

Abstract
The EFSA Panel on Plant Health (PLHP) performed a pest categorisation of Saperda tridentata (Coleoptera: Cerambycidae) for the EU. S. tridentata (elm borer) occurs in eastern North America. Ulmus americana and U. rubra are almost exclusively reported as hosts, apart from two 19th century records from the USA of larvae from Acer sp. and Populus sp. The Panel does not exclude the possibility of a post‐entry shift in host range to European Ulmus or Acer and Populus. S. tridentata infests trees that are already weakened, and severe infestations can result in tree death. S. tridentata occurs across a range of climate types in North America that occur also in Europe. Between 2016 and 2019, S. tridentata larvae were intercepted with North American Ulmus logs imported into the EU. In the EU, American Ulmus species are mainly found in arboreta and as ornamental specimen trees. If only North American Ulmus are hosts, establishment is unlikely. However, if European Ulmus, Populus or Acer species become hosts, establishment is much more likely, with impact confined to already weakened trees. The information currently available on geographical distribution, biology, impact and potential entry pathways of S. tridentata has been evaluated against the criteria for it to qualify as potential Union quarantine pest or as Union regulated non‐quarantine pest (RNQP). Since the pest is not reported in EU, it does not meet the criteria assessed by EFSA to qualify as potential Union RNQP. S. tridentata satisfies the criterion for quarantine pest regarding entry into the EU territory. Due to the scarcity of data, the Panel is unable to conclude if S. tridentata meets the post‐entry criteria of establishment, spread and potential impact.
Keywords: Elm borer, longhorn beetle, pest risk, plant health, plant pest, quarantine, host range uncertainty
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
1.1.1. Background
Council Directive 2000/29/EC1 on protective measures against the introduction into the Community of organisms harmful to plants or plant products and against their spread within the Community establishes the present European Union plant health regime. The Directive lays down the phytosanitary provisions and the control checks to be carried out at the place of origin on plants and plant products destined for the Union or to be moved within the Union. In the Directive's 2000/29/EC annexes, the list of harmful organisms (pests) whose introduction into or spread within the Union is prohibited, is detailed together with specific requirements for import or internal movement.
Following the evaluation of the plant health regime, the new basic plant health law, Regulation (EU) 2016/2031 on protective measures against pests of plants, was adopted on 26 October 2016 and will apply from 14 December 2019 onwards, repealing Directive 2000/29/EC. In line with the principles of the above mentioned legislation and the follow‐up work of the secondary legislation for the listing of EU regulated pests, EFSA is requested to provide pest categorisations of the harmful organisms included in the annexes of Directive 2000/29/EC, in the cases where recent pest risk assessment/pest categorisation is not available.
1.1.2. Terms of Reference
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide a scientific opinion in the field of plant health.
EFSA is requested to prepare and deliver a pest risk assessment (step 1 and step 2 analysis) for Saperda tridentata. The opinion should address all entry pathways, spread, establishment and risk reduction options. As explained in the background, please pay particular attention to the pathway of wood of Ulmus.
1.1.2.1. Background
The new Plant Health Regulation (EU) 2016/2031, on the protective measures against pests of plants, will be applying from 14 December 2019. Provisions within the above Regulation are in place for the listing of “high risk plants, plant products and other objects” (Article 42) on the basis of a preliminary assessment, and to be followed by a risk assessment. Commission Implementing Regulation (EU) 2018/2019 is establishing a provisional list of high risk plants, plant products or other objects, within the meaning of Article 42 of Regulation (EU) 2016/2031. The import of commodities included in the above mentioned list will be banned from 14 December 2019, awaiting the outcome of a risk assessment. Wood of Ulmus L. originating from third countries or areas of third countries where S. tridentata is known to occur, is included in this list and its introduction into the Union shall be provisionally prohibited.
1.2. Interpretation of the Terms of Reference
European Food Safety Authority (EFSA) is to prepare and deliver a two‐step pest risk assessment for S. tridentata Olivier (Coleoptera: Cerambycidae). The first step will be to subject S. tridentata to the process of pest categorisation to determine whether it fulfils the criteria, which are within the remit for EFSA to assess, for it to be regarded as a quarantine pest or of a regulated non‐quarantine pest for the area of the European Union (EU) excluding Ceuta, Melilla and the outermost regions of Member States referred to in Article 355(1) of the Treaty on the Functioning of the European Union (TFEU), other than Madeira and the Azores. The second step will require EFSA to conduct a more detailed assessment of all entry pathways, spread, establishment and risk reduction options. Particular attention will be given to the pathway of wood of Ulmus.
The new Plant Health Regulation (EU) 2016/2031,2 on the protective measures against pests of plants, will be applying from December 2019. The regulatory status sections (Section 3.3) of the present opinion are still based on Council Directive 2000/29/EC, as the document was adopted in November 2019.
2. Data and methodologies
2.1. Data
2.1.1. Literature search
A comprehensive literature search on S. tridentata was conducted at the beginning of the categorisation in the ISI Web of Science bibliographic database, using the scientific name and synonyms (Chapter 3.1.1) of the pest as search term. Relevant papers were reviewed, and further references and information were obtained from experts, as well as from citations within the references and grey literature.
Because of uncertainty around host plants, additional searches in the Biodiversity Heritage Library database (https://www.biodiversitylibrary.org/), and the JSTOR database (https://www.jstor.org/), were conducted using the scientific name of the pest and synonyms as a search term (more information in Appendix D).
2.1.2. Database search
Pest information, on host(s) and distribution, was retrieved from the European and Mediterranean Plant Protection Organization (EPPO) Global Database (EPPO, online) and relevant publications.
Data for the estimation of the import of elm wood were obtained from https://www.americanhardwood.org/ and Eurostats.
Data about the area of hosts grown in the EU were obtained from the European Atlas of Forest Tree Species (https://forest.jrc.ec.europa.eu/en/european-atlas/atlas-download-page/).
The Europhyt database was consulted for pest‐specific notifications on interceptions and outbreaks. Europhyt is a web‐based network run by the Directorate General for Health and Food Safety (DG SANTÉ) of the European Commission and is a subproject of PHYSAN (Phyto‐Sanitary Controls) specifically concerned with plant health information. The Europhyt database manages notifications of interceptions of plants or plant products that do not comply with EU legislation, as well as notifications of plant pests detected in the territory of the Member States (MS) and the phytosanitary measures taken to eradicate or avoid their spread.
2.2. Methodologies
The Panel performed the pest categorisation for S. tridentata, following guiding principles and steps presented in the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018) and in the International Standard for Phytosanitary Measures No 11 (FAO, 2014) and No 21 (FAO, 2004).
This work was initiated following an evaluation of potential high risk commodities to determine whether S. tridentata satisfied the criteria for it to qualify as a potential Union quarantine pest or a potential Union regulated non‐quarantine pest in accordance with Regulation (EU) 2016/2031 on protective measures against pests of plants, and includes additional information required in accordance with the specific terms of reference received by the European Commission. In addition, for each conclusion, the Panel provides a short description of its associated uncertainty.
Table 1 presents the Regulation (EU) 2016/2031 pest categorisation criteria on which the Panel bases its conclusions. All relevant criteria have to be met for the pest to potentially qualify either as a potential quarantine pest or as a potential regulated non‐quarantine pest. If one of the criteria is not met, the pest will not qualify. A pest that does not qualify as a quarantine pest may still qualify as a regulated non‐quarantine pest that needs to be addressed in the opinion.
Table 1.
Pest categorisation criteria under evaluation, as defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest | Criterion in Regulation (EU) 2016/2031 regarding Union regulated non‐quarantine pest |
|---|---|---|
| Identity of the pest (Section 3.1 ) | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? |
| Absence/presence of the pest in the EU territory (Section 3.2 ) |
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU? Describe the pest distribution briefly! |
Is the pest present in the EU territory? If not, it cannot be a regulated non‐quarantine pest. (A regulated non‐quarantine pest must be present in the risk assessment area). |
| Regulatory status (Section 3.3 ) | If the pest is present in the EU but not widely distributed in the risk assessment area, it should be under official control or expected to be under official control in the near future | Is the pest regulated as a quarantine pest? If currently regulated as a quarantine pest, are there grounds to consider its status could be revoked? |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4 ) | Is the pest able to enter into, become established in, and spread within, the EU territory? If yes, briefly list the pathways! |
Is spread mainly via specific plants for planting, rather than via natural spread or via movement of plant products or other objects? Clearly state if plants for planting is the main pathway! |
| Potential for consequences in the EU territory (Section 3.5 ) | Would the pests’ introduction have an economic or environmental impact on the EU territory? | Does the presence of the pest on plants for planting have an economic impact, as regards the intended use of those plants for planting? |
| Available measures (Section 3.6 ) | Are there measures available to prevent the entry into, establishment within or spread of the pest within the EU such that the risk becomes mitigated? | Are there measures available to prevent pest presence on plants for planting such that the risk becomes mitigated? |
| Conclusion of pest categorisation (Section 4 ) | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential quarantine pest were met and (2) if not, which one(s) were not met | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential regulated non‐quarantine pest were met, and (2) if not, which one(s) were not met |
It should be noted that the Panel's conclusions are formulated respecting its remit and particularly with regard to the principle of separation between risk assessment and risk management (EFSA founding regulation (EU) No 178/2002); therefore, instead of determining whether the pest is likely to have an unacceptable impact, the Panel will present a summary of the observed pest impacts. Economic impacts are expressed in terms of yield and quality losses and not in monetary terms; addressing social impacts is outside the remit of the Panel (Table 1).
3. Pest categorisation
3.1. Identity and biology of the pest
3.1.1. Identity and taxonomy
3.1.1.1.
Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? (Yes or No)
Yes, S. tridentata is established as a recognised species.
S. tridentata Olivier, 1795 is a coleopteran in the family Cerambycidae. Synonyms include Eutetrapha tridentata (Olivier, 1795), Compsidea tridentata (Olivier, 1795) and Saperda dubiosa (Haldeman, 1847). In North America, S. tridentata is known as the elm borer (Bosik, 1997) or the elm tree borer.
The genus Saperda Fabricius, 1775, consists of around 42 species. Felt and Joutel (1904) reported 16 species from North America, but Bezark (2016) revised the genus and suggests that there are 15 species of Saperda in North America.
3.1.2. Biology of the pest
Much of the literature on the biology of S. tridentata was published in the late 19th century, when S. tridentata was first recognised as a pest of American elms, and during the first half of the 20th century when it was thought to vector Ophiostoma ulmi, a causal agent of Dutch elm disease. The disease arrived in North America in the 1930s but was primarily spread by the native elm bark beetle, Hylurgopinus rufipes. Dutch elm disease killed millions of American elm trees and they are no longer a prominent feature in natural or urban landscapes (Allen and Humble, 2002). Consequent to the great loss of elms, and decrease in the supply for elm wood, almost no studies have been conducted on S. tridentata in recent decades. Much of the recent literature that mentions S. tridentata are simply records of its spatial expansion, e.g. westwards into Saskatchewan (Heffern, 1998; Bousquet et al., 2013), Colorado (Heffern, 1998) and Idaho (Rice et al., 2017). The following is therefore taken mainly from older literature describing the natural history of the species rather than from papers reporting experimental studies.
Oviposition takes place at night. Females chew a slit in the bark of branches of hosts or on logs of U. americana, U. rubra or U. crassifolia in which a single egg is oviposited. A female can chew slits quite close together (Pechuman, 1940; Drooz, 1985; Campbell et al., 1989). Females living for a month or so can lay 50–60 eggs (Pechuman, 1940). Branches that have recently died or are weakened, stressed or are otherwise low in vitality are preferred for oviposition (Baker, 1941). Host trees that have been recently felled are also favoured (Craighead, 1923). Tucker (1907) found no evidence that S. tridentata attacked healthy trees. Pechuman (1940) also suggested that healthy trees are not attacked.
Eggs hatch after a few days and larvae chew through the bark into the sapwood (Perkins, 1890). Unlike many other cerambycid species, larvae of S. tridentata do not burrow deeply but tend to remain between the sapwood and bark. As larvae grow, the tunnels they create meander in all directions within the sapwood and inner bark. There are at least three larval instars (Perkins, 1890). Feeding loosens the bark which can be peeled away easily (Perkins, 1890; Pechuman, 1940). When abundant, tunnels can girdle branches and the host's trunk (Hoffmann, 1942). Once established in a damaged or weakened branch of a host, larvae can move to healthy areas (Pechuman, 1940) although Baker (1941) reported that only occasionally were larvae found in live branches.
Larvae overwinter under the bark and in sapwood. In the spring, they bore a small distance into sapwood to form a chamber 5–6 mm into the wood. Here they develop into pupae. However, in unfavourable conditions, some remain as larvae for another year before pupae are formed the following spring.
After eclosure adults can remain in the pupal chamber for up to 7 days. They eventually exit either via the larval tunnel leading into the chamber or gnaw a new tunnel through the wood, leaving via roundish exit holes (Pechuman, 1940). Adults emerge in late spring and early summer (Campbell et al., 1989). Adults emerging in May develop from pupae that take on average 24–27 days to develop whereas adults emerging in June develop from pupae that develop in 15–18 days (Pechuman, 1940). Males emerge first. Adults feed on the mid‐rib and larger veins of host leaves, the surrounding leaf material, leaf petiole and the bark of young twigs (Craighead, 1923; Pechuman, 1940; Drooz, 1985). Adults can mate after 3 or 4 days of maturation feeding (Pechuman, 1940). Adults can fly and are most active at night; both males and females can be caught by light traps from May to August (Solomon et al., 1972; Gosling and Gosling, 1977). During the day adults shelter amongst foliage or bark (Pechuman, 1940). Adults live for 1 or 2 months.
There is usually one generation per year, but larvae hatching from eggs laid later in the summer e.g. during late July and August usually require an additional year to develop (Pechuman, 1940). When conditions are unfavourable for larval development, it may also take 2 years to complete development (Campbell et al., 1989). In wood that is dried out development may take 2 or 3 years (Pechuman, 1940).
3.1.2.1. Intraspecific diversity
Bousquet et al. (2013) list four subspecies of S. tridentata. However, no biologically relevant information could be found to justify considering them separately within this pest categorisation. The four are S. tridentata dubiosa Haldeman, 1847, S. tridentata rubronotata Fitch, 1858, S. tridentata intermedia Fitch, 1858 and S. tridentata trifasciata Casey, 1913.
3.1.3. Detection and identification of the pest
3.1.3.1.
Are detection and identification methods available for the pest?
Yes, light traps can capture adults. Juvenile stages can be detected by visual inspections. Traditional morphological keys are available to identify larvae and adults to species.
Detection
As a species that spends the majority of its life within its host, S. tridentata is not easy to detect until the host shows symptoms. Symptoms of S. tridentata larval infestation include the premature yellowing of leaves by a month or so (Tucker, 1907), thinning foliage, a few high branches dying before the rest of the crown, frass and sawdust on branches. When heavily infested, bark becomes loosened and can easily be peeled back to reveal larval tunnels. The inner bark can be heavily mined (Felt and Joutel, 1904). Careful inspection of trees can reveal roundish emergence holes 4–4.5 mm in diameter. Large holes in leaf tissue surrounding the larger veins and young twigs dangling by a strip of bark as a result of being chewed by adults (Pechuman, 1940) could indicate adults infesting an elm.
Identification
Traditional morphological keys can be used to identify adults and larvae. Craighead (1923) provides a species key to larvae in the genus Saperda. Packard (1890) provides a detailed description of the larvae. Craighead (1923) provides a description of the pupae. Felt and Joutel (1904) describe the adult. An image of the adult is provided in Drooz (1985).
Mature larvae are whitish legless and rather flattened, 12–25 mm long (Drooz, 1985; Solomon, 1995). Adults are 9–17 mm long; males have antennae almost as long as their body, the antennae of females are shorter. The body is grey with three orange‐yellow oblique bands on the elytra. There are twin black spots on each side of the pronotum and at the base of the elytra (Solomon, 1995).
3.2. Pest distribution
3.2.1. Pest distribution outside the EU
The genus Saperda occurs in temperate parts of the northern hemisphere (Felt and Joutel, 1904). S. tridentata is common in deciduous forests of eastern North America although it has been found as far west as Idaho (Rice et al., 2017). S. tridentata is not known to have spread outside North America. Figure 1 shows the known global distribution of S. tridentata.
Figure 1.
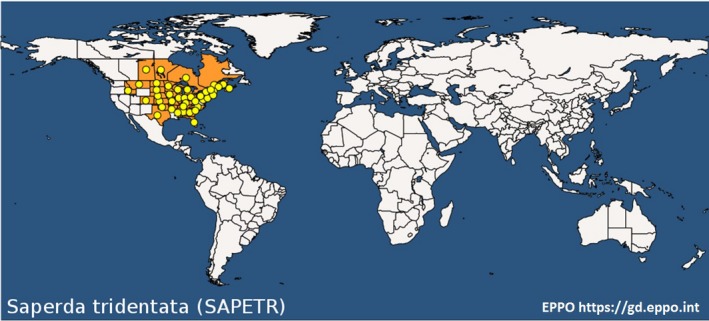
Global distribution of S. tridentata (last updated 7 November 2019). Extracted from the EPPO Global Database
3.2.2. Pest distribution in the EU
3.2.2.1.
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU?
No. S. tridentata is not known to be present in the EU.
3.3. Regulatory status
3.3.1. Council Directive 2000/29/EC
S. tridentata is not listed in Council Directive 2000/29/EC.
3.3.2. Legislation addressing the hosts of Saperda tridentata
The hosts range of S. tridentata is very uncertain (see Section 3.4.1). As a precaution, all legislation related to Ulmus species was included. Ulmus L. species comprise the major reported hosts of S. tridentata. Wood, cut branches and isolated bark of Ulmus davidiana Planch are controlled within Directive 2000/29/EC. Legislation relating to the genus Ulmus concerns the control of Agrilus planipennis (Annex IV A1, point 2.3, 2.4, 2.5), Stegophora ulmea, a pest listed in Annex II A I of the plant health directive 2000/29/EC, and Candidatus Phytoplasma ulmi (an Annex I A II pest). Tables 2 and 3 provide details of requirements pertaining to plants of Ulmus and Candidatus Phytoplasma ulmi).
Table 2.
Regulated hosts and commodities that may involve S. tridentata in Annex IV of Council Directive 2000/29/EC
|
Annex IV Part A |
Special requirements which must be laid down by all Member States for the introduction and movement of plants, plant products and other objects into and within all member states | |
| Section I | Plants, plant products and other objects originating outside the community | |
| Plants, plant products and other objects | Special requirements | |
| 2.3 | Whether or not listed among CN codes in Annex V, Part B, wood of […], Ulmus davidiana Planch. […] including wood which has not kept its natural round surface, and furniture and other objects made of untreated wood, originating in Canada, […] and USA |
Official statement that: (a) the wood originates in an area recognised as being free from Agrilus planipennis Fairmaire […], or (b) the bark and at least 2.5 cm of the outer sapwood are removed in a facility authorised and supervised by the national plant protection organisation, or (c) the wood has undergone ionizing irradiation to achieve a minimum absorbed dose of 1 kGy throughout the wood. |
| 2.4 | Whether or not listed among CN codes in Annex V, Part B, wood in the form of chips, particles, sawdust, shavings, wood waste and scrap obtained in whole or in part from […], Ulmus davidiana Planch. […] originating in Canada, […] USA | Official statement that the wood originates in an area recognised as being free from Agrilus planipennis Fairmaire […]. |
| 2.5 | Whether or not listed among CN codes in Annex V, Part B, isolated bark and objects made of bark of […] Ulmus davidiana Planch. […] originating in Canada, […] and USA | Official statement that the bark originates in an area recognised as being free from Agrilus planipennis Fairmaire […] |
| 14. | Plants of Ulmus L., intended for planting, other than seeds, originating in North American countries | Without prejudice to the provisions applicable to the plants in Annex IV (A)(I)(11.4), official statement that no symptoms of Candidatus Phytoplasma ulmi have been observed at the place of production or in its immediate vicinity since the beginning of the last complete cycle of vegetation. |
Table 3.
Regulated hosts and commodities that may involve S. tridentata in Annex V of Council Directive 2000/29/EC
| Annex V | Plants, plant products and other objects which must be subject to a plant health inspection ([…] — in the country of origin or the consignor country, if originating outside the Community) before being permitted to enter the community | |
| PART B | Plants, plant products and other objects originating in territories, other than those territories referred to in Part A | |
| Section 1 | Plants, plant products and other objects which are potential carriers of harmful organisms of relevance for the entire Community | |
| 6. (b) | Wood within the meaning of the first subparagraph of Article 2(2), where it meets one of the following descriptions laid down in Annex I, Part two to Council Regulation (EEC) No 2658/87: | |
| CN Code | Description | |
| ex 4403 99 | Non‐coniferous wood (other than tropical wood specified in subheading note 1 to Chapter 44 or other tropical wood, oak (Quercus spp.), beech (Fagus spp.) or birch (Betula L.)), in the rough, whether or not stripped of bark or sapwood, or roughly squared, not treated with paint, stains, creosote or other preservatives | |
| ex 4407 99 | Non‐coniferous wood (other than tropical wood specified in subheading note 1 to Chapter 44 or other tropical wood, oak (Quercus spp.), beech (Fagus spp.), maple (Acer spp.), cherry (Prunus spp.) or ash (Fraxinus spp.)), sawn or chipped lengthwise, sliced or peeled, whether or not planed, sanded or end‐jointed, of a thickness exceeding 6 mm | |
Commission Implementing Decision (UE) 2015/893 targeting Anoplophora glabripennis contains requirements to inspect plants and wood of A. glabripennis hosts which include Ulmus sp.
3.4. Entry, establishment and spread in the EU
3.4.1. Host range
Felt and Joutel (1904) report that S. tridentata feeds almost exclusively on white elm (Ulmus americana) although they recognise that red elm (U. rubra) can also be attacked. Solomon (1995) states that cedar elm (U. crassifolia) is also a host. MacRae (1993) reported finding adults of S. tridentata on Ulmus alata (winged elm, native to south central and south east USA). Whether or not U. alata is a host suitable for breeding is unknown.
The literature does not report species of European elm introduced into North America as hosts of S. tridentata (Table 4). Felt and Joutel (1904) state that there is no evidence that S. tridentata attacks European elms, U. glabra or U. minor. Campbell et al. (1989) cite Metcalf et al. (1951) when reporting that S. tridentata does not attack English elm (U. minor) or wych elm (U. glabra) two of the three most common elm species in Europe (Caudullo and de Rigo, 2016). The Swedish Unit for Risk Assessment of Plant Pests (Boberg and Björklund, 2018) drafted a short document citing Krischik and Davidson (2013) which reports ‘American elms, slippery elm and other elms’ as host plants. However ‘other elms’ are not identified and it is assumed that Krischik and Davidson (2013) were referring to North American species (listed as the first four in Table 4). No evidence was found that S. tridentata attacks the third European elm species (U. laevis). Hence, S. tridentata literature does not regard European Ulmus species as hosts. However, the literature is based on observational evidence. There have been no formal experiments, such as feeding choice studies, or oviposition choice experiments to categorically confirm that S. tridentata could, or particularly could not, develop on European elms.
Table 4.
Binomial and common names of key elm species referred to within this pest categorisation and notes related to their occurrence in Europe
| Ulmus species | Common name | Host statusa | Notes in relation to occurrence in Europe |
|---|---|---|---|
| U. americana |
American elm white elm water elm |
Major | This species has been introduced in Europe but does not grow well as it is more susceptible to insect foliage damage than native European elms and is susceptible to Dutch elm disease. It grows in European arboreta (Source: Botanic Gardens Conservation International database ‐ https://tools.bgci.org/global_tree_search.php), could be present in private gardens and parks as it is sold in horticultural trade. Abundance and density uncertain |
| U. crassifolia |
cedar elm Texas cedar elm |
Minor | Grows in European garden arboreta (Source: Botanic Gardens Conservation International database – https://tools.bgci.org/global_tree_search.php) |
| U. rubra |
slippery elm red elm |
Minor | Introduced to Europe in 1830 (White and Moore, 2003) |
| U. alata | Winged elm | Uncertain; minor host if a host at all | One of three American elm species known to be cultivated in UK as an ornamental in the early 1800s (Main, 1839), now rare. Could be present in private gardens and parks as it is sold in horticultural trade also as a bonsai. Abundance and density uncertain |
| U. glabra |
mountain elm wych elm Scots elm Scotch elm |
No evidence of being a host |
Wide range across most of Europe, from UK to Siberia, including Turkey. Previously widely planted as an ornamental in urban parks and along roadsides but no longer used as such due to its susceptibility to Dutch elm disease. |
| U. minor |
field elm smooth‐leaved English elm (in US?) |
No evidence of being a host |
Mainly in southern European regions by banks of small streams |
| U. laevis |
European white elm Russian elm |
No evidence of being a host |
Occurs across Central and Eastern Europe and is relatively rare. |
Major = most common host reported in literature. Minor = very few references in the literature.
Figure 2 illustrates the number of pieces of literature that name plants as hosts of S. tridentata. Out of 304 references about S. tridentata, 183 refer to association with a named host. 94% of these associations were with Ulmus sp. (elm, Ulmus and four American species). Two generic reports refer to Siberian elm (U. pumila) introduced into North America.
Figure 2.
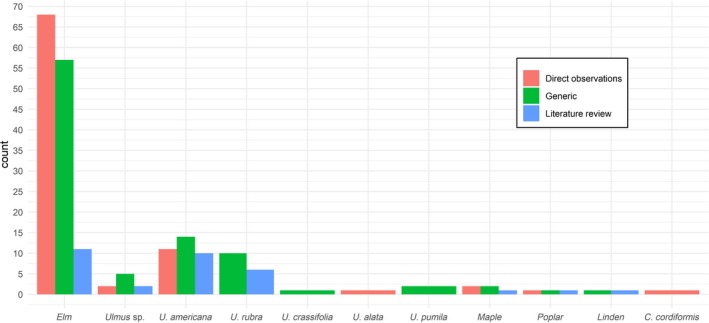
Number of pieces of literature that name plants as hosts of S. tridentata. Red: Count of literature with direct observations (e.g. documents reporting S. tridentata and its host(s) in open field or under laboratory conditions); Green: literature with generic observations (e.g. manuals and textbooks without supporting references); Blue: literature reviews. Names are quoted as found in the reference (Appendix D). Source: Biodiversity Heritage Library database (https://www.biodiversitylibrary.org/), the JSTOR database (https://www.jstor.org/), ISI Web of Knowledge. Last access to databases: 21 October 2019
However, five other broad‐leafed tree groups were mentioned in association with the pest, i.e. maple (Acer sp., n = 2), poplar (Populus sp., n = 1), linden (Tilia sp., n = 1) and bitternut hickory (Carya cordiformis, n = 1) which require particular attention.
Maple: A repeatedly cited observation originally by Harrington (1883) described that he was ”…stripping the bark from a large prostrate maple on May 22nd,… The tree yielded… some pupae of Saperda tridentata, Oliv., from which imagos emerged on June 15th’’. Felt and Joutel (1904) noted (without further reference) that there is a record of S. tridentata emerging from maple (i.e. Acer). They write that the ‘infestation must have been abnormal or else the record was founded on an erroneous observation’. Ehrmann (1897) mentions that on ‘June 3. S. tridentata is found in numbers on the Elm and Acer’ without further description e.g. of distance between the elm and maple specimens, or S. tridentata phenological stage (adults or larvae). Hence, the observation by Ehrmann does not constitute a reliable record that Acer is a host on which S. tridentata development takes place.
Poplar (Populus): The only reference is by Washburn (1910) who annexed the following sentence to a chapter on S. tridentata on elm: ‘We have reared this same borer, S. tridentata, from Poplars.’ Details about the wood offered, the rearing conditions and the successful development of imagos are lacking.
Linden (Tilia): Lugger (1899) recognised that ‘S. vestida Say (linden borer), S. tridentata Oliv., and S. lateralis Fab., occur upon a variety of forest trees, such as linden, poplar and others’. Unfortunately, it remains ambiguous which borer referred to which tree, and whether by ‘others’ he referred to elm.
Hickory (Carya cordiformis): The report of Park (1931) regarding relations of Coleopterans to plants for food and shelter mentions for a mesophytic oak–elm–hickory subclimax that ‘Saperda tridentata Oliv. has been beaten from the foliage of the bitter‐nut hickory in numbers on July I7, and this is in the seasonal range for the species as given by Blatchley (1910, p. 1087). It has been repeatedly beaten from elm foliage. The larvae bore into this tree (Felt 1906, I, pp. 67‐70)’. Recall that during the day adults shelter amongst foliage or bark (Pechuman, 1940). Hence, the observation by Park (1931) does not constitute a reliable record that hickory is a host on which S. tridentata development takes place.
The potential of S. tridentata to change host selection preference given host limitation was not reported in literature. Therefore, host range in other species of the genus Saperda was addressed for American and non‐American species. The reported hosts of the other 14 North American Saperda species were compiled into a table (Appendix B). Half of the 14 species are reported to feed on hosts in more than one family. Of the seven remaining species at least five feed on hosts from two or more host genera. Two of the 14 species are recorded as having a single genus as a host. Appendix C lists 22 Saperda species from outside North America and records the host plants where known. Most feed on multiple genera. Three species, S. octomaculata, S. scalaris and S. subobliterara, feed on Ulmus. In Asia, S. octomaculata and S. scalaris are reported as feeding on Ulmus and other unspecified deciduous trees. S. subobliterata is an Asian species only recorded feeding on Ulmus japonica and U. laciniata (Appendix C). Examining the hosts of other Saperda species, four out of five species feeding on Ulmus have hosts in other Families too.
3.4.2. Entry
3.4.2.1.
Is the pest able to enter the EU territory? (Yes or No) If yes, identify and list the pathways.
Yes, wood is a pathway, S. tridentata has been intercepted in the EU with U. rubra wood from USA on seven occasions since records began being centrally collected via Europhyt in 1995.
3.4.2.2. Interceptions
EUROPHYT data indicate that between 1995 and June 2019, there were six interceptions of S. tridentata (once in 2016, three times in 2017, once in 2018 and once in 2019) and one interception of Saperda sp. All were intercepted in Italy associated with U. rubra wood and bark from states close to the Great Lakes in USA (Ohio, Illinois, Iowa).
Prior to this pest categorisation, a 2016 interception report in EUROPHYT reported S. tridentata with a consignment of Ulmus rubra and of Juglans nigra roundwood (logs with bark) which led to J. nigra being referenced as a possible host in subsequent literature. A personal communication with the plant protection officer who conducted the original inspection has clarified that the larvae of S. tridentata were only found in U. rubra logs. The report in EUROPHYT has been clarified to prevent future misinterpretation. Consequently, the PLH Panel does not consider J. nigra as host of S. tridentata
Two records of Saperda sp. interceptions where no species is provided relate to interceptions of pallets and wood packing material from China to Germany. S. tridentata is not known to occur in China. We assume these interceptions were not S. tridentata but Asian Saperda species.
3.4.2.3. Pathways
S. tridentata could potentially enter the EU within different commodities comprising plant material including wood products (Table 5).
Table 5.
Traded wood and wood products that might comprise elm timber. Suggested pathways are listed with legislative measures which may result in S. tridentata detection during inspections required for other sanitary reasons
| CN Codea | Description | Regulatory measures |
|---|---|---|
| Solid wood packaging material (SWPM) if constructed using recently felled host timber | Managed by ISPM 15 (need to treat wood materials of a thickness greater than 6 mm, used to ship products between countries) | |
| ex 0602 | Plants for planting, other than seeds, in vitro material and naturally or artificially dwarfed woody plants for planting, originating from all third countries |
Included in Commission Implementing Regulation EU 2018/2019. Ulmus plants for planting from North America are inspected […](2000/29 EC, Annex IV A I 14.). |
| 4401 12 00 | Fuel wood, in logs, in billets, in twigs, in faggots or in similar forms. Non‐coniferous | Requires inspection (Annex V B) |
| ex 4401 22 00 | Wood in chips or particles. Non‐coniferous | Included in Commission Implementing Regulation EU 2018/2019 |
| ex 4401 39 00 | Sawdust and wood waste and scrap, agglomerated in logs, briquettes, pellets or similar forms. Others. | Included in Commission Implementing Regulation EU 2018/2019 |
| ex 4403 12 00 | Wood in the rough, whether or not stripped of bark or sapwood, or roughly squared. Treated with paint, stains, creosote or other preservatives. Non‐coniferous | Included in Commission Implementing Regulation EU 2018/2019 |
| ex 4403 99 00 | Wood in the rough, whether or not stripped of bark or sapwood, or roughly squared. Others. |
Included in Commission Implementing Regulation EU 2018/2019. Requires inspection (See 2000/29 EC Annex V B Section I 6 b). |
| 4404 20 00 | Hoopwood; split poles; piles, pickets and stakes of wood, pointed but not sawn lengthwise; wooden sticks, roughly trimmed but not turned, bent or otherwise worked, suitable for the manufacture of walking sticks, umbrellas, tool handles or the like; chipwood and the like. Non‐coniferous | Requires inspection (Annex V B) |
| 4406 12 00 | Railway or tramway sleeps (cross‐ties) of wood. Not impregnated. Non‐coniferous | Requires inspection (Annex V B) |
| ex 4407 99 | Wood sawn or chipped lengthwise, sliced or peeled, whether or not planed, sanded or end‐jointed, of a thickness exceeding 6 mm. Other |
Included in Commission Implementing Regulation EU 2018/2019. Requires inspection (See 2000/29 EC Annex V B Section I 6 b) |
| 4408 90 | Sheets for veneering (including those obtained by slicing laminated wood), for plywood or for similar laminated wood and other wood, sawn lengthwise, sliced or peeled, whether or not planed, sanded, spliced or end‐jointed, of a thickness not exceeding 6 mm. Other | No phytosanitary measures required. Highly processed. |
| 4409 29 | Wood (including strips and friezes for parquet flooring, not assembled) continuously shaped (tongued, grooved, rebated, chamfered, V‐jointed, beaded, moulded, rounded or the like) along any of its edges, ends or faces, whether or not planed, sanded or end‐jointed. Non‐coniferous. Other | No phytosanitary measures required. Highly processed |
| 4410 | Particle board, oriented strand board (OSB) and similar board (e.g. waferboard) of wood or other ligneous materials, whether or not agglomerated with resins or other organic binding substances | No phytosanitary measures required. Highly processed |
| 4411 | Fibreboard of wood or other ligneous materials, whether or not bonded with resins or other organic substances | No phytosanitary measures required. Highly processed |
| 4412 | Plywood, veneered panels and similar laminated wood | No phytosanitary measures required. Highly processed |
Chapter 44. Commission Implementing Regulation (EU) 2018/1602 of 11 October 2018 amending Annex I to Council Regulation (EEC) No 2658/87 on the tariff and statistical nomenclature and on the Common Customs Tariff. Official Journal of the European Union L273, 61, 31 October 2018. (https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=OJ:L:2018:273:FULL%26from=ENCN Code)
3.4.2.4. Elm wood imports
Elm wood is relatively scarce in trade and only a small amount of elm is exported from USA (Cassens, 2007). Nevertheless, rough‐wood logs are shipped from North America to Europe.
Elm wood, Ulmus spp., is not itemised in trade nomenclature. The international nomenclature and codification system used to record trade statistics are not sufficiently detailed to determine the amount of trade specifically in elm wood. Table 5 details import classifications sub‐summing elm wood and derived products. CN 4403 (wood in the rough, whether or not stripped of bark or sapwood, or roughly squared, not treated) recognises subclass CN 4403 99 00 for rough hardwood not itemised and thus would include elm; CN 4407 (Wood sawn or chipped lengthwise, sliced or peeled, whether or not planed, sanded or end‐jointed, of a thickness exceeding 6 mm.) recognises a sub‐class CN 4407 99 for all sawn non‐tropical hardwoods not itemised and thus would include elm.
3.4.2.5. Import volume
Eurostat statistics based on customs records of import do not explicitly itemise elm wood. Estimation thus combines secondary indicators. Luppold and Thomas (1991) estimated the volume of annual exports of individual hardwoods from USA to European countries for the years 1981–1989. The mean volume of elm hardwood was 311 m3 per year (minimum 24 m3; maximum 807 m3). The density of elm wood can be estimated by assuming 12% moisture and is estimated as 593 kg m−3 (source: http://www.americanhardwood.org). Hence, between 1981 and 1989, a mean of 185 tonnes of elm wood was imported annually into the EU (range 14–479 tonnes; Table 6). 75% of US elm exports to the EU went to Italy, 18% to France, 5% to UK and 2% to BENELUX (Luppold and Thomas, 1991) (Table 6).
Table 6.
Estimated annual volumes and weight of elm wood imported into EU from USA 1981–1989 (Source: Luppold and Thomas, 1991)
| Year | Volume (m3) | Weight (tonnes) |
|---|---|---|
| 1981 | 300 | 178 |
| 1982 | 404 | 240 |
| 1983 | 191 | 113 |
| 1984 | 175 | 104 |
| 1985 | 807 | 479 |
| 1986 | 24 | 14 |
| 1987 | 28 | 17 |
| 1988 | 300 | 78 |
| 1989 | 573 | 340 |
| Annual mean | 311 | 185 |
| Minimum | 24 | 14 |
| Maximum | 807 | 479 |
In more recent years, the import volume of two main wood categories sub‐summing elm wood (CN 4403 99 and 4407 99) is available and is reported by weight (Table 7).
Table 7.
Import of rough wood (CN 4403 99) and sawn wood (CN 4407 99) potentially containing Ulmus species, from USA 2014 to 2018, into EU members states previously known to import US elm wood, (tonnes). Source: Eurostat
| Code/wood type | Importer | 2014 | 2015 | Year 2016 | 2017 | 2018 |
|---|---|---|---|---|---|---|
| CN 4403 99 rough wood | Italy | 33,976 | 39,910 | 37,627 | 37,259 | 34,354 |
| France | 284 | 102 | 366 | 19 | 0 | |
| UK | 345 | 307 | 158 | 213 | 93 | |
| sum | 34,605 | 40,319 | 38,151 | 37,492 | 34,447 | |
| CN 4407 99 sawn wood | Italy | 30,091 | 22,992 | 21,198 | 19,787 | 18,835 |
| France | 24,205 | 22,963 | 24,205 | 7,472 | 9,162 | |
| UK | 777 | 731 | 791 | 409 | 485 | |
| sum | 55,072 | 46,686 | 46,194 | 27,667 | 28,481 | |
| CN 4403 99 + CN 4407 99 | 89,677 | 87,005 | 84,345 | 65,159 | 62,928 |
The two data sources (Tables 6 and 7) were combined using earlier available Eurostat data from 1988. In 1988, a combined total of 148,332 tonnes of ‘rough wood, other’ (CN 4403 99) and ‘sawn wood, other’ (CN 4407 99) were imported into Italy, France and UK from USA. In 1989, the figure was 156,245 tonnes. According to Table 6 (Luppold and Thomas, 1991), approximately 178 tonnes of elm wood was imported into the EU (Italy, France and UK) in 1988 and 340 tonnes in 1989. Assuming all elm wood from USA was classified as either CN 4403 99 or CN 4407 99 then in 1988 0.12% (178/148,332) of rough and sawn wood ‘other’ entering the EU from USA was elm. Similarly, in 1989, approximately 0.22% (340/156,245) may have been elm.
Applying these estimates of 0.12% and 0.22% to the recent imports (Table 8) would suggest that between 63 and almost 180 tonnes of elm wood arrived in the EU each year between 2014 and 2018 from the USA, although the trend is downwards (Table 8).
Table 8.
Estimated annual amount of elm wood in tonnes imported from USA into the EU (Italy, France, UK), 2014–2018
| Elm wood | Year | ||||
|---|---|---|---|---|---|
| 2014 | 2015 | 2016 | 2017 | 2018 | |
| Combined total of rough wood and sawn wood ‘other’ | 89,677 | 87,005 | 84,345 | 65,159 | 62,928 |
| Elm estimate assuming 0.12% | 90 | 87 | 84 | 65 | 63 |
| Elm estimate assuming 0.22% | 179 | 174 | 169 | 130 | 126 |
EU member states were approached by EFSA and asked for information on the amount of elm logs imported annually from North America since 2016 (more information in Appendix F). Results are shown in Table 9.
Table 9.
Annual weight in tonnes of elm logs imported into EU member states from North America 2016–2019. (Note that 2019 is not a full year). (Source: replies to EFSA from individual member states, October 2019). Data were rounded to the nearest whole tonne. In some cases, weight was derived assuming a reference density (see Appendix F)
| EU member state | 2016 | 2017 | 2018 | 2019 |
|---|---|---|---|---|
| Italy | 133 | 113 | 41 | 143 |
| Germany | 38 | 62 | 31 | 26 |
| Portugal | 0 | 0 | 5 | 0 |
| Croatia | 0 | 0 | 0 | 0 |
| Cyprus | 0 | 0 | 0 | 0 |
| Estonia | 0 | 0 | 0 | 0 |
| Finland | 0 | 0 | 0 | 0 |
| France | 0 | 0 | 0 | 0 |
| Latvia | 0 | 0 | 0 | 0 |
| Lithuania | 0 | 0 | 0 | 0 |
| Malta | 0 | 0 | 0 | 0 |
| Poland | 0 | 0 | 0 | 0 |
| Spain | 0 | 0 | 0 | 0 |
| Sweden | 0 | 0 | 0 | 0 |
| The Netherlands | 0 | 0 | 0 | 0 |
| UK | 0 | 0 | 0 | 0 |
| Sum | 171 | 175 | 76 | 169 |
| Denmarka | 98 | 7,827 | 32 | 2 |
Data for Denmark refers to any type of wood of Ulmus, not only logs.
From replies received from individual member states, annual imports of elm logs into the EU from North America ranged from 76 to 175 tonnes (Table 9). These figures are similar to the estimates made in Table 8 suggesting between 63 and almost 180 tonnes of elm wood arrived in the EU each year between 2014 and 2018.
Denmark was unable to provide data on the tonnage of logs of Ulmus imported but was able to provide data on the imported volume (m3) including all types of wood of Ulmus which are under phytosanitary import provisions and have been inspected on import.
3.4.3. Establishment
3.4.3.1.
Is the pest able to become established in the EU territory? (Yes or No)
If European elms and/or maple and poplar species are hosts of S. tridentata, establishment is possible.
3.4.3.2. EU distribution of main host plants
Within Europe, there are limited number of arboreta where American species of elm such as U. americana and U. rubra grow (Table 10). Data of individual stand distributions are not accessible. Arboreta are assumed to be isolated and widely spread out (i.e. distances between arboreta exceed the range of adult flight capacity). There may be other American elms in private gardens and parks, but abundance and density are not known (see e.g. https://www.gbif.org/species/5361872). It is suggested that S. tridentata is unlikely to be able to establish if American elms are the only hosts. European elm species are more abundant but not known to be hosts. However, it has also not been proven that European elms are not hosts. The argument is valid also with Acer and Populus. If European elms, Acer and Populus are genuine hosts (see Section 3.4.1) and S. tridentata demonstrated a shift in host preference to European elms, Acer (maple) and Populus (poplar) in the EU, then hosts would be ubiquitous, and conditions would be conducive to establishment.
Table 10.
Number of gardens/arboreta in EU MS that are recorded as having North American elm species. Whether sites have only single specimens or multiple plantings is not known (Source: https://tools.bgci.org/global_tree_search.php)
| Country | U. americana | U. rubra | U. crassifolia | U. thomasii | U. serotina | U. alata | Sum |
|---|---|---|---|---|---|---|---|
| United Kingdom | 5 | 3 | 2 | 2 | 3 | 3 | 18 |
| Germany | 7 | 3 | 10 | ||||
| Belgium | 2 | 1 | 1 | 4 | |||
| Poland | 1 | 1 | 1 | 1 | 4 | ||
| Finland | 2 | 1 | 3 | ||||
| France | 2 | 1 | 3 | ||||
| Sweden | 1 | 2 | 3 | ||||
| Czech Republic | 1 | 1 | 2 | ||||
| Latvia | 1 | 1 | 2 | ||||
| Spain | 2 | 2 | |||||
| Denmark | 1 | 1 | |||||
| Estonia | 1 | 1 | |||||
| Sum | 26 | 10 | 4 | 5 | 4 | 4 | 53 |
3.4.3.3. Climatic conditions affecting establishment
The distribution and abundance of an organism that cannot control or regulate its body temperature are largely determined by host distribution and climate.
Comparing climates from the known distribution of an organism with climates in the risk assessment area can inform judgements regarding the potential distribution and abundance of an organism in the risk assessment area (Sutherst and Maywald, 1985; Ehrlén and Morris, 2015). The global Köppen–Geiger climate zone categories, and subsequent modifications made by Trewartha and Horn (1980), describe terrestrial climate in terms of factors such as average minimum winter temperatures and summer maxima, amount of precipitation and seasonality (rainfall pattern) (Trewartha and Horn, 1980; Kottek et al., 2006) and can inform judgements of aspects of establishment during pest categorisation (MacLeod and Korycinska, 2019).
Climate types found in regions of North America where S. tridentata occurs are also found in Europe (Figures 1 and 3) suggesting that climate would support the establishment of S. tridentata in large parts of the EU.
Figure 3.
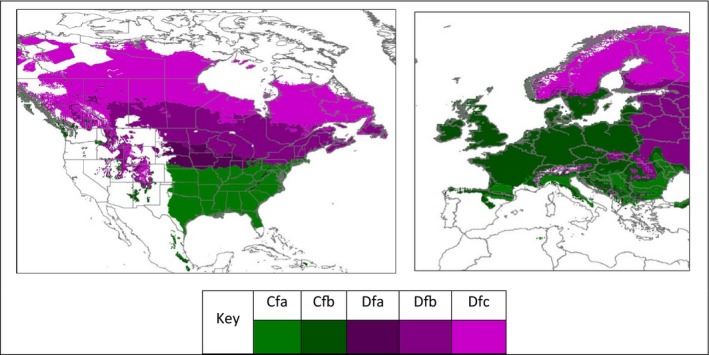
Occurrence of Köppen–Geiger climates (Trewartha and Horn, 1980) in North America (left pane) within which S. tridentata occurs and the same climates in Europe (right pane). The pane at the bottom shows the key scheme colour used for the different climates: Cfa = Humid subtropical climate, Cfb = Temperate oceanic climate, Dfa = Hot summer humid continental climate, Dfb = Warm summer humid continental climate, Dfc = Subarctic climate
3.4.4. Spread
3.4.4.1.
Is the pest able to spread within the EU territory following establishment?
Yes. If S. tridentata did establish it could spread. The species is a free‐living organism, adults can fly.
RNQPs: Is spread mainly via specific plants for planting, rather than via natural spread or via movement of plant products or other objects?
No. S. tridentata would not mainly spread via plants for planting.
Adults fly during the night. They tend not to disperse far if hosts are available locally (Hanks, 1999). The dispersal of S. tridentata, like that of other cerambycids such as S. candida and S. inornata, is influenced by the dietary requirements of adults and host requirements of the larvae. Females may oviposit on their natal host if its condition has not declined too greatly; such behaviour results in adults appearing to be relatively sedentary with a disinclination to disperse (Hanks, 1999).
S. tridentata has slowly spread within the USA and Canada. The species was known to occur in the eastern states of the USA and in eastern Canada in the late 1790s and was recorded in Idaho in 2017 (Rice et al., 2017). Whether recorded spread is related to recording effort, natural spread or spread on host plant material, such as infested Ulmus wood, is unknown.
3.5. Impacts
3.5.1.
Would the pests’ introduction have an economic or environmental impact on the EU territory?
The preference for weakened trees might generally limit the potential impact.
No, if S. tridentata does not feed on European species of elm impacts would be limited to isolated species of American elm growing in Europe, assuming S. tridentata could locate them.
Yes, if weakened trees of elms, poplars, or maples are hosts for S. tridentata there is likely to be an impact in the EU territory.
RNQPs: Does the presence of the pest on plants for planting have an economic impact, as regards the intended use of those plants for planting?
Yes. The occurrence of S. tridentata on plants for planting could have an economic impact on the intended use of those plants. Infested plants would be introducing a potentially serious pest that could spread and kill other hosts close by. However, the stem size of such plants for planting may limit final development of the pest to the adult stage.
As the pest is not present in the EU impact estimates are based on reports for North America.
S. tridentata prefers trees suffering a lack of nutrients or moisture (Campbell et al., 1989) or are otherwise stressed or weakened. Tucker (1907) reported that there was no evidence that adults oviposit in healthy trees. Hanks (1999) considers S. tridentata to be a species that specialises in attacking host plants whose defences have been compromised in some way, such as by chronically poor growing conditions or pathogen infestation. Urban habitats (typical for elm) might make trees more prone to attack by S. tridentata. The impact of S. tridentata could be to speed up the decline of a weakened tree that was already deteriorating.
Environmental impacts would be confined to already weakened or stressed individual hosts that were located and attacked. Whilst adult feeding damages leaves and twigs, it is the feeding and tunnelling of larvae within the cambium and phloem, inner bark and sapwood, of already weakened or stressed hosts that causes most damage. Infested trees tend to die slowly, a branch at a time (Drooz, 1985) although all parts can be attacked from small branches to the main trunk. The upper branches are usually affected first. Subsequent generations will work down an infected tree. Larval mines can girdle branches killing them; when abundant larval tunnelling can girdle the trunk causing the death of the tree (Hoffmann, 1942; Krischik and Davidson, 2013). Trees can be killed after 2 or 3 years (Packard, 1890). Roots are not attacked (Packard, 1890). Rows of elms in urban areas would gradually be attacked as subsequent generations of adult S. tridentata spread from tree to tree.
S. tridentata is more injurious in USA than in Canada (Campbell et al., 1989) perhaps because larvae develop more slowly further north in cooler climates.
Elm is a valued component of the urban forest as a shade provider although Dutch elm disease has greatly reduced the population of elms both in North America and Europe (Allen and Humble, 2002). Historically elm wood was used for barrel hoops and staves, boxes, crates, curved wooden parts of furniture, and panelling, more recently, in areas where the amount of elm wood available has declined (due to Dutch elm disease), it is used for pallets and solid wood packing material (Cassens, 2007). Elm wood is also used to make hockey sticks, veneer, wood pulp and in papermaking (https://www.wood-database.com/american-elm/). When sawn, elm is mainly used to make planks 25.4 mm thick.
Extrapolating information on consequences from the native range of the S. tridentata, a potential introduction of the organism into the EU territory likely is limited to weakened trees. If S. tridentata does neither feed on European species of elm nor on Acer or Populus impacts would be limited to isolated species of American elm growing in EU, assuming S. tridentata could locate them. If, however, S. tridentata feed on trees of elms, poplars or maples, there is likely to be an environmental and/or economic impact in the EU territory. Scarce evidence on precise host range of S. tridentata in the EU territory does not support any of the impact scenarios over the other.
3.6. Availability and limits of mitigation measures
3.6.1.
Are there measures available to prevent the entry into, establishment within or spread of the pest within the EU such that the risk becomes mitigated?
Yes, measures used for Agrilus planipennis could be appropriate (Table 2) and those in 3.6.1. However, parameters of mechanical treatment (i.e. removal of 2.5 cm in sapwood) need to be confirmed.
RNQPs: Are there measures available to prevent pest presence on plants for planting such that the risk becomes mitigated?
Yes, plant from planting from pest free areas.
3.6.2. Identification of additional measures
Phytosanitary measures are currently applied to Ulmus plants for planting from North America for the control of elm phloem necrosis (see Section 3.3).
Rough wood and sawn wood consignments require inspection (see Section 3.3.2). The existing general requirements are not specific to S. tridentata but inhibit the entry of many pests.
3.6.2.1. Additional control measures
Potential additional control measures are listed in Table 11.
Table 11.
Selected control measures (a full list is available in EFSA PLH Panel et al., 2018) for pest entry/establishment/spread/impact in relation to currently unregulated hosts and pathways. Control measures are measures that have a direct effect on pest abundance
| Information sheet title (with hyperlink to information sheet if available) | Control measure summary | Risk component (entry/establishment/spread/impact) |
|---|---|---|
| https://zenodo.org/record/1311026#.XchtZ-eTLVo |
Use of chemical compounds that may be applied to plants or to plant products after harvest, during process or packaging operations and storage. The treatments addressed in this information sheet are: a) fumigation; b) spraying/dipping pesticides; c) surface disinfectants; d) process additives; e) protective compounds |
Entry. Fumigants available. In principle applicable but challenging to put into practice for large timbers. |
| https://zenodo.org/record/1311058#.XchuAueTLVo | This information sheet deals with the following categories of physical treatments: irradiation/ionisation; mechanical cleaning (brushing, washing); sorting and grading, and removal of plant parts (e.g. debarking wood). This information sheet does not address: heat and cold treatment (information sheet 1.14); roughing and pruning (information sheet 1.12) |
Entry. Rough wood should be bark free rather than debarked to reduce the likelihood of entry |
| Heat and cold treatments | Controlled temperature treatments aimed to kill or inactivate pests without causing any unacceptable prejudice to the treated material itself. The measures addressed in this information sheet are: autoclaving; steam; hot water; hot air; cold treatment |
Entry. Equipment available. For large timbers complex in routine practice. |
| Roguing and pruning of infested brunches | If detected sufficiently early, pruning and burning of infested branches could remove infestations within individual trees | Establishment, Spread. |
| Felling and burning of infested trees | Felling and burning of infested trees to reduce the spread to neighbouring hosts. | Spread. |
| Insecticides targeting adults | Insecticides targeting adults could be applied to foliage; insecticides with long‐lasting residual activity could be applied to trunks | Spread. Reduce establishment. |
3.6.2.2. Additional supporting measures
Potential additional supporting measures are listed in Table 12.
Table 12.
Selected supporting measures (a full list is available in EFSA PLH Panel, 2018) in relation to currently unregulated hosts and pathways. Supporting measures are organisational measures or procedures supporting the choice of appropriate risk reduction options that do not directly affect pest abundance
| Information sheet title (with hyperlink to information sheet if available) | Supporting measure summary | Risk component (entry/establishment/spread/impact) |
|---|---|---|
| https://zenodo.org/record/1311135#.XchvGueTLVo |
Inspection is defined as the official visual examination of plants, plant products or other regulated articles to determine if pests are present or to determine compliance with phytosanitary regulations (ISPM 5). The effectiveness of sampling and subsequent inspection to detect pests may be enhanced by including trapping and luring techniques. |
Light traps at ports of entry and inland at sites for wood distribution could be used to monitor for adults emerging from imported wood. Establishment |
| Encourage tree health and vigour | S. tridentata attacks weakened trees therefore practices that encourage tree health and vigour such as site selection, mulching and watering. | Establishment, spread, impact. |
3.7. Uncertainty
The precise hosts of S. tridentata in the EU are unknown and judgements about the expected establishment and/or spread come with high uncertainty. These critical uncertainties are not expected to lessen by conducting risk assessment.
Host diversity and plasticity of S. tridentata are unknown due to inconclusive anecdotal evidence and lack of experimental studies. Indicative reports on diverse species, other than North American elms, are isolated, mostly inconclusive/speculative in biological relevance, and not representative of any situation in the EU. Categorisation of the potential to establish within EU is highly uncertain and driven by assumptions regarding hosts.
Assuming European elms could be hosts of S. tridentata in the EU, still the occurrence, density and distribution of European elms in Europe is uncertain. The categorisation of capability to spread is highly uncertain as to whether there are enough hosts at local reach to enable the establishment of S. tridentata (i.e. perpetuate for the foreseeable future).
4. Conclusions
Table 13 provides a summary of the conclusions of each part of this pest categorisation.
Table 13.
The Panel's conclusions on the pest categorisation criteria defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Panel's conclusions against criteria in Regulation (EU) 2016/2031 regarding Union quarantine pest | Key uncertainties |
|---|---|---|
| Identity of the pests (Section 3.1 ) | S. tridentata Olivier, (Coleoptera: Cerambycidae) is a well‐established and recognised pest | None |
| Absence/presence of the pest in the EU territory (Section 3.2 ) | S. tridentata is not known to occur in the EU. The pest occurs in eastern North America (USA and Canada) | A few interceptions have been recorded |
| Regulatory status (Section 3.3 ) | S. tridentata is not under official control | |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4 ) |
Entry is possible (proven by interceptions). If European elms and/or maple and poplar species are hosts of S. tridentata, then establishment is possible. Spread is possible as S. tridentata is a free‐living organism and adults can fly |
High uncertainty level. No experimental data; only anecdotal evidence; inadequate information on the precise host range in the EU. |
| Potential for consequences in the EU territory (Section 3.5 ) | If S. tridentata does not feed on European species of elm, impacts would be limited to isolated species of American elm growing in Europe, assuming S. tridentata could locate them. The preference for weakened trees might generally limit the potential impact. If European elms, poplars or maples are hosts for S. tridentata, there is likely to be an impact in the EU territory | High uncertainty level. No experimental evidence (e.g. choice experiments) to anticipate behaviour when known hosts (American elms) are scarce. |
| Available measures (Section 3.6 ) | Measures for wood‐inhabiting beetles with similar biology are available | Efficacy of measures unknown against S. tridentata |
|
Conclusion on pest categorisation (Section 4 ) |
Information on geographical distribution, biology, epidemiology, impact and potential entry pathways of S. tridentata has been evaluated against the criteria for it to qualify as potential Union quarantine pest or as Union regulated non‐quarantine pest (RNQP). Since the pest is not reported in EU or plants for planting are not the principal means of spread, it does not meet the criteria assessed by EFSA to qualify as potential Union regulated non‐quarantine pest. S. tridentata satisfies the criterion regarding entry into the EU territory. Due to the scarcity of data, the Panel is unable to conclude if S. tridentata meets the post‐entry criteria of establishment, spread and potential impact | Uncertainty regarding the precise host range and behaviour in the EU cannot be reduced by any subsequent risk assessment without new experimental evidence in these regards |
| Aspects of the assessment to focus on/scenarios to address in the future if appropriate | As it is, there is little reason to believe there are hosts on the European continent except those American elms in arboreta, but a host shift cannot be excluded as a possibility. A firm conclusion on all criteria of the categorisation can be achieved through gathering conclusive experimental evidence about the precise host range and behaviour in the EU | |
Glossary
- Containment (of a pest)
Application of phytosanitary measures in and around an infested area to prevent spread of a pest (FAO, 1995, 2017)
- Control (of a pest)
Suppression, containment or eradication of a pest population (FAO, 1995, 2017)
- Entry (of a pest)
Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2017)
- Eradication (of a pest)
Application of phytosanitary measures to eliminate a pest from an area (FAO, 2017)
- Establishment (of a pest)
Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2017)
- Impact (of a pest)
The impact of the pest on the crop output and quality and on the environment in the occupied spatial units
- Introduction (of a pest)
The entry of a pest resulting in its establishment (FAO, 2017)
- Measures
Control (of a pest) is defined in ISPM 5 (FAO 2017) as “Suppression, containment or eradication of a pest population” (FAO, 1995). Control measures are measures that have a direct effect on pest abundance. Supporting measures are organisational measures or procedures supporting the choice of appropriate Risk Reduction Options that do not directly affect pest abundance.
- Pathway
Any means that allows the entry or spread of a pest (FAO, 2017)
- Phytosanitary measures
Any legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2017)
- Protected zones (PZ)
A Protected zone is an area recognised at EU level to be free from a harmful organism, which is established in one or more other parts of the Union.
- Quarantine pest
A pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2017)
- Regulated non‐quarantine pest
A non‐quarantine pest whose presence in plants for planting affects the intended use of those plants with an economically unacceptable impact and which is therefore regulated within the territory of the importing contracting party (FAO, 2017)
- Risk reduction option (RRO)
A measure acting on pest introduction and/or pest spread and/or the magnitude of the biological impact of the pest should the pest be present. A RRO may become a phytosanitary measure, action or procedure according to the decision of the risk manager
- Spread (of a pest)
Expansion of the geographical distribution of a pest within an area (FAO, 2017)
Abbreviations
- EPPO
European and Mediterranean Plant Protection Organization
- FAO
Food and Agriculture Organization
- IPPC
International Plant Protection Convention
- ISPM
International Standards for Phytosanitary Measures
- MS
Member State
- PLH
EFSA Panel on Plant Health
- PZ
Protected Zone
- RNQP
regulated non‐quarantine pest
- TFEU
Treaty on the Functioning of the European Union
- ToR
Terms of Reference
Appendices to the Pest Categorisation on Saperda tridentata
To further inform decision‐making regarding the quarantine status of S. tridentata in the EU, the EFSA PLH Panel collected accessible information and observational knowledge supporting conclusions. The outcome is collated within the Appendices (Appendices A–F). The extra information and data were thought useful given the uncertainties about the host range of S. tridentata in EU. Nevertheless, interpretation of the data and information requires caution due to the non‐systematic collection, patchy accessibility and observational character (data assembled for different purposes).
List of Appendices:
Appendix A – Detailed Saperda tridentata global distribution
Appendix B – Host plants of North American Saperda species, other than S. tridentata
Appendix C – Host plants of non‐North American Saperda species
Appendix D – Literature search and review on S. tridentata hosts
Appendix E – Scoping review on the flight capacity of adult Saperda tridentata and other Cerambycidae, and analysis of spatial separation of European elm trees in Northern Italy
Appendix F – Ulmus logs imports in Europe.
Appendix A – Detailed Saperda tridentata global distribution
1.
Table A.1.
S. tridentata global distribution according to the online EPPO Global database (https://gd.eppo.int/). Last access: 7 November 2019
| Continent | Country | Sub‐national distribution | Status |
|---|---|---|---|
| North America | Canada | Present, restricted distribution | |
| Manitoba | Present, no details | ||
| New Brunswick | Present, no details | ||
| Nova Scotia | Present, no details | ||
| Ontario | Present, no details | ||
| Québec | Present, no details | ||
| Saskatchewan | Present, no details | ||
| USA | Present, restricted distribution | ||
| Alabama | Present, no details | ||
| Arkansas | Present, no details | ||
| Colorado | Present, no details | ||
| Connecticut | Present, no details | ||
| Florida | Present, restricted distribution | ||
| Georgia | Present, no details | ||
| Idaho | Present, no details | ||
| Illinois | Present, no details | ||
| Indiana | Present, no details | ||
| Iowa | Present, no details | ||
| Kansas | Present, no details | ||
| Kentucky | Present, no details | ||
| Maine | Present, no details | ||
| Maryland | Present, no details | ||
| Massachusetts | Present, no details | ||
| Michigan | Present, no details | ||
| Minnesota | Present, no details | ||
| Mississippi | Present, no details | ||
| Missouri | Present, no details | ||
| Montana | Present, restricted distribution | ||
| Nebraska | Present, no details | ||
| New Hampshire | Present, no details | ||
| New Jersey | Present, no details | ||
| New York | Present, no details | ||
| North Carolina | Present, no details | ||
| North Dakota | Present, no details | ||
| Ohio | Present, no details | ||
| Oklahoma | Present, no details | ||
| Pennsylvania | Present, no details | ||
| Rhode Island | Present, no details | ||
| South Carolina | Present, no details | ||
| South Dakota | Present, no details | ||
| Tennessee | Present, no details | ||
| Texas | Present, no details | ||
| Vermont | Present, no details | ||
| West Virginia | Present, no details | ||
| Wisconsin | Present, no details |
Appendix B – Host plants of North American Saperda species, other than S. tridentata
1.
Table B.1 includes a list of host plants of North American Saperda species, other than S. tridentata based on Felt and Joutel (1904), Zasada and Phipps (1990), Nord et al. (1972), and the http://titan.gbif.fr website.
Table B.1.
List of host plants of North American Saperda species, other than S. tridentata (Sources: a = Felt and Joutel, 1904; b = http://titan.gbif.fr/sel_plantes1.php?numplantes=7140; c = Zasada and Phipps, 1990; d = Nord et al., 1972)
| HOST FAMILY Genus species | North American Saperda spp. | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. imitans | S. discoidea | S. lateralis | S. vestita | S. candida | S. puncticolis | S. moesta | S. cretata | S. fayi | S. calcarata | S. obliqua | S. inornata | S. horni | S. mutica | |
| ACERACEAE | ||||||||||||||
| Acer sp. | b | |||||||||||||
| ANACARDIACEAE | ||||||||||||||
| Rhus radicans | a | |||||||||||||
| Rhus toxicodendron | a | |||||||||||||
| Toxicodendron radicans | a | |||||||||||||
| BETULACEAE | ||||||||||||||
| Alnus serrulata | a | |||||||||||||
| Betula sp. | b | a | ||||||||||||
| BURCERACEAE | ||||||||||||||
| Cammiphora opobalsamum | a | |||||||||||||
| CARYOCARACEAE | ||||||||||||||
| Caryocar sp. | a | |||||||||||||
| CORNACEAE | ||||||||||||||
| Cornus sp. | a | |||||||||||||
| JUGLANDACEAE | ||||||||||||||
| Carya amara | a | |||||||||||||
| Carya cordiformis | a | a | ||||||||||||
| Carya glabra | a | a | ||||||||||||
| Carya ovata | a | |||||||||||||
| Juglans nigra | a | |||||||||||||
| ROSACEAE | ||||||||||||||
| Amelanchier alnifolia | a | |||||||||||||
| Amelanchier arborea | a | |||||||||||||
| Amelanchier canadensis | a | |||||||||||||
| Aronia sp. | a | |||||||||||||
| Cotoneaster sp. | a | |||||||||||||
| Crataegus crus‐galli | a | |||||||||||||
| Crataegus oxyacantha | a | |||||||||||||
| Crataegus phaenopyrum | a | |||||||||||||
| Crataegus sp. | a | a | a | a | ||||||||||
| Crataegus tomentosa | a | |||||||||||||
| Cydonia oblonga | a | |||||||||||||
| Malus sp | a | a | a | |||||||||||
| Prunus avium | a | |||||||||||||
| Prunus domestica | a | |||||||||||||
| Prunus sp. | a | |||||||||||||
| Pyracantha sp. | a | |||||||||||||
| Pyrus communis | a | |||||||||||||
| Sorbus americana | a | |||||||||||||
| SALICACEAE | ||||||||||||||
| Populus balsamifera | a | c | a | |||||||||||
| Populus deltoides | a | |||||||||||||
| Populus grandidentata | a | |||||||||||||
| Populus nigra | a | |||||||||||||
| Populus sp. | a | a | ||||||||||||
| Populus tremuloides | a | d | ||||||||||||
| Salix lasiolepis | a | |||||||||||||
| Salix scouleriana | a | |||||||||||||
| Salix bebbiana | a | |||||||||||||
| Salix concolor | a | |||||||||||||
| Salix discolor | a | |||||||||||||
| Salix humilis | a | |||||||||||||
| Salix interior | a | |||||||||||||
| Salix petiolaris | a | |||||||||||||
| Salix sp. | a | a | a | a | a | a | ||||||||
| TILIACEAE | ||||||||||||||
| Tilia americana | a | |||||||||||||
| Tilia sp. | a | |||||||||||||
| ULMACEAE | ||||||||||||||
| Ulmus sp. | a | |||||||||||||
| Ulmus rubra | a | |||||||||||||
| VITACEAE | ||||||||||||||
| Parthenocissus engelmannii | a | |||||||||||||
| Parthenocissus quinquefolia | a | |||||||||||||
| Vitis sp. | a | |||||||||||||
| Number of host families | 5 | 4 | 3 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Number of host genera | 5 | 5 | 3 | 3 | 11 | 4 | 3 | 3 | 2 | 2 | 2 | 2 | 1 | 1 |
References
Felt EM, and Joutel L, 1904. “Monograph of the genus Saperda,” New York State Education Dept., Albany.
Nord JC, Grimble DG, and Knight FB, 1972. Biology of Saperda inornata (Coleoptera: Cerambycidae) in Trembling Aspen, Populus Iremuloides1. Annals of the Entomological Society of America 65, 127–135.
Zasada JC, and HPhipps HM, 1990. Populus balsamifera L. ‐ Balsam Poplar. In “Silvics of North America ‐ Volume 2, Hardwoods” (R. M. Burns and B. H. Honkala, eds.), pp. 518–529. Forest Service, United Service Department of Agriculture, Washington, DC.
Appendix C – Host plants of non‐North American Saperda species
1.
Table C.1 includes a list of host plants of non‐North American Saperda species.
Table C.1.
List of host plants of non‐North American Saperda species
| Saperda species | Distribution | Hosts | References |
|---|---|---|---|
| S. alberti | Asia (Japan, North Korea, China, Mongolia, Russia) |
Salix Populus |
Cherepanov (1991) |
| S. bacillicornis | China | Salix |
Wallin et al. (2017) Tavakilian and Chevillotte (2019) |
| S. balsamifera | Siberia, NE China, Korea, Japan |
Populus alba Salix |
Sheng and Hilszczanski (2009) Cherepanov (1991) |
| S. bilineatocollis | China, Russia | Unknown |
Danilevsky (2010) Tavakilian and Chevillotte (2019) |
| S. carcharias | Europe, northern Asia, China, Korea |
Alnus Populus Prunus Quercus Salix |
Cherepanov (1991) Tavakilian and Chevillotte (2019) |
| S. facetula | Vietnam | Unknown ‐ | Holzschuh (1999) |
| S. gilanense | Iran | Unknown | Tavakilian and Chevillotte (2019) |
| S. gleneoides | Laos, Vietnam | Unknown | Tavakilian and Chevillotte (2019) |
| S. interrupta | Siberia, NE China, Korea, Japan |
Abies (fir) Picea (spruce) Pinus (pine) other conifers |
Cherepanov (1991) |
| S. kojimai | Taiwan | Unknown | Tavakilian and Chevillotte (2019) |
| S. maculosa |
Iran, Transcaspia (=Azerbaijan, Kazakhstan, Turkmenistan Uzbekistan) |
Unknown | Tavakilian and Chevillotte (2019) |
| S. octomaculata | Northern Asia, NE China, Korea, Japan |
Ulmus others deciduous trees |
Cherepanov (1991) |
| S. octopunctata | Europe, Russia | Unknown | Tavakilian and Chevillotte (2019) |
| S. ohbayashii | Japan | Unknown | Wikipedia |
| S. perforata | Europe, northern Asia |
Populus tremuloides rarely other Populus spp.. Salix Sorbus |
Cherepanov (1991) |
| Saperda populnea | Europe, North Asia |
Populus Salix |
Felt and Joutel (1904) Cherepanov (1991) |
| S. quercus | Europe (Hungary, Greece, Bulgaria, Serbia, Montenegro) | Unknown | Tavakilian and Chevillotte (2019) |
| S. scalaris | Europe, Siberia, NE China, Korea |
Mainly Betula, rarely other deciduous species such as Prunus padus, Salix, Sorbus and Ulmus |
Cherepanov (1991) |
| S. similis | Northern Mongolia, NE China, Korea | Salix | Cherepanov (1991) |
| S. simulans | China | Unknown | Tavakilian and Chevillotte (2019) |
| S. subobliterata | China, Japan, Russia |
Ulmus japonica Ulmus laciniata |
Tavakilian and Chevillotte (2019) |
| S. viridipennis | China | Unknown | Tavakilian and Chevillotte (2019) |
References
Cherepanov AI, 1991. “Cerambycidae of Northern Asia, Volume 3, Lamiinae, Part III,” Oxonian Press PVT., New Delhi and Calcutta.
Danilevsky ML, 2010. Additions and corrections to the new Catalogue of Palaearctic Cerambycidae (Coeloptera) edited by I. Löbl and A. Smetana. Russian Entomological Journal 19, 215–239.
Felt EM, and Joutel L, 1904. “Monograph of the genus Saperda,” New York State Education Dept., Albany.
Holzschuh C, 1999. Beschreibung von 71 neuen Bockkäfern aus Asien, vorwiegend aus China, Laos, Thailand und Indien (Coleoptera, Cerambycidae). In “Schriftenreihe der Forstlichen Bundesversuchanstalt (FBVA‐Berichte)”, pp. 1–64, Wien, Austria.
Mao‐Ling S, and Hilszczański J, 2009. Two new species of genus Xorides (Hymenoptera: Ichneumonidae) parasitizing Saperda balsamifera Motschulsky and Asias halodendri (Pallas) (Coleoptera: Cerambycidae) in China. Annales Zoologici, 59, 165–170.
Tavakilian G, and Chevilotte H, 2019. Titan database about Longhorns or Timber‐Beetles (Cerambycidae). Available online: http://titan.gbif.fr/accueil_uk.html [Accessed 04 August 2019].
Wallin H, Kvamme T, and Bergsten J, 2017. To be or not to be a subspecies: description of Saperda populnea lapponica ssp. n. (Coleoptera, Cerambycidae) developing in downy willow (Salix lapponum L.). ZooKeys, 691, 103.
Appendix D – Literature search and review on S. tridentata hosts
1.
A comprehensive literature review was conducted in order to explore the reported host range of S. tridentata.
The following databases and platforms were explored (last access: 21 October 2019):
-
–
JSTOR (https://www.jstor.org)
-
–
BHL ‐ Biodiversity heritage library (https://www.biodiversitylibrary.org)
-
–
WOS ‐ Web of Science (http://apps.webofknowledge.com, all databases)
These databases include a variety of documents including among others scientific and technical papers, technical bulletins, governmental reports and documents, conference proceedings, experts’ personal reports, manuals, textbooks, field fact sheets. Catalogues of insects (from museums or laboratories) with no indication of host(s) and documents including duplicates of other documents (i.e. not being literature review) were not considered.
The review was focused on retrieving information regarding the hosts of S. tridentata, i.e. other information related to the biology of the insect were not considered for this review.
Considering that the literature focusing on the hosts of S. tridentata is very poor, and in order to retrieve as much information as possible, the web search comprised the main scientific name and synonyms (Section 3.1.1) of the pest, i.e.:
-
–
“Saperda tridentata”,
-
–
“Eutetrapha tridentata”,
-
–
“Compsidea tridentata”,
-
–
“Saperda dubiosa”.
The documents considered for the review were assigned to four categories:
-
–
No information: documents mentioning S. tridentata but not its host.
-
–
Direct observation: documents reporting the direct observation of S. tridentata and its host(s) (in open field or under laboratory conditions).
-
–
Generic: document mentioning S. tridentata and its host(s) but neither based on direct observation nor referencing other documents, e.g. manuals, textbooks, technical sheets.
-
–
Literature review: documents explicitly refereeing to other documents which mention S. tridentata and its host(s).
Table D.1 summarise the total documents found in JSTOR, WOS and BHL, using the four different search key, before removing duplicates and catalogues.
Table D.1.
Number of documents found in JSTOR, WOS and BHL, using the four search keys, before removing duplicates and catalogues
| Search key | JSTOR | WOS | BHL |
|---|---|---|---|
| “Saperda tridentata” | 0 | 39 | 416 |
| “Eutetrapha tridentata” | 0 | 2 | 4 |
| “Compsidea tridentata” | 2 | 0 | 37 |
| “Saperda dubiosa” | 0 | 0 | 2 |
Most of the documents were found in the BHL database; however, many of those documents were duplicates. Table D.2 shows the results of the literature review. After removal of duplicates and insect catalogues, the total number of documents reviewed was 304 of which 121 gathered no information, 78 were generic documents, 22 literature reviews and 83 direct observations.
Table D.2.
Documents found in the JSTOR, WOS and BHL database after removal of duplicates and catalogues and their classification according to the source of information and the reported hosts
| Reference | Type of source information | Reported hosts |
|---|---|---|
| 1. Abdullah, M., Abdullah, A. (1966). Saperda Fabricius, 1775 = Eutetrapha Bates, 1884, syn. n. (Coleoptera: Cerambycidae, Lamiinae), with a catalogue, new records, colour variation and a key to the species. In Proceedings of the Royal Entomological Society of London. Series B, Taxonomy, Vol 35(7–8): 87–94 | Direct observation | Elm |
| 2. Academy of Natural Sciences of Philadelphia Entomological Section, 1904. “Entomological news and proceedings of the Entomological Section of the Academy of Natural Sciences of Philadelphia” Entomological Rooms of the Academy of Natural Sciences, Philadelphia, Vol 15 | Direct observation | Elm |
| 3. Academy of Natural Sciences of Philadelphia Entomological Section, 1902. Entomological news and proceedings of the entomological section of the Academy of Natural Sciences of Philadelphia. Entomological News, Vol 13. | No information | No information |
| 4. Academy of Natural Sciences of Philadelphia Entomological Section, 1915. “Entomological news and proceedings of the Entomological Section of the Academy of Natural Sciences of Philadelphia”, Vol 26 | No information | No information |
| 5. Adams CC, 1915. “An ecological study of prairie and forest invertebrates,” Illinois State Laboratory of Natural History, Urbana, Ill. | No information | No information |
| 6. Alexander C, 1944. “Investigation of materials which promise value in insect control,” Annual report of the Massachussetts Agricultural Experimental Station, Rep. No. 0097‐6776. Massachusetts Agricultural Experiment Station, Amherst, Mass., No 417 | Direct observation | Elm |
| 7. American Association of Economic Entomologists, 1902. “Proceedings of the fourteenth annual meeting of the Association of Economic Entomologists,” U.S. Dept. of Agriculture, Division of Entomology, Washington, D.C., Vol 37 | Direct observation | Elm |
| 8. American Microscopical Society, 1918. “Transactions of the American Microscopical Society, Organized 1878, Incorporated 1891” Columbus, Ohio, Vol 37 | No information | No information |
| 9. Anonymous, 1935. Eleventh National Shade Tree Conference Proceedings (Abstract). Eleventh National Shade Tree Conference Proceedings., 167 pp | No information | No information |
| 10. Atkinson MD, 1992. Betula Pendula Roth (B. Verrucosa Ehrh.) and B. Pubescens Ehrh. Journal of Ecology, 80, 837–870. | No information | No information |
| 11. Bailey LH, and Miller W, 1906. “Cyclopedia of American horticulture, comprising suggestions for cultivation of horticultural plants, descriptions of the species of fruits, vegetables, flowers and ornamental plants sold in the United States and Canada, together with geographical and biographical sketches, and a synopsis of the vegetable kingdom,” 4th ed., Ed. Doubleday, Page & Company, New York. | Generic | Elm |
| 12. Baker WC, 1941. Type of wood preferred by Coleoptera commonly found in decadent parts of living elm trees. Journal of Economic Entomology, 34, 475–476. | Direct observation | Elm |
| 13. Baker WL, 1972. “Eastern forest insects ‐ Miscellaneous publication,” U.S. Dept. of Agriculture, Forest Service;, Washington, No 1175 | Generic | U. americana, U. rubra |
| 14. Banks N, and Henshaw S, 1889. Bibliography of the more important contributions to American economic entomology, Govt. print. off., Washington | No information | No information |
| 15. Barnes J, 1988. “Asa Fitch and the emergence of American entomology”. University of the State of New York, State Education Dept., Albany, N.Y., pp 122 | No information | No information |
| 16. Barnes J, 2003. “Natural History of the Albany Pine Bush: Albany and Schenectady Counties, New York,” New York State Museum. | No information | No information |
| 17. Barr M, Rogerson C, Smith S, and Haines J, 1986. “An annotated catalogue of the Pyrenomycetes described by Charles H. Peck,” University of the State of New York, State Education Dept., Albany, N.Y. | No information | No information |
| 18. Beutenmüller W, 1891. Bibliographical catalogue of the described transformations of North American Coleoptera. Journal of the New‐York Microscopical Society, 7, 1–52. | No information | No information |
| 19. Beutenmuller W, 1896. Food‐Habits of North American Cerambycidæ. Journal of the New York Entomological Society, 4, 73–81. | Generic | Elm |
| 20. Blatchley WS, 1910. “An illustrated descriptive catalogue of the Coleoptera or beetles (exclusive of the Rhynchophora) known to occur in Indiana,” Nature Pub. Co., Indianapolis. | Generic | U. americana, U. rubra |
| 21. Board of Regents of the University of the State of New York, 1889. “Annual report of the Regents,” J.B. Lyon, State Printer, Albany, NY, Vol 43 | No information | No information |
| 22. Board of Regents of the University of the State of New York, 1894. “48th Annual report of the Regents of the New York State Museum,” J.B. Lyon, State Printer, Albany, NY, Vol 48 | Literature review | Elm |
| 23. Board of Regents of the University of the State of New York, 1898. “Annual report of the Regents,” J.B. Lyon, State Printer, Albany, NY, Vol 52. | Generic | Elm |
| 24. Board of Regents of the University of the State of New York, 1899. “53d Annual report of the Regents 1899,” J.B. Lyon, State Printer, Albany, NY, Vol 53. | Generic | Elm |
| 25. Board of Regents of the University of the State of New York (1900). “Annual report of the Regents,” J.B. Lyon, State Printer, Albany, NY, Vol 54 | Generic | Elm |
| 26. Board of Regents of the University of the State of New York, 1903. “55th Annual report of the Regents 1901,” J.B. Lyon, State Printer, Albany, NY, Vol 55. | Literature review | U. americana |
| 27. Boiteau G, 1983. “The arthropod fauna of potato fields: composition and abundance.”, Fredericton, New Brunswick, CA, Contribution 1983‐16E. | No information | No information |
| 28. Breuning S, 1952. Revision einiger Gattungen aus der Gruppe der Saperdini Muls. (Col. Cerambycidae). Entomologische Arbeiten aus dem Museum G. Frey Tutzing bei München, 3, 107–213. | No information | No information |
| 29. Britton WE, 1933. “Connecticut State Entomologits thirty‐second report 1932,” Connecticut Agricultural Experiment Station, New Haven, CT, Vol 349 | Direct observation | Elm |
| 30. Britton WE, 1935. “Report of the State Entomologist of Connecticut for the year,” New Haven, CT, Vol 383 | Direct observation | Elm |
| 31. Britton WE, 1936. “Connecticut state entomologist thirty‐sixth report ‐ 1936,” New Haven. | Direct observation | Elm |
| 32. Britton WE, 1937. “Connecticut State Entomologist thirty‐seven report,” New Haven. | No information | No information |
| 33. Britton WE, 1939. “Report of the State Entomologist of Connecticut for the year 1938,” New Haven, CT, Vol 428 | Direct observation | Elm |
| 34. Britton WE, Clinton G, and Filley W, 1924. “Second report of the Tree Protection Examining Board,” Connecticut Agricultural Experiment Station, New Haven, CT. | Generic | Elm |
| 35. Bromley SW, 1948. Tests on Control of Elm Scolytus. Journal of Economic Entomology, 41, 327. | No information | No information |
| 36. Bromley SW, 1950. Tests on elm Scolytus 1949. Journal of Economic Entomology, 43, 397–398. | No information | No information |
| 37. Brooklyn Entomological Society, 1937. “Bulletin of the Brooklyn Entomological Society,” The Brooklyn Entomological Society, Brooklyn, N.Y., Vol 32–33 | Generic | Elm |
| 38. Buchanan WD, and May C, 1942. Technique for artificially feeding Scolytus multistriatus and Saperda tridentata spores of Ceratosto‐mella ulmi and other substances. Phytopathology, 32, 95–97. | No information | No information |
| 39. Buck WR, Heald SV, Holmgren NH, and Zanoni TA, 1996. Index to American Botanical Literature. Brittonia, 48, 124–160. | No information | No information |
| 40. Campbell J, Sarazin M., and Lyons D, 1989. “Canadian beetles (Coleoptera) injurious to crops, ornamentals, stored products, and buildings,” Agriculture Canada, Ottawa, Ontario. | Literature review | U. americana, U. rubra |
| 41. Casey TL, 1913. Further studies among the American Longicornia. Memoirs on the coleoptera, 4, 193–388. | No information | No information |
| 42. Chadbourne A, and Savage H, 1883. “The journal of the Boston Zoological Society,” The Society, Boston, Vol 2. | No information | No information |
| 43. Clarke J, and Felt EM, 1905. “20th Report of the State Entomologist on injurious and other insects of the State of New York, 1904” New York State Education Dept., Albany, Bulletin 97, Entomology 24. | No information | No information |
| 44. Clarke J, and Felt EM, 1907. “22d Report of the State Entomologist on injurious and other insects of the State of New York, 1906” New York State Education Dept., Albany, Bulletin 110. | No information | No information |
| 45. Clarke J, and Felt EP, 1923. “35th report of the State Entomologist on injurious and other insects of the state of New York, 1921” Weed, Parsons and Co.; University of the State of New York, Albany. | No information | No information |
| 46. Clemens J, and Pettman D, 2004. Public Transport: Jaunting from the Spaceship Nomad to the HSS Tampa. In “Avoiding the Subject”, pp. 109–128. Amsterdam University Press. | No information | No information |
| 47. Coleman TW, and Seybold SJ, 2011. Collection History and Comparison of the Interactions of the Goldspotted Oak Borer, Agrilus Auroguttatus Schaeffer (Coleoptera: Buprestidae), with Host Oaks in Southern California and Southeastern Arizona, U.S.A. The Coleopterists Bulletin, 65, 93–108. | No information | No information |
| 48. Collins CW, 1935. Insect vectors of the Dutch Elm disease caused by the fungus Ceratostomella ulmi (Schwarz) Buisman. Proceedings of the National Shade Tree Conference (US), 127–132. | Generic | Elm |
| 49. Collins CW, 1941. Studies of elm insects associated with Dutch elm disease fungus. Journal of Economic Entomology, 34, 369–372. | Direct observation | Elm |
| 50. Collins CW, Buchanan WD, Whitten RR, and Hoffmann CH, 1936. Bark beetles and other possible insect vectors of the Dutch elm disease Ceratostomella ulmi (Schwarz) Buisman. Journal of Economic Entomology, 29, 169–176. | Direct observation | Elm |
| 51. Commissioners of Fisheries, Game and Forest of the State of New York (1900). “Fifth annual report of the Commissioners of Fisheries, Game and Forest of the State of New York,” Albany, NY. | Literature review | Elm |
| 52. Committee of the Natural History Society of Montreal, 1860. “The Canadian Naturalist and Geologist and proceedings of the natural history society of Montreal,” Dawson & Son, Montreal, CA., Vol 5 | No information | No information |
| 53. Committee of the Natural History Society of Montreal, 1861. The Canadian Naturalist and Geologist and proceedings of the natural history society of Montreal. Vol 6. | No information | No information |
| 54. Connecticut Agricultural Experiment Station, 1900. “Protection of shade trees in cities and towns” Connecticut Agricultural Experiment Station, New Haven, Connecticut, Bulletin 131 | Generic | Elm |
| 55. Craighead FC, 1923. “North American cerambycid larvae : a classification and the biology of North American cerambycid larvae,”, Bulletin 27 – New Series, Dept. of Agriculture, Ottawa | Generic | Ulmus sp. |
| 56. Craighead FC, 1950. “Insect enemies of eastern forests,” U.S. Dept. of Agriculture, Washington, D.C., Vol 657 | Generic | Elm |
| 57. Craighead FC, and Middleton W, 1930. “An annotated list of the important North American forest insects,” U.S. Dept. of Agriculture, Washington, D. C. | Literature review | Elm |
| 58. Cranshaw W, 2004. Trunk and Branch Borers. In “Garden Insects of North America”, pp. 460–499. Princeton University Press. | Generic | U. americana |
| 59. Cranshaw W, and Shetlar D, 2018. Insects associated with large branches and the trunk of trees and shrubs. In “Garden Insects of North America”, pp. 424–463. Princeton University Press. | Generic | U. americana |
| 60. Currie RP, 1905. “Catalogue of the exhibit of economic entomology at the Lewis and Clark Centennial Exposition, Portland, Oregon, 1905,” U.S. Dept. of Agriculture, Bureau of Entomology, Washington, D.C., Vol 53 | Generic | Elm |
| 61. Davenport Academy of Natural Sciences, 1897. “Proceedings of the Davenport Academy of Natural Sciences.” Vol 6 | No information | No information |
| 62. Davey J, 1914. “Shade Tree Insects, Lesson number ten,” Kent, Ohio. | Generic | Elm |
| 63. Daviault L, 1946. Forest Entomology. In Report of the Minister of Lands and Forests of the Province of Quebec for the year ending March 31st, 1945, Redempti Paradis, pp 113–122. | Direct observation | Elm |
| 64. Derek SS, and Reginald PW, 2005. Bioinventory of Rhode Island Coleoptera: 45 New Records. The Coleopterists Bulletin, 59, 311–327. | No information | No information |
| 65. Devereaux W, 1879. Captures of Noctuidæ at Clyde, Wayne Co., N.Y. The Canadian entomologist, 11, 105–110. | No information | No information |
| 66. Dougherty EC, 1951. Evolution of Zoöparasitic Groups in the Phylum Nematoda, with Special Reference to Host‐Distribution. The Journal of Parasitology, 37, 353–378. | No information | No information |
| 67. Douglass BW, 1912. “Fourth annual report of the State entomologist of Indiana,” W.B. Burford, Indianapolis :. | Generic | Elm |
| 68. Downing ER, 1922. “A naturalist in the Great Lakes region,” The University of Chicago Press, Chicago, Ill. | No information | No information |
| 69. Drooz A, 1985. “Insects of eastern forests,” US Department of Agriculture, Forest Service, Miscellaneous publication No 1426 | Generic | U. americana, U. rubra |
| 70. D'Urban WSM, and Bell R, 1860. “Contributions to Canadian natural history” Lovell, Montreal. | No information | No information |
| 71. Ehrmann E, 1897. Coleopterological notes from my brother's diary. Entomological news, and proceedings of the Entomological Section of the Academy of Natural Sciences of Philadelphia, 8, 168–170. | Direct observation | Elm, Maple |
| 72. Elias TS, 1970. The genera of ulmaceae in the southeastern United States. Journal of the Arnold Arboretum, 51, 18–40. | Literature review | Elm |
| 73. Emmons E, 1846. “Agriculture of New York : comprising an account of the classification, composition and distribution of the soils and rocks together with a condensed view of the climate and the agricultural productions of the state,” C. Van Benthuysen, Albany, Vol 5 | No Information | No Information |
| 74. Emmons E, 1854. “Coleoptera of the state of New York,” C. Van Benthuysen & Co., Albany, NY. | No Information | No Information |
| 75. English LL, 1976. “Illinois trees and shrubs: their insect enemies,” 6th print., with revisions, Illinois Natural History Survey, Urbana, Ill. | Generic | Elm |
| 76. Entomological Society of Canada, 1879. “The Canadian entomologist,” Entomological Society of Canada, Ottawa, Vol 11 | Direct observation | Elm |
| 77. Entomological Society of Canada, 1908. “The Canadian entomologist,” Entomological Society of Canad, Ottawa, Vol 40. | Direct observation | Elm |
| 78. Entomological Society of Ontario, 1890. “Twenty‐first annual report of the entomologicla society of Ontario,” Entomological Society of Ontario, Toronto, Canada. | Generic | Elm |
| 79. Entomological Society of Ontario, 1897a. “Twenty‐seventh annual report of the Entomological Society of Ontario 1896,” Entomological Society of Ontario, Toronto. | No information | No information |
| 80. Entomological Society of Ontario, 1897b. “Twenty‐eight annual report of the Fruit Growers’ Association of Ontario 1896,” Fruit Growers’ Association of Ontario, Toronto. | No information | No information |
| 81. Entomological Society of Ontario, 1901, Forty‐ninth annual report of the Entomological Society of Ontario 1900. The Entomological Society of Ontario, Vol 31. | Literature review | Linden, Poplar |
| 82. Entomological Society of Ontario, 1904. “Thirty‐fourth annual report of the Entomological Society of Ontario 1903,” Entomological Society of Ontario, Toronto. | Literature review | Elm |
| 83. Entomological Society of Ontario, 1906. “Thirty‐sixth annual report of the Entomological Society of Ontario,1905.” Entomological Society of Ontario, Toronto, Vol 36 | Generic | Elm |
| 84. Entomological Society of Ontario, 1919. “Forty‐ninth annual report of the Entomological Society of Ontario 1918” Entomological Society of Ontario, Toronto. | Direct observation | Elm |
| 85. Entomological Society of Washington, 1905. “Proceedings of the Entomological Society of Washington,” Entomological Society of Washington, Washington, Vol 7 | Direct observation | Elm |
| 86. Evans AV, 2014. “Beetles of Eastern North America,” Princeton University Press. | Generic | Elm |
| 87. Evans AV, and Hogue JN, 2006. Family accounts. In “Field Guide to Beetles of California”, pp. 49–278. University of California Press. | No information | No information |
| 88. Fellin DG, 1973. “Weevils attracted to thinned lodgepole pine stands in Montana,” Intermountain Forest and Range Experiment Station, Forest Service, U.S. Dept. of Agriculture, [Ogden, Utah] | No information | No information |
| 89. Felt EP, 1898b. “Elm‐leaf beetle in New York State,” New York State Education Dept., Albany. | Generic | Elm |
| 90. Felt EP, 1898c. “14th Report of the State Entomologist on injurious and other insects of the state of New York”. Bulletin of the New York State Museum, University of the State of New York, Albany, NY, Vol 5, No 23 | No information | No information |
| 91. Felt EP, 1899a. “Shade tree pests in New York State,” New York State Education Dept., Albany. | No information | No information |
| 92. Felt EP, 1899b. “Memorial of life and entomologic work of Joseph Albert Lintner, state entomologist, 1874–98,” University of the State of New York, Albany | Literature review | U. americana |
| 93. Felt EP, 1900a. “15th Report of the state entomologist on injurious and other insects of the State of New York,” New York State Education Dept., Albany | Direct observation | Elm |
| 94. Felt EP, 1900b. Illustrated descriptive catalogue of some of the more important injurious and beneficial insects of New York State. Bulletin of the New York State Museum, Vol 8(37) | Generic | Elm |
| 95. Felt EP, 1901. “16th Report of the state entomologist on injurious and other insects of the State of New York,” New York State Education Dept., Albany. | Generic | Elm |
| 96. Felt EP, 1904. “Bulletin of the New York State Museum,” New York State Education Dept., Albany, Bulletin 76, Entomology 21 | No information | No information |
| 97. Felt EP, 1905. “Insects affecting park and woodland trees,” New York State Education Dept., Albany. | Generic | Elm |
| 98. Felt EP, 1930. The economic importance of shade tree insects. Journal of Economic Entomology, 23, 109–113. | No information | No information |
| 99. Felt EP, 1932. Shade Trees Threatened by Insect Pests. The Scientific Monthly, 35, 59–62. | No information | No information |
| 100. Felt EP, and Bromley SW, 1930. Shade tree problems (Abstract). Proceedings of the Annual Meeting of the National Shade Tree Conference, 6, 13–23. | Generic | Elm |
| 101. Felt EP, and Bromley SW, 1937. Scientific Contributions (Abstract). Bulletin of the Bartlett Tree Research Laboratory, 24 pp. | Direct observation | Elm |
| 102. Felt EP, and Joutel L, 1904. “Monograph of the genus Saperda,” New York State Education Dept., Albany. | Literature review | U. americana, U. rubra |
| 103. Fiske WF, 1905. Catogenus rufus, a coleopterous parasite. Proceedings of the Entomological Society of Washington, 7, 90–92. | No information | No information |
| 104. Fitch A, 1885. “Fifth report on the noxious and other insects of the State of New York.” | Generic | U. rubra |
| 105. Fitch HS, 1965. “Miscellaneous publication ‐ University of Kansas, Museum of Natural History,” University of Kansas, Lawrence, KS, Vol 42 | No information | No information |
| 106. Forbes SA, 1912a. “Report of the State Entomologist on the noxious and beneficial insects of the state of Illinois,” State Entomologist. | Direct observation | U. americana |
| 107. Forbes SA, 1912b. “What is the matter with the elms in Illinois?” Bulletin of the Agricultural Experiment Station of the University of Illinois, Bulletin 154. | Generic | Elm |
| 108. Forbes SA, 1885. “Fourteenth report of the State Entomologist on the noxious and beneficial insects of the state of Illinois,” State Entomologist, Springfield, Ill, Vol 14 | Direct observation | U. americana |
| 109. Franklin BH, 1882. “The elements of forestry designed to afford information concerning the planting and care of forest trees for ornament or profit and giving suggestions upon the creation and care of woodlands with the view of securing the greatest benefit for the longest time, particularly adapted to the wants and conditions of the United States”, Robert Clarke & Co, Cincinnati (OH) | Generic | Elm |
| 110. Fruit Growers’ Association of Ontario, 1891. “Twenty‐second annual report of the Fruit Growers’ Association of Ontario 1890,” The Fruit Growers’ Association, Toronto, Vol 22 | Generic | Elm |
| 111. Fyles, T. W. (1905). Forest insects. In Report of the sixth annual meeting of the Canadian forestry association, Quebec. | Generic | Elm |
| 112. Garman H, 1915. “The locust borer (Cyllene robiniae) and other insect enemies of the black locust”, Reprint form the 2nd Biennial Report of the State Forester of Kentucky, The State Journal Company, Frankfort, KY. | No information | No information |
| 113. Goodey T, 1951. “Soil and freshwater nematodes.”, John Wiley & Sons, London (UK), New York (USA) | No information | No information |
| 114. Haack RA, 2017. Feeding biology of Cerambycids. In “Cerambycidae of the World: Biology and Pest Management” (Q. Wang, ed.), pp. 105–124. CRC Press, Boca Raton, Florida (US). | Generic | Ulmus sp. |
| 115. Haldeman SS, 1847. Corrections and Additions to His Paper on the Longicornia of the United States. Proceedings of the American Philosophical Society, 4, 371–376. | No information | No information |
| 116. Haldeman SS, 1853. Materials towards a History of the Coleoptera Longicornia of the United States. Transactions of the American Philosophical Society, 10, 27–66. | No information | No information |
| 117. Hamilton J, and Schmitt PJ, 1895. Catalogue of the Coleoptera of Southwestern Pennsylvania, with Notes and Descriptions. Transactions of the American Entomological Society (1890‐), 22, 317–381. | Generic | Elm |
| 118. Hanson HC, 1958. Principles Concerned in the Formation and Classification of Communities. Botanical Review, 24, 65–125. | No information | No information |
| 119. Hard JS, 1964. “The identification of primary red pine cone insects,” Lake States Forest Experiment Station, Forest Service, U.S. Department of Agriculture, [St. Paul] | No information | No information |
| 120. Harrington WH, 1883. Correspondence. The Canadian entomologist, 15, 79–80. | Direct observation | Maple |
| 121. Harrington WH, 1887. “Insects infesting Maple trees”, Report of the Fruit Gowers’ Association of Ontario for the Year 1886, Toronto. | Generic | Elm, Maple |
| 122. Harris TW, Flint CL, Sonrel A, and Agassiz L, 1862. “A treatise on some of the insects injurious to vegetation,” Edited by Charles L. Flint, secretary of the Massachusetts State Board of Agriculture, Orange Judd and Company, New York. | Direct observation | Elm |
| 123. Harrison GD, 1896. Life History of Ichthyura Strigosa Grote. Psyche, 7, 424–425. | No information | No information |
| 124. Hart CJ, Cope JS, and Ivie MA, 2013. A Checklist of the Cerambycidae (Coleoptera) of Montana, USA, with Distribution Maps. The Coleopterists Bulletin, 67, 133–148. | No information | No information |
| 125. Herrick GW, 1920. “Insects of economic importance,” New and rev. ed., Ed. The Macmillan company, New York, N.Y. | No information | No information |
| 126. Hoebeke ER, 1992. Obituary: In Memoriam LaVerne L. Pechuman 1913‐1992. Journal of the New York Entomological Society, 100, 641–651. | No information | No information |
| 127. Hoffmann CH, 1939. Observations on the biology of Saperda tridentata Oliv. and Magdalis armicollis Say (Coleoptera). Journal of Economic Entomology, 32, 848–851. | Direct observation | Elm |
| 128. Hoffmann CH, 1940. Additions to annotated lists of insects reared from elm bark and wood. Bulletin of the Brooklyn Entomological Society, 35, 54–63. | Direct observation | Elm |
| 129. Hollrung M, 1899. “Jahresbericht über die Neuerungen und Leistungen auf dem Gebiete des Pflanzenschutzes”, Verlagsbuchhandling Paul Parey, Berlin, Germany | Direct observation | U. americana |
| 130. Holt BD, 2013. A Preliminary Checklist of the Cerambycidae and Disteniidae (Coleoptera) of Alabama. The Coleopterists Bulletin, 67, 241–256. | No information | No information |
| 131. Hopkins AD, 1904. “Catalogue of exhibits of insect enemies of forests and forest products at the Louisiana Purchase Exposition, St. Louis, Mo., 1904,” U.S. Dept. of Agriculture, Division of Entomology, Washington, D.C. | Generic | Elm |
| 132. Horn GH, 1886. A Review of the Species Described by Olivier in the “Entomologie”. Transactions of the American Entomological Society and Proceedings of the Entomological Section of the Academy of Natural Sciences, 13, 135–144. | No information | No information |
| 133. Howard L, 1899. “Three insect enemies of shade trees”, Government Printing Office, Washington D.C., Vol 99 | Direct observation | U. americana |
| 134. Huard VA, 1905. “Le Naturaliste canadien” Éditions l'Ardoise, Neufchâtel, QC, Vol 32(4) | Generic | Elm |
| 135. Hubbard HG, 1874. Notes on the Habits of Magdalinus armicpllis Say. Psyche, 1, 5–6. | Direct observation | Elm |
| 136. Hubbard HG, Schwarz EA, and LeConte JL, 1878. The Coleoptera of Michigan. Proceedings of the American Philosophical Society, 17, 593–669. | No information | No information |
| 137. Illinois State Laboratory of Natural History, 1915. “Bulletin of the Illinois State Laboratory of Natural History” The Laboratory, Bloomington, Ill., Vol 11. | Generic | Elm |
| 138. Illinois State Laboratory of Natural History, 1918. “Bulletin of the Illinois State Laboratory of Natural History” Urbana, Ill., Vol 12. | Direct observation | Elm |
| 139. Imperial Bureau of Entomology, 1916. “The review of applied entomology. Series A: Agricultural,” Imperial Bureau of Entomology, London, UK. | No information | No information |
| 140. International Institute of Agriculture, 1916. “International review of the science and practice of agriculture ‐ Monthly bulletin of Agricultural Intelligence and plant diseases” International Institute of Agriculture, Bureau of Agricultural Intelligence and Plant Diseases, Rome, Year VII, Number 7, July 1916. | No information | No information |
| 141. Jacot AP, 1934. Acarina as possible vectors of the Dutch elm disease. Journal of Economic Entomology, 27, 858–859. | Generic | Elm |
| 142. Johannsen OA, 1912. “Insect notes for 1912,” Maine Agricultural Experiment Station, Orono, ME, Bulletin 207 | Direct observation | Elm |
| 143. Johnson H, 1915. Coleoptera found in the vicinity of Meriden, Connecticut. Entomological news, and proceedings of the Entomological Section of the Academy of Natural Sciences of Philadelphia, 26, 307–319. | No information | No information |
| 144. Kansas Academy of Science, 1907. “Transactions of the Kansas Academy of Science,” W.Y. Morgan, Topeka, Kan., Vol 21. | Direct observation | Elm |
| 145. Kaston BJ, 1939. “The native elm bark beetle, Hylurgopinus rufipes (Eichhoff), in Connecticut,” Connecticut Agricultural Experiment Station, New Haven. | Generic | Elm |
| 146. Keenan W, 1949. “Dutch Elm Disease Symposium” Entomologicla Society of Ontario, 79, 54–57 | Generic | Elm |
| 147. Kethidi DR, Roden DB, Ladd TR, Krell PJ, Retnakaran A, and Feng QL, 2003. Development of SCAR markers for the DNA‐based detection of the Asian long‐horned beetle, Anoplophora glabripennis (Motschulsky). Archives of Insect Biochemistry and Physiology, 52, 193–204. | No information | No information |
| 148. Kingsley JS, 1884. “The standard natural history. Vol II Crustacea and insects,” Boston, Mass. | Generic | Elm |
| 149. Klingeman WE, Youssef NN, Oliver JB, and Basham JP, 2017. The longhorned beetles (Coleoptera: Cerambycidae) of Tennessee: distribution of species, seasonal adult activity, and new state records. The Florida Entomologist, 100, 292–302. | No information | No information |
| 150. Krinsky WL, and Godwin PA, 1996. Long‐Horned Beetles from the Forest Canopy in New England and New York (Coleoptera:Cerambycidae). The Coleopterists Bulletin, 50, 236–240. | No information | No information |
| 151. Krischik V, and Davidson J, 2013. Elm Borer. In IPM (Integrated Pest Management) of Midwest Landscapes, V. Krischik and J. Davidson, eds., Minnesota Agricultural Experiment Station. pp. 115–116. | Generic | U. americana, U. rubra, Ulmus sp. |
| 152. Kulzer H, 1952. “Entomologische Arbeiten aus dem Museum G. Frey, München,” Eigenverlag., Münich, DE, Band 3, Heft 1 | No information | No information |
| 153. Lambert H, 1979. Notes sur Saperda tridentata Oliv. (Coleoptera: Cerambycidae). Febreries, 6, pp 37 | Direct observation | U. americana |
| 154. LeConte JL, 1850. An attempt to classify the Longicorn Coleoptera of the part of America North Mexico. Journal of the Academy of Natural Sciences of Philadelphia, Vol 2, Second series, Part 1, 2–178 | No information | No information |
| 155. Leng CW, and Hamilton J, 1896. The Lamiinæ of North America. Transactions of the American Entomological Society (1890‐), 23, 101–178. | Literature review | U. americana, U. rubra, Maple |
| 156. Leonard MD, 1928. “A List of the insects of New York: with a list of the spiders and certain other allied groups,” Cornell University, Ithaca, N.Y. | No information | No information |
| 157. Lingafelter SWL, and Horner NV, 1993. The Cerambycidae of North‐Central Texas. The Coleopterists Bulletin, 47, 159–191. | Direct observation | Elm |
| 158. Linsley EG, 1958. The role of cerambycidae in forest, urban, and agricultural environments. The Pan‐Pacific Entomologist, 34, 105–124. | No information | No information |
| 159. Lintner JA, 1892. “Report of the State Entomologist on injurious and other insects of the state of New York,” Weed, Parsons and Co.; University of the State of New York, Albany. | No information | No information |
| 160. Lintner JA, 1897. “Twelfth report on the injurious and other insects of the state of New York for the year 1896 (From the fitieth report on the New York State Museum),” University of the State of New York, Albany. | Literature review | Elm, U. americana, U. rubra |
| 161. Lintner JA, 1895. “Report of the State Entomologist on injurious and other insects of the state of New York, 1895” Weed, Parsons and Co.; University of the State of New York, Albany, NY | Literature review | Elm, U. americana |
| 162. Lochhead W, 1919. “Class book of economic entomology, with special reference to the economic insects of the northern United States and Canada,” Blakiston, Philadelphia | Generic | Ulmus sp. |
| 163. Long BL, Kurta A, and Clemans DL, 2013. Analysis of DNA from Feces to Identify Prey of Big Brown Bats (Eptesicus fuscus) Caught in Apple Orchards. American Midland Naturalist, 170, 287–297. | No information | No information |
| 164. Lugger O, 1899. Beetles injurious to fruit‐producing plants. Bulletin of the Agricultural Experiment Station of the University of Minnesota 66. | Generic | Poplar, Linden |
| 165. MacAloney HJ, and Ewan HG, 1964. “Identification of hardwood insects by type of tree injury, north‐central region,” Lake States Forest Experiment Station, St. Paul, Vol 11. | Generic | Elm |
| 166. MacRae TC, 1993. Annotated checklist of the longhorned beetles (Coleoptera: Cerambycidae and Disteniidae) occurring in Missouri. Insecta mundi, 7, 223–252. | Direct observation | U. americana, U. alata |
| 167. Malloch J, 1918. A preliminary classification of Diptera, exclusive of Pupipara, based upon larval and pupal characters, with keys to imagines in certain families. Part 1. Bulletin of the Illinois State Laboratory of Natural History, 12, 161–409. | No information | No information |
| 168. Martin CH, 1936. Preliminary report of trap‐log studies on elm bark beetles. Journal of Economic Entomology, 29, 297–306. | Direct observation | Elm |
| 169. Maine Agricultural Experiment Station, 1913. “Twenty‐eighth annual report of the Maine Agricultural Experiment Station.” Maine Agricultural Experiment Station, Orono, Maine, Vol 28. | Literature review | Elm |
| 170. Massachusetts Agricultural Experiment Station, 1936. “Annual Report” Massachusetts Agricultural Experiment Station, Amherst, No 327. | Generic | Elm |
| 171. Massachusetts Agricultural Experiment Station, 1937. “Annual Report,” Rep. No. 0097‐6776. Massachusetts Agricultural Experiment Station, Amherst, Vol 346. | No information | No information |
| 172. Massachusetts Agricultural Experiment Station, 1944. Insect infestation of stored logs. Bulletin of the Massachusetts Agricultural Experimental Station, 41–43. | Direct observation | Elm |
| 173. Massachusetts Board of Agriculture, 1858. “Annual report of the Secretary of the Massachusetts Board of Agriculture,” W. White, Boston, Mass., Vol 6. | No information | No information |
| 174. May C, 1935. The Dutch Elm disease from the research standpoint. Proceedings of the national Shade Tree Conference (US)., 122–127. | Generic | Elm |
| 175. Mayer P, and Giesbrecht W, 1886. “Zoologischer Jahresbericht fur 1885. Herausgegeben von der zoologischen station zu Neapel.” | No information | No information |
| 176. Merril FI, and Felt EM, 1902. “17th Report of the State Entomologist on injurious and other insects of the State of New York – 1901”, New York State Education Dept., Albany, NY, Bulletin 53, Entomology 14. | Generic | U. americana |
| 177. Merril FI, and Felt EM, 1902b. “Elm leaf beetle in New York State,” New York State Education Dept., Albany, NY. | Generic | Elm |
| 178. Merril FI, and Felt EM, 1904. “19th Report of the State Entomologist on injurious and other insects of the state of New York 1903,” Weed, Parsons and Co.; University of the State of New York, Albany, NY. | No information | No Information |
| 179. Merril FI, and Felt EP, 1903. “18th Report of the State Entomologist on injurious and other insects of the State of New York ‐ 1902,” Weed, Parsons and Co.; University of the State of New York, Albany, NY, Bulletin 64, Entomology 17. | No information | No Information |
| 180. Merril J, 1916. Life‐history and habits of two new nematodes parasitic to insects. Journal of Economic Entomology, 9, 148–149. | Direct observation | Elm |
| 181. Metcalf RL and Metcalf RA, 1993. Destructive and useful insects ‐ Their habitats and control. McGraw‐Hill, Inc. | Generic | U. americana, U. rubra |
| 182. Middleton W, 1929b. Factors influencing the activity of shade‐tree insects and the utilization of these in control work. In”Fourth international congress of entomology” (K. Jordan and W. Horn, eds.), Vol. 1, Ithaca, NY. | Generic | Elm |
| 183. Miller WJ, 1921. “New York State Museum bulletin,” University of the State of New York, Albany, N.Y., 245–248. | No information | No information |
| 184. Mitchell RS, 1988. “Platanaceae through Myricaceae of New York State,” University of the State of New York, State Education Dept., Albany, N.Y., Vol 464. | Generic | Ulmus sp. |
| 185. Montandon AL, 1906. Notes sur la faune entomologique de la Roumanie. Buletinul Societății de Științe din București‐România/Bulletin de la Société des Sciences de Bucarest‐Roumanie, 15, 30–80. | No information | No information |
| 186. Mook PV, and Wolfenbarger DO, 1943. Distribution of Beauveria bassiana on elm insects in the United States. Phytopathology, 33, 76–77. | Direct observation | Elm |
| 187. Moore AE, 1957. “Bibliography of forest disease research in the Department of Agriculture,” U.S. Dept. of Agriculture, Washington, D.C. | No information | No information |
| 188. Morris FJA, 1909. “Some beetle haunts” by an amateur botanist. Thirty‐ninth annual report of the Entomological Society of Ontario, 1908, 39. | Direct observation | Elm |
| 189. Morris FJA, 1916. “Reports on Insects of the Year; Division no. 5, Port Hope District (Abstract)”. 46th Annual Report of the Entomological Society of Ontario 1915, pp17–21. | Direct observation | Elm |
| 190. Morris FJA, 1920. Popular and Practical Entomology. Familiar Haunts. The Canadian Entomologist, 52, 73–76. | Direct observation | Elm |
| 191. Muséum national d'histoire naturelle, 1973. “Bulletin du Muséum national d'histoire naturelle. Série 3, n° 123, Zoologie 95, Paris, France. | No information | No information |
| 192. New York Entomological Society, 1902. Proceedings of the New York Entomological Society. Journal of the New York Entomological Society, 10, 170–176. | No information | No information |
| 193. New York Entomological Society, 1908. Proceedings of the New York Entomological Society. Journal of the New York Entomological Society, 16, 115–124. | No information | No information |
| 194. New York Legislature Assembly, 1840. Communication from the governor, transmitting several reports relative to the geological survey of the State. In “Hume Tracts”. | No information | No information |
| 195. New York State Museum, 1896. “Annual report of the Regents,” J.B. Lyon, State Printer, Albany. | Literature review | U. americana, U. rubra |
| 196. New York State Museum, 1903. “Bulletin of the New York State Museum,” New York State Education Dept., Albany, Bulletin 66, Miscellaneous 2. | No Information | No information |
| 197. New York State Museum, 1904. 56th Annual Report 1902. Vol 3. | Generic | Elm |
| 198. New York State Museum, 1905. “57th Annual report 1903” University of the State of New York, Albany, Vol 1, Part 2 | Generic | Elm |
| 199. New York State Museum, 1906. “58th Annual report 1904” University of the State of New York, Albany, Vol 5 Appendix 7. | Generic | Elm |
| 200. New York State Museum, 1907. “59th Annual report 1905” University of the State of New York, Albany, Vol 3 Appendix 7. | Generic | Elm |
| 201. Olivier GA, 1795. “Entomologie ou Histoire naturelle des insectes, avec leurs caractères génériques et spécifiques, leur description, leur synonumie et leur figure enluminee, Coléoptères Tomes I‐V” Imp. Baudouin, Paris, Vol 4. | No information | No information |
| 202. Ottawa Field‐Naturalists Club, 1879. “Transactions ‐ Ottawa Field‐Naturalists’ Club,” Ottawa Field‐Naturalists’ Club., Ottawa., Vol 1. | Generic | Elm |
| 203. Packard AS, 1870. The Borers of Certain Shade Trees. The American Naturalist, 4, 588–594. | No information | No information |
| 204. Packard AS, 1881. Insects Injurious to Forest and Shade Trees. Bulletin of the US Entomological Commission, 7, 275. | Generic | Elm, U. rubra |
| 205. Packard AS, 1890. “Fifth report of the United States Entomological Commission: being a revised and enlarged edition of Bulletin no. 7, on insects injurious to forest and shade trees,” G.P.O., Washington, Vol 5. | Generic | Elm, U. americana, U. rubra, Maple |
| 206. Packard AS, Riley CV, and Thomas C, 1883. “Third report of the United States Entomological Commission, relating to the Rocky Mountain locust, the western cricket, the army worm, canker worms, and the Hessian fly; together with descriptions of larvae of injurious forest insects, studies on the embryological development of the locust and of other insects, and on the systematic position of the orthoptera in relation to other orders of insects; with maps and illustrations” Washington Government Printing Office | No information | No information |
| 207. Packard AS, 1869. Guide to the study of insects, and a treatise on those injurious and beneficial to crops: for the use of colleges, farm‐schools, and agriculturists. Naturalist's Book Agency, Salem, Mass. | Literature review | Elm |
| 208. Packard AS, 1869. Record of American entomology for the year 1868. Naturalist's Book Agency, Salem, Mass. | No information | No information |
| 209. Packard AS, 1873. Our common insects. A popular account of the insects of our fields, forests, gardens and houses. Naturalists’ agency, Salem, Mass. | Direct observation | Elm |
| 210. Park O, 1931. Studies in the Ecology of Forest Coleoptera. Ecology, 12, 188–207. | Direct observation | Elm, Carya cordiformis |
| 211. Pechuman LL, 1937. An annotated list of insects found in the bark and wood of Ulmus americana L. in New York State. Bulletin of the Brooklyn Entomological Society, 32, 8–21. | Direct observation | U. americana |
| 212. Pechuman LL, 1940. Notes on the feeding and breeding habits of Saperda tridentata Oliv. Bulletin of the Brooklyn Entomological Society, 35, 113–116. | Direct observation | Elm |
| 213. Perkins GH, 1890. “Insects injurious to the American elm,” Argus, Montpelier. | Generic | Elm |
| 214. Pinchot G, and Ashe WW, 1897. “Timber trees and forests of North Carolina,” M. I. & J. C. Stewart, public printers, Winston, Vol 6. | Generic | U. americana |
| 215. Popenoe EA, 1877. A List of Kansas Coleoptera. Transactions of the Kansas Academy of Science (1872‐1880), 5, 21–40. | No information | No information |
| 216. Quebec Society for the Protection of Plants from Insects and Fungous Diseases, 1915. “Annual report of the Quebec Society for the Protection of Plants from Insects and Fungous Diseases,” The Society, Quebec, 7. | Generic | Elm |
| 217. Rau P, 1923. Ecological and behaviour notes on Missouri insects. Transactions of the Academy of Science of St.Louis, 25. | No information | No information |
| 218. Readio PA, 1935. The entomological phases of the Dutch elm disease. Journal of Economic Entomology, 28, 341–353. | Direct observation | Elm |
| 219. Remillet M, and Waerebeke DV, 1973. Eudronema intestinalis n. g., n. sp. (Nematoda, Rhabditidae), parasite de l'intestin postérieur d'Eudromus striaticollis Brullé (Coleoptera, Harpalidae). Bulletin du Muséum national d'histoire naturelle, 123, 555–560. | No information | No information |
| 220. Rice ME, Merickel F, and MacRae TC, 2017b. The longhorned beetles (coleoptera: Cerambycidae) of idaho. Coleopterists Bulletin, 71, 667–678. | No information | No information |
| 221. Riley C, 1880. Food habits of the longicorn beetles or wood borers. The American entomologist, 3, 270–271. | Literature review | Elm, U. rubra |
| 222. Rivista di Patologia Vegetale, 1939. Brevi notizie e note pratiche. Rivista di Patologia Vegetale, 29, 393–398. | Literature review | U. americana |
| 223. Robert A, 1958. Recherches sur le cycle evolutif de la Saperde de l'orme, Saperda tridentata (Oliv.)‐(Coleopteres‐Cerambycidae). Annales de la Societe Entomologique de Quebec, 3, 46–67. | Direct observation | Elm |
| 224. Rose A, and Lindquist O, 1997. “Insects of eastern hardwood trees,” Natural Resources Canada, Canadian Forest Service, Forestry Technical Report 29 | Generic | Elm |
| 225. Russel L, 1909. “Annual report of the Nebraska State Horticultural Society” The Nebraska State Horticultural Society, Lincoln, Neb. | Direct observation | Elm |
| 226. Russo R, 2006. The gall inducers. In “Field Guide to Plant Galls of California and Other Western States”, pp. 37–71. University of California Press. | No information | No information |
| 227. Sabrosky CW, 1951. A Revision of the Nearctic Species of the Genus Gaurax (Diptera, Chloropidae). The American Midland Naturalist, 45, 407–431. | Literature review | Elm |
| 228. Schenck CA, 1909. “Forest protection; guide to lectures delivered at the Biltmore Forest School,” The Inland Press, Asheville, N. C. | Literature review | Ulmus sp. |
| 229. Schenck CA, and Oberholser HC, 1907. “Baltimore lectures on sylviculture,” Brandow Printing Co., Albany, N.Y. | Literature review | Ulmus sp. |
| 230. Schiefer TL, 1998. A Preliminary List of the Cerambycidae and Disteniidae (Coleoptera) of Mississippi. Transactions of the American Entomological Society (1890‐), 124, 113–131. | No information | No information |
| 231. Schiefer TL, and Newell P, 2010. A Distinctive New Subspecies of Saperda lateralis F. (Coleoptera: Cerambycidae) from the Southeastern United States. The Coleopterists Bulletin, 64, 329–336. | No information | No information |
| 232. Shade Tree Commission of Newark, 1913. Protection of street trees. Park and cemetery and landscape gardening, Vol 23 | Generic | U. americana |
| 233. Smith RC, and Kelly EG, 1933. A Summary of the Population of Injurious Insects in Kansas for 1932. Journal of the Kansas Entomological Society, 6, 37–60. | No information | No information |
| 234. Smith RC, and Kelly EG, 1935. The Fourth Annual Summary of the More Important Insects of Kansas Covering the Year 1934. Transactions of the Kansas Academy of Science (1903‐), 38, 171–185. | No information | No information |
| 235. Snyder LL, Logier E, Kurata T, Urquhart F, and Brimley J, 1941. “A faunal investigation of Prince Edward County, Ontario” University of Toronto Press, Toronto. | Direct observation | Elm |
| 236. Solomon JD, 1995. “Guide to insect borers in North American broadleaf trees and shrubs,” USDA, Service Agriculture Handbook AH‐706. | Generic | U. americana, U. crassifolia, U. rubra |
| 237. Solomon JD, Doolittle RE, and Spilman TJ, 1976. Cerambycid Beetles Captured in Sticky‐Traps in Mississippi. The Coleopterists Bulletin, 30, 289–290. | No information | No information |
| 238. Stein JD, 1976. “Insects : a guide to their collection, identification, preservation, and shipment,” Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. | Generic | U. pumila |
| 239. Stein JD, and Kennedy PC, 1972. “Key to shelterbelt insects in the Northern Great Plains,” Rocky Mountain Forest and Range Experiment Station, Forest Service, U.S. Dept. of Agriculture, Fort Collins, Colo. | Generic | U. americana, U. pumila |
| 240. Swaine JM, and Hutchings CB, 1926. “The more important shade tree insects of eastern Canada and their control,” Minister of Agriculture, Ottawa, CA, Vol 63 | Generic | Elm |
| 241. Tehon LR, 1934. Elm disease in Illinois. National Shade Tree Conference Proceedings ‐ Annual Meeting, 10, 105–111. | Generic | Elm |
| 242. Thomas C, 1875. “Sixth report of the State Entomologist on the noxious and beneficial insects of the state of Illinois,” State Entomologist, Springfield, Ill. Vol 6. | Generic | Elm |
| 243. Townes H, Townes M, Walley G, Walkley L, Habeck D, and Townes G, 1960. “Ichneumon‐flies of America North of Mexico: 2. Subfamilies Ephialtinae, Xoridinae, Acaenitinae,” Washington, D.C. | Direct observation | Ulmus sp., U. americana |
| 244. Townsend C HT, 1893. Some notes on Michigan insects, principally Coleoptera, affecting forest trees. The Canadian Entomologist, 25, 201–204. | Direct observation | Elm |
| 245. Troop JB, 1905. Report of the state entomologist. Indiana State Board of Agriculture 1904‐1905, 46. | Generic | Elm |
| 246. Tucker ES, 1907. Concerning Some Insects Collected and Bred from Dead and Dying Elm. Transactions of the Kansas Academy of Science (1903‐), 21, 158–162. | Direct observation | Elm |
| 247. Tyler LJ, Parker KG, and Pechuman LL, 1939. The relation of Saperda tridentata to infection of American elm by Ceratostomella ulmi. Phytopathology, 29, 547–549. | Direct observation | U. americana |
| 248. United States Bureau of Entomology and Plant Quarantine, 1935. “Newsletter.” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 3(1). | Generic | Elm |
| 249. USDA ‐ Bureau of Entomology, 1925. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 5 | Direct observation | Elm |
| 250. USDA ‐ Bureau of entomology and plant quarantine, 1939. “Newsletter.” USDA, Vol 7 | No information | No information |
| 251. USDA ‐ Bureau of entomology and plant quarantine, 1940. “Newsletter.” USDA, Vol 6 | Direct observation | Elm |
| 252. USDA ‐ Bureau of Entomology, 1921a. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 1 | Direct observation | Elm |
| 253. USDA ‐ Bureau of Entomology, 1921b. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 10 | Direct observation | Elm |
| 254. USDA ‐ Bureau of Entomology, 1922. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 2. | Direct observation | Elm |
| 255. USDA ‐ Bureau of Entomology, 1925. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 5. | Direct observation | Elm |
| 256. USDA ‐ Bureau of Entomology, 1928. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 8. | Direct observation | Elm |
| 257. USDA ‐ Bureau of Entomology, 1930. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 10. | No information | No information |
| 258. USDA ‐ Bureau of Entomology, 1932a. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 12. | Direct observation | Elm |
| 259. USDA ‐ Bureau of Entomology, 1932b. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 12 | Direct observation | Ulmus americana |
| 260. USDA ‐ Bureau of Entomology, 1935a. “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 2 | Direct observation | Elm |
| 261. USDA ‐ Bureau of Entomology, 1935b. “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 15. | Direct observation | Elm |
| 262. USDA ‐ Bureau of Entomology, 1935c. “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 15. | Direct observation | Elm |
| 263. USDA ‐ Bureau of Entomology, 1936a. “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 3. | Literature review | Elm |
| 264. USDA ‐ Bureau of Entomology, (1936b). “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 3 | Direct observation | Elm |
| 265. USDA ‐ Bureau of Entomology, 1937. “News letter” Bureau of Entomology and Plant Quarantine, Washington, D.C., Vol 4. | No information | No information |
| 266. USDA ‐ Bureau of Entomology, 1937. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 17. | Direct observation | Elm |
| 267. USDA ‐ Bureau of Entomology, 1940. “The Insect pest survey bulletin” Bureau of Entomology, U.S. Dept. of Agriculture, Washington, D.C., Vol 20. | Direct observation | Elm |
| 268. USDA, 1907. Insects injurious to shade and ornamental trees. In “Yearbook of the USDA1906”. | No information | No information |
| 269. USDA, 1954a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 4 | Direct observation | U. americana |
| 270. USDA, 1954b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 4. | No information | No information |
| 271. USDA, 1955. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 5. | Direct observation | Elm |
| 272. USDA, 1955. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 5. | No information | No information |
| 273. USDA, 1956a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 6. | No information | No information |
| 274. USDA, 1956b. “Cooperative economic insect report”, Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 6. | No information | No information |
| 275. USDA, 1957a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 7. | No information | No information |
| 276. USDA, 1957b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 7. | No information | No information |
| 277. USDA, 1957c. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 7. | Direct observation | Elm |
| 278. USDA, 1957d. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 7. | Direct observation | Elm |
| 279. USDA, 1957e. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 7. | Direct observation | Elm |
| 280. USDA (1958a). “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 8(27) | No information | No information |
| 281. USDA, 1958a. Cooperative economic insect report. Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., 8. | Direct observation | Elm |
| 282. USDA, 1958b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 8. | Direct observation | Elm |
| 283. USDA, 1959a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 9. | No information | No information |
| 284. USDA, 1959b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 9. | No information | No information |
| 285. USDA, 1961. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 11. | Direct observation | Elm |
| 286. USDA, 1963a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 13. | No information | No information |
| 287. USDA, 1963b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 13. | No information | No information |
| 288. USDA, 1964a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 14. | Direct observation | Elm |
| 289. USDA, 1964b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 14. | Direct observation | Elm |
| 290. USDA, 1967a. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C., Vol 17. | Direct observation | Elm |
| 291. USDA, 1967b. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C..Vol 17. | Direct observation | Elm |
| 292. USDA, 1968. “Cooperative economic insect report” Plant pest control branch, Agricultural Research Service, USDA, Washington, D.C. | Direct observation | Elm |
| 293. USDA, 1985. “Insects of Eastern forests”, U.S. Forest Service, Washington, D.C., Vol 1426. | Generic | U. americana, U. rubra |
| 294. USDA‐ Bureau of Entomology, 1909. “The principal injurious insects of the year 1908”. Yearbook of the USDA1908. Washington. | No information | No information |
| 295. Vlasak J, and Vlasakova K, 2002. Records of Cerambycidae (Coleoptera) in Massachusetts with Notes on Larval Hosts. The Coleopterists Bulletin, 56, 203–219. | Direct observation | Ulmus sp. |
| 296. Washburn F, 1910. “Thirteenth Report of the State Entomologist of Minnesota to the Governor for the years 1909 and 1910,” Agricultural Experiment Station, St. Anthony Park, Minn, Vol 13. | Direct observation | Poplar |
| 297. Washburn FL, 1918. “Injurious insects and useful birds, successful control of farm pests,” J.B Lippincott company, Philadelphia. | Generic | Elm |
| 298. Weed CM, 1894. Entomology. The American Naturalist, 28. | Generic | Elm |
| 299. Welch PS, 1916. Entomological Notes. Transactions of the American Microscopical Society, 35, 257–268. | No information | No information |
| 300. Welch PS, and Wehrle LP, 1918. Observations on Reproduction in Certain Parthenogenetic and Bisexual Nematodes Reared in Artificial Media. Transactions of the American Microscopical Society, 37, 141–176. | No information | No information |
| 301. Werner F, 1927. Zur kenntnis der fauna einer xerothermischen lokalität in Niederösterreich (unteres kamptal). Zeitschrift für Morphologie und Ökologie der Tiere, 9, 1–96. | No information | No information |
| 302. Wickham HF, 1897. A list of Coleoptera from the southern shore of Lake Superior, with remarks on geographical distribution. Proceedings of the Davenport Academy of Natural Sciences, 6, 125–169. | No information | No information |
| 303. Wood SL, and Bright DE, 1987. A Catalog of Scolytidae and Platypodidae (Coleoptera), Part 1: Bibliography. Great Basin Naturalist Memoirs, 1–685. | No information | No information |
| 304. Yanega D, 1996. “Field guide to northeastern longhorn beetles (Coleoptera: Cerambycidae),” Illinois Natural History Survey, Champaign, Ill. | Generic | Elm |
Appendix E – Scoping review on the flight capacity of adult Saperda tridentata and other Cerambycidae, and analysis of spatial separation of European elm trees in Northern Italy
Scoping review on the flight capacity of adult S. tridentata and other Cerambycidae
Adult S. tridentata emerge from hosts and quickly mate (Solomon, 1995). However, unless females feed no eggs are laid (Pechuman, 1940). A mated female is, therefore, behaviourally required to locate a suitable host tree for feeding and, subsequently, for egg laying. Since egg laying takes place preferentially on weakened trees, females are likely to respond to cues given off by such trees which suggest they are capable to find suitable hosts even if they are uncommon in the landscape.
No literature reporting experimental studies examining flight distance of S. tridentata could be found. We therefore draw on literature describing the flight and dispersal of other cerambycid species feeding on deciduous hosts, including literature on other species of Saperda.
Short distance flight is typical of many cerambycids and the natural spread rate of longhorn beetles is generally considered to be quite slow because adults reproduce on, or close to, the natal host (Hack et al., 2010; Rhainds et al., 2011). For example, Felt and Joutel (1904) report that Saperda fayi usually oviposit on the tree they inhabited as borers and fly only a little, leading to their distribution being considered localised.
Where host density is moderate to high, there is no need for slow‐growing cerambycids to spread far. For example, Sawyer et al. (2011) examined the spread of Anoplophora glabripennis within US outbreak sites. In areas with abundant hosts, the population remained on a few trees within a few metres of each other for the first few years, e.g. 90% of oviposition sites were within 140 m of the nearest site of emergence and 99% were within 300 m. This contrasted with spread of approximately 3,200 m over 5 years in a region with much lower host density. Hence, the lack of hosts encouraged wider dispersal. Similar distances are reported for A. glabripennis in Italy where survey results used in a model suggest 80% of dispersal was within 300 m of the nearest infested hosts although some adults did spread more than 2 km from the nearest infested host (Favaro et al., 2015).
If there is no suitable host in the vicinity, adult A. glabripennis can fly several hundred metres or more in a single flight. Lopez et al. (2017) conducted flight mill experiments to measure the dispersal potential of male and female adult A. glabripennis. From a study of 162 individuals, the mean flight distance during a 24 hour period was 2,272 m although the maximum distance an individual flew was 13,667 m. However, such experiments do not represent natural conditions and the distances flown in a mill are difficult to relate to those in the natural environment. A similar study carried out on invasive A. glabripennis in Europe confirmed that some individuals were capable of flying an accumulated distance of around 14 km during their lifetime (Javal et al. 2018). The authors speculate that the longer flights were prompted by searches for oviposition sites and scarcity of local resources. Analysis of patterns of attack by A. glabripennis in a large infestation in Worcester, Massachusetts by Trotter & Hull‐Sanders (2015) under different assumed attack scenarios suggested that longer distance dispersal occurred when local resources near natal trees were scarce.
In China, mark–release–recapture studies demonstrated that, although 72% of A. glabripennis adults were recaptured within 300 m of release points, some beetles were recaptured up to 2,600 m away (Smith et al., 2004).
There are observations on single flight distances of Saperda candida (the round‐headed apple tree borer) by Hess (1940). Hess reports capturing adults then releasing them individually from the hand and watching how far they flew in a single flight. Although Hess (1940) reports distances in yards, his estimates are reported in the table below in SI units.
Table E.1.
Observed distances of single flights by adult Saperda candida (Hess, 1940)
| Number of adults tested | Mean distance (m) | Maximum distance (m) | |
|---|---|---|---|
| Males | 52 | 10.1 | > 82.3 |
| Female | 61 | 6.4 | 20.1 |
Hess (1940) suggested that S. candida could spread within North America due to the presence of apple orchards and apples trees in peoples’ gardens. However, areas without hosts for ‘even a few miles’ would act as natural barriers and prevent spread since without human assistance the species would not be able to cross such barriers. The same is thought to be likely for S. tridentata. The distance between hosts will be a key factor influencing whether S. tridentata could establish in the EU.
Abiotic environmental conditions such as temperature, humidity, light level and wind speed are factors likely to influence adult flight. However, the specific criteria for initiating flight are unknown for S. tridentata. S. tridentata most often fly at night and, therefore, host location is more likely to be to olfactory rather than visual cues.
Biotic factors such as the density of individuals on the natal host, and the quality of the remaining host resources are also likely factors that influence whether an adult will fly and disperse from the host it emerged from. Adult A. glabripennis disperse in response to the number of beetles that emerge from a tree. The increasing abundance of adults leads to greater migration although they only move a short distance (Bancroft & Smith, 2005).
The Sawyer (2007) study of A. glabripennis outbreaks in New Jersey (USA) concluded that there were a few beetles that first established; they remained localised for many years and spread slowly until at some point, the local resources becomes overexploited at which time dispersal increased from a few hundred meters to over 1 km.
If S. tridentata is able to utilise European Ulmus species, Acer or Populus as hosts, something similar could be expected for S. tridentata in the EU although the likelihood that such trees could become hosts remains highly uncertain.
Spatial separation of European elm trees in Northern Italy
The scarce and ambiguous information regarding the host range of S. tridentata hampered firm conclusions on the criteria of the pest categorisation regarding establishment, spread and impact. Nevertheless, arising from the search for scientific or anecdotal evidence about host range, other data were deemed useful to improve the assessment once host range (in EU) was determined.
The observational evidence regarding entry (see Section 3.4.2 in main document) refers to annual interceptions of immature stages of the insect in imported wood through the port of Livorno, Italy. All S. tridentata infested wood imports originated from the USA were associated with round or sawn wood of American elm (Ulmus sp.). The Panel is uncertain whether the concentration of interceptions at only one port in EU is the matter of fact or due to improved testing procedures at the Livorno location.
It was deemed informative to consider the spatial separation of elm species in the near vicinity of the Italian ports and the distance profile of elm trees on greater spatial scales. The first proxies the capacity of the regional elm population to provide accessible habitat around Italian ports, while the second provides an indication whether mature beetles translocated to a random elm tree in the area (e.g. anthropogenically mediated) are likely reaching secondary elm trees for future establishment of successive generations by natural flight distance (dispersal mobility).
However, the value of the following considerations remains uncertain because dependent on the unknown host range of S. tridentata in the EU possibly anywhere between only American Ulmus and any Acer + Populus trees (see conclusions of the pest categorisation in Table 13).
Stand data of elm trees (Ulmus sp)
Northern Italy provides the only example region where S. tridentata interceptions are recorded at the port of Livorno (Tuscany Region, Italy), and (at least patchy) retrieval of elm tree stand distributions was possible. For the purposes of these considerations, EU species of elm were assumed to provide a more plausible theoretical host range compared to Acer sp. and/or Populus sp. while the actual host range of the beetle in EU remains unknown.
EFSA did combine independent data sources to compile the stand distribution shown in Figure E.1. The data sources are heterogeneous regarding geographical coverage, absence reporting and tree size included
Figure E.1.
Data on observed presences and absences of elm trees in Northern Italy. The information compiles different data sources as coloured. Neither of the data sources is deemed comprehensive regarding area coverage or observed absences. Additionally, the stem diameter of recognised tree presences is not equal across data sources and might comprise small trees not representing habitat for reproduction of the beetle (establishment). Symbols related to the Italian Forest Inventory represent the south‐west corners of 1 km × 1 km grid cells including (red circles) or not (grey crosses) Ulmus plants. Squares indicate centroids of polygons (Emilia Romagna, Lombardy, Piedmont) or grid cells (Tuscany) with records of the presence of Ulmus plants. Yellow triangles represent Ulmus plants inspected within the Tuscany phytosanitary monitoring program
Distance spread from the point of entry
Usual flight distances reported for cerambycids in laboratory and field studies were around a few hundred meters. In situations of host scarcity, however, mobility distance was reported to increase substantially. Extreme flight distances reported for cerambycid species suggest capacity under host limitations of up to 3 km (see above): 3,200 m over 5 years; more than 2 km; up to 2,600 m; over 1 km; laboratory 13,667 m).
The following map visually combines the tree stand data of Figure E.2 with the hypothetical mobility range of the elm borer. Concentric rings illustrate the chance of reaching an elm tree if a mature beetle starts from either port in the mapped region. Whether finding the tree allows the beetle to reproduce remains uncertain due to unknown host range of S. tridentata in the EU.
Figure E.2.
Panel A: Spatial distribution of elm trees (Ulmus sp.) of unknown stem diameter in the vicinity of Northern Italian ports of La Spezia and Livorno (blue stars) being locations of plausible pest interception. Different assumptions of flight capability are represented by radial distance of 1 km (red area), 3 km (orange area) and 10 km (green area) to the port of La Spezia (left top) and Livorno (middle bottom). Additionally, in Panel B, secondary distances are encircled by dotted lines representing the available host for further spread once established on any elm in the surroundings of the two ports. Symbols related to the Italian Forest Inventory represent the south‐west corners of 1 km x 1 km grid cells including (red circles) or not (grey crosses) Ulmus plants. Squares indicate centroids of polygons (Emilia Romagna) or grid cells (Tuscany) with records of the presence of Ulmus plants. Triangles represent Ulmus plants inspected within the Tuscany phytosanitary monitoring programme
If theoretically S. tridentata can utilise European Ulmus species as host, the maps reveal repeated extreme dispersal flights are required following entry to reach first trees and subsequent spatial expansion. However, with Acer or Populus as hosts, ubiquitous availability would alter the picture, although the likelihood that such trees could become hosts in the EU remains uncertain.
Distance spread from a random location of establishment
Neat scenario assumes that the mature insect was translocated to a random elm tree in Northern Italy (e.g. by transportation of infested wood out of a port). Allowing for natural spread the insect needs to search for and locate secondary tree stands for exploitation. The problem of interest is the expected distance a dispersing beetle might be confronted with when starting from its tree of larval development. Such patterns usually inform about colonisation potential of a species in an environment of patchy habitat. The shown distance distributions make the unreasonable assumption that the beetle could locate any near neighbour tree in a straight‐line flight. Insects, however, are deemed to search only locally using chemical cues while larger scale movement is driven by ecological pattern recognitions or chance search (Webster and Cardé, 2017). Hence, the distance between any elm tree and its nearest neighbour (next‐neighbour distance distribution) is an optimistic proxy for the capability of the beetles to colonise other trees (Figure E.3).
Figure E.3.
Nearest, second and third nearest neighbour distribution for the elm tree stands reported in the Italian Forest Inventory data from Northern Italy. Same data are presented as absolute frequencies (left column) and as cumulative frequencies (right column). The median of the distribution and the respective distance value are marked in red. In yellow, the respective 25 and 75 percentiles are highlighted
Maximising the geographic coverage of the analysis, only the data of the Italian forest inventory were used. The advantage of the data is the 1 km scale and the sample locations i.e. absence locations are of value. Substantial uncertainty remains whether the sample‐based approach inherently may exclude close neighbour patches i.e. trees in the distance range of up to 3 km which is the most interesting distance according to the flight distance data collected above.
About 47 of the 330 recorded grid cells containing Ulmus trees (first three frequency values) have a neighbouring grid cell within 3 km distance. To find a new tree with ~25% chance (lower yellow line), beetle should be able to cover flight distances up to 4,530 m assuming straight flight without deviation. Beetles may miss the ideal direction to the nearest tree (chance usually determined by the angle segment the beetle could search while moving away of the tree, unknown for this species). Starting on 12 (only one) grid cell beetles may still find a potential host, second (third) nearest neighbour, within 3 km distance if choosing the respective angle segment for the search.
If S. tridentata can utilise European Ulmus species, Acer and Populus as hosts in the EU, there would be widespread availability of trees to colonise. The actual possible host range in the EU, however, remains uncertain.
References
Bancroft JS, and Smith MT, 2005. Dispersal and influences on movement for Anoplophora glabripennis calculated from individual mark‐recapture. Entomologia Experimentalis et Applicata, 116, 83–92.
Favaro R, Wichmann L, Ravn HP and Faccoli M, 2015. Spatial spread and infestation risk assessment in the Asian longhorned beetle, Anoplophora glabripennis. Entomologia Experimentalis et Applicata, 155, 95–101.
Haack RA, Hérard F, Sun J and Turgeon JJ, 2010. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: a worldwide perspective. Annual Review of Entomology, 55, 521–546.
Hess AD, 1940. “The biology and control of the round‐headed apple‐tree borer, Saperda Candida Fabricius,” Geneva, NY.
Javal M, Roux G, Roques A, and Sauvard D, 2018. Asian Long‐horned Beetle dispersal potential estimated in computer‐linked flight mills. Journal of Applied Entomology, 142, 282–286.
Lopez VM, Hoddle MS, Francese JA, Lance DR, and Ray AM, 2017. Assessing Flight Potential of the Invasive Asian Longhorned Beetle (Coleoptera: Cerambycidae) With Computerized Flight Mills. Journal of Economic Entomology, 110, 1070–1077.
Pechuman LL, 1940. Notes on the feeding and breeding habits of Saperda tridentata Oliv. Bulletin of the Brooklyn Entomological Society, 35, 113–116.
Rhainds M, Mackinnon WE, Porter KB, Sweeney JD and Silk PJ, 2011. Evidence for limited spatial spread in an exotic longhorn beetle, Tetropium fuscum (Coleoptera: Cerambycidae). Journal of Economic Entomology, 104, 1928–1933.
Sawyer A, 2007. Infestation dynamics of the Asian Longhorned Beetle in the United States. Proceedings, 17th U.S. Department of Agriculture interagency research forum on gypsy moth and other invasive species 2006.
Sawyer A, 2008. “Asian longhorned beetle: annotated host list ‐ https://www.fs.fed.us/foresthealth/technology/pdfs/alb_hostlist.pdf.” USDA‐APHIS‐PPQ, Otis Pest Survey Detection & Exclusion Laboratory.
Sawyer AJ, Panagakos WS, Horner AE and Freeman KJ, 2011. Asian longhorned beetle, over the river and through the woods: habitat‐dependent population spread. In: McManus, Katherine A; Gottschalk, Kurt W., eds. 2010. Proceedings. 21st US Department of Agriculture interagency research forum on invasive species 2010; 2010 January 12‐15; Annapolis, MD. Gen. Tech. Rep. NRS‐P‐75. Newtown Square, PA: US Department of Agriculture, Forest Service, Northern Research Station: 52‐54. 2011 (pp. 52–54).
Smith MT, Bancroft J, Li G, Gao R, and Teale S, 2001. Dispersal of Anoplophora glabripennis (Cerambycidae). Environmental Entomology, 30, 1036–1040.
Smith MT, Tobin PC, Bancroft J, Li G, and Gao R, 2004. Dispersal and Spatiotemporal Dynamics of Asian Longhorned Beetle (Coleoptera: Cerambycidae) in China. Environmental Entomology, 33, 435–442.
Trotter RT, and Hull‐Sanders HM, 2015. Quantifying Dispersal of the Asian longhorned beetle (Anoplophora glabripennis, Coleoptera) with incomplete data and behavioral knowledge. Biological Invasions, 17, 3359–3369.
Webster B, and Cardé RT, 2017. Use of habitat odour by host‐seeking insects. Biological Reviews, 92, 1241–1249.
Appendix F – Ulmus logs imports in Europe
1.
Member States were contacted by EFSA to have data about import of Ulmus logs in the respective countries from 2016 to 2019.
Table F.1.
Ulmus logs Import data from North America to European countries from 2016 to 2019. Derived variable was calculated using as a reference the density of U. rubra wood: 600 kg m−3 (source: https://www.wood-database.com/)
| Country | Port | Year | Imported | Inspected | Derived variable | ||||
|---|---|---|---|---|---|---|---|---|---|
| Containers (#) | Weight (tonnes) | Volume (m3) | Containers (#) | Weight (tonnes) | Volume (m3) | ||||
| Italy | Livorno | 2016 | 8 | 61.63 | 102.71 | 4 | 45.17 | 75.29 | weight |
| Italy | Livorno | 2017 | 16 | 113.57 | 189.29 | 14 | 106.40 | 177.33 | weight |
| Italy | Livorno | 2018 | 5 | 40.63 | 67.71 | 3 | 31.27 | 52.12 | weight |
| Italy | Livorno | 2019 | 1 | 10.83 | 18.05 | 1 | 10.83 | 18.05 | weight |
| Italy | La Spezia | 2016 | 4 | 71.00 | 118.00 | 4 | 4.00 | 118.00 | volume |
| Italy | La Spezia | 2017 | 0 | 0.00 | 0.00 | 0 | 0.00 | 0.00 | volume |
| Italy | La Spezia | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | volume |
| Italy | La Spezia | 2019 | 7.00 | 132.00 | 220.00 | 7.00 | 132.00 | 220.00 | volume |
| Italy | Napoli | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Napoli | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Napoli | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Napoli | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Venezia | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Venezia | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Venezia | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Italy | Venezia | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Estonia | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Estonia | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Estonia | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Estonia | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Germany | na | 2016 | na | 37.67 | 62.78 | na | 37.67 | 62.78 | weight |
| Germany | na | 2017 | na | 61.54 | 102.57 | na | 51.63 | 86.06 | weight |
| Germany | na | 2018 | na | 30.88 | 51.47 | na | 30.35 | 50.58 | weight |
| Germany | na | 2019 | na | 26.10 | 43.51 | na | 26.10 | 43.51 | weight |
| Spain | Barcelona | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Barcelona | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Barcelona | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Barcelona | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Valencia | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Valencia | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Valencia | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Valencia | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Bilbao | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Bilbao | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Bilbao | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Spain | Bilbao | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Finland | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Finland | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Finland | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Finland | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Portugal | Leixões | 2016 | 0.00 | 0.00 | 0.00 | na | na | na | – |
| Portugal | Leixões | 2017 | 0.00 | 0.00 | 0.00 | na | na | na | – |
| Portugal | Leixões | 2018 | 0.00 | 4.65 | 7.76 | na | na | na | volume |
| Portugal | Leixões | 2019 | 0.00 | 0.00 | 0.00 | na | na | na | – |
| Latvia | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Latvia | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Latvia | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Latvia | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Cyprus | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Cyprus | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Cyprus | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Cyprus | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Lithuania | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Lithuania | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Lithuania | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Lithuania | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Croatia | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Croatia | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Croatia | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Croatia | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| UK | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| UK | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| UK | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| UK | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Denmark | na | 2016 | na | 97.80 | 163.00 | na | 97.80 | 163.00 | weight |
| Denmark | na | 2017 | na | 7827.00 | 13045.00 | na | 7827.00 | 13045.00 | weight |
| Denmark | na | 2018 | na | 31.80 | 53.00 | na | 31.80 | 53.00 | weight |
| Denmark | na | 2019 | na | 2.40 | 4.00 | na | 2.40 | 4.00 | weight |
| The Netherlands | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| The Netherlands | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| The Netherlands | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| The Netherlands | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| France | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| France | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| France | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| France | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Sweden | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Sweden | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Sweden | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Sweden | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Poland | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Poland | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Poland | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Poland | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Malta | na | 2016 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Malta | na | 2017 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Malta | na | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
| Malta | na | 2019 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | – |
na: not available
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard C, Dehnen‐Schmutz K, Di Serio F, Gonthier P, Jacques M‐A, Jaques Miret JA, Justesen AF, MacLeod A, Magnusson CS, Milonas P, Navas‐Cortés JA, Parnell S, Potting R, Reignault PL, Van der Werf W, Civera AV, Yuen J, Zappalà L, Evans H, Maiorano A and Thulke H‐H, 2020. Scientific Opinion on the pest categorisation of Saperda tridentata . EFSA Journal 2020;18(1):5940, 59 pp. 10.2903/j.efsa.2020.5940
Requestor: European Commission
Question number: EFSA‐Q‐2019‐00170
Panel members: Claude Bragard, Katharina Dehnen‐Schmutz, Francesco Di Serio, Paolo Gonthier, Marie‐Agnès Jacques, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A Navas‐Cortés, Stephen Parnell, Roel Potting, Philippe L. Reignault, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent, Jonathan Yuen and Lucia Zappalà.
Acknowledgements: The Panel thanks for the information and/or data provided to this scientific output: Walter Baruzzo (Regione Liguria Phytosanitary Service, Italy), Martin Damus (Canadian Food Inspection Agency), Dalia Del Nista (Regione Toscana Phytosanitary Service, Italy), Massimo Fiorini (Federlegnoarredo, Italy), Gianni Franchini (Regione Toscana Phytosanitary Service, Italy), Patrizia Gasparini (CREA‐Research Centre for Forestry and Wood, Trento, Italy), Diego Guidotti (Aedit s.r.l., Italy), Robert Haack (USDA, Forest Service Northern Research Station, US), Paolo Marseglia (Regione Toscana Phytosanitary Service, Italy), Christian Morasso (Morasso Consulenze & Perizie), Guelfi Moreno (Regione Liguria Phytosanitary Service, Italy), Davide Paradiso (Conlegno Consortium, Italy), Marco Pattuelli (Regione Emilia Romagna, Italy), Fabrizio Pennacchio (CREA‐DC, Florence, Italy), Domenico Rizzo (Regione Toscana Phytosanitary Service, Italy), Suzanne Sharrock (Botanic Gardens Conservation International), Eduardo Ucciero (Regione Campania Phytosanitary Service, Italy), Marco Vettorazzo (Regione Veneto Phytosanitary Service, Italy). EFSA acknowledges all European competent institutions, Member State bodies and other organisations that provided data for this scientific output.
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder: Figure 1: © EPPO
Adopted: 20 November 2019
Notes
Council Directive 2000/29/EC of 8 May 2000 on protective measures against the introduction into the Community of organisms harmful to plants or plant products and against their spread within the Community. OJ L 169/1, 10.7.2000, p. 1–112.
Regulation (EU) 2016/2031 of the European Parliament of the Council of 26 October 2016 on protective measures against pests of plants, amending Regulations (EU) 228/2013, (EU) 652/2014 and (EU) 1143/2014 of the European Parliament and of the Council and repealing Council Directives 69/464/EEC, 74/647/EEC, 93/85/EEC, 98/57/EC, 2000/29/EC, 2006/91/EC and 2007/33/EC. OJ L 317, 23.11.2016, pp. 4–104.
References
- Allen EA and Humble LM, 2002. Nonindigenous species introductions: a threat to Canada's forests and forest economy1. Canadian Journal of Plant Pathology, 24, 103–110. [Google Scholar]
- Baker WC, 1941. Type of wood preferred by Coleoptera commonly found in decadent parts of living elm trees. Journal of Economic Entomology, 34, 475–476. [Google Scholar]
- Bezark LG, 2016. Checklist of the Oxypeltidae, Vesperidae, Disteniidae and Cerambycidae, (Coleoptera) of the Western Hemisphere(https://www.zin.ru/Animalia/Coleoptera/rus/whemis16.htm). pp. 497.
- Boberg J and Björklund N, 2018. “Saperda tridentata ‐ risks associated with trade of wood of Ulmus spp”. Swedish University of Agricultural Sciences ‐ Unit for Risk Assessment of Plant Pests.
- Bosik JJ, 1997. Common Names of Insects and Related Organisms. Entomological Society of America, Lanham, Maryland, US. [Google Scholar]
- Bousquet Y, Bouchard P, Davies A and Sikes D, 2013. Checklist of Beetles (Coleoptera) of Canada and Alaska. Second edition. ZooKeys. p. 360 [DOI] [PMC free article] [PubMed]
- Campbell J, Sarazin M and Lyons D, 1989. Canadian Beetles (Coleoptera) Injurious to Crops, Ornamentals, Stored Products, and Buildings. Agriculture Canada, Ottawa, Ontario. [Google Scholar]
- Cassens DL, 2007. Hardwood lumber and Veneer Series: Elm. Purdue University Extension, FNR‐283‐W.
- Caudullo G. and de Rigo D, 2016. Ulmus ‐ elms in Europe: distribution, habitat, usage and threats. In “European Atlas of Forest Tree Species” (San‐Miguel‐Ayanz J, de Rigo D, Caudullo G, T. Houston Durrant and A. Mauri, eds.). Publ. Off. EU, Luxembourg.
- Craighead FC, 1923. North American Cerambycid Larvae: A Classification and the Biology of North American Cerambycid Larvae. Dept. of Agriculuture, Ottawa. [Google Scholar]
- Drooz A, 1985. Insects of eastern forests, US Department of Agriculture, Forest Service.
- EFSA PLH Panel (EFSA Panel on Plant Health), Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Grégoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Hart A, Schans J, Schrader G, Suffert M, Kertész V, Kozelska S, Mannino MR, Mosbach‐Schulz O, Pautasso M, Stancanelli G, Tramontini S, Vos S and Gilioli G, 2018. Guidance on quantitative pest risk assessment. EFSA Journal 2018;16(8):e05350 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrlén J and Morris WF, 2015. Predicting changes in the distribution and abundance of species under environmental change. Ecology Letters, 18, 303–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrmann E, 1897. Coleopterological notes from my brother's diary. Entomological News, and Proceedings of the Entomological Section of the Academy of Natural Sciences of Philadelphia., 8, 168–170. [Google Scholar]
- EPPO (European and Mediterranean Plant Protection Organization), online. EPPO Global Database. Available online: https://www.eppo.int/ [Accessed: 7 November 2019].
- FAO (Food and Agriculture Organization of the United Nations), 1995. ISPM (International standards for phytosanitary measures) No 4. Requirements for the establishment of pest free areas. Available online: https://www.ippc.int/en/publications/614/
- FAO (Food and Agriculture Organization of the United Nations), 2004. Pest risk analysis for regulated non‐quarantine pests. In “International standards for phytosanitary measures” (FAO, ed.), Rome, Italy.
- FAO (Food and Agriculture Organization of the United Nations), 2014. ISPM No. 11, Pest risk analysis for quarantine pests. In “International Standards for Phytosanitary Measures”. FAO, Rome, Italy.
- FAO (Food and Agriculture Organization of the United Nations), 2017. ISPM (International standards for phytosanitary measures) No 5. Glossary of phytosanitary terms. Available online: https://www.ippc.int/en/publications/622/
- Felt EM and Joutel L, 1904. Monograph of the Genus Saperda. New York State Education Dept, Albany. [Google Scholar]
- Gosling DCL and Gosling NM, 1977. An annotated list of the cerambycidae of Michigan (Coleoptera) Part II, the Subfamilies Lepturinae and Lamiinae. The Great Lakes. Entomologist, 10. [Google Scholar]
- Haldemann , 1847. Corrections and additions to his paper on the Longicornia of the United States. Proceedings of the American Philosophical Society, 4, 371–376. [Google Scholar]
- Hanks LM, 1999. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annual Review of Entomology, 44, 483–505. [DOI] [PubMed] [Google Scholar]
- Harrington WH, 1883. Correspondence. The Canadian Entomologist., 15, 79–80. [Google Scholar]
- Heffern DJ, 1998. “Insects of Western North America 1. A survey of the Cerambycidae (Coleoptera) or longhorned beetles of Colorado”.
- Hoffmann CH, 1942. Annotated List of elm Insects in the United States. U.S. Dept. of Agriculture, Washington, D.C.. [Google Scholar]
- Kottek M, Grieser J, Beck C, Rudolf B and Rubel F, 2006. World Map of the Köppen‐Geiger climate classification updated. Meteorologische Zeitschrift, 15, 259–263. [Google Scholar]
- Krischik V and Davidson J, 2013. In: Krischik V. and Davidson J. (eds). IPM (Integrated Pest Management) of Midwest Landscapes. Agricultural Experiment Station, Minnesota: pp. 115–116. [Google Scholar]
- Lugger O, 1899. Beetles injurious to fruit‐producing plants. Bulletin of the Agricultural Experiment Station of the University of Minnesota, 66.
- Luppold WG and Thomas RE, 1991. New estimates of hardwood lumber exports to Europe and Asia. USDA Forest Service, Research Paper NE‐652.
- MacLeod A and Korycinska A, 2019. Detailing Köppen–Geiger climate zones at sub‐national to continental scale: a resource for pest risk analysis. EPPO Bulletin, 49, 73–82. [Google Scholar]
- MacRae TC, 1993. Annotated checklist of the longhorned beetles (Coleoptera: Cerambycidae and Disteniidae) occurring in Missouri. Insecta Mundi, 7, 223–252. [Google Scholar]
- Main J, 1839. The Forest Planter and Pruner's Assistant: Being a Practical Treatise on the Management of the Native and Exotic Forest Trees Commonly Cultivated in Great Britain. Ridgway, Piccadilly, London. [Google Scholar]
- Metcalf RL, Metcalf CL and Flint WF, 1951. “Destructive and useful Insects: their habitats and control,” Third/Ed.
- Olivier GA, 1795. “Entomologie ou Histoire naturelle des insectes, avec leurs caractères génériques et spécifiques, leur description, leur synonumie et leur figure enluminee… Coléoptères Tomes I‐V,” Imp. Baudouin, Paris.
- Packard AS, 1890. Fifth Report of the United States Entomological Commission: Being a Revised and Enlarged Edition of Bulletin no. 7, on Insects Injurious to Forest and Shade Trees, G.P.O., Washington.
- Park O, 1931. Studies in the Ecology of Forest Coleoptera. Ecology, 12, 188–207. [Google Scholar]
- Pechuman LL, 1940. Notes on the feeding and breeding habits of Saperda tridentata Oliv. Bulletin of the Brooklyn Entomological Society, 35, 113–116. [Google Scholar]
- Perkins GH, 1890. Insects Injurious to the American elm. Argus, Montpelier. [Google Scholar]
- Rice ME, Merickel F and MacRae TC, 2017. The longhorned beetles (coleoptera: Cerambycidae) of Idaho. Coleopterists Bulletin, 71, 667–678. [Google Scholar]
- Solomon JD, 1995. Guide to Insect Borers in North American Broadleaf Trees and Shrubs. Service Agriculture Handbook AH‐706. USDA, Wasghington, D.C. [Google Scholar]
- Solomon JD, Newsome L and Darwin WN, 1972. Carpenterworm moths and cerambycid hardwood borers caught in light traps. Journal of the Georgia Entomological Society, 7, 76–79. [Google Scholar]
- Sutherst RW and Maywald GF, 1985. A computerised system for matching climates in ecology. Agriculture, Ecosystems & Environment, 13, 281–299. [Google Scholar]
- Trewartha GT and Horn LH, 1980. “An Introduction to Climate”.
- Tucker ES, 1907. Concerning Some Insects Collected and Bred from Dead and Dying Elm. Transactions of the Kansas Academy of Science (1903‐), 21, 158–162. [Google Scholar]
- Washburn F, 1910. Thirteenth Report of the State Entomologist of Minnesota to the Governor for the Years 1909 and 1910. Agricultural Experiment Station, St. Anthony Park, Minn. [Google Scholar]
- White J and Moore D, 2003. Trees of Britain and Northern Europe. Orion Pub Co, Cassell, London. [Google Scholar]