

Abstract
The Plant Health Panel reviewed the paper by Guarnaccia et al. (2017) and compared their findings with previous predictions on the establishment of Phyllosticta citricarpa. Four species of Phyllosticta were found by Guarnaccia et al. (2017) in Europe. P. citricarpa and P. capitalensis are well‐defined species, with P. citricarpa recorded for the first time in Europe, confirming predictions by Magarey et al. (2015) and EFSA (2008, 2014, 2016) that P. citricarpa can establish in some European citrus‐growing regions. Two new species P. paracitricarpa and P. paracapitalensis were also described, with P. paracitricarpa (found only in Greece) shown to be pathogenic on sweet orange fruits. Genotyping of the P. citricarpa isolates suggests at least two independent introductions, with the population in Portugal being different from that present in Malta and Italy. P. citricarpa and P. paracitricarpa were isolated only from leaf litter in backyards. However, since P. citricarpa does not infect or colonise dead leaves, the pathogen must have infected the above living leaves in citrus trees nearby. Guarnaccia et al. (2017) considered introduction to be a consequence of P. citricarpa having long been present or of illegal movement of planting material. In the Panel's view, the fruit pathway would be an equally or more likely origin. The authors did not report how surveys for citrus black spot (CBS) disease were carried out, therefore their claim that there was no CBS disease even where the pathogen was present is not supported by the results presented. From previous simulations, the locations where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa were conducive for P. citricarpa establishment, with number of simulated infection events by pycnidiospores comparable to sites of CBS occurrence outside Europe. Preliminary surveys by National Plant Protection Organisations (NPPOs) have not confirmed so far the findings by Guarnaccia et al. (2017) but monitoring is still ongoing.
Keywords: leaf litter, backyard, citrus black spot, Phyllosticta citricarpa, Phyllosticta paracitricarpa, European Union, single mating type
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
The European Commission has been recently informed by EFSA of the availability on line (http://www.sciencedirect.com/science/article/pii/S0166061617300234) of an accepted manuscript that will be published in the near future in Studies of Mycology entitled “First report of Phyllosticta citricarpa and description of two new species, P. paracapitalensis and P. paracitricarpa, from citrus in Europe”. This document described the identification of Phyllosticta citricarpa in Italy, Malta and Portugal and other species found in Europe.
EFSA is requested, pursuant to Article 29 of Regulation (EC) No 178/2002, to prepare an urgent evaluation of this scientific paper, i.e. before the end of September 2017. In particular, EFSA is asked, in view of current scientific knowledge, to review the methodologies used in this paper, as well as the analysis of the results and the related discussion. In addition, EFSA is requested to report and discuss how these findings fit the predictions of the model simulations on the establishment of P. citricarpa in the EU.
1.2. Interpretation of the Terms of Reference
Phyllosticta citricarpa (McAlpine) van der Aa is officially absent from the EU and is a harmful organism listed in Directive 2000/29/EC, whereas Phyllosticta capitalensis Henn., which is morphologically similar to P. citricarpa, is a non‐pathogenic endophyte, commonly isolated from citrus tissues and other hosts and is known to occur in the European Union (Wikee et al., 2013). P. paracapitalensis Guarnaccia & Crous, sp. nov. and P. paracitricarpa Guarnaccia & Crous, sp. nov. are proposed by Guarnaccia et al. (2017) as new fungal species and have never been previously reported in Europe.
In this Scientific Opinion, the Panel has evaluated the paper by Guarnaccia et al. (2017), but it has not assessed the regulatory status of the harmful plant pathogenic fungus P. citricarpa. Such activities are within the competence of the National Plant Protection Organisations (NPPOs).
The Terms of reference required EFSA to undertake two distinct tasks, and these are assessed in separate chapters of this opinion:
the evaluation of the paper by Guarnaccia et al. (2017) (see Section 3.1)
comparison of the findings by Guarnaccia et al. (2017) with previous model predictions on the establishment of P. citricarpa in the European Union (see Section 3.2).
Since the newly described species P. paracitricarpa has been shown by Guarnaccia et al. (2017) to induce symptoms of citrus black spot (hereafter referred to as CBS) in artificially inoculated sweet orange (Citrus sinensis Osbeck) fruits, for the second task the Panel considered P. paracitricarpa as well as P. citricarpa.
1.3. Additional information
Before the publication of this Scientific Opinion, EFSA has already provided risk assessment and scientific advice on P. citricarpa for the EU territory. The principal EFSA outputs on this topic are listed below:
EFSA (European Food Safety Authority), 2008. Scientific Opinion of the Panel on Plant Heath (PLH) on a request from the European Commission on Guignardia citricarpa Kiely. EFSA Journal 2008;7(1):925, 108 pp. https://doi.org/10.2903/j.efsa.2009.925
EFSA PLH Panel (EFSA Plant Health Panel), 2014. Scientific Opinion on the risk of Phyllosticta citricarpa (Guignardia citricarpa) for the EU territory with identification and evaluation of risk reduction options. EFSA Journal 2014;12(2):3557, 243 pp. https://doi.org/10.2903/j.efsa.2014.3557
EFSA PLH Panel (EFSA Plant Health Panel), 2016. Evaluation of new scientific information on Phyllosticta citricarpa in relation to the EFSA PLH Panel (2014) Scientific Opinion on the plant health risk to the EU. EFSA Journal 2016;14(6):4513, 53 pp. https://doi.org/10.2903/j.efsa.2016.4513
To reduce the key risk assessment uncertainties, EFSA also outsourced research on the splash dispersal of P. citricarpa asexual spores (conidia, hereafter referred to as pycnidiospores) from artificially infected citrus fruit. Such work can be consulted in the following publications:
Perryman SAM and West JS, 2014. Splash dispersal of Phyllosticta citricarpa pycnidiospores from infected citrus. EFSA supporting publication 2014‐EN‐560, 30 pp.
Perryman SAM, Clark SJ and West JS, 2014. Splash dispersal of Phyllosticta citricarpa conidia from infected citrus fruit. Scientific Reports, 4, 6568.
2. Data and methodologies
2.1. Data
The data used in this Scientific Opinion are provided:
in the Tables published by Guarnaccia et al. (2017); the geographical locations are given in Table 1 of the paper by Guarnaccia et al. (2017);
in publications reporting the results of previous model simulations for the establishment of P. citricarpa in Europe (EFSA, 2008; Yonow et al., 2013; EFSA PLH Panel, 2014, 2016; Magarey et al., 2015).
Table 1.
Summary of locations and characteristics of the investigated sites as from Table 1 by Guarnaccia et al. (2017)
| Country | Total | Number of sites investigated by Guarnaccia et al. (2017) | ||||
|---|---|---|---|---|---|---|
| Backyards | Botanical gardens | Experimental orchards | Nurseries | Orchards | ||
| Greece | 22 | 2 | 0 | 0 | 0 | 20 |
| Italy | 36 | 11 | 0 | 0 | 5 | 20 |
| Malta | 6 | 4 | 0 | 0 | 0 | 2 |
| Portugal | 10 | 4 | 0 | 0 | 0 | 6 |
| Spain | 21 | 1 | 2 | 3 | 0 | 15 |
| Total | 95 | 22 | 2 | 3 | 5 | 63 |
In Table C.1 of Appendix C – Data provenance, the locations are shown for the data files on which the maps of this Scientific Opinion are based on. The data files are deposited on http://zenodo.org, as 10.5281/zenodo.1003121 (https://doi.org/10.5281/zenodo.1003121).
Table C.1.
Locations of data files on which figures of this Scientific Opinion are based on
| Figure | map file name | input data file |
|---|---|---|
| 1 | locationDensity.png | locations.csv |
| euCitrusSurface.rds | ||
| 3 | maghareyAscLocations.png | euCitrusSurface.rds |
| mag2015_table 1.csv | ||
| locations.csv | ||
| 4 | asc3_15_5m.png | locations.csv |
| mag2015_table 1.csv | ||
| 5 | asc3_15_7m.png | locations.csv |
| asco_avg_WSPR.zip/Asco_3_15_R7_Model_AVG_WSPR.xlsx | ||
| 6 | asc3_15_12m.png | locations.csv |
| asco_avg_WSPR.zip/Asco_3_15_R7_Model_AVG_WSPR.xlsx | ||
| 7 | maghareyPycLocations.png | euCitrusSurface.rds |
| locations.csv | ||
| mag2015_table 1.csv | ||
| 8 | pyc3_10_5m.png | locations.csv |
| pycnidio_avg_WR.zip/Pycnidio_3_10_Model_AVG_WR.xlsx | ||
| 9 | pyc3_10_7m.png | pyc3_10_7m.png |
| pycnidio_avg_WR.zip/Pycnidio_3_10_Model_AVG_WR.xlsx | ||
| 10 | pyc3_10_12m.png | locations.csv |
| pycnidio_avg_WR.zip/Pycnidio_3_10_Model_AVG_WR.xlsx | ||
| 12 | climateGI_climex.png | Yonow_et_al_2013_CLIMEX_with_10minresolution_1961‐1990.zip |
| locations.csv | ||
| 13 | climateEI_climex.png | Yonow_et_al_2013_CLIMEX_with_10minresolution_1961‐1990.zip |
| locations.csv | ||
| 14 | climateGI_jrc.png | Yonow_et_al_2013_CLIMEX_with_10minresolution_1961‐1990.zip |
| locations.csv | ||
| 15 | climateEI_jrc.png | Yonow_et_al_2013_CLIMEX_with_10minresolution_1961‐1990.zip |
2.2. Methodologies
When conducting this assessment, the Panel also took into consideration the following EFSA horizontal guidance documents:
Guidance of the Scientific Committee on Transparency in the Scientific Aspects of risk assessments carried out by EFSA. Part 2: General Principles (EFSA, 2009);
Guidance on the structure and content of EFSA's scientific opinions and statements (EFSA Scientific Committee, 2014);
Guidance on uncertainty (EFSA Scientific Committee, 2016).
3. Assessment
3.1. Evaluation of a paper by Guarnaccia et al. (2017)
The paper has three principal components that have been evaluated according to: (1) the methodology used; (2) the results; (3) the discussion of the results; additionally, (4) some other considerations relating to the evaluation have been raised. The paper has four distinct but inter‐related lines of enquiry: the sampling sites used for isolation of Phyllosticta spp.; the identification of known and new species using morphological and DNA‐based criteria; the inferences made on the origin of introduction; and the relationships between the isolations made and CBS disease at the sampling sites. The Panel first notes that the Institute involved in the designation of the two new Phyllosticta species is at the forefront of fungal taxonomy and the Panel is of the view that the new species designations are well‐founded and make a major contribution to the taxonomy of the genus Phyllosticta. The Panel also notes that the ‘One fungus‐one name’ principle in nomenclature has been criticised by some mycologists (Gams and Jaklitsch, 2011). As in other PLH Panel opinions, the Panel accepts the principle of ‘one fungus‐one name’.
3.1.1. Evaluation of Methodology
The paper by Guarnaccia et al. (2017) describes results obtained from samples taken from 95 locations in a wide range of citrus hosts and associated environments such as leaf litter in southern Europe from 2015 to 2017 to assess for the presence of Phyllosticta species.
The locations of sample sites were reported with GPS coordinates in decimal degrees to 6 decimal places, allowing them to be mapped and revisited if necessary, but without mention of the method used to record these coordinates. This implies a spatial resolution of approximately 0.1 m. However, to obtain such a level of precision requires professional GPS equipment with differential correction. The accuracy of the GPS on mobile phones depends on the model and the location. Zandbergen and Barbeau (2011) found that the mobile phones they tested had a median horizontal error of between 5.0 and 8.5 m. Thus, depending on the type of GPS used by Guarnaccia et al. (2017) and the type of GPS available to other teams undertaking the resampling of the locations, it may be difficult to re‐identify the precise tree that yielded the positive samples found by Guarnaccia et al. (2017), particularly if several trees are located in the near vicinity.The sampling produced 64 isolates of Phyllosticta, of which 52 were selected for further morphological and molecular characterisation. A multilocus phylogenetic analysis, using partial DNA sequences from genes encoding the rDNA internal transcribed spacer region (ITS), actin (actA), the translation elongation factor 1‐α (tef1), glyceraldehyde‐3‐phosphate dehydrogenase (gapdh), the 28S large subunit nrDNA (LSU) and the RNA polymerase II second largest subunit (rpb2), was carried out on many Phyllosticta species (99 isolates in total, including the 52 selected test isolates). Growth rates at 9–39°C at 3°C intervals were assessed for a subset of isolates of the Phyllosticta species collected. The number of isolates of each species in the subset was not given.
Some details of the methods used were either missing or unclear:
It was not clear from the methodology presented in the paper how the sample locations and sites (95 in total) were chosen as well as how different fruits (approximately 25), twig portions (25), living leaves (50) and litter layer leaves (50) from each Citrus host were collected at each site.
The dates of sampling for the isolations done in 2015 and 2016 were not reported. The number of host plants sampled at each location was not given. It was only stated that a certain number of leaves, twigs or fruit per host was sampled, i.e. we do not know whether the 50 living leaves were from one tree or many trees found at a site or whether leaves from the leaf litter were collected beneath one tree or many trees. The approach of trying to isolate from plant material has been shown to be less effective in detecting the presence of the pathogen than established molecular diagnostic methods using macerated or powdered leaves (Bonants et al., 2003; Meyer et al., 2012; Hu et al., 2014). With the methods they used, symptoms would only be expected to be visible on certain plant tissues at particular times of the year depending on the location, citrus species and cultivar.
New field visits, in the sites where P. citricarpa or P. paracitricarpa were found in 2015 and 2016, were made between December 2016 and January 2017 to confirm the initial findings and to assay for the presence of symptoms on fruit, leaves and twigs. However, the Panel notes that this period would have been too early for symptoms to be visible on fruit for most citrus varieties in southern Europe (see Figure 51 in EFSA PLH Panel, 2014). These follow‐up visits in December 2016 to January 2017 were simply reported in the methods section but no results were presented in the paper (from personal communication from the corresponding author, the Panel was informed that no samples were taken; for details, please see Other considerations, Section 3.1.4).
The number of isolation points attempted per sampled unit (leaf, fruit, twig) was not given, although the discussion mentions that P. citricarpa was found at a very low frequency in a few of the sites investigated. As P. citricarpa was only found in leaf litter leaves and approximately 50 were tested for the presence of P. citricarpa at each site, a frequency of the percentage of positive isolations could have been provided.
In the material and methods, it was stated that a total of 111 Phyllosticta (incl. 64 European) isolates were included in the study (Table 2 of Guarnaccia et al., 2017) of which 100 (including the out‐group) were used in the phylogenetic analysis. However, the discussion indicated that a subset of 52 European isolates was compared to several reference isolates using partial gene sequences of six different loci, as well as morphological characteristics. This was based on a comparison with sequences retrieved from GenBank of an additional 43 isolates. It is not stated why 12 European isolates were not sequenced (eight P. citricarpa isolates and four P. paracitricarpa isolates were not used). For the four isolates from Malta that underwent multilocus sequence analysis, it was also not clear whether the isolates used came from both or only one of the two possible locations in Malta, since the isolate location was not given in Table 2.
-
No explanation is given for how the CBS disease surveys were conducted:
-
–
whether the numbers of trees and locations were chosen systematically or randomly;
-
–
whether disease surveys were conducted in or close to the vicinity of the sampling sites;
-
–
which methods were used to decide on the time the surveys were conducted. Symptoms would only be expected to be visible on certain plant tissues at certain times of the year, varying with location, citrus species and cultivar;
-
–
whether the second set of field visits conducted in December 2016 and January 2017 was accompanied by a disease survey. It was intended to revisit the sites that had previously tested positive in order to confirm the findings and to assay for the presence of symptoms on fruit, leaves and twigs. However, this period, as explained above, depending on the citrus cultivar, may be not optimal for symptom expression on fruit;
-
–
why some important citrus production areas and sites at risk of infection, e.g. those identified by Magarey et al. (2015) or EFSA PLH Panel (2014), were not visited. In order to relate the findings to climate‐based risk, information on the type of irrigation system used at the sites selected would have been very useful.
-
–
Without properly constituted disease surveys, there is little support for the conclusion that the P. citricarpa isolates did not naturally cause disease.
The pathogenicity tests did not include a positive control, viz. inoculation with a known pathogenic isolate; the tests were terminated after 25 days, while a relatively longer period might have been needed to obtain typical CBS lesions and subsequent pycnidia (Perryman et al., 2014).
A minor technical point is that it is stated that 100 mL of spore suspension was inoculated onto fruit but this must have been 100 μL.
Table 2.
Summary of differences in the spatial and temporal factors among the three models
| Models, locations and climatic data | Yonow et al. (2013) and its applications by EFSA PLH Panel (2014) | EFSA PLH Panel (2014) | Magarey et al. (2015) |
|---|---|---|---|
| Models | CLIMEX Compare Locations |
Ascospore maturation and release Fourie et al. (2013) Ascospore/pycnidiospore infection Magarey et al. (2005) |
Ascospore maturation and release Fourie et al. (2013) Ascospore/pycnidiospore infection Magarey et al. (2005) |
| Locations modelled, spatial resolution and extent |
Global 0.5° latitude × 0.5° longitude grid cells. The EFSA PLH Panel (2014) used the same model also for 1518 25 × 25 km citrus grid cells and for 0.1° latitude and 0.1° latitude grid cells for the EU. |
One thousand five hundred and eighteen 25 × 25 km grid cells in Europe covering all European NUTS Level 3 regions with citrus production |
36 locations in Europe: Cyprus (1), France (2), Greece (7), Italy (15), Malta (1), Portugal (5) and Spain (5) |
| Source of weather/climatic data |
CRU CL1.0 New et al. (1999). The EFSA PLH Panel (2014) used the same model with MCYFS climatic data and with CRU CL1.0 1961–1990 climatology for 0.1° latitude and 0.1° latitude grid cells (New et al., 2002). |
The JRC Mars Crop Yield Forecasting System (MCYFS) grid cells interpolated to 25 × 25 km based on the mean agricultural altitude Agri4castWiki (2015) | NCEP CFSR global database interpolated to 38 × 38 km based on the mean grid elevation |
| Time period of weather/climatic data |
1961–1990. The EFSA PLH Panel (2014) used the same model with data from 1998 to 2007 |
1989–2009 | 2003–2011 |
| Temporal resolution of weather/climatic period utilised |
Monthly. The EFSA PLH Panel (2014) used the same model with daily MCYFS data. |
Daily for MCYFS data. Bregaglio et al. (2010) was used to generate hourly values of relative humidity. Hourly for 10 Italian and 14 Spanish agro‐meteorological stations |
Hourly |
3.1.2. Evaluation of Results
3.1.2.1. New species/taxonomy
A Bayesian tree based on ITS partitioning constructed for the genus Phyllosticta (isolates and/or sequences obtained from 7 European and 12 non‐European countries) resolved 15 species, of which two, P. paracapitalensis and P. paracitricarpa (previously considered a subclade of P. citricarpa), named after their close resemblance to P. capitalensis and P. citricarpa, respectively, were new species. Of the 15 species resolved, only these four were found from the locations in Europe (P. citricarpa was isolated from leaf litter in Italy, Malta and Portugal; P. paracitricarpa from leaf litter in Greece). Sequence analysis showed the presence of two previously described species, P. capitalensis (from asymptomatic living leaves of Citrus spp.) in Greece, Italy, Malta, Portugal and Spain, and P. citricarpa (from leaf litter of C. sinensis and C. limon) in Italy, Malta and Portugal. In addition, two new species were described, P. paracapitalensis (from asymptomatic living leaves of Citrus spp.) in Italy and Spain, and P. paracitricarpa (from leaf litter of C. limon) in Greece. As indicated above, the Panel is of the view that the new species designations are well‐founded.
Nucleotide variation was observed when comparing P. paracapitalensis and P. capitalensis (20 fixed nucleotide changes) and between P. paracitricarpa and P. citricarpa (seven fixed nucleotide changes). One nucleotide change distinguishing P. capitalensis from P. paracapitalensis, position 628, was wrongly marked in bold (Table 3) but it was not counted as one of the 20 nucleotide changes. Morphological observations also supported the distinction between the two known species and their sister novel species. The species P. capitalensis and P. paracapitalensis grew faster than P. citricarpa and P. paracitricarpa in the mycelial growth rate study, although all four species had similar cardinal temperatures. Formal descriptions of the two new species are provided, including the designation of holotypes and culture ex‐types. Mating type identifications were made for P. citricarpa but not for P. paracitricarpa. The P. citricarpa isolates found at sites in Italy and Malta were identical multilocus genotypes (MLG) with mating type MAT1‐2‐1, while additionally isolates from sites in Portugal were a different single MLG (with mating type MAT1‐1‐1). At each site, only a single mating type was found.
Table 3.
Model outputs from Yonow et al. (2013) for grid cells corresponding to the locations in Europe where Guarnaccia et al. (2017) found Phyllosticta citricarpa or P. paracitricarpa, run with different spatial and temporal resolution interpolated climate datasets: 0.5° climate dataset applied by Yonow et al. (2013), with the 0.1° and JRC Mars 25 km
| Locations where P. citricarpa or P. paracitricarpa were found by Guarnaccia et al. (2017) | Yonow et al. (2013), run on 0.1° climatic dataset | Yonow et al. (2013), run on 25 km JRC Mars climatic dataset | |||||
|---|---|---|---|---|---|---|---|
| Country | Location | Latitude | Longitude | Growth Index | Ecoclimatic Index | Growth Index | Ecoclimatic Index |
| Malta | Zurrieq | 35.823845 | 14.505099 | n.a. | n.a. | 4 | 2 |
| Malta | Gozo | 36.055138 | 14.259907 | 3 | 1 | 5 | 2 |
| Italy | Trebisacce | 39.910122 | 16.564824 | 3 | 0 | 6 | 1 |
| Portugal | Monchique | 37.326195 | −8.526232 | 1 | 0 | 2 | 1 |
| Greece | Arta | 39.160465 | 20.918257 | 6 | 1 | 7 | 2 |
3.1.2.2. Pathogenicity tests
Lesions developed on sweet orange fruits inoculated with two isolates of P. citricarpa. The lesions were considered by the authors to be atypical compared to those seen in CBS disease. The pathogen was consistently re‐isolated from these lesions. The Panel, however, observes that these lesions resembled some of those in the plates of the EPPO/OEPP (2009) and ISPM 27 diagnostic protocols and therefore were not atypical. Known pathogenic P. citricarpa isolates from other continents were not included in the pathogenicity tests; under the experimental conditions used, known pathogenic isolates could have induced similar or more typical lesions compared to the European isolates. No symptoms were observed by the authors on fruits inoculated with two isolates of P. capitalensis or two of P. paracapitalensis; however, this is not immediately obvious from their Figures 6C and 6D. Moreover, the water control was not shown. No attempt was made to re‐isolate from asymptomatic fruit (i.e. water controls or inoculated with P. capitalensis or P. paracapitalensis). The two isolates of P. paracitricarpa included in the pathogenicity tests were able to induce lesions (necrosis) in artificially inoculated mature sweet orange fruit and were consistently re‐isolated from these lesions.
3.1.2.3. Genotypic evaluation/connectivity of P. citricarpa
A distinction needs to be made between the isolates obtained and the populations from which the samples were taken. The isolate sampled may not be representing the diversity of the populations, especially when the number of isolates (64 across all sampling sites) is small. For this reason, as recognised by the authors, the populations can only be described as putative. The 20 P. citricarpa isolates obtained from the four localities in the three countries (as shown in Table 2 of Guarnaccia et al., 2017) were genotyped and two MLG were identified. The isolates from Malta (2) and from Italy (1) shared one MLG, the isolates from Portugal had a distinct MLG. The isolates from Portugal shared an MLG with three isolates from Australia. The MLG shared between Malta and Italy was different from those of isolates obtained from the other countries.
3.1.2.4. Sampling results
According to the sampling and isolation results, 64 Phyllostica isolates were recovered from 11 sites. Of these, 32 were obtained from fresh leaves, 28 were associated with leaf litter and 4 with leaf spot symptoms. Table 2 of Guarnaccia et al. (2017) shows that all 20 ‘pathogenic’ Phyllostica isolates (12 isolates of P. citricarpa from C. limon and C. sinensis and 8 isolates of P. paracitricarpa from C. limon) originated from leaf litter and the 4 ‘leaf spot’ isolates were associated with P. capitalensis on C. medica var. sarcodactylis (Wikee et al., 2013 suggest that P. capitalensis can be associated with lesions caused by insects) at a site in Italy.
Guarnaccia et al. (2017) indicated that during the periods when samples were taken from the different locations, no CBS symptoms were observed. However, the basis for this statement is not given. The methodology for disease surveys has been clearly described for epidemiological studies, including where first detection is the objective of the survey (Weltzien, 2012), and in the particular case where the results are to conform to IPPC requirements (Kalaris et al., 2014). Guarnaccia et al. (2017) describe their sampling procedures for isolating Phyllostica; however, no survey methodology for detecting disease is described. The symptoms of CBS are variable in appearance and often resemble those caused by other citrus pathogens or by insects, mechanical or cold damage, particularly in the case of freckle spot (EPPO/OEPP, 2009; ISPM 27, 2016). Symptoms that might be confused with those of CBS have previously been reported in the Mediterranean Basin, but these were caused by other pathogens, pests or abiotic disorders (Amat, 1988; Agustí et al., 2004; Vacante and Calabrese, 2009; Agustí, 2012).
After the initial samplings in 2015 and 2016, Guarnaccia et al. (2017) revisited during December 2016 and January 2017 the sites where P. citricarpa or P. paracitricarpa had been found. Details and results of the isolations eventually performed in this confirmatory sampling were not reported in the paper. However, from a personal communication by the corresponding author of the paper (see Other considerations, Section 3.1.4), the Panel was informed that no samples were taken. Guarnaccia et al. (2017) provided an overall statement saying that during the ‘surveys’ no CBS symptoms had been observed. Presumably this also applied to the second set of visits. However, as in 2015 and 2016, no information was provided on how these disease surveys were conducted.
Table 1 below summarises the 95 locations investigated by Guarnaccia et al. (2017) in Greece, Italy, Malta, Portugal and Spain. In Figure 1, the 95 locations are shown over a European map showing the density of citrus production areas. The countries with the highest number of locations investigated were Italy (36), Greece (22) and Spain (21). Portugal and Malta were represented by 10 and 6 locations, respectively. A total of 63 orchards were investigated and 22 backyards.1 Two botanical gardens and three experimental orchards were investigated in Spain. Five nurseries were investigated in Italy. Regarding the condition of the investigated locations, 48 orchards were cultivated and 15 were ‘unkept’ (defined as ‘plants abandoned’). From the 22 backyards investigated, 10 were cultivated and 12 were ‘unkept’. One botanical garden was cultivated and another ‘unkept’. The three experimental orchards and five nurseries investigated were all cultivated. Most of the locations investigated (69) had trees between 10 and 30 years old. Only 14 locations had trees younger than 10 years and 12 locations were older than 30 years. Based on this summary, it would appear that Guarnaccia et al. (2017) followed a stratified if uneven sampling scheme. For instance, some site types such as nurseries, botanical gardens and experimental orchards, were only sampled in two countries. Likewise, the number of backyards sampled was highly variable, from 1 out of 21 in Spain to 4 out of 6 in Malta.
Figure 1.
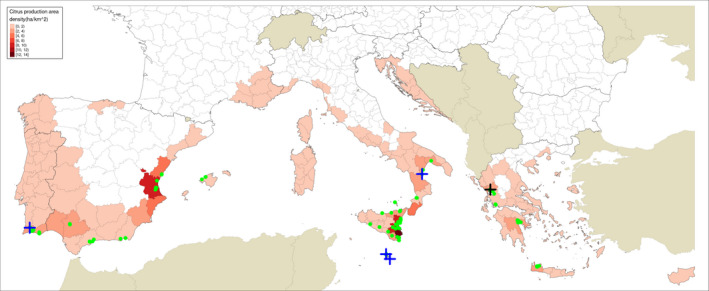
Locations sampled by Guarnaccia et al. (2017): blue crosses indicate sites where P. citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece; green dots indicate sites of sampling without positive finding. The map provides in the background the density of citrus production areas (in ha of citrus production areas per km2 of total area) for the European Union NUTS3 regions (as extracted from national statistical databases of Portugal, Spain, France, Italy, Malta, Croatia, Greece and Cyprus for year 2007; for details, see Appendix F of EFSA PLH Panel, 2014)
Guarnaccia et al. (2017) found P. citricarpa and P. paracitricarpa only in leaf litter samples. In Malta, P. citricarpa was detected in leaf litter samples in two backyards, one cultivated site with 60–70 years old trees at Gozo and another ‘unkept’ location with 15–25 years old trees at Zurrieq. In Italy, P. citricarpa was detected in leaf litter in a cultivated backyard with 20–30 years old trees at Trebisacce. In Portugal, P. citricarpa was detected at Monchique, in an ‘unkept’ backyard with 30–40 years old trees. The newly described species, P. paracitricarpa was detected in Arta, Greece, in a leaf litter sample from an ‘unkept’ backyard with 30–40 years old trees. Photos of the backyards and smallholdings from the sites corresponding to the geographical coordinates indicated by Guarnaccia et al. (2017) for its findings of P. citricarpa are shown in Figure 2.
Figure 2.

Photos of backyards and smallholdings from sites corresponding to the geographical coordinates indicated in Guarnaccia et al. (2017) for their findings of P. citricarpa. Top: (a) site in Zurrieq, Malta (Source: Google Earth, 5.823845 N, 14.505099 W, Eye altitude 200 m, Image: CNES/Airbus 2017 with approximate scale 1: 250); and (b) site at Zurrieq, Malta (by courtesy of Malta NPPO); Middle: (c) site at Zhagra, Gozo (Source: Google Earth, 36.055138 N, 14.259907 W, Eye altitude 200 m, Image: CNES/Airbus 2017 scale 1: 250); and (d) site at Zhagra, Gozo, (by courtesy of Malta NPPO); bottom; (e) site at Monchique, Algarve (by courtesy of Portugal NPPO); (f) site at Trebisacce, Calabria (Source: Google Earth, 39.910122 N, 16.564824 W, Eye altitude 80 m, Image: TerraMetrics 2017 with approximate scale 1:250)
P. citricarpa and P. paracitricarpa were found by Guarnaccia et al. (2017) in 4.2% and 1%, respectively, of the locations investigated. In Italy, P. citricarpa was found in 2.8% of all sites investigated (9% when considering only the backyards). Equivalent figures for Malta were 33.3% overall and 50% in the backyards. In Portugal, the findings were 10% overall and 25% in the backyards. The overall percentages for P. paracitricarpa in Greece were 4.5% overall and 50% when considering only the backyards. Despite the relative low number of sampled sites, the overall percentage of backyard sites where P. citricarpa was found was relatively high, particularly in Malta. The NPPOs are making follow up investigations at these locations.
As indicated in Section 3.1.1, the evaluations by Guarnaccia et al. (2017) were conducted by sampling fruits, twigs, living leaves and leaf litter from each citrus host present at each site investigated. P. citricarpa or P. paracitricarpa were only detected by Guarnaccia et al. (2017) in the leaf litter but not in living leaves in the canopy of the trees nearby. Isolations from the leaf litter are much more difficult to make than those from living leaves in the canopy because of the presence of soil microorganisms that may contaminate the samples (Frankland et al., 1990; Waller et al., 1998). Moreover, Truter et al. (2007) indicated that P. citricarpa pycnidiospores were not able to infect and colonise freshly detached green leaves or natural leaf litter. Therefore, leaf infections by P. citricarpa are restricted to those on living leaves in the canopy. Infected leaves will eventually fall down and form the leaf litter on the soil surface; therefore, it is unclear why no isolations were obtained from the living leaves.
3.1.3. Evaluation of Discussion
This study is a result of fresh collections of several Phyllosticta isolates from citrus in Europe and of a comprehensive DNA sequence analysis of Phyllosticta isolates from all continents; however, as discussed in Section 3.1.2 above, it does not represent a disease survey. This study is the first report of P. citricarpa in Europe and the first description of the new species P. paracitricarpa from eight isolates from Greece and two others from China, previously identified as P. citricarpa. The discussion by Guarnaccia et al. (2017) demonstrates the importance of this study and the need for further research, particularly on the relevance of P. paracitricarpa as a citrus pathogen.
The P. citricarpa isolates from Portugal and Malta/Italy represent two distinct putative clones based on MLGs and mating types. They differed also in their degree of ‘connectedness’ with isolates from other countries. Populations from Portugal and Australia were closely connected but those from Malta/Italy appeared distinct from other populations. The authors claim that this study establishes the presence of P. paracitricarpa only in Asia and Europe; however, the Panel notes the lack of any systematic investigation in citrus‐producing areas worldwide on this newly described species.
No statistically based sampling procedure was employed by Guarnaccia et al. (2017), so the representativeness of the samples and the resulting population structure of P. citricarpa and P. paracitricarpa in Europe are not conclusive. Indeed, Guarnaccia et al. (2017) refer to ‘putative’ populations due to the low number of isolates obtained and the sampling strategy employed. Moreover, as indicated in Section 3.1.1, no information was provided about the number of isolation points in each sample and the corresponding association of the isolates obtained with them. For instance, isolates obtained from the same pycnidium in a dead leaf are likely to belong to the same mating type and MLG.
The species P. citricarpa and P. paracitricarpa were detected only in leaf litter samples, whereas they were apparently not detected on living leaves at the same location. It is known that CBS leaf symptoms are seldom seen on living leaves except on lemon leaves, a very susceptible host species. Nevertheless, isolations from leaf litter are usually less successful with respect to isolation from living plant material due to the presence of soil microorganisms and contaminants (Frankland et al., 1990; Waller et al., 1998). Moreover, Truter et al. (2007) indicated that pycnidiospores of P. citricarpa cannot infect and colonise detached leaves or leaf litter. Consequently, it can be assumed that, in order for the leaf litter to be colonised by P. citricarpa, the leaves in the canopy should have been first infected by the pathogen before falling down to form the leaf litter. This raises the questions as to how P. citricarpa persisted over time in these locations and how it was able to colonise other substrates as a saprophyte, a life style that has not previously been described for this species. Nevertheless, the use of detection methods based on polymerase chain reaction (PCR) (Meyer et al., 2012; Hu et al., 2014) applied to a broader sample of living leaves would have improved the sensitivity of the analyses to clarify this issue. Phyllosticta species are often associated with endophytic infections (Johnston, 1998) in other plant species but it is not clear whether the presence of a fungus in a citrus plant is truly endophytic or symptomless infection of a pathogen, which may still reduce yield and sporulate, without causing visible symptoms (as in the case of Rhynchosporium commune 2 on barley reported by Looseley et al., 2012). This raises a question over the final statement made in the abstract, that Phyllosticta species did not cause disease.
The authors indicated that the putative ‘populations’ of P. citricarpa from Italy and Malta represented a single clone of mating type MAT1‐2‐1, and that from Portugal another of mating type MAT1‐1‐1. The presence of only one mating type in each country suggests that P. citricarpa would reproduce only asexually through pycnidiospores in these sites. These asexual spores are formed in fruit and twig lesions as well as on leaf litter (Kotzé, 1981; Kotzé, 2000; Silva et al., 2016), although Guarnaccia et al. (2017) only detected P. citricarpa in the leaf litter.
Pycnidiospores of P. citricarpa are dispersed by water splash and they have typically been considered as a short‐range dispersal mechanism of minor epidemiological relevance when compared with the airborne ascospores (Kotzé, 1981; Kotzé, 2000). However, more recent studies under laboratory conditions demonstrated that P. citricarpa pycnidiospores could reach longer distances than previously thought (Perryman and West, 2014; Perryman et al., 2014). In addition, infected leaf litter leaves could be dispersed by wind over relatively long distances. Furthermore, pycnidiospores are believed to play a major epidemiological role in Florida, where the population of P. citricarpa is clonal and only one mating type (MAT1‐2) is present (Wang et al., 2016; Hendricks et al., 2017). Studies conducted in Brazil also suggested that pycnidiospores are of major importance in CBS epidemics (Spósito et al., 2007; Spósito et al., 2008), even in the presence of complementary mating types allowing for sexual reproduction through ascospores (Amorim et al., 2016).
Some evidence worldwide also illustrates the importance of pycnidiospores during the early stages of invasion. At the beginning of the epidemics in Zimbabwe, it was shown that most infections originated from pycnidiospores, while ascospores were only found in very small numbers (Whiteside, 1967). Later in 1978, when epidemics were well established, Kotzé (1981) found that ascospores were abundant in the same area. In Argentina, Garrán (1996) indicated that attempts to detect the sexual stage by weekly sampling of dead leaves in plots affected by CBS were unsuccessful. In Florida, CBS was first observed in 2010 (Schubert et al., 2012). As indicated above, disease establishment and spread in this area were attributed to pycnidiospores (Wang et al., 2016; Hendricks et al., 2017).
Citrus regions in Florida and Brazil are characterised by having high rainfall amounts during the growing season, creating conditions very favourable for splash dispersal and subsequent infections by pycnidiospores. On the contrary, citrus regions in the Mediterranean Basin are characterised by semi‐arid climates with much lower rainfall amounts than Florida or Brazil. The ways in which P. citricarpa has been able to establish and persist with asexual water‐splashed pycnidiospores under the particular environmental conditions of the Mediterranean Basin should be further studied. In any case, as Guarnaccia et al. (2017) pointed out, a broader sampling and a larger number of isolates would be needed to determine whether other complementary mating types are present in these areas.
Guarnaccia et al. (2017) suggested that P. citricarpa has probably been introduced in the Mediterranean basin with ancient importations of citrus plants, as early as the 5th century BC. However, the putative clonal populations of P. citricarpa found over all three area sites by the authors do not support the conclusion made in the paper about the hypothesis of ancient introductions. Many studies have documented clonal population structures throughout the entire introduced range. Shifts to clonality and loss of sexual reproduction are frequent in the life‐history traits of introduced fungi (Gladieux et al., 2015). Bottlenecks and founder effects may hamper the coexistence of complementary mating types so that the pathogen can only reproduce asexually. However, the persistence of isolated clonal populations in the long term would be possible only in the absence of subsequent introductions of the pathogen, potentially carrying other complementary mating types leading to sexual reproduction. Assuming their hypothesis of introduction with ancient citrus plant imports, it is unlikely that no further introductions of P. citricarpa occurred in such a long period of time. Although clonal populations of pathogens such as Phytophthora infestans (Mont.) de Bary may persist over centuries, this was associated with a large pathogen population, leading to devastating outbreaks of late blight disease. Clonal populations of P. citricarpa, in the claimed absence of CBS disease, could be indicative of more recent introductions.
Infected fruit was not considered a pathway for P. citricarpa by Guarnaccia et al. (2017), although the potential of this pathway as a means of introduction was thoroughly documented in the pest risk assessment by EFSA (EFSA PLH Panel, 2014). Relatively large amounts of citrus fruit are imported every year from CBS‐affected areas into the EU. These imports are subjected to specific phytosanitary measures to provide a level of confidence that fruits are free from CBS symptoms, but P. citricarpa interceptions are reported by border inspections in the EU (EFSA PLH Panel, 2014). Guarnaccia et al. (2017) indicated that compost heap waste disposal and fruit processing plants were not observed near the sites where P. citricarpa was detected. However, compost heaps and fruit processing plants could have been present in the area before 2015, when the samplings were initiated. Even within the 2015–2017 sampling period, the frequency and duration of the visits to the locations investigated might not have been sufficient to ensure that infected fruits or peel were not deposited in close proximity to the plots. Interestingly, P. citricarpa was detected only in backyard trees, which are more likely to come in close contact with household fruit refuse than commercial orchards.
The Panel considers that citrus fruit will be equally or more likely to introduce a single mating type clonal genotype of P. citricarpa, compared to an infected plant for planting. This is because P. citricarpa only reproduces on fruit through asexual pycnidiospores (Kotzé, 2000). Moreover, fruit is short‐lived compared to an infected plant and the chance of an infection establishing by splash dispersal from a single lesion of a transient fruit would be lower (EFSA PLH Panel, 2014), leading to a greater likelihood that only one genotype would successfully establish a founding population. Tran et al. (2017) suggest only one mating type being present in a single disease lesion in aboveground parts. A plant‐for‐planting pathway would be in general a more likely route of introduction for both mating types from a citrus production area with both mating types, as the plant will be a persistent source of inoculum and sexual ascospores can be produced on the citrus leaves in the leaf litter.
As indicated in Section 3.1, the methods described for disease survey by Guarnaccia et al. (2017) raised some concerns. No information about the citrus cultivars present in the investigated locations was provided. No information was provided on the number of trees assessed for CBS symptoms or how they were selected for assessment (e.g. systematically or at random). The dates when the locations were visited were not reported, except for the second field visit that was conducted during December 2016 and January 2017 at sites where P. citricarpa or P. paracitricarpa were found during the initial samplings. Leaf symptoms of CBS are rare and infected leaves remain generally asymptomatic (Kotzé, 2000). In fruit, CBS is characterised by a relatively long incubation period, and fruit symptoms become visible several months after infection. Lesion formation is driven by phenology and environmental factors and CBS symptoms are visible when fruit mature and reach the ripening stage (Timmer, 1999; Spósito et al., 2004; Sousa and de Goes, 2010). As shown in the harvesting calendar in Figure 51 by EFSA PLH Panel (2014), citrus fruit in the Mediterranean Basin may ripen from September to July depending on the cultivar. Therefore, fruit symptoms may be easily overlooked if surveys were not conducted to coincide with the specific ripening season for each cultivar.
The CBS disease is characterised by a long lag phase and so the initial symptoms of the disease may remain undetected during the early stages of introduction. As Kotzé (1981) indicated, the causal fungus may be present for many years in a particular area before symptoms are observed. Inoculum build‐up from the time of introduction to the onset of an epidemic may take decades. It may take 5–30 years from the time the first symptoms are noticed until the disease reaches epidemic proportions, depending on climate conditions and host susceptibility (Kotzé, 1981). Indeed, CBS symptoms in South Africa were observed for over 30 years before control measures became necessary (Kotzé, 1981). Likewise, experiences from Zimbabwe and Argentina are also indicative of the long lag phase of CBS. In Zimbabwe, CBS was first discovered in 1961 and in 1967 was still considered rare and very localised (Whiteside, 1967). It was not until 1978 that an epidemic was considered to be properly established in this country (Kotzé, 1981). In the north‐east of Entre Rios, Argentina, CBS was first reported in 1986, but in 1996, it was not yet prevalent in the area with only a few foci showing slight to moderate disease severity (Garrán, 1996). The report of P. citricarpa in some European citrus locations by Guarnaccia et al. (2017) and the presumed absence of CBS symptoms could be explained by the known long lag phase of the CBS disease and might be indicative of recent introduction events.
The penultimate sentence of the paper emphasises that the climate modelling studies by Paul et al. (2005), Yonow et al. (2013) and Magarey et al. (2015) were parameterised according to the presence, absence, distribution and severity of CBS, and not the potential presence of the fungus in the absence of disease. With the exception of this paper by Guarnaccia et al. (2017), the known geographical distribution of P. citricarpa worldwide matches with that of CBS. Consequently, P. citricarpa and CBS distributions cannot be disaggregated within the datasets used by these previous climate model studies (Paul et al., 2005; Yonow et al., 2013; Magarey et al., 2015). Paul et al. (2005) indicated that areas of CBS presence were mapped where P. citricarpa has been isolated from symptomatic citrus fruit and/or leaves, or where the disease has been observed by field specialists. Likewise, Yonow et al. (2013) indicated that the distribution of P. citricarpa in South Africa was based on the data reported by Paul et al. (2005) revised in accordance with recent surveys, quoting E. Carstens, pers. comm. Sampling and isolations for P. citricarpa are described by Carstens et al. (2012) and are further reviewed by the EFSA PLH Panel (2016). Based on Carstens et al. (2012), P. citricarpa absence in South Africa was defined in citrus areas without CBS symptoms. For Taiwan and Australia, Yonow et al. (2013) used official sources and reports for P. citricarpa. Disease prevalence data from South Africa and Australia were used by Magarey et al. (2015) but data of P. citricarpa distribution by Carstens et al. (2012) and Yonow et al. (2013) were used for model validation. Moreover, as indicated above, in these two countries the current geographical distributions on P. citricarpa and CBS are coincident.
3.1.4. Other considerations
The Panel notes that notifications of the findings of the quarantine plant pathogen3 Phyllosticta citricarpa by Guarnaccia et al. (2017) were not sent to the NPPOs of the respective countries and that the NPPOs of Italy, Malta and Portugal became aware of these findings only by the publication online of the paper by Guarnaccia et al. (2017) on 29 May 2017.4 Hence, measures, in the sites of the findings by Guarnaccia et al. (2017), could only be taken by NPPOs starting from summer 2017.
Following a request for clarifications, the corresponding author of the paper has informed EFSA that, during the field visits undertaken in December 2016 and January 2017 to confirm the initial findings and to assay the presence of symptoms on fruit, leaves and twigs, as there were no symptoms, no samples were collected.5 Samples are currently being taken by the NPPOs of Italy, Malta and Portugal, at the sites where Guarnaccia et al. (2017) reported the finding of P. citricarpa. Preliminary results of surveys conducted by the NPPOs during the summer 2017 in Malta6 and Portugal,7 using an established DNA‐based diagnostic protocol and repeating the methods by Guarnaccia et al. (2017), did not confirm the findings. Further samplings will be continued in the following seasons.
3.2. Comparisons of findings by Guarnaccia et al. (2017) with previous model predictions
As explained in Sections 1.1 and 1.2, EFSA is requested to report and discuss how the findings of P. citricarpa by Guarnaccia et al. (2017) in Europe fit the predictions of previous model simulations on the establishment of P. citricarpa.
The potential of P. citricarpa to establish in Europe has been studied previously (Paul et al., 2005; EFSA, 2008; Yonow et al., 2013; EFSA PLH Panel, 2014, 2016; Magarey et al., 2015). The EFSA PLH Panel (2014) combined previous modelling work on P. citricarpa ascospore maturation and release (Fourie et al., 2013) and a generic infection model (Magarey et al., 2005), to predict ascospore maturation, release and infection in EU citrus‐growing areas. The EFSA PLH Panel (2014) primarily focused on the sexual reproduction cycle, as this was expected to be the principal way for this pathogen to establish and spread in the risk assessment area. However, this work also included the modelling of pycnidiospore infections taking into account experimental studies on pycnidiospore splash dispersal (Perryman and West, 2014; Perryman et al., 2014).
The geographical coordinates of the findings by Guarnaccia et al. (2017) of P. citricarpa and P. paracitricarpa were superimposed on the results for those sites from previous model simulations for the establishment of P. citricarpa in Europe. The models considered were the applications of the generic infection model for foliar fungal pathogens by Magarey et al. (2005), as extended for ascospore infections simulations by the ascospore maturation and release model of Fourie et al. (2013) (EFSA, 2008; EFSA PLH Panel, 2014, 2016; Magarey et al., 2015). The CLIMEX model by Yonow et al. (2013), further implemented by the EFSA PLH Panel (2014) with interpolated climate datasets of higher spatial and temporal resolution, was not the most appropriate for P. citricarpa but is also considered here as its outcomes were discussed by Guarnaccia et al. (2017).
3.2.1. Review of models previously applied to simulate P. citricarpa establishment
Models to simulate P. citricarpa establishment have been thoroughly reviewed by EFSA (EFSA, 2008; EFSA PLH Panel, 2014, 2016). A summary of results taken from these reviews is presented here.
Yonow et al. (2013) used CLIMEX, a model which predicts an organism's potential distribution based on climatic conditions in its current distribution and any known climatic responses (Table 2). The overall suitability of the location is given by the ecoclimatic index (EI), which is the product of growth and stress indices. The application of CLIMEX for the modelling of P. citricarpa establishment in Europe was evaluated in Section 2.1.3 of EFSA (2008) and in Section 3.3.2.2 of EFSA PLH Panel (2014). The key shortcoming noted in CLIMEX was that the calculation of the EI to provide an indicator of the overall suitability of the location does not take into account the periods of the year when the host is at a susceptible stage and inoculum is potentially available. Moreover, it cannot directly take into account the effect of leaf wetness, a critical microclimatic environmental variable for the successful infection of most fruit and foliage fungal pathogens including P. citricarpa. Nevertheless, the Panel found that the potential for establishment represented by the EI in some EU citrus‐growing areas varied according to both the spatial resolution and the time period covered by the input climate data with the maximum EI ranging from 3 (1961–1990, 0.5° resolution) to 4 (1961–1990 0.1° resolution) and 11 (1998–2007, 25 km resolution). These EI values were similar to those obtained for Addo in the Eastern Cape province of South Africa where P. citricarpa persists but according to Yonow et al. (2013) ‘does not flourish’. Nevertheless, fungicide sprays are applied for CBS control in this area (Schutte, 1995; Korf, 1998; Grout, 2015).
In order to overcome these limitations of CLIMEX, process‐based models were later used to simulate the potential establishment of P. citricarpa. Models by Fourie et al. (2013) were used to simulate the maturation and release of P. citricarpa ascospores (i.e. inoculum availability) and the generic infection model by Magarey et al. (2005) to simulate infections by ascospores or pycnidiospores (EFSA PLH Panel, 2014; Magarey et al., 2015). Sections 3.1.1.2 and 4.2, and Table 1 of EFSA PLH Panel (2016) provided a detailed comparison of the data inputs and modelling approaches adopted by EFSA PLH Panel (2014) and Magarey et al. (2015) for ascospore maturation, release and infection as well as pycnidiospore infection. They found a poor correlation between the models, although both studies simulated infection by ascospores or pycnidiospores with essentially the same equations and parameter values and also utilised the Gompertz equation (Fourie et al., 2013) to describe the dynamics of ascospore production. However, there were a number of differences in the weather and climatic data as well as in model assumptions (Table 2). Actually, the models were shown to be highly sensitive to these assumptions and more precise definitions of the model components are needed to allow direct comparisons to be made.
Magarey et al. (2015) published a rule to classify the suitability of sites for ascospore or pycnidiospore infections based on the average predicted infection period score at the lowest ranked site with a moderate observed disease prevalence (Addo, South Africa). Magarey et al. (2015) stated that the infection score represents the total number of predicted ascospore or pycnidiospore infection periods over a 9‐year period. However, because they presented the model outputs as averages with a standard deviation, the logical interpretation is that they were actually providing the average number of infection periods per year.
3.2.2. Applications of the models to the locations sampled by Guarnaccia et al. (2017)
3.2.2.1. CLIMEX model
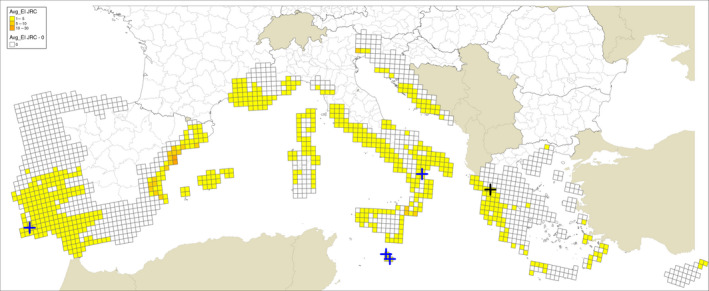
Regarding the CLIMEX growth index and EI for the grid cells where the sites investigated by Guarnaccia et al. (2017) are located, the results presented are from the model by Yonow et al. (2013) run over the 1961–1990 climatic dataset by New et al. (2002) (Appendix A Figures A.1 and A.2) and JRC Mars 1989–2009 interpolated climatic dataset (Appendix A Figures A.3 and A.4) for the cells of the EU citrus‐growing area. Table A.1 in Appendix A shows that none of the 95 locations sampled by Guarnaccia et al. (2017) in southern Portugal, southern and eastern Spain, Majorca, Sicily, southern Italy, Malta, mainland Greece and Crete were in 25 km grid cells with EIs greater than 4 and 26 cells had zero EIs, as in Yonow et al. (2013). Samples were not taken from the Ebro Delta Region of eastern Spain, where the EFSA PLH Panel (2014) found the highest EI values.
Figure A.1.
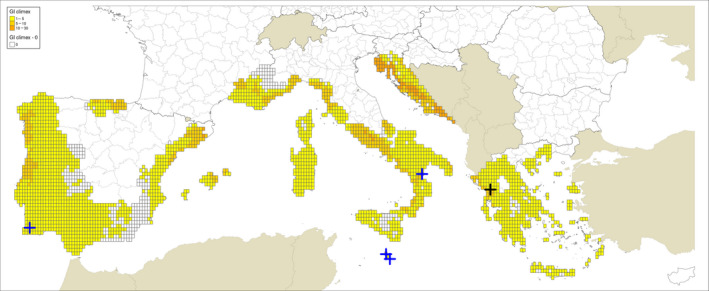
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where P. citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the Growth Indexes (GI) calculated based on CLIMEX model by Yonow et al. (2013), run with 0.1° latitude 0.1° longitude 1961–1990 climatology (New et al., 2002), which is expressing the climatic growth potential of the organism inferred from the temperature and humidity conditions
Figure A.2.
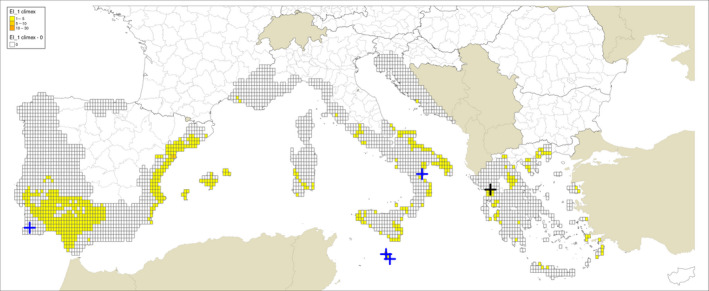
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where P. citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the Ecoclimatic Indexes (EI) calculated based on CLIMEX model by Yonow et al. (2013) run with 0.1° latitude 0.1° longitude 1961–1990 climatology (New et al., 2002). The EI is the growth potential (GI) with the predicted effect of stress factors subtracted
Figure A.3.
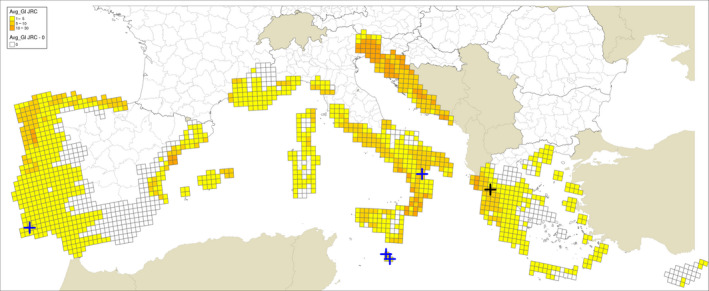
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where P. citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the Growth Indexes (GI) calculated based on CLIMEX model by Yonow et al. (2013), run with 25 × 25 km JRC Mars interpolated climate data, which is expressing the climatic growth potential of the organism inferred from the temperature and humidity conditions
Figure A.4.
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where P. citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the Ecoclimatic Indexes (EI) calculated based on CLIMEX model by Yonow et al. (2013), run with 25 km × 25 km JRC Mars interpolated climate data. The EI is the growth potential (GI) with the predicted effect of stress factors subtracted
Table A.1.
Locations sampled for investigation of presence of Phyllosticta spp. by Guarnaccia et al. (2017) compared with model results according to Yonow et al. (2013), who parameterised a CLIMEX model to predict the potential global distribution of the citrus black spot disease and the risk posed to Europe. The table provides the CLIMEX model outputs for Ecoclimatic index and Growth index for the investigated sites indicated in Table 1 of Guarnaccia et al. (2017). The CLIMEX model was run with climate data at 25 km resolution for the time period 1998–2007
| City | Country | e | n | Type | Grid 0.1° | G. I. 0.1° | E.I. 0.1° | Grid 25 km JRC Mars | G.I 25 Km JRC Mars | EI 25 Km JRC Mars |
|---|---|---|---|---|---|---|---|---|---|---|
| Arta‐1a | Greece | 20.92959 | 39.16172 | P. paracitricarpa | 17753 | 6 | 1 | 52147 | 7 | 2 |
| Gozo‐1b | Malta | 14.25991 | 36.05514 | P. citricarpa. | 12142 | 3 | 1 | 36125 | 5 | 2 |
| Monchique‐1b | Portugal | −8.52623 | 37.3262 | P. citricarpa | 213 | 1 | 0 | 49044 | 2 | 1 |
| Trebisacce‐1b | Italy | 16.56482 | 39.91012 | P. citricarpa | 14311 | 3 | 0 | 54132 | 6 | 1 |
| Zurrieq‐1b | Malta | 14.5051 | 35.82385 | P. citricarpa | NA | NA | NA | 35126 | 4 | 2 |
| Acitrezza‐1 | Italy | 15.16109 | 37.56108 | Other | 12886 | 2 | 0 | 43128 | 1 | 1 |
| Agia‐1 | Greece | 23.92124 | 35.46598 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| Algemesi‐1 | Spain | −0.44977 | 39.20764 | Other | 3928 | 2 | 2 | 52073 | 2 | 2 |
| Algemesi‐2 | Spain | −0.47082 | 39.1969 | Other | 3928 | 2 | 2 | 52073 | 2 | 2 |
| Alginet‐1 | Spain | −0.45803 | 39.26007 | Other | 3928 | 2 | 2 | 52073 | 2 | 2 |
| Alginet‐2 | Spain | −0.41642 | 39.25141 | Other | 3928 | 2 | 2 | 52073 | 2 | 2 |
| Alhaurin_El_Grande‐1 | Spain | −4.67709 | 36.64537 | Other | 1648 | 2 | 0 | 43057 | 2 | 1 |
| Alhaurin_El_Grande‐2 | Spain | −4.69818 | 36.66469 | Other | 1648 | 2 | 0 | 43057 | 2 | 1 |
| Alikianos‐1 | Greece | 23.90863 | 35.45666 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| Alikianos‐2 | Greece | 23.90437 | 35.46238 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| Alikianos‐3 | Greece | 23.9198 | 35.44644 | Other | 20794 | 1 | 0 | 37160 | 1 | 0 |
| Alikianos‐4 | Greece | 23.94556 | 35.46622 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| Almeria‐1 | Spain | −2.40293 | 36.83464 | Other | 2846 | 0 | 0 | 43065 | 0 | 0 |
| Almeria‐2 | Spain | −2.40289 | 36.82883 | Other | NA | NA | NA | 43065 | 0 | 0 |
| Alzira‐1 | Spain | −0.49072 | 39.15696 | Other | 3927 | 2 | 2 | 52073 | 2 | 2 |
| Amfilochia‐1 | Greece | 21.17164 | 38.96138 | Other | 18035 | 3 | 0 | 51148 | 7 | 2 |
| Argo‐1 | Greece | 22.74218 | 37.62865 | Other | 19593 | 2 | 1 | 46154 | 1 | 0 |
| Argo‐2 | Greece | 22.73931 | 37.65556 | Other | 19593 | 2 | 1 | 46154 | 1 | 0 |
| Argos‐1 | Greece | 22.66172 | 37.68659 | Other | 19411 | 1 | 0 | 46154 | 1 | 0 |
| Arta‐2 | Greece | 20.90379 | 39.15566 | Other | 17753 | 6 | 1 | 52147 | 7 | 2 |
| Arta‐3 | Greece | 20.91826 | 39.16047 | Other | 17753 | 6 | 1 | 52147 | 7 | 2 |
| Barcellona P.G.‐1 | Italy | 15.13679 | 38.11056 | Other | 12889 | 2 | 0 | 45127 | 0 | 0 |
| Brucoli‐1 | Italy | 15.11052 | 37.29482 | Other | 12884 | 2 | 1 | 42128 | 1 | 1 |
| Canicattí‐1 | Italy | 13.8409 | 37.35843 | Other | 11855 | 0 | 0 | 42123 | 1 | 0 |
| Carruba‐1 | Italy | 15.19094 | 37.68463 | Other | 13039 | 4 | 1 | 43128 | 1 | 1 |
| Castello‐1 | Spain | −0.0862 | 39.90392 | Other | 4097 | 3 | 3 | 55075 | 1 | 1 |
| Castello‐2 | Spain | −0.08823 | 39.88386 | Other | 4097 | 3 | 3 | 55075 | 1 | 1 |
| Castello‐3 | Spain | −0.09095 | 39.88401 | Other | 4097 | 3 | 3 | 55075 | 1 | 1 |
| Cefalú‐1 | Italy | 14.01227 | 38.02948 | Other | NA | NA | NA | 45124 | 6 | 2 |
| Chania‐1 | Greece | 24.05114 | 35.49315 | Other | 20965 | 1 | 0 | 38161 | 2 | 1 |
| Chania‐2 | Greece | 23.94806 | 35.47789 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| citricarpa‐1 | NA | −8.51451 | 37.33241 | Other | 213 | 1 | 0 | 49044 | 2 | 1 |
| Comiso‐1 | Italy | 14.63716 | 36.94398 | Other | 12430 | 1 | 0 | 40126 | 1 | 0 |
| Conceicao‐1 | Portugal | −7.91693 | 37.04848 | Other | 447 | 1 | 0 | 47046 | 2 | 1 |
| Curiglia‐1 | Italy | 16.20376 | 38.76773 | Other | 13985 | 4 | 0 | 48131 | 12 | 2 |
| El_Ejido‐1 | Spain | −2.71999 | 36.79521 | Other | 2669 | 0 | 0 | 43064 | 0 | 0 |
| Estellencs‐1 | Spain | 2.481876 | 39.6535 | Other | 5090 | 2 | 1 | 53083 | 1 | 1 |
| Faro‐1 | Portugal | −7.91681 | 37.10846 | Other | 447 | 1 | 0 | 48046 | 1 | 1 |
| Faro‐2 | Portugal | −7.91743 | 37.06264 | Other | 447 | 1 | 0 | 47046 | 2 | 1 |
| Giarratana‐1 | Italy | 14.97442 | 36.88344 | Other | 12726 | 1 | 0 | 40127 | 3 | 1 |
| Gouria‐1 | Greece | 21.25765 | 38.45498 | Other | 18032 | 4 | 1 | 49149 | 7 | 2 |
| Gozo‐2 | Malta | 14.2593 | 36.04907 | Other | 12142 | 3 | 1 | 36125 | 5 | 2 |
| Gozo‐3 | Malta | 14.26012 | 36.03753 | Other | 12142 | 3 | 1 | 36125 | 5 | 2 |
| Gozo‐4 | Malta | 14.27936 | 36.04965 | Other | 12142 | 3 | 1 | 36125 | 5 | 2 |
| Gozo‐5 | Malta | 14.28445 | 36.05817 | Other | 12142 | 3 | 1 | 36125 | 5 | 2 |
| Grotte‐1 | Italy | 15.17701 | 37.67993 | Other | 13039 | 4 | 1 | 43128 | 1 | 1 |
| Guardia‐1 | Italy | 15.17592 | 37.66271 | Other | NA | NA | NA | 43128 | 1 | 1 |
| Kirtomados‐1 | Greece | 23.91666 | 35.47875 | Other | 20794 | 1 | 0 | 38160 | 2 | 1 |
| Leni‐1 | Italy | 14.59752 | 38.04442 | Other | 12437 | 2 | 0 | 45126 | 1 | 1 |
| Leni‐2 | Italy | 14.82713 | 38.55289 | Other | NA | NA | NA | 47126 | 1 | 0 |
| Lentini‐1 | Italy | 15.0209 | 37.32058 | Other | 12884 | 2 | 1 | 42127 | 1 | 0 |
| Malaga‐1 | Spain | −4.42706 | 36.76176 | Other | 1801 | 1 | 0 | 44058 | 1 | 0 |
| Mascali‐1 | Italy | 15.1925 | 37.76768 | Other | 13039 | 4 | 1 | 44128 | 1 | 1 |
| Mascali‐2 | Italy | 15.19464 | 37.76826 | Other | 13039 | 4 | 1 | 44128 | 1 | 1 |
| Massafra‐1 | Italy | 17.14411 | 40.54476 | Other | 14761 | 1 | 0 | 57134 | 3 | 1 |
| Mastro‐1 | Greece | 21.28054 | 38.43029 | Other | 18032 | 4 | 1 | 49149 | 7 | 2 |
| Mesquita‐1 | Portugal | −8.28949 | 37.21367 | Other | 328 | 1 | 0 | 48045 | 2 | 1 |
| Mesquita‐2 | Portugal | −8.29781 | 37.20453 | Other | 328 | 1 | 0 | 48045 | 2 | 1 |
| Mineo‐1 | Italy | 14.69086 | 37.35072 | Other | 12580 | 2 | 1 | 42126 | 0 | 0 |
| Moncada‐1 | Spain | −0.39458 | 39.58855 | Other | 3930 | 2 | 2 | 54074 | 3 | 2 |
| Monchique‐2 | Portugal | −8.50369 | 37.33623 | Other | 214 | 1 | 0 | 49044 | 2 | 1 |
| Monchique‐3 | Portugal | −8.49223 | 37.33224 | Other | 270 | 1 | 0 | 49044 | 2 | 1 |
| Motta_S._Anastasia‐1 | Italy | 14.88602 | 37.4821 | Other | 12729 | 2 | 2 | 42127 | 1 | 0 |
| Motta_S._Anastasia‐2 | Italy | 14.95416 | 37.46971 | Other | 12729 | 2 | 2 | 42127 | 1 | 0 |
| Nafplio‐1 | Greece | 22.78527 | 37.58931 | Other | 19593 | 2 | 1 | 46155 | 0 | 0 |
| Nafplio‐2 | Greece | 22.69559 | 37.5751 | Other | 19593 | 2 | 1 | 46154 | 1 | 0 |
| Nafplio‐3 | Greece | 22.6968 | 37.58229 | Other | 19593 | 2 | 1 | 46154 | 1 | 0 |
| Nafplio‐4 | Greece | 22.87484 | 37.5888 | Other | 19771 | 1 | 0 | 46155 | 0 | 0 |
| Nicolosi‐1 | Italy | 15.02948 | 37.61127 | Other | 12886 | 2 | 0 | 43127 | 0 | 0 |
| Niscemi‐1 | Italy | 14.3934 | 37.13978 | Other | 12287 | 1 | 0 | 41125 | 1 | 0 |
| Noto‐1 | Italy | 15.09545 | 36.8465 | Other | 12882 | 2 | 1 | 40128 | 7 | 3 |
| Pachino‐1 | Italy | 15.08699 | 36.72003 | Other | 12881 | 3 | 1 | 39128 | 9 | 4 |
| Pachino‐2 | Italy | 15.08941 | 36.72233 | Other | 12881 | 3 | 1 | 39128 | 9 | 4 |
| Pedara‐1 | Italy | 15.06654 | 37.60871 | Other | 12886 | 2 | 0 | 43127 | 0 | 0 |
| Pizzo Calabro‐1 | Italy | 16.22601 | 38.76039 | Other | 13985 | 4 | 0 | 48131 | 12 | 2 |
| Ribera‐1 | Italy | 13.24185 | 37.49711 | Other | 11271 | 2 | 1 | 42121 | 5 | 2 |
| Ribera‐2 | Italy | 13.25207 | 37.50442 | Other | 11272 | 1 | 0 | 42121 | 5 | 2 |
| Riposto‐1 | Italy | 15.19935 | 37.69647 | Other | 13039 | 4 | 1 | 43128 | 1 | 1 |
| Rocca_Imperiale‐1 | Italy | 16.61795 | 40.10839 | Other | 14312 | 3 | 0 | 54132 | 6 | 1 |
| San_Gregorio‐1 | Italy | 15.10097 | 37.5623 | Other | 12886 | 2 | 0 | 43127 | 0 | 0 |
| Scordia‐1 | Italy | 14.86915 | 37.28153 | Other | 12728 | 2 | 1 | 42127 | 1 | 0 |
| Seville‐1 | Spain | −5.96282 | 37.50854 | Other | 1207 | 3 | 1 | 48053 | 4 | 3 |
| Seville‐2 | Spain | −5.95491 | 37.48298 | Other | 1206 | 4 | 2 | 48053 | 4 | 3 |
| Sikoula‐1 | Greece | 21.0834 | 39.08593 | Other | 17891 | 5 | 1 | 52148 | 6 | 1 |
| Silves‐1 | Portugal | −8.39084 | 37.16415 | Other | 269 | 1 | 0 | 48044 | 2 | 1 |
| Soller‐1 | Spain | 2.709609 | 39.76453 | Other | 5211 | 3 | 0 | 53084 | 1 | 1 |
| Soller‐2 | Spain | 2.7266 | 39.77012 | Other | 5211 | 3 | 0 | 53084 | 1 | 1 |
| Terme_Vigliatore‐1 | Italy | 15.16324 | 38.1458 | Other | 12889 | 2 | 0 | 45128 | 0 | 0 |
| Torremolinos‐1 | Spain | −4.50413 | 36.67272 | Other | 1721 | 2 | 0 | 43057 | 2 | 1 |
| Trebisacce‐2 | Italy | 16.56063 | 39.90671 | Other | 14311 | 3 | 0 | 54132 | 6 | 1 |
Guarnaccia et al. (2017) found P. citricarpa in Zurrieq, Gozo, Trebisacce and Monchique, and P. paracitricarpa in Arta (Figure 1). The grid cells with the five locations where Guarnaccia et al. (2017) found P. citricarpa either had EIs of 1 or 2 (Table 3 below and Figures A.1, A.2, A.3 and A.4 in Appendix A). However, the EFSA PLH Panel (2014) noted that the CLIMEX growth index, the combination of the temperature index and moisture index, may be a more appropriate indicator of climate suitability for P. citricarpa since these show the extent to which suitable temperatures and moistures coincide in the spring and autumn and this ranged from 2 to 7 in the 25 km grid cells where Guarnaccia et al. (2017) found P. citricarpa (Table 3). Although the EFSA PLH Panel (2014) has already shown that CLIMEX has limited applicability in determining the area of potential establishment for P. citricarpa, it can be concluded that nearly 75% of the samples taken by Guarnaccia et al. (2017), including the five positive samples, were in 25 km grid cells shown to be marginally suitable following the methods used by Yonow et al. (2013).
3.2.2.2. Models for ascospore maturation, release and infection (EFSA PLH Panel, 2014; Magarey et al., 2015)
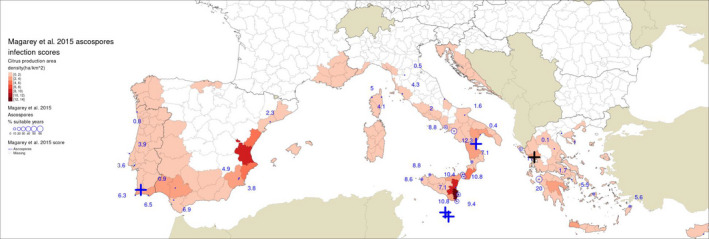
The simulations by Magarey et al. (2015) for ascospore maturation and infection did not include the sites investigated by Guarnaccia et al. (2017); therefore, the values reported in Table 4 for ascospore infection scores are from the nearest location in the site dataset of Magarey et al. (2015): i.e. for Zurrieq and Gozo, the values are from Luqa in Malta; for Trebisacce the values are from Marina di Ginosa; for Monchique, the values are from Sagres; for Arta, the values are from Kerkyra (Figure 3). The lowest score of 0.4 was recorded for Trebisacce in Italy and the highest of 11 at Arta in Greece. The threshold for a site predicted to be suitable for ascosporic infection by Magarey et al. (2015) was 13, based on the score reported for Addo, South Africa. Nevertheless, different conclusions would have been reached if other sites (EFSA PLH Panel, 2016) or periods (see paragraph below) had been chosen as the benchmark (EFSA PLH Panel, 2016). Magarey et al. (2015) considered Addo as of moderate disease prevalence, however fungicide sprays are applied for CBS control there (Korf, 1998; Grout, 2015).
Table 4.
Model outputs from Magarey et al. (2015) for locations in Europe near to sites where Guarnaccia et al. (2017) found Phyllosticta citricarpa or P. paracitricarpa
| Locations where P. citricarpa or P. paracitricarpa were found by Guarnaccia et al. (2017) | Near location from Table 1 of Magarey et al. (2015) | From Table 2 of Magarey et al. (2015) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ascospores | Pycnidiospores | |||||||||
| Infection on days with PAT | Infection | |||||||||
| Country | Location | Latitude | Longitude | Location, Country | Mean | Std. Dev. | % suitable years | Mean | Std. Dev. | % suitable years |
| Malta | Zurrieq | 35.823845 | 14.505099 | Luqa, Malta | 4.8 | 2.2 | 0 | 9.4 | 5.3 | 0 |
| Malta | Gozo | 36.055138 | 14.259907 | Luqa, Malta | 4.8 | 2.2 | 0 | 9.4 | 5.3 | 0 |
| Italy | Trebisacce | 39.910122 | 16.564824 | Marina di Ginosa, Italy | 0.4 | 0.9 | 0 | 15.9 | 9.3 | 0 |
| Portugal | Monchique | 37.326195 | −8.526232 | Sagres, Portugal | 6.3 | 3.2 | 0 | 15.7 | 9.2 | 0 |
| Greece | Arta | 39.160465 | 20.918257 | Kerkyra, Greece | 11 | 5.6 | 22 | 21.6 | 7.9 | 0 |
PAT: proportion of ascospores trapped.
Figure 3.
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the ascospores infection scores for P. citricarpa as calculated by Magarey et al. (2015) over EU NUTS3 citrus production area density (as extracted from national statistical databases of Portugal, Spain, France, Italy, Malta, Croatia, Greece and Cyprus for year 2007; for details, see Appendix F of EFSA PLH Panel, 2014)
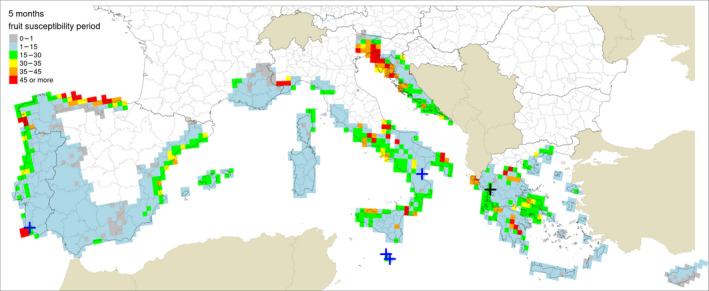
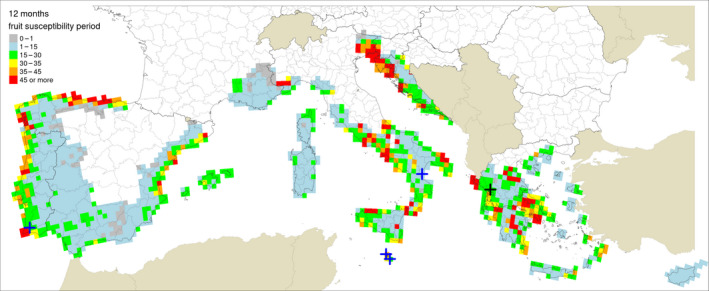
Table 5 shows the outputs from the application of the model by Fourie et al. (2013) for ascospore maturation and release and the generic infection model (Magarey et al., 2005) for P. citricarpa ascospores (EFSA PLH Panel, 2014) for the locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa. The outputs from the locations where neither species was found are given in Appendix B. The results are shown as the number of infections accumulated from May to September considering a fruit susceptibility of 5 months (as per Magarey et al., 2015); accumulated from May to November taking into account studies indicating a period of fruit susceptibility of at least 6–7 months (Reis et al., 2003; Baldassari et al., 2006; Brentu et al., 2012; Lanza et al., 2017); and accumulated for the whole year considering that susceptible leaves are always present in the citrus canopy (Truter et al., 2004a,b; Truter, 2010). Values for the 25 × 25 km grid cells of the JRC Mars 1989 interpolated climatic dataset for the cells of the EU citrus‐growing area are shown in Figures 4–6, including the sites where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa.
Table 5.
Model outputs for Phyllosticta citricarpa ascospore infections from EFSA PLH Panel (2014) (D 50 = 3; T min = 15) for locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa, considering different periods of host susceptibility: five months (May–September) of fruit susceptibility; seven months (May–November) of fruit susceptibility; 12 months (January–December) of presence of susceptible leaves
| Locations and coordinates where P. citricarpa (Zurrieq, Gozo, Trebisacce and Monchique) or P. paracitricarpa (Arta) were found by Guarnaccia et al. (2017) | Ascospores (D 50 = 3 h; T min = 15°C) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of ascospore infections in the period May–September (5 months fruit susceptibility) in days with PAT | Number of ascospore infections in the period May–November (7 months fruit susceptibility) in days with PAT | Number of ascospores infections in the period January–December (12 months leaf susceptibility) in days with PAT | ||||||||||
| Country | Location | Latitude | Longitude | Mean | Std. Dev. | % suitable years | Mean | Std. Dev. | % suitable years | Mean | Std. Dev. | % suitable years |
| Malta | Zurrieq | 35.823845 | 14.505099 | 9 | 11 | 76 | 22 | 25 | 95 | 22 | 25 | 95 |
| Malta | Gozo | 36.055138 | 14.259907 | 10 | 9 | 90 | 29 | 33 | 100 | 29 | 33 | 100 |
| Italy | Trebisacce | 39.910122 | 16.564824 | 9 | 15 | 67 | 10 | 16 | 81 | 10 | 16 | 81 |
| Portugal | Monchique | 37.326195 | −8.526232 | 15 | 32 | 62 | 30 | 49 | 81 | 30 | 49 | 81 |
| Greece | Arta | 39.160465 | 20.918257 | 12 | 20 | 71 | 18 | 26 | 76 | 18 | 26 | 76 |
PAT: proportion of ascospores trapped.
Figure 4.
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the ascospores infection as calculated by EFSA PLH Panel (2014) for D 50 = 3 h and T min = 15°C (Fourie 0.7), considering a 5‐month fruit susceptibility period (from May to September)
Figure 6.
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the ascospores infection by EFSA PLH Panel (2014) for D 50 = 3 h and T min =15°C (Fourie 0.7), considering a 12‐month leaves susceptibility period
The mean number of ascospore infections was higher during the period from May to November compared with those from May to September. The highest mean number of ascospore infections from May to November was 15 at Monchique, Portugal, and the lowest was 9 at Zurrieq, Malta, and Trebisacce, Italy. From May to November, the mean number of ascospore infections ranged from 10 at Trebisacce, Italy, to 30 at Monchique, Portugal. Numbers of ascospore infections from January to December and from May to November did not differ. The percentage of suitable years for ascospore infections ranged from 62% at Monchique, Portugal, for the period May–September to 100% at Gozo, Malta, for the periods May–November or January–December.
In conclusion, simulations by EFSA PLH Panel (2014) and Magarey et al. (2015) suggested that climatic conditions at the locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa may sustain to some extent infections by P. citricarpa ascospores.
3.2.2.3. Models for pycnidiospore infection (EFSA, 2008; EFSA PLH Panel, 2014; Magarey et al., 2015)
As for ascospores, the simulations for pycnidiosporic infection by Magarey et al. (2015) did not include the sites investigated by Guarnaccia et al. (2017); therefore, the values reported in Table 3 are for the nearest locations in each case (Figure 7). The scores ranged from 9.4 at Luqa (nearest location to Zurrieq and Gozo) in Malta to 21.6 at Kerkyra (nearest location to Arta) in Greece. The threshold set by Magarey et al. (2015) for pycnidiosporic infection was 48, based on the average score for Addo, South Africa. The score values obtained for the locations where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa were somewhat lower that the threshold set by Magarey et al. (2015). However, Guarnaccia et al. (2017) only found one mating type of P. citricarpa in each location; thus, the pathogen is likely to thrive through asexual reproduction by pycnidiosporic infection. Nevertheless, as indicated above, different conclusions may be reached by choosing other thresholds.
Figure 7.
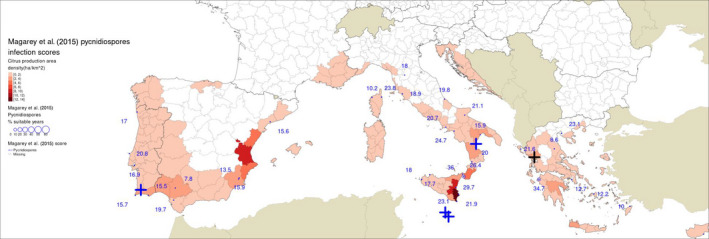
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece; green dots indicate sites of sampling without positive finding. The map provides the pycnidiospores infection scores for P. citricarpa as calculated by Magarey et al. (2015) over EU NUTS3 citrus production area density (as extracted from national statistical databases of Portugal, Spain, France, Italy, Malta, Croatia, Greece and Cyprus for year 2007; for details, see Appendix F of EFSA PLH Panel, 2014)
Table 6 shows the model outputs from the application of the generic infection model (Magarey et al., 2005) for pycnidiospores (EFSA PLH Panel, 2014) for the locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa. The outputs from the locations where neither species was found are given in Appendix B. As for ascospores, the results are shown as the number of infections accumulated from May to September considering a fruit susceptibility of 5 months (as per Magarey et al., 2015); accumulated from May to November taking into account studies indicating a period of fruit susceptibility of at least 6–7 months (Reis et al., 2003; Baldassari et al., 2006; Brentu et al., 2012; Lanza et al., 2017); and accumulated for the whole year considering that susceptible leaves are always present in the citrus canopy (Truter et al., 2004a,b; Truter, 2010). This is particularly important in the case of P. citricarpa pycnidiospores, because significant numbers of infection events were predicted by the model in October and November and from December to May (see Figures 29, 32–35 in EFSA (2008) and Figure 19 in EFSA PLH Panel (2014)).
Table 6.
Model outputs for Phyllosticta citricarpa pycnidiospore infections from EFSA PLH Panel (2014) (D 50 = 3; T min = 10) for the locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa, considering different periods of host susceptibility: 5 months (May–September) of fruit susceptibility; 7 months (May–November) of fruit susceptibility; 12 months (January–December) of presence of susceptible leaves
| Locations and coordinates where P. citricarpa (Zurrieq, Gozo, Trebisacce and Monchique) or P. paracitricarpa (Arta) were found by Guarnaccia et al. (2017) | Pycnidiospores (D 50 = 3 h; T min = 10°C) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of infections in the period May–September (5 months fruit susceptibility, as per Magarey et al., 2015) | Number of infections in the period May–November (7 months fruit susceptibility, as per EFSA PLH Panel 2014, 2016) | Number of infections in the period January–December (12 months leaf susceptibility, as per EFSA PLH Panel 2014, 2016) | ||||||||||
| Country | Location | Latitude | Longitude | Mean | Std. Dev. | % suitable years | Mean | Std. Dev. | % suitable years | Mean | Std. Dev. | % suitable years |
| Malta | Zurrieq | 35.823845 | 14.505099 | 25 | 17 | 95 | 92 | 59 | 100 | 152 | 75 | 100 |
| Malta | Gozo | 36.055138 | 14.259907 | 26 | 18 | 95 | 91 | 67 | 100 | 145 | 76 | 100 |
| Italy | Trebisacce | 39.910122 | 16.564824 | 34 | 29 | 95 | 56 | 40 | 100 | 61 | 43 | 100 |
| Portugal | Monchique | 37.326195 | −8.526232 | 55 | 51 | 95 | 150 | 93 | 100 | 241 | 128 | 100 |
| Greece | Arta | 39.160465 | 20.918257 | 36 | 33 | 95 | 96 | 54 | 100 | 112 | 63 | 100 |
When considering the 5‐month period from May to September, the mean number of pycnidiospore infections ranged from 25 at Zurrieq, Malta, to 55 at Monchique, Portugal (Table 6, Figure 8). Mean values increased for the 7‐month period of May–November, with the lowest value of 91 at Gozo, Malta, and the highest of 150 at Monchique, Portugal (Table 6, Figure 9). The highest values were obtained for the 12‐month period, ranging from 61 at Trebisacce, Italy, to 241 at Monchique, Portugal (Table 6, Figure 10). The percentage of suitable years for pycnidiospore infections was 95% for the period May–September and 100% for the periods May–November or January–December.
Figure 8.
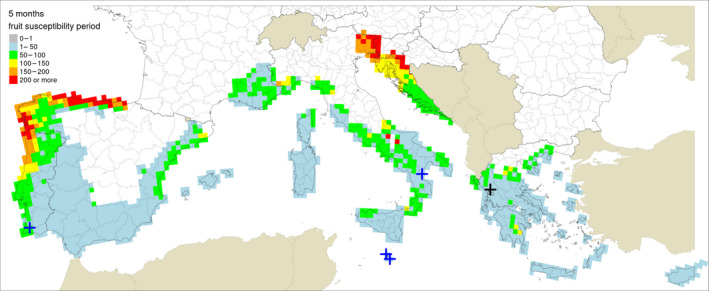
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the pycnidiospores infection by EFSA PLH Panel (2014) for D 50 = 3 h and T min = 10°C, considering a 5‐month fruit susceptibility period (from May to September)
Figure 9.
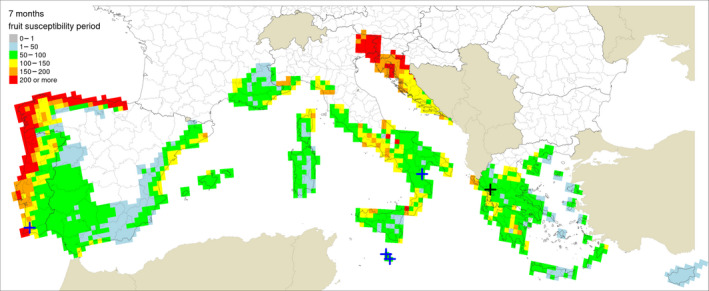
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the pycnidiospores infection by EFSA PLH Panel (2014) for D 50 = 3 h and T min = 10°C, considering a 7‐month fruit susceptibility period (May to November)
Figure 10.
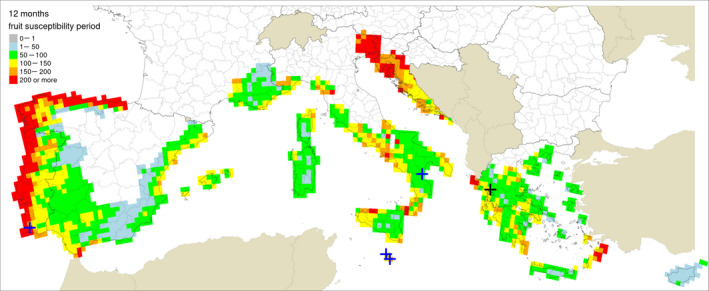
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the pycnidiospores infection by EFSA PLH Panel (2014) for D 50 = 3 h and T min = 10°C, considering a 12‐month leaves susceptibility period
Taking into account the 5‐month period of May–September, relatively lower number of pycnidiospore infection events was reported by Magarey et al. (2015) (Table 4) when compared with those by EFSA PLH Panel (2014) in Table 6. Both studies used the same generic infection model (Magarey et al., 2005), the same parameter values: T min = 10°C, T opt = 25°C, T max = 35°C and D 50 = 3 hours, and incorporated a rain threshold to account for the splash requirement in pynidiospore dispersal. However, in Magarey et al. (2015), this threshold was set to a default value of 0.2 mm per hour, whereas the presence of at least one rain event per day was considered by EFSA PLH Panel (2014). Moreover, different underlying climatic datasets were used and locations drawn from EFSA PLH Panel (2014) were just proxies of those in Magarey et al. (2015).
EFSA (2008) simulated infections by P. citricarpa pycnidiospores for 12‐month periods using the same generic infection model, parameter values, and rain threshold for the splash dispersal. The number of potential infection events obtained by EFSA (2008) for locations in Portugal, Greece, Spain and Italy (Figure 25 of EFSA, 2008) were in line with those by EFSA PLH Panel (2014) from January to December for the locations in Europe where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa (Table 6). Likewise EFSA (2008) concluded, the number of infection events by EFSA PLH Panel (2014) for these locations in Europe (Table 6) was comparable to those from sites of current CBS occurrence (Figure 25 of EFSA, 2008). These results indicated that climatic conditions at these sites are favourable for P. citricarpa to complete its asexual cycle through infections by pycnidiospores.
Figure 5.
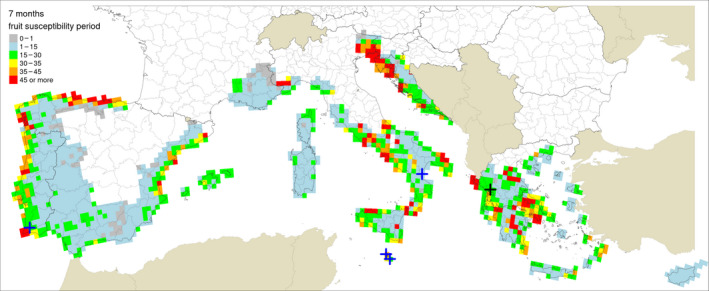
Locations indicated by Guarnaccia et al. (2017) for their samplings: blue crosses indicate sites where Phyllosticta citricarpa was reported in the leaf litter; a black cross indicates the site where P. paracitricarpa was reported in leaf litter in Greece. The map provides the ascospores infection by EFSA PLH Panel (2014) for D 50 = 3 h and T min =15°C (Fourie 0.7), considering a 7‐month fruit susceptibility period (from May to November)
3.3. Uncertainties
3.3.1. Uncertainties identified in the evaluation of the paper by Guarnaccia et al. (2017)
The main uncertainties identified in the evaluation of the paper by Guarnaccia et al. (2017) are:
the apparent lack of a CBS disease survey by the authors;
the fact that the NPPOs were not alerted at the time of the findings and as a consequence a confirmation of the findings by Guarnaccia et al. (2017) by the NPPO is still pending, although preliminary results are not positive;
how to reconcile the novelty of the findings by Guarnaccia et al. (2017) (all findings are in leaf litter and backyards only, all reporting only one mating type) with the known life cycle of P. citricarpa;
the representativeness of the isolates to the populations from which they were sampled;
the origin of P. citricarpa in Europe;
the ecology and pathogenicity of P. citricarpa in the field, if confirmed in the Mediterranean area;
P. paracitricarpa distribution worldwide and its pathogenicity under field conditions.
3.3.2. Uncertainties on the fitting of the previous model simulations to the findings by Guarnaccia et al. (2017)
There are general uncertainties on the comparison of previous model simulations to the findings of P. citricarpa in Europe by Guarnaccia et al. (2017):
These findings present some atypical aspects which were not considered in the previous models for P. citricarpa: in all samplings, P. citricarpa was only found in the leaf litter but never on living plant tissues; in each location, all isolates belong to a single mating type, hence, if representative of the fungal population in each site, they should not be able to produce ascospores but only pycnidiospores.
In addition, P. citricarpa was found by Guarnaccia et al. (2017) only in backyard landscapes (two backyards near a swimming pool, one in a vegetable garden on the road side and one on a terraced slope) where water splashes and runoff might happen more likely than in commercial orchards; however, all previous models did not account for the effect of irrigation.
All model predictions (EFSA, 2008; Yonow et al., 2013; EFSA PLH Panel 2014, 2016; Magarey et al., 2015) were parameterised and conducted for the fungus P. citricarpa. For this reason, the extrapolation of the results of these simulations to the newly described species P. paracitricarpa is highly uncertain, because nothing is known about the biology and ecology of P. paracitricarpa.
There are uncertainties on the model parameters for the application to P. citricarpa of the generic infection model for foliar fungal pathogens by Magarey et al. (2005). Such uncertainties are due to the lack of epidemiological studies on P. citricarpa ascospore and pycnidiospore infection and are discussed in details in previous EFSA opinions (EFSA, 2008; EFSA PLH Panel, 2014).
4. Conclusions
The study reported in Guarnaccia et al. (2017) has identified four species of Phyllosticta present on citrus in Europe. Two, P. citricarpa and P. capitalensis, are well‐defined species. P. citricarpa is recorded for the first time in Europe, but only in citrus leaf litter, and confirms the predictions made by Magarey et al. (2015) and by EFSA (2008) and EFSA PLH Panel (2014, 2016) that P. citricarpa can establish in some citrus‐growing regions of Europe. Two new species P. paracitricarpa and P. paracapitalensis are described using a combination of morphological and molecular phylogeny techniques. P. paracitricarpa is shown to have similar pathogenicity to P. citricarpa on fruits of C. sinensis.
Genotyping of the P. citricarpa isolates, assuming they are representative of the population sampled, suggests at least two independent introductions, with the population in Portugal being different from that present in Malta and Italy, based on MLG and mating type idiomorph. The sampling protocols as described in the paper are not explained clearly. Although most of the major citrus‐growing areas in Europe were covered, access to the locations seemed to be a key feature determining the choice of locations. All findings of P. citricarpa were in backyards and in a high percentage of these locations there were positive identifications (positive identifications were overall in 18% of the sampled backyards, ranging from 9% of the Italian backyards sampled to 25% in Portugal and 50% in Malta).
The authors only report findings of P. citricarpa and P. paracitricarpa from leaf litter, but no positive detection from living plants (fruit, leaves or twigs) was obtained in the same plants. However, since leaf litter only originates from living leaves and since P. citricarpa has not been shown to infect or colonise dead leaves (Truter et al., 2007), the P. citricarpa must have infected the above living leaves.
The origin of the introductions remains obscure. Guarnaccia et al. (2017) considered introduction to be a consequence of P. citricarpa having been present for a considerable time through ancient plants or of illegal movement of planting/propagating material. In the Panel's view, there are problems with the first of these interpretations. If the pathogen had been present for a considerable time, then it can only have persisted if there were repeated asexual cycles involving living leaves and possibly other tissues (particularly twigs) and the resulting leaf litter. Assuming only a single mating type population was present at each location, as suggested by the isolates obtained, then no sexual reproduction involving ascospores would have occurred. Persistence of the pathogen over extended time involving only mycelium and pycnidiospores in an endophytic/saprophytic cycle would be highly problematic, especially as no P. citricarpa isolates were obtained from living leaves from plants.
In the case of illegal movement of planting/propagating material, it would be more likely that the two mating types would be present on the intact plants than would occur with a sporadic occurrence on fruits which, with only asexual reproduction, would involve more probably a single mating type. Tran et al. (2017) suggest only one mating type being present in a single disease lesion in aboveground parts. In the Panel's view, the fruit pathway would be an equally or more likely origin of these introductions.
It was not reported how surveys for CBS disease were carried out by the authors, and whether these were in or close to the locations where samples were taken and positive identifications made. Thus, the claim that there was no CBS disease even where the pathogen was present is not supported by the results presented. The persistence of P. citricarpa in leaf litter also means that the requirement for conducive climatic conditions over a sequence of years would no longer apply as a persistent endophytic/saprophytic cycle could result in CBS disease being manifest in a single year if conditions were favourable.
Results from the simulation models indicated that climatic conditions at the locations where Guarnaccia et al. (2017) found P. citricarpa or P. paracitricarpa were conducive for P. citricarpa establishment. The number of infection events by P. citricarpa pycnidiospores obtained for these locations in Europe was comparable to those from sites of current CBS occurrence outside Europe. This is particularly relevant considering that only one mating type of P. citricarpa was found by Guarnaccia et al. (2017) in each site; thus, only asexual reproduction by pycnidiospores would be expected. Further epidemic development and impacts would be boosted by the concurrence of complementary mating types of P. citricarpa in the same location, leading to sexual reproduction by ascospores, subsequent inoculum build up and airborne dispersal.
The results by Guarnaccia et al. (2017) are based on the sampling conducted during the initial field visits. Following a request for clarifications, the corresponding author of the paper has informed EFSA that, during the second set of field visits undertaken in December 2016 and January 2017 to confirm the initial findings, no symptoms were observed and therefore no confirmatory samples were collected by the authors. Preliminary results of surveys conducted during the summer 2017 in Malta and Portugal by the NPPOs, using an established DNA‐based diagnostic protocol and repeating the methods by Guarnaccia et al. (2017), did not so far confirm the findings by Guarnaccia et al. (2017). Further samplings will be continued in all sites in the following seasons.
Abbreviations
- actA
actin
- CBS
citrus black spot
- CLIMEX
Software to predict the effect of climate on species
- EI
ecoclimatic Index
- gapdh
glyceraldehyde‐3‐phosphate dehydrogenase
- GI
growth index
- GPS
Global Positioning System
- ITS
internal transcribed spacer region
- LSU
large subunit nrDNA
- Mars
Monitoring Agriculture Resources System
- MLG
multilocus genotype
- MCYFS
Mars Crop Yield Forecasting System
- NPPO
National Plant Protection Organisation
- PCR
polymerase chain reaction
- PLH
EFSA Plant Health Panel
- rpb2
RNA polymerase II second largest subunit
- tef1
translation elongation factor 1‐α
Appendix A – CLIMEX results for locations sampled by Guarnaccia et al. (2017)
1.
Appendix B – Ascospores and pycnidiospores infection
1.
Table B.1.
Locations sampled for investigation of presence of Phyllosticta spp. by Guarnaccia et al. (2017) (as indicated in Table 1 of Guarnaccia et al., 2017), compared with model results for ascospore and pycnidiospore infections according to the EFSA PLH Panel (2014)
| City | Country | e | n | Grid_Code | Type of finding by Guarnaccia et al. (2017) | Pycnidiospores (D 50 = 3 hours; T min = 10°C) | Ascospores (D 50 = 3 hours; T min = 15°C; Fourie 0.7) | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean number of infections in the period May–September (5 months fruit susceptibility) | Mean number of infections in the period May–November (7 months fruit susceptibility) | Mean number of yearly infection January–December (12 months leaf susceptibility) | Mean number of infections in the period May–September (5 months fruit susceptibility) in days with PAT | Mean number of infections in the period May–November (7 months fruit susceptibility) in days with PAT | Mean number of yearly infection January–December (12 months leaf susceptibility) in days with PAT | ||||||
| Arta‐1 | Greece | 20.92959 | 39.16172 | 52147 | P. paracitricarpa | 36.09524 | 95.80952 | 111.9852 | 17.90476 | 24.52381 | 24.52381 |
| Gozo‐1 | Malta | 14.25991 | 36.05514 | 36125 | P. citricarpa | 25.85714 | 90.90476 | 145.3727 | 13.2381 | 36.42857 | 36.42857 |
| Monchique‐1 | Portugal | −8.52623 | 37.3262 | 49044 | P. citricarpa | 54.57143 | 150.0476 | 241.1149 | 15.04762 | 30 | 30 |
| Trebisacce‐1 | Italy | 16.56482 | 39.91012 | 54132 | P. citricarpa | 34.42857 | 56.19048 | 60.90476 | 10 | 11.52381 | 11.52381 |
| Zurrieq‐1 | Malta | 14.5051 | 35.82385 | 35126 | P. citricarpa | 24.66667 | 91.90476 | 151.9951 | 10.47619 | 25.42857 | 25.42857 |
| Acitrezza‐1 | Italy | 15.16109 | 37.56108 | 43128 | Other | 16.71429 | 45.09524 | 57.61412 | 6.285714 | 8.095238 | 8.095238 |
| Agia‐1 | Greece | 23.92124 | 35.46598 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| Algemesi‐1 | Spain | −0.44977 | 39.20764 | 52073 | Other | 60.19048 | 105.2857 | 122.1264 | 19.52381 | 28.80952 | 28.80952 |
| Algemesi‐2 | Spain | −0.47082 | 39.1969 | 52073 | Other | 60.19048 | 105.2857 | 122.1264 | 19.52381 | 28.80952 | 28.80952 |
| Alginet‐1 | Spain | −0.45803 | 39.26007 | 52073 | Other | 60.19048 | 105.2857 | 122.1264 | 19.52381 | 28.80952 | 28.80952 |
| Alginet‐2 | Spain | −0.41642 | 39.25141 | 52073 | Other | 60.19048 | 105.2857 | 122.1264 | 19.52381 | 28.80952 | 28.80952 |
| Alhaurin_El_Grande‐1 | Spain | −4.67709 | 36.64537 | 43057 | Other | 39.33333 | 111.619 | 171.133 | 12.80952 | 25.61905 | 25.90476 |
| Alhaurin_El_Grande‐2 | Spain | −4.69818 | 36.66469 | 43057 | Other | 39.33333 | 111.619 | 171.133 | 12.80952 | 25.61905 | 25.90476 |
| Alikianos‐1 | Greece | 23.90863 | 35.45666 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| Alikianos‐2 | Greece | 23.90437 | 35.46238 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| Alikianos‐3 | Greece | 23.9198 | 35.44644 | 37160 | Other | 16.33333 | 53.47619 | 63.04762 | 4.142857 | 6.809524 | 6.809524 |
| Alikianos‐4 | Greece | 23.94556 | 35.46622 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| Almeria‐1 | Spain | −2.40293 | 36.83464 | 43065 | Other | 11.2381 | 31.38095 | 42.56814 | 3.857143 | 8.333333 | 8.333333 |
| Almeria‐2 | Spain | −2.40289 | 36.82883 | 43065 | Other | 11.2381 | 31.38095 | 42.56814 | 3.857143 | 8.333333 | 8.333333 |
| Alzira‐1 | Spain | −0.49072 | 39.15696 | 52073 | Other | 60.19048 | 105.2857 | 122.1264 | 19.52381 | 28.80952 | 28.80952 |
| Amfilochia‐1 | Greece | 21.17164 | 38.96138 | 51148 | Other | 27.90476 | 82.57143 | 95.70936 | 13 | 17.61905 | 17.61905 |
| Argo‐1 | Greece | 22.74218 | 37.62865 | 46154 | Other | 15.47619 | 59 | 63.04762 | 4.238095 | 10.90476 | 10.90476 |
| Argo‐2 | Greece | 22.73931 | 37.65556 | 46154 | Other | 15.47619 | 59 | 63.04762 | 4.238095 | 10.90476 | 10.90476 |
| Argos‐1 | Greece | 22.66172 | 37.68659 | 46154 | Other | 15.47619 | 59 | 63.04762 | 4.238095 | 10.90476 | 10.90476 |
| Arta‐2 | Greece | 20.90379 | 39.15566 | 52147 | Other | 36.09524 | 95.80952 | 111.9852 | 17.90476 | 24.52381 | 24.52381 |
| Arta‐3 | Greece | 20.91826 | 39.16047 | 52147 | Other | 36.09524 | 95.80952 | 111.9852 | 17.90476 | 24.52381 | 24.52381 |
| Barcellona P.G.‐1 | Italy | 15.13679 | 38.11056 | 45127 | Other | 32.61905 | 68.09524 | 72.19048 | 7.952381 | 10 | 10 |
| Brucoli‐1 | Italy | 15.11052 | 37.29482 | 42128 | Other | 22.33333 | 60.47619 | 81.28407 | 8.857143 | 12.95238 | 12.95238 |
| Canicattí‐1 | Italy | 13.8409 | 37.35843 | 42123 | Other | 15.95238 | 43.90476 | 49.61248 | 6.380952 | 9.333333 | 9.333333 |
| Carruba‐1 | Italy | 15.19094 | 37.68463 | 43128 | Other | 16.71429 | 45.09524 | 57.61412 | 6.285714 | 8.095238 | 8.095238 |
| Castello‐1 | Spain | −0.0862 | 39.90392 | 55075 | Other | 57.66667 | 100.5238 | 124.1429 | 23.71429 | 34.14286 | 34.38095 |
| Castello‐2 | Spain | −0.08823 | 39.88386 | 55075 | Other | 57.66667 | 100.5238 | 124.1429 | 23.71429 | 34.14286 | 34.38095 |
| Castello‐3 | Spain | −0.09095 | 39.88401 | 55075 | Other | 57.66667 | 100.5238 | 124.1429 | 23.71429 | 34.14286 | 34.38095 |
| Cefalú‐1 | Italy | 14.01227 | 38.02948 | 45124 | Other | 43.2381 | 93.33333 | 107.468 | 21.42857 | 33.14286 | 33.14286 |
| Chania‐1 | Greece | 24.05114 | 35.49315 | 38161 | Other | 13.09524 | 72.19048 | 105.1429 | 8.285714 | 19.90476 | 19.90476 |
| Chania‐2 | Greece | 23.94806 | 35.47789 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| citricarpa‐1 | NA | −8.51451 | 37.33241 | 49044 | Other | 54.57143 | 150.0476 | 241.1149 | 15.04762 | 30 | 30 |
| Comiso‐1 | Italy | 14.63716 | 36.94398 | 40126 | Other | 21.71429 | 56.14286 | 64.09524 | 12.09524 | 15.80952 | 15.80952 |
| Conceicao‐1 | Portugal | −7.91693 | 37.04848 | 47046 | Other | 36.14286 | 99 | 177.4778 | 12.2381 | 25.42857 | 25.42857 |
| Curiglia‐1 | Italy | 16.20376 | 38.76773 | 48131 | Other | 70.85714 | 140.1429 | 163.3563 | 19.04762 | 24.80952 | 24.80952 |
| El_Ejido‐1 | Spain | −2.71999 | 36.79521 | 43064 | Other | 17.14286 | 32.66667 | 38.57143 | 2.142857 | 3.571429 | 3.571429 |
| Estellencs‐1 | Spain | 2.481876 | 39.6535 | 53083 | Other | 45.71429 | 95.04762 | 103.4762 | 10.66667 | 12.90476 | 12.90476 |
| Faro‐1 | Portugal | −7.91681 | 37.10846 | 48046 | Other | 29.80952 | 84.47619 | 132.6929 | 14.80952 | 23.28571 | 23.28571 |
| Faro‐2 | Portugal | −7.91743 | 37.06264 | 47046 | Other | 36.14286 | 99 | 177.4778 | 12.2381 | 25.42857 | 25.42857 |
| Giarratana‐1 | Italy | 14.97442 | 36.88344 | 40127 | Other | 26 | 69 | 74.47619 | 9.428571 | 14 | 14 |
| Gouria‐1 | Greece | 21.25765 | 38.45498 | 49149 | Other | 35.95238 | 107.4286 | 137.9179 | 18 | 30.90476 | 30.90476 |
| Gozo‐2 | Malta | 14.2593 | 36.04907 | 36125 | Other | 25.85714 | 90.90476 | 145.3727 | 13.2381 | 36.42857 | 36.42857 |
| Gozo‐3 | Malta | 14.26012 | 36.03753 | 36125 | Other | 25.85714 | 90.90476 | 145.3727 | 13.2381 | 36.42857 | 36.42857 |
| Gozo‐4 | Malta | 14.27936 | 36.04965 | 36125 | Other | 25.85714 | 90.90476 | 145.3727 | 13.2381 | 36.42857 | 36.42857 |
| Gozo‐5 | Malta | 14.28445 | 36.05817 | 36125 | Other | 25.85714 | 90.90476 | 145.3727 | 13.2381 | 36.42857 | 36.42857 |
| Grotte‐1 | Italy | 15.17701 | 37.67993 | 43128 | Other | 16.71429 | 45.09524 | 57.61412 | 6.285714 | 8.095238 | 8.095238 |
| Guardia‐1 | Italy | 15.17592 | 37.66271 | 43128 | Other | 16.71429 | 45.09524 | 57.61412 | 6.285714 | 8.095238 | 8.095238 |
| Kirtomados‐1 | Greece | 23.91666 | 35.47875 | 38160 | Other | 15.80952 | 88.7619 | 130.2381 | 6.238095 | 18.42857 | 18.66667 |
| Leni‐1 | Italy | 14.59752 | 38.04442 | 45126 | Other | 29.90476 | 68 | 75.09524 | 11.66667 | 14.42857 | 14.42857 |
| Leni‐2 | Italy | 14.82713 | 38.55289 | 47126 | Other | NA | NA | NA | NA | NA | NA |
| Lentini‐1 | Italy | 15.0209 | 37.32058 | 42127 | Other | 27.2381 | 64.33333 | 81.36617 | 39.04762 | 50.09524 | 50.09524 |
| Malaga‐1 | Spain | −4.42706 | 36.76176 | 44058 | Other | 23.19048 | 48.2381 | 50.95238 | 3.714286 | 4.571429 | 4.571429 |
| Mascali‐1 | Italy | 15.1925 | 37.76768 | 44128 | Other | 20.14286 | 45.71429 | 54.33333 | 9.619048 | 10.66667 | 10.66667 |
| Mascali‐2 | Italy | 15.19464 | 37.76826 | 44128 | Other | 20.14286 | 45.71429 | 54.33333 | 9.619048 | 10.66667 | 10.66667 |
| Massafra‐1 | Italy | 17.14411 | 40.54476 | 57134 | Other | 39.04762 | 62.14286 | 62.57143 | 9.333333 | 10.95238 | 10.95238 |
| Mastro‐1 | Greece | 21.28054 | 38.43029 | 49149 | Other | 35.95238 | 107.4286 | 137.9179 | 18 | 30.90476 | 30.90476 |
| Mesquita‐1 | Portugal | −8.28949 | 37.21367 | 48045 | Other | 41.38095 | 109.5238 | 189.0903 | 12.90476 | 31.04762 | 31.04762 |
| Mesquita‐2 | Portugal | −8.29781 | 37.20453 | 48045 | Other | 41.38095 | 109.5238 | 189.0903 | 12.90476 | 31.04762 | 31.04762 |
| Mineo‐1 | Italy | 14.69086 | 37.35072 | 42126 | Other | 17.71429 | 45 | 54.2381 | 11 | 14.47619 | 14.47619 |
| Moncada‐1 | Spain | −0.39458 | 39.58855 | 54074 | Other | 56.61905 | 97.57143 | 115.0443 | 17 | 24.95238 | 24.95238 |
| Monchique‐2 | Portugal | −8.50369 | 37.33623 | 49044 | Other | 54.57143 | 150.0476 | 241.1149 | 15.04762 | 30 | 30 |
| Monchique‐3 | Portugal | −8.49223 | 37.33224 | 49044 | Other | 54.57143 | 150.0476 | 241.1149 | 15.04762 | 30 | 30 |
| Motta_S._Anastasia‐1 | Italy | 14.88602 | 37.4821 | 42127 | Other | 27.2381 | 64.33333 | 81.36617 | 39.04762 | 50.09524 | 50.09524 |
| Motta_S._Anastasia‐2 | Italy | 14.95416 | 37.46971 | 42127 | Other | 27.2381 | 64.33333 | 81.36617 | 39.04762 | 50.09524 | 50.09524 |
| Nafplio‐1 | Greece | 22.78527 | 37.58931 | 46155 | Other | 21.14286 | 52.28571 | 73.66174 | 10.71429 | 15.71429 | 15.71429 |
| Nafplio‐2 | Greece | 22.69559 | 37.5751 | 46154 | Other | 15.47619 | 59 | 63.04762 | 4.238095 | 10.90476 | 10.90476 |
| Nafplio‐3 | Greece | 22.6968 | 37.58229 | 46154 | Other | 15.47619 | 59 | 63.04762 | 4.238095 | 10.90476 | 10.90476 |
| Nafplio‐4 | Greece | 22.87484 | 37.5888 | 46155 | Other | 21.14286 | 52.28571 | 73.66174 | 10.71429 | 15.71429 | 15.71429 |
| Nicolosi‐1 | Italy | 15.02948 | 37.61127 | 43127 | Other | 18.33333 | 42 | 52.09524 | 8.047619 | 9.285714 | 9.285714 |
| Niscemi‐1 | Italy | 14.3934 | 37.13978 | 41125 | Other | 20.28571 | 53.2381 | 65.33333 | 10 | 17.14286 | 17.14286 |
| Noto‐1 | Italy | 15.09545 | 36.8465 | 40128 | Other | 25.47619 | 76.47619 | 100.7603 | 9 | 18.14286 | 18.14286 |
| Pachino‐1 | Italy | 15.08699 | 36.72003 | 39128 | Other | 24.66667 | 107.1905 | 165.33 | 14.90476 | 39.85714 | 39.85714 |
| Pachino‐2 | Italy | 15.08941 | 36.72233 | 39128 | Other | 24.66667 | 107.1905 | 165.33 | 14.90476 | 39.85714 | 39.85714 |
| Pedara‐1 | Italy | 15.06654 | 37.60871 | 43127 | Other | 18.33333 | 42 | 52.09524 | 8.047619 | 9.285714 | 9.285714 |
| Pizzo Calabro‐1 | Italy | 16.22601 | 38.76039 | 48131 | Other | 70.85714 | 140.1429 | 163.3563 | 19.04762 | 24.80952 | 24.80952 |
| Ribera‐1 | Italy | 13.24185 | 37.49711 | 42121 | Other | 42.47619 | 98.04762 | 124.7094 | 21.57143 | 30.95238 | 30.95238 |
| Ribera‐2 | Italy | 13.25207 | 37.50442 | 42121 | Other | 42.47619 | 98.04762 | 124.7094 | 21.57143 | 30.95238 | 30.95238 |
| Riposto‐1 | Italy | 15.19935 | 37.69647 | 43128 | Other | 16.71429 | 45.09524 | 57.61412 | 6.285714 | 8.095238 | 8.095238 |
| Rocca_Imperiale‐1 | Italy | 16.61795 | 40.10839 | 54132 | Other | 34.42857 | 56.19048 | 60.90476 | 10 | 11.52381 | 11.52381 |
| San_Gregorio‐1 | Italy | 15.10097 | 37.5623 | 43127 | Other | 18.33333 | 42 | 52.09524 | 8.047619 | 9.285714 | 9.285714 |
| Scordia‐1 | Italy | 14.86915 | 37.28153 | 42127 | Other | 27.2381 | 64.33333 | 81.36617 | 39.04762 | 50.09524 | 50.09524 |
| Seville‐1 | Spain | −5.96282 | 37.50854 | 48053 | Other | 22.7619 | 88.47619 | 144.9557 | 9.809524 | 27.14286 | 27.14286 |
| Seville‐2 | Spain | −5.95491 | 37.48298 | 48053 | Other | 22.7619 | 88.47619 | 144.9557 | 9.809524 | 27.14286 | 27.14286 |
| Sikoula‐1 | Greece | 21.0834 | 39.08593 | 52148 | Other | 27.7619 | 65.57143 | 71.47619 | 14.33333 | 16.33333 | 16.33333 |
| Silves‐1 | Portugal | −8.39084 | 37.16415 | 48044 | Other | 52.33333 | 128.0476 | 234.5599 | 16.33333 | 33.19048 | 33.19048 |
| Soller‐1 | Spain | 2.709609 | 39.76453 | 53084 | Other | 46.33333 | 91.28571 | 101.2742 | 17.2381 | 20.66667 | 20.66667 |
| Soller‐2 | Spain | 2.7266 | 39.77012 | 53084 | Other | 46.33333 | 91.28571 | 101.2742 | 17.2381 | 20.66667 | 20.66667 |
| Terme_Vigliatore‐1 | Italy | 15.16324 | 38.1458 | 45128 | Other | 27.19048 | 54.7619 | 62.8555 | 6.809524 | 8.095238 | 8.095238 |
| Torremolinos‐1 | Spain | −4.50413 | 36.67272 | 43057 | Other | 39.33333 | 111.619 | 171.133 | 12.80952 | 25.61905 | 25.90476 |
| Trebisacce‐2 | Italy | 16.56063 | 39.90671 | 54132 | Other | 34.42857 | 56.19048 | 60.90476 | 10 | 11.52381 | 11.52381 |
Appendix C – Data provenance
1.
The following table shows for each map on which data files it is based on.The data files are deposited on http://zenodo.org, as 10.5281/zenodo.1003121 (https://doi.org/10.5281/zenodo.1003121)
The data files are available as part of the upper mentioned deposit.
The identity of the data files can be verified with the following md5 checksums:
| 984cd97090f1d84cce8659f3cf9855c0 | asco_avg_WSPR.zip |
| 3181603536251722eb5046758941fe01 | euCitrusSurface.rda |
| 09bddeda76d4f428a66701658bd68f10 | Guarnaccia_et al 2017 ‐ comparison with model predictions.xlsx |
| fc76546c6ad19fd7c43b0b292290cb47 | locations.csv |
| 34117b40e8ab9a456a1076b7aa5acf63 | mag2015_table 1.csv |
| 5fa0b207f62e2e2c3e1bd17aa1aa1f7f | mag2015_table 2.csv |
| 41d56dec5592f8187982af679cc9cc4a | martinez2015.zip |
| bfdf72bb018eb8b6d9e00220d58d83b4 | pycnidio_avg_WR.zip |
| 9fc7144478dee24db65f48a6f18c9bda | spores2.zip |
| 74be3b7bb8b56ba29b7f52e0dcc47990 | Yonow_et_al_2013_CLIMEX_with_10minresolution_1961‐1990.zip |
| 4dd6a44c815cd33db4adf5285eddd737 | Yonow_et_al_2013_CLIMEX_with_JRC.zip |
The complete computing environment, which allows easy reproduction of all maps, is deposited as well as a Docker container: Doi: 10.5281/zenodo.1044096 (https://doi.org/10.5281/zenodo.1044096)
The tables mag2015_table 1.csv and mag2015_table 2.csv are originated from Magarey et al. (2015), accessible here: http://www.sciencedirect.com/science/article/pii/S0261219415300387
Suggested citation: EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger M, Bragard C, Caffier D, Candresse T, Chatzivassiliou E, Dehnen‐Schmutz K, Gilioli G, Grégoire J‐C, Jaques Miret JA, MacLeod A, Navajas Navarro M, Niere B, Parnell S, Potting R, Rafoss T, Rossi V, Urek G, Van Bruggen A, Van Der Werf W, West J, Winter S, Baker R, Fraaije B, Vicent A, Behring C, Mosbach Schulz O and Stancanelli G, 2018. Scientific Opinion on the evaluation of a paper by Guarnaccia et al. (2017) on the first report of Phyllosticta citricarpa in Europe. EFSA Journal 2018;16(1):5114, 48 pp. 10.2903/j.efsa.2018.5114
Requestor: European Commission
Question number: EFSA‐Q‐2017‐00540 and EFSA‐Q‐2017‐00541
Panel members: Claude Bragard, David Caffier, Thierry Candresse, Elisavet Chatzivassiliou, Katharina Dehnen‐Schmutz, Gianni Gilioli, Jean‐Claude Grégoire, Josep Anton Jaques Miret, Michael Jeger, Alan MacLeod, Maria Navajas Navarro, Bjoern Niere, Stephen Parnell, Roel Potting, Trond Rafoss, Vittorio Rossi, Gregor Urek, Ariena Van Bruggen, Wopke Van Der Werf, Jonathan West and Stephan Winter.
Acknowledgements: The Panel wishes to thank the following for the support provided to this scientific output: Mitesha Aukhojee, Ciro Gardi and Emanuela Tacci. The Panel wishes to acknowledge all European competent institutions, Member State bodies and other organisations that provided data for this scientific output.
Adopted: 21 November 2017
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder:
Figure 2a © 2017 Google Earth, CNES/Airbus; Figure 2b: © Malta NPPO; Figure 2c: © 2017 Google Earth, CNES/Airbus; Figure 2d: © Malta NPPO; Figure 2e: © Portugal NPPO; Figure 2f: © 2017 Google Earth, TerraMetrics
Notes
When the locations sampled by Guarnaccia et al. (2017) were examined in Google Earth or by field visits by the NPPOs, those locations described as backyards appear to vary from large hotel gardens to relatively few citrus trees in a small garden.
Current name according to Index Fungorum (Anonymous, 2016) is Rhynchosporium graminicola Heinsen (syn. R. commune used by Looseley et al., 2012).
As required by Directive 2000/29/EC as last amended, in Articles 15a and 16, and addressed in national plant health legislations (for Italy, see the Decreto Legislativo 19 agosto 2005, n. 214, art. 8; for Malta, see the Maltese Subsidiary Legislation 433.03 Regulation 5b; for Portugal, see the Portuguese Plant Health Law ‐ Decree no 154/2005 and its amendments, article 30). Moreover, the species P. citricarpa is a listed harmful organism by Directive 2000/29/EC. Therefore, the conditions under which P. citricarpa may be moved within the EU for scientific purposes are clearly established by Commission Directive 2008/61/EC.
Personal communication from Paula Cruz de Carvalho, Direcção‐Geral de Alimentação e Veterinária (DGAV), Lisboa (PT), 7 June 2017; Personal communication from Sharlo Camilleri, Veterinary and Phytosanitary Regulation Department, Plant Health Directorate, Lija (MT), 8 June 2017; Personal communication from Michele Ghezzi, Direzione Generale dello Sviluppo Rurale, Ufficio DISR V, Servizio Fitosanitario Centrale, Roma (IT), 16 June 2017.
Personal communication from Pedro Crous, Westerdijk Fungal Biodiversity Institute, Utrecht (NL), 9 November 2017.
Note dated 11 September 2017 of the Plant Health Directorate, Lija (MT).
Note dated 18 September 2017 of the DGAV, Lisboa (PT).
References
- Agri4castWiki , 2015. Joint Research Center, Interpolation of observed weather. Available online: http://marswiki.jrc.ec.europa.eu/agri4castwiki/index.php/Interpolation_of_observed_weather
- Agustí M, 2012. Citricultura. Ediciones Mundi‐Prensa, Madrid. p. 422. [Google Scholar]
- Agustí M, Almela V and Juan M, 2004. Alteraciones fisiológicas de los frutos cítricos. Secretaría General Técnica, MAPA, Madrid, 126 pp. [Google Scholar]
- Amat SG, 1988. Defectos y alteraciones de los frutos cítricos en su comercialización. Comité de Gestión para la Exportación de Cítricos, Almassora, 153 pp.
- Amorim R, Daiani CS, Ferreira‐Maba L, Aluizio R, Goulin EH, Takita MA, Machado MA and Glienke C, 2016. MAT gene idiomorphs suggest a heterothallic sexual cycle in the citrus pathogen Phyllosticta citricarpa . European Journal of Plant Pathology, 147, 325–337. [Google Scholar]
- Anonymous , 2016. Available online: http://www.speciesfungorum.org/Names/SynSpecies.asp?RecordID=155274
- Baldassari RB, Reis RF and de Goes A, 2006. Susceptibility of fruits of the Valência and Natal sweet orange varieties to Guignardia citricarpa and the influence of the coexistence of healthy and symptomatic fruits. Fitopatologia Brasileira, 31, 337–341. [Google Scholar]
- Bonants PJM, Carroll GC, de Weerdt M, van Brouwershaven IR and Baayen RP, 2003. Development and validation of a fast PCR‐based detection method for pathogenic isolates of the citrus black spot fungus, Guignardia citricarpa . European Journal of Plant Pathology, 109, 503–513. [Google Scholar]
- Bregaglio S, Donatelli M, Confalonieri R, Acutis M and Orlandini S, 2010. An integrated evaluation of thirteen modelling solutions for the generation of hourly values of air relative humidity. Theoretical and Applied Climatology, 102, 429–438. 10.1007/s00704-010-0274-y [DOI] [Google Scholar]
- Brentu FC, Oduro KA, Offei SK, Odamtten GT, Vicent A, Peres NA and Timmer LW, 2012. Crop loss, aetiology, and epidemiology of citrus black spot in Ghana. European Journal of Plant Pathology, 133, 657–670. [Google Scholar]
- Carstens E, Le Roux HF, Holtzhausen MA, Van Rooyen L, Coetzee J, Wentzel R, Laubscher W, Dawood Z, Venter E, Schutte G, Fourie P and Hattingh V, 2012. Citrus black spot is absent in the Western Cape, Northern Cape and Free State Provinces. South African Journal of Science, 108, 71–77. [Google Scholar]
- EFSA (European Food Safety Authority), 2008. Scientific Opinion of the Panel on Plant Heath (PLH) on a request from the European Commission on Guignardia citricarpa Kiely. EFSA Journal 2008;7(1):925, 108 pp. 10.2903/j.efsa.2009.925 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2009. Guidance on the transparency in the Scientific Aspects of Risk Assessments carried out by EFSA. Part 2: general principles. EFSA Journal 2009;7(5):1051, 22 pp. 10.2903/j.efsa.2009.1051 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Plant Health Panel), 2014. Scientific Opinion on the risk of Phyllosticta citricarpa (Guignardia citricarpa) for the EU territory with identification and evaluation of risk reduction options. EFSA Journal 2014;12(2):3557, 243 pp. 10.2903/j.efsa.2014.3557 [DOI] [Google Scholar]
- EFSA PLH Panel (EFSA Plant Health Panel), 2016. Evaluation of new scientific information on Phyllosticta citricarpa in relation to the EFSA PLH Panel (2014) Scientific Opinion on the plant health risk to the EU. EFSA Journal 2016;14(6):4513, 53 pp. 10.2903/j.efsa.2016.4513 [DOI] [Google Scholar]
- EFSA Scientific Committee , 2014. Guidance on the structure and content of the EFSA scientific opinions and statements. EFSA Journal 2014;12(9):3808, 10 pp. 10.2903/j.efsa.2014.3808 [DOI] [Google Scholar]
- EFSA Scientific Committee , 2016. Guidance on uncertainty in EFSA Scientific Assessment (draft submitted to public consultation on 18 June 2015
- EPPO/OEPP (European and Mediterranean Plant Protection Organization), 2009. Standards: diagnostic protocols for regulated pests: PM 7/17(2) Guignardia citricarpa . EPPO Bulletin, 39, 318–327. [Google Scholar]
- Fourie P, Schutte T, Serfontein S and Swart F, 2013. Modelling the effect of temperature and wetness on Guignardia pseudothecium maturation and ascospore release in citrus orchards. Phytopathology, 103, 281–292. [DOI] [PubMed] [Google Scholar]
- Frankland JC, Dighton J and Boddy L, 1990. Methods for studying fungi in soil and forest litter. Methods in Microbiology, 22, 343–404. [Google Scholar]
- Gams W and Jaklitsch W and 77 signatories, 2011. A critical response to the ‘Amsterdam Declaration’. Mycotaxon, 116, 1–12. [Google Scholar]
- Garrán SM, 1996. Citrus black spot in the Northeast of Entre Rios: Etiology, epidemiology and control. Proceedings of the International Society of Citriculture, Sun City, South Africa, 1, 466–471. [Google Scholar]
- Gladieux P, Feurtey A, Hood ME, Snirc A, Clavel J, Dutech C, Roy M and Giraud T, 2015. The population biology of fungal invasions. Molecular Ecology, 24, 1969–1986. [DOI] [PubMed] [Google Scholar]
- Grout TG, 2015. The status of citrus IPM in South Africa. Acta Horticulture, 1065, 1091–1095. [Google Scholar]
- Guarnaccia V, Groenewald JZ, Li H, Glienke C, Carstens E, Hattingh V, Fourie PH and Crous PW, 2017. First report of Phyllosticta citricarpa and description of two new species, P. paracapitalensis and P. paracitricarpa, from citrus in Europe. Studies in Mycology, 87, 161–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendricks KE, Christman MC and Roberts PD, 2017. Spatial and temporal patterns of commercial citrus trees affected by Phyllosticta citricarpa in Florida. Scientific Reports, 7, 1641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu J, Johnson EG, Wang NY, Davoglio T and Dewdney MM, 2014. qPCR quantification of pathogenic Guignardia citricarpa and non‐pathogenic G. mangiferae in citrus. Plant Disease, 98, 112–120. [DOI] [PubMed] [Google Scholar]
- ISPM 27 (International Standards for Phytosanitary Measures), 2016. Diagnostic protocols for regulated pests DP 5: Phyllosticta citricarpa (McAlpine) Aa on fruit. International Plant Protection Convention (IPPC), FAO, Rome. [Google Scholar]
- Johnston RP, 1998. Leaf endophytes of manuka (Leptospermum scoparium). Mycological Research, 102, 1009–1016. [Google Scholar]
- Kalaris T, Fieselmann D, Magarey R, Colunga‐Garcia M, Roda A, Hardie D, Cogger N, Hammond N, Martin PAT and Whittle P, 2014. Chapter 11: the Role of surveillance methods and technologies in plant biosecurity. Other Publications in Zoonotics and Wildlife Disease. 172. http://digitalcommons.unl.edu/zoonoticspub/172
- Korf HJG, 1998. Survival of Phyllostica citricarpa, anamorph of the citrus black spot pathogen. MSc Thesis, University of Pretoria
- Kotzé JM, 1981. Epidemiology and control of citrus black spot in South Africa. Plant Disease, 65, 945–950. [DOI] [PubMed] [Google Scholar]
- Kotzé JM, 2000. Black spot. In: Garnsey SM and Graham JH (eds). Compendium of Citrus Diseases (Timmer LW, 2nd edition. The American Phytopathological Society, St. Paul, MN, USA. pp. 23–25. [Google Scholar]
- Lanza FEE, Metzker TG, Vinhas T, Behlau F and Silva GJJ, 2017. Critical fungicide spray period for citrus black spot control in São Paulo state, Brazil. Plant Disease, 10.1094/pdis-04-17-0537-re [DOI] [PubMed] [Google Scholar]
- Looseley ME, Newton AC, Atkins SD, Fitt BDL, Fraaije BA, Thomas WTD, Keith R, Macaulay M, Lynott J and Harrap D, 2012. Genetic basis of control of Rhynchosporium secalis infection and symptom expression in barley. Euphytica, 184, 47–56. [Google Scholar]
- Magarey RD, Sutton TB and Thayer CL, 2005. A simple generic infection model for foliar fungal plant pathogens. Phytopathology, 95, 92–100. [DOI] [PubMed] [Google Scholar]
- Magarey RD, Hong SC, Fourie PH, Christie DN, Miles AK, Schutte GC and Gottwald TR, 2015. Prediction of Phyllosticta citricarpa using an hourly infection model and validation with prevalence data from South Africa and Australia. Crop Protection, 75, 104–114. [Google Scholar]
- Meyer L, Jacobs R, Kotzé JM, Truter M and Korsten L, 2012. Detection and molecular identification protocols for Phyllosticta citricarpa from citrus matter. South African Journal of Science, 108, 53–59. [Google Scholar]
- New M, Hulme M and Jones PD, 1999. Representing twentieth century space‐time climate variability. Part 1: development of a 1961–90 mean monthly terrestrial climatology. Journal of Climate, 12, 829–856. [Google Scholar]
- New M, Lister D, Hulme M and Makin I, 2002. A high‐resolution data set of surface climate over global land areas. Climate Research, 21, 1–25. [Google Scholar]
- Paul I, van Jaarsveld AS, Korsten L and Hattingh V, 2005. The potential global geographical distribution of Citrus Black Spot caused by Guignardia citricarpa Kiely: likelihood of disease establishment in the European Union. Crop Protection, 24, 297–308. [Google Scholar]
- Perryman SAM and West JS, 2014. Splash dispersal of Pyllosticta citricarpa pycnidiospores from infected citrus. EFSA supporting publication 2014‐EN‐560, 30 pp.
- Perryman SAM, Clark SJ and West JS, 2014. Splash dispersal of Phyllosticta citricarpa conidia from infected citrus fruit. Scientific Reports, 4, 6568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reis RF, de Goes A and Pereira GT, 2003. Efeito da aplicaçao de oxicloreto de cobre em diferentes épocas no controle da mancha preta dos citros causada por Guignardia citricarpa . Summa Phytopathologica, 29, 12–18. [Google Scholar]
- Schubert TS, Dewdney MM, Peres NA, Palm ME, Jeyaprakash A, Sutton B, Mondal SN, Wang N‐Y, Rascoe J and Picton DD, 2012. First report of Guignardia citricarpa associated with citrus black spot on sweet orange (Citrus sinensis) in North America. Plant Disease, 96, 1225. [DOI] [PubMed] [Google Scholar]
- Schutte GC, 1995. Evaluation of control strategies for Citrus Black Spot in Southern Africa. PhD Thesis, pp. 151. University of Pretoria, Pretoria, South Africa, September 1995.
- Silva AD, Savi DC, Raiser PHS, Gonçalves FP, Kava V, Galli‐Terasawa LV and Glienke C, 2016. Epidemiological aspects of Phyllosticta citricarpa colonization and viability in Citrus sinensis . Journal of Plant Diseases and Protection, 124, 73–80. [Google Scholar]
- Sousa PFC and de Goes A, 2010. Reação de laranjeiras‐doces quanto à resistência a Guignardia citricarpa . Revista Brasileira de Fruticultura, 32, 718–725. [Google Scholar]
- Spósito MB, Bassanezin RB and Amorim L, 2004. Resistance to citrus black spot by the analyses of disease progress curves. Fitopatologia Brasileira, 29, 532–537. [Google Scholar]
- Spósito MB, Amorim L, Ribeiro PJ, Bassanezi RB and Krainski ET, 2007. Spatial pattern of trees affected by black spot in citrus groves in Brazil. Plant Disease, 91, 36–40. [DOI] [PubMed] [Google Scholar]
- Spósito MB, Amorim L, Bassanezi RB, Bergamin Filho A and Hau B, 2008. Spatial pattern of black spot incidence within citrus trees related to disease severity and pathogen dispersal. Plant Pathology, 57, 103–108. [Google Scholar]
- Timmer LW, 1999. Diseases of fruit and foliage. In: Timmer LW and Duncan LW (eds.). Citrus health management. APS Press, St. Paul, MN, USA. pp. 107–115. [Google Scholar]
- Tran NT, Miles AK, Dietzgen RG, Dewdney MM, Zhang K, Rollins JA and Drenth A, 2017. Sexual reproduction in the citrus black spot pathogen, Phyllosticta citricarpa . Phytopathology, 107, 732–739. [DOI] [PubMed] [Google Scholar]
- Truter MP, 2010. Epidemiology of citrus black spot disease in South Africa and its impact on phytosanitary trade restrictions. PhD Thesis University of Pretoria, South Africa. 148 pp
- Truter M, Kotzé JM, van Rensberg TNJ and Korsten L, 2004a. A sampler to determine available Guignardia citricarpa [=Phyllosticta citricarpa] inoculum on citrus leaf litter. Biosystems Engineering, 89, 515–519. [Google Scholar]
- Truter MP, Labuschagne M and Korsten L, 2004b. Inoculation of citrus leaves at different leaf ages with Guignardia citricarpa. In: Third Citrus Research Symposium, Citrus Research International, Modimolle, South Africa, p. 37.
- Truter M, Labuschagne PM, Kotze JM, Meyer L and Korsten L, 2007. Failure of Phyllosticta citricarpa pycnidiospores to infect Eureka lemon leaf litter. Australasian Plant Pathology, 36, 87–93. [Google Scholar]
- Vacante V and Calabrese F, 2009. Citrus: trattato di agrumicultura. Edagricole, Milan, Italy, 496 pp. [Google Scholar]
- Waller JM, Ritchie BJ and Holderness M, 1998. Plant clinic handbook. CAB International, Oxfordshire, United Kingdom. [Google Scholar]
- Wang NY, Zhang K, Huguet‐Tapia JC, Rollins JA and Dewdney MM, 2016. Mating type and simple sequence repeat markers indicate a clonal population of Phyllosticta citricarpa in Florida. Phytopathology, 106, 1300–1310. [DOI] [PubMed] [Google Scholar]
- Whiteside JO, 1967. Sources of inoculum of the black spot fungus, Guignardia citricarpa, in infected Rhodesian citrus orchards. The Rhodesia, Zambia and Malawi Journal of Agricultural Research, 5, 171–177. [Google Scholar]
- Wikee S, Lombard L, Crous PW, Nakashima C, Motohashi K, Chukeatirote E, Alias SA, McKenzie EHC and Hyde KD, 2013. Phyllosticta capitalensis, a widespread endophyte of plants. Fungal Diversity, 60, 91–105. [Google Scholar]
- Yonow T, Hattingh V and de Villiers M, 2013. CLIMEX modelling of the potential global distribution of the citrus black spot disease caused by Guignardia citricarpa and the risk posed to Europe. Crop Protection, 44, 18–28. 10.1016/j.cropro.2012.10.006 [DOI] [Google Scholar]
- Weltzien HC, 2012. Use of geophytopathological information. Chapter 16 In: Kranz J and Rotem J (eds.). Experimental techniques in plant disease epidemiology. Springer Science & Business Media, Berlin, Germany. pp. 237‐242. [Google Scholar]
- Zandbergen PA and Barbeau SJ, 2011. Positional accuracy of assisted GPS data from high‐sensitivity GPS‐enabled mobile phones. Journal of Navigation, 64, 381–399. https://www.cambridge.org/core/journals/journal-of-navigation/article/positional-accuracy-of-assisted-gps-data-from-highsensitivity-gpsenabled-mobile-phones/E1EE20CD1A301C537BEE8EC66766B0A9 [Google Scholar]