

Abstract
The Panel on Plant Health performed a pest categorisation of non‐EU Scolytinae (Coleoptera: Curculionidae) of coniferous hosts (hereafter NESC). NESC occur worldwide, and some species are important forest pests. Species can be identified using taxonomic keys and molecular methods. Most NESC species (bark beetles) live in the inner bark of their hosts (phloem and cambium), while the remaining species mostly colonise the sapwood (ambrosia beetles). Bark‐ and ambrosia beetles are often associated with symbiotic fungi, which behave as pathogens towards the host trees, or are used as food by ambrosia beetle larvae. The larvae live in individual tunnels or in communal chambers. Pupation occurs in the wood or in the bark. Some species are semi‐ or multivoltine, others are monovoltine. Some species attack and kill living, apparently healthy trees. Other species specialise in weakened, dying or dead trees. The pathways for entry are cut branches, cones, round wood with or without bark, sawn wood with or without bark, wood packaging material, bark, manufactured wood items and wood chips and plants for planting (including seeds) of conifers. Availability of host plants and suitable climate would allow the establishment in the EU of NESC. Measures are in place to prevent their introduction through the pathways described above. NESC satisfy all the criteria to be considered as Union quarantine pests. As NESC are not present in the EU and plants for planting are not their major pathway for spread, they do not meet the criteria to be considered as regulated non‐quarantine pests.
Keywords: Bark beetles, ambrosia beetles, European Union, pest risk, plant health, plant pest, quarantine
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
1.1.1. Background
Council Directive 2000/29/EC1 on protective measures against the introduction into the Community of organisms harmful to plants or plant products and against their spread within the Community establishes the present European Union plant health regime. The Directive lays down the phytosanitary provisions and the control checks to be carried out at the place of origin on plants and plant products destined for the Union or to be moved within the Union. In the Directive's 2000/29/EC annexes, the list of harmful organisms (pests) whose introduction into or spread within the Union is prohibited, is detailed together with specific requirements for import or internal movement.
Following the evaluation of the plant health regime, the new basic plant health law, Regulation (EU) 2016/20312 on protective measures against pests of plants, was adopted on 26 October 2016 and will apply from 14 December 2019 onwards, repealing Directive 2000/29/EC. In line with the principles of the above mentioned legislation and the follow‐up work of the secondary legislation for the listing of EU regulated pests, EFSA is requested to provide pest categorisations of the harmful organisms included in the annexes of Directive 2000/29/EC, in the cases where recent pest risk assessment/pest categorisation is not available.
1.1.2. Terms of Reference
EFSA is requested, pursuant to Article 22(5.b) and Article 29(1) of Regulation (EC) No 178/20023, to provide scientific opinion in the field of plant health.
EFSA is requested to prepare and deliver a pest categorisation (step 1 analysis) for each of the regulated pests included in the appendices of the annex to this mandate. The methodology and template of pest categorisation have already been developed in past mandates for the organisms listed in Annex II Part A Section II of Directive 2000/29/EC. The same methodology and outcome is expected for this work as well.
The list of the harmful organisms included in the annex to this mandate comprises 133 harmful organisms or groups. A pest categorisation is expected for these 133 pests or groups and the delivery of the work would be stepwise at regular intervals through the year as detailed below. First priority covers the harmful organisms included in Appendix 1, comprising pests from Annex II Part A Section I and Annex II Part B of Directive 2000/29/EC. The delivery of all pest categorisations for the pests included in Appendix 1 is June 2018. The second priority is the pests included in Appendix 2, comprising the group of Cicadellidae (non‐EU) known to be vector of Pierce's disease (caused by Xylella fastidiosa), the group of Tephritidae (non‐EU), the group of potato viruses and virus‐like organisms, the group of viruses and virus‐like organisms of Cydonia Mill., Fragaria L., Malus Mill., Prunus L., Pyrus L., Ribes L., Rubus L. and Vitis L., and the group of Margarodes (non‐EU species). The delivery of all pest categorisations for the pests included in Appendix 2 is end 2019. The pests included in Appendix 3 cover pests of Annex I part A section I and all pest categorisations should be delivered by end 2020.
For the above mentioned groups, each covering a large number of pests, the pest categorisation will be performed for the group and not the individual harmful organisms listed under “such as” notation in the Annexes of the Directive 2000/29/EC. The criteria to be taken particularly under consideration for these cases, is the analysis of host pest combination, investigation of pathways, the damages occurring and the relevant impact.
Finally, as indicated in the text above, all references to ‘non‐European’ should be avoided and replaced by ‘non‐EU’ and refer to all territories with exception of the Union territories as defined in Article 1 point 3 of Regulation (EU) 2016/2031.
1.1.2.1. Terms of Reference: Appendix 1
List of harmful organisms for which pest categorisation is requested. The list below follows the annexes of Directive 2000/29/EC.
| Annex IIAI | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Aleurocanthus spp. | Numonia pyrivorella (Matsumura) |
| Anthonomus bisignifer (Schenkling) | Oligonychus perditus Pritchard and Baker |
| Anthonomus signatus (Say) | Pissodes spp. (non‐EU) |
| Aschistonyx eppoi Inouye | Scirtothrips aurantii Faure |
| Carposina niponensis Walsingham | Scirtothrips citri (Moultex) |
| Enarmonia packardi (Zeller) | Scolytidae spp. (non‐EU) |
| Enarmonia prunivora Walsh | Scrobipalpopsis solanivora Povolny |
| Grapholita inopinata Heinrich | Tachypterellus quadrigibbus Say |
| Hishomonus phycitis | Toxoptera citricida Kirk. |
| Leucaspis japonica Ckll. | Unaspis citri Comstock |
| Listronotus bonariensis (Kuschel) | |
| (b) Bacteria | |
| Citrus variegated chlorosis | Xanthomonas campestris pv. oryzae (Ishiyama) Dye and pv. oryzicola (Fang. et al.) Dye |
| Erwinia stewartii (Smith) Dye | |
| (c) Fungi | |
| Alternaria alternata (Fr.) Keissler (non‐EU pathogenic isolates) | Elsinoe spp. Bitanc. and Jenk. Mendes |
| Anisogramma anomala (Peck) E. Müller | Fusarium oxysporum f. sp. albedinis (Kilian and Maire) Gordon |
| Apiosporina morbosa (Schwein.) v. Arx | Guignardia piricola (Nosa) Yamamoto |
| Ceratocystis virescens (Davidson) Moreau | Puccinia pittieriana Hennings |
| Cercoseptoria pini‐densiflorae (Hori and Nambu) Deighton | Stegophora ulmea (Schweinitz: Fries) Sydow & Sydow |
| Cercospora angolensis Carv. and Mendes | Venturia nashicola Tanaka and Yamamoto |
| (d) Virus and virus‐like organisms | |
| Beet curly top virus (non‐EU isolates) | Citrus tristeza virus (non‐EU isolates) |
| Black raspberry latent virus | Leprosis |
| Blight and blight‐like | Little cherry pathogen (non‐ EU isolates) |
| Cadang‐Cadang viroid | Naturally spreading psorosis |
| Tatter leaf virus | Palm lethal yellowing mycoplasm |
| Witches’ broom (MLO) | Satsuma dwarf virus |
| Annex IIB | |
| (a) Insect mites and nematodes, at all stages of their development | |
| Anthonomus grandis (Boh.) | Ips cembrae Heer |
| Cephalcia lariciphila (Klug) | Ips duplicatus Sahlberg |
| Dendroctonus micans Kugelan | Ips sexdentatus Börner |
| Gilphinia hercyniae (Hartig) | Ips typographus Heer |
| Gonipterus scutellatus Gyll. | Sternochetus mangiferae Fabricius |
| Ips amitinus Eichhof | |
| (b) Bacteria | |
| Curtobacterium flaccumfaciens pv. flaccumfaciens (Hedges) Collins and Jones | |
| (c) Fungi | |
| Glomerella gossypii Edgerton | Hypoxylon mammatum (Wahl.) J. Miller |
| Gremmeniella abietina (Lag.) Morelet | |
1.1.2.2. Terms of Reference: Appendix 2
List of harmful organisms for which pest categorisation is requested per group. The list below follows the categorisation included in the annexes of Directive 2000/29/EC.
| Annex IAI | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Group of Cicadellidae (non‐EU) known to be vector of Pierce's disease (caused by Xylella fastidiosa), such as: | |
| 1) Carneocephala fulgida Nottingham | 3) Graphocephala atropunctata (Signoret) |
| 2) Draeculacephala minerva Ball | |
| Group of Tephritidae (non‐EU) such as: | |
| 1) Anastrepha fraterculus (Wiedemann) | 12) Pardalaspis cyanescens Bezzi |
| 2) Anastrepha ludens (Loew) | 13) Pardalaspis quinaria Bezzi |
| 3) Anastrepha obliqua Macquart | 14) Pterandrus rosa (Karsch) |
| 4) Anastrepha suspensa (Loew) | 15) Rhacochlaena japonica Ito |
| 5) Dacus ciliatus Loew | 17) Rhagoletis fausta (Osten‐Sacken) |
| 6) Dacus curcurbitae Coquillet | 18) Rhagoletis indifferens Curran |
| 7) Dacus dorsalis Hendel | 19) Rhagoletis mendax Curran |
| 8) Dacus tryoni (Froggatt) | 20) Rhagoletis pomonella Walsh |
| 9) Dacus tsuneonis Miyake | 21) Rhagoletis suavis (Loew) |
| 10) Dacus zonatus Saund. | |
| 11) Epochra canadensis (Loew) | |
| (c) Viruses and virus‐like organisms | |
| Group of potato viruses and virus‐like organisms such as: | |
| 1) Andean potato latent virus | 4) Potato black ringspot virus |
| 2) Andean potato mottle virus | 5) Potato virus T |
| 3) Arracacha virus B, oca strain | 6) non‐EU isolates of potato viruses A, M, S, V, X and Y (including Yo, Yn and Yc) and Potato leafroll virus |
| Group of viruses and virus‐like organisms of Cydonia Mill., Fragaria L., Malus Mill., Prunus L., Pyrus L., Ribes L., Rubus L. and Vitis L., such as: | |
| 1) Blueberry leaf mottle virus | 8) Peach yellows mycoplasm |
| 2) Cherry rasp leaf virus (American) | 9) Plum line pattern virus (American) |
| 3) Peach mosaic virus (American) | 10) Raspberry leaf curl virus (American) |
| 4) Peach phony rickettsia | 11) Strawberry witches’ broom mycoplasma |
| 5) Peach rosette mosaic virus | 12) Non‐EU viruses and virus‐like organisms of Cydonia Mill., Fragaria L., Malus Mill., Prunus L., Pyrus L., Ribes L., Rubus L. and Vitis L. |
| 6) Peach rosette mycoplasm | |
| 7) Peach X‐disease mycoplasm | |
| Annex IIAI | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Group of Margarodes (non‐EU species) such as: | |
| 1) Margarodes vitis (Phillipi) | 3) Margarodes prieskaensis Jakubski |
| 2) Margarodes vredendalensis de Klerk | |
1.1.2.3. Terms of Reference: Appendix 3
List of harmful organisms for which pest categorisation is requested. The list below follows the annexes of Directive 2000/29/EC.
| Annex IAI | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Acleris spp. (non‐EU) | Longidorus diadecturus Eveleigh and Allen |
| Amauromyza maculosa (Malloch) | Monochamus spp. (non‐EU) |
| Anomala orientalis Waterhouse | Myndus crudus Van Duzee |
| Arrhenodes minutus Drury | Nacobbus aberrans (Thorne) Thorne and Allen |
| Choristoneura spp. (non‐EU) | Naupactus leucoloma Boheman |
| Conotrachelus nenuphar (Herbst) | Premnotrypes spp. (non‐EU) |
| Dendrolimus sibiricus Tschetverikov | Pseudopityophthorus minutissimus (Zimmermann) |
| Diabrotica barberi Smith and Lawrence | Pseudopityophthorus pruinosus (Eichhoff) |
| Diabrotica undecimpunctata howardi Barber | Scaphoideus luteolus (Van Duzee) |
| Diabrotica undecimpunctata undecimpunctata Mannerheim | Spodoptera eridania (Cramer) |
| Diabrotica virgifera zeae Krysan & Smith | Spodoptera frugiperda (Smith) |
| Diaphorina citri Kuway | Spodoptera litura (Fabricus) |
| Heliothis zea (Boddie) | Thrips palmi Karny |
| Hirschmanniella spp., other than Hirschmanniella gracilis (de Man) Luc and Goodey | Xiphinema americanum Cobb sensu lato (non‐EU populations) |
| Liriomyza sativae Blanchard | Xiphinema californicum Lamberti and Bleve‐Zacheo |
| (b) Fungi | |
| Ceratocystis fagacearum (Bretz) Hunt | Guignardia laricina (Saw.) Yamamoto and Ito |
| Chrysomyxa arctostaphyli Dietel | Gymnosporangium spp. (non‐EU) |
| Cronartium spp. (non‐EU) | Inonotus weirii (Murril) Kotlaba and Pouzar |
| Endocronartium spp. (non‐EU) | Melampsora farlowii (Arthur) Davis |
| Mycosphaerella larici‐leptolepis Ito et al. | Septoria lycopersici Speg. var. malagutii Ciccarone and Boerema |
| Mycosphaerella populorum G. E. Thompson | Thecaphora solani Barrus |
| Phoma andina Turkensteen | Trechispora brinkmannii (Bresad.) Rogers |
| Phyllosticta solitaria Ell. and Ev. | |
| (c) Viruses and virus‐like organisms | |
| Tobacco ringspot virus | Pepper mild tigré virus |
| Tomato ringspot virus | Squash leaf curl virus |
| Bean golden mosaic virus | Euphorbia mosaic virus |
| Cowpea mild mottle virus | Florida tomato virus |
| Lettuce infectious yellows virus | |
| (d) Parasitic plants | |
| Arceuthobium spp. (non‐EU) | |
| Annex IAII | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Meloidogyne fallax Karssen | Rhizoecus hibisci Kawai and Takagi |
| Popillia japonica Newman | |
| (b) Bacteria | |
| Clavibacter michiganensis (Smith) Davis et al. ssp. sepedonicus (Spieckermann and Kotthoff) Davis et al. | Ralstonia solanacearum (Smith) Yabuuchi et al. |
| (c) Fungi | |
| Melampsora medusae Thümen | Synchytrium endobioticum (Schilbersky) Percival |
| Annex I B | |
| (a) Insects, mites and nematodes, at all stages of their development | |
| Leptinotarsa decemlineata Say | Liriomyza bryoniae (Kaltenbach) |
| (b) Viruses and virus‐like organisms | |
| Beet necrotic yellow vein virus | |
1.2. Interpretation of the Terms of Reference
Scolytinae spp. are listed as Scolytidae spp. (non‐EU) in the Appendices to the Terms of Reference (ToR) to be subject to pest categorisation to determine whether it fulfils the criteria of a quarantine pest or those of a regulated non‐quarantine pest (RNQP) for the area of the EU excluding Ceuta, Melilla and the outermost regions of Member States (MSs) referred to in Article 355(1) of the Treaty on the Functioning of the European Union (TFEU), other than Madeira and the Azores. Although the leading taxonomists in the 2000s (Wood, 1982; Bright and Skidmore, 2002) still considered the Scolytidae to be a family distinct from the Curculionidae according to morphological criteria, modern phylogenetics supports the position of scolytine beetles (Scolytinae) within the family Curculionidae (Knizek and Beaver, 2004; Hulcr et al., 2015). This is reflected by the growing number of citations in Scopus (2019) referring to Scolytinae (18 in 1990 vs. 210 in 2018), as opposed to citations referring to Scolytidae (50 in 1990 vs. 16 in 2018). The Scolytinae includes two subcategories, the ‘bark beetles’ which live in the phloem, and the ‘ambrosia beetles’ which live in the sapwood. This categorisation focused on non‐EU Scolytinae as defined in the opinion on the ‘List of non‐EU Scolytinae of coniferous hosts’ (EFSA PLH Panel, 2020). From the list of 705 non‐EU Scolytinae on coniferous hosts identified in Appendix A of the ‘List of non‐EU Scolytinae of coniferous hosts’, 222 species for which sufficient information regarding biology is available and which colonise conifer genera which are widely represented in Europe were selected as the basis for further pest categorisation. Further information on those 222 species can be found in Appendix B and in the supporting document which includes detailed data (link to the excel shortlist). The information about the feeding habits and host range of the remaining 483 species can be found in the supporting document (link to excel Full list).
2. Data and methodologies
2.1. Data
2.1.1. Literature search, data collection and selection of species for pest categorisation
The data for the current opinion are based on preparatory work (literature review and data collection) conducted by the University of Padova (Department of Agronomy, Food, Natural Resources, Animals and Environment) and on information provided by MSs on the distribution of Scolytinae species on territory. For further details we would like to refer the Scientific Opinion on the List of non‐EU Scolytinae of coniferous hosts where all data are published (EFSA PLH Panel, 2002).
2.2. Methodologies
The Panel performed the pest categorisation for Scolytinae spp., following guiding principles and steps presented in the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018) and in the International Standard for Phytosanitary Measures No 11 (FAO, 2013) and No 21 (FAO, 2004).
This work was initiated following an evaluation of the EU plant health regime. Therefore, to facilitate the decision‐making process, in the conclusions of the pest categorisation, the Panel addresses explicitly each criterion for a Union quarantine pest and for a Union RNQP in accordance with Regulation (EU) 2016/2031 on protective measures against pests of plants, and includes additional information required in accordance with the specific terms of reference received by the European Commission. In addition, for each conclusion, the Panel provides a short description of its associated uncertainty.
Table 1 presents the Regulation (EU) 2016/2031 pest categorisation criteria on which the Panel bases its conclusions. All relevant criteria have to be met for the pest to potentially qualify either as a quarantine pest or as a RNQP. If one of the criteria is not met, the pest will not qualify. A pest that does not qualify as a quarantine pest may still qualify as a RNQP that needs to be addressed in the opinion. For the pests regulated in the protected zones only, the scope of the categorisation is the territory of the protected zone; thus, the criteria refer to the protected zone instead of the EU territory.
Table 1.
Pest categorisation criteria under evaluation, as defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest | Criterion in Regulation (EU) 2016/2031 regarding protected zone quarantine pest (articles 32–35) | Criterion in Regulation (EU) 2016/2031 regarding Union regulated non‐quarantine pest |
|---|---|---|---|
| Identity of the pest (Section 3.1) | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? | Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible? |
| Absence/presence of the pest in the EU territory (Section 3.2) |
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU? Describe the pest distribution briefly! |
Is the pest present in the EU territory? If not, it cannot be a protected zone quarantine organism | Is the pest present in the EU territory? If not, it cannot be a RNQP. (A regulated non‐quarantine pest must be present in the risk assessment area) |
| Regulatory status (Section 3.3) | If the pest is present in the EU but not widely distributed in the risk assessment area, it should be under official control or expected to be under official control in the near future |
The protected zone system aligns with the pest free area system under the International Plant Protection Convention (IPPC) The pest satisfies the IPPC definition of a quarantine pest that is not present in the risk assessment area (i.e. protected zone) |
Is the pest regulated as a quarantine pest? If currently regulated as a quarantine pest, are there grounds to consider its status could be revoked? |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4) | Is the pest able to enter into, become established in, and spread within, the EU territory? If yes, briefly list the pathways! |
Is the pest able to enter into, become established in, and spread within, the protected zone areas? Is entry by natural spread from EU areas where the pest is present possible? |
Is spread mainly via specific plants for planting, rather than via natural spread or via movement of plant products or other objects? Clearly state if plants for planting is the main pathway! |
| Potential for consequences in the EU territory (Section 3.5) | Would the pests’ introduction have an economic or environmental impact on the EU territory? | Would the pests’ introduction have an economic or environmental impact on the protected zone areas? | Does the presence of the pest on plants for planting have an economic impact as regards the intended use of those plants for planting? |
| Available measures (Section 3.6) | Are there measures available to prevent the entry into, establishment within or spread of the pest within the EU such that the risk becomes mitigated? |
Are there measures available to prevent the entry into, establishment within or spread of the pest within the protected zone areas such that the risk becomes mitigated? Is it possible to eradicate the pest in a restricted area within 24 months (or a period longer than 24 months where the biology of the organism so justifies) after the presence of the pest was confirmed in the protected zone? |
Are there measures available to prevent pest presence on plants for planting such that the risk becomes mitigated? |
| Conclusion of pest categorisation (Section 4) | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential quarantine pest were met and (2) if not, which one(s) were not met | A statement as to whether (1) all criteria assessed by EFSA above for consideration as potential protected zone quarantine pest were met, and (2) if not, which one(s) were not met | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential RNQP were met, and (2) if not, which one(s) were not met |
It should be noted that the Panel's conclusions are formulated respecting its remit and particularly with regard to the principle of separation between risk assessment and risk management (EFSA founding regulation (EU) No 178/2002); therefore, instead of determining whether the pest is likely to have an unacceptable impact, the Panel will present a summary of the observed pest impacts. Economic impacts are expressed in terms of yield and quality losses and not in monetary terms, whereas addressing social impacts is outside the remit of the Panel.
The Panel will not indicate in its conclusions of the pest categorisation whether to continue the risk assessment process, but following the agreed two‐step approach, will continue only if requested by the risk managers. However, during the categorisation process, experts may identify key elements and knowledge gaps that could contribute significant uncertainty to a future assessment of risk. It would be useful to identify and highlight such gaps so that potential future requests can specifically target the major elements of uncertainty, perhaps suggesting specific scenarios to examine.
3. Pest categorisation
3.1. Identity and biology of the pest
3.1.1. Identity and taxonomy
3.1.1.1.
Is the identity of the pest established, or has it been shown to produce consistent symptoms and to be transmissible?
Yes, the identity of the pests is well established, and the non‐EU Scolytinae are described in worldwide, regional or national catalogues and faunae.
The non‐EU Scolytinae of coniferous hosts (hereafter NESC) are coleopteran insects belonging to the subfamily Scolytinae, a subgroup of the family Curculionidae (weevils). They have been fully listed in the world catalogue of (Wood and Bright, 1992) and in further publications (e.g. Bright and Skidmore, 2002; Bright, 2014), as well as in the online catalogue of Atkinson (2019) for species native to, or introduced in, North America. Keys and documentation have been published e.a. by Wood (1982, 2007), Gomez et al. (2018), respectively for North and Central America, and South America, and by (Schedl et al. (1981) for Central Europe.
3.1.2. Biology of the pest
A general introduction to the biology of bark‐ and ambrosia beetles is provided by Raffa et al. (2015) and Kirkendall et al. (2015). Out of the 705 NESC species, 580 species (bark beetles) are living in the inner bark of conifers (phloem and cambium) and often also slightly engraving the outer sapwood and 100 species are living in the sapwood. The remaining species are pith‐feeding (myelophagous) or seed‐feeding (spermatophagous). Although 15 of the sapwood‐inhabiting species are xylophagous (i.e. feeds directly on the wood), the other 85 species are xylomycetophagous, feeding on symbiotic fungi that they grow in the galleries or the chambers that they excavate in the wood. They are called ambrosia beetles, because of their association with ambrosia fungi, a vernacular name for several fungal genera associated with wood boring insects (Alamouti et al., 2009). Some species are polygamous (harem polygyny): the males leave their natal system, create a new gallery and attract one or several females. In the monogamous species, each female initiates a new gallery and attracts one male.
Out of the 222 NESC species for which detailed biological information is available, 194 species (86.9%) are outbreeding (mate outside of their family) and 29 species (13%) are inbreeding (the females mate with a brother and initiates alone a new colony). Furthermore, 22 of these inbreeding species (10.3%) are haplodiploid (in the absence of males, the female parthenogenetically produces a male and afterward mates with her son). This differs very much from the general proportions within the Scolytinae, with 1627 inbreeding species (Kirkendall et al., 2015), corresponding to 26.9% of the 6056 Scolytinae species known so far (Hulcr et al., 2015).
Some species (e.g. in the genera Dendroctonus and Ips) attack and kill living, apparently healthy trees. Other species specialise in weakened, dying or dead trees. Scolytinae are associated with various symbionts that play a role in nutrient acquisition by the insects, including pathogenic fungi which contribute to overwhelming the defences of living trees (Raffa et al., 2015; Hofstetter et al. (2015)). The galleries or the brood chambers vary in shape and size between the different species, often creating specific patterns. Each female excavates an egg gallery or an egg chamber. The eggs are either laid individually in niches along the gallery, or in batches along the gallery or in a chamber. The larvae develop either in individual galleries at the end of which they pupate, or gregariously in a common chamber. In some species, the young adults must proceed to maturation feeding, before or after emergence from the natal tree. In this latter case, they may feed on fresh bark tissues or on young twigs. Dispersal occurs by flight, except for the males of many inbreeding species, which do not fly but are sometimes able to walk into a neighbouring gallery in the same tree.
The chemical ecology of Scolytinae is very complex (Raffa et al., 2015). Some species only respond to primary attractants from their hosts, such as alpha‐pinene, and ethanol when the hosts are dying or dead and their tissues start to ferment. In addition to or instead of, primary attractants, many species produce aggregation pheromones that attract conspecifics of both sexes. This results in mass‐colonisation of the hosts.
3.1.3. Intraspecific diversity
Some subspecies have been identified (e.g. Pityophthorus buyssoni angeri; Pityophthorus buyssoni buyssoni).
3.1.4. Detection and identification of the pest
3.1.4.1.
Are detection and identification methods available for the pest?
Yes, non‐EU Scolytinae can be detected visually from external symptoms as well as from the shape of the galleries or brood chambers. Pheromones or other attractants are also available for detection for several species. Descriptions and illustrations are available for identification at species level. For some genera, molecular tools can also be used.
Bark and ambrosia beetles cause typical symptoms on the host plants including crown discoloration (yellow, red or grey), bark shedding, resin emission, white sawdust, brown sawdust, shoot tunnelling. They can also be detected by the shape of galleries or brood chambers.
They could also be detected using traps baited with pheromones or attractants originating from the host plants. Identification is based on taxonomic keys and descriptions and illustrations of the species are available. In some taxa, species can also be distinguished using molecular tools (Stauffer et al., 2001).
3.2. Pest distribution
3.2.1. Pest distribution outside the EU
Most of the 705 NESC occur in North America and Asia. In North America,401 species have been recorded and in Asia 222 (Figure 1). Moreover, 320 and 188 species are distributed exclusively in North America and Asia respectively (Figure 2). There are 102 species that occur in more than one continent, and one species (Xyleborus perforans) has a very wide distribution being recorded in 7 continents (Figure 3). Detailed information on distribution of these species and the corresponding references can be found in (EFSA PLH Panel, 2020).
Figure 1.
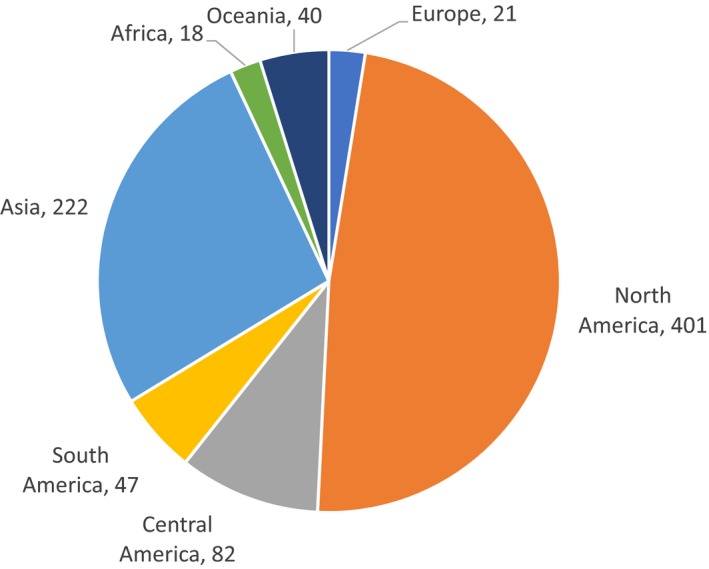
Number of non‐EU Scolytinae reported from each continent
Figure 2.
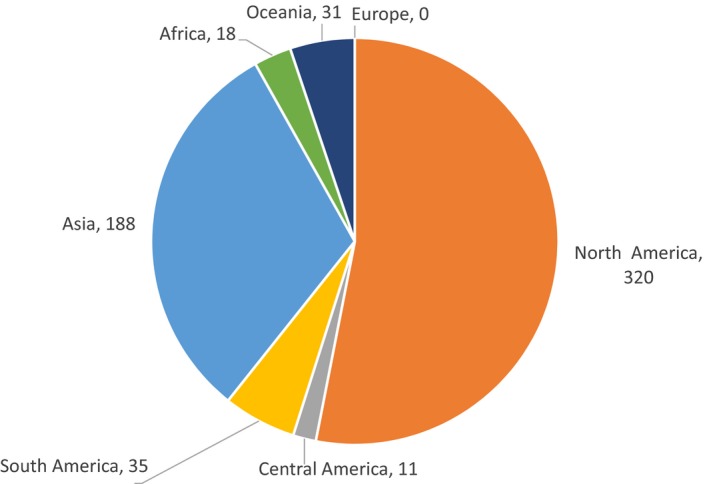
Number of species reported exclusively from one continent
Figure 3.
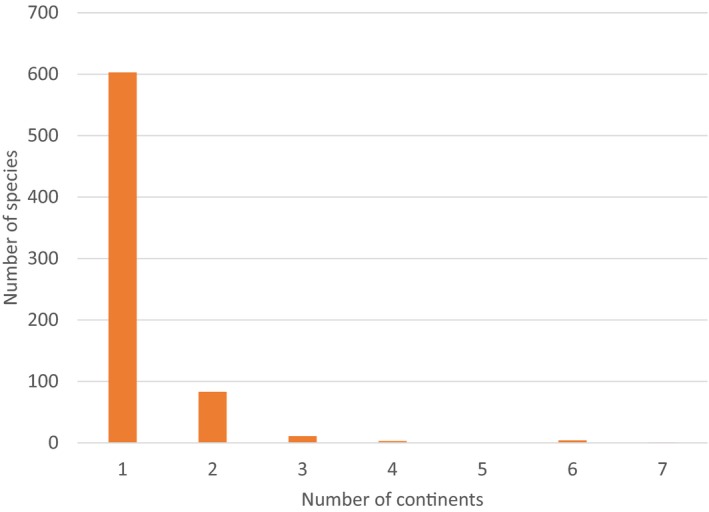
Number of species distributed in world continents
3.2.2. Pest distribution in the EU
3.2.2.1.
Is the pest present in the EU territory? If present, is the pest widely distributed within the EU?
From the total of 705 non‐EU Scolytinae there are 16 species that are present in the EU (in a few MSs).
There are 16 non‐EU Scolytinae species that are also present in EU (Table 2). These species have limited distribution in up to three EU MSs. Eleven species occur in one EU MS, three species in two and two in three EU MSs. All these species are also present in other countries outside of the EU.
Table 2.
Current distribution of non‐EU Scolytinae in the 28 EU MS based on information from the EPPO Global Database and other sources
| Species | Presence in EU MS | Presence outside EU | Comments/Uncertainties | Reference | |
|---|---|---|---|---|---|
| 1 | Carphoborus henscheli | Cyprus | Georgia, Turkey, Israel, Syria | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) | |
| 2 | Carphoborus marani | Greece, Hungary | European Russia | Presence in Hungary only mentioned once and never confirmed later (Milos Knizek, pers. comm.) | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) |
| 3 | Crypturgus dubius | France, Spain | Turkey, Iran | Presence in France and Spain is doubtful | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) |
| 4 | Cyrtogenius luteus | Italy | China, Japan, South Korea, South America | Introduced species | Atkinson (2019), Gomez et al. (2012) |
| 5 | Hylastes batnensis batnensis | Italy | Algeria, Morocco | Likely to be inroduced | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) |
| 6 | Orthotomicus tridentatus | Austria | Turkey | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) | |
| 7 | Phloeosinus armatus | Greece, Italy, Cyprus | Turkey, Libya, Iran, Israel, Jordan, Lebanon, Syria | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014), Pennacchio (2013) | |
| 8 | Phloeosinus cedri | Spain | Turkey, Algeria, Morocco, India | Introduced species | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) |
| 9 | Phloeosinus gillerforsi | Azores (Portugal) | Canary Islands | May be endemic in either or both groups of Macaronesic archipelagos | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) |
| 10 | Pityogenes pennidens | Greece, Cyprus | Russia, Israel, Syria | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) | |
| 11 | Pityophthorus mauretanicus | France | Algeria, Egypt, Libya, Morocco, Tunisia | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) | |
| 12 | Pityophthorus pityographus cribratus | Greece | Russia, Turkey | Alonso‐Zarazaga et al. (2017), de Jong et al. (2014) | |
| 13 | Pityophthorus solus | Spain | North America | Introduced species | Alonso‐Zarazaga et al. (2017) |
| 14 | Xyleborinus gracilis | Azores (Portugal) | North America, Central America, South America | Introduced species | Atkinson (2019), Alonso‐Zarazaga et al. (2017) |
| 15 | Xyleborus perforans | Azores (Portugal) | North America, Central America, South America, Asia, North Africa, Sub‐Saharan Africa, Oceania | Alonso‐Zarazaga et al. (2017) | |
| 16 | Xylosandrus compactus | Italy, France, Greece | North America, Central America, South America, Asia, Sub‐Saharan Africa | Wood and Bright (1992), Wood (2007), Spanou et al. (2019), Anses (2017), Garonna et al. (2012) |
3.3. Regulatory status
3.3.1. Council Directive 2000/29/EC
Non‐EU Scolytinae species are listed in Council Directive 2000/29/EC as Scolytidae spp. (non‐European). Details are presented in Tables 3 and 4.
Table 3.
Non‐EU Scolytinae spp. in Council Directive 2000/29/EC
| Annex II, Part A | Harmful organisms whose introduction into, and spread within, all member states shall be banned if they are present on certain plants or plant products | |
|---|---|---|
| Section I | Harmful organisms not known to occur in the community and relevant for the entire community | |
| (a) | Insects, mites and nematodes, at all stages of their development | |
| Species | Subject of contamination | |
| 28. | Scolytidae spp. (non‐European) | Plants of conifers (Coniferales), over 3 m in height, other than fruit and seeds, wood of conifers (Coniferales) with bark, and isolated bark of conifers (Coniferales), originating in non‐European countries |
3.3.2. Legislation addressing the hosts of Scolytinae spp
Table 4.
Regulated hosts and commodities that may involve Scolytinae spp. in Annexes III, IV and V of Council Directive 2000/29/EC
| Annex III, Part A | Plants, plant products and other objects the introduction of which shall be prohibited in all Member States | |
|---|---|---|
| Description | Country of origin | |
| Plants of Abies Mill., Cedrus Trew, […], Larix Mill., Picea A. Dietr., Pinus L., Pseudotsuga Carr. and Tsuga Carr., other than fruit and seeds | Non‐European countries | |
| Annex IV, Part A | Special requirements which shall be laid down by all member states for the introduction and movement of plants, plant products and other objects into and within all member states | |
| Section I | Plants, plant products and other objects originating outside the community | |
| Plants, plant products and other objects | Special requirements | |
| 1.5 |
Whether or not listed among the CN codes in Annex V, Part B, wood of conifers (Coniferales), other than in the form of: – chips, particles, sawdust, shavings, wood waste and scrap obtained in whole or part from these conifers, – wood packaging material, in the form of packing cases, boxes, crates, drums and similar packings, pallets, box pallets and other load boards, pallet collars, dunnage, whether actually in use or not in the transport of objects of all kinds, except dunnage supporting consignments of wood, which is constructed from wood of the same type and quality as the wood in the consignment and which meets the same Union phytosanitary requirements as the wood in the consignment, but including that which has not kept its natural round surface, originating in Russia, Kazakhstan and Turkey. |
Official statement that the wood: (a) originates in areas known to be free from: — Pissodes spp. (non‐European) The area shall be mentioned on the certificates referred to in Article 13.1.(ii), under the rubric ‘place of origin,’ or […] or (c) has undergone kiln‐drying to below 20% moisture content, expressed as a percentage of dry matter, achieved through an appropriate time/temperature schedule. There shall be evidence thereof by a mark ‘kiln‐dried’ or ‘K.D’. or another internationally recognised mark, put on the wood or on any wrapping in accordance with the current usage, or (d) has undergone an appropriate heat treatment to achieve a minimum temperature of 56 °C for a minimum duration of 30 continuous minutes throughout the entire profile of the wood (including at its core). There shall be evidence thereof by a mark ‘HT’ put on the wood or on any wrapping in accordance with current usage, and on the certificates referred to in Article 13.1.(ii), or (e) has undergone an appropriate fumigation to a specification approved in accordance with the procedure laid down in Article 18.2. There shall be evidence thereof by indicating on the certificates referred to in Article 13.1.(ii), the active ingredient, the minimum wood temperature, the rate (g/m 3) and the exposure time (h), or (f) has undergone an appropriate chemical pressure impregnation with a product approved in accordance with the procedure laid down in Article 18.2. There shall be evidence thereof by indicating on the certificates referred to in Article 13.1.(ii), the active ingredient, the pressure (psi or kPa) and the concentration (%). |
| 1.7 |
Whether or not listed among the CN codes listed in Annex V, Part B, wood in the form of chips, particles, sawdust, shavings, wood waste and scrap obtained in whole or in part from conifers (Coniferales), originating in — Russia, Kazakhstan and Turkey, — non‐European countries other than Canada, China, Japan, the Republic of Korea, Mexico, Taiwan and the USA, where Bursaphelenchus xylophilus (Steiner et Bührer) Nickle et al. is `known to occur. |
Official statement that the wood: (a) originates in areas known to be free from: — Pissodes spp. (non‐European) The area shall be mentioned on the certificates referred to in Article 13.1.(ii), under the rubric ‘place of origin,’ or (b) has been produced from debarked round wood, or (c) has undergone kiln‐drying to below 20% moisture content, expressed as a percentage of dry matter, achieved through an appropriate time/temperature schedule, or (d) has undergone an appropriate fumigation to a specification approved in accordance with the procedure laid down in Article 18.2. There shall be evidence of the fumigation by indicating on the certificates referred to in Article 13.1.(ii), the active ingredient, the minimum wood temperature, the rate (g/m 3) and the exposure time (h), or (e) has undergone an appropriate heat treatment to achieve a minimum temperature of 56 °C for a minimum duration of 30 continuous minutes throughout the entire profile of the wood (including at its core), the latter to be indicated on the certificates referred to in Article 13.1.(ii). |
| 8.1. | Plants of conifers (Coniferales), other than fruit and seeds, originating in non‐European countries | Without prejudice to the prohibitions applicable to the plants listed in Annex III(A)(1), where appropriate, official statement that the plants have been produced in nurseries and that the place of production is free from Pissodes spp. (non‐ European). |
| Annex V, Part A |
Plants, plant products and other objects which must be subject to a plant health inspection (at the place of production if originating in the community, before being moved within the community in the country of origin or the consignor country, if originating outside the community) before being permitted to enter the community Plants, plant products and other objects originating in the community |
|
| Section II | Plants, plant products and other objects which are potential carriers of harmful organisms of relevance for certain protected zones, and which must be accompanied by a plant passport valid for the appropriate zone when introduced into or moved within that zone | |
| 1.11. | Isolated bark of conifers (Coniferales) | |
| Annex V, Part B | Plants, plant products and other objects originating in territories, other than those territories referred to in part a | |
| Section II | Plants, plant products and other objects which are potential carriers of harmful organisms of relevance for the entire Community | |
| 2. |
Parts of plants, other than fruits and seeds of: – conifers (Coniferales) |
|
| 5. |
Isolated bark of: – conifers (Coniferales) |
|
3.4. Entry, establishment and spread in the EU
3.4.1. Host range
All non‐EU Scolytinae feeding on conifers attack plant species belonging mainly to four botanical families: Araucariaceae, Cupressaceae, Pinaceae and Taxaceae. The number of species reported to attack various conifer plants are shown in Figure 4. Most common host plants belong to the genera Abies, Picea and Pinus (Figure 4). Other host plants are from the genera Cupressus, Larix, Juniperus, Cedrus, Chamaecyparis, Pseudotsuga, Tsuga and Thuja (see Annex A of EFSA PLH Panel, 2020). The number of species reported here includes also those that have been reported from more than one plant genera. Specifically, 24% of the species are reported to attack more than one plant genus (Figure 5). From the 397 species reported attacking Pinus, 117 are reported to attack other host genera as well. For 14 species only the plant family is reported and not specific host plant species or genera.
Figure 4.
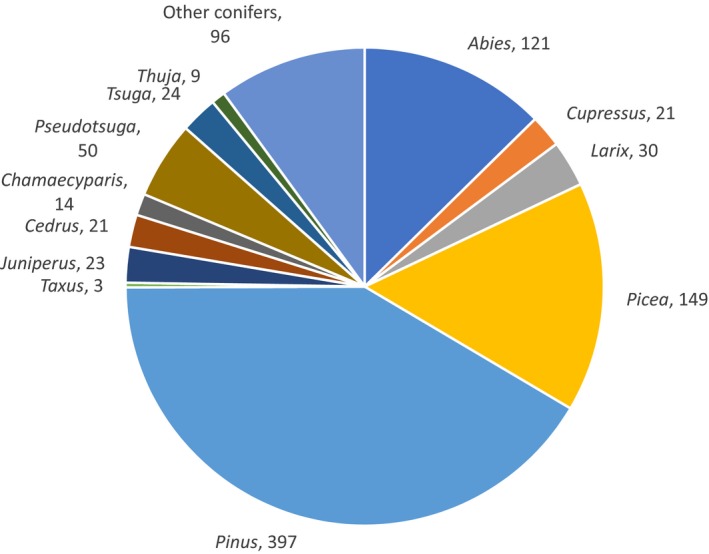
Species reported on each host plant genera
Figure 5.
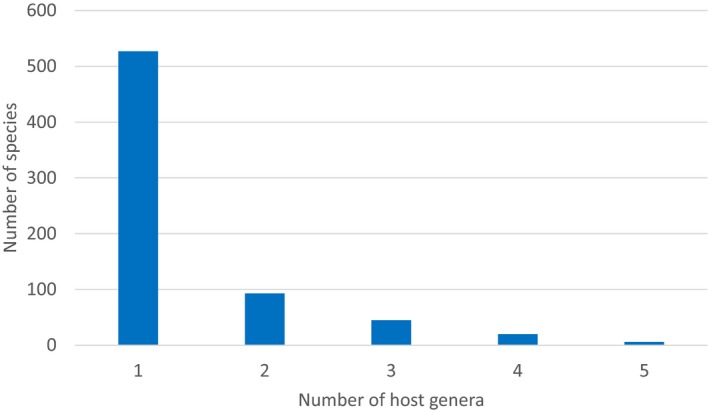
Number of species reported attacking one or more plant genera
3.4.2. Entry
3.4.2.1.
Is the pest able to enter into the EU territory? If yes, identify and list the pathways.
Yes, non‐Eu Scolytinae are able to enter at any developmental stage on wood and plants for planting.
The main pathways for entry of the non‐EU Scolytinae are:
plants for planting (including seeds) of conifers, with or without soil
cut branches of conifer plants
fruits (including cones of conifers) of conifer plants
round wood with bark of conifer plants
round wood without bark of conifer plants
sawn wood without bark
sawn wood with bark
wood packaging material
bark of conifers
manufactured wood items
wood chips.
For the pathways listed above, the following prohibitions (Annex III) or special requirements (Annex IV) are in place:
Plants for planting
Plants of Abies, Cedrus, Chamaecyparis, Juniperus, Larix, Picea, Pinus, Pseudotsuga, Tsuga, are prohibited from non‐European countries (Annex IIIA 1.)
Plants of conifers other than fruit and seeds – special requirements for import (Annex IVAI 8.1., 8.2.)
Plants of Pinus L., intended for planting, other than seeds – special requirements in relation to other pests (Annex IVAI 9.)
Plants of Abies, Larix, Picea, Pinus, Pseudotsuga, Tsuga, intended for planting, other than seeds – special requirements in relation to other pests (Annex IVAI 10.)
Wood
Wood of conifers – special requirements for import (Annex IVAI 1.1., 1.2., 1.3., 1.4., 1.5.)
Bark
Isolated bark of conifers is prohibited from non‐European countries (Annex IIIA 4.)
Isolated bark of conifers – special requirements (Annex IVAI 7.3.)
There are also records of interception for several species around the world on various pathways (Brockerhoff et al., 2006; Haack, 2006). The most common pathway is round wood with bark followed by wood packaging material and sawn wood with bark (Figure 6).
Figure 6.
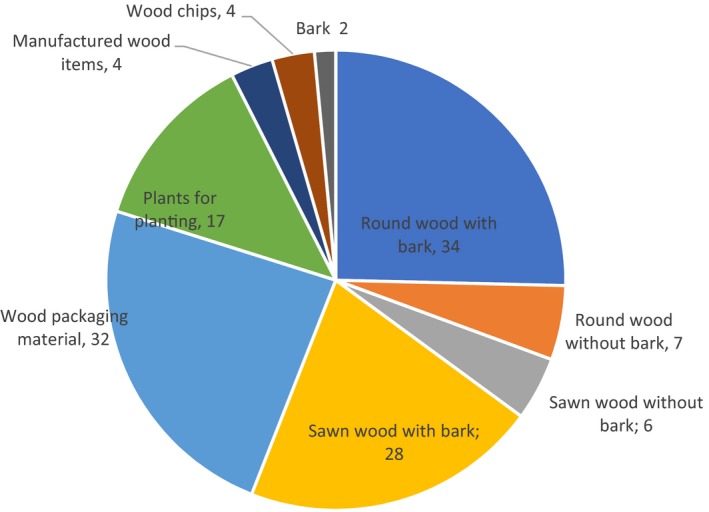
Number of species intercepted on different pathways worldwide
3.4.3. Establishment
3.4.3.1.
Is the pest able to become established in the EU territory?
Yes, the Non‐EU Scolytinae species are considered to be able to establish in the EU territory
3.4.3.2. EU distribution of main host plants
Non‐EU Scolytinae species feed on various coniferous hosts that are distributed throughout the EU territory (see Figures 7, 8, 9, 10, 11, 12).
Figure 7.
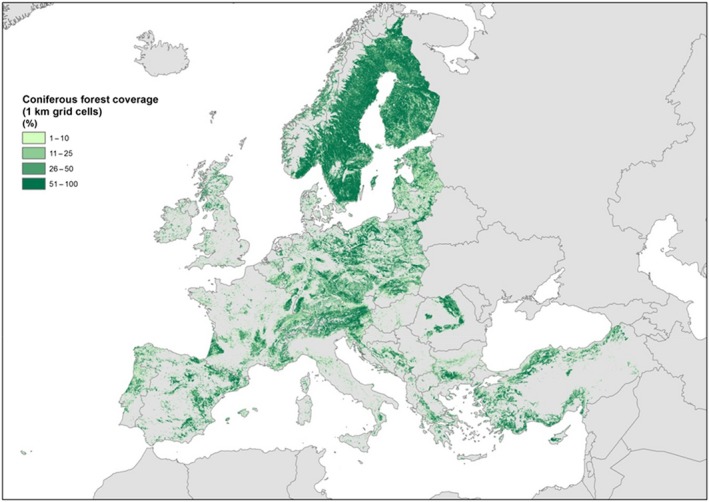
The cover percentage of coniferous forests in Europe with a range of values from 0 to 100 at 1 km resolution (source: Corine Land Cover year 2012 version 18.5 by European Environment Agency)
Figure 8.
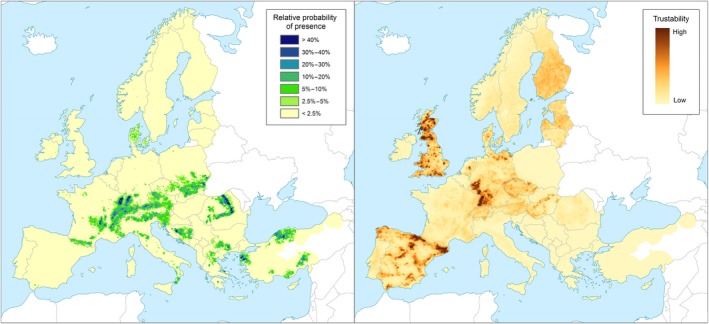
Left panel: Relative probability of presence (RPP) of the genus Abies (based on data from the species: Abies alba, Abies cephalonica, Abies borisii‐regis, Abies nordmanniana, Abies cilicica, Abies pinsapo, Abies numidica, Abies nebrodensis, Abies grandis, Abies procera) in Europe, mapped at 100 km2 resolution. The underlying data are from European‐wide forest monitoring data sets and from national forestry inventories based on standard observation plots measuring in the order of hundreds m². RPP represents the probability of finding at least one individual of the taxon in a standard plot placed randomly within the grid cell. For details, see Appendix A (courtesy of JRC, 2017). Right panel: Trustability of RPP. This metric expresses the strength of the underlying information in each grid cell and varies according to the spatial variability in forestry inventories. The colour scale of the trustability map is obtained by plotting the cumulative probabilities (0‐1) of the underlying index (for details see Appendix A)
Figure 9.
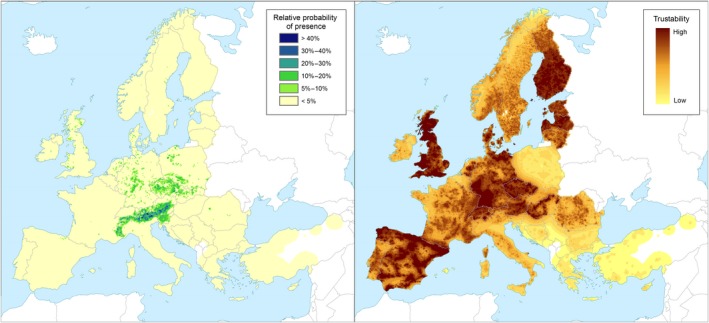
Left panel: Relative probability of presence (RPP) of the genus Larix (based on data from the species: Larix decidua, Larix kaempferi, Larix sibirica) in Europe, mapped at 100 km2 resolution. The underlying data are from European‐wide forest monitoring data sets and from national forestry inventories based on standard observation plots measuring in the order of hundreds m². RPP represents the probability of finding at least one individual of the taxon in a standard plot placed randomly within the grid cell. For details, see Appendix A (courtesy of JRC, 2017). Right panel: Trustability of RPP. This metric expresses the strength of the underlying information in each grid cell and varies according to the spatial variability in forestry inventories. The colour scale of the trustability map is obtained by plotting the cumulative probabilities (0–1) of the underlying index (for details see Appendix A)
Figure 10.
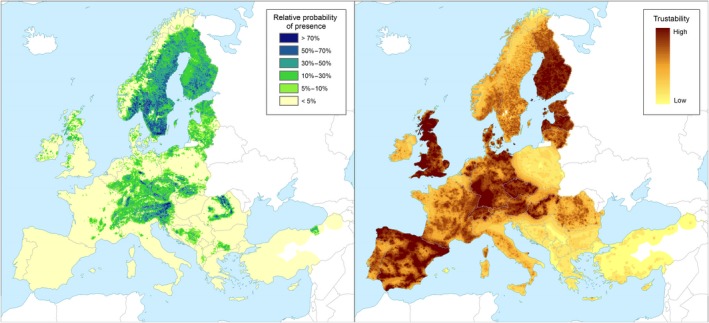
Left panel: Relative probability of presence (RPP) of the genus Picea (based on data from the species: Picea abies, Picea sitchensis, Picea glauca, Picea engelmannii, Picea pungens, Picea omorika, Picea orientalis) in Europe, mapped at 100 km2 resolution. The underlying data are from European‐wide forest monitoring data sets and from national forestry inventories based on standard observation plots measuring in the order of hundreds m². RPP represents the probability of finding at least one individual of the taxon in a standard plot placed randomly within the grid cell. For details, see Appendix A (courtesy of JRC, 2017). Right panel: Trustability of RPP. This metric expresses the strength of the underlying information in each grid cell and varies according to the spatial variability in forestry inventories. The colour scale of the trustability map is obtained by plotting the cumulative probabilities (0‐1) of the underlying index (for details see Appendix A)
Figure 11.
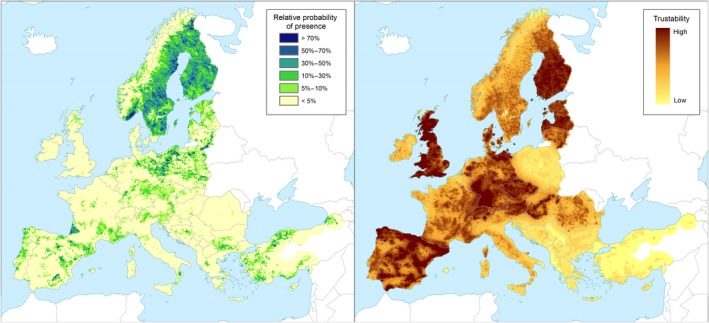
Left panel: Relative probability of presence (RPP) of the genus Pinus (based on data from the species: Pinus sylvestris, Pinus pinaster, Pinus halepensis, Pinus nigra, Pinus pinea, Pinus contorta, Pinus cembra, Pinus mugo, Pinus radiata, Pinus canariensis, Pinus strobus, Pinus brutia, Pinus banksiana, Pinus ponderosa, Pinus heldreichii, Pinus leucodermis, Pinus wallichiana) in Europe, mapped at 100 km2 resolution. The underlying data are from European‐wide forest monitoring data sets and from national forestry inventories based on standard observation plots measuring in the order of hundreds m². RPP represents the probability of finding at least one individual of the taxon in a standard plot placed randomly within the grid cell. For details, see Appendix A (courtesy of JRC, 2017). Right panel: Trustability of RPP. This metric expresses the strength of the underlying information in each grid cell and varies according to the spatial variability in forestry inventories. The colour scale of the trustability map is obtained by plotting the cumulative probabilities (0–1) of the underlying index (for details see Appendix A)
Figure 12.
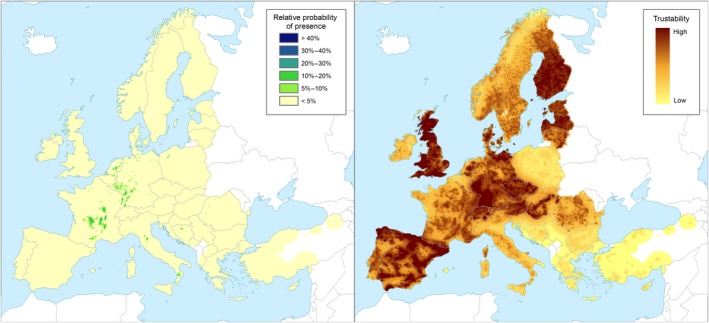
Left panel: Relative probability of presence (RPP) of the species Pseudotsuga menziesii in Europe, mapped at 100 km2 resolution. The underlying data are from European‐wide forest monitoring data sets and from national forestry inventories based on standard observation plots measuring in the order of hundreds m². RPP represents the probability of finding at least one individual of the taxon in a standard plot placed randomly within the grid cell. For details, see Appendix A (courtesy of JRC, 2017). Right panel: Trustability of RPP. This metric expresses the strength of the underlying information in each grid cell and varies according to the spatial variability in forestry inventories. The colour scale of the trustability map is obtained by plotting the cumulative probabilities (0–1) of the underlying index (for details see Appendix A)
3.4.3.3. Climatic conditions affecting establishment
Non‐EU Scolytinae are distributed worldwide in all continents. Most of the species are present in North America where Koppen–Geiger climate zones (Figure 13) such as Cfb, Cfa and Csa occur. These climate zones also occur in large areas in the EU. Moreover, 31% of the non‐EU Scolytinae are known to be distributed in Asia. Climatic zones such as Dfb, Cfa and Csa that occur in Asia also occur in EU (MacLeod and Korycinska, 2019). Climatic conditions are not expected to limit the ability for establishment of non‐EU Scolytinae.
Figure 13.
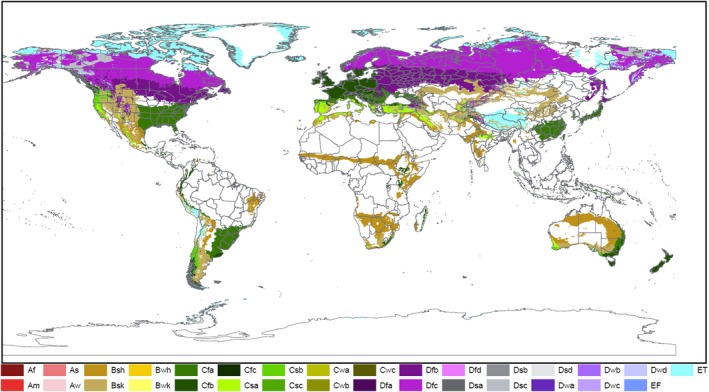
Köppen‐Geiger climatic zones in Europe and worldwide, 13 climate types in EU 28: Bsh, Bsk, Cfa, Cfb, Cfc, Csa, Csb, Csc, Dfb, Dfc, Dsb, Dsc, ET (according to MacLeod and Korycinska (2019))
3.4.4. Spread
3.4.4.1.
Is the pest able to spread within the EU territory following establishment?
Yes, non‐EU Scolytinae are able to spread by natural flight, or with various commodities.
RNQPs: Is spread mainly via specific plants for planting, rather than via natural spread or via movement of plant products or other objects?
Yes, a few species (e.g. Xylosandrus compactus) are spreading mainly via plants for planting.
Although generally little is known about the flight capacity of Scolytinae, for several species data have been collected with flight mills for several species (Eveden et al., 2014; Forsse and Solbreck, 1985) (Jactel and Gaillard, 1991), or direct observations (Nilssen, 1984; Chase et al., 2017). The species studied were able to cover 20–50 km by flight. In British Columbia, Dendroctonus ponderosae was observed by aerial captures with fixed balloons and weather radar measurements to cover 30–110 km per day up to 800 m above canopy (Jackson et al., 2008). Many species can also travel in all sorts of wood commodities (see Section 3.4.2 on entry). Bark‐ and ambrosia beetles travel in round wood with bark, firewood, wood chips and plants for planting; ambrosia beetles travel with debarked round wood, squared wood and objects made of wood; bark beetles can travel with bark alone. Since its first report in Germany in 1951, the Asian species Xylosandrus germanus spread to 21 neighbouring countries (Galko et al., 2019). Plants for planting are suspected to be the main pathway for Xylosandrus compactus, which rapidly spread all along the Tyrrhenian coasts up to France since its first occurrence in Italy (ANSES, 2017).
3.5. Impacts
3.5.1.
Would the pests’ introduction have an economic or environmental impact on the EU territory?
Yes, the introduction of non‐EU Scolytinae species would have an economic or environmental impact on the EU territory.
RNQPs: Does the presence of the pest on plants for planting have an economic impact, as regards the intended use of those plants for planting?
Yes, the presence of the pest on plants for planting can have an economic impact, as regards the intended use of those plants for planting.
Tree killing is an obvious impact of Scolytinae species, Dendroctonus ponderosae has caused the most damaging outbreak in history, killing more than 450 million m3 of Pinus contorta in British Columbia between 2000 and 2015 (British Columbia Government, 2019). In addition to killing trees, Scolytinae also reduce their commercial value, by the loss of mechanical properties and/or aesthetic value caused by the galleries of ambrosia beetles in the sapwood or the introduction of pathogenic or lignophagous associated fungi by bark and ambrosia beetles. Fungi associated with non‐native bark‐ or ambrosia beetles could prove extremely dangerous for some new host plants colonised in a new area, as recently observed after the recent introduction into South‐eastern USA of the Redbay ambrosia beetle, Xyleborus glabratus with a fungal symbionts, Raffaelea lauricola, that caused the death of millions of avocados, Persea americana, and redbay, Persea borbonia (Hughes et al., 2017) Other types of impacts include reduction of ecosystem services when forest biodiversity, water balance or soil properties are affected, or socio‐economic impact when amenity trees or whole landscapes are modified by the disappearance of tree species (Grégoire et al., 2015).
Details about species with reported impact can be found in the short list excel file (see Annex B in EFSA PLH Panel, 2020). There are 566 species for which no information is available about their potential economic or environmental impact.
3.6. Availability and limits of mitigation measures
3.6.1.
Are there measures available to prevent the entry into, establishment within or spread of the pest within the EU such that the risk becomes mitigated?
Yes, the existing measures (see Sections 3.3 and 3.6.1) can mitigate the risks of entry, establishment, and spread of non‐EU Scolytinae species within the EU
RNQPs: Are there measures available to prevent pest presence on plants for planting such that the risk becomes mitigated?
Yes, plants for planting from pest free areas and grown in isolation would mitigate the risk in case non‐EU Scolytinae entered the EU.
3.6.2. Identification of additional measures
Phytosanitary measures are currently applied to coniferous plants (see Sections 3.3 and 3.4.2).
3.6.2.1. Additional control measures
Potential additional control measures are listed in Table 5.
Table 5.
Selected control measures (a full list is available in EFSA PLH Panel, 2018) for pest entry/establishment/spread/impact in relation to currently unregulated hosts and pathways. Control measures are measures that have a direct effect on pest abundance
| Information sheet title (with hyperlink to information sheet if available) | Control measure summary | Risk component (entry/establishment/spread/impact) |
|---|---|---|
| Growing plants in isolation | Description of possible exclusion conditions that could be implemented to isolate the crop from pests and if applicable relevant vectors. E.g. a dedicated structure such as glass or plastic greenhouses | Entry/spread |
| Chemical treatments on consignments or during processing |
Use of chemical compounds that may be applied to plants or to plant products after harvest, during process or packaging operations and storage The treatments addressed in this information sheet are: a) fumigation; b) spraying/dipping pesticides; c) surface disinfectants; d) process additives; e) protective compounds |
Entry/spread |
| Chemical treatments on crops including reproductive material | Application of insecticides on nurseries for plants for planting may be considered to reduce the presence of the pest | Entry/spread |
3.6.2.2. Additional supporting measures
Since the import of conifers plants, wood and wood products are currently regulated no additional supporting measures are specified.
3.6.2.3. Biological or technical factors limiting the effectiveness of measures to prevent the entry, establishment and spread of the pest
There is always the possibility of recolonisation of wood commodities and plants for planting after a fumigation treatment.
3.7. Uncertainty
There are gaps in the scientific knowledge on the biology of many Scolytinae species (e.g. associations with a new host or with new pathogenic fungi). Therefore, it is difficult to know all the possible impacts that could be expected. Potential establishment of some species occurring in tropical areas is unknown.
4. Conclusions
Out of the 705 non‐EU Scolytinae species which were considered for pest categorisation the panel identified 139 species of non‐EU Scolytinae which meet all the criteria assessed by EFSA for consideration as potential quarantine pests for the EU territory. No information was available on the potential impact for 566 species. However, this does not exclude that those species could have an economic or environmental impact when they are introduced into a new environment (see for example the case of Xyleborous glabratus in Section 3.5).
Table 6.
The Panel's conclusions on the pest categorisation criteria defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column)
| Criterion of pest categorisation | Panel's conclusions against criterion in Regulation (EU) 2016/2031 regarding Union quarantine pest | Panel's conclusions against criterion in Regulation (EU) 2016/2031 regarding Union regulated non‐quarantine pest | Key uncertainties |
|---|---|---|---|
| Identity of the pests (Section 3.1) | The identity of the NESC is well established and the 705 species considered in the pest categorisation are described in worldwide, regional or national catalogues and faunae | The identity of the NESC is well established and the 222 species considered in the pest categorisation are described in worldwide, regional or national catalogues and faunae | None |
| Absence/presence of the pest in the EU territory (Section 3.2) | From the total of 705 species considered for the pest categorisation there are 16 species also present in a few EU Member States | From the total of 705 species considered for the pest categorisation there are 16 species also present in a few EU Member States | None |
| Regulatory status (Section 3.3) | Non‐EU Scolytinae species are listed in Council Directive 2000/29/EC as Scolytidae (non European) in Annex II, Part A, Section I | Non‐EU Scolytinae are regulated as quarantine pests and legislation is in place addressing the hosts of Scolytinae spp (Annex III, Part A and Annex IV, part A, Section I of Council Directive 2000/29/EC) | None |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4) | NESC are able to enter into the EU through plants for planting (including seeds) of conifers, with or without soil, cut branches of conifer plants, fruits (including cones of conifers) of conifer plants, round wood with bark of conifer plants, round wood without bark of conifer plants, sawn wood without bark, sawn wood with bark, wood packaging material, bark of conifers, manufactured wood items, wood chips. Establishment is possible as host plants are available and climatic conditions similar to their native range do occur in the EU. Dispersal by flight and movement of commodities are the main means for spread | The spread is mainly by natural flight or with various commodities. Some species are spread by plants for planting | Potential establishment of some species occurring in tropical areas is unknown |
| Potential for consequences in the EU territory (Section 3.5) | NESC would have an economic or environmental impact if they were introduced in the EU territory. For 566 species there is no information available about potential impact | Yes, the presence of the pest on plants for planting can have an economic impact, as regards the intended use of those plants for planting | For several species the potential impact is not known although impact is commonly observed for species outside from their native range |
| Available measures (Section 3.6) | Yes, the existing measures can mitigate the risks of entry, establishment, and spread within the EU | Yes, plants for planting from pest free areas and grown in isolation would mitigate the risk in case non‐EU Scolytinae entered the EU | None |
| Conclusion on pest categorisation (Section 4) | 139 non‐EU Scolytinae feeding on conifers meet all criteria assessed by EFSA above for quarantine pests. For 566 species information about impact is lacking and the panel was unable to conclude | The species considered in the current categorisation are non‐EU Scolytinae (regulated in annex IIAI) and as such do not meet the corresponding criterion evaluated by EFSA to qualify as potential regulated non‐quarantine pests | There are gaps in the scientific knowledge on the biology of many Scolytinae species. |
| Aspects of assessment to focus on/scenarios to address in future if appropriate | – |
Abbreviations
- CLC
Corine Land Cover
- C‐SMFA
constrained spatial multi‐scale frequency analysis
- EPPO
European and Mediterranean Plant Protection Organization
- EUFGIS
European Information System on Forest Genetic Resources
- FAO
Food and Agriculture Organization
- GD2
Georeferenced Data on Genetic Diversity
- IPPC
International Plant Protection Convention
- ISPM
International Standards for Phytosanitary Measures
- MS
Member State
- NESC
non‐EU Scolytinae of coniferous hosts
- PLH
EFSA Panel on Plant Health
- PZ
Protected Zone
- RNQP
regulated non‐quarantine pest
- RPP
relative probability of presence
- TFEU
Treaty on the Functioning of the European Union
- ToR
Terms of Reference
Glossary
- Containment (of a pest)
Application of phytosanitary measures in and around an infested area to prevent spread of a pest (FAO, 1995, 2017)
- Control (of a pest)
Suppression, containment or eradication of a pest population (FAO, 1995, 2017)
- Entry (of a pest)
Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2017)
- Eradication (of a pest)
Application of phytosanitary measures to eliminate a pest from an area (FAO, 2017)
- Establishment (of a pest)
Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2017)
- Impact (of a pest)
The impact of the pest on the crop output and quality and on the environment in the occupied spatial units
- Introduction (of a pest)
The entry of a pest resulting in its establishment (FAO, 2017)
- Measures
Control (of a pest) is defined in ISPM 5 (FAO, 2017) as ‘Suppression, containment or eradication of a pest population’ (FAO, 1995). Control measures are measures that have a direct effect on pest abundance. Supporting measures are organisational measures or procedures supporting the choice of appropriate Risk Reduction Options that do not directly affect pest abundance
- Pathway
Any means that allows the entry or spread of a pest (FAO, 2017)
- Phytosanitary measures
Any legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2017)
- Protected zones (PZ)
A Protected zone is an area recognised at EU level to be free from a harmful organism, which is established in one or more other parts of the Union
- Quarantine pest
A pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2017)
- Regulated non‐quarantine pest
A non‐quarantine pest whose presence in plants for planting affects the intended use of those plants with an economically unacceptable impact and which is therefore regulated within the territory of the importing contracting party (FAO, 2017)
- Risk reduction option (RRO)
A measure acting on pest introduction and/or pest spread and/or the magnitude of the biological impact of the pest should the pest be present. A RRO may become a phytosanitary measure, action or procedure according to the decision of the risk manager
- Spread (of a pest)
Expansion of the geographical distribution of a pest within an area (FAO, 2017)
Appendix A – Methodological notes on Figures 7, 8, 9, 10, 11, 12–12
1.
The relative probability of presence (RPP) reported here for Quercus spp. in Figure 2 and in the European Atlas of Forest Tree Species (de Rigo et al., 2016; San‐Miguel‐Ayanz et al., 2016) is the probability of that genus to occur in a given spatial unit (de Rigo et al., 2017). In forestry, such a probability for a single taxon is called ‘relative’. The maps of RPP are produced by means of the constrained spatial multi‐scale frequency analysis (C‐SMFA) (de Rigo et al., 2014, 2017) of species presence data reported in geo‐located plots by different forest inventories.
A.1. Geolocated plot databases
The RPP models rely on five geo‐databases that provide presence/absence data for tree species and genera: four European‐wide forest monitoring data sets and a harmonised collection of records from national forest inventories (de Rigo et al., 2014, 2016, 2017). The databases report observations made inside geolocalised sample plots positioned in a forested area, but do not provide information about the plot size or consistent quantitative information about the recorded species beyond presence/absence.
The harmonisation of these data sets was performed within the research project at the origin of the European Atlas of Forest Tree Species (de Rigo et al., 2016; San‐Miguel‐Ayanz, 2016; San‐Miguel‐Ayanz et al., 2016). Given the heterogeneity of strategies of field sampling design and establishment of sampling plots in the various national forest inventories (Chirici et al. 2011a,b), and also given legal constraints, the information from the original data sources was harmonised to refer to an INSPIRE compliant geospatial grid, with a spatial resolution of 1 km2 pixel size, using the ETRS89 Lambert Azimuthal Equal‐Area as geospatial projection (EPSG: 3035, http://spatialreference.org/ref/epsg/etrs89-etrs-laea/).
A.1.1. European National Forestry Inventories database
This data set was derived from National Forest Inventory data and provides information on the presence/absence of forest tree species in approximately 375,000 sample points with a spatial resolution of 1 km²/pixel, covering 21 European countries (de Rigo et al., 2014, 2016).
A.1.2. Forest Focus/Monitoring data set
This project is a Community scheme for harmonised long‐term monitoring of air pollution effects in European forest ecosystems, normed by EC Regulation No. 2152/20034. Under this scheme, the monitoring is carried out by participating countries on the basis of a systematic network of observation points (Level I) and a network of observation plots for intensive and continuous monitoring (Level II). For managing the data, the JRC implemented a Forest Focus Monitoring Database System, from which the data used in this project were taken (Hiederer et al., 2007; Houston Durrant and Hiederer, 2009). The complete Forest Focus data set covers 30 European Countries with more than 8,600 sample points.
A.1.3. BioSoil data set
This data set was produced by one of a number of demonstration studies performed in response to the ‘Forest Focus’ Regulation (EC) No 2152/2003 mentioned above. The aim of the BioSoil project was to provide harmonised soil and forest biodiversity data. It comprised two modules: a Soil Module (Hiederer et al., 2011) and a Biodiversity Module (Houston Durrant et al., 2011). The data set used in the C‐SMFA RPP model came from the Biodiversity module, in which plant species from both the tree layer and the ground vegetation layer were recorded for more than 3,300 sample points in 19 European Countries.
A.1.4. European Information System on Forest Genetic Resources (EUFGIS)
EUFGIS (http://portal.eufgis.org) is a smaller geodatabase providing information on tree species composition in over 3,200 forest plots in 34 European countries. The plots are part of a network of forest stands managed for the genetic conservation of one or more target tree species. Hence, the plots represent the natural environment to which the target tree species are adapted.
A.1.5. Georeferenced Data on Genetic Diversity (GD2)
GD2 (http://gd2.pierroton.inra.fr) provides information about 63 species of interest for genetic conservation. The database covers 6,254 forest plots located in stands of natural populations that are traditionally analysed in genetic surveys. While this database covers fewer species than the others, it covers 66 countries in Europe, North Africa and the Middle East, making it the data set with the largest geographic extent.
A.2. Modelling methodology
For modelling, the data were harmonised in order to have the same spatial resolution (1 km²) and filtered to a study area comprising 36 countries in the European continent. The density of field observations varies greatly throughout the study area and large areas are poorly covered by the plot databases. A low density of field plots is particularly problematic in heterogeneous landscapes, such as mountainous regions and areas with many different land use and cover types, where a plot in one location is not representative of many nearby locations (de Rigo et al., 2014). To account for the spatial variation in plot density, the model used here (C‐SMFA) considers multiple spatial scales when estimating RPP. Furthermore, statistical resampling is systematically applied to mitigate the cumulated data‐driven uncertainty.
The presence or absence of a given forest tree species then refers to an idealised standard field sample of negligible size compared with the 1 km2 pixel size of the harmonised grid. The modelling methodology considered these presence/absence measures as if they were random samples of a binary quantity (the punctual presence/absence, not the pixel one). This binary quantity is a random variable having its own probability distribution which is a function of the unknown average probability of finding the given tree species within a plot of negligible area belonging to the considered 1 km2 pixel (de Rigo et al., 2014). This unknown statistic is denoted hereinafter with the name of ‘probability of presence’.
C‐SMFA performs spatial frequency analysis of the geo‐located plot data to create preliminary RPP maps (de Rigo et al., 2014). For each 1 km² grid cell, the model estimates kernel densities over a range of kernel sizes to estimate the probability that a given species is present in that cell. The entire array of multi‐scale spatial kernels is aggregated with adaptive weights based on the local pattern of data density. Thus, in areas where plot data are scarce or inconsistent, the method tends to put weight on larger kernels. Wherever denser local data are available, they are privileged ensuring a more detailed local RPP estimation. Therefore, a smooth multi‐scale aggregation of the entire arrays of kernels and data sets is applied instead of selecting a local ‘best performing’ one and discarding the remaining information. This array‐based processing, and the entire data harmonisation procedure, are made possible thanks to the semantic modularisation which defines the Semantic Array Programming modelling paradigm (de Rigo, 2012).
The probability to find a single species (e.g. a particular coniferous tree species) in a 1 km² grid cell cannot be higher than the probability of presence of all the coniferous species combined. The same logical constraints applied to the case of single broadleaved species with respect to the probability of presence of all the broadleaved species combined. Thus, to improve the accuracy of the maps, the preliminary RPP values were constrained so as not to exceed the local forest‐type cover fraction with an iterative refinement (de Rigo et al., 2014). The forest‐type cover fraction was estimated from the classes of the Corine Land Cover (CLC) maps which contain a component of forest trees (Bossard et al., 2000; Büttner et al. 2012).
The resulting probability of presence is relative to the specific tree taxon, irrespective of the potential co‐occurrence of other tree taxa with the measured plots, and should not be confused with the absolute abundance or proportion of each taxon in the plots. RPP represents the probability of finding at least one individual of the taxon in a plot placed randomly within the grid cell, assuming that the plot has negligible area compared with the cell. As a consequence, the sum of the RPP associated with different taxa in the same area is not constrained to be 100%. For example, in a forest with two co‐dominant tree species which are homogeneously mixed, the RPP of both may be 100% (see e.g. the Glossary in San‐Miguel‐Ayanz et al. (2016), http://forest.jrc.ec.europa.eu/media/atlas/Glossary.pdf).
The robustness of RPP maps depends strongly on sample plot density, as areas with few field observations are mapped with greater uncertainty. This uncertainty is shown qualitatively in maps of ‘RPP trustability’. RPP trustability is computed on the basis of the aggregated equivalent number of sample plots in each grid cell (equivalent local density of plot data). The trustability map scale is relative, ranging from 0 to 1, as it is based on the quantiles of the local plot density map obtained using all field observations for the species. Thus, trustability maps may vary among species based on the number of databases that report a particular species (de Rigo et al., 2014, 2016).
The RPP and relative trustability range from 0 to 1 and are mapped at a 1 km spatial resolution. To improve visualisation, these maps can be aggregated to coarser scales (i.e. 10 × 10 pixels or 25 × 25 pixels, respectively summarising the information for aggregated spatial cells of 100 and 625 km2) by averaging the values in larger grid cells.
Appendix B – Non‐EU Scolytinae species identified which meet all critera for quarantine pests
1.
The appendix lists the 139 non‐EU Scolytinae species for which sufficient information on their biology and impact is available to conclude that they meet all criteria for quarantine species. Their host plant family and the number of interceptions are listed for each species. More detailed data (e.g. on host plant species, the raw data and further details on their biology, capacity to spread, economic and environmental impact) can be found in the supporting publication. Annex B of EFSA, 2020.
| ID | Species | Host plant family | Number of interceptions | Reference for interception |
|---|---|---|---|---|
| 1. | Carphoborus bifurcus | Pinaceae | 1 | Haack and Rabaglia (2013) |
| 2. | Carphoborus costatus | Pinaceae | ||
| 3. | Carphoborus zhobi | Pinaceae | ||
| 4. | Cnestus mutilatus | Pinaceae | 8 | Haack and Rabaglia (2013) |
| 5. | Conophthorus conicolens | Pinaceae | ||
| 6. | Conophthorus coniperda | Pinaceae | ||
| 7. | Conophthorus michoacanae | Pinaceae | ||
| 8. | Conophthorus monophyllae | Pinaceae | ||
| 9. | Conophthorus ponderosae | Pinaceae | ||
| 10. | Conophthorus radiatae | Pinaceae | ||
| 11. | Conophthorus resinosae | Pinaceae | ||
| 12. | Conophthorus teocotum | Pinaceae | ||
| 13. | Corthylus schaufussi | Araucariaceae | ||
| 14. | Cryphalus fulvus | Pinaceae | ||
| 15. | Cryphalus lipingensis | Pinaceae | ||
| 16. | Cryphalus montanus | Pinaceae | ||
| 17. | Cryphalus piceus | Pinaceae | ||
| 18. | Cryphalus ruficollis | Pinaceae | ||
| 19. | Crypturgus borealis | Pinaceae | 17 | Brockerhoff et al. (2014) |
| 20. | Cyrtogenius luteus | Pinaceae | 10 | Haack and Rabaglia (2013) |
| 21. | Dendroctonus adjunctus | Pinaceae | ||
| 22. | Dendroctonus approximates | Pinaceae | ||
| 23. | Dendroctonus armandi | Pinaceae | ||
| 24. | Dendroctonus brevicomis | Pinaceae | ||
| 25. | Dendroctonus frontalis | Pinaceae | 3 | Haack and Rabaglia (2013) |
| 26. | Dendroctonus jeffreyi | Pinaceae | ||
| 27. | Dendroctonus mesoamericanus | Pinaceae | ||
| 28. | Dendroctonus mexicanus | Pinaceae | 28 | Haack and Rabaglia (2013) |
| 29. | Dendroctonus murrayanae | Pinaceae | ||
| 30. | Dendroctonus parallelocollis | Pinaceae | ||
| 31. | Dendroctonus ponderosae | Pinaceae | 17 | Brockerhoff et al. (2006) |
| 32. | Dendroctonus pseudotsugae barragani | Pinaceae | ||
| 33. | Dendroctonus pseudotsugae pseudotsugae | Pinaceae | 12 | Brockheroff et al. (2003), Haack and Rabaglia (2013) |
| 34. | Dendroctonus punctatus | Pinaceae | ||
| 35. | Dendroctonus rhizophagus | Pinaceae | ||
| 36. | Dendroctonus rufipennis | Pinaceae | 11 | Brockerhoff et al. (2006) |
| 37. | Dendroctonus simplex | Pinaceae | ||
| 38. | Dendroctonus terebrans | Pinaceae | ||
| 39. | Dendroctonus valens | Pinaceae | ||
| 40. | Dendroctonus vitei | Pinaceae | ||
| 41. | Dryocoetes affaber | Pinaceae | 17 | Brockerhoff et al. (2014) |
| 42. | Dryocoetes caryi | Pinaceae | ||
| 43. | Dryocoetes confuses | Pinaceae | ||
| 44. | Gnathotrichus retusus | PInaceae | 11 | Brockerhoff et al. (2006) |
| 45. | Gnathotrichus sulcatus | Pinaceae | 62 | Haack and Rabaglia (2013) |
| 46. | Hylastes gracilis | Pinaceae | ||
| 47. | Hylastes longicollis | Pinaceae | ||
| 48. | Hylastes macer | Pinaceae | ||
| 49. | Hylastes nigrinus | Pinaceae | 11 | Brockerhoff et al. (2006) |
| 50. | Hylastes parallelus | Pinaceae | ||
| 51. | Hylastes porculus | Pinaceae | ||
| 52. | Hylastes salebrosus | Pinaceae | ||
| 53. | Hylastes tenuis | Pinaceae | ||
| 54. | Hylurdrectonus araucariae | Araucariaceae | ||
| 55. | Hylurgops longipillus | Pinaceae | ||
| 56. | Hylurgops pinifex | Pinaceae | ||
| 57. | Hylurgops porosus | Pinaceae | ||
| 58. | Hylurgops reticulatus | Pinaceae | ||
| 59. | Hylurgops rugipennis | Pinaceae | 6 | Brockerhoff et al. (2014) |
| 60. | Hypothenemus seriatus | Pinaceae | ||
| 61. | Ips apache | Pinaceae | 9 | Haack and Rabaglia (2013) |
| 62. | Ips avulsus | Pinaceae | 11 | Brockerhoff et al. (2006) |
| 63. | Ips bonanseai | Pinaceae | 27 | Haack and Rabaglia (2013) |
| 64. | Ips calligraphus | Pinaceae | 62 | Brockerhoff et al. (2006) |
| 65. | Ips confuses | Pinaceae | 6 | Brockerhoff et al. (2006) |
| 66. | Ips grandicollis | Pinaceae | 287 | Brockerhoff et al. (2006), Lawson et al. (2018) |
| 67. | Ips hauseri | Pinaceae | ||
| 68. | Ips hoppingi | Pinaceae | ||
| 69. | Ips knausi | Pinaceae | ||
| 70. | Ips lecontei | Pinaceae | 43 | Haack and Rabaglia (2013) |
| 71. | Ips nitidus | Pinaceae | 1 | Brockerhoff et al. (2014) |
| 72. | Ips paraconfusus | Pinaceae | ||
| 73. | Ips perturbatus | Pinaceae | ||
| 74. | Ips pini | Pinaceae | 43 | Brockerhoff et al. (2006); |
| 75. | Ips plastographus maritimus | Pinaceae | ||
| 76. | Ips plastographus plastographus | Pinaceae | 1 | Brockerhoff et al. (2014) |
| 77. | Ips schmutzenhoferi | Pinaceae | ||
| 78. | Ips shangrila | Pinaceae | ||
| 79. | Ips stebbingi | Pinaceae | ||
| 80. | Ips subelongatus | Pinaceae | 1 | Hellrigl (2002) |
| 81. | Ips tridens tridens | Pinaceae | ||
| 82. | Orthotomicus caelatus | Pinaceae | 45 | Brockerhoff et al. (2014) |
| 83. | Orthotomicus chaokhao | Pinaceae | ||
| 84. | Orthotomicus latidens | Pinaceae | ||
| 85. | Orthotomicus tridentatus | Pinaceae | ||
| 86. | Pachysquamus subcostulatus | Pinaceae | ||
| 87. | Phloeosinus armatus | Cupressaceae | 1 | Brockerhoff et al. (2006) |
| 88. | Phloeosinus cristatus | Cupressaceae | ||
| 89. | Phloeosinus cupressi | Cupressaceae | ||
| 90. | Phloeosinus dentatus | Cupressaceae | ||
| 91. | Phloeosinus scopulorum scopulorum | Cupressaceae | ||
| 92. | Phloeosinus sequoia | Cupressaceae | ||
| 93. | Phloeosinus sinensis | Cupressaceae | ||
| 94. | Pityogenes japonicus | Pinaceae | ||
| 95. | Pityogenes pennidens | Pinaceae | 1 | Brockerhoff et al. (2014) |
| 96. | Pityogenes scitus | Pinaceae | ||
| 97. | Pityogenes spessivtsevi | Pinaceae | ||
| 98. | Pityokteines marketae | Pinaceae | ||
| 99. | Pityokteines sparsus | Pinaceae | 11 | Brockerhoff et al. (2014) |
| 100. | Pityophthorus absonus | Pinaceae | ||
| 101. | Pityophthorus cariniceps | Pinaceae | ||
| 102. | Pityophthorus carmeli | Pinaceae | ||
| 103. | Pityophthorus confertus | Pinaceae | ||
| 104. | Pityophthorus confuses | Pinaceae | ||
| 105. | Pityophthorus micrographus sibiricus | Pinaceae | ||
| 106. | Pityophthorus nitidulus | Pinaceae | ||
| 107. | Pityophthorus opaculus | Pinaceae | ||
| 108. | Pityophthorus orarius | Pinaceae | ||
| 109. | Pityophthorus pityographus cribratus | Pinaceae | ||
| 110. | Pityophthorus puberulus | Pinaceae | ||
| 111. | Pityophthorus pulchellus | Pinaceae | ||
| 112. | Pityophthorus pulicarius | Pinaceae | ||
| 113. | Pityophthorus sculptor | Pinaceae | ||
| 114. | Pityophthorus setosus | Pinaceae | ||
| 115. | Polygraphus jezoensis | Pinaceae | ||
| 116. | Polygraphus major | Pinaceae | ||
| 117. | Polygraphus Proximus | Pinaceae | 3 | Haack and Rabaglia (2013) |
| 118. | Polygraphus rufipennis | Pinaceae | 125 | Brockerhoff et al. (2006) |
| 119. | Pseudips mexicanus | Pinaceae | 6 | Haack and Rabaglia (2013) |
| 120. | Pseudips orientalis a | Pinaceae | ||
| 121. | Pseudohylesinus nebulosus nebulosus | Pinaceae | 6 | Brockerhoff et al. (2014) |
| 122. | Pseudohylesinus pini | Pinaceae | ||
| 123. | Pseudohylesinus sericeus | Pinaceae | ||
| 124. | Scolytus morawitzi | Pinaceae | ||
| 125. | Scolytus mundus | Pinaceae | ||
| 126. | Scolytus reflexus | Pinaceae | ||
| 127. | Scolytus subscaber | Pinaceae | ||
| 128. | Scolytus tsugae | Pinaceae | ||
| 129. | Scolytus unispinosus | Pinaceae | ||
| 130. | Scolytus ventralis | Pinaceae | ||
| 131. | Tomicus armandii | Pinaceae | ||
| 132. | Tomicus brevipilosus | Pinaceae | ||
| 133. | Tomicus yunnanensis | Pinaceae | ||
| 134. | Xyleborus ferrugineus | Pinaceae, Araucariaceae | 16 | Brockerhoff et al. (2006), Lawson et al. (2018) |
| 135. | Xyleborus intrusus | Pinaceae | 10 | Brockerhoff et al. (2006) |
| 136. | Xyleborus perforans | Pinaceae, Araucariaceae | 205 | Brockerhoff et al. (2003) |
| 137. | Xyleborus seriatus | Cupressaceae, Pinaceae | ||
| 138. | Xylosandrus compactus | Cupressaceae, Pinaceae | ||
| 139. | Xyloterinus politus | Pinaceae | 3 | Brockerhoff et al. (2006) |
Cognato AI, 2000. Phylogenetic reveals new genus of Ipini bark beetle (Scolytidae). Annals of the Entomological Society of America, 93, 362–366. and the species Ips (=Orthotomicus) orientalis Wood & Yin, 1986 was moved under Pseudips orientalis (Wood & Yin, 1986). The recent catalogue of the Palaearctic species of Alonso‐Zarazaga et al. (2017) adopts this new classification.
Appendix C – Non‐EU Scolytinae species for which information on the impact is missing
1.
The appendix lists the 83 non‐EU Scolytinae species for which sufficient information on their biology and impact is available but for which no information on their impact is available. Their host plant family and the number of interceptions are listed for each species. More detailed data (e.g. on host plant species, the raw data and further details on their biology, capacity to spread) can be found in the supporting publication: Annex B of EFSA, 2020.
| ID | Species | Host plant family | Number of interceptions | Reference for interception |
|---|---|---|---|---|
| 1. | Ambrosiodmus hagedorni | Pinaceae | ||
| 2. | Ambrosiodmus lecontei | Pinaceae | ||
| 3. | Ambrosiodmus lewisi | Pinaceae | ||
| 4. | Coccotrypes advena | Pinaceae | 1 | Haack and Rabaglia (2013) |
| 5. | Cryphalus redikorzevi | Pinaceae | ||
| 6. | Cryphalus rubentis | Pinaceae | ||
| 7. | Dolurgus pumilus | Pinaceae | ||
| 8. | Dryocoetes granicollis | Pinaceae | ||
| 9. | Dryocoetes striatus | Pinaceae | 1 | Brockerhoff et al. (2014) |
| 10. | Dryocoetes uniseriatus | Pinaceae | ||
| 11. | Euwallacea interjectus | Pinaceae | ||
| 12. | Euwallacea validus | Pinaceae | 24 | Brockerhoff et al. (2006) |
| 13. | Gnathotrichus pilosus | Pinaceae | ||
| 14. | Hylastes obscurus | Pinaceae | ||
| 15. | Hylastes ruber | Pinaceae | ||
| 16. | Hylurgops inouyei | Pinaceae | ||
| 17. | Hylurgops interstitialis | Pinaceae | 1 | Brockerhoff et al. (2014) |
| 18. | Hypothenemus crudiae | Araucariaceae, Pinaceae | ||
| 19. | Hypothenemus interstitialis | Pinaceae | ||
| 20. | Ips borealis borealis | Pinaceae | 6 | Brockerhoff et al. (2006) |
| 21. | Ips borealis lanieri | Pinaceae | ||
| 22. | Ips borealis swainei | Pinaceae | ||
| 23. | Ips borealis thomasi | Pinaceae | ||
| 24. | Ips chinensis | Pinaceae | ||
| 25. | Ips cribricollis | Pinaceae | 39 | Haack and Rabaglia (2013) |
| 26. | Ips emarginatus | Pinaceae | ||
| 27. | Ips integer | Pinaceae | 82 | Haack and Rabaglia (2013) |
| 28. | Ips montanus | Pinaceae | 6 | Haack and Rabaglia (2013) |
| 29. | Ips perroti | Pinaceae | ||
| 30. | Ips pilifrons pilifrons | Pinaceae | ||
| 31. | Ips pilifrons sulcifrons | Pinaceae | ||
| 32. | Ips pilifrons thatcheri | Pinaceae | ||
| 33. | Ips pilifrons utahensis | Pinaceae | ||
| 34. | Ips tridens engelmanni | Pinaceae | ||
| 35. | Orthotomicus nobilis | Pinaceae | ||
| 36. | Pachycotes grandis | Araucariaceae | ||
| 37. | Phloeosinus pini | Pinaceae | 6 | Brockerhoff et al. (2014) |
| 38. | Phloeosinus scopulorum neomexicanus | Cupressaceae | ||
| 39. | Pityogenes carinulatus | Pinaceae | ||
| 40. | Pityogenes hopkinsi | Pinaceae | 17 | Brockerhoff et al. (2014) |
| 41. | Pityogenes knechteli | Pinaceae | ||
| 42. | Pityogenes seirindensis | Pinaceae | ||
| 43. | Pityokteines elegans | Pinaceae | ||
| 44. | Pityokteines minutus | Pinaceae | ||
| 45. | Pityophthorus balsameus | Pinaceae | ||
| 46. | Pityophthorus deletus | Pinaceae | ||
| 47. | Pityophthorus grandis | Pinaceae | ||
| 48. | Pityophthorus jucundus | Pinaceae | ||
| 49. | Pityophthorus lautus | Pinaceae | ||
| 50. | Pityophthorus murrayanae | Pinaceae | ||
| 51. | Pityophthorus serratus | Pinaceae | ||
| 52. | Pityophthorus solus | Pinaceae | ||
| 53. | Polygraphus verrucifrons | Pinaceae | ||
| 54. | Pseudips concinnus | Pinaceae | ||
| 55. | Pseudohylesinus tsugae | Pinaceae | ||
| 56. | Scierus pubescens | Pinaceae | ||
| 57. | Scolytoplatypus daimio | Pinaceae, Taxaceae | ||
| 58. | Scolytoplatypus raja | Pinaceae | ||
| 59. | Scolytoplatypus shogun | Pinaceae | ||
| 60. | Scolytoplatypus tycoon | Pinaceae | ||
| 61. | Scolytus oregoni | Pinaceae | ||
| 62. | Scolytus piceae | Pinaceae | ||
| 63. | Scolytus praeceps | Pinaceae | ||
| 64. | Tomicus pilifer | Pinaceae | ||
| 65. | Tomicus puellus | Pinaceae | ||
| 66. | Trypodendron proximum | Pinaceae | ||
| 67. | Trypodendron rufitarsus | Pinaceae | 1 | Brockerhoff et al. (2006) |
| 68. | Trypodendron scabricollis | Pinaceae | ||
| 69. | Xyleborinus gracilis | Pinaceae | ||
| 70. | Xyleborinus linearicollis | Araucariaceae | ||
| 71. | Xyleborinus spinifer | Pinaceae | ||
| 72. | Xyleborus aquilus | Pinaceae | ||
| 73. | Xyleborus detectus | Pinaceae | ||
| 74. | Xyleborus emarginatus | Pinaceae | ||
| 75. | Xyleborus festivus | Pinaceae | ||
| 76. | Xyleborus pinicola | Pinaceae | 1 | Browne (1980) |
| 77. | Xyleborus pubescens | Pinaceae | ||
| 78. | Xyleborus septentrionalis | Pinaceae | ||
| 79. | Xyleborus spinulosus | Pinaceae | ||
| 80. | Xyleborus volvulus | Araucariaceae | 15 | Haack and Rabaglia (2013) |
| 81. | Xylechinosomus lucianae | Araucariaceae | ||
| 82. | Xylechinus araucariae | Araucariaceae | ||
| 83. | Xylechinus montanus | Pinaceae |
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard C, Dehnen‐Schmutz K, Di Serio F, Gonthier P, Jacques M‐A, Jaques Miret JA, Justesen AF, MacLeod A, Magnusson CS, Navas‐Cortes JA, Parnell S, Potting R, Reignault PL, Thulke H‐H, Van der Werf W, Civera AV, Yuen J, Zappalà L, Grégoire J‐C, Kertész V, Streissl F and Milonas P, 2020. Scientific Opinion on the pest categorisation of non‐EU Scolytinae of coniferous hosts. EFSA Journal 2020;18(1):5934, 39 pp. 10.2903/j.efsa.2020.5934
Requestor: European Commission
Question number: EFSA‐Q‐2019‐00425
Panel members: Claude Bragard, Katharina Dehnen‐Schmutz, Francesco Di Serio, Paolo Gonthier, Marie‐Agnès Jacques, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L Reignault, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent Civera, Jonathan Yuen and Lucia Zappalà.
Acknowledgments: EFSA wishes to acknowledge the contribution of Massimo Faccoli from the Department of Agronomy, Food, Natural Resources, Animals and the Environment at the University of Padua who contributed via a contract for a literature review and data collection to this opinion.
Adopted: 20 November 2019
Notes
Council Directive 2000/29/EC of 8 May 2000 on protective measures against the introduction into the Community of organisms harmful to plants or plant products and against their spread within the Community. OJ L 169/1, 10.7.2000, p. 1–112.
Regulation (EU) 2016/2031 of the European Parliament of the Council of 26 October 2016 on protective measures against pests of plants. OJ L 317, 23.11.2016, p. 4–104.
Regulation (EC) No 178/2002 of the European Parliament and of the Council of 28 January 2002 laying down the general principles and requirements of food law, establishing the European Food Safety Authority and laying down procedures in matters of food safety. OJ L 31/1, 1.2.2002, p. 1–24.
Council of the European Union, 2003. Regulation (EC) No 2152/2003 of the European Parliament and of the Council of 17 November 2003 concerning monitoring of forests and environmental interactions in the Community (Forest Focus). Official Journal of the European Union 46 (L 324), 1–8.
References
- Alamouti SM, Tsui CK and Breuil C, 2009. Multigene phylogeny of filamentous ambrosia fungi associated with ambrosia and bark beetles. Mycological Research, 113, 822–835. 10.1016/j.mycres.2009.03.003 [DOI] [PubMed] [Google Scholar]
- Alonso‐Zarazaga MA, Barrios H, Borovec R, Bouchard P, Caldara R, Colonnelli E, Gültekin L, Hlavác P, Korotyaev B, Lyal CHC, Machado A, Meregalli M, Pierotti H, Ren L, Sánchez‐Ruiz M, Sforzi A, Silfverberg H, Skuhrovec J, Trýzna M, de Castro Velázquez AJ and Yunakov NN, 2017, Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea. Monografías electrónicas S.E.A., vol. 8, Sociedad Entomológica Aragonesa S.E.A., Zaragoza, Spain, 729 pp. Available online: https://www.researchgate.net/publication/320895385_Cooperative_Catalogue_of_Palaearctic_Coleoptera_Curculionoidea [Accessed January 15 2019]
- ANSES , 2017. ANSES Opinion on “a request for an express risk assessment (ERA) on Xylosandrus compactus (Eichhoff) identified in metropolitan France”. Available online https://www.anses.fr/en/content/anses-opinion-request-express-risk-assessment-era-xylosandrus-compactus-eichhoff-identified
- Atkinson T, 2019. Bark and Ambrosia Beetles web‐page on Check‐lists of scolytids from Americas. Available online: https://www.barkbeetles.info/index.php
- Bossard M, Feranec J and Otahel J, 2000. CORINE land cover technical guide ‐ Addendum 2000. Tech. Rep. 40, European Environment Agency. Available online: https://www.eea.europa.eu/ds_resolveuid/032TFUPGVR, INRMM‐MiD:13106045
- Bright DE, 2014. A Catalog of Scolytidae and Platypodidae (Coleoptera), Supplement 3 (2000‐2010), with notes on subfamily and tribal reclassifications. Insecta Mundi, 861 Available online: http://digitalcommons.unl.edu/insectamundi/861 [Google Scholar]
- Bright DE and Skidmore RE, 2002. A catalogue of Scolytidae and Platypodidae (Coleoptera) Supplement 2 (1995–1999). NRC Research Press.
- British Columbia Government , 2019. MPB projections. Available online: https://www2.gov.bc.ca/gov/content/industry/forestry/managing-our-forest-resources/forest-health/forest-pests/bark-beetles/mountain-pine-beetle/mpb-projections?keyword=mountain%26keyword=pine%26keyword=beetle%26keyword=2016 [Accessed 21 July 2019] [Google Scholar]
- Brockerhoff GE, Bain J, Kimberley M and Knízek M, 2006. Interception frequency of exotic bark and ambrosia beetles (Coleoptera: Scolytinae) and relationship with establishment in New Zealand and worldwide. Canadian Journal of Forest Research, 36, 289–298. [Google Scholar]
- Brockerhoff EG, Kimberley M, Liebhold AM, Haack RA and Cavey JF, 2014. Predicting how altering propagule pressure changes establishment rates of biological invaders across species pools. Ecology, 95(3), 594–601. [DOI] [PubMed] [Google Scholar]
- Büttner G, Kosztra B, Maucha G and Pataki R, 2012. Implementation and achievements of CLC2006. Tech. rep., European Environment Agency. Available online: http://www.eea.europa.eu/ds_resolveuid/GQ4JECM8TB, INRMM‐MiD:14284151
- Chase KD, Kelly D, Liebhold AM, Bader MKF and Brockerhoff EG, 2017. Long‐distance dispersal of non‐native pine bark beetles from host resources. Ecological Entomology, 42, 173–183. 10.1111/een.12371 [DOI] [Google Scholar]
- Chirici G, Bertini R, Travaglini D, Puletti N and Chiavetta U, 2011a. The common NFI database In: Chirici G, Winter S. and McRoberts RE. (eds.). National forest inventories: contributions to forest biodiversity assessments. Springer, Berlin: pp. 99–119. [Google Scholar]
- Chirici G, McRoberts RE, Winter S, Barbati A, Brändli U‐B, Abegg M, Beranova J, Rondeux J, Bertini R, Alberdi Asensio I and Condés S, 2011b. Harmonization tests In: Chirici G, Winter S. and McRoberts RE. (eds.). National forest inventories: contributions to forest biodiversity assessments. Springer, Berlin: pp. 121–190. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health), Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Gregoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Hart A, Schans J, Schrader G, Suffert M, Kertesz V, Kozelska S, Mannino MR, Mosbach‐Schulz O, Pautasso M, Stancanelli G, Tramontini S, Vos S and Gilioli G, 2018. Guidance on quantitative pest risk assessment. EFSA Journal 2018;16(8):5350, 86 pp. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health), Bragard C, Dehnen‐Schmutz K, Di Serio F, Gonthier P, Jacques M‐A, Jaques Miret JA, Justesen AF, MacLeod A, Magnusson CS, Navas‐Cortes JA, Parnell S, Potting R, Reignault PL, Thulke H‐H, Van der Werf W, Civera AV, Yuen J, Zappalà L, Grégoire JC, Kertész V and Streissl Fand Milonas P, 2020. Scientific opinion on the list of non‐EU Scolytinae of coniferous hosts. In preparation.
- EPPO (European and Mediterranean Plant Protection Organization), online. EPPO Global Database. Available online: https://gd.eppo.int [Accessed January 9, 2019]
- Eveden ML, Whitehouse CM and Sykes J, 2014. Factors Influencing Flight Capacity of the Mountain Pine Beetle (Coleoptera: Curculionidae: Scolytinae). Environmental Entomology, 43, 187–196. [DOI] [PubMed] [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations), 1995. ISPM (International standards for phytosanitary measures) No 4. Requirements for the establishment of pest free areas. Available online: https://www.ippc.int/en/publications/614/
- FAO (Food and Agriculture Organization of the United Nations), 2004. ISPM (International Standards for Phytosanitary Measures) 21—Pest risk analysis of regulated non‐quarantine pests. FAO, Rome, 30 pp. Available online: https://www.ippc.int/sites/default/files/documents//1323945746_ISPM_21_2004_En_2011-11-29_Refor.pdf
- FAO (Food and Agriculture Organization of the United Nations), 2013. ISPM (International Standards for Phytosanitary Measures) 11—Pest risk analysis for quarantine pests. FAO, Rome, 36 pp. Available online: https://www.ippc.int/sites/default/files/documents/20140512/ispm_11_2013_en_2014-04-30_201405121523-494.65%20KB.pdf
- FAO (Food and Agriculture Organization of the United Nations), 2017. ISPM (International standards for phytosanitary measures) No 5. Glossary of phytosanitary terms. Available online: https://www.ippc.int/en/publications/622/
- Forsse E and Solbreck CH, 1985. Migration in the bark beetle Ips typographus L.: duration, timing and height of flight. Zeitschrift für angewandte Entomologie, 100, 47–57. [Google Scholar]
- Galko J, Dzurenko M, Ranger CM, Kulfan J, Kula E, Nikolov C, Zúbrik M and Zach P, 2019. Distribution, Habitat Preference, and Management of the Invasive Ambrosia Beetle Xylosandrus germanus (Coleoptera: Curculionidae, Scolytinae) in European Forests with an Emphasis on the West Carpathians. Forests, 10, 10 10.3390/f10010010 [DOI] [Google Scholar]
- Garonna AP, Dole SA, Mazzoleni S and Cristinzio G, 2012. First record of the black twig borer Xylosandrus compactus (Eichhoff) (Coleoptera: Curculionidae, Scolytinae) from Europe. Zootaxa, 3251, 64–68. [Google Scholar]
- Gomez D, Martinez G and Beaver R, 2012. First record of Cyrtogenius lutus (Blandford) (Coleoptera: Curculionidae: Scolytinae) in the Americas and its distribution in Uruguay. The Coleopterists Bulletin, 66, 362–364. [Google Scholar]
- Gomez DF, Rabaglia RJ, Fairbanks KEO and Hulcr J, 2018. North American Xyleborini north of Mexico: a review and key to genera and species (Coleoptera, Curculionidae, Scolytinae). ZooKeys, 19–68. 10.3897/zookeys.768.24697 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grégoire J‐C, Raffa KF and Lindgren BS, 2015. Economics and politics of bark beetles In: Vega FE. and Hofstetter RW. (eds.). Bark Beetles. Biology and Ecology of Native and Invasive Species. Elsevier/Academic Press, London, UK: pp. 585–613. [Google Scholar]
- Haack RA, 2006. Exotic bark‐ and wood‐boring Coleoptera in the United States: recent establishments and interceptions. Canadian Journal of Forest Research, 36, 269–288. [Google Scholar]
- Haack RA and Rabaglia RJ, 2013. Exotic bark and ambrosia beetles In: Pena JE. (ed.). Potential invasive pests of agricultural crops. CABI International, Wallingford, UK, Chapter 3. pp. 48–74. [Google Scholar]
- Hiederer R, Houston Durrant T, Granke O, Lambotte M, Lorenz M, Mignon B and Mues V, 2007. Forest focus monitoring database system ‐ validation methodology. Vol. EUR 23020 EN of EUR – Scientific and Technical Research. Office for Official Publications of the European Communities. 10.2788/51364 [DOI]
- Hiederer R, Houston Durrant T and Micheli E, 2011. Evaluation of BioSoil demonstration project ‐ Soil data analysis. Vol. 24729 of EUR ‐ Scientific and Technical Research. Publications Office of the European Union. 10.2788/56105 [DOI]
- Hofstetter RW, Dinkins‐Bookwalter J, Davis TS and Klepzig KD, 2015. Symbiotic Associations of Bark Beetles In Vega FE, Hofstetter RW. (eds.). Bark Beetles. Biology and Ecology of Native and Invasive Species. Elsevier/Academic Press, London, UK, pp. 209–245. [Google Scholar]
- Houston Durrant T and Hiederer R, 2009. Applying quality assurance procedures to environmental monitoring data: a case study. Journal of Environmental Monitoring, 11, 774–781. [DOI] [PubMed] [Google Scholar]
- Houston Durrant T, San‐Miguel‐Ayanz J, Schulte E and Suarez Meyer A, 2011. Evaluation of BioSoil demonstration project: forest biodiversity ‐ Analysis of biodiversity module. Vol. 24777 of EUR – Scientific and Technical Research. Publications Office of the European Union. 10.2788/84823 [DOI]
- Hughes MA, Riggins JJ, Koch FH, Cognato AI, Anderson C, Formby JP, Dreaden T, Ploetz R and Smith JA, 2017. No rest for the laurels: symbiotic invaders cause unprecedented damage to southern USA forests. Biological invasions, 19, 2143–2157. [Google Scholar]
- Hulcr J, Atkinson TH, Cognato AI, Jordal BH and McKenna DD, 2015. Morphology, taxonomy, and phylogenetics of bark beetles In Vega FE and Hofstetter RW (eds.). Bark Beetles. Biology and Ecology of Native and Invasive Species. Elsevier/Academic Press, London, UK, pp. 41–84. [Google Scholar]
- Jackson PL, Straussfogel D, Lindgren BS, Mitchell S and Murphy B, 2008. Radar observation and aerial capture of mountain pine beetle, Dendroctonus ponderosae Hopk. (Coleoptera: Scolytidae) in flight above the forest canopy. Canadian Journal of Forest Research, 38, 2313–2327. 10.1139/X08-066 [DOI] [Google Scholar]
- Jactel H and Gaillard J, 1991. A preliminary study of the dispersal potential of Ips sexdentatus (Boern)(Col., Scolytidae) with an automatically recording flight mill. Journal of Applied Entomology, 112, 138–145. [Google Scholar]
- de Jong Y, Verbeek M, Michelsen V, Bjørn P, Los W, Steeman F, Bailly N, Basire C, Chylarecki P, Stloukal E, Hagedorn G, Wetzel F, Glöckler F, Kroupa A, Korb G, Hoffmann A, Häuser C, Kohlbecker A, Müller A, Güntsch A, Stoev P and Penev L, 2014. Fauna Europaea ‐ all European animal species on the web. Biodiversity DataJournal, 2, e4034. 10.3897/bdj.2.e4034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkendall LR, Biedermann PHW and Jordal BH, 2015Evolution and Diversity of Bark and Ambrosia Beetles In Vega FE. and Hofstetter RW. (eds.), Bark Beetles. Biology and Ecology of Native and Invasive Species. Elsevier/Academic Press, London, UK: pp.85–156. [Google Scholar]
- Knizek M and Beaver R, 2004. Taxonomy and systematics of bark and ambrosia beetles In: Lieutier F, Day K, Battisti A, Grégoire JC. and Evans H. (eds.). Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis. Kluwer, Dordrecht. pp. 41–54. 10.1007/978-1-4020-2241-8_11 [DOI] [Google Scholar]
- Lawson SA, Carnegie AJ, Cameron N, Wardlaw T and Venn TJ, 2018. Risk of exotic pests to the Australian forest industry. Australian Forestry, 81(1), 3–13. [Google Scholar]
- MacLeod A and Korycinska A, 2019. Detailing Köppen‐Geiger climate zones at sub‐national to continental scale: a resource for pest risk analysis. EPPO Bullletin, 49, 73–82. [Google Scholar]
- Nilssen AC, 1984. Long‐range aerial dispersal of bark beetles and bark weevils (Coleoptera, Scolytidae and Curculionidae) in northern Finland. Annales Entomologici Fennici, 50, 37–42. [Google Scholar]
- Raffa KF, Gregoire J‐C and Lindgren BS, 2015. Natural history and ecology of bark beetles In Vega FE. and, Hofstetter RW. (eds.). Bark Beetles. Biology and Ecology of Native and Invasive Species. Elsevier/Academic Press, London, UK: pp. 1–40. [Google Scholar]
- de Rigo D, 2012. Semantic Array Programming for environmental modelling: application of the Mastrave library. In: Seppelt R, Voinov AA, Lange S and Bankamp D (eds.). International Environmental Modelling and Software Society (iEMSs) 2012 International Congress on Environmental Modelling and Software ‐ Managing Resources of a Limited Planet: Pathways and Visions under Uncertainty, Sixth Biennial Meeting. pp. 1167–1176.
- de Rigo D, Caudull G, Busetto L, San‐Miguel‐Ayanz J, 2014. Supporting EFSA assessment of the EU environmental suitability for exotic forestry pests: final report. EFSA Supporting Publications 11 (3), EN‐434+. 10.2903/sp.efsa.2014.en-434 [DOI] [Google Scholar]
- de Rigo D, Caudullo G, Houston Durrant T and San‐Miguel‐Ayanz J, 2016. The European Atlas of Forest Tree Species: modelling, data and information on forest tree species. In: San‐Miguel‐Ayanz J, de Rigo D, Caudullo G, Houston Durrant T and Mauri A (eds.). European Atlas of Forest Tree Species. Publ. Off. EU, Luxembourg, pp. e01aa69+ Available online: https://w3id.org/mtv/FISE-Comm/v01/e01aa69
- de Rigo D, Caudullo G, San‐Miguel‐Ayanz J and Barredo JI, 2017. Robust modelling of the impacts of climate change on the habitat suitability of forest tree species. Publication Office of the European Union, 58 pp. ISBN:978‐92‐79‐66704‐6, 10.2760/296501 [DOI]
- San‐Miguel‐Ayanz J, 2016. The European Union Forest Strategy and the Forest Information System for Europe. In: San‐Miguel‐Ayanz J, de Rigo D, Caudullo G, Houston Durran T and Mauri A (eds.). European Atlas of Forest Tree Species Publ. Off. EU, Luxembourg, pp. e012228+ Available online: https://w3id.org/mtv/FISE-Comm/v01/e012228
- San‐Miguel‐Ayanz J, de Rigo D, Caudullo G, Houston Durrant T and Mauri A (eds.), 2016. European Atlas of Forest Tree Species. Publication Office of the European Union, Luxembourg. ISBN: 978‐92‐79‐36740‐3, Available online: https://w3id.org/mtv/FISE-Comm/v01
- Schedl KJF, Harde L and Die Käfer Mitteleuropas B, 1981. Scolytidae, Borkenkäfer. 10, 34–101. Goecke & Evers. [Google Scholar]
- Spanou K, Marathianou M, Gouma M, Dimou D, Nikoletos L, Milonas PG and Papachristos DP, 2019. First record of black twig borer Xylosandrus compactus (Coleoptera: Curculionidae) in Greece. 18th Panhellenic Entomological Congress, KOMOTINI 15‐17/10/2019, abstract page 77. [Google Scholar]
- Stauffer C, Kirisits T, Nussbaumer C, Pavlin R and Wingfield MJ, 2001. Phylogenetic relationships between the European and Asian eight spined larch bark beetle populations (Coleoptera: Scolytidae) inferred from DNA sequences and fungal associates. European Journal of Entomology, 98, 99–106. [Google Scholar]
- Wood SL, 1982. The bark and ambrosia beetles of North and Central America (Coleoptera Scolytidae), a Taxonomic monograph. Great Basin Nat. Mem., 6, 1–1356. [Google Scholar]
- Wood SL, 2007. Bark and Ambrosia Beetles of South America. (Coleoptera: Scolytidae). Bark and ambrosia beetles of South America. Monte L. Bean Sci. Mus., Provo, Utah: 1–900.
- Wood SL and Bright DE, 1992. A catalogue of Scolytidae and Platypodidae Part 2 Taxonomic index (Volumes A, B). Great Basin Nat. Mem., 13, 1–1553. [Google Scholar]