier R.E. The aroma of grapes I. Extraction and determination of free and glycosidically bound fractions of some grape aroma components. J. Chromatogr. A. 1985;331:83–90. doi: 10.1016/0021-9673(85)80009-1. [DOI] [Google Scholar]
- 8.Ibarz M.J., Ferreira V., Hernandez-Orte P., Loscos N., Cacho J. Optimization and evaluation of a procedure for the gas chromatographic-mass spectrometric analysis of the aromas generated by fast acid hydrolysis of flavor precursors extracted from grapes. J. Chromatogr. A. 2006;1116:217–229. doi: 10.1016/j.chroma.2006.03.020. [DOI] [PubMed] [Google Scholar]
- 9.Hampel D., Robinson A.L., Johnson A.J., Ebeler S.E. Direct hydrolysis and analysis of glycosidically bound aroma compounds in grapes and wines: Comparison of hydrolysis conditions and sample preparation methods. Aust. J. Grape Wine Res. 2014;20:361–377. doi: 10.1111/ajgw.12087. [DOI] [Google Scholar]
- 10.Gunata Y.Z., Bayonove C.L., Baumes R.L., Cordonnier R.E. Changes in free and bound fractions of aromatic components in vine leaves during development of muscat grapes. Phytochemistry. 1986;25:943–946. doi: 10.1016/0031-9422(86)80032-2. [DOI] [Google Scholar]
- 11.Carro N., López E., Günata Z.Y., Baumes R.L., Bayonove C.L. Free and glycosidically bound aroma compounds in grape must of four non-floral Vitis vinifera varieties. Analusis. 1996;24:254–258. [Google Scholar]
- 12.Loscos N., Hernandez-Orte P., Cacho J., Ferreira V. Comparison of the Suitability of Different Hydrolytic Strategies to Predict Aroma Potential of Different Grape Varieties. J. Agric. Food Chem. 2009;57:2468–2480. doi: 10.1021/jf803256e. [DOI] [PubMed] [Google Scholar]
- 13.Francis I.L., Sefton M.A., Williams P.J. Sensory Descriptive Analysis of the Aroma of Hydrolyzed Precursor Fractions from Semillon, Chardonnay and Sauvignon Blanc Grape Juices. J. Sci. Food Agric. 1992;59:511–520. doi: 10.1002/jsfa.2740590414. [DOI] [Google Scholar]
- 14.Loscos N., Hernandez-Orte P., Cacho J., Ferreira V. Evolution of the aroma composition of wines supplemented with grape flavour precursors from different varietals during accelerated wine ageing. Food Chem. 2010;120:205–216. doi: 10.1016/j.foodchem.2009.10.008. [DOI] [Google Scholar]
- 15.Alegre Y., Arias-Pérez I., Hernandez-Orte P., Ferreira V. Development of a new strategy for studying the aroma potential of winemaking grapes through the accelerated hydrolysis of phenolic and aromatic fractions (PAFs) Food Res. Int. 2019 doi: 10.1016/j.foodres.2019.108728. in press. [DOI] [PubMed] [Google Scholar]
- 16.Noordermeer M.A., Veldink G.A., Vliegenthart J.F.G. Fatty acid hydroperoxide lyase: A plant cytochrome P450 enzyme involved in wound healing and pest resistance. ChemBioChem. 2001;2:494–504. doi: 10.1002/1439-7633(20010803)2:7/8<494::AID-CBIC494>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 17.Podolyan A., White J., Jordan B., Winefield C. Identification of the lipoxygenase gene family from Vitis vinifera and biochemical characterisation of two 13-lipoxygenases expressed in grape berries of Sauvignon Blanc. Funct. Plant Biol. 2010;37:767–784. doi: 10.1071/FP09271. [DOI] [Google Scholar]
- 18.Starkenmann C., Le Calve B., Niclass Y., Cayeux I., Beccucci S., Troccaz M. Olfactory Perception of Cysteine-S-Conjugates from Fruits and Vegetables. J. Agric. Food Chem. 2008;56:9575–9580. doi: 10.1021/jf801873h. [DOI] [PubMed] [Google Scholar]
- 19.Munoz-Gonzalez C., Cueva C., Pozo-Bayon M.A., Moreno-Arribas M.V. Ability of human oral microbiota to produce wine odorant aglycones from odourless grape glycosidic aroma precursors. Food Chem. 2015;187:112–119. doi: 10.1016/j.foodchem.2015.04.068. [DOI] [PubMed] [Google Scholar]
- 20.Parker M., Black C.A., Barker A., Pearson W., Hayasaka Y., Francis I.L. The contribution of wine-derived monoterpene glycosides to retronasal odour during tasting. Food Chem. 2017;232:413–424. doi: 10.1016/j.foodchem.2017.03.163. [DOI] [PubMed] [Google Scholar]
- 21.Hatanaka A. The Biogeneration of Green Odor by Green Leaves. Phytochemistry. 1993;34:1201–1218. doi: 10.1016/0031-9422(91)80003-J. [DOI] [Google Scholar]
- 22.Joslin W.S., Ough C.S. Cause and fate of certain C6 compounds formed enzymatically in macerated grape leaves during harvest and wine fermentation. Am. J. Enol. Vitic. 1978;29:11–17. [Google Scholar]
- 23.Wang D., Duan C.Q., Shi Y., Zhu B.Q., Javed H.U., Wang J. Free and glycosidically bound volatile compounds in sun-dried raisins made from different fragrance intensities grape varieties using a validated HS-SPME with GC-MS method. Food Chem. 2017;228:125–135. doi: 10.1016/j.foodchem.2017.01.153. [DOI] [PubMed] [Google Scholar]
- 24.Slegers A., Angers P., Ouellet E., Truchon T., Pedneault K. Volatile Compounds from Grape Skin, Juice and Wine from Five Interspecific Hybrid Grape Cultivars Grown in Quebec (Canada) for Wine Production. Molecules. 2015;20:10980–11016. doi: 10.3390/molecules200610980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Fan W.L., Xu Y., Jiang W.G., Li J.M. Identification and Quantification of Impact Aroma Compounds in 4 Nonfloral Vids vinifera Varieties Grapes. J. Food Sci. 2010;75:S81–S88. doi: 10.1111/j.1750-3841.2009.01436.x. [DOI] [PubMed] [Google Scholar]
- 26.Mayr C.M., Parker M., Baldock G.A., Black C.A., Pardon K.H., Williamson P.O., Herderich M.J., Francis I.L. Determination of the importance of in-mouth release of volatile phenol glycoconjugates to the flavor of smoke-tainted wines. J. Agric. Food Chem. 2014;62:2327–2336. doi: 10.1021/jf405327s. [DOI] [PubMed] [Google Scholar]
- 27.Parker M., Barker A., Black C.A., Hixson J., Williamson P., Francis I.L. Don’t miss the marc: Phenolic-free glycosides from white grape marc increase flavour of wine. Aust. J. Grape Wine Res. 2019;25:212–223. doi: 10.1111/ajgw.12390. [DOI] [Google Scholar]
- 28.Ribereaugayon P., Boidron J.N., Terrier A. Aroma of muscat grape varieties. J. Agric. Food Chem. 1975;23:1042–1047. doi: 10.1021/jf60202a050. [DOI] [Google Scholar]
- 29.Wu Y.S., Zhang W.W., Yu W.J., Zhao L.P., Song S.R., Xu W.P., Zhang C.X., Ma C., Wang L., Wang S.P. Study on the volatile composition of table grapes of three aroma types. LWT Food Sci. Technol. 2019;115:108450. doi: 10.1016/j.lwt.2019.108450. [DOI] [Google Scholar]
- 30.Wu Y.S., Zhang W.W., Duan S.Y., Song S.R., Xu W.P., Zhang C.X., Bondada B., Ma C., Wang S.P. In-Depth Aroma and Sensory Profiling of Unfamiliar Table-Grape Cultivars. Molecules. 2018;23:1703. doi: 10.3390/molecules23071703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wu Y.S., Duan S.Y., Zhao L.P., Gao Z., Luo M., Song S.R., Xu W.P., Zhang C.X., Ma C., Wang S.P. Aroma characterization based on aromatic series analysis in table grapes. Sci. Rep. 2016;6:31116. doi: 10.1038/srep31116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Guth H. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 1997;45:3027–3032. doi: 10.1021/jf970280a. [DOI] [Google Scholar]
- 33.Ong P.K.C., Acree T.E. Similarities in the aroma chemistry of Gewürztraminer variety wines and Lychee (Litchi chinesis Sonn.) Fruit. J. Agric. Food Chem. 1999;47:665–670. doi: 10.1021/jf980452j. [DOI] [PubMed] [Google Scholar]
- 34.Yamamoto T., Matsuda H., Utsumi Y., Hagiwara T., Kanisawa T. Synthesis and odor of optically active rose oxide. Tetrahedron Lett. 2002;43:9077–9080. doi: 10.1016/S0040-4039(02)02311-0. [DOI] [Google Scholar]
- 35.Girard B., Fukumoto L., Mazza G., Delaquis P., Ewert B. Volatile terpene constituents in maturing Gewurztraminer grapes from British Columbia. Am. J. Enol. Vitic. 2002;53:99–109. [Google Scholar]
- 36.Fenoll J., Manso A., Hellin P., Ruiz L., Flores P. Changes in the aromatic composition of the Vitis vinifera grape Muscat Hamburg during ripening. Food Chem. 2009;114:420–428. doi: 10.1016/j.foodchem.2008.09.060. [DOI] [Google Scholar]
- 37.Ruiz-Garcia L., Hellin P., Flores P., Fenoll J. Prediction of Muscat aroma in table grape by analysis of rose oxide. Food Chem. 2014;154:151–157. doi: 10.1016/j.foodchem.2014.01.005. [DOI] [PubMed] [Google Scholar]
- 38.Skinkis P.A., Bordelon B.P., Wood K.V. Comparison of Monoterpene Constituents in Traminette, Gewurztraminer, and Riesling Winegrapes. Am. J. Enol. Vitic. 2008;59:440–445. [Google Scholar]
- 39.Shure K.B., Acree T.E. Changes in the Odor-Active Compounds in Vitis-Labruscana Cv Concord During Growth And Development. J. Agric. Food Chem. 1994;42:350–353. doi: 10.1021/jf00038a022. [DOI] [Google Scholar]
- 40.Kobayashi H., Sasaki K., Tanzawa F., Matsuyama S., Suzuki S., Takata R., Saito H. Impact of harvest timing on 4-hydroxy-2,5-dimethyl-3(2H)-furanone concentration in ‘Muscat Bailey A’ grape berries. Vitis. 2013;52:9–11. [Google Scholar]
- 41.Sale J.W., Wilson J.B. Distribution of volatile flavor in grapes and grape juices. J. Agric. Res. 1926;33:0301–0310. doi: 10.1016/S0016-0032(26)91735-3. [DOI] [Google Scholar]
- 42.Acree T.E., Lavin E.H., Nishida R., Watanabe S. o-Amino Acetophenone the Foxy Smelling Component of Labruscana Grapes. In: Bessiere Y., Thomas A.F., editors. Flavour Science and Technology—6th Weurmann Symposium. Wiley; Hoboken, NJ, USA: 1990. pp. 49–52. [Google Scholar]
- 43.Massa M.J., Robacker D.C., Patt J. Identification of grape juice aroma volatiles and attractiveness to the Mexican fruit fly (Diptera: Tephritidae) Fla. Entomol. 2008;91:266–276. doi: 10.1653/0015-4040(2008)91[266:IOGJAV]2.0.CO;2. [DOI] [Google Scholar]
- 44.Rapp A., Versini G., Ullemeyer H. 2-Aminoacetophenone—Causal Component of Untypical Aging Flavor (Naphthalene Note, Hybrid Note) Of Wine. Vitis. 1993;32:61–62. [Google Scholar]
- 45.Baek H.H., Cadwallader K.R., Marroquin E., Silva J.L. Identification of predominant aroma compounds in muscadine grape juice. J. Food Sci. 1997;62:249–252. doi: 10.1111/j.1365-2621.1997.tb03978.x. [DOI] [Google Scholar]
- 46.Baek H.H., Cadwallader K.R. Contribution of free and glycosidically bound volatile compounds to the aroma of muscadine grape juice. J. Food Sci. 1999;64:441–444. doi: 10.1111/j.1365-2621.1999.tb15059.x. [DOI] [Google Scholar]
- 47.Yang C.X., Wang Y.J., Wu B.H., Fang J.B., Li S.H. Volatile compounds evolution of three table grapes with different flavour during and after maturation. Food Chem. 2011;128:823–830. doi: 10.1016/j.foodchem.2010.11.029. [DOI] [Google Scholar]
- 48.Buttery R.G., Teranishi R., Ling L.C., Turnbaugh J.G. Quantitative and Sensory Studies on Tomato Paste Volatiles. J. Agric. Food Chem. 1990;38:336–340. doi: 10.1021/jf00091a074. [DOI] [Google Scholar]
- 49.Takeoka G.R., Flath R.A., Mon T.R., Teranishi R., Guentert M. Volatile Constituents of Apricot (Prunus-Armeniaca) J. Agric. Food Chem. 1990;38:471–477. doi: 10.1021/jf00092a031. [DOI] [Google Scholar]
- 50.Buttery R.G., Ling L.C. Importance Of 2-Aminoacetophenone to the Flavor of Masa Corn Flour Products. J. Agric. Food Chem. 1994;42:1–2. doi: 10.1021/jf00037a001. [DOI] [Google Scholar]
- 51.Hirvi T., Honkanen E. The Volatiles of 2 New Strawberry Cultivars, Annelie and Alaska Pioneer, Obtained by Backcrossing of Cultivated Strawberries with Wild Strawberries, Fragaria-Vesca, Rugen and Fragaria-Virginiana. Z. Lebensmittel Unters. Forsch. 1982;175:113–116. doi: 10.1007/BF01135046. [DOI] [Google Scholar]
- 52.Iyer M.M., Sacks G.L., Padilla-Zakour O.I. Assessment of the Validity of Maturity Metrics for Predicting the Volatile Composition of Concord Grape Juice. J. Food Sci. 2012;77:C319–C325. doi: 10.1111/j.1750-3841.2011.02592.x. [DOI] [PubMed] [Google Scholar]
- 53.Depinho P.G., Bertrand A. Analytical Determination of Furaneol (2,5-Dimethyl-4-Hydroxy-3(2h)-Furanone)—Application to Differentiation of White Wines from Hybrid and Various Vitis-Vinifera Cultivars. Am. J. Enol. Vitic. 1995;46:181–186. [Google Scholar]
- 54.Rapp A., Engel L. Determination and Detection of Furaneol (2,5-Dimethyl-4-Hydroxy-3-Furanon) in Wines from Vitis-Vinifera Varieties. Vitis. 1995;34:71–72. [Google Scholar]
- 55.Drappier J., Thibon C., Rabot A., Geny-Denis L. Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming-Review. Crit. Rev. Food Sci. Nutr. 2019;59:14–30. doi: 10.1080/10408398.2017.1355776. [DOI] [PubMed] [Google Scholar]
- 56.Ruiz M.J., Moyano L., Zea L. Changes in aroma profile of musts from grapes cv. Pedro Ximenez chamber-dried at controlled conditions destined to the production of sweet Sherry wine. LWT Food Sci. Technol. 2014;59:560–565. doi: 10.1016/j.lwt.2014.04.056. [DOI] [Google Scholar]
- 57.Wang D., Cai J., Zhu B.Q., Wu G.F., Duan C.Q., Chen G., Shi Y. Study of free and glycosidically bound volatile compounds in air-dried raisins from three seedless grape varieties using HS-SPME with GC-MS. Food Chem. 2015;177:346–353. doi: 10.1016/j.foodchem.2015.01.018. [DOI] [PubMed] [Google Scholar]
- 58.Javed H.U., Wang D., Wu G.F., Kaleem Q.M., Duan C.Q., Shi Y. Post-storage changes of volatile compounds in air- and sun-dried raisins with different packaging materials using HS-SPME with GC/MS. Food Res. Int. 2019;119:23–33. doi: 10.1016/j.foodres.2019.01.007. [DOI] [PubMed] [Google Scholar]
- 59.Campo E., Cacho J., Ferreira V. The chemical characterization of the aroma of dessert and sparkling white wines (Pedro Ximenez, Fino, Sauternes, and Cava) by gas chromatography-olfactometry and chemical quantitative analysis. J. Agric. Food Chem. 2008;56:2477–2484. doi: 10.1021/jf072968l. [DOI] [PubMed] [Google Scholar]
- 60.Coelho E., Rocha S.M., Delgadillo I., Coimbra M.A. Headspace-SPME applied to varietal volatile components evolution during Vitis vinifera L. cv. ‘Baga’ ripening. Anal. Chim. Acta. 2006;563:204–214. doi: 10.1016/j.aca.2005.11.018. [DOI] [Google Scholar]
- 61.Yuan F., Qian M.C. Development of C13-norisoprenoids, carotenoids and other volatile compounds in Vitis vinifera L. Cv. Pinot noir grapes. Food Chem. 2016;192:633–641. doi: 10.1016/j.foodchem.2015.07.050. [DOI] [PubMed] [Google Scholar]
- 62.Lukic I., Radeka S., Grozaj N., Staver M., Persuric D. Changes in physico-chemical and volatile aroma compound composition of Gewurztraminer wine as a result of late and ice harvest. Food Chem. 2016;196:1048–1057. doi: 10.1016/j.foodchem.2015.10.061. [DOI] [PubMed] [Google Scholar]
- 63.Luo J.Q., Brotchie J., Pang M., Marriott P.J., Howell K., Zhang P.Z. Free terpene evolution during the berry maturation of five Vitis vinifera L. cultivars. Food Chem. 2019;299:125101. doi: 10.1016/j.foodchem.2019.125101. [DOI] [PubMed] [Google Scholar]
- 64.Šuklje K., Zhang X., Antalick G., Clark A.C., Deloire A., Schmidtke L.M. Berry Shriveling Significantly Alters Shiraz (Vitis vinifera L.) Grape and Wine Chemical Composition. J. Agric. Food Chem. 2016;64:870–880. doi: 10.1021/acs.jafc.5b05158. [DOI] [PubMed] [Google Scholar]
- 65.Chou H.C., Šuklje K., Antalick G., Schmidtke L.M., Blackman J.W. Late-Season Shiraz Berry Dehydration That Alters Composition and Sensory Traits of Wine. J. Agric. Food Chem. 2018;66:7750–7757. doi: 10.1021/acs.jafc.8b01646. [DOI] [PubMed] [Google Scholar]
- 66.Slaghenaufi D., Ugliano M. Norisoprenoids, Sesquiterpenes and Terpenoids Content of Valpolicella Wines During Aging: Investigating Aroma POtential in Relationship to Evolution of Tobacco and Balsamic Aroma in Aged Wine. Front. Chem. 2018;6:66. doi: 10.3389/fchem.2018.00066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lee S.H., Seo M.J., Riu M., Cotta J.P., Block D.E., Dokoozlian N.K., Ebeler S.E. Vine microclimate and norisoprenoid concentration in cabernet sauvignon grapes and wines. Am. J. Enol. Vitic. 2007;58:291–301. [Google Scholar]
- 68.Song J.Q., Smart R., Wang H., Dambergs B., Sparrow A., Qian M.C. Effect of grape bunch sunlight exposure and UV radiation on phenolics and volatile composition of Vitis vinifera L. cv. Pinot noir wine. Food Chem. 2015;173:424–431. doi: 10.1016/j.foodchem.2014.09.150. [DOI] [PubMed] [Google Scholar]
- 69.Pineau B., Barbe J.-C., Van Leeuwen C., Dubourdieu D. Which impact for beta-damascenone on red wines aroma? J. Agric. Food Chem. 2007;55:4103–4108. doi: 10.1021/jf070120r. [DOI] [PubMed] [Google Scholar]
- 70.San-Juan F., Ferreira V., Cacho J., Escudero A. Quality and Aromatic Sensory Descriptors (Mainly Fresh and Dry Fruit Character) of Spanish Red Wines can be Predicted from their Aroma-Active Chemical Composition. J. Agric. Food Chem. 2011;59:7916–7924. doi: 10.1021/jf1048657. [DOI] [PubMed] [Google Scholar]
- 71.Juan F.S., Cacho J., Ferreira V., Escudero A. Aroma Chemical Composition of Red Wines from Different Price Categories and Its Relationship to Quality. J. Agric. Food Chem. 2012;60:5045–5056. doi: 10.1021/jf2050685. [DOI] [PubMed] [Google Scholar]
- 72.Ferreira V., Lopez R., Cacho J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000;80:1659–1667. doi: 10.1002/1097-0010(20000901)80:11<1659::AID-JSFA693>3.0.CO;2-6. [DOI] [Google Scholar]
- 73.Sacks G.L., Gates M.J., Ferry F.X., Lavin E.H., Kurtz A.J., Acree T.E. Sensory threshold of 1,1,6-trimethyl-1,2-dihydronaphthalene (TDN) and concentrations in young Riesling and non-Riesling wines. J. Agric. Food Chem. 2012;60:2998–3004. doi: 10.1021/jf205203b. [DOI] [PubMed] [Google Scholar]
- 74.Black C., Francis L., Henschke P., Capone D., Anderson S., Day M., Holt H., Pearson W., Herderich M., Johnson D. Aged Riesling and the development of TDN. Wine Vitic. J. 2012;27:20–26. [Google Scholar]
- 75.Janusz A., Capone D.L., Puglisi C.J., Perkins M.V., Elsey G.M., Sefton M.A. (E)-1-(2,3,6-trimethylphenyl)buta-1,3-diene: A potent grape-derived odorant in wine. J. Agric. Food Chem. 2003;51:7759–7763. doi: 10.1021/jf0347113. [DOI] [PubMed] [Google Scholar]
- 76.Cox A., Capone D.L., Elsey G.M., Perkins M.V., Sefton M.A. Quantitative analysis, occurrence, and stability of (E)-1-(2,3,6-Trimethylphenyl)buta-1,3-diene in wine. J. Agric. Food Chem. 2005;53:3584–3591. doi: 10.1021/jf0479057. [DOI] [PubMed] [Google Scholar]
- 77.Sarrazin E., Dubourdieu D., Darriet P. Characterization of key-aroma compounds of botrytized wines, influence of grape botrytization. Food Chem. 2007;103:536–545. doi: 10.1016/j.foodchem.2006.08.026. [DOI] [Google Scholar]
- 78.Tosi E., Fedrizzi B., Azzolini M., Finato F., Simonato B., Zapparoli G. Effects of noble rot on must composition and aroma profile of Amarone wine produced by the traditional grape withering protocol. Food Chem. 2012;130:370–375. doi: 10.1016/j.foodchem.2011.07.053. [DOI] [Google Scholar]
- 79.Furdikova K., Machynakova A., Drtilova T., Klempova T., Durcanska K., Spanik I. Comparison of volatiles in noble-rotten and healthy grape berries of Tokaj. LWT Food Sci. Technol. 2019;105:37–47. doi: 10.1016/j.lwt.2019.01.055. [DOI] [Google Scholar]
- 80.Pons A., Lavigne V., Eric F., Darriet P., Dubourdieu D. Identification of volatile compounds responsible for prune aroma in prematurely aged red wines. J. Agric. Food Chem. 2008;56:5285–5290. doi: 10.1021/jf073513z. [DOI] [PubMed] [Google Scholar]
- 81.Allamy L., Darriet P., Pons A. Molecular interpretation of dried-fruit aromas in Merlot and Cabernet Sauvignon musts and young wines: Impact of over-ripening. Food Chem. 2018;266:245–253. doi: 10.1016/j.foodchem.2018.06.022. [DOI] [PubMed] [Google Scholar]
- 82.Bowen A.J., Reynolds A.G. Aroma compounds in Ontario Vidal and Riesling icewines. I. Effects of harvest date. Food Res. Int. 2015;76:540–549. doi: 10.1016/j.foodres.2015.06.046. [DOI] [PubMed] [Google Scholar]
- 83.Javed H.U., Wang D., Shi Y., Wu G.F., Xie H., Pan Y.Q., Duan C.Q. Changes of free-form volatile compounds in pre-treated raisins with different packaging materials during storage. Food Res. Int. 2018;107:649–659. doi: 10.1016/j.foodres.2018.03.019. [DOI] [PubMed] [Google Scholar]
- 84.Pons A., Allamy L., Lavigne V., Dubourdieu D., Darriet P. Study of the contribution of massoia lactone to the aroma of Merlot and Cabernet Sauvignon musts and wines. Food Chem. 2017;232:229–236. doi: 10.1016/j.foodchem.2017.03.151. [DOI] [PubMed] [Google Scholar]
- 85.D’Onofrio C. Changes in volatile compounds. In: Mencarelli F., Tonutti P., editors. Sweet, Reinforced, and Fortified Wines. Wiley & Sons; Chichester, UK: 2013. pp. 91–103. [Google Scholar]
- 86.Noguerol-Pato R., González-Álvarez M., González-Barreiro C., Cancho-Grande B., Simal-Gándara J. Evolution of the aromatic profile in Garnacha Tintorera grapes during raisining and comparison with that of the naturally sweet wine obtained. Food Chem. 2013;139:1052–1061. doi: 10.1016/j.foodchem.2012.12.048. [DOI] [PubMed] [Google Scholar]
- 87.D’Onofrio C., Matarese F., Scalabrelli G., Boss P. Functional characterization of terpene synthases of ‘aromatic’ and ‘non-aromatic’ grapevine varieties; Proceedings of the 10th International Conference on Grapevine Breeding and Genetics; Geneva, NY, USA. 1–5 August 2010; pp. 557–563. [Google Scholar]
- 88.Ruiz M.J., Zea L., Moyano L., Medina M. Aroma active compounds during the drying of grapes cv. Pedro Ximenez destined to the production of sweet Sherry wine. Eur. Food Res. Technol. 2010;230:429–435. doi: 10.1007/s00217-009-1183-0. [DOI] [Google Scholar]
- 89.Schelezki O.J., Smith P.A., Hranilovic A., Bindon K.A., Jeffery D.W. Comparison of consecutive harvests versus blending treatments to produce lower alcohol wines from Cabernet Sauvignon grapes: Impact on polysaccharide and tannin content and composition. Food Chem. 2018;244:50–59. doi: 10.1016/j.foodchem.2017.10.024. [DOI] [PubMed] [Google Scholar]
- 90.Bellincontro A., De Santis D., Botondi R., Villa I., Mencarelli F. Different postharvest dehydration rates affect quality characteristics and volatile compounds of Malvasia, Trebbiano and Sangiovese grapes for wine production. J. Sci. Food Agric. 2004;84:1791–1800. doi: 10.1002/jsfa.1889. [DOI] [Google Scholar]
- 91.Franco M., Peinado R.A., Medina M., Moreno J. Off-vine grape drying effect on volatile compounds and aromatic series in must from Pedro Ximénez grape variety. J. Agric. Food Chem. 2004;52:3905–3910. doi: 10.1021/jf0354949. [DOI] [PubMed] [Google Scholar]
- 92.Bayonove C., Cordonnier R., Dubois P. Study of an aromatic characteristic fraction of cabernet sauvignon grape variety, identification of 2-methoxy-3-isobutyl-pyrazine. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences. 1975;281:75–78. [Google Scholar]
- 93.Lacey M.J., Allen M.S., Harris R.L.N., Brown W.V. Methoxypyrazines in Sauvignon Blanc Grapes and Wines. Am. J. Enol. Vitic. 1991;42:103–108. [Google Scholar]
- 94.De Boubee D.R., Van Leeuwen C., Dubourdieu D. Organoleptic impact of 2-methoxy-3-isobutylpyrazine on red Bordeaux and Loire wines. Effect of environmental conditions on concentrations in grapes during ripening. J. Agric. Food Chem. 2000;48:4830–4834. doi: 10.1021/jf000181o. [DOI] [PubMed] [Google Scholar]
- 95.Belancic A., Agosin E. Methoxypyrazines in grapes and wines of Vitis vinifera cv. Carmenere. Am. J. Enol. Vitic. 2007;58:462–469. [Google Scholar]
- 96.Mendez-Costabel M.P., Wilkinson K.L., Bastian S.E.P., McCarthy M., Ford C.M., Dokoozlian N. Seasonal and Regional Variation of Green Aroma Compounds in Commercial Vineyards of Vitis vinifera L. Merlot in California. Am. J. Enol. Vitic. 2013;64:430–436. doi: 10.5344/ajev.2013.12109. [DOI] [Google Scholar]
- 97.Falcao L.D., de Revel G., Perello M.C., Moutsiou A., Zanus M.C., Bordignon-Luiz M.T. A survey of seasonal temperatures and vineyard altitude influences on 2-methoxy-3-isobutylpyrazine, C-13-norisoprenoids, and the sensory profile of Brazilian Cabernet Sauvignon wines. J. Agric. Food Chem. 2007;55:3605–3612. doi: 10.1021/jf070185u. [DOI] [PubMed] [Google Scholar]
- 98.Ryona I., Pan B.S., Intrigliolo D.S., Lakso A.N., Sacks G.L. Effects of Cluster Light Exposure on 3-Isobutyl-2-methoxypyrazine Accumulation and Degradation Patterns in Red Wine Grapes (Vitis vinifera L. Cv. Cabernet Franc) J. Agric. Food Chem. 2008;56:10838–10846. doi: 10.1021/jf801877y. [DOI] [PubMed] [Google Scholar]
- 99.Gregan S.M., Jordan B. Methoxypyrazine Accumulation and O-Methyltransferase Gene Expression in Sauvignon Blanc Grapes: The Role of Leaf Removal, Light Exposure, and Berry Development. J. Agric. Food Chem. 2016;64:2200–2208. doi: 10.1021/acs.jafc.5b05806. [DOI] [PubMed] [Google Scholar]
- 100.Helwi P., Habran A., Guillaumie S., Thibon C., Hilbert G., Gomes E., Delrot S., Darriet P., van Leeuwen C. Vine Nitrogen Status Does Not Have a Direct Impact on 2-Methoxy-3-isobutylpyrazine in Grape Berries and Wines. J. Agric. Food Chem. 2015;63:9789–9802. doi: 10.1021/acs.jafc.5b03838. [DOI] [PubMed] [Google Scholar]
- 101.Koegel S., Botezatu A., Hoffmann C., Pickering G. Methoxypyrazine composition of Coccinellidae-tainted Riesling and Pinot noir wine from Germany. J. Sci. Food Agric. 2015;95:509–514. doi: 10.1002/jsfa.6760. [DOI] [PubMed] [Google Scholar]
- 102.Gracia-Moreno E. Ph.D. Thesis. Universidad de Zaragoza; Zaragoza, Spain: 2015. Nuevos Métodos Analíticos para la Determinación Selectiva de Pirazinas, Ácidos y Otros Compuestos de Interés Aromático Presentes en Cantidades Traza. [Google Scholar]
- 103.Buttery R.G., Seifert R.M., Guadagni D.G., Ling L.C. Characterization of some volatile constituents of bell peppers. J. Agric. Food Chem. 1969;17:1322–1327. doi: 10.1021/jf60166a061. [DOI] [PubMed] [Google Scholar]
- 104.Pickering G.J., Karthik A., Inglis D., Sears M., Ker K. Detection thresholds for 2-isopropyl-3-methoxypyrazine in Concord and Niagara grape juice. J. Food Sci. 2008;73:S262–S266. doi: 10.1111/j.1750-3841.2008.00847.x. [DOI] [PubMed] [Google Scholar]
- 105.Allen M.S., Lacey M.J., Harris R.L.N., Brown W.V. Contribution of Methoxypyrazines to Sauvignon Blanc Wine Aroma. Am. J. Enol. Vitic. 1991;42:109–112. [Google Scholar]
- 106.Sidhu D., Lund J., Kotseridis Y., Saucier C. Methoxypyrazine Analysis and Influence of Viticultural and Enological Procedures on their Levels in Grapes, Musts, and Wines. Crit. Rev. Food Sci. Nutr. 2015;55:485–502. doi: 10.1080/10408398.2012.658587. [DOI] [PubMed] [Google Scholar]
- 107.Kotseridis Y., Baumes R. Identification of impact odorants in Bordeaux red grape juice, in the commercial yeast used for its fermentation, and in the produced wine. J. Agric. Food Chem. 2000;48:400–406. doi: 10.1021/jf990565i. [DOI] [PubMed] [Google Scholar]
- 108.Oliveira J.M., Faria M., Sa F., Barros F., Araujo I.A. C-6-alcohols as varietal markers for assessment of wine origin. Anal. Chim. Acta. 2006;563:300–309. doi: 10.1016/j.aca.2005.12.029. [DOI] [Google Scholar]
- 109.Noguerol-Pato R., Gonzalez-Barreiro C., Cancho-Grande B., Martinez M.C., Santiago J.L., Simal-Gandara J. Floral, spicy and herbaceous active odorants in Gran Negro grapes from shoulders and tips into the cluster, and comparison with Brancellao and Mouraton varieties. Food Chem. 2012;135:2771–2782. doi: 10.1016/j.foodchem.2012.06.104. [DOI] [PubMed] [Google Scholar]
- 110.Meng J.F., Xu T.F., Song C.Z., Li X.L., Yue T.X., Qin M.Y., Fang Y.L., Zhang Z.W., Xi Z.M. Characteristic free aromatic components of nine clones of spine grape (Vitis davidii Foex) from Zhongfang County (China) Food Res. Int. 2013;54:1795–1800. doi: 10.1016/j.foodres.2013.09.039. [DOI] [Google Scholar]
- 111.Feng H., Yuan F., Skinkis P.A., Qian M.C. Influence of cluster zone leaf removal on Pinot noir grape chemical and volatile composition. Food Chem. 2015;173:414–423. doi: 10.1016/j.foodchem.2014.09.149. [DOI] [PubMed] [Google Scholar]
- 112.Yuan F., Schreiner R.P., Qian M.C. Soil Nitrogen, Phosphorus, and Potassium Alter β-Damascenone and Other Volatiles in Pinot noir Berries. Am. J. Enol. Vitic. 2018;69:157–166. doi: 10.5344/ajev.2017.17071. [DOI] [Google Scholar]
- 113.Buttery R.G., Turnbaugh J.G., Ling L.C. Contribution of volatiles to rice aroma. J. Agric. Food Chem. 1988;36:1006–1009. doi: 10.1021/jf00083a025. [DOI] [Google Scholar]
- 114.Hansen M., Cantwell M.I., Buttery R.G., Stern D.J., Ling L.C. Broccoli Storage under Low-Oxygen Atmosphere: Identification of Higher Boiling Volatiles. J. Agric. Food Chem. 1992;40:850–852. doi: 10.1021/jf00017a029. [DOI] [Google Scholar]
- 115.Teranishi R., Buttery R.G., Guadagni D.G. Odor quality and chemical structure in fruit and vegetable flavors. Ann. N. Y. Acad. Sci. 1974;237:209–216. doi: 10.1111/j.1749-6632.1974.tb49855.x. [DOI] [PubMed] [Google Scholar]
- 116.Preston L.D., Block D.E., Heymann H., Soleas G., Noble A.C., Ebeler S.E. Defining vegetal aromas in Cabernet Sauvignon using sensory and chemical evaluations. Am. J. Enol. Vitic. 2008;59:137–145. [Google Scholar]
- 117.Escudero A., Campo E., Farina L., Cacho J., Ferreira V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007;55:4501–4510. doi: 10.1021/jf0636418. [DOI] [PubMed] [Google Scholar]
- 118.Capone D.L., Jeffery D.W., Sefton M.A. Vineyard and fermentation studies to elucidate the origin of 1,8-cineole in Australian red wine. J. Agric. Food Chem. 2012;60:2281–2287. doi: 10.1021/jf204499h. [DOI] [PubMed] [Google Scholar]
- 119.Poitou X., Thibon C., Darriet P. 1,8-Cineole in French Red Wines: Evidence for a Contribution Related to Its Various Origins. J. Agric. Food Chem. 2017;65:383–393. doi: 10.1021/acs.jafc.6b03042. [DOI] [PubMed] [Google Scholar]
- 120.Capone D.L., Sefton M.A., Jeffery D.W., Francis I.L. Terroir or terpenoid transformation: The origin of 1,8-cineole (eucalyptol) in wine; Proceedings of the 10th Wartburg Symposium on Flavor Chemistry and biology; Eisenach, Germany. 16–19 April 2013; pp. 130–136. [Google Scholar]
- 121.Farina L., Boido E., Carrau F., Versini G., Dellacassa E. Terpene compounds as possible precursors of 1,8-cineole in red grapes and wines. J. Agric. Food Chem. 2005;53:1633–1636. doi: 10.1021/jf040332d. [DOI] [PubMed] [Google Scholar]
- 122.Stevens K.L., Bomben J.L., McFadden W.H. Volatiles from Grapes. Vitis Vinifera (Linn.) Cultivar Grenache. J. Agric. Food Chem. 1967;15:378–380. doi: 10.1021/jf60151a029. [DOI] [Google Scholar]
- 123.Gomez E., Martinez A., Laencina J. Changes in volatile compounds during maturation of some grape varieties. J. Sci. Food Agric. 1995;67:229–233. doi: 10.1002/jsfa.2740670213. [DOI] [Google Scholar]
- 124.Ferrandino A., Carlomagno A., Baldassarre S., Schubert A. Varietal and pre-fermentative volatiles during ripening of Vitis vinifera cv Nebbiolo berries from three growing areas. Food Chem. 2012;135:2340–2349. doi: 10.1016/j.foodchem.2012.06.061. [DOI] [PubMed] [Google Scholar]
- 125.Genovese A., Lamorte S.A., Gambuti A., Moio L. Aroma of Aglianico and Uva di Troia grapes by aromatic series. Food Res. Int. 2013;53:15–23. doi: 10.1016/j.foodres.2013.03.051. [DOI] [Google Scholar]
- 126.Perestrelo R., Caldeira M., Camara J.S. Solid phase microextraction as a reliable alternative to conventional extraction techniques to evaluate the pattern of hydrolytically released components in Vitis vinifera L. grapes. Talanta. 2012;95:1–11. doi: 10.1016/j.talanta.2012.03.005. [DOI] [PubMed] [Google Scholar]
- 127.Loscos N., Hernandez-Orte P., Cacho J., Ferreira V. Release and formation of varietal aroma compounds during alcoholic fermentation from nonfloral grape odorless flavor precursors fractions. J. Agric. Food Chem. 2007;55:6674–6684. doi: 10.1021/jf0702343. [DOI] [PubMed] [Google Scholar]
- 128.Wood C., Siebert T.E., Parker M., Capone D.L., Elsey G.M., Pollnitz A.P., Eggers M., Meier M., Vossing T., Widder S., et al. From wine to pepper: Rotundone, an obscure sesquiterpene, is a potent spicy aroma compound. J. Agric. Food Chem. 2008;56:3738–3744. doi: 10.1021/jf800183k. [DOI] [PubMed] [Google Scholar]
- 129.Zhang P., Barlow S., Krstic M., Herderich M., Fuentes S., Howell K. Within-Vineyard, Within-Vine, and Within-Bunch Variability of the Rotundone Concentration in Berries of Vitis vinifera L. cv. Shiraz. J. Agric. Food Chem. 2015;63:4276–4283. doi: 10.1021/acs.jafc.5b00590. [DOI] [PubMed] [Google Scholar]
- 130.Geffroy O., Descôtes J., Levassseur-Garcia C., Debord C., Denux J.-P., Dufourcq T. A 2-year multisite study of viticultural and environmental factors affecting rotundone concentration in Duras red wine. OENO One. 2019;53:457–470. doi: 10.20870/oeno-one.2019.53.3.2341. [DOI] [Google Scholar]
- 131.Huang A.C., Burrett S., Sefton M.A., Taylor D.K. Production of the pepper aroma compound, (-)-rotundone, by aerial oxidation of alpha-guaiene. J. Agric. Food Chem. 2014;62:10809–10815. doi: 10.1021/jf504693e. [DOI] [PubMed] [Google Scholar]
- 132.Cullere L., Ontanon I., Escudero A., Ferreira V. Straightforward strategy for quantifying rotundone in wine at ngL(-1) level using solid-phase extraction and gas chromatography-quadrupole mass spectrometry. Occurrence in different varieties of spicy wines. Food Chem. 2016;206:267–273. doi: 10.1016/j.foodchem.2016.03.039. [DOI] [PubMed] [Google Scholar]
- 133.Geffroy O., Descôtes J., Serrano E., Li Calzi M., Dagan L., Schneider R. Can a certain concentration of rotundone be undesirable in Duras red wine? A study to estimate a consumer rejection threshold for the pepper aroma compound. Aust. J. Grape Wine Res. 2018;24:88–95. doi: 10.1111/ajgw.12299. [DOI] [Google Scholar]
- 134.Roberts D.D., Mordehai A.P., Acree T.E. Detection and Partial Characterization of 8 Beta-Damascenone Precursors in Apples (Malus-Domestica Borkh, Cv Empire) J. Agric. Food Chem. 1994;42:345–349. doi: 10.1021/jf00038a021. [DOI] [Google Scholar]
- 135.Picard M., de Revel G., Marchand S. First identification of three p-menthane lactones and their potential precursor, menthofuran, in red wines. Food Chem. 2017;217:294–302. doi: 10.1016/j.foodchem.2016.08.070. [DOI] [PubMed] [Google Scholar]
- 136.Carlomagno A., Schubert A., Ferrandino A. Screening and evolution of volatile compounds during ripening of ‘Nebbiolo’, ‘Dolcetto’ and ‘Barbera’ (Vitis vinifera L.) neutral grapes by SBSE-GC/MS. Eur. Food Res. Technol. 2016;242:1221–1233. doi: 10.1007/s00217-015-2626-4. [DOI] [Google Scholar]
- 137.Garcia-Carpintero E.G., Sanchez-Palomo E., Gallego M.A.G., Gonzalez-Vinas M.A. Free and bound volatile compounds as markers of aromatic typicalness of Moravia Dulce, Rojal and Tortosi red wines. Food Chem. 2012;131:90–98. doi: 10.1016/j.foodchem.2011.08.035. [DOI] [Google Scholar]
- 138.Ugliano M., Moio L. Free and hydrolytically released volatile compounds of Vitis vinifera L. cv. Fiano grapes as odour-active constituents of Fiano wine. Anal. Chim. Acta. 2008;621:79–85. doi: 10.1016/j.aca.2008.03.002. [DOI] [PubMed] [Google Scholar]
- 139.Botelho G., Mendes-Faia A., Climaco M.C. Characterisation of free and glycosidically bound odourant compounds of Aragonez clonal musts by GC-O. Anal. Chim. Acta. 2010;657:198–203. doi: 10.1016/j.aca.2009.10.030. [DOI] [PubMed] [Google Scholar]
- 140.Cabrita M.J., Freitas A.M.C., Laureano O., Di Stefano R. Glycosidic aroma compounds of some Portuguese grape cultivars. J. Sci. Food Agric. 2006;86:922–931. doi: 10.1002/jsfa.2439. [DOI] [Google Scholar]
- 141.Schneider R., Razungles A., Augier C., Baumes R. Monoterpenic and norisoprenoidic glycoconjugates of Vitis vinifera L. cv. Melon B. as precursors of odorants in Muscadet wines. J. Chromatogr. A. 2001;936:145–157. doi: 10.1016/S0021-9673(01)01150-5. [DOI] [PubMed] [Google Scholar]
- 142.Lopez R., Ezpeleta E., Sanchez I., Cacho J., Ferreira V. Analysis of the aroma intensities of volatile compounds released from mild acid hydrolysates of odourless precursors extracted from Tempranillo and Grenache grapes using gas chromatography-olfactometry. Food Chem. 2004;88:95–103. doi: 10.1016/j.foodchem.2004.01.025. [DOI] [Google Scholar]
- 143.Oliveira I., Ferreira V. Modulating Fermentative, Varietal and Aging Aromas of Wine Using non-Saccharomyces Yeasts in a Sequential Inoculation Approach. Microorganisms. 2019;7:164. doi: 10.3390/microorganisms7060164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Wirth J., Guo W.F., Baumes R., Gunata Z. Volatile compounds released by enzymatic hydrolysis of glycoconjugates of leaves and grape berries from Vitis vinifera Muscat of Alexandria and Shiraz cultivars. J. Agric. Food Chem. 2001;49:2917–2923. doi: 10.1021/jf001398l. [DOI] [PubMed] [Google Scholar]
- 145.Torchio F., Giacosa S., Vilanova M., Segade S.R., Gerbi V., Giordano M., Rolle L. Use of response surface methodology for the assessment of changes in the volatile composition of Moscato bianco (Vitis vinifera L.) grape berries during ripening. Food Chem. 2016;212:576–584. doi: 10.1016/j.foodchem.2016.05.191. [DOI] [PubMed] [Google Scholar]
- 146.Crespo J., Rigou P., Romero V., Garcia M., Arroyo T., Cabellos J.M. Effect of seasonal climate fluctuations on the evolution of glycoconjugates during the ripening period of grapevine cv. Muscat a petits grains blancs berries. J. Sci. Food Agric. 2018;98:1803–1812. doi: 10.1002/jsfa.8656. [DOI] [PubMed] [Google Scholar]
- 147.D’Onofrio C., Matarese F., Cuzzola A. Study of the terpene profile at harvest and during berry development of Vitis vinifera L. aromatic varieties Aleatico, Brachetto, Malvasia di Candia aromatica and Moscato bianco. J. Sci. Food Agric. 2017;97:2898–2907. doi: 10.1002/jsfa.8126. [DOI] [PubMed] [Google Scholar]
- 148.Sefton M.A., Francis I.L., Williams P.J. The Volatile Composition of Chardonnay Juices—A Study by Flavor Precursor Analysis. Am. J. Enol. Vitic. 1993;44:359–370. [Google Scholar]
- 149.Picard M., Lytra G., Tempere S., Barbe J.C., de Revel G., Marchand S. Identification of Piperitone as an Aroma Compound Contributing to the Positive Mint Nuances Perceived in Aged Red Bordeaux Wines. J. Agric. Food Chem. 2016;64:451–460. doi: 10.1021/acs.jafc.5b04869. [DOI] [PubMed] [Google Scholar]
- 150.Garcia-Munoz S., Asproudi A., Cabello F., Borsa D. Aromatic characterization and enological potential of 21 minor varieties (Vitis vinifera L.) Eur. Food Res. Technol. 2011;233:473–481. doi: 10.1007/s00217-011-1538-1. [DOI] [Google Scholar]
- 151.Garcia-Carpintero E.G., Sanchez-Palomo E., Gallego M.A.G., Gonzalez-Vinas M.A. Volatile and sensory characterization of red wines from cv. Moravia Agria minority grape variety cultivated in La Mancha region over five consecutive vintages. Food Res. Int. 2011;44:1549–1560. doi: 10.1016/j.foodres.2011.04.022. [DOI] [Google Scholar]
- 152.Garcia-Carpintero E.G., Sanchez-Palomo E., Gonzalez-Vinas M.A. Aroma characterization of red wines from cv. Bobal grape variety grown in La Mancha region. Food Res. Int. 2011;44:61–70. doi: 10.1016/j.foodres.2010.11.013. [DOI] [Google Scholar]
- 153.Gracia-Moreno E., Lopez R., Ferreira V. Determination of 2-, 3-, 4-methylpentanoic and cyclohexanecarboxylic acids in wine: Development of a selective method based on solid phase extraction and gas chromatography-negative chemical ionization mass spectrometry and its application to different wines and alcoholic beverages. J. Chromatogr. A. 2015;1381:210–218. doi: 10.1016/j.chroma.2014.12.074. [DOI] [PubMed] [Google Scholar]
- 154.Hjelmeland A.K., Ebeler S.E. Glycosidically Bound Volatile Aroma Compounds in Grapes and Wine: A Review. Am. J. Enol. Vitic. 2015;66:1–11. doi: 10.5344/ajev.2014.14104. [DOI] [Google Scholar]
- 155.Black C.A., Parker M., Siebert T.E., Capone D.L., Francis I.L. Terpenoids and their role in wine flavour: Recent advances. Aust. J. Grape Wine Res. 2015;21:582–600. doi: 10.1111/ajgw.12186. [DOI] [Google Scholar]
- 156.Liu J., Zhu X.-L., Ullah N., Tao Y.-S. Aroma Glycosides in Grapes and Wine. J. Food Sci. 2017;82:248–259. doi: 10.1111/1750-3841.13598. [DOI] [PubMed] [Google Scholar]
- 157.Bowles D., Isayenkova J., Lim E.-K., Poppenberger B. Glycosyltransferases: Managers of small molecules. Curr. Opin. Plant Biol. 2005;8:254–263. doi: 10.1016/j.pbi.2005.03.007. [DOI] [PubMed] [Google Scholar]
- 158.Jones P., Vogt T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta. 2001;213:164–174. doi: 10.1007/s004250000492. [DOI] [PubMed] [Google Scholar]
- 159.Song C.K., Hartl K., McGraphery K., Hoffmann T., Schwab W. Attractive but Toxic: Emerging Roles of Glycosidically Bound Volatiles and Glycosyltransferases Involved in Their Formation. Mol. Plant. 2018;11:1225–1236. doi: 10.1016/j.molp.2018.09.001. [DOI] [PubMed] [Google Scholar]
- 160.Bonisch F., Frotscher J., Stanitzek S., Ruhl E., Wust M., Bitz O., Schwab W. A UDP-Glucose: Monoterpenol Glucosyltransferase Adds to the Chemical Diversity of the Grapevine Metabolome. Plant Physiol. 2014;165:561–581. doi: 10.1104/pp.113.232470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Hjelmeland A.K., Zweigenbaum J., Ebeler S.E. Profiling monoterpenol glycoconjugation in Vitis vinifera L. cv. Muscat of Alexandria using a novel putative compound database approach, high resolution mass spectrometry and collision induced dissociation fragmentation analysis. Anal. Chim. Acta. 2015;887:138–147. doi: 10.1016/j.aca.2015.06.026. [DOI] [PubMed] [Google Scholar]
- 162.Godshaw J., Hjelmeland A.K., Zweigenbaum J., Ebeler S.E. Changes in glycosylation patterns of monoterpenes during grape berry maturation in six cultivars of Vitis vinifera. Food Chem. 2019;297 doi: 10.1016/j.foodchem.2019.05.195. [DOI] [PubMed] [Google Scholar]
- 163.Gunata Y.Z., Bayonove C.L., Baumes R.L., Cordonnier R.E. The Aroma of Grapes—Localization and Evolution of Free and Bound Fractions of Some Grape Aroma Components Cv Muscat During 1st Development And Maturation. J. Sci. Food Agric. 1985;36:857–862. doi: 10.1002/jsfa.2740360915. [DOI] [Google Scholar]
- 164.Razungles A., Gunata Z., Pinatel S., Baumes R., Bayonove C. Quantitative studies on terpenes, norisoprenoides and their precursors in several varieties of grapes. Sci. Aliments. 1993;13:59–72. [Google Scholar]
- 165.Maicas S., Mateo J.J. Hydrolysis of terpenyl glycosides in grape juice and other fruit juices: A review. Appl. Microbiol. Biotechnol. 2005;67:322–335. doi: 10.1007/s00253-004-1806-0. [DOI] [PubMed] [Google Scholar]
- 166.Genisheva Z., Oliveira J.M. Monoterpenic Characterization of White Cultivars from Vinhos Verdes Appellation of Origin (North Portugal) J. Inst. Brew. 2009;115:308–317. doi: 10.1002/j.2050-0416.2009.tb00386.x. [DOI] [Google Scholar]
- 167.Lamorte S.A., Gambuti A., Genovese A., Selicato S., Moio L. Free and glycoconjugated volatiles of V. vinifera grape ‘Falanghina’. Vitis. 2008;47:241–243. [Google Scholar]
- 168.Winterhalter P. 1,1,6-Trimethyl-1,2-Dihydronaphthalene (Tdn) Formation in Wine. 1. Studies on the Hydrolysis of 2,6,10,10-Tetramethyl-1-Oxaspiro [4.5]Dec-6-Ene-2,8-Diol Rationalizing the Origin of Tdn and Related C-13 Norisoprenoids in Riesling Wine. J. Agric. Food Chem. 1991;39:1825–1829. doi: 10.1021/jf00010a027. [DOI] [Google Scholar]
- 169.Salinas M.R., De La Hoz K.S., Zalacain A., Lara J.F., Garde-Cerdán T. Analysis of red grape glycosidic aroma precursors by glycosyl glucose quantification. Talanta. 2012;89:396–400. doi: 10.1016/j.talanta.2011.12.050. [DOI] [PubMed] [Google Scholar]
- 170.Yuan F., Qian M.C. Aroma Potential in Early- and Late-Maturity Pinot noir Grapes Evaluated by Aroma Extract Dilution Analysis. J. Agric. Food Chem. 2016;64:443–450. doi: 10.1021/acs.jafc.5b04774. [DOI] [PubMed] [Google Scholar]
- 171.Chatonnet P., Dubourdieu D., Boidron J.N., Lavigne V. Synthesis of Volatile Phenols by Saccharomyces-Cerevisiae in Wines. J. Sci. Food Agric. 1993;62:191–202. doi: 10.1002/jsfa.2740620213. [DOI] [Google Scholar]
- 172.Vanbeneden N., Saison D., Delvaux F., Delvaux F.R. Decrease of 4-Vinylguaiacol during Beer Aging and Formation of Apocynol and Vanillin in Beer. J. Agric. Food Chem. 2008;56:11983–11988. doi: 10.1021/jf8019453. [DOI] [PubMed] [Google Scholar]
- 173.Cho J.Y., Kim S.J., Lee H.J., Moon J.H. Two novel glycosyl cinnamic and benzoic acids from Korean black raspberry (Rubus coreanus) wine. Food Sci. Biotechnol. 2014;23:1081–1085. doi: 10.1007/s10068-014-0148-7. [DOI] [Google Scholar]
- 174.Sasaki K., Takase H., Tanzawa F., Kobayashi H., Saito H., Matsuo H., Takata R. Identification of Furaneol Glucopyranoside, the Precursor of Strawberry-like Aroma, Furaneol, in Muscat Bailey A. Am. J. Enol. Vitic. 2015;66:91–94. doi: 10.5344/ajev.2014.14072. [DOI] [Google Scholar]
- 175.Sasaki K., Takase H., Kobayashi H., Matsuo H., Takata R. Molecular cloning and characterization of UDP-glucose: Furaneol glucosyltransferase gene from grapevine cultivar Muscat Bailey A (Vitis labrusca × V. vinifera) J. Exp. Bot. 2015;66:6167–6174. doi: 10.1093/jxb/erv335. [DOI] [PubMed] [Google Scholar]
- 176.Strauss C.R., Wilson B., Williams P.J. Novel Monoterpene Diols and Diol Glycosides in Vitis-Vinifera Grapes. J. Agric. Food Chem. 1988;36:569–573. doi: 10.1021/jf00081a041. [DOI] [Google Scholar]
- 177.Strauss C.R., Dimitriadis E., Wilson B., Williams P.J. Studies on the Hydrolysis of 2 Megastigma-3,6,9-Triols Rationalizing the Origins of Some Volatile C-13 Norisoprenoids of Vitis-Vinifera Grapes. J. Agric. Food Chem. 1986;34:145–149. doi: 10.1021/jf00067a039. [DOI] [Google Scholar]
- 178.Sefton M.A., Skouroumounis G.K., Massywestropp R.A., Williams P.J. Norisoprenoids in Vitis-Vinifera White Wine Grapes and the Identification of A Precursor of Damascenone in These Fruits. Aust. J. Chem. 1989;42:2071–2084. doi: 10.1071/CH9892071. [DOI] [Google Scholar]
- 179.Puglisi C.J., Elsey G.M., Prager R.H., Skouroumounis G.K., Sefton M.A. Identification of a precursor to naturally occurring beta-damascenone. Tetrahedron Lett. 2001;42:6937–6939. doi: 10.1016/S0040-4039(01)01411-3. [DOI] [Google Scholar]
- 180.Puglisi C.J., Daniel M.A., Capone D.L., Elsey G.M., Prager R.H., Sefton M.A. Precursors to damascenone: Synthesis and hydrolysis of isomeric 3,9-dihydroxymegastigma-4,6,7-trienes. J. Agric. Food Chem. 2005;53:4895–4900. doi: 10.1021/jf050327p. [DOI] [PubMed] [Google Scholar]
- 181.Daniel M.A., Puglisi C.J., Capone D.L., Elsey G.M., Sefton M.A. Rationalizing the formation of damascenone: Synthesis and hydrolysis of damascenone precursors and their analogues, in both aglycone and glycoconjugate forms. J. Agric. Food Chem. 2008;56:9183–9189. doi: 10.1021/jf8018134. [DOI] [PubMed] [Google Scholar]
- 182.Lloyd N.D.R., Capone D.L., Ugliano M., Taylor D.K., Skouroumounis G.K., Sefton M.A., Elsey G.M. Formation of Damascenone under both Commercial and Model Fermentation Conditions. J. Agric. Food Chem. 2011;59:1338–1343. doi: 10.1021/jf103741n. [DOI] [PubMed] [Google Scholar]
- 183.Hayasaka Y., Wilkinson K.L., Elsey G.A., Raunkjaer M., Sefton M.A. Identification of natural oak lactone precursors in extracts of American and french oak woods by liquid chromatography-tandem mass Spectrometry. J. Agric. Food Chem. 2007;55:9195–9201. doi: 10.1021/jf072171u. [DOI] [PubMed] [Google Scholar]
- 184.Wilkinson K.L., Prida A., Hayasaka Y. Role of Glycoconjugates of 3-Methyl-4-hydroxyoctanoic Acid in the Evolution of Oak Lactone in Wine during Oak Maturation. J. Agric. Food Chem. 2013;61:4411–4416. doi: 10.1021/jf400175h. [DOI] [PubMed] [Google Scholar]
- 185.Gracia-Moreno E., Lopez R., Ferreira V. Quantitative determination of five hydroxy acids, precursors of relevant wine aroma compounds in wine and other alcoholic beverages. Anal. Bioanal. Chem. 2015;407:7925–7934. doi: 10.1007/s00216-015-8959-9. [DOI] [PubMed] [Google Scholar]
- 186.Pena-Gallego A., Hernandez-Orte P., Cacho J., Ferreira V. S-Cysteinylated and S-glutathionylated thiol precursors in grapes. A review. Food Chem. 2012;131:1–13. doi: 10.1016/j.foodchem.2011.07.079. [DOI] [Google Scholar]
- 187.Roland A., Schneider R., Razungles A., Cavelier F. Varietal Thiols in Wine: Discovery, Analysis and Applications. Chem. Rev. 2011;111:7355–7376. doi: 10.1021/cr100205b. [DOI] [PubMed] [Google Scholar]
- 188.Tominaga T., Murat M.L., Dubourdieu D. Development of a method for analyzing the volatile thiols involved in the characteristic aroma of wines made from Vitis vinifera L. cv. Sauvignon Blanc. J. Agric. Food Chem. 1998;46:1044–1048. doi: 10.1021/jf970782o. [DOI] [Google Scholar]
- 189.Mateo-Vivaracho L., Zapata J., Cacho J., Ferreira V. Analysis, Occurrence, and Potential Sensory Significance of Five Polyfunctional Mercaptans in White Wines. J. Agric. Food Chem. 2010;58:10184–10194. doi: 10.1021/jf101095a. [DOI] [PubMed] [Google Scholar]
- 190.Cerreti M., Esti M., Benucci I., Liburdi K., de Simone C., Ferranti P. Evolution of S-cysteinylated and S-glutathionylated thiol precursors during grape ripening of Vitis vinifera L. cvs Grechetto, Malvasia del Lazio and Sauvignon Blanc. Aust. J. Grape Wine Res. 2015;21:411–416. doi: 10.1111/ajgw.12152. [DOI] [Google Scholar]
- 191.Thibon C., Boecker C., Shinkaruk S., Moine V., Darriet P., Dubourdieu D. Identification of S-3-(hexanal)-glutathione and its bisulfite adduct in grape juice from Vitis vinifera L. cv. Sauvignon blanc as new potential precursors of 3SH. Food Chem. 2016;199:711–719. doi: 10.1016/j.foodchem.2015.12.069. [DOI] [PubMed] [Google Scholar]
- 192.Tominaga T., Peyrot des Gachons C., Dubourdieu D. A new type of flavor precursors in Vitis vinifera L. cv. Sauvignon Blanc: S-cysteine conjugates. J. Agric. Food Chem. 1998;46:5215–5219. doi: 10.1021/jf980481u. [DOI] [PubMed] [Google Scholar]
- 193.Fedrizzi B., Pardon K.H., Sefton M.A., Elsey G.M., Jeffery D.W. First Identification of 4-S-Glutathionyl-4-methylpentan-2-one, a Potential Precursor of 4-Mercapto-4-methylpentan-2-one, in Sauvignon Blanc Juice. J. Agric. Food Chem. 2009;57:991–995. doi: 10.1021/jf802799w. [DOI] [PubMed] [Google Scholar]
- 194.Subileau M., Schneider R., Salmon J.-M., Degryse E. New insights on 3-mercaptohexanol (3MH) biogenesis in sauvignon Blanc wines: Cys-3MH and (E)-Hexen-2-al are not the major precursors. J. Agric. Food Chem. 2008;56:9230–9235. doi: 10.1021/jf801626f. [DOI] [PubMed] [Google Scholar]
- 195.Grant-Preece P.A., Pardon K.H., Capone D.L., Cordente A.G., Sefton M.A., Jeffery D.W., Elsey G.M. Synthesis of Wine Thiol Conjugates and Labeled Analogues: Fermentation of the Glutathione Conjugate of 3-Mercaptohexan-1-ol Yields the Corresponding Cysteine Conjugate and Free Thiol. J. Agric. Food Chem. 2010;58:1383–1389. doi: 10.1021/jf9037198. [DOI] [PubMed] [Google Scholar]
- 196.Bonnaffoux H., Roland A., Rémond E., Delpech S., Schneider R., Cavelier F. First identification and quantification of S-3-(hexan-1-ol)-γ-glutamyl-cysteine in grape must as a potential thiol precursor, using UPLC-MS/MS analysis and stable isotope dilution assay. Food Chem. 2017;237:877–886. doi: 10.1016/j.foodchem.2017.05.116. [DOI] [PubMed] [Google Scholar]
- 197.Concejero B., Pena-Gallego A., Fernandez-Zurbano P., Hernandez-Orte P., Ferreira V. Direct accurate analysis of cysteinylated and glutathionylated precursors of 4-mercapto-4-methyl-2-pentanone and 3-mercaptohexan-1-ol in must by ultrahigh performance liquid chromatography coupled to mass spectrometry. Anal. Chim. Acta. 2014;812:250–257. doi: 10.1016/j.aca.2014.01.004. [DOI] [PubMed] [Google Scholar]
- 198.Roland A., Schneider R., Charrier F., Cavelier F., Rossignol M., Razungles A. Distribution of varietal thiol precursors in the skin and the pulp of Melon B. and Sauvignon Blanc grapes. Food Chem. 2011;125:139–144. doi: 10.1016/j.foodchem.2010.08.050. [DOI] [Google Scholar]
- 199.Kobayashi H., Matsuyama S., Takase H., Sasaki K., Suzuki S., Takata R., Saito H. Impact of Harvest Timing on the Concentration of 3-Mercaptohexan-1-ol Precursors in Vitis vinifera Berries. Am. J. Enol. Vitic. 2012;63:544–548. doi: 10.5344/ajev.2012.12051. [DOI] [Google Scholar]
- 200.Wang L., Harada J., Endo Y., Hisamoto M., Saito F., Okuda T. Diurnal Changes in Amino Acid Concentrations in Riesling and Chardonnay Grape Juices and a Possible Role of Sunlight. Am. J. Enol. Vitic. 2014;65:435–442. doi: 10.5344/ajev.2014.13144. [DOI] [Google Scholar]
- 201.Olejar K.J., Fedrizzi B., Kilmartin P.A. Influence of harvesting technique and maceration process on aroma and phenolic attributes of Sauvignon blanc wine. Food Chem. 2015;183:181–189. doi: 10.1016/j.foodchem.2015.03.040. [DOI] [PubMed] [Google Scholar]
- 202.Allen T., Herbst-Johnstone M., Girault M., Butler P., Logan G., Jouanneau S., Nicolau L., Kilmartin P.A. Influence of Grape-Harvesting Steps on Varietal Thiol Aromas in Sauvignon blanc Wines. J. Agric. Food Chem. 2011;59:10641–10650. doi: 10.1021/jf2018676. [DOI] [PubMed] [Google Scholar]
- 203.Maggu M., Winz R., Kilmartin P.A., Trought M.C.T., Nicolau L. Effect of skin contact and pressure on the composition of Sauvignon Blanc must. J. Agric. Food Chem. 2007;55:10281–10288. doi: 10.1021/jf072192o. [DOI] [PubMed] [Google Scholar]
- 204.Capone D.L., Black C.A., Jeffery D.W. Effects on 3-Mercaptohexan-1-ol Precursor Concentrations from Prolonged Storage of Sauvignon Blanc Grapes Prior to Crushing and Pressing. J. Agric. Food Chem. 2012;60:3515–3523. doi: 10.1021/jf300054h. [DOI] [PubMed] [Google Scholar]
- 205.Larcher R., Nicolini G., Tonidandel L., Villegas T.R., Malacarne M., Fedrizzi B. Influence of oxygen availability during skin-contact maceration on the formation of precursors of 3-mercaptohexan-1-ol in Muller-Thurgau and Sauvignon Blanc grapes. Aust. J. Grape Wine Res. 2013;19:342–348. doi: 10.1111/ajgw.12039. [DOI] [Google Scholar]
- 206.Capone D.L., Sefton M.A., Jeffery D.W. Application of a Modified Method for 3-Mercaptohexan-1-ol Determination To Investigate the Relationship between Free Thiol and Related Conjugates in Grape Juice and Wine. J. Agric. Food Chem. 2011;59:4649–4658. doi: 10.1021/jf200116q. [DOI] [PubMed] [Google Scholar]
- 207.Darriet P., Tominaga T., Demole E., Dubourdieu D. Evidence of the Presence of a 4-Mercapto-4-Methylpentan-2-One Precursor in Vitis-Vinifera Sauvignon Blanc Grape Variety. C. R. Acad. Sci. III-Vie. 1993;316:1332–1335. [Google Scholar]
- 208.Segurel M.A., Razungles A.J., Riou C., Salles M., Baumes R.L. Contribution of dimethyl sulfide to the aroma of Syrah and Grenache Noir wines and estimation of its potential in grapes of these varieties. J. Agric. Food Chem. 2004;52:7084–7093. doi: 10.1021/jf049160a. [DOI] [PubMed] [Google Scholar]
- 209.Lytra G., Tempere S., Zhang S., Marchand S., de Revel G., Barbe J.-C. Olfactory Impact of Dimethyl Sulfide on Red Wine Fruity Esters Aroma Expression in Model Solution. OENO One. 2014;48:75–85. doi: 10.20870/oeno-one.2014.48.1.1660. [DOI] [Google Scholar]
- 210.Segurel M.A., Razungles A.J., Riou C., Trigueiro M.G.L., Baumes R.L. Ability of possible DMS precursors to release DMS during wine aging and in the conditions of heat-alkaline treatment. J. Agric. Food Chem. 2005;53:2637–2645. doi: 10.1021/jf048273r. [DOI] [PubMed] [Google Scholar]
- 211.Loscos N., Segurel M., Dagan L., Sommerer N., Marlin T., Baumes R. Identification of S-methylmethionine in Petit Manseng grapes as dimethyl sulphide precursor in wine. Anal. Chim. Acta. 2008;621:24–29. doi: 10.1016/j.aca.2007.11.033. [DOI] [PubMed] [Google Scholar]
- 212.Dupre N.D.R., Schneider R., Payan J.C., Salancon E., Razungles A. Effects of Vine Water Status on Dimethyl Sulfur Potential, Ammonium, and Amino Acid Contents in Grenache Noir Grapes (Vitis vinifera) J. Agric. Food Chem. 2014;62:2760–2766. doi: 10.1021/jf404758g. [DOI] [PubMed] [Google Scholar]
- 213.Thibon C., Dubourdieu D., Darriet P., Tominaga T. Impact of noble rot on the aroma precursor of 3-sulfanylhexanol content in Vitis vinifera L. cv Sauvignon blanc and Semillon grape juice. Food Chem. 2009;114:1359–1364. doi: 10.1016/j.foodchem.2008.11.016. [DOI] [Google Scholar]
- 214.Thibon C., Shinkaruk S., Jourdes M., Bennetau B., Dubourdieu D., Tominaga T. Aromatic potential of botrytized white wine grapes: Identification and quantification of new cysteine-S-conjugate flavor precursors. Anal. Chim. Acta. 2010;660:190–196. doi: 10.1016/j.aca.2009.10.018. [DOI] [PubMed] [Google Scholar]
- 215.Sadoughi N., Schmidtke L.M., Antalick G., Blackman J.W., Steel C.C. Gas Chromatography-Mass Spectrometry Method Optimized Using Response Surface Modeling for the Quantitation of Fungal Off-Flavors in Grapes and Wine. J. Agric. Food Chem. 2015;63:2877–2885. doi: 10.1021/jf505444r. [DOI] [PubMed] [Google Scholar]
- 216.Morales-Valle H., Silva L.C., Paterson R.R.M., Venancio A., Lima N. Effects of the origins of Botrytis cinerea on earthy aromas from grape broth media further inoculated with Penicillium expansum. Food Microbiol. 2011;28:1048–1053. doi: 10.1016/j.fm.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 217.Krstic M.P., Johnson D.L., Herderich M.J. Review of smoke taint in wine: Smoke-derived volatile phenols and their glycosidic metabolites in grapes and vines as biomarkers for smoke exposure and their role in the sensory perception of smoke taint. Aust. J. Grape Wine Res. 2015;21:537–553. doi: 10.1111/ajgw.12183. [DOI] [Google Scholar]
- 218.Kennison K.R., Wilkinson K.L., Williams H.G., Smith J.H., Gibberd M.R. Smoke-derived taint in wine: Effect of postharvest smoke exposure of grapes on the chemical composition and sensory characteristics of wine. J. Agric. Food Chem. 2007;55:10897–10901. doi: 10.1021/jf072509k. [DOI] [PubMed] [Google Scholar]
- 219.Kennison K.R., Gibberd M.R., Pollnitz A.P., Wilkinson K.L. Smoke-derived taint in wine: The release of smoke-derived volatile phenols during fermentation of Merlot juice following grapevine exposure to smoke. J. Agric. Food Chem. 2008;56:7379–7383. doi: 10.1021/jf800927e. [DOI] [PubMed] [Google Scholar]
- 220.Hayasaka Y., Dungey K.A., Baldock G.A., Kennison K.R., Wilkinson K.L. Identification of a beta-d-glucopyranoside precursor to guaiacol in grape juice following grapevine exposure to smoke. Anal. Chim. Acta. 2010;660:143–148. doi: 10.1016/j.aca.2009.10.039. [DOI] [PubMed] [Google Scholar]
- 221.Hayasaka Y., Baldock G.A., Parker M., Pardon K.H., Black C.A., Herderich M.J., Jeffery D.W. Glycosylation of smoke-derived volatile phenols in grapes as a consequence of grapevine exposure to bushfire smoke. J. Agric. Food Chem. 2010;58:10989–10998. doi: 10.1021/jf103045t. [DOI] [PubMed] [Google Scholar]
- 222.Dungey K.A., Hayasaka Y., Wilkinson K.L. Quantitative analysis of glycoconjugate precursors of guaiacol in smoke-affected grapes using liquid chromatography-tandem mass spectrometry based stable isotope dilution analysis. Food Chem. 2011;126:801–806. doi: 10.1016/j.foodchem.2010.11.094. [DOI] [Google Scholar]
- 223.Ristic R., van der Hulst L., Capone D.L., Wilkinson K.L. Impact of Bottle Aging on Smoke-Tainted Wines from Different Grape Cultivars. J. Agric. Food Chem. 2017;65:4146–4152. doi: 10.1021/acs.jafc.7b01233. [DOI] [PubMed] [Google Scholar]