

Abstract
Fine balance between loss-of water and gain-of water is essential for maintaining body fluid homeostasis. The development of neural manipulation and mapping tools has opened up new avenues to dissect the neural circuits underlying body fluid regulation. Recent studies have identified several nodes in the brain that positively and negatively regulate thirst. The next step forward would be to elucidate how neural populations interact with each other to control drinking behavior.
Introduction
Thirst is an instinctive drive that prompts animals toward intense water seeking and consumption to restore body fluid balance. Precise tuning of fluid balance is essential for survival. Classical and contemporary studies across species have unveiled the basic principles of thirst regulation.
In this review, we will briefly overview fluid regulatory mechanisms conserved in many species, and summarize gain-of-function and loss-of-function studies of neural populations and circuitry. Second, we will describe our current understanding of the mechanisms involved in thirst quenching and satiety.
Driving of thirst
Thirst driving system across species and gain of function studies
In many species, Angiotensin II (Ang II) along with systemic osmolality plays important roles in driving thirst and drinking behavior. The dipsogenic effect of Ang II was first demonstrated in rodents: intracranial injection of Ang II into the third ventricle and other brain areas immediately induced water drinking behavior [1]. Besides rodents, rhesus monkeys [2–4], goats [5,6], cows [7], sheep [8], pigs [9,10], dogs [11,12], and cats [13–15] have been found to drink water in response to intracranial administration of Ang II. These studies were followed up by electrical stimulation of the lamina terminalis (LT), a major site of Ang II action that increases fluid intake in several mammalian species (Table 1). Ang II also causes water drinking behavior in reptiles, amphibians and birds [1,16]. Interestingly, studies in aquatic amphibious fishes revealed a unique system of Ang II-related thirst regulation. In mudskipper, Ang II stimulates the area postrema (AP), a hindbrain nucleus, to induce swallowing action [17,18]. Recent studies with contemporary neural manipulation tools have expanded our knowledge of thirst circuits at a finer resolution. In particular, optogenetic manipulation using light-sensitive channels [19], and Designer Receptors Exclusively Activated by Designer Drugs (DREADD)-based chemogenetic manipulation [20] allowed to link the activity of a specific neural population with behavioral outcome. The LT is composed of three structures; the subfornical organ (SFO), the organum vasculosum of the lamina terminalis (OVLT) and the median preoptic nucleus (MnPO). Of these, SFO and OVLT lack the normal blood–brain barrier, and have direct access to circulation. Recently, several genetically defined neural populations related to thirst have been identified in the LT [21,22] (Table 1). Stimulation of a glutamatergic population of SFO neurons marked by the expression of the transcription factor ETV-1 [23••], nitric oxide synthase 1 (nNOS) [23••,24••,25] and Ca2+/calmodulin-dependent kinase II (CamKII) [23••,26], evoked voracious drinking of water. The MnPO also has excitatory populations expressing nNOS or adenylate cyclase activating polypeptide 1 (Adcyap1) that positively regulate drinking behavior [27••,28]. Similarly, in the OVLT, nNOS – or angiotensin 1A receptor (Agtr1a) – expressing neurons were identified as thirst-promoting neurons [27••,28].
Table 1.
Gain of function studies related with the LT
| Brain part or cell types | Method of manipulation | Phenotype | Species | Ref. |
|---|---|---|---|---|
| SFO | Ang II injection | Water intake ↑ | Rat | [68,69] |
| Japanese quail | [70] | |||
| Electrical stimulation | Water intake ↑ | Rat | [71,72] | |
| AVP secretion ↑ | Rat | [73] | ||
| POA | Ang II injection | Water intake ↑ | Cat | [14] |
| Water intake ↑; AVP secretion ↑ | Rhesus monkey | [3,4] | ||
| Water intake ↑ | Japanese quail | [74] | ||
| SFO, POA | Ang II injection | Water intake ↑ | Dog | [11] |
| MnPO | Ang II injection | Water intake ↑ | Rat | [75] |
| SFOnNOS, sFOcamkII, SFOETV1 | ChR2 | Time-locked water intake upon photostimulation | Mouse | [23••,24••,25,27••] |
| SFOnNOS, SFOcamkII | Gq-DREADDs | Water intake ↑ | Mouse | [25,26,27••] |
| MnPO/OVLTGLUT | ChR2 | Water intake ↑ | Mouse | [50•] |
| MnPOnNOS | ChR2 | Time-locked water intake upon photostimulation | Mouse | [27••] |
| MnPOAdcyap1 | ChR2 | Water but not NaCl intake | Mouse | [28] |
| OVLTnNOS | ChR2 | Time-locked water intake upon photostimulation | Mouse | [27••] |
| OVLTAgtr1a | ChR2 | Water but not NaCl intake | Mouse | [28] |
| SFOGLUT neurons projecting to the MnPO | ChR2 | Time-locked water intake upon photostimulation | Mouse | [24••] |
| SFOGLUT neurons projecting to the OVLT | ChR2 | Time-locked water intake upon photostimulation | Mouse | [33] |
| SCNavp neurons projecting to the OVLT | ChR2 | Time-locked water intake upon photostimulation | Mouse | [29•] |
| SFOGABA | ChR2 | Water intake ↓ (dehydration) | Mouse | [23••] |
| MnPO/OVLTGABA | ChR2 | Water intake ↓ (dehydration) | Mouse | [50•] |
| MnPOGLP1R | ChR2 | Water intake ↓ (dehydration) | Mouse | [27••] |
Adcyap1, adenylate cyclase activating polypeptide 1; Agtr1a, angiotensin 1A receptor; Ang II, Angiotensin II; AVP, arginine vasopressin; AVP secretion ↑; induction of AVP secretion; Camk2, calcium/calmodulin-dependent protein kinase type 2; ChR2, channelrhodopsin2; DREADDs, designer receptors exclusively activated by designer drugs; Etv1, ETS translocation variant 1; GABA, GABAergic; GLUT, glutamatergic; MnPO, median preoptic nucleus; nNOS, neuronal nitric oxide synthase; OVLT, organum vasculosum of the lamina terminalis; POA, preoptic area; SCN, suprachiasmatic nucleus; SFO, subfornical organ; Water intake ↑; induction of water intake; Water intake ↓, reduction of water intake; (dehydration), under water-restricted condition.
Outside the LT, vasopressin-expressing neurons in the suprachiasmatic nucleus (SCN) that project to thirst neurons in the OVLT have been shown to mediate anticipatory thirst before sleep [29•]. The lateral hypothalamic area (LHA) is also known for regulating ingestive behavior [30]. A recent study reported that activation of neurotensin (Nts)-expressing LHA neurons promotes fluid but not food intake [31]. Because LHA is one of the major downstream target of thirst neurons in the MnPO [27••,28,32•], it is possible that Nts neurons may receive direct inputs from the LT to relay thirst information to the next brain station.
We note that the LT is also implicated in the regulation of sodium ingestion. A study showed that SFO neurons expressing Agtr1a mediate sodium intake [33]. It is intriguing that SFOAgtr1a neurons seem to be a subset of SFOnNOS neurons, but their activation drives sodium intake instead of water. Future studies are required to reconcile how appetites for water and sodium are encoded by partially overlapping SFO excitatory neurons.
Loss-of-function studies of the LT
In parallel to gain-of-function studies, several loss-of-function techniques have been employed in the past few decades to study the functional necessity of a given neural circuit (Table 2). Early studies demonstrated that radio-frequency lesions of forebrain areas including the LT disrupted fluid balance and thirst mechanisms in goats [34] and rats [35,36]. The physical destruction of the neural connection between the SFO and MnPO has been shown to attenuate water intake in rats [37,38]. Consistently, in rats with lesions of the SFO and OVLT, there was a decrease in the number of Fos-like immuno-reactive neurons in the MnPO, following intravenous infusion of hypertonic saline solution [39]. Moreover, ablation of the MnPO by multiple techniques such as electrolytic ablation [40] or ibotenic acid [41] all blocked drinking behavior. A comprehensive study in sheep also supports this view: radiofrequency ablation of MnPO reduced drinking behavior, but ablation of the OVLT or SFO alone had minor effects on drinking in response to hypertonic saline [42]. Taken together, ablation of individual nuclei of the LT (with the most profound effects by MnPO ablation) has an impact on angiotensin-induced or hypertonicity-induced drinking.
Table 2.
Loss of function studies related with the LT
| Brain part or cell types | Method of lesions or inhibition | Phenotype | AVP secretion | Species | Ref. |
|---|---|---|---|---|---|
| AV3V | Radio-frequency lesions | Adipsia | ↓ | Goat | [34] |
| ↓ [76] | Rat | [35,36] | |||
| Fibers between the SFO and MnPO | Physical cut | Water intake ↓ (Ang II) Normal ad-lib drinking |
– | Rat | [37,38] |
| SFO | Electrolytic lesions | Water intake ↓ (Ang II) | ↓ [77] | Rat | [44] |
| Recovered after 2 weeks | |||||
| Water intake ↓ (Ang II) | – | Dog | [80] | ||
| Radio-frequency lesions | No effect (high NaCl) | ↓ [78] | Sheep | [42,45] | |
| MnPO | Electrolytic lesions | Water intake ↓ (Ang II and high NaCl) | ↓ [79] | Rat | [81] |
| Ibotenate lesions | Water intake ↓ (Ang II and high NaCl) | – | Rat | [41] | |
| Radio-frequency lesions | Water intake ↓ (high NaCl) | ↓ [78] | Sheep | [42,45] | |
| OVLT | Electrolytic lesions | Water intake ↓ (high NaCl) | ↓ | Dog | [82] |
| No effect (high NaCl) | – | Rat | [83] | ||
| Radio-frequency lesions | No effect (high NaCl) | ↓ [78] | Sheep | [42,45] | |
| SFO and MnPO | Radio-frequency lesions | Water intake ↓ (high NaCl) | – | Sheep | [42,45] |
| SFO and OVLT | Radio-frequency lesions | Water intake ↓ (high NaCl) | ↓ [78] | Sheep | [42,45] |
| SFO and AV3V | Electrolytic lesions | C-fos in MnPO ↓ (high NaCl) | – | Rat | [39] |
| SFO,MnPO and OVLT | Radio-frequency lesions | Abolished water intake (high NaCl) | ↓ [78] | Sheep | [42,45,84] |
| MnPO and OVLT | Radio-frequency lesions | Abolished water intake (high NaCl) | – | Sheep | [42,45] |
| SFOnNOS | Arch | Abolished water intake (dehydration) | – | Mouse | [24••] |
| SFO neurons projecting to the OVLT | Arch | Water intake ↓ (dehydration) | – | Mouse | [33] |
| MnPOnNOS | Caspase ablation | Water intake ↓ (SFOnNOS or OVLTnNOS stimulation) | – | Mouse | [27••] |
| MnPOGLP1R | Caspase ablation | Liquid intake (saline) ↑ | – | Mouse | [27••] |
| MnPOnNOS | Gi-DREADDs | Abolished water intake (dehydration or SFOnNOS stimulation water) | – | Mouse | [27••] |
| MnPOGLP1R | Gi-DREADDs | Liquid intake (saline) ↑ | – | Mouse | [27••] |
| SCNAVP neurons projecting to the OVLT | Arch | Abolished water intake (before sleep) | – | Mouse | [29•] |
Arch, archaerhodopsin; AV3V, periventricular tissue surrounding the anteroventral third ventricle; AVP, arginine vasopressin; DREADDs, designer receptors exclusively activated by designer drugs; GLP1R, glucagon-like peptide 1 receptor); MnPO, median preoptic nucleus; nNOS, neuronal nitric oxide synthase; N.I., not indicated; OVLT, organum vasculosum of the lamina terminalis; SCN, suprachiasmatic nucleus; SFO, subfornical organ; Water intake ↓, around 50–95% of reduction of water intake; Abolished water intake, around 95–100% of reduction of water intake; (Ang II), response to Ang II administration; (high NaCl), response to hypertonic NaCl administration; (dehydration), under water-restricted condition.
Classical loss-of-function approaches generally affect all neurons in a given brain area, whereas recent manipulation tools can be used to silence/ablate a genetically defined neural population. Augustine et al. applied cell-type-specific ablation by caspase [43] to individual nuclei of the LT. The study revealed that MnPOnNOS neurons are essential for driving drinking behavior induced by photostimulation of SFOnNOS neurons [27••]. By contrast, stimulation of MnPOnNOS neurons after ablating SFOnNOS/OVLTnNOS still triggered drinking. Thus, the MnPOnNOS neuronal population is required to integrate the signals from thirst-driving neurons of the SFO [27••]. However, there are some discrepancies regarding the requirement of SFO neurons for drinking. Permanent ablation of the SFO [42,44,45] or SFOnNOS neurons (unpublished) had temporally/minimum effect on water intake. In contrast, acute optogenetic inhibition of SFOnNOS neurons [24••], or the SFO → OVLT projection [33] using archaerhodopsin significantly attenuated water intake. Although precise mechanisms are unknown, it is feasible that the function of SFOnNOS neurons may be required for drinking in short periods, but permanent ablation induces neural plasticity over time that compensates for the loss of SFO neurons to maintain body fluid balance.
Quenching of thirst
Functional studies of the LT
Thirsty animals including humans stop drinking water before the systemic environment recovers (rehydration). This early termination of drinking was known both behaviorally and endocrinologically. For instance, studies in dogs [46,47] and rhesus monkeys [48] with a gastric fistula showed that only sham-drinking rapidly inhibited vasopressin release from the brain. On the basis of these observations, it was proposed that there may be thirst-quenching neural circuits that respond to drinking. More recently, drinking water has been shown to inhibit the activity of thirst-related neurons [24••,27••,32••,49•]. These studies suggested that water intake stimulates thirst-quenching signals in the brain that leads to rapid drinking termination.
At the circuit level, we have just begun to get handles on thirst-quenching neurons mainly in the LT. It was demonstrated that the activation of GABAergic neurons in the SFO (SFOVGAT) suppresses water intake in thirsty animals [23••]. Similarly, optogenetic activation of inhibitory neurons in the MnPO/OVLT suppressed water intake [27••,50•]. Thus, it appears that stimulation of inhibitory populations in the LT quenches thirst in general.
Although LT inhibitory neurons are sufficient to inhibit drinking, few studies have addressed their physiological role related to thirst quenching. Our laboratory tackled this question by optical recording from individual neural populations in the LT. We showed that MnPO inhibitory neurons that coexpress glucagon-like peptide 1 receptor (MnPOGLP1R) are activated upon gulping action regardless of liquid type, and they send mono-synaptic inhibition to SFOnNOS neurons [27••]. These experiments implicated that MnPOGLP1R neurons receive signals from the oropharyngeal areas in response to drinking. Sensory stimuli from the oropharyngeal area are generally transported through cranial nerves V, VII, IX, and X to the central pattern generator within the nucleus tractus solitarius (NTS), which elicits swallowing action. Although fluid-sensing mechanisms in the peripheral area remain poorly understood, MnPOGLP1R neurons may receive oropharyngeal inputs from other brain regions such as the NTS, and provide rapid quenching of thirst circuits.
Functional studies of other brain regions
Several other brain regions are also implicated in thirst-quenching. The NTS receives peripheral inputs including visceral and baroreceptor signals [51–54], and is known to regulate various appetites [53,55,56]. For example, optogenetic stimulation of cholecystokinin (CCK) expressing NTS neurons decreases feeding [57]. Lesions of NTS neurons have been reported to cause overdrinking in rats [51,52] suggesting that the NTS plays a role in optimizing water intake. However, distinct neural populations that control water intake remain unknown. Another hindbrain structure related to thirst-quenching is the parabrachial nucleus (PBN). Stimulation of oxytocin receptor-expressing neurons in the PBN (OxtrPBN neurons) has been shown to suppress fluid intake, but not food or salt intake [58•]. The PBN is one of the major projection sites of NTS neurons [59] that forms reciprocal connections with forebrain areas [60–63]. It is possible that the PBN relays thirst-quenching signals from the NTS to forebrain regions such as the MnPO.
Although recent studies have pinpointed multiple thirst-quenching nodes, it is still unclear how activation of these neurons suppresses drinking. Do they reduce the valence of water, or the motivation to drink? Currently, understanding of emotional and conscious processing of thirst is severely limited. On the basis of anatomical connections in rodents [64] and functional studies in humans [65], the LT-thalamus-insular/cingulate cortex axis may be related to the genesis of thirst perception.
Conclusions and future directions
-
Efferent connections from the LT to other brain regions for driving thirst.
The brain regions including the LT, NTS, PBN, and insular/cingulate cortex have been all implicated in fluid regulation. However, how interoceptive information from the LT is processed in downstream neural circuits remains unknown. Future studies should focus on identifying the specific circuitry from the LT and functionally annotate individual neural populations to behavioral and hormonal outputs related to thirst.
-
How do peripheral organs sense drinking and send signals to thirst-quenching neurons?
Water intake stimulates multiple sensory signals including taste [66], oropharyngeal gulping motion, and osmolality changes in the gut (Figure 1) [67]. Interestingly, one thirst-quenching population (e.g. MnPOGLP1R neurons) represents a specific aspect of drinking behavior (in this case, liquid gulping). However, there are still important questions remain to be answered. Are there distinct types of thirst-quenching neurons that detect other sensory stimuli evoked by water intake? How does each neural population ‘know’ that peripheral drinking events have occurred? Recent development of in vivo optical recording, manipulation, and tracing tools will help identify molecular and neural basis of thirst-quenching and satiety.
Figure 1.
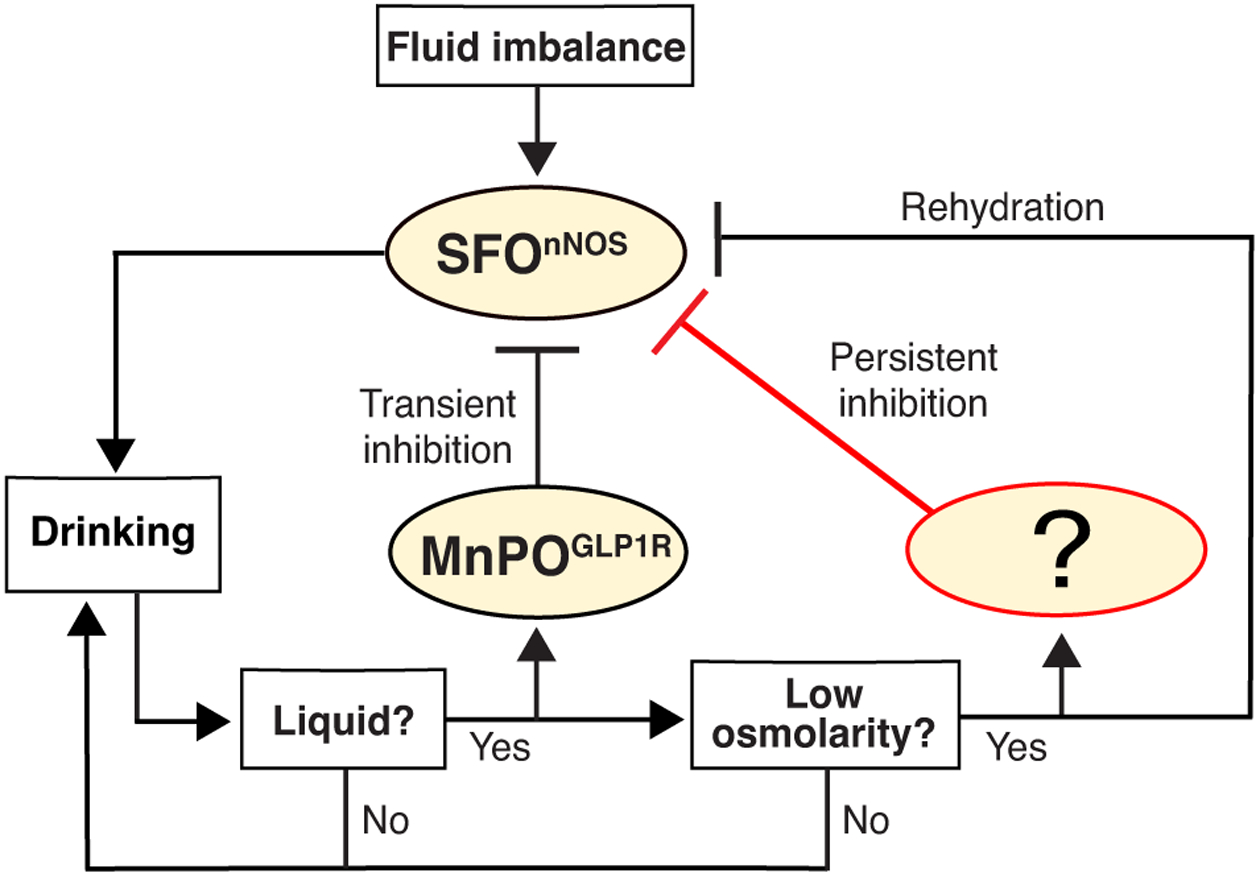
Flow chart of the inhibitory effects on the SFO thirst driving neurons. After ingestion, the MnPOGLP1r neurons quickly respond to drinking of any types of liquid, and provide transient inhibition to the SFOnNOS neurons. It remains unknown which neurons are specifically activated by hypo-osmolarity induced by water ingestion and send the persistent inhibitory signal to the SFO.
Acknowledgements
We thank the members of the Oka laboratory for helpful comments. Y.O. is supported by Startup funds from California Institute of Technology, the Searle Scholars Program, the Mallinckrodt Foundation, the McKnight Foundation and the Klingenstein-Simons Foundation, and NIH (U01NS099717, R56MH113030).
Footnotes
Conflict of interest statement
Nothing declared.
References and recommended reading
Papers of particular interest, published within the period of review, have been highlighted as:
• of special interest
•• of outstanding interest
- 1.Fitzsimons JT: Angiotensin, thirst, and sodium appetite. Physiol Rev 1998, 78:583–686. [DOI] [PubMed] [Google Scholar]
- 2.Setler P: Drinking induced by injection of angiotensin II into the hypothalamus of the rhesus monkey. J Physiol 1971, 217:59P–60P. [PubMed] [Google Scholar]
- 3.Sharpe LG, Swanson LW: Drinking induced by injections of angiotensin into forebrain and mid-brain sites of the monkey. J Physiol 1974, 239:595–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Simonnet G, Rodriguez F, Fumoux F, Czernichow P, Vincent JD: Vasopressin release and drinking induced by intracranial injection of angiotensin II in monkey. Am J Physiol 1979, 237: R20–R25. [DOI] [PubMed] [Google Scholar]
- 5.Andersson B, Westbye O: Synergistic action of sodium and angiotensin on brain mechanisms controlling water and salt balance. Nature 1970, 228:75. [DOI] [PubMed] [Google Scholar]
- 6.Thornton SN, Baldwin BA: Drinking in the goat in response to simultaneous peripheral and central infusions of angiotensin II. Physiol Behav 1985, 35:753–755. [DOI] [PubMed] [Google Scholar]
- 7.Blair-West JR, Denton DA, McKinley MJ, Weisinger RS: Angiotensin-related sodium appetite and thirst in cattle. Am J Physiol 1988, 255:R205–R211. [DOI] [PubMed] [Google Scholar]
- 8.Weisinger RS, Denton DA, Di Nicolantonio R, McKinley MJ, Muller AF, Tarjan E: Role of angiotensin in sodium appetite of sodium-deplete sheep. Am J Physiol 1987, 253:R482–R488. [DOI] [PubMed] [Google Scholar]
- 9.Baldwin BA, Thornton SN: Operant drinking in pigs following intracerebroventricular injections of hypertonic solutions and angiotensin II. Physiol Behav 1986, 36:325–328. [DOI] [PubMed] [Google Scholar]
- 10.Mutter J, Lemoine J, Tsang B, Kucharczyk J: Central angiotensin-induced water intake and salt appetite in the pig. Brain Res 1984, 322:374–377. [DOI] [PubMed] [Google Scholar]
- 11.Fitzsimons JT, Kucharczyk J: Drinking and haemodynamic changes induced in the dog by intracranial injection of components of the renin-angiotensin system. J Physiol 1978, 276:419–434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ramsay DJ, Reid IA: Some central mechanisms of thirst in the dog. J Physiol 1975, 253:517–525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sturgeon RD, Brophy PD, Levitt RA: Drinking elicited by intracranial microinjection of angiotensin in the cat. Pharmacol Biochem Behav 1973, 1:353–355. [DOI] [PubMed] [Google Scholar]
- 14.Brophy PD, Levitt RA: Dose-response analysis of angiotensin and renin-induced drinking in the cat. Pharmacol Biochem Behav 1974, 2:509–514. [DOI] [PubMed] [Google Scholar]
- 15.Cooling MJ, Day MD: Drinking behaviour in the cat induced by renin, angiotensin I, II and isoprenaline. J Physiol 1975, 244:325–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kobayashi H, Uemura H, Wada M, Takei Y: Ecological adaptation of angiotensin-induced thirst mechanism in tetrapods. Gen Comp Endocrinol 1979, 38:93–104. [DOI] [PubMed] [Google Scholar]
- 17.Katayama Y, Sakamoto T, Saito K, Tsuchimochi H, Kaiya H, Watanabe T, Pearson JT, Takei Y: Drinking by amphibious fish: convergent evolution of thirst mechanisms during vertebrate terrestrialization. Sci Rep 2018, 8:625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Katayama Y, Sakamoto T, Takanami K, Takei Y: The amphibious mudskipper: a unique model bridging the gap of central actions of osmoregulatory hormones between terrestrial and aquatic vertebrates. Front Physiol 2018, 9:1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K: Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci 2005, 8:1263–1268. [DOI] [PubMed] [Google Scholar]
- 20.Roth BL: DREADDs for neuroscientists. Neuron 2016, 89:683–694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zimmerman CA, Leib DE, Knight ZA: Neural circuits underlying thirst and fluid homeostasis. Nat Rev Neurosci 2017, 18:459–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Augustine V, Gokce SK, Oka Y: Peripheral and central nutrient sensing underlying appetite regulation. Trends Neurosci 2018, 41:526–539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Oka Y, Ye M, Zuker CS: Thirst driving and suppressing signals encoded by distinct neural populations in the brain. Nature 2015, 520:349–352. [DOI] [PMC free article] [PubMed] [Google Scholar]; •• First study to show that cell type-specific manipulations of the SFO. The authors demonstrate that optogenetic activation of ETV-1 expressing neurons in the SFO evokes drinking behavior, whereas activation of GABA expressing neurons suppresses drinking behavior.
- 24.Zimmerman CA, Lin YC, Leib DE, Guo L, Huey EL, Daly GE, Chen Y, Knight ZA: Thirst neurons anticipate the homeostatic consequences of eating and drinking. Nature 2016, 537:680–684. [DOI] [PMC free article] [PubMed] [Google Scholar]; •• This study shows a mechanism of anticipatory control of thirst satiation. Optical recordings of calcium dynamics in SFOnNOS neurons reveal that SFOnNOS neurons are rapidly inhibited during drinking and transiently inhibited by oral cooling, whereas activated during feeding.
- 25.Betley JN, Xu S, Cao ZFH, Gong R, Magnus CJ, Yu Y, Sternson SM: Neurons for hunger and thirst transmit a negative-valence teaching signal. Nature 2015, 521:180–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nation HL, Nicoleau M, Kinsman BJ, Browning KN, Stocker SD: DREADD-induced activation of subfornical organ neurons stimulates thirst and salt appetite. J Neurophysiol 2016, 115:3123–3129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Augustine V, Gokce SK, Lee S, Wang B, Davidson TJ, Reimann F, Gribble F, Deisseroth K, Lois C, Oka Y: Hierarchical neural architecture underlying thirst regulation. Nature 2018, 555:204–209. [DOI] [PMC free article] [PubMed] [Google Scholar]; •• This study shows a hierarchical circuit organization in the LT showing that the MnPOnNOS neurons are essential for the integration of the signals from thirst-driving neurons of the SFO. This study also showed that the inhibitory neurons expressing glucagon-like peptide 1 receptor (GLP1r) in the MnPO respond to drinking actions and transiently inhibits thirst.
- 28.Leib DE, Zimmerman CA, Poormoghaddam A, Huey EL, Ahn JS, Lin YC, Tan CL, Chen Y, Knight ZA: The forebrain thirst circuit drives drinking through negative reinforcement. Neuron 2017, 96:1272–1281 e1274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gizowski C, Zaelzer C, Bourque CW: Clock-driven vasopressin neurotransmission mediates anticipatory thirst prior to sleep. Nature 2016, 537:685–688. [DOI] [PubMed] [Google Scholar]; • The first study to demonstrate that the suprachiasmatic nucleus (SCN) vasopressin neurons that project to the OVLT are more active and increase AVP release before sleep and prevent night-time dehydration.
- 30.Bernardis LL, Bellinger LL: The lateral hypothalamic area revisited: ingestive behavior. Neurosci Biobehav Rev 1996, 20:189–287. [DOI] [PubMed] [Google Scholar]
- 31.Kurt G, Woodworth HL, Fowler S, Bugescu R, Leinninger GM: Activation of lateral hypothalamic area neurotensin-expressing neurons promotes drinking. Neuropharmacology 2018. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Allen WE, DeNardo LA, Chen MZ, Liu CD, Loh KM, Fenno LE, Ramakrishnan C, Deisseroth K, Luo L: Thirst-associated preoptic neurons encode an aversive motivational drive. Science 2017, 357:1149–1155. [DOI] [PMC free article] [PubMed] [Google Scholar]; • This study characterizes neurons activated under water deprivation in MnPO using activity-dependent genetic labeling and single-cell transcriptional profiling. Optogenetic activation of these neurons induces drinking of water and aversive thirst motivational state.
- 33.Matsuda T, Hiyama TY, Niimura F, Matsusaka T, Fukamizu A, Kobayashi K, Kobayashi K, Noda M: Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ. Nat Neurosci 2017, 20:230–241. [DOI] [PubMed] [Google Scholar]
- 34.Andersson B, Leksell LG, Lishajko F: Perturbations in fluid balance induced by medially placed forebrain lesions. Brain Res 1975, 99:261–275. [DOI] [PubMed] [Google Scholar]
- 35.Buggy J, Jonhson AK: Preoptic-hypothalamic periventricular lesions: thirst deficits and hypernatremia. Am J Physiol 1977, 233:R44–R52. [DOI] [PubMed] [Google Scholar]
- 36.Johnson AK, Buggy J: Periventricular preoptic-hypothalamus is vital for thirst and normal water economy. Am J Physiol 1978, 234:R122–R129. [DOI] [PubMed] [Google Scholar]
- 37.Eng R, Miselis RR: Polydipsia and abolition of angiotensin-induced drinking after transections of subfornical organ efferent projections in the rat. Brain Res 1981, 225:200–206. [DOI] [PubMed] [Google Scholar]
- 38.Lind RW, Johnson AK: Subfornical organ-median preoptic connections and drinking and pressor responses to angiotensin II. J Neurosci 1982, 2:1043–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hochstenbach SL, Ciriello J: Effect of lesions of forebrain circumventricular organs on c-fos expression in the central nervous system to plasma hypernatremia. Brain Res 1996, 713:17–28. [DOI] [PubMed] [Google Scholar]
- 40.Gardiner TW, Stricker EM: Hyperdipsia in rats after electrolytic lesions of nucleus medianus. Am J Physiol 1985, 248:R214–R223. [DOI] [PubMed] [Google Scholar]
- 41.Cunningham JT, Beltz T, Johnson RF, Johnson AK: The effects of ibotenate lesions of the median preoptic nucleus on experimentally-induced and circadian drinking behavior in rats. Brain Res 1992, 580:325–330. [DOI] [PubMed] [Google Scholar]
- 42.McKinley MJ, Mathai ML, Pennington G, Rundgren M, Vivas L: Effect of individual or combined ablation of the nuclear groups of the lamina terminalis on water drinking in sheep. Am J Physiol 1999, 276:R673–R683. [DOI] [PubMed] [Google Scholar]
- 43.Yang CF, Chiang MC, Gray DC, Prabhakaran M, Alvarado M, Juntti SA, Unger EK, Wells JA, Shah NM: Sexually dimorphic neurons in the ventromedial hypothalamus govern mating in both sexes and aggression in males. Cell 2013, 153:896–909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Buggy J, Fisher AE, Hoffman WE, Johnson AL, Phillips MI: Ventricular obstruction: effect on drinking induced by intracranial injection of angiotensin. Science 1975, 190:72–74. [DOI] [PubMed] [Google Scholar]
- 45.McKinley MJ, Cairns MJ, Denton DA, Egan G, Mathai ML, Uschakov A, Wade JD, Weisinger RS, Oldfield BJ: Physiological and pathophysiological influences on thirst. Physiol Behav 2004, 81:795–803. [DOI] [PubMed] [Google Scholar]
- 46.Towbin EJ: Gastric distention as a factor in the satiation of thirst in esophagostomized dogs. Am J Physiol 1949, 159:533–541. [DOI] [PubMed] [Google Scholar]
- 47.Thrasher TN, Nistal-Herrera JF, Keil LC, Ramsay DJ: Satiety and inhibition of vasopressin secretion after drinking in dehydrated dogs. Am J Physiol 1981, 240:E394–E401. [DOI] [PubMed] [Google Scholar]
- 48.Maddison S, Wood RJ, Rolls ET, Rolls BJ, Gibbs J: Drinking in the rhesus monkey: peripheral factors. J Comp Physiol Psychol 1980, 94:365–374. [DOI] [PubMed] [Google Scholar]
- 49.Mandelblat-Cerf Y, Kim A, Burgess CR, Subramanian S, Tannous BA, Lowell BB, Andermann ML: Bidirectional anticipation of future osmotic challenges by vasopressin neurons. Neuron 2017, 93:57–65. [DOI] [PMC free article] [PubMed] [Google Scholar]; • This study shows an anticipatory control of AVP neurons in the SON and PVH. AVP neurons are rapidly inhibited both upon presentation of water-predicting cues and during drinking of water. In contrast, food availability increases AVP neuron activity only following feeding onset.
- 50.Abbott SB, Machado NL, Geerling JC, Saper CB: Reciprocal control of drinking behavior by median preoptic neurons in mice. J Neurosci 2016, 36:8228–8237. [DOI] [PMC free article] [PubMed] [Google Scholar]; • The authors demonstrate that the glutamate and GABA neurons in the MnPO/OVLT in mice positively and negatively regulate water drinking, respectively.
- 51.Hyde TM, Miselis RR: Area postrema and adjacent nucleus of the solitary tract in water and sodium balance. Am J Physiol 1984, 247:R173–R182. [DOI] [PubMed] [Google Scholar]
- 52.Ohman LE, Johnson AK: Brain stem mechanisms and the inhibition of angiotensin-induced drinking. Am J Physiol 1989, 256:R264–R269. [DOI] [PubMed] [Google Scholar]
- 53.Zheng H, Patterson LM, Rhodes CJ, Louis GW, Skibicka KP, Grill HJ, Myers MG Jr, Berthoud HR: A potential role for hypothalamomedullary POMC projections in leptin-induced suppression of food intake. Am J Physiol Regul Integr Comp Physiol 2010, 298:R720–R728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Vivas L, Godino A, Dalmasso C, Caeiro XE, Macchione AF, Cambiasso MJ: Neurochemical circuits subserving fluid balance and baroreflex: a role for serotonin, oxytocin, and gonadal steroids In Neurobiology of Body Fluid Homeostasis: Transduction and Integration. Edited by De Luca LA Jr, Menani JV, Johnson AK. 2014. Frontiers in Neuroscience. [PubMed] [Google Scholar]
- 55.Huo L, Maeng L, Bjorbaek C, Grill HJ: Leptin and the control of food intake: neurons in the nucleus of the solitary tract are activated by both gastric distension and leptin. Endocrinology 2007, 148:2189–2197. [DOI] [PubMed] [Google Scholar]
- 56.Powley TL: Vagal circuitry mediating cephalic-phase responses to food. Appetite 2000, 34:184–188. [DOI] [PubMed] [Google Scholar]
- 57.Roman CW, Derkach VA, Palmiter RD: Genetically and functionally defined NTS to PBN brain circuits mediating anorexia. Nat Commun 2016, 7:11905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ryan PJ, Ross SI, Campos CA, Derkach VA, Palmiter RD: Oxytocin-receptor-expressing neurons in the parabrachial nucleus regulate fluid intake. Nat Neurosci 2017, 20:1722–1733. [DOI] [PMC free article] [PubMed] [Google Scholar]; • This study shows that Oxytocin receptor-expressing neurons in the parabrachial nucleus (OxtrPBN neurons) have a selective role in fluid satiation. OxtrPBN neurons are rapidly activated after drinking of water, but not after drinking of caloric liquid diet, suggesting that OxtrPBN neurons relay pre-absorptive inputs.
- 59.Menani JV, De Luca LA Jr, Johnson AK: Role of the lateral parabrachial nucleus in the control of sodium appetite. Am J Physiol Regul Integr Comp Physiol 2014, 306:R201–R210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Johnson AK, Thunhorst RL: The neuroendocrinology of thirst and salt appetite: visceral sensory signals and mechanisms of central integration. Front Neuroendocrinol 1997, 18:292–353. [DOI] [PubMed] [Google Scholar]
- 61.Jhamandas JH, Petrov T, Harris KH, Vu T, Krukoff TL: Parabrachial nucleus projection to the amygdala in the rat: electrophysiological and anatomical observations. Brain Res Bull 1996, 39:115–126. [DOI] [PubMed] [Google Scholar]
- 62.MenaniJV ColombariDS, BeltzTG ThunhorstRL, JohnsonAK:Salt appetite: interaction of forebrain angiotensinergic and hindbrain serotonergic mechanisms. Brain Res 1998, 801:29–35. [DOI] [PubMed] [Google Scholar]
- 63.Menezes MF, Barbosa SP, De Andrade CA, Menani JV, De Paula PM: Purinergic mechanisms of lateral parabrachial nucleus facilitate sodium depletion-induced NaCl intake. Brain Res 2011, 1372:49–58. [DOI] [PubMed] [Google Scholar]
- 64.Hollis JH, McKinley MJ, D’Souza M, Kampe J, Oldfield BJ: The trajectory of sensory pathways from the lamina terminalis to the insular and cingulate cortex: a neuroanatomical framework for the generation of thirst. Am J Physiol Regul Integr Comp Physiol 2008, 294:R1390–R1401. [DOI] [PubMed] [Google Scholar]
- 65.Saker P, Farrell MJ, Adib FR, Egan GF, McKinley MJ, Denton DA: Regional brain responses associated with drinking water during thirst and after its satiation. Proc Natl Acad Sci USA 2014, 111:5379–5384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zocchi D, Wennemuth G, Oka Y: The cellular mechanism for water detection in the mammalian taste system. Nat Neurosci 2017, 20:927–933. [DOI] [PubMed] [Google Scholar]
- 67.Ramsay DJ, Booth D: In Thirst: Physiological and Psychological Aspects. Edited by Ramsay DJ, Booth D. Springer; 1991. Ch. 5, 6, 9–12, 19. [Google Scholar]
- 68.Simpson JB, Routtenberg A: Subfornical organ: site of drinking elicitation by angiotensin II. Science 1973, 181:1172–1175. [DOI] [PubMed] [Google Scholar]
- 69.Mangiapane ML, Simpson JB: Subfornical organ: forebrain site of pressor and dipsogenic action of angiotensin II. Am J Physiol 1980, 239:R382–R389. [DOI] [PubMed] [Google Scholar]
- 70.Takei Y: The role of the subfornical organ in drinking induced by angiotension in the Japanese quail, coturnix coturnix japonica. Cell Tissue Res 1977, 185:175–181. [DOI] [PubMed] [Google Scholar]
- 71.Robertson A, Kucharczyk J, Mogenson GJ: Drinking behavior following electrical stimulation of the subfornical organ in the rat. Brain Res 1983, 274:197–200. [DOI] [PubMed] [Google Scholar]
- 72.Smith PM, Beninger RJ, Ferguson AV: Subfornical organ stimulation elicits drinking. Brain Res Bull 1995, 38:209–213. [DOI] [PubMed] [Google Scholar]
- 73.Ferguson AV, Kasting NW: Electrical stimulation in subfornical organ increases plasma vasopressin concentrations in the conscious rat. Am J Physiol 1986, 251:R425–R428. [DOI] [PubMed] [Google Scholar]
- 74.Takei Y: Angiotensin and water intake in the Japanese quail (Conturnix coturnix japonica). Gen Comp Endocrinol 1977, 31:364–372. [DOI] [PubMed] [Google Scholar]
- 75.O’Neill TP, Brody MJ: Role for the median preoptic nucleus in centrally evoked pressor responses. Am J Physiol 1987, 252: R1165–R1172. [DOI] [PubMed] [Google Scholar]
- 76.Bealer SL, Phillips MI, Johnson AK, Schmid PG: Anteroventral third ventricle lesions reduce antidiuretic responses to angiotensin II. Am J Physiol 1979, 236:E610–E615. [DOI] [PubMed] [Google Scholar]
- 77.Mangiapane ML, Thrasher TN, Keil LC, Simpson JB, Ganong WF: Role for the subfornical organ in vasopressin release. Brain Res Bull 1984, 13:43–47. [DOI] [PubMed] [Google Scholar]
- 78.McKinley MJ, Mathai ML, McAllen RM, McClear RC, Miselis RR, Pennington GL, Vivas L, Wade JD, Oldfield BJ: Vasopressin secretion: osmotic and hormonal regulation by the lamina terminalis. J Neuroendocrinol 2004, 16:340–347. [DOI] [PubMed] [Google Scholar]
- 79.Gardiner TW, Verbalis JG, Stricker EM: Impaired secretion of vasopressin and oxytocin in rats after lesions of nucleus medianus. Am J Physiol 1985, 249:R681–R688. [DOI] [PubMed] [Google Scholar]
- 80.Thrasher TN, Simpson JB, Ramsay DJ: Lesions of the subfornical organ block angiotensin-induced drinking in the dog. Neuroendocrinology 1982, 35:68–72. [DOI] [PubMed] [Google Scholar]
- 81.Mangiapane ML, Thrasher TN, Keil LC, Simpson JB, Ganong WF: Deficits in drinking and vasopressin secretion after lesions of the nucleus medianus. Neuroendocrinology 1983, 37:73–77. [DOI] [PubMed] [Google Scholar]
- 82.Thrasher TN, Keil LC, Ramsay DJ: Lesions of the organum vasculosum of the lamina terminalis (OVLT) attenuate osmotically-induced drinking and vasopressin secretion in the dog. Endocrinology 1982, 110:1837–1839. [DOI] [PubMed] [Google Scholar]
- 83.Fitts DA, Freece JA, Van Bebber JE, Zierath DK, Bassett JE: Effects of forebrain circumventricular organ ablation on drinking or salt appetite after sodium depletion or hypernatremia. Am J Physiol Regul Integr Comp Physiol 2004, 287:R1325–R1334. [DOI] [PubMed] [Google Scholar]
- 84.May CN, McAllen RM, McKinley MJ: Renal nerve inhibition by central NaCl and ANG II is abolished by lesions of the lamina terminalis. Am J Physiol Regul Integr Comp Physiol 2000, 279: R1827–R1833. [DOI] [PubMed] [Google Scholar]