

Abstract
Although the retrosplenial cortex (RSC) is involved in spatial learning and memory, it appears to make less significant contributions to learning and memory for discrete cues. For example, damage to the RSC does not impair Pavlovian delay fear conditioning to a discrete auditory cue (e.g., tone), when RSC manipulation occurs just prior or shortly after conditioning. In contrast, when lesions of the RSC occur following a substantial retention interval (e.g., 28 days), the RSC is necessary for retrieval of fear to the tone. Thus, the RSC makes time-dependent contributions to memory retrieval for discrete auditory cues. The purpose of the current experiment was to assess if the time-dependent involvement of the RSC in cue-specific fear memory extended to cues of other sensory modalities. Rats firsts underwent fear conditioning to a visual conditioning stimulus, and lesions of the RSC subsequently occurred 1- or 28-days later. Lesions of the RSC impaired fear expression when made 28-days after conditioning, but not when made 1-day following conditioning. Coupled with previous findings, the current results suggest the RSC is necessary for retrieval of remotely acquired cued fear memories across multiple modalities.
Keywords: retrosplenial, remote memory, fear conditioning, visual cue
Over the past decade there has been a surge of research on the contributions of the retrosplenial cortex (RSC) to learning, memory, and behavior. Although there is still much to understand and unify regarding the function of this region and its interactions with medial temporal lobe structures, it has become clear that the RSC is especially involved in spatial navigation as well as contextual learning and memory (for reviews see Miller, Vedder, Law & Smith, 2014; Todd and Bucci, 2015; Vann, Aggleton, & McGuire, 2009). For example, in vivo electrophysiological recording studies have demonstrated that RSC neurons process critical information related to landmarks, trajectories, and reward location (Vedder et al., 2017), as well as the overall spatial structure of complex routes (Alexander & Nitz, 2017; Clark, 2017). More so, the RSC is critically involved in contextual fear conditioning (e.g., Corcoran, 2011; Kwapis et al., 2015; Todd et al., 2017) a process that likely involves communication between the RSC and hippocampus (Tayler et al., 2013). Importantly, both spatial navigation and contextual learning and memory typically require the integration of information from multiple cues in the environment.
Apart from spatial navigation and contextual learning and memory, there is renewed interest in understanding how the RSC contributes to learning and memory for discrete stimuli. While an earlier body of work by Gabriel and colleagues focused on the role of the RSC in avoidance discrimination learning involving discrete auditory stimuli (Gabriel, Sparenborg, & Stolar, 1987; Gabriel & Sparenborg, 1987), recent studies have tested the involvement of the RSC in Pavlovian fear conditioning, in which a single auditory cue is associated with mild foot-shock. In rodents, these studies have consistently demonstrated little, if any, contribution of the RSC to encoding or retrieval of “delay” auditory fear conditioning, in which a tone is presented for a short period of time (e.g., 10 s) and co-terminates with a mild foot-shock (Corcoran et al., 2011; Keene & Bucci, 2008b; Kwapis et al., 2014, 2015). The fact that the RSC is not necessary for learning and memory for a single cue in Pavlovian delay fear conditioning procedures has led some to suggest that the RSC is specifically involved in situations that require the processing and integration of multiple cues (e.g., Keene & Bucci, 2008a; Bucci & Robinson, 2014), perhaps consistent with the RSC’s important role in spatial and contextual learning and memory.
There are however, exceptions, in which the RSC does contribute significantly to learning and memory for single cues in Pavlovian fear conditioning procedures. For example, insertion of a short interval between the end a Pavlovian conditioned stimulus (CS) and presentation of shock (so called “trace” conditioning”) appears to recruit the RSC (see Kwapis et al., 2014; 2015). Additionally, and of relevance to the current set of experiments, we recently found that as cue-specific memories age, they become dependent upon the RSC for their retrieval. For example, disrupting RSC function with lesions or temporary inactivation impairs the retrieval of conditioned fear to a tone that was acquired 28 days earlier (i.e., remote fear memory; Todd et al., 2016a), but has no effect on tone fear conditioning that occurred 1 day earlier (i.e., recent memory; Keene & Bucci, 2008b). Although the nature of this time-dependent shift is still unclear, these findings are consistent with the fact that the RSC receives direct projections from secondary auditory cortex (Todd, et al., 2016a, Vogt & Miller, 1983), a region necessary for the retrieval of remote, but not recently, acquired fear memories (Sacco & Sachettic, 2010).
In addition to its connections with auditory cortex, the RSC has strong reciprocal connections with visual cortex (van Groen & Wyss, 1990, 1992, 1993). Consistent with these connections, RSC neurons are responsive to visual cues (e.g., Vedder et al., 2016), and RSC damage attenuates discrimination learning between multiple visual cues (e.g., Todd et al., 2016b; Bussey et al., 1996; 1997). Nevertheless, studies to date indicate that the RSC is not necessary for Pavlovian delay conditioning with a single visual cue. For example, pre-training lesions of the RSC have no impact on the acquisition of Pavlovian delay conditioning to a visual conditioned stimulus with either shock (Todd et al., 2016c, Experiment 1) or food (Keene & Bucci, 2008) as the reinforcer. Likewise, lesions made shortly after Pavlovian fear conditioning have no measurable impact on the expression of conditioned fear to a visual stimulus (Todd et al., 2016c, Experiment 2). However, no study has examined the involvement of the RSC in the retrieval of remotely acquired memory for a visual conditioned stimulus. To address this, lesions of the RSC were carried out either 1 day (Experiment 1) or 28 days (Experiment 2) after fear conditioning in which a light was paired with foot shock. We hypothesized that, similar to the involvement of the RSC in auditory fear conditioning, remotely, but not recently-acquired fear memories for a visual stimulus would be RSC dependent.
Experiment 1
Methods
Subjects
The subjects were 18 naïve male Long–Evans rats (~60 days old at start of training), obtained from Envigo Laboratories, Inc., (Indianapolis, IN). Rats were housed individually and allowed at least 6 days to acclimate to the vivarium prior to surgery. Food and water was available ad libitum (Purina standard rat chow: Nestle Purina, St. Louis, MO). Throughout the study, rats were maintained on a 14:10 light-dark cycle and monitored and cared for in compliance with association for Assessment and Accreditation of laboratory Animal Care guidelines and the Dartmouth College Institutional Animal Care and Use Committee.
Surgery
Subjects were anesthetized with isoflurane gas (1.5%–3% in oxygen) and placed in a Kopf stereotaxic apparatus. The skin was retracted and holes were drilled through the skull above each of the intended lesion sites using the rat brain atlas of Paxinos and Watson (2009). RSC lesion rats received bilateral electrolytic lesions (2.5 mA, 15 s at each site) of RSC 24 hours (n = 10) after behavioral training using the stereotaxic coordinates outlined in Table 1. Control rats received sham lesions (n = 8) consisting of a craniotomy and shallow, non-puncturing burr holes to minimize damage to underlying cortex either 24 hours after training. All rats were allowed to recover for 10 days before testing began.
Table 1.
Stereotaxic coordinates for restrosplenial cortex (RSC) lesions.
| AP | ML | DV |
|---|---|---|
| −2.0 | ± 0.3 | −2.0 and −2.7 |
| −3.5 | ± 0.4 | −2.0 and −2.7 |
| −5.0 | ± 0.4 and ± 1.0 | −2.0 and −2.7 (medial site) and −2.0 (lateral site) |
| −6.5 | ± 0.8 and ± 1.5 | −2.0 and −2.8 (medial site) and −3.4 (lateral site) |
| −8.0 | ± 1.6 and ± 2.4 | −2.5 (medial site) and −3.1 (lateral site) |
| −9.0 | ± 3.4 | −4.0 |
Note. All anterior-posterior (AP), medial-lateral (ML) and dorsal-ventral (DV) measurements are derived from bregma, midline, and skull surface, respectively (measurements are in mm).
Behavioral Apparatus
Two sets of four conditioning chambers served as the two contexts (counterbalanced). All chambers were of the same standard design (Med Associates, Inc., St. Albans, VT, ENV-007; 24 cm W × 30.5 cm L × 29 cm H) and each was housed in its own sound attenuation chamber (Med Associates, ENV-017M; 66 cm W × 56 cm L × 56 cm H) and outfitted with an exhaust fan to provide airflow and background noise (~68 dB). Each chamber was outfitted with a food cup, recessed in the center of the front wall, and a retractable lever (Med Associates, ENV-112CM) positioned to the right of the food cup, which remained retracted throughout the experiment. Each chamber had a panel light (Med Associates, ENV-221M) mounted approximately 16 cm above the grid floor centered over the food cup, and a house light (Med Associates, ENV-215M) mounted approximately 24 cm above the grid floor on the back wall of the chamber. A speaker (Med Associates, ENV-224AM) was located 20 cm above and to the right of the food cup. Both sets of chambers were illuminated with a 2.8 W bulb (with a red cover), mounted to the ceiling of the sound attenuating chamber.
In one set of chambers, the sidewalls and ceiling were made of clear acrylic plastic and the front and rear walls were made of brushed aluminum. The grid floor was stainless steel rods (5 mm diameter) spaced 1.5 cm apart (center-to-center). In the second set of chambers the grids of the floor were staggered such that odd- and even-numbered grids were mounted in two separate planes, one 0.5 cm above the other. The staggered grid floor provided a distinct tactile feature. In these chambers, the ceiling and door were covered with laminated black and white checkerboard paper (1 cm squares) to provide distinct visual cues.
Because these two sets of chambers were located within the same room of the laboratory, in order to prevent diffusion of the olfactory cues, one olfactory cue was used for Context A sessions, and a second olfactory cue was used for Context B sessions. During Context A sessions, 3 mL of Pine-Sol (Clorox, Co., Oakland, CA) was placed in the chamber tray below the grid floor, and for Context B sessions approximately 0.5 grams of Vicks Vaporub (Proctor & Gamble, Cincinnati, OH) was smeared along the chamber tray below the grid floor.
The panel light mounted to the front wall served as the visual stimulus. During CS presentation, it flashed twice per second for 10 seconds. Footshocks (1 mA, 1 s) were generated by a Med Associates shock generator (ENV-414) connected to each chamber. The apparatus was controlled by computer equipment located in an adjacent room. Surveillance cameras located inside the sound-attenuating chambers were used to monitor the rats’ behavior.
Behavioral Procedures
The training session consisted of three 10-second presentations of the light co-terminating with the foot shock. The interval from shock to the next light (intertrial interval, ITI) was 64 seconds. The first trial began 3 minutes after the rat was placed in the chamber. Following recovery from surgery, rats were then re-exposed to the original training chamber (Context A) for a single 20-minute context test session during which no tones or shocks were presented. Twenty-four hours after this context test, a light test session was carried out by placing the rats in Context B and presenting the light 20 times (10 s each, 30 s ITI) beginning 3 minutes after the rat was placed in the chamber. Again, no shock was delivered during this test session.
Behavioral Observations
Freezing served as the index of conditioned fear and was operationally defined as total motor immobility except for breathing (Blanchard & Blanchard, 1969; Fanselow, 1980). On the training day, the incidence of freezing behavior was recorded during the 64 second period prior to the first trial (baseline freezing) and during the 64 second period following each trial (post-shock freezing). The rats’ behavior was scored every 8 seconds during the 64 second epochs and the mean percent freezing across the three post-shock epochs was calculated for each rat. For the context test session, each rat was scored every 8 seconds for the first 8 min and 32 s, yielding 64 observations for each rat (Maren et al., 1997). For the light test session, freezing was recorded every 2 seconds during each 10 second presentation of the light. For each rat, the data was used to calculate the average freezing during the context test session and the average freezing during the light test session. The frequency of freezing behavior was converted to a percentage of total observations. A single primary observer, blind to treatment condition, scored all the behavioral data, while a second observer scored a subset of the data to assess objectivity. The observations from both observers were highly correlated (r ≥ 0.9).
Lesion verification and analysis.
After the behavioral procedures were completed rats were deeply anesthetized with an overdose of sodium pentobarbital and transcardially perfused with 0.9% saline for 2 min, followed by 10% buffered formalin for 6 min. Coronal brain sections (60 μm) were collected using a freezing microtome and were Nissl-stained using thionin. Using a compound microscope (Axioskop I, Zeiss, Inc.), we identified gross tissue damage as necrosis, missing tissue, or marked thinning of the cortex. Outlines of the lesions were drawn onto digital images adapted from Paxinos and Watson (2009) using PowerPoint at 6 levels along the rostro-caudal extent of the RSC (−1.8, −3.0, −4.2, −5.4, −6.6, and −7.8 mm from bregma). At each level, area measurements where then made with ImageJ, including the total area of the target region and the area of the target region that exhibited gross tissue damage. From these measurements, we report the average percentage of RSC that was damaged. In addition, we report the average percentage of sections across the rostro-caudal plane that exhibited RSC damage (out of ~24 sections collected for each rat), the average percentage of sections with damage outside the RSC, and the number of rats with damage to regions outside the RSC. Finally, we report the average percentage of secondary visual cortex that was damaged, at points −4.2, −5.4, −6.6, and −7.8 from bregma.
Results and Discussion
Histology
Two rats were excluded from statistical analysis due to substantial external damage. The remaining groups sizes were: Sham (n = 8) and RSC (n = 8). Figure 1a shows a photomicrograph of a representative RSC lesion. In Figure 1b (Experiment 1), lesion drawings are stacked onto a single atlas image for each of the 6 coronal levels. Bilateral damage to the RSC was observed in all animals. The average area of RSC damage on each section analyzed was 70.6 % (SEM = 2.83). Damage to the RSC was present on 90.7% (SEM = 1.02) of sections collected, indicating that damage extended throughout the rostro-caudal extent of the RSC. In all rats, there was minor damage to some areas outside the RSC (e.g., visual cortex, motor cortex, cingulum bundle, forceps major corpus callosum). The average area of secondary visual cortex damage was 6.4%. Damage to the secondary visual cortex was exclusively within the medial portion; there was no damage to lateral secondary visual cortex.
Figure 1.
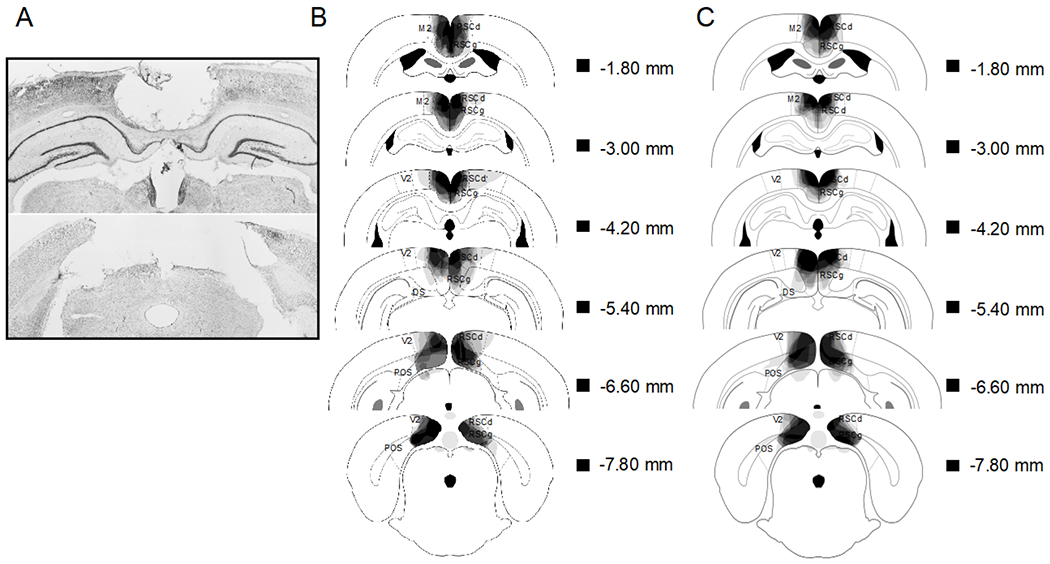
A: Photomicrograph of a representative RSC lesion. B (Experiment 1) and C (Experiment 2): drawings of lesions at six levels along the rostro-caudal extent of the RSC (−1.8, −3.0, −4.2, −5.4, −6.6, and −7.8 posterior to bregma). At each level, lesion drawings were stacked onto a single image. The darkness of an area indicates the number of lesions cases that include that area. Grey boxes (next to the bregma values) represents the expected darkness for overlap from all subjects. DS = dorsal subiculum , POS = postsubiculum, M2 = secondary motor cortex, RSCd = restrosplenial dysgranular, RSCg = retrosplenial granula, V2 = secondary visual cortex.
Behavior
The results from Experiment 1 are presented in Figure 2. Freezing during the baseline period and the post-shock period were analyzed with a 2 (Lesion: sham vs. RSC) × 2 (Period: baseline vs. post-shock) ANOVA. The analysis revealed a main effect of period, F(1, 14) = 248.17, p < .001, indicating more freezing during the post-shock period than the baseline period. Neither the main effect of lesion, F(1, 14) < 1, p = .95, nor the lesion by period interaction, F(1, 14) < 1, p = .95 were significant. During the context test session, RSC-lesioned rats froze significantly less than Sham lesioned rats, F(1, 14) = 28.53, p < .001. Data from the light test session are presented in the right panel of Figure 2. Freezing to the light itself is presented in 5-trial blocks. (Note: the first 5-trial block is also plotted in bar form in the left panel). Freezing during the light test was analyzed with a 2 (Lesion: sham vs. RSC) × 5 (Period: baseline, Block 1 - 4) ANOVA. This analysis revealed a main effect of period, F(4, 56) = 39.37, p < .001. Neither the main effect of lesion, F(1, 14) < 1, p = .58, nor the lesion by period interaction, F(4, 56) < 1, p = .80 were significant.
Figure 2.

Left panel: results of Experiment 1. “BL” = freezing during the baseline period (prior to light-shock pairings) in the training session, “Post-Shock” = freezing during the three post-shock periods of the training session, “Context Test” = freezing during the test session in Context A, “Light Test” = freezing during the first 5 trial-block of the light test in Context B. Right panel: results from the Light Test session plotted in 5-trial blocks. “BL”: freezing during the baseline period just prior to the presentation of the first light stimulus. Sham = sham lesioned rats, RSC = retrosplenial lesioned rats. Error bars represent ±1 SEM. *p < .05.
The findings from the current experiment are consistent with previous studies examining the role of the RSC in learning and memory for discrete auditory cues. As in previous studies, lesions made shortly after conditioning impaired fear expression to the context, but not the discrete cue (Keene & Bucci, 2008). Further, while lesions made shortly after conditioning have no impact on freezing to a tone conditioned stimulus (e.g., Keene & Bucci, 2008), the current experiment demonstrates RSC lesions, likewise, do not impair freezing to a visual conditioned stimulus. Thus, although the RSC is necessary for the retrieval of contextual fear memory, it is not necessary for retrieval of recently acquired fear to a visual conditioned stimulus (see also Todd et al., 2016b).
Experiment 2
Subjects and surgery
The subjects were 20 experimentally naïve adult male Long Evans rats, purchased from the same vendor as those in the previous experiment and maintained under the same conditions. Surgical procedures for Sham and RSC lesions were the same as Experiment 1.
Behavioral apparatus, procedures and observations
The apparatus and procedures were the same as those used in Experiment 1, with the exception that all lesions occurred 28-days after behavioral training.
Lesion verification and analysis.
RSC lesions were verified and analyzed using the same procedures as Experiment 1.
Results and Discussion
Histology
One rat was excluded from statistical analysis due to substantial extra-RSC damage. In addition, 1 Sham and 1 RSC lesioned rat died during surgery. The remaining groups sizes were: Sham (n = 8) and RSC (n = 9). In Figure 1c (Experiment 2), lesion drawings are stacked onto a single atlas image for each of the 6 coronal levels. Bilateral damage to the RSC was observed in all animals. The average area of RSC damage on each section analyzed was 65.2% (SEM = 2.81). Damage to the RSC was present on 87.9% (SEM = 1.27) of sections collected, indicating that damage extended throughout the rostro-caudal extent of the RSC and was very similar to Experiment 1. In all rats, there was minor damage outside the RSC (e.g., visual cortex, motor cortex, cingulum bundle, forceps major corpus callosum). The average area of secondary visual cortex damage was 5.6%. Damage to the secondary visual cortex was exclusively within the medial portion; there was no damage to lateral secondary visual cortex.
Behavior
The results from Experiment 2 are presented in Figure 3. Freezing during the baseline period and the post-shock period were analyzed with a 2 (Lesion: sham vs. RSC) × 2 (Period: baseline vs. post-shock) ANOVA. The analysis revealed a main effect of period, F(1, 15) = 511.90, p < .001, indicating more freezing during the post-shock period than the baseline period. Neither the main effect of lesion, F(1, 15) < 1, p = .70, nor the lesion by period interaction, F(1, 15) < 1, p = .70 were significant. During the context test session, RSC-lesioned rats froze significantly less than Sham lesioned rats, F(1, 15) = 39.13, p < .001. Data from the light test session are presented in the right panel of Figure 3. Freezing to the light itself is presented in 5-trial blocks. (Note: the first 5-trial block is also plotted in bar form in the left panel). Freezing during the light test was analyzed with a 2 (Lesion: sham vs. RSC) × 5 (Period: baseline, Block 1 – 4) ANOVA. This analysis revealed a main effect of period, F(4, 60) = 30.75, p < .001. Although the main effect of lesion was not significant, F(1, 15) = 2.16, p = .16, the interaction between lesion and period approached significance, F(4, 60) = 2.46, p = .055. The effect of lesion was significant during Block 1, F(1, 14) = 7.10, p = .018, but not Block 2, Block 3, or Block 4 (all ps > .24). The level of freezing did not differ between groups during the baseline period of the light test F(1, 14) = 2.51, p = .14.
Figure 3.
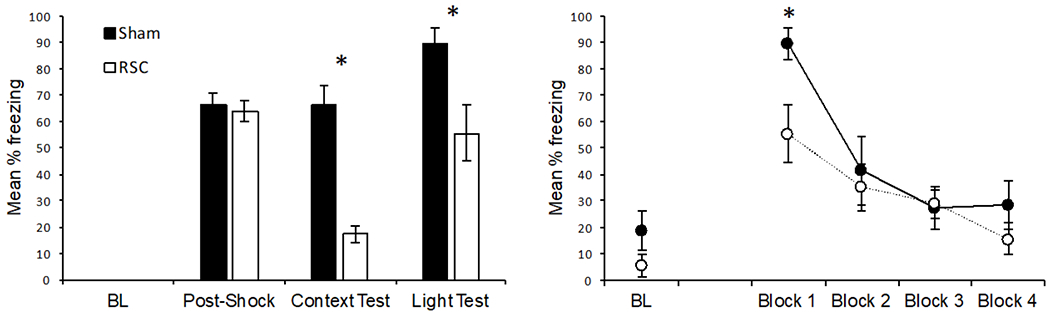
Left panel: results of Experiment 2. “BL” = freezing during the baseline period (prior to light-shock pairings) in the training session, “Post-Shock” = freezing during the three post-shock periods of the training session, “Context Test” = freezing during the test session in Context A, “Light Test” = freezing during the first 5 trial-block of the light test in Context B. Right panel: results from the Light Test session plotted in 5-trial blocks. “BL”: freezing during the baseline period just prior to the presentation of the first light stimulus. Sham = sham lesioned rats, RSC = retrosplenial lesioned rats. Error bars represent ±1 SEM. *p < .05.
Lesions of the RSC impaired retrieval of remote contextual fear, consistent with previous findings (Corcoran et al., 2011; Todd et al., 2016a, Experiment 1). Further, lesions made following a substantial retention interval (e.g., 28 days) also impaired retrieval of fear to a visual conditioned stimulus. This finding is in contrast to Experiment 1, where lesions made 1 day after conditioning had no impact on fear expression to the visual cue. Overall, these results are consistent with, and extend the results of Todd et al. (2016a), demonstrating the RSC has a time-dependent role in memory retrieval for discrete cues.
Sacco & Sachetti (2010) have previously reported that lesions of the secondary visual cortex impair remote memory for visual cues (Sacco & Sachetti, 2010). Thus, it is notable that we observed incidental damage to the medial portion of the secondary visual cortex in all rats. We note, however, that the incidental damage to the medial secondary visual cortex was very minor (~ 6%). In addition, we note that Sacco & Sachetti (2010) lesioned the lateral secondary visual cortex. There was no damage to this region in the current experiment. Moreover, there was no significant correlation between the amount of damage to secondary visual cortex and freezing to the light (p = .36).
General Discussion
Our previous studies have demonstrated a role for the RSC in the retrieval of remotely, but not recently, acquired fear memories in a Pavlovian delay fear conditioning procedure with an auditory CS (Keene & Bucci, 2008; Todd et al., 2016a). The purpose of the current experiment was to assess if the time-dependent involvement of the RSC in cue-specific fear memory extended to cues of other sensory modalities, or if it was unique to auditory stimuli. Lesions of the RSC made 1-day after training had no detectable impact on freezing to a visual cue (Experiment 1); however, lesions made 28-days after training did attenuate fear elicited by the visual cue (Experiment 2). These data provide evidence that the RSC is necessary specifically for the retrieval remotely acquired fear memories to visual cues. In both experiments, freezing to the context was impaired, consistent with prior studies demonstrating a time-independent role for RSC in contextual fear memory (e.g., Corcoran et al., 2011; Todd et al., 2016a, Experiment 1).
The present findings are consistent with the notion that the neurocircuitry of remote cued fear memory differs from that of recent cued fear memory (for a review see Bergstrom, 2016). In contrast to recently acquired cued fear memories, remotely acquired cued fear memories depend upon secondary sensory cortices for retrieval (Sacco & Sachetti, 2010). For example, post-training lesions of the auditory cortex impair the retrieval of remote, but not recent, memory for auditory cues. Likewise, lesions of the piriform cortex impair remote memory retrieval for olfactory cues, and lesions of visual cortex impair remote memory retrieval for visual cues (Sacco & Sachetti, 2010). In most cases, the observed retrieval deficits were sensory-modality specific, leading Sacco & Sachetti (2010) to conclude that each secondary sensory cortex is involved in remote memory storage and retrieval for a specific sensory modality. The role of the RSC, however, appears to be more general. Along with the findings from Todd et al. (2016a), the current findings indicate the RSC is necessary for the retrieval of both visual and auditory remotely acquired cued fear memories. Thus, the RSC is necessary for retrieval of remotely acquired cued fear memories across multiple modalities.
The fact that the RSC has direct connections with both auditory cortex and visual cortex makes it well-suited to contribute to memory retrieval for cues from both modalities. It is, however, interesting to note that not all polymodal cortical areas are necessary for retrieval of remotely acquired cued fear memories. For example, although the perirhinal cortex receives both visual and auditory information (e.g., Furtak et al., 2007; Kealy & Commins, 2011), post-training lesions of the posterior portions of the perirhinal cortex do not impact retrieval of remotely acquired fear memories (Sacco & Sachetti, 2010). This suggests a functional dissociation between the role of the RSC and perirhinal cortex in the retrieval of remotely acquired cued fear memories. However, since the precise role of the RSC in remote memory retrieval is still unclear (see Todd et al., 2016a for a discussion), future research is necessary to determine not only the exact role of the RSC, but how RSC function differs from regions such as the perirhinal cortex.
Although we observed a time-dependent effect on the retrieval of memory for a visual cue, lesions of the RSC attenuated freezing to the context at both time points. This finding is consistent with prior research demonstrating that the RSC is both active (Tayler et al., 2013) and necessary (Corcoran et al., 2011) for the retrieval of contextual fear memories at both recent and remote time points. Nevertheless, it suggests a fundamental dissociation between contextual fear memories and cue specific fear memories. On the one hand, contextual fear memories are RSC dependent at both recent and remote time points, whereas on the other hand cued fear memories are only RSC dependent at remote time points.
In the current experiments, RSC damage was produced via electrolytic lesions made either 1- or 28-days following initial Pavlovian conditioning. Although electrolytic lesions damage both cell bodies and fibers of passage, it seems unlikely that the results observed are simply due to damage of fibers pathways, since our prior studies have demonstrated that either neurotoxic or electrolytic lesions of the RSC, or temporary silencing of RSC activity produce specific deficits in remote memory for auditory cues (Todd et al., 2016a).
In summary, the present findings add to a growing literature aimed at identifying the precise circumstances under which RSC is necessary for cue-specific learning and memory. For instance, the involvement of RSC in remote but not recent fear memory for visual or auditory CSs suggests a temporal dimension to its role in cue-specific memory, however, it is unclear whether RSC involvement is dichotomous (i.e., recent vs. remote) or varies along a gradient. While additional studies are necessary to fully define the functional contributions of RSC to learning and memory, one possibility is that the RSC serves to integrate information from multiple modalities in the service of long-term memory.
Acknowledgments
This work was supported by National Science Foundation Grant IOS1353137 (D.J.B.) and National Institutes of Health Grant F32MH105125 (T.P.T.).
References
- Alexander AS, & Nitz DA (2017). Spatially Periodic Activation Patterns of Retrosplenial Cortex Encode Route Sub-spaces and Distance Traveled. Current Biology, 27(11), 1551–1560. [DOI] [PubMed] [Google Scholar]
- Bergstrom HC (2016). The neurocircuitry of remote cued fear memory. Neuroscience and Biobehavioral Reviews, 71, 409–417. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, & Blanchard DC (1969). Crouching as an index of fear. Journal of Comparative and Physiological Psychology, 67(3), 370–375. [DOI] [PubMed] [Google Scholar]
- Bussey TJ, Muir JL, Everitt BJ, & Robbins TW (1997). Triple dissociation of anterior cingulate, posterior cingulate, and medial frontal cortices on visual discrimination tasks using a touchscreen testing procedure for the rat. Behavioral Neuroscience, 111(5), 920–936. [DOI] [PubMed] [Google Scholar]
- Bussey TJ, Muir JL, Everitt BJ, & Robbins TW (1996). Dissociable effects of anterior and posterior cingulate cortex lesions on the acquisition of a conditional visual discrimination: Facilitation of early learning vs. impairment of late learning. Behavioural Brain Research, 82(1), 45–56. [DOI] [PubMed] [Google Scholar]
- Clark BJ (2017). Spatial Navigation: Retrosplenial Cortex Encodes the Spatial Structure of Complex Routes. Current Biology, 27(13), 649–651. [DOI] [PubMed] [Google Scholar]
- Corcoran KA, Donnan MD, Tronson NC, Guzman YF, Gao C, Jovasevic V, … Radulovic J (2011). NMDA Receptors in Retrosplenial Cortex Are Necessary for Retrieval of Recent and Remote Context Fear Memory. Journal of Neuroscience, 31(32), 11655–11659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fanselow MS (1980). Conditional and unconditional components of post-shock freezing. The Pavlovian Journal of Biological Science, 15(4), 177–182. [DOI] [PubMed] [Google Scholar]
- Furtak SC, Wei SM, Agster KL, & Burwell RD (2007). Functional neuroanatomy of the parahippocampal region in the rat: The perirhinal and postrhinal cortices. Hippocampus. 17(9), 709–722. [DOI] [PubMed] [Google Scholar]
- Gabriel M, Sparenborg SP, & Stolar N (1987). Hippocampal control of cingulate cortical and anterior thalamic information processing during learning in rabbits. Experimental Brain Research, 67(1), 131–152. [DOI] [PubMed] [Google Scholar]
- Gabriel M, & Sparenborg S (1987). Posterior cingulate cortical lesions eliminate learning-related unit activity in the anterior cingulate cortex. Brain Research, 409(1), 151–157. [DOI] [PubMed] [Google Scholar]
- Kealy J, & Commins S (2011). The rat perirhinal cortex: A review of anatomy, physiology, plasticity, and function. Progress in Neurobiology, 93(4), 522–548. [DOI] [PubMed] [Google Scholar]
- Keene CS, & Bucci DJ (2008a). Contributions of the retrosplenial and posterior parietal cortices to cue-specific and contextual fear conditioning. Behavioral Neuroscience, 122(1), 89–97. [DOI] [PubMed] [Google Scholar]
- Keene CS, & Bucci DJ (2008b). Involvement of the retrosplenial cortex in processing multiple conditioned stimuli. Behavioral Neuroscience, 122(3), 651–658. [DOI] [PubMed] [Google Scholar]
- Kwapis JL, Jarome TJ, Lee JL, Gilmartin MR, & Helmstetter FJ (2014). Extinguishing trace fear engages the retrosplenial cortex rather than the amygdala. Neurobiology of Learning and Memory, 113, 41–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwapis JL, Jarome TJ, Lee JL, & Helmstetter FJ (2015). The retrosplenial cortex is involved in the formation of memory for context and trace fear conditioning. Neurobiology of Learning and Memory, 123, 110–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maren S, Aharonov G, & Fanselow MS (1997). Neurotoxic lesions of the dorsal hippocampus and Pavlovian fear conditioning in rats. Behavioural brain research, 88(2), 261–274. [DOI] [PubMed] [Google Scholar]
- Miller AM, Vedder LC, Law LM, & Smith DM (2014). Cues, context, and long-term memory: the role of the retrosplenial cortex in spatial cognition. Frontiers in human neuroscience, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paxinos G, & Watson C (1998). The Rat Brain in Stereotaxic Coordinates. Academic Press, (1), 1–474. [Google Scholar]
- Robinson S, Todd TP, Pasternak AR, Luikart BW, Skelton PD, Urban DJ, & Bucci DJ (2014). Chemogenetic silencing of neurons in retrosplenial cortex disrupts sensory preconditioning. J Neuroscience, 34(33), 10982–10988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacco T, & Sacchetti B (2010). Role of secondary sensory cortices in emotional memory storage and retrieval in rats. Science, 329, 649–656. [DOI] [PubMed] [Google Scholar]
- Tayler KK, Tanaka KZ, & Reijmers LG (2014). Reactivation of neural ensembles during the retrieval of recent and remote memory. Current Biology, 23, 99–106. [DOI] [PubMed] [Google Scholar]
- Todd TP, & Bucci DJ (2015). Retrosplenial cortex and long-term memory: molecules to behavior. Neural plasticity, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, Mehlman ML, Keene CS, DeAngeli NE, & Bucci DJ (2016a). Retrosplenial cortex is required for the retrieval of remote memory for auditory cues. Learning & Memory, 23, 278–288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, Huszár R, DeAngeli NE, & Bucci DJ (2016b). Higher-order conditioning and the retrosplenial cortex. Neurobiology of Learning and Memory, 133, 257–264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, DeAngeli NE, Jiang MY, & Bucci DJ (2017). Retrograde amnesia of contextual fear conditioning: Evidence for retrosplenial cortex involvement in configural processing. Behavioral Neuroscience, 131, 46–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Groen T, & Michael Wyss J (1990). Connections of the retrosplenial granular a cortex in the rat. Journal of Comparative Neurology, 300, 593–606. [DOI] [PubMed] [Google Scholar]
- van Groen T, Vogt BA, & Wyss JM (1993). Interconnections Between the Thalamus and Retrosplenial Cortex in the Rodent Brain In Vogt BA & Gabriel M (Eds.), Neurobiology of Cingulate Cortex and Limbic Thalamus: A Comprehensive Handbook (pp. 123–150). Boston, MA. [Google Scholar]
- Vann SD, Aggleton JP, & Maguire EA (2009). What does the retrosplenial cortex do? Nature Reviews. Neuroscience, 10, 792–802. [DOI] [PubMed] [Google Scholar]
- Vedder LC, Miller AMP, Harrison MB, & Smith DM (2017). Retrosplenial cortical neurons encode navigational cues, trajectories and reward locations during goal directed navigation. Cerebral Cortex, 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogt BA, & Miller MW (1983). Cortical connections between rat cingulate cortex and visual, motor, and postsubicular cortices. The Journal of Comparative Neurology, 216(2), 192–210. [DOI] [PubMed] [Google Scholar]
- Wyss JM, & Van Groen T (1992). Connections between the retrosplenial cortex and the hippocampal formation in the rat: a review. Hippocampus, 2(1), 1–11. [DOI] [PubMed] [Google Scholar]