An official website of the United States government
Here's how you know
Official websites use .gov
A
.gov website belongs to an official
government organization in the United States.
Secure .gov websites use HTTPS
A lock (
) or https:// means you've safely
connected to the .gov website. Share sensitive
information only on official, secure websites.
As a library, NLM provides access to scientific literature. Inclusion in an NLM database does not imply endorsement of, or agreement with,
the contents by NLM or the National Institutes of Health.
Learn more:
PMC Disclaimer
|
PMC Copyright Notice
. Author manuscript; available in PMC: 2020 Feb 10.
1Department of Dermatology, University of Pennsylvania, 226 Clinical Res Building, 415 Curie Boulevard, Philadelphia, PA 19104-6142, USA. Fax:+12155732116
1Department of Dermatology, University of Pennsylvania, 226 Clinical Res Building, 415 Curie Boulevard, Philadelphia, PA 19104-6142, USA. Fax:+12155732116
The publisher's version of this article is available at Exp Dermatol
Abstract
Conventional textbook wisdom portrays the skin as an organ that literally enwraps whatever each of us stands for as a more or less functional, individual member of the mammalian species, and has it that the skin primarily establishes, controls and transmits contacts with the external world. In addition, the skin has long been recognized to protect the organism from deleterious environmental impacts (physical, chemical, microbiological), and is well-known as crucial for the maintenance of temperature, electrolyte and fluid balance.
Now, ever more studies are being published that show the skin to also operate as a huge and highly active biofactory for the synthesis, processing and/or metabolism of an astounding range of e.g. structural proteins, glycans, lipids and signaling molecules. Increasingly, it becomes appreciated that the skin, furthermore, is an integral component of the immune, nervous and endocrine systems, with numerous lines of crosstalk between these systems established intracutaneously (e.g. Ann NY Acad Sci Vol 885, 1999; Endocrine Rev 21:457–487, 2000; Physiol Rev 80: 980–1020, 2001; Exp Dermatol 10: 349–367, 2001).
All these emerging cutaneous functions beyond the classical image of the skin as a barrier and sensory organ are immediately relevant for many of the quandaries that clinical dermatology, dermatopathology, and dermatopharmacology are still struggling with to-date, and offer the practising dermatologist attractive new targets for therapeutic intervention. Yet, many of these skin functions are not even mentioned in dermatology textbooks and await systematic therapeutic targeting. Following a suggestion by Enno Christophers, the current ‘Controversies’ feature brings together an unusually diverse council of biologists and clinicians, who share their thought-provoking views with the readers and allow us to peek into the future of research in cutaneous biology, not the least by reminding us of the – often ignored – evolutionary and embryonal origins of our favorite organ. Hopefully, this unique discussion feature will foster an understanding of the ‘true’ skin functions that is both more comprehensive and more profound than conventional teaching on this topic, and will stimulate more than ‘skin-deep’ reflections on the full range of skin functions.
Introduction
What is the true function of skin?
The above title is full of mischief! Including the word ‘true’ suggests that the skin has ‘false’ functions.
Secondly, the skin has not one but many functions, which are expanding rapidly as our knowledge progresses at its current dizzying pace.
One might invert the title and ask ‘Is there anything that the skin can’t do’? To assay an answer would take a small textbook.
We have in the past greatly underestimated the number and diversity of the skin’s functions. Protection and thermoregulation are embedded in didactic rock but comprise only a small part of the total biologic enterprise.
Perhaps a short list of these newly acquired acquisitions is better than none:
Immunologic
Endocrine
Metabolic
Psycho-social
Neuro-psycho-immunologic
And so on, depending on your particular interests.
In short, the question is impossible to answer in any condensed form. Besides, we live in many skins, from head to foot.
Let us take a biological approach to discuss the function of the skin.
The origin of skin
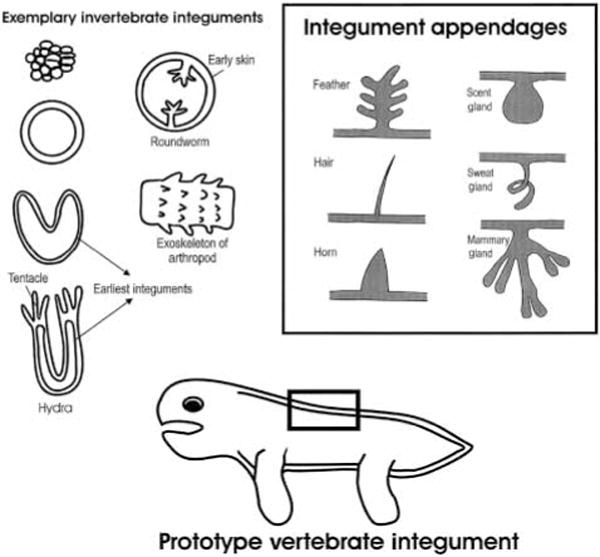
When did the skin start to exist? Early life forms appeared as single cells or groups of cells, and there were no tissues to be named as ‘skin’. About 500–600 million years ago, early multicellular organisms started to try different ways of organization and formed many different animal phyla (1). One of the primitive body plans that have the earliest appearing integument is Cnidaria (2). The organism such as hydra and jelly fish folds into a two layered cylindrical or structure, the ectoderm (epidermis) and the endoderm (gastrodermis). This single layer epidermis can be considered as the earliest form of skin (Fig. 1). Or, let us call it ‘integument’ which has a broader meaning than skin. It indicates the outermost layer of the organism, can be simple or complex. It includes skin or the outer epithelial layer and associated structures.
Prototype of invertebrate integument, vertebrate integument and its appendages.
The first and most basic function of the integument is to set up a boundary between the organism and the environment. It provides the scaffold that originally defines the form of the animal. It sets up as a mechanical as well as chemical barrier for protection of the organism from the harsh environment (Table 1). Within the boundary, cells, tissues and organs are arranged in order and internal homeostasis has to be maintained.
Table 1.
Selective functions of the integument
Protection from the environment
Mechanical, keep the form and internal organs in position and away from damage
Chemical, internal homeostasis in water and land (barrier)
Physical: UV (melanocyte on human)
Keep moist (amphibian) and oily (sebaceous gland)
To be worn off
To heal and regenerate (cytokines)
Defense
Exoskeleton (arthropod)
Armor (turtle, armadillo)
Spiny appendages (porcupine quills)
Inflammatory response (prostaglandin, etc.)
Immune function, with memory of previous stimuli
Weapons
Sting cell of hydra and jelly fish
Claws
Poisonous glands
Communication with outside organisms
Display of messages (pigment pattern, painted skin of human)
To mark territory
Pheromones for sexual attraction
For pack behavior coordination
To scare enemies away
To mimic
Communication with inside organs
Sense the environment (human skin, mouse vibrissa)
Tactile or thermo senses go in through nerves
Endocrine-like function through secretion (neuro-endocrines, endorphin, growth factors, etc.)
Respiration
Insects
Some frogs
Chemical reaction
Vitamin D
Locomotion
Swim (tentacles of hydra, jelly fish and octopus; tube feet of sea cucumber)
Crawl (belly scale of snake)
Glide (skin flap of Pterosaur, bat)
Fly (feathers)
Thermoregulation
Hairs (mammals)
Sweat gland
Dermal blood vessels
Feathers (feathered dinosaurs, birds)
Progeny bearing
Skin flap in toads and abdominal pouch in kangaroos
The second function likely to form is sensory. Organisms have to feel the environment and make appropriate responses for survival. Although organs dedicated to specialized sensory functions soon evolve, the skin retains a major sensory function even today. Reflex responses to heat and other ominous stimuli from nerves to the spinal cord are essential for survival.
Communication with other organisms through display is another function that uses the integument surface as a canvas to scare away predators, to attract each other, or to convey a certain message for collaborative effort.
In invertebrates, the integument and their appendages also work with muscles or a hydraulic system for locomotion. This can be seen in the tentacles of hydra, jelly fish, octopus, etc. Other major categories are for defense (not to be eaten) or weaponry for predation (to eat for survival). In arthropods, external skeletons have evolved to provide a strong armor or framework for protection, locomotion and weaponry. In many animals, the integument is also used for respiration. A summary of these exemplary functions can be seen in Table 1.
Vertebrate integument
In the vertebrate animals, the integument gradually became more complex (3). In the cartilagenous fish, thick skin provided strong protection. In the bony fish, scales became the major form of integument. The overlapping scales provide a more effective way for protection, movement and repair. In many fishes, the surface also became a canvas for pigmentation patterning. The striking pattern displays carry messages for communication. In amphibians, the integument has evolved to serve this class of animals that live between the water and the land. Keeping moisture is of vital importance and therefore skin glands are highly evolved in amphibian skin. From here they also have evolved several specialized functions such as a poisonous gland, fancy colors, or even pouches to carry their youngsters.
The rise of reptiles marks the conquering of the land. The first major evolutionary novelty in this aspect is the evolution of skin barriers. The formation of an effective barrier prevented water loss through the body surface and allowed the animal to go onto the land. Gradually, the reptile scale evolved as a unit of skin for more effective protection. The majority of the scales are short and bumpy structures, although some scales can become elongated and sharp for protection. The scale arrangement pattern and pigmentation (such as snakes) can be complex, suggesting some elaborate communication functions. The reptile integument has evolved for locomotion for crawling on the land (snake ventral scales) or gliding in the sky (Pterosaur). Powerful and sharp claws are the result of weapon competition during dinosaur evolution.
All mammals and birds and some dinosaurs are warm-blooded animals. Two methods for maintaining temperature evolved to offer these animals selective advantages: either to increase heat production or to decrease heat loss.
One of the most effective ways to prevent heat loss is to grow skin appendages that trap the air effectively and maintain body temperature. This need drives the formation of elongated skin appendages such as hairs and down feathers. Although hairs and feathers most likely evolved independently, both form follicles with stem cells hidden in the follicle and both have the ability to cycle and regenerate. In mammals, the evolution of hair has served this function well. Indeed, the length of the hairs has also been changing due to changes of the environment. As elephants migrated north to become the wooly mammoth, the hairs became longer and longer. This may be adjusted by regulating the length of anagen, simply through the activity of FGF5 (4). When whales went back to live in the ocean, hairs were lost because they became an ineffective burden. Instead, blubber, a thick layer of fat, evolved for insulation in the cold water. On the other side of the thermo-regulation are the sweat glands that decrease body temperature through evaporation as needed.
The highest form of integument evolution is achieved in the bird. The elongated and cylindrical appendages branch out to make more complex structures. First, the appendages branch to form barbs that provide a more fluffy down feather coat, highly efficient in thermal regulation. Second, the rachis forms to define feathers of different types, shapes and sizes, and feather is more effective in protection and communication. Third, the barbules form that interweave barbs into a vane and allow the birds to launch into the sky and fly. This opens the whole space in the sky for the Aves class. The recent discovery of the fossils in China provides a window to look at the formative stages of the feathers (reviewed in 5, 6). Some of these skin appendages on the dinosaur are intermediate forms between elongated skin appendages and feathers. When feathers of today’s forms are achieved, these less stable, less efficient intermediate forms were selected out.
Let us come back to look at our own skin. Both characters that define the mammal, hairs and mammary glands, are skin appendage derivatives. The primates have long hairs covering most parts of the body. Humans have terminal hair covering the scalp but most of the body surface is covered by villus hair. This loss of hair during evolution is probably the result of the loss of need for maintaining temperature. Human civilizations have relied upon animal furs and other primitive forms of clothing that have evolved. Growing hairs all over the body is energy consumptive and can also be a problem in hygiene. So humans probably became naked apes gradually, over the few million years of human evolution. Melanin pigment provides protection from UV light and is actually influenced by where those human beings inhabit. In Africa, the need to protect the organism from sunshine favored the formation of dark skin. The humans in the northern climes developed white skin to absorb the rare sunshine for Vitamin D synthesis. Communication with others through display may be taken over simply by changing clothes (rather than changing skin). On the other hand, the communication to other internal organs gradually became more sophisticated, and the skin can have cross talk with neural, psychiatric, endocrine and cardiovascular systems, as well as being a window to show changes in the function of the liver, lung, etc.
True function of the integument
So what are the true functions of the skin? As we have surveyed the animal kingdom, in the beginning it set the boundary between the organism and the environment. It is needed to keep the osmotic pressure, and to prevent heat/water loss. Then comes the basic function of protection. In addition to the mechanical protection, skin also gives UV radiation protection through the melanin pigment. More advanced protection can be seen in the immune system cells in mammalian skin, and the antibiotics secreted in frog skin. Communication has always been important for animals to display messages on their body surfaces. Sensory function also comes early, as animals need to feel the environment. Later specialized sense organs have evolved, but the skin remains as an important sensory organ. It is the most peripheral organ and functions as a detector that communicates with nearly every central-positioned organ through nerves, endocrine systems, cytokines, etc. The formation of skin appendages leads to more elaborate skin functions (7). Secretory glands invaginate in and are important to properties of the skin, attracting the opposite sex, maintaining temperature (sweat gland), and for progeny bearing (mammary gland). Other types of skin appendage protrude from the body, represented by hair and feather. They function in thermo-regulation. Finally, locomotion function served by the skin has been seen in the invertebrate tentacles, snake ventral scales and bat skin flap, but the ultimate form is the feather that allows the birds to fly.
What is the future direction of integument evolution? If the environment changes, we can only hope that the integument of our biological system will have enough plasticity to adapt to the new environment and be as successful as it has been in the last 2 billion years. Therefore, there is no one true function of the skin, but there are many diverse functions of the skin. The weight of different functions depends on the perspectives of different species.
2.Campbell NA, et al. Biology. 5. New York, NY: Addison-Wesley Longman; 1999. [Google Scholar]
3.Bereiter-Hahn J, et al., editors. Biology of the Integument of Vertebrates Berlin. Heidelberg, New York, Tokyo: Springer-Verlag; 1986. [Google Scholar]
1Skin Cancer Research Laboratories, Cardinal Bernardin Cancer Center, Loyola University Medical Center, 2160S. First Avenue, Maywood, IL 60153-5385, USA
1Skin Cancer Research Laboratories, Cardinal Bernardin Cancer Center, Loyola University Medical Center, 2160S. First Avenue, Maywood, IL 60153-5385, USA
In my opinion, the best way to understand the true function of human skin is to examine the structure, cellular configuration, and confederacy of cell types present in normal human skin (Fig. 1). The underlying premise for this perspective is that each resident cell that comprises the skin plays a unique functional role, contributing to the overall homeostasis of this massive organ that covers the body. Because of space constraints, we will only focus on the epidermal compartment, but it should be recognized that both an epidermal immune system and dermal immune system exist, and complement each other (1). To understand the function of skin, it is important to highlight its unique anatomical location – serving as an interface tissue type, subjected not only to the internally generated vicissitudes that all other organ systems sharing a common cardiovascular system are exposed, but it is also being consistently barraged by externally derived factors. The two greatest challenges of this biological interface system are to effectively shield against infectious agents (gram positive/negative bacteria, fungi, viruses); and to defend the body from adverse effects of the sun – including the prevention of desiccation, and at the same time avoidance of premature epidermal cell death so as to preserve the barrier function of skin (2).
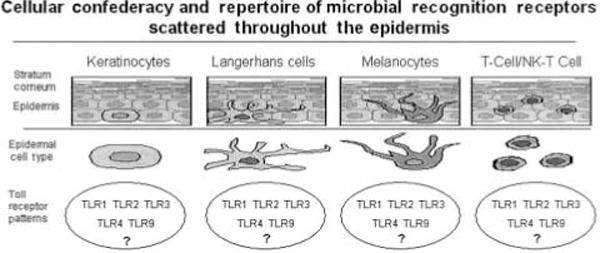
At least 5 different cell types in human epidermis are poised to help defend the epidermis and underlying dermis from attack by biological, chemical, or solar-derived hostile agents. This figure portrays each cell type and potentially important Toll-like receptors that may be expressed by more than one cell type. Collectively, it is implied that the overall pattern of Toll-like receptor expression will embue the epidermis with the capability to recognize and respond to a vast array of infectious insults, so as to rapidly restore cutaneous homeostasis.
Human skin – the ultimate biological shield
How does the skin subserve its anti-infection function? The first layer of the ‘protective coat’ is the stratum corneum, which represents a thin, but highly effective collection of dead keratinocytes, in the outermost aspect of the epidermis (3). It is somewhat ironic that the vitality of skin, and our overall well-being, is entrusted to only a few layers of essentially non-living keratinocytes in the stratum corneum. It is probably not a coincidence that certain genes that encode for proteins importantly involved in formation of the stratum corneum, such as corneodesmosin, are actually located on 6p21.3, which is home for many other genes that regulate immune responses (HLA genes). Thus, I believe it is legitimate to portray the stratum corneum as one of the components of the integument-related innate immune system. Should either bacteria, viruses or fungi break through this biological Saran-wrap, other constituents of the innate immune system are well positioned, and wellequipped to respond. There are at least five different cell types in normal human epidermis that can be called into action once an infectious agent penetrates through the stratum corneum, including: keratinocytes, Langerhans cells, melanocytes, conventional T cells and NK-T cells. These cell types are likely to posses various pathogen recognition alert signaling systems, and can rapidly be triggered to produce a wide assortment of anti-infectious agents as summarized in Table 1 (4).
Table 1.
A summary of antimicrobial agents that can be produced by various cellular constituents of the epidermis that contribute to the ‘Biological Shield’.
The entire field of innate immunity has become one of the most rapidly advancing lines of inquiry during the past 5 years (5). Recently, it has become apparent that plant defense mechanisms against infection share many molecular and cellular components resembling mammalian cutaneous responses (6). A common theme emerging from many laboratories is the fundamental importance of Toll signaling pathways that regulate susceptibility to infection (7). The identification of numerous Toll-like receptors (TLRs) in mammals, and their contribution to innate immune responses, provides new opportunities to dissect out and understand the molecular basis by which human skin can subserve its true function as providing a protective shield for the body (8). While this field is in its infancy as regards TLRs and human epidermis, I predict that each of the various cellular constituents of the epidermis will express discrete (but partially overlapping) patterns of specific TLRs. Thus, keratinocytes will express certain TLRs, that may be different than those TLRs expressed by Langerhans cells, melanocytes, or T cells and NK-T cells (Fig. 1).
I suspect that while any individual epidermal cell type may not be capable of recognizing each class of pathogen (i.e. gram positive vs. gram negative bacteria; or fungi vs. viral infection), when all the confederacy of cell types are taken into account, the full spectrum of TLRs will be represented by their collective presence in human epidermis. This hypothesis can be tested since mAbs and other reagents are becoming available to permit localization for each TLR on individual cell types in normal and diseased human skin samples. It will be of great interest not only to define which resident epidermal cell type express a specific TLR profile, but also to determine which signal transduction pathway is engaged upon challenge with the inciting infections agent. Currently, considerable interest is centered on the NF-κB signaling pathway (9), and at least one autoimmune disease (i.e. Crohn’s disease) has been definitely linked to a genetically defined abnormality in this pathway (10, 11). I raise this point because some patients with Crohn’s disease also suffer from psoriasis, and we have previously postulated a pathogenic role for various cellular components of the innate immune response in this common and enigmatic skin disease (12, 13).
While it is clear that the true function of skin is to subserve an infection-defense function, it is also important to consider the protective function against the adverse effects of the ubiquitous carcinogen and potentially hazardous entity – the sun. Once again, many new insights are being gained by investigators attempting to understand exactly how human epidermal keratinocytes resist apoptosis when exposed to UV light. After all, if there is premature apoptosis of keratinocytes, this interferes with proper barrier formation, and hence may render the individual highly susceptible to infectious assaults following excessive sun exposure (14).
The senescent switch and psoriasis
Returning to psoriasis as our model, we may also gain some new insight into this area of investigative skin biology. I have focused on psoriasis because it is a rather unique disease in which the keratinocytes in the plaque are simultaneously resistant to apoptosis induced by UV light, and at the same time, resistant to transformation (15, 16). In every other clinical scenario, when cells acquire a resistance to apoptosis, this predisposes the tissue containing such cells to transformation; with emergence of malignant clones. To reconcile this apparent paradox, I have suggested that perhaps keratinocytes within psoriatic plaques respond to the presence of chronic inflammation and cytokines produced by the local immune response by undergoing a ‘senescent switch’. That is, they become irreversibly growth arrested, and concomitantly acquire a striking resistance to apoptosis (17). Indeed, I suspect keratinocytes may be distinguished from many other cell types such as endothelial cells, fibroblasts, T cells, etc., by reacting to stress, not by undergoing apoptosis, but by entering a senescent state. By senescence, I do not mean to imply a state of decrepitude. Rather, I believe the senescent state is energy requiring and characterized by a distinct genetic program with active expression of specific genes (18). Perhaps the most important gene that governs senescence in keratinocytes is the p16INK4a locus (17, 19). The main reason for postulating this scenario relates back to the fundamental role of epidermal keratinocytes in creating and maintaining a biological shield in the integument. Thus, keratinocytes cannot die, but must survive and keep the barrier intact.
Keratinocytes, unlike other cell types such as hepatocytes in the liver, cannot die when confronted by a life-threatening challenge, because this would compromise the barrier function of skin. While many hepatocytes can be deleted without producing a clinically significant impact on the metabolic function of the liver, keratinocytes must be considerably more resilient, and have devised a nonapoptotic strategy (i.e. senescent switch) when exposed to pro-apoptotic stimuli either emanating from the sun, or infectious agents, or chemical agents (20).
Summary
In conclusion, it should be clear that the true function of skin is to provide a protective shield by using all of the confederacy of cell types, particularly keratinocytes, to thwart infections and non-infectious threats to maintain the homeostasis of the body. Many new avenues of exploration are now open for investigators, including studies concentrating on TLRs, NF-κB signaling, and p16, to name a few. As new knowledge is gained in this area, it will be possible to devise new highly targeted molecular strategies to preserve, protect and restore barrier function in human skin. Such new insights may also have therapeutic implication beyond barrier function, to include treatment of various dermatoses as well as skin cancers.
Cutaneous protective functions are of paramount importance both under normal conditions and for the vast majority of dermatologic patients. Yet, because dermatologists commonly encounter inflammatory dermatoses in their practices, it is standard dogma to both view and treat these disorders as if they had an immunopathogenic basis (1, 2). Although T cell abnormalities occur in more common dermatoses, such as psoriasis contact dermatitis and atopic dermatitis, these diseases often require external perturbations to provoke disease expression (e.g. Koebner phenomenon in psoriasis). Moreover, true primary, immunologic disorders of the skin (i.e. lupus erythematosus, pemphigus vulgaris, pemphigoid, and vitiligo) are actually quite uncommon. Thus, almost all epidermal functions can be considered protective, and more specifically, defensive (Table 1), and of these functions, most reside in the stratum corneum (SC).
Table 1.
Protective functions of the stratum corneum
Functions
Structural basis
Biochemical mechanisms
Mechanical integrity/resilience
Cornified envelope, cytosolic filaments
Cross-linked peptides; e.g. loricrin; keratin filaments
Even immune phenomena involved in primary defense are triggered by the release of a preformed pool of pro-IL-1a and pro-IL-1b, stored within the corneocyte cytosol. These primary cytokines are poised for release in response to minimal external perturbations (3, 4), and following their release, they signal divergent, downstream pathways that initiate both homeostatic (repair-related) and pro-inflammatory processes (5, 6). This often-pathogenic sequence is based upon the cutaneous defensive function of greatest importance; i.e. the requirement to maintain a competent permeability barrier in a hostile terrestrial environment. These cytokines, along with other signaling molecules, stimulate a variety of metabolic responses aimed at a rapid restoration of normal barrier function by downstream recruitment/entrapment of inflammatory cells. Yet, they often simultaneously initiate a cytokine cascade that stimulates epidermal hyperplasia, inflammation, and perhaps a further barrier abnormality (Fig. 1). Accordingly, many cutaneous inflammatory phenomena, including disease-specific T-cell responses, are recruited merely as incidental participants in a defensive sequence aimed at normalizing SC function. Of course, immunologic mechanisms, once recruited, can further compromise barrier function, leading to the vicious circle shown in Figure 1. Finally, abrogation of the barrier also impacts cutaneous immune functions by limiting (or allowing) the ingress of xenobiotes, including antigens and pathogenic microorganisms.
Logically then, bolstering the skin’s barrier status should increase resistance to inflammation, and further decrease susceptibility to diseases, which are triggered, sustained, or exacerbated by external perturbations, such as atopic dermatitis, contact dermatitis, and psoriasis. Hence, the recent emergence of ‘barrier repair’ strategies to decrease the susceptibility to these disorders. These repair approaches can be classified into three subcategories (6) (Table 2): 1. mixtures of all three physiologic lipids (ceramides, cholesterol, and free fatty acids) in appropriate molar ratios; 2. one or more non-physiologic lipids (e.g. petrolatum, lanolin); 3. dressings, either vapor-permeable, which allow metabolic (repair) processes to continue in the underlying epidermis, or vapor-impermeable, which shut down metabolic responses in the underlying epidermis. But caveat emptor – the term ‘barrier repair’ is frequently applied loosely to emollients, often based upon petrolatum alone, and often inappropriately to formulations that can do more harm than good; i.e., incomplete mixtures of physiologic lipids that impede rather than allow or facilitate normalization of barrier function. Nevertheless, we are now in the era of ‘choice of barrier’ (6), and it now should be possible to select the most logical barrier repair strategy for a specific clinical indication, based upon knowledge of disease pathogenesis (Table 2). Thus, the cutaneous permeability barrier and inflammatory signaling are paramount among the defensive functions of the epidermis.
The outer layers of the epidermis also mediate several other, critical protective functions, including:
water repellency
integrity/cohesion/desquamation
mechanical resistance
resistance to xenobiotics
antimicrobial defense
UV filtration
antioxidant defense
SC hydration
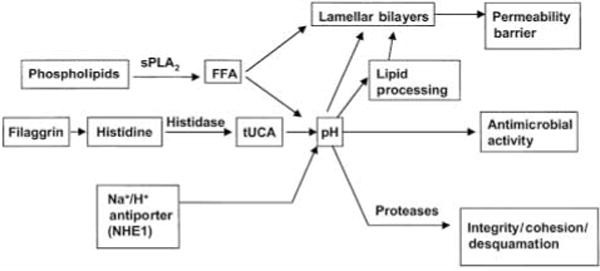
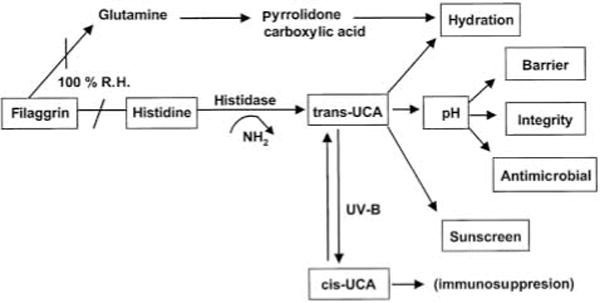
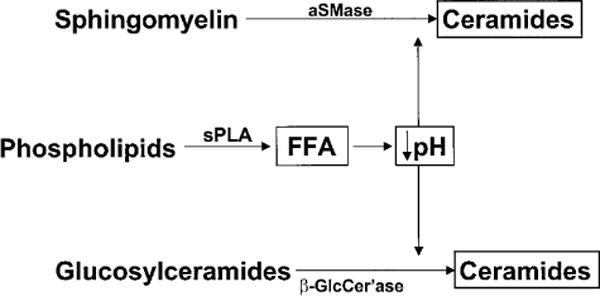
These protective functions are listed in Table 1, which also summarizes current concepts about their structural and biochemical bases. Of these functions, we will focus here on those that are potentially regulated by the pH of the SC. These include not only permeability barrier function and inflammation, as discussed above, but also the related functions of SC integrity, cohesion and des-quamation, as well as antimicrobial defense. Normal SC demonstrates a markedly acidic pH (‘acid mantle’) (7). While the origins of the acidic pH of the SC are incompletely understood, exogenous influences, such as lactic acid in sweat, microbial metabolites, and free fatty acids from sebum, have been considered the likely sources. But recent studies point, instead, to three unrelated, endogenous pathways that contribute to SC acidification (Fig. 2). The first acidifying mechanism results from the deimination of filaggrin-derived histidine to trans-urocanic acid (tUCA) by the enzyme, histidase. This key metabolite, in turn, could impact several pH-related and non-pH-related functions (8, 9) (Fig. 3). With regard to non-pH-dependent, defensive cutaneous functions, tUCA is an effective UV-filter, but as tUCA absorbs incident UV-B, it isomerizes to cis-UCA, a potent immunosup-pressive molecule that is hypothesized to allow development of UV-B-and UV-A-induced skin cancer (10). tUCA is also a potent endogenous humectant; i.e. an important source of SC hydration (7), which in turn regulates skin flexibility, as well as downstream effects on epidermal proliferation (11) (Fig. 3). The second pathway, phospholipid hydrolysis by lamellar body-derived, secretory phospholipase A2 (sPLA2), generates a pool of free fatty acids (FFA), that contributes not only to SC acidification, but also to SC integrity and cohesion (12) (Fig. 2). Third and finally, a sodium-proton membrane antiporter, NHE1, is expressed in the outer nucleated layers of the epidermis, where it acidifies localized membrane domains at the SG– SC interface, with a lesser contribution to the bulk pH of the SC (13) (Fig. 2).
How does the pH of the SC modulate these many functions? With regard to permeability barrier function, after acute insults, the barrier recovers more slowly when exposed to neutral vs. acidic buffers (14). This delay in recovery can be explained by the pH optima of certain key lipid processing enzymes in the SC interstices. While sPLA2 displays a neutral pH optimum, two other key lipid processing enzymes, β-glucocerebrosidase (β-GlcCer’ase) and acidic sphingomyelinase (aS-Mase), are activated at an acidic pH (15). Hence, phospholipid catabolism yields locally acidifying products (12) (i.e. FFA), that, in turn, probably activate β-GlcCer’ase and aSMase, generating ceramides, one of the three key SC barrier lipids, from their polar precursors (Fig. 4). Acidification also regulates SC integrity/cohesion, thereby restricting premature desquamation (12). The basis for this activity relates to corneodesmosome degradation within the SC interstices, a process that requires two serine proteases, the SC chymotryptic and tryptic enzymes, which exhibit neutral-to-alkaline pH optima (e.g. 16). Thus, at an acidic pH, low protease activity presumably restricts corneocyte detachment to the low rates that accompany normal desquamation. Further, the release and activation of IL-1α and IL-1β from their preformed, precursor pools in the SC also requires serine protease activity, including the SC chymotryptic enzyme (17). Thus, the earliest, cutaneous pro-inflammatory events may be triggered by loss of normal SC acidification. Finally, the defensive, antimicrobial function of the skin appears to be dependent upon SC acidification (e.g., 18). Whereas normal flora, such as microccoci and co-rynebacteriae, grow better at an acidic pH, pathogenic organisms, such as staphylococci, streptococci, and candida, proliferate more avidly at a neutral pH (18). Thus, SC pH appears to regulate several of the SC’s key defensive functions.
In summary, we propose that many immune functions of the skin are both secondary and downstream. In fact, immunotherapy is currently highly fashionable, immune processes are of no more importance than a host of other defensive functions embedded in the outer layers of the epidermis, that are equally deserving of therapeutic interventions. In fact, we propose that the cohort of defensive functions of the epidermis, residing largely in the normally acidic SC, are the paramount functions of the skin.
For decades understanding of immune mechanisms has been a central issue in dermatology. As a consequence, a significant number of skin diseases have started to loose their enigmatic features. Early highlights were bullous autoimmune disorders and allergic contact dermatitis and, more recently, others including atopic dermatitis and psoriasis. In these conditions, cutaneous immune reactivity is mirrored by the presence of activated lymphocytes as a causative cell type. In addition, skin is the principal arena for allergies which frequently present as generalized or localized (fixed) drug reactions and result from the interaction of pharmaceutical compounds with the cutaneous immune system. Thus reactive immune responses, drug reactions and autoimmunity represent a majority of unwanted chronic skin disorders.
Since activated T cells are all important players in the field, this has resulted in great admiration for the principles of acquired (adaptive) immunity in skin, and dermatologists tend to eagerly follow the route of thinking delineated by immunologists and transplantation biologists. Although there is little doubt about the essential role of skin in protecting the organism against microbial and parasitic attack, the concept that the tools for survival primarily consist of mechanisms of the adaptive (acquired) immunity has become the prevailing philosophy. This concept of cutaneous function is transmitted to medical students and nurses day by day.
Unfortunately, the idea that protective functions in skin could also be provided by non-T-cell-mediated (innate) pathways has rarely been considered. The fact that cutaneous integrity is largely based upon innate immune mechanisms has nearly been forgotten – despite the fact that skin susceptibility to infection, colonization of skin with potent pathogens and the microbial ecology of skin have all been of major interest, years ago (1, 2).
Considering the myriads of pathogens (and potential pathoens) living on skin and mucous surfaces, one wonders how adaptive immunity by way of it’s complex and time consuming (days) armentarium can possibly provide effective, immediate and continuous protection.
In the following, we will briefly discuss the role of keratinocytes as sentinels at the forefront of microbial invasion in skin. These cells appear vital in recognizing danger from microbial invasion and provide potent means of protection.
Antimicrobial Immunity in Skin
The resident flora of human skin mainly consists of aerobic species with a density ranging from 102 to 107 cells/cm2 (3). Aerobic species are present predominantly in sebaceous areas (face, scalp, mid-line) at a densitiy of 104–106/cm2 (4). Other pathogens of the resident flora include propionibacteria which may cause inflammation of the sebaceous glands and deeper skin sites following surgery. Among fungal elements Pityrosporum, yeasts and temporarily Candida species predominate. On normal skin colony counts of 106 organisms per cm2 remain without the sequelae of inflammation or wound infection.
In view of the distribution and relative density of pathogens it comes as a surprise that skin infections are relatively rare (except in the tropics or at skin sites with a tropical microenvironment like toe webs). Yet even under ‘normal’ conditions, human skin is constantly traumatized, especially around the nail folds, the infundibula of hair, the oral, nasal and anal orifices. This causes exposure of (living) keratinocytes to the cutaneous microflora.
Since the presence of microbes at such sites is not causing infection it follows that some form of protective immununity is powerful enough to prevent infection. This type of protective inmmity of skin should be constantly expressed (as in mucous membranes) or up-regulated following wounding and invasion of pathogens. In addition, the antimicrobial system should be site-specific, as pathogens demonstrate site specificity. As an example Staphylococcus aureus has been located in the toe webs, nasal cavity and perineum as commensals (5, 6). Others, including Pseudomonas aeruginosa prefer perianal regions (3).
Thirdly skin should be able to recognize a potential pathogenic agent using recognition facilities (receptors) whereby pathogens (and non-pathogens of the resident flora) are noted as potential danger.
Lessons from psoriasis
Psoriasis is a hyperproliferative skin disorder with massive scale production as characteristic feature (7). Colonization of skin with pathogenic bacteria is common in psoriatic, but also in eczematous skin. In psoriatic and eczematous skin S.aureus, has shown to be the most common pathogen. Thus impetigo, folliculitis and furuncles caused by S. aureus occur frequently in atopic patients.
Nevertheless, in lesional skin of patients with psoriasis containing abundant numbers of bacteria, notably again S.aureus (8), complications caused by bacterial infections are rare. In fact, using a patient-oriented databank at the Kiel Department of Dermatology, we could show that psoriasis patients, even with severe and wide-spread involvement, demonstrate any type of pyoderma only half as often as do patients with other inflammatory skin diseases. This indicates that in dealing with microbial invasion some fundamental functional difference exists between the skin in psoriasis and atopic eczema.
In theory, lack of susceptibility for infections in psoriasis – in contrast to atopic skin – could be linked with a Th 1-regulated response pattern, which is known to be expressed in psoriasis (9). Th 1-dominated immune responses predominantly activate a phagocyte-dependent type of inflammation directed against infections sustained by intracellular bacteria and certain viruses (10). Increased production of IFNγ, TNFα, IL-1 and IL-8 is characteristic for psoriatic skin. Interleukin 8 proved to be one of the most potent chemo-attractants for neutrophils, and significant amounts are produced by human keratinocytes (11). However, Th 1-directed immune responses or phagocytosis by neutrophils takes place within the inflammatory sites of living tissue, not at the surface of the skin where pathogens are located.
In psoriasis, there is no evidence that microbial agents are able to transmigrate into the subcorneal living layers of the epidermis. In atopic skin, the situation is different in that depletion of the stratum corneum by scratching and scratching-induced microwounds (fissures) are common. Loss of the barrier enables microbes to enter, and favors bacterial growth with exsudation of serum. Furthermore, the predominating Th 2 responses of atopic skin has suppressive effects on Th 1-regulated antimicrobial defense so that susceptibility to infections caused by bacteria (and viruses) can be understood.
The question remains by what mechanism(s) infectious agents are kept under control in psoriasis and also why in ordinary wounds keratinocytes are able to sustain in the presence of pathogens. These questions are of fundamental importance for any multicellular organism, and have first been raised in lower animals and plants (reviewed in 12).
Sentinel role of keratinnocytes
In lower organisms, which, as commonly accepted, do not contain any adaptive immune system, the epithelium represents the major defense organ. It is believed that their epithelia constitutively produce a number of antimicrobial compounds, including gene-encoded antimicrobial peptides, to control the normal microflora. In addition, they mount a rather pathogen-specific defense reaction by inducing the synthesis of more or less pathogen-specific antimicrobial peptides, when they come into contact with pathogens. This mechanism is not well-understood, but implicates that plants and invertebrates must be able to recognize ‘dangerous’ microorganisms and to discriminate between these and commensales.
But where are the differences between virulent pathogens and non-pathogens? The discovery of human homologues to Drosophila Toll-receptors, so-called ‘Toll-like receptors, TLRs’, which are believed to mediate innate immune responses via recognition of bacterial products such as Gram-negative bacteria-derived LPS (TLR-4), Gram-positive bacteria-derived lipopeptides (TLR-2), bacterial flagellin (TLR-5) and bacterial DNA (TLR-9), invites one to speculate on the role of these receptors to signal keratinocytes the presence of microorganisms.
Indeed, there are hints that these receptors, which are all found to be expressed on macrophages, are also expressed – at least in part – on keratinocytes (13). Whether these mediate the epithelial induction of antimicrobial peptides such as human β-Defensin-2 (hBD-2) is not yet clear. hBD-2 is the first human inducible epithelial peptide-antibiotic recently discovered in our laboratory, which is active predominantly against Gram-negative bacteria and yeasts, but not S.aureus (14). We have seen that tracheal epithelial cells (15) as well a epidermal keratinocytes (unpublished results) respond towards LPS by inducing hBD-2. The concentration of more than 10μg/ml necessary to induce hBD-2, however, seems to be too high to be relevant in vivo. But what could be the relevant stimulus instead of LPS?
Recently we saw that different strains of microorganisms had a different capacity to induce hBD-2. Remarkably, mucoid strains of P. aeruginosa nearly always induced hBD-2, whereas only rarely non-mucoid P.aeruginosa showed this behavior. Because mucoid strains of P. aeruginosa always developed a biofilm, it is possible that human epidermal keratinocytes recognize molecules only produced by this virulent form and thus molecules involved in the biofilm formation, and mount a defense, but do not attack the non-virulent planctonic strains.
It is likely that virulence factors that are connected with the formation of biofilms, represent pathogen-associated molecules, which tell the keratinocyte the presence of dangerous, biofilm-forming Gram-negative bacteria. In other words, keratinocytes mount a rather Gram-negative bacteria-selective defense answer when they recognize P. aeruginosa. This mechanism would be very similar to innate defense reactions seen in Drosophila, where the contact of these insects with fungi induces the fungus-specific antifungal peptide drosomycin (16).
If this hypothesis is true one would expect that also human epidermal keratinocytes should be able to induce the production of antimicrobial peptides directed against different microorganisms, i.e. Gram-negative bacteria like P. aeruginos or Gram-positive bacteria such as S.aureus. We recently could prove this hypothesis, when we discovered hBD-2, which is an epithelial inducible and rather Gram-negative selective antimicrobial peptide (14) and very recently hBD-3, a potent antimicrobial peptide, that efficiently kills Gram-positive bacteria and which is also inducible in keratinocytes (17).
With these findings in mind, one may speculate that keratinocytes have the capacity to produce a number of antimicrobial peptides and proteins, which have been optimized by evolution in order to kill different microorganism targets. Preliminary data of our investigations clearly show that this is indeed true.
The observation that (unlike macrophages) keratinocytes apparently attack only biofilm-forming microorganisms further leads us to speculate that keratinocytes could also interfere with the formation of biofilms, i.e. by disturbing the ‘quorum-sensing’ of bacteria (which precedes the formation of biofilms) and/or its adhesion to the substratum, that is necessary for the start of the biofilm-formation with subsequent colonization. The finding that biofilm-forming microorganisms also induce primary cytokines such as TNF-α and IL-1β or IL-8 in keratinocytes (our unpublished results), which is seen to a much lesser extent in single cell suspensions of planctonic grown bacteria, may be interpreted as an induction of recruitment of neutrophils for help to defend infection.
The question remains why a normal human flora can colonize on the epidermis. In any multicellular organism, epithelial linings show different functional characteristics with variable microbial colonization. It is abundant in the intestines and external mucous membranes, whereas in mammalian epidermis, due to the stratum corneum, keratinocytes live in a sterile microenvironment. However, the stratum corneum provides only limited protection against mechanical trauma (wounding) and, as discussed above, microwounds, fissures, minor and often unnoticed epithelial lesions are common.
Thus, being located at the outermost body surface keratinocytes play a fundamental role not only in wound closure but also in recognition of danger from infectious agents. As shown in gut epithelia and by recent work in human keratinocytes these cells respond by expressing two separate defense strategies: 1. production of peptide antibiotics and other antimicrocial substances and 2. secretion of cytokines and chemokines. Both, antimicrobials and signal substances (cytokines, chemokines) are generated within a short time and in significant amounts following activation.
Innate immunity in previous years was thought to be provided by ‘classical’ members of this system, e.g. macrophages, neutrophils and NK cells (18, 19). As demonstrated, keratinocytes need now to be included. Due to their position of infinite exposure to the environment and in view of their possession of an effective armentarium for killing and signaling, they serve as sentinels at one of the major sites of microbial entry into the mammalian body.
When considering these epithelial functional activities it may occur as a puzzle that in keratinocytes one fundamental function of innate defense seems to be lacking, i.e. phagocytosis. Although keratinocytes may occasionally be noted to phagocytose (i.e. Candida albicans (21)) this remains an exception. Intracellular uptake and killing is restricted to professional phagocytes, neutrophils and macrophages. It is therefore not surprising to find neutrophils in close proximity to the epidermis whenever wounding (and subsequent microbial invasion) occur. At such sites secretion of interleukin 8 by keratinocytes provides potent signals for leukocyte attraction (22). In addition to wounding, intraepidermal neutrophils are commonly seen in any type of spongiform as well as psoriasiforme dermatitis. In fact, neutrophils appear to be the most common cell type invading inflamed epidermis, their number outweighing lymphocytes by far. Thus beneath the stratum corneum, whenever this delicate barrier is broken down, epidermal keratinocytes are able to recognize danger from microbial invasion, start secreting antimicrobial peptides and, finally, open the door for the professional phagocytes, e.g. neutrophils.
In summary, epidermal protection is two-armed (keratinocytes: signaling, killing and repair, neutrophils: migration, phagocytosis and killing). This provides the basis for functional synergism of these two cell types, keratinocytes and neutrophils (Fig. 1). Understanding the sentinel function of keratinocytes in innate immunity of skin enables us to envision epithelial-phagocyte synergisms as a key feature guaranteeing organisational integrity and survival in a hostile microbial world. This system of cutaneous immune protection developed long before the adaptive immune system, dermatologist’s beloved playground, made its appearance during evolution.
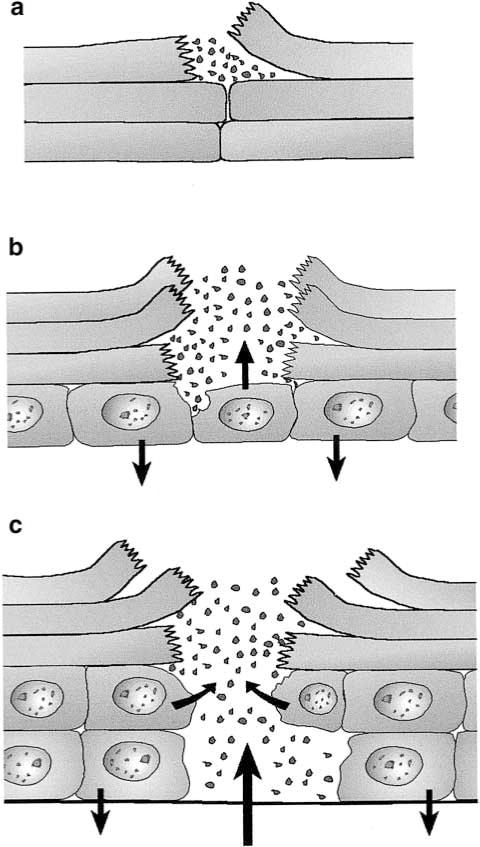
Different degrees of epidermal wounding elicit variable response patterns. (a) Intracorneal break up may activate antimicrobials from intra-and intercellular pools. This has not been studied so far, although the events are common. (b) Epidermal wound (fissure) extending into non-keratinized cell layer. This causes secretion of antimicrobials into the defect. Proinflammatory cytokines are released by neighboring keratinocytes. (c) Deep epidermal wound extending onto the basemen membrane. In addition to the aforementioned responses, chemokines released into the dermis will now attract neutrophils with the ability to phagocytose invading pathogens.
References
1.Noble WC. Microbiology of human skin. London: Lloyd-Luke; 1981. [Google Scholar]
2.Noble WC, editor. The Skin Microflora and Microbial Skin Disease. Cambridge: Cambridge University Press; 1993. [Google Scholar]
3.Kligman AM. The bacteriology of normal skin. In: Maibach HI, Hildrich-Smith G, editors. Skin Bacteria: Their Role in Infection. New York: McGraw-Hill; 1965. pp. 13–31. [Google Scholar]
4.Leyden JJ, et al. Invest Dermatol. 1988;88:65–72. [Google Scholar]
5.Noble WC. Staphylococci as pathogens. In: Noble WC, editor. The Skin Microflora and Microbial Skin Disease. Cambridge: Cambridge University Press; 1993. [Google Scholar]
11.Schröder JM, Christophers E. The biology of NAP-1/IL-8, a neutrophil-activating cytokine. In: Coffey RG, editor. Granulocyte Responses to Cytokines. Basic and clinical research. M Dekker Inc; 1992. pp. 387–416. [PubMed] [Google Scholar]
The skin can think: a modest proposal and its critique by NIH
Specific Aim
The hypothesis that skin is a thinking organ will be critically tested using standard and advanced biological techniques. Establishing the skin as a thinking organ will allow for new diagnostic and therapeutic techniques for many skin diseases as well as general ailments of the body and the psyche in children and adults.
Background and Preliminary Data
The skin has the embryological derivation and the structural basis required for it to be a thinking organ. The functions of the skin will be interpreted in terms of those exhibiting memory, intelligence and ultimately consciousness, all of which will be required for considering higher level thinking. Finally, an experimentally testable approach for this hypothesis will be outlined. The skin is considered to include the epidermis, dermis and subcutaneous tissues, skin appendages, and its vascular and neurological structures.
The epidermis and the neural crest, are derived from the same embryonic ectoderm that forms the brain and peripheral nervous system. The number of neurocutanteous syndromes reinforces the skin nervous system relationships.
The epidermis and the rest of the skin are richly innervated with both sensory and motor nerves, general and specialized receptors, for various sensations, and even the immune system of the skin has prominent neurological connections. There are formal efferent and afferent nerve pathways throughout the skin which are repetitive and can be thought of as the physical strata of a Turing Machine (1). This Turing Machine can read and process a great deal of information about the body’s internal environment as well as of the external environment. Other morphological and functional networks involve the skin including the endocrine system, e.g. via Vitamin D production in the epidermis, its modification by two different hydroxylations; a 25-hydroxylation in the liver, and a 1-hydroxylation in the kidney, forming 1,25-dihydroxy Vitamin D which then interacts with the epithelium of the hair follicle and the keratinocytes.
The immune system through its antigen detecting and processing cell in the epidermis, the Langerhans cell, is the afferent arm of the immune system. There is involvement of keratinocytes in antigen presentation producing tolerance; UVB modifies the entire immune process, and ultimately the efferent portions of the immune system either through cells directly or through their lymphokines and chemokines, thus forming an integrated neural-like network in the skin.
With this richness of biological and physiological structure and function, why hasn’t the hypothesis of the skin thinking been addressed in the past? Exhaustive searching through Medline has failed to reveal any published discussions when searching on skin and thinking, skin consciousness, or skin and thought. The phrase ‘think skin’ in Goggle gave many pornographic sites but did not address the issues being considered in this manuscript.
Two issues must be considered: the role of the skin in the body’s economy and in the body’s internal political system. I have previously discussed the challenge of skin being the body’s largest organ (2); physiological systems will counter the natural tendency of organs to use their anatomical positions to take over resources from the rest of the body; this is the internal politics of bodily functioning. This state requires large numbers of regulatory systems to keep the skin in check and to keep it in its place. These essentially ‘negative’ influences on the physiology of the skin keep the skin from expressing the higher functions of memory, consciousness and expression which are usually considered part of thinking.
The skin can obviously communicate in an expressive fashion with vascular dilation, patterning and thickening and thinning of its various strata, pigmentary patterns resembling a semaphore system and pheromones to stimulate the vomeronasal organ. The skin can ripple through the contraction of its arrector pili muscles and produce low frequency sounds that are primitive forms of speech.
Is there evidence that the skin is conscious and that the skin can know itself? Consciousness is the holy grail for those interested in thought, the mind and epistemology (3). Since consciousness is difficult to define short of a tome, I will say that there is no evidence either way on the conscious nature of the skin. Clinicians often see the cleverness of the skin in changing its antigens and metabolic pathways to escape antineoplastic therapy, the stubbornness exhibited by skin disease such as psoriasis, but there has not been the detailed analysis and testing required for the demonstration of consciousness. I often hear a gentle humming from my skin late at night as it contemplates the body over which it resides and rules, a sign of a sentient organ and probably a form of consciousness.
Experimental Plan
The essential element of the experimental approach that is proposed is to leave the skin relatively intact but to free it from the inhibitory systems of the general body. Using the Hannibal Lecter (4) full skin dermatome the entire skin will be removed from an animal severing all connections of the vascular, neurological and immune systems between the skin and the rest of the body. The skin will be removed from the animal under a protocol approved by the institutional committee on animal resources. These experiments can not be performed on tissue culture preparations of skin or skin equivalents that are excellent for many physiological studies but have not been optimized for studies of the skin’s thinking ability. The inner surface of the skin will be perfused with an optimum metabolite preparation whose composition will be determined in preliminary experiments.
After stabilization of the preparation, the skin will be tested using standard protocols for neurophysiological studies such as learning, memory, recall and the integration of information. Information will be presented in the form of vibratory (20–20 000 cycles/s), temperature (4°–40°) and electromagnetic irradiation from 250–1150nm. Appropriate controls will be included in all experiments. Responses from the skin include changes in galvinic skin response, pigment cell distribution, epidermal barrier function and phermone production using standard techniques.
Preliminary data will be analyzed by paired t-tests and Wilcoxon non-parametric techniques, when appropriate, and dose–response-curves. Specimens for males, females, children and pregnant femalesofthe speciestobe examined. Since the hypothesis being tested considers the potential for skin to be a thinking organ the research will be presented to a broad-based bioethical review board as well as the Institutional Committee on Animal Resources.
If thinking can be demonstrated in the skin this will lead to new, safe and effective therapies and approaches for skin disorders that have plagued mankind for generations.
NIH Summary Review Statement
The principle investigator has broad general knowledge of the skin but has never studied thinking before. There is no preliminary data presented to support the novel approach that is presented. The experiments proposed are technically feasible. The budget is within guidelines.
The thinking skin has broad national important since it will become necessary to win the hearts, minds and now the skins of those the government wishes to influence domestically and around the world. Failure in American policy in the past may have been due to failure to recognize that the skin thinks. For these reasons the Departments of Defense and State are the appropriate organs of the government which should be reviewing this proposal for funding. The decision not to fund this proposal by NIH should not be taken as a statement against the scientific rational or approach the investigator proposed.
References
1.Berlinksi D. The Imaginary Machine The Advent of the Algorithm. New York: Harcourt, Inc; 2000. [Google Scholar]
The skin as an organ is performing a whole lot of functions, each of which being truly indispensable for the maintenance of life. Amongst them is, though not considered first, its function as a membrane. Therefore, I shall focus here on exploring in more detail the as yet underappreciated function of the skin as a permeable biologic membrane.
‘Intact healthy skin is a remarkably good barrier to the mass transport of topically applied substances, yet it allows some permeation of almost every substance’ (1). This is due to the particular structure of the skin, built up into a composite membrane by several layers of different tissues: the stratum corneum (10mm), the viable epidermis (100mm), and the papillary layer of the dermis (100–200mm). The actual permeability barrier resides in the stratum corneum. Since this horny layer is composed of fully keratinized epidermal cells that are metabolically inactive, skin permeation and percutaneous absorption appear to be controlled by the passive diffusion of substances through this tissue rather than by filtration or pinocytosis. Diffusion through intact skin is in no way dependent on cellular metabolic activity (2).
Diffusion occurs as a result of the tendency of substances within a single phase to equalize their concentrations (3). Molecules absorbed at the surface of the stratum corneum diffuse through it, subsequently do so more rapidly through the viable epidermis and the papillary dermis, and lastly reach the capillary plexus therewith entering the circulating blood. The rate-limiting process is diffusion through the stratum corneum. Some labile substances might diffuse through the stratum corneum unaltered, only to be oxidized, hydrolysed or metabolized by the viable dermis prior to the absorption into sytemic circulation (1).
A host of substances has been shown to be absorbed percutaneously (4). Included are non-toxic substances such as water and electrolytes as well as toxic compounds such as insecticides and ‘war gases’. Also topically applied medications such as phenol, boric acid, elemental sulfur, resorcin, tar, and mercury fall into this group. Because of their toxicity they have nowadays been banned from external therapy.
Permeation through the skin is, of course, no one-way road. The permeability of skin to water, for example, can be measured in vivo from a non-sweating region of the forearm as transepidermal water loss (5), or in vitro from excised skin supported as a diaphragm over a reservoir of water (6). The former process is commonly called ‘insensible perspiration’, the latter is simple permeation. Remarkably, in both cases the measured amounts of water are closely similar. When the skin is diseased or damaged, the transepidermal water loss can increase by several orders of magnitude. The property of water sealing is an important physiological function of the skin and resides solely in the stratum corneum.
Of particular dermatological interest is the penetration of hapten molecules. Owing to their low molecular weight and depending on their lipid or water solubility they can more or less easily enter the skin. There are literally thousands of different substances that by this way can produce delayed hypersensitivity reactions. Following penetration they are known to couple to endogenous proteins, thereby forming an antigen which can then elicit an immunological response.
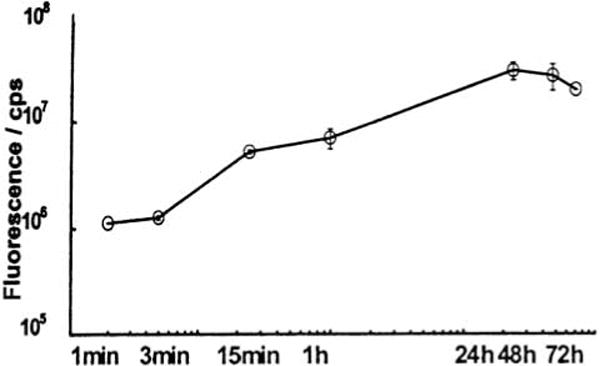
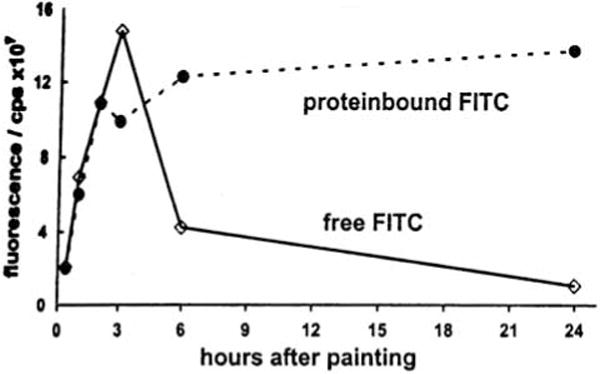
We have studied the percutaneous absorption of lipophilic haptensby employing the fluorescent contact sensitizer fluorescein-isothiocyanate (FITC) (7). It was painted once only in a sensitizing dosage (500μg) onto one ear of normal BALB/c mice. Already 1min later, blood drawn from the axillary vein exhibited significant fluorescence emission at 517nm, when examined by means of fluorescence spectrophotometry (Fig. 1). FITC fluorescence in the circulating blood could be traced up to 72h (Fig. 2). In order to discriminate free hapten from protein bound molecules, blood samples were subjected to Sephadex G-25 column chromatography. It was found that free hapten molecules entered the circulation for more than 24h, although decreasingly, but while circulating they gradually combined with plasma proteins (Fig. 3). Meanwhile, however, free hapten molecules were dispersed throughout the body. (The findings presented have immunological consequences which in the context of this essay are not considered further).
Fluorescence spectra of blood plasma samples after topical application of fluorescein-isothiocyanate (FITC) (upper curve) or solvent only (lower curve). The excitation wavelength was 488nm (band pass 2nm), the integration time was 1s. Data were taken at the emission maximum of FITC at about 517nm.
Samples of mouse plasma were subjected to Sephadex G-25 column chromatography. Fluorescein-isothiocyanate (FITC) molecules bound to plasma proteins were separated from free FITC molecules. Curves illustrate that free hapten combines increasingly with plasma proteins.
These findings indicate that normal mouse skin acts as a permeable biologic membrane. The lipophilic hapten FITC does indeed penetrate the permeability barrier very quickly, but stays within the membrane for some hours. During this time coupling to skin proteins is supposed to occur. However, because coupling is a time-dependent process, molecules still uncoupled leave the membrane and enter the circulating blood as early as 1min after painting and for as long as 1 day. This suggests that the permeating substance, after entry, accumulates in the membrane thus serving as a reservoir which is then gradually discharged into systemic circulation.
References
1.Scheuplein R. The skin as a barrier. In: Jarrett A, editor. The Physiology and Pathophysiology of the Skin. Vol. 5. London-New York-San Francisco: Academic Press; 1978. pp. 1669–1692. [Google Scholar]
3.Scheuplein R. Skin permeation. In: Jarrett A, editor. The Physiology and Pathophysiology of the Skin. Vol. 5. London-NewYork-San Francisco: Academic Press; 1978. pp. 1693–1730. [Google Scholar]
4.Malkinson FD, Rothman S. Percutaneous Absorption. In: Marchionini A, Spier HW, editors. Erg Werk Bd. 1/3. 1963. pp. 90–156. [Google Scholar]
The phylum Vertebrata emerged 550million years “Before Present” (BP): initially, all classes were exclusively aquatic, but for the last 380million years, four additional classes have lived on land (1). Throughout the entire period, fossil and comparative anatomical data reveal diversity of skin form but its functions as an interface between internal and external environments have remained unchanged. This essay reviews general integumentary form/function relationships, compares and contrasts ways in which the skin of aquatic and terrestrial vertebrates reflects physical differences of their worlds and concludes with an evolutionary biologist’s perspective on the unique features of human skin and their relevance to the choice of ‘animal models’ in dermatological research.
Basic morphology
Unlike other animals, vertebrates have a multilayered epidermis (2). In fish its cells are primarily mucogenic but in tetrapods keratins replace mucins as primary structural proteins. The subjacent dermis comprises collagenous connective tissues that house blood vessels, nerves, pigment cells and sometimes calcified elements.
Over any or all body regions, the surface may be folded into scales that overlap to varying degrees, or it may comprise appendages defined as: ‘localized centers of specialized epidermal and/or dermal cell proliferation and differentiation [surrounded by] an otherwise generalized integument’ (3). The fundamental distinction between scaled and appendage-bearing skin is critical to an understanding of the evolution of its functions.
Primary and secondary functions
Due to somatic muscle action, a vertebrate’s body, continuously changes shape. Whether we speak of a fish’s swimming movements or human thoracic cage expansion and contraction during respiration, a major requirement for skin is that any resultant deformations are congruent with sustained integrity of those tissues responsible for primary barrier functions (3). The latter include 1. prevention of pathogen ingress; 2. maintenance of physiological homeostasis and 3. mechanical protection. Because epidermis contacts the external environment, viability of its cells is continuously at risk and direct damage is always possible. Therefore all vertebrates show a genetically based pattern of epidermal cell turnover and a capacity for wound repair because, without such, function would be compromised and death would result. The skin also performs many secondary functions that always involve appendages formed in the embryo. Although they show cellular turnover, when damaged by trauma they are rarely replaced (4), their place being taken by tissues that restore only barrier functions. Such damage is rarely life threatening, although we humans may find scars aesthetically undesirable. These generalizations permit considering how skin form reflects habitat.
Piscine vertebrates and aquatic life
Aquatic life has many advantages. Continuous ‘washing’ of the body surface during swimming decreases the probability of pathogen ingress aided by antibiotic properties of mucins whose additional role is reduction of drag (5). Relative to biomass, the environment’s volume is so great that the few problems of physical abrasion are met by very rapid epithelial proliferation (6) – only in high population densities of poorly maintained aquaria or pisciculture does physical contact between individuals cause excessive skin trauma later exacerbated by fungal infections. Water buoyancy permits many fish to bear a heavily armored skin. Water provides an inexhaustible supply of oxygen and percutaneous gas exchange is important in fish (7): because water loss is never a problem, diffusion is unimpeded by barrier lipids. Aquatic environments are thermally, constant so that piscine homeostasis is unaffected by diurnal temperature fluctuations.
Tetrapod vertebrates and terrestrial life
Land living is difficult for animals. Many – several phyla of ‘worms’, slugs, a few fish, frogs, salamanders, etc., ‘live on, or take excursions onto, land’ but are confined to very humid, even wet, microhabitats. Among 35 known phyla, only arthropods (mainly insects) and amniote vertebrates (reptiles, birds and mammals) solved the problem of allowing shape change within a skin that also resists water loss and physical abrasion. Arthropods sacrifice mobility: much of their body surface is inflexible. Amniotes are covered by a mechanically flexible, lipid/protein complex. Products of lipogenic lamellar bodies (amniote innovations (8)) deposited in intra and/or extracellular domains of a pluristratified, α-keratogenic epidermis form a barrier that protects the internal milieu from air’s dehydrating effects. However, the delicate barrier tissues whose relative dimensions in a lizard, a chicken, a mouse or a cow do not track the sizes of the organisms, would be susceptible to environmental abrasion were they not protected by overlying β-keratogenic tissues (lizard scale surfaces or avian feathers) or hair (9).
Most biology texts address only insulatory properties of mammalian pelage and avian plumage, aerodynamic aspects of feathers and some comment on coloration. In comparative anatomy texts, accompanying figures are often legendary in the pejorative sense! Thus, the extraordinary, diversity of functional roles of epidermal systems in toto is neglected. Some examples of completely unexplored issues warrant mention. Is it that hairs and feathers provide such excellent protection for both barrier tissues and the entire organism that, in contrast to all other vertebrates, few mammals (armadillos) and no birds (not even defenseless, flightless species) possess dermal sclerifications? Why is it that while many mammals, large (elephants, rhinoceri, even Homo sapiens) and small (some rodents) have reduced or lost the pelage, no bird lacks a plumage? Denizens of cold climes and/or aquatic habitats (among mammals – mink, seals, beaver, among birds – ducks, geese) have evolved secondary or tertiary hairs or feathers that enhance their skin’s insulatory properties, but their development remains undocumented. Lack of a systematic study of gland distribution and morphology precludes explaining the evolution of sweating as a component of mammalian thermoregulation.
The practical importance of an evolutionary perspective
Of all the body’s organ systems our knowledge of form/function relations in skin is arguably the poorest. This assertion has important implications for biomedical pedagogy and research.
Students are accustomed to a deluge of data concerning physiological control mechanisms in other systems. They find it difficult to accept that acute dehydration – a phenomenon they associate with our proclivity for excessive physical activity on hot summer days – is 1. an inevitable possibility in the face of the remorseless, dehydrating effects of air, 2. completely uncontrollable and 3. a major, daily problem for a large percentage of earth’s human population!
An evolutionary perspective is illuminating with respect to clinical dermatology, and research programs seeking remedies. In no other taxon does skin structure in one species differ as dramatically from that of closely related species as does that of Homo sapiens compared to that of the great apes, let alone primates in general. In fact, diversity of skin phenotype within our species has no equal and this presents a plethora of fascinating evolutionary problems. Some deem the questions inherently unanswerable because we do not, and never will, have direct fossil evidence of the origin, distribution and environmental context of soft tissue subtleties. Ignoring the questions – a permissible option for academic biologists or physical anthropologists – is a luxury, not available to clinical dermatologists because of the association between skin ‘differences’ between subspecies (colloquially = ‘races’) of Homo sapiens and differential proclivities to integumentary dysfunctions. It would seem prudent to seek research models in which Nature makes available varying phenotypes that might emulate human skin form/function relations. Apart from the fact that granting agencies are loath to consider proposals involving ‘exotic species’ i.e. anything other than E.coli, laboratory rodents and rabbits, and the occasional small carnivore (dogs or cats), basic biology tells us the choice is verv limited.
The body of data concerning non-human primates does not solve the problem – rather it serves to emphasize it by, showing how their skin is, in so many important respects, unlike that of humans. For example, most species inhabit tropical or subtropical climes. However, many medium to large-sized species cavort cheerfully in outdoor cages in temperate or even subtemperate zoos because their pelage protects them from potentially lethal heat loss. To find mammals whose body heat is conserved primarily by cutaneous fat deposits, one has to look to porpoises and whales – unfortunately, they have lived in water for over 50million years and keratinocyte cytodifferentiation in them is totally different from that in terrestrial mammals. Two problems are thus exemplified. First, if we accept the premise that skin form must accommodate multiple, interrelated functions, we see that any species expresses a unique combinatorial pattern of ‘compromises’. Second, such patterns must be viewed within the context of a species’ natural environment. How could this affect a research program?
A program of study of epidermal lipids might involve, in part, measurement of rates of evaporative water loss through skin. Such yields different absolute values for different species. However, this does not permit the conclusion that ‘species X is better at conserving body water than species Y’. The fact that both species exist establishes that both have evolved barrier functions that fulfill at least the minimal requirements to sustain life in Nature. Suppose the researcher then determines that both quantity and quality of lipid molecules varies between and among those species. Establishing the fact that one or more ‘lipid profile(s)’ is congruent with barrier function(s) might not necessarily imply a direct causal relationship. Many species employ semiochemical signals of epidermal origin in their behavioral repertoires. Without data concerning e.g. the biochemistry of skin bacteria, behavioral tests, etc., etc., those profiles cannot be fully understood. If one were comparing epidermal lipids in related species of small rodents one should be aware of yet another problem: burrows in different soils, with different particle sizes may, necessitate different grooming behaviors.
Space limitations preclude listing the many unanswered questions in comparative skin biology, whose potential clinical significance must be emphasized. Recent advances in molecular genetics facilitate our understanding of evolutionary issues germane to skin (8) and the same data are applicable to clinical dermatology (10). The time has come to extend our choice of ‘animal models’ beyond laboratory rodents, species in which skin form and function is possibly as unique as our own.
In summary, although vertebrate skin has shown great morphological diversity over 550million years, its functions as an interface between internal and external environments have largely been unchanged although the specifics of the challenges presented by aquatic vs. terrestrial habitats differ. Greater emphasis must be placed on consideration of the multiple interrelated roles performed by scales and epidermal appendages.
References
1.Pough FH, et al. Vertebrate Life. 4. Upper Saddle River, NJ: Prentice Hall; 1996. [Google Scholar]
2.Bereiter-Hahn J, et al., editors. Biology of the Integument. Vol. 2. Berlin: Springer-Verlag; 1986. [Google Scholar]
3.Maderson PFA. Am Zool. 1972;12:159–171. [Google Scholar]
5.Whitear M. The skin of fishes including cylostomes. In: Bereiter-Hahn J, et al., editors. Biology of the Integument. Vol. 2. Berlin: Springer-Verlag; 1986. [Google Scholar]
6.Quilhac A, Sire J-Y. J Exp Zool. 1998;281:305–327. [Google Scholar]
The first real question should be what is mammalian skin, then what does it do? What is (or is not) mammalian skin? The classic essay by Lowell Goldsmith (1) defining the skin as the largest of the intermediate sized organs assumes, as most dermatology, anatomy, and pathology books state, that the skin and hair is an entity unto itself. Just as hair is an integral part of mammalian skin so too is skin an integrated part of the body. Since cutaneous nerves connect to the spinal cord and/or brain (central nervous system, CNS), the skin is an extension of the brain just as cutaneous blood vessels are an integral part of the circulatory system. The skin, hair and nails therefore represent the entire animal on the outside. By inference the gut lumen is the epithelial interface or ‘skin’ on the inside.
What does mammalian skin ‘do’? The appropriate but limited metaphor of cutting tree limbs, losing leaves and debarking (‘skinning’) a tree may be invoked as to what mammalian skin does. If you remove the limbs and debark or ‘skin’ a tree, you expose the viable tissue that covers the dead structural tissue that supports the tree. By analogy mammalian skin therefore is the outer layer that covers and protects the viable, ‘critical’, and commonly considered more important organs and tissues below. However, except for Sampson’s loss of strength after getting his hair cut by Delilah, loss of hair in humans does not cause death or serious problems but does cause serious problems in other mammals. If you ‘skin’ a bear, you better hope the bear does not ‘skin’ you!
Is mammalian skin an effective somatosensory organ? Hair is an extension of the somatosensory system of the skin in that tylotrich follicles on the body of many mammals have specifically evolved for helping the mammal define where it is in its environment. Most mammals, the notable exception being Homo sapiens, have vibrissae, highly specialized sensory hairs found at key locations, around the head and feet. These specialized hair follicles and fibers combine blood sinuses and large nerves (systematic integration) with long, thick hairs that make eyes obsolete. In Silicon Valley terms, the skin represents a rapid response, solid state computer to integrate input from multiple sources (ionizing radiation, reactive chemicals, etc.) and transmits these signals to the central storage unit, the CNS. This CNS integrative function is performed while conducting its other functions such as holding the animal together, providing protection from inside and out, and manufacturing proteins and vitamins, such as Vitamin D, needed by other organs for survival. From the veterinarian’s perspective (John P. Sundberg), hair therefore is a critical part in all mammals.
The physician’s interpretation (Lloyd E. King) is just the opposite. Evolution may have rewarded a progressive loss of hair in Homo sapiens so that the skin is a more responsive computer in humans (and favored a more extensive development of eccrine glands, see below). These CNS mutations led to humans making clothes from other animal’s skins, fire, electricity, houses, cars and other contraptions to protect humans from the environment and allowed them to be more mobile. More surface area magnifies the ability to respond to minimal signals in the short term but carries the penalty of more vulnerability and decreased wound healing ability as compared to haired animals. Long-term responses to the environment forces the CNS to become more flexible and deal with more data storage capacity, thereby increasing memory and the ability to deal with complex, often competing signals (therefore requiring highly developed eyes since the hair as a sensory input is all but gone).
Why do humans lack more hair than other mammals? What functions do hair and hair follicles serve in mammalian skin? Darwin’s revolutionary idea of survival of the fittest and evolution (2) supports the importance of hair for survival of most mammals. Living in harsh environments requires protection from the elements (heat, cold, sun, rain, bugs, etc.). Some mammals adapt by building homes in the ground, trees, etc. but are still exposed. So why, did Homo sapiens, a species generally regarded as one of the weakest mammals, progressively lose a major defense mechanism, a ‘fur coat’ or pelage? Did evolution favor the development of the CNS, temperature regulation, and/or other functions? Maybe it was simply an adaptation against ectoparasitism (3).
In many non-human mammals the hair follicles cycle in seasonal shedding patterns. This physiological shedding of hair somewhat compensated for marked seasonal temperature differences. However, human hair cycles are irregular or random and bear no known relationship to seasonal temperature variations. Perhaps the answer may relate to known physiological functions of hair follicles that produce sebaceous gland secretions to coat the hair shafts in all mammals. Furthermore, at special sites apocrine glands are attached to the hair follicles.
Why did eccrine glands develop independently of hair follicles and apocrine glands? In humans there is a much more extensive distribution of eccrine sweat glands than in other mammals. A major purpose of the eccrine sweat glands is heat dissipation but another critical function is to provide moisture to help distribute the lipid contents of the sebaceous glands to provide the necessary components of the epidermal water barrier. Even in sites with predominantly vellus hairs, the sebaceous lipids move outward along the hair shaft to reach the epidermal surface. Lack of moisture, as normally produced by eccrine sweat glands, leads to abnormal epidermal differentiation, cornification, and heat intolerance.
What then is the biological advantage of being a ‘Naked Ape’ (4)? Do the increased number of eccrine glands play a major role in detoxifying internal or external noxious agents? Do they only serve as the major avenue of heat dissipation that allows humans to adapt more readily to differing environmental situations, with or without fur as optional clothing? Perhaps less muscle strength leads to more efficient and adaptable temperature control and permits a high level of persistent muscular activity compensated for by sweating rather than panting. Is hibernation of furred animals linked to the predictable demands to maintain brute muscle strength at times when food sources are difficult to locate? Humans may gain biological advantage by not having such a high metabolic demand due to less bulky muscles and their ability to more rapidly adapt to changing climates.
People often look at the exceptions in the biology of human skin as compared to other mammals. In general the skin functions in a predictable manner in most mammals. However, from a phylogenetic perspective, most mammals have a defined hair cycle consisting of a relatively short growth phase (anagen) followed by long periods of resting (telogen). Most follicles remain in telogen, the result being that the hair grows to a certain length and then stops. By contrast, human hair follicles remain in anagen, the growing phase, for very long periods. Could this be to make up for the general sparcity of hair over most of their body? Why is this only the scalp hair and not hairs covering the rest of the body except for sex hormone determined sites? Progressive hair growth is rarely found in mammals and often limited to those created by selective breeding for this purpose, such as poodle dogs and Merino sheep. Perhaps regulation of follicular cycling by the Vitamin D receptor may determine the location of dense hair growth in sex hormone dependent sites (5, 6). The toxic effects of UV light may be limited by hair growth, thereby favoring those species that have prolonged anagen or this feature limited to exposed sites such as the scalp in humans.
Skin, hair, and nails serve as courtship and reproductive signals. In mammals hair has evolved for many specialized purposes including courtship. Humans are the exception, or are they? Skin is the primary non-reproductive organ involved in courtship. As a species, humans spend an inordinate amount of money and effort at maintaining the quality of their skin and its appendages, hair and nails. Sexual dichotomy defines hair cycle length which ultimately translates into socially acceptable short hair for males and long hair for females. Sub-populations that have tried to switch this feature are either fraught with failure, as has been the case for both the authors, or met with social indignation. The perception is that hair length (long in men, short in women) is not (sexually) attractive but socially accepted length represents biological function of the organ. Therefore at a gross level, social and sexual signals provided by the skin are direct reflections of normal and optimal function, not simply cosmetic issues.
Fanciers have kept interesting animals in captivity for centuries, many of which were common species with abnormal physical attributes. The most easily observed differences (phenotypic deviants in today’s phenomic, genomic, proteonomicera) are skin, hair, and nail changes. These mutant animals have finally been recognized as important natural experiments. For example, spontaneous mutations such as the first report of the rhino mouse by Gaskoin in 1856 (7), a mutation in the mouse hairless gene. Mutants due to genetic engineering, such as insertional mutagenesis at the hairless locus created by Jones in 1993 (8), also help to define the minute details of how the mammalian skin functions. Explaining the genetic, biochemical, physiological, and pathological bases of the phenotypic changes in skin, hair and nails is the focus of modern dermatological research. Mammalian skin is an integral part of every mammal and should remind us to search for its interconnections with all of its constitutive parts as well as all internal organs.
At first, allow me to restrict my discussion to human skin. To understand the true function of our skin, we should imagine the situations in which some of the skin components are missing.
Among the three structural components of the skin, the lack of the deepest tissue (subcutis) does not cause any serious problems. In fact, the lack of subcutaneous fat in male genital skin causes no inconveniences. In the case of children or adult females, the absence of subcutaneous fat may damage the physical appearance as observed in patients with lipodystrophy, because they look as muscular as those well trained male athletes. Functionally, the skin offers only poor protection against cold without the subcutaneous fatty tissue, analogous to skin without the sweat apparatus that would make us unable to bear a hot environment. However, these are comparatively minor inconveniences for our survival.
In contrast, the lack of the surface portion of the skin, i.e. the epidermis together with the upper dermis, would cause a life-threatening problem because of the absence of the protective barrier, the stratum corneum (1, 2). Just imagine clinical situations in which the skin surface is extensively lacking such as in pemphigus vulgaris, epidermolysis bullosa, TEN or burn. They all present a serious problem for survival.
Although the stratum corneurn is thin, being less than 20microns in thickness at most sites, it effectively protects our body from desiccation. To sustain life, all of our body organs must be soaked in tissue fluid. Without such a skin barrier we would become as dry as a mummy in a short time. In addition, our body is protected efficiently by this barrier function from external invasion of various injurious agents such as poisonous chemicals, microorganisms and even the ultraviolet light of sunlight. Thus, the presence of the stratum corneum, not to mention the epidermis whose keratinocytes constantly keep producing corneocytes with the intercellular lipids and the upper dermis that sustains the existence of the epidermal keratinocytes, is essential for our existence in this world.
This vanguard of our protective system of the skin is further supported by subsidiary mechanisms that are also unique to the skin. The skin immune system and pigment production system, represented by two different kinds of dendritic cells in the epidermis, Langerhans cells and melanocytes, more specifically protect us from the injurious effects of invading exogenous substances and ultraviolet light, respectively (3, 4). In addition, the sensory nerve system that has direct contact with the brain makes us aware of imminent risks from the environment. However, these are less essential for us to sustain life than the stratum corneum. Even without these functions, we could still live if an artificial living condition that protects against these environmental effects is available. In fact, we can treat patients with grave immune-mediated dermatoses using intensive immunosuppressive therapy by placing them in a sterilized room. Likewise, those with extensive vitiligo or albinism whose skin cannot produce melanin pigment can live out their allotted span of life as long as they are well protected from the sunlight with sunscreen or remain in a dark room, like patients with xeroderma pigmentosum whose complete avoidance of the sun can assure a normal lifespan.
Thus, the minimum essential requirement for our existence in this world is the presence of the skin surface tissues that produce the stratum corneum. Then, what is the purpose for living creatures to exist in this world? Biologically, it is to produce as many offspring in as good a condition as possible, and to increase their habitats by overpowering other species. It is apparent that we humans cannot defeat wild beasts without weapons or that we cannot fly freely, swim swiftly or run very fast. Despite these physical weak points, nobody would deny that humans have succeeded in conquering this world and, to some extent, even outer space. It is with their intellectual powers of the brain that the sublime culmination of animal evolution is demonstrated by the ability to invent a suitable living condition under any circumstances on the earth.
As we perform comparative studies of various animals, we notice that the skin, which, interestingly, is also an organ whose epidermis is also derived from the ectoderm like the brain, has evolved to its best form in human beings, not only in terms of the function but also the external appearance. In attracting the attention of our fellow humans, the properties of the stratum corneum play a crucial role. Its appropriate amount of water content determines the smoothness, softness and texture of our skin surface. Therefore, in humans, like the brain, the skin will be the last organ for which transplants from other animals would be considered.
From the biological purpose of having good offspring by attracting desirable members of the opposite sex, it is natural for the human skin to reach the best aesthetic condition around adolescence, under the influence of hormones. Although some suffer from acne, sebum production is important to maintain the skin surface in a well-hydrated condition. The beauty of young skin has repeatedly become a subject of art.
Then, what biological meaning is there for the remaining part of our lives after leaving offspring? Is it just a period of waiting for physical decline, namely aging? In the case of humans, the civilization developed by them has grown in complexity to a point far exceeding what could be managed by the naive brain ability of young people. Only human beings remain far more active even in the later stages of their social life, after reaching adulthood by using their experience and judgement. In addition, they are the only animals that have used cosmetics or skin care products such as moisturizers, makeup products, perfume, soap, hair color, hair bleach, hair growth promoter or depilator and nail enamel to maintain their charm even in advanced age by hiding the decline in the skin appearance. Recently, the development of skin science has enabled us to even make disappear the signs of aging such as wrinkles and pigmented spots due to photoaging, the worst environmental damage for the skin that can even lead to the development of skin malignancies.
In conclusion, the basic function of the skin is to protect our body from environmental harm by producing the thin but most efficient biological wrapping structure on its surface. The immune and pigment production systems of the skin may form a secondary defense line against the environmental damage. and the tertiary one is the presence of the subcutaneous fatty tissue, sensory, nerves and skin appendages that produce hair, nail, sweat and sebum.
However, probably, modern science will soon succeed in producing an environment where we can live even in the total absence of the skin. Nevertheless, nobody would want to live there facing the scene in which our fellow humans moving around without their skin as seen in a horror movie. Who will fall in love with a fellow member of the opposite sex with the appearance of a figure found in a textbook of anatomy?
So, whatever may happen, there remains a very important function for the skin: to be appreciated by our fellow humans. In this context we cannot disregard the importance of hair, because hair loss has continued to be a serious issue only in human society.
References
1.Kligman AM. The biology of the stratum corneum. In: Montagna W, Lobitz WC, editors. The Epidermis. Orlando, Florida: Academic Press Inc; 1964. pp. 387–433. [Google Scholar]
3.Bos J. Skin Immune System (SIS) Cutaneous Immunology and Clinical Immunodermatology. 2. Boca Raton: CRC Press; 1997. [Google Scholar]
4.Jimbow K, et al. Biology of melanocytes. In: Freedberg IM, et al., editors. Fitzpatrick’s Dermatology in General Medicine. 5. New York: McGraw-Hill; 1999. pp. 192–220. [Google Scholar]
Skin is an intricate ectodermo-mesodermal covering organ of higher Eumetazoa – the Vertebrates (1). Its true function is to be a barrier between the internal and the external environment of the organism, but consequences of this apparently obvious statement reach far beyond the popular view on the problem.
Firstly, because the primary function of this barrier is neither the protection of the organism, nor its coloration. and secondly, as the possession of skin might have led to the evolutionary development of such a sophisticated organ like the human brain. Let me explain.
A cellular membrane suffices to protect the intracellular environment of many quite sizeable organisms. For example, plasmodial slime moulds (Myxomycetes) often reach a diameter of tens of centimeters and a mass of hundreds of grams. Their cytoplasm is usually strongly pigmented. Being still unicellular organisms covered by a single phospholipid bilayer, they may live both on land and in the water, and perfectly do without any covering tissue (2). Therefore, the primary function of skin is neither the protection nor the coloration of organisms.
The skin evolved gradually, after the emergence of the third germ layer – the mesoderm. The ectoderm, the primordial covering tissue, is present in all Eumetazoa (1). Higher Vertebrates enjoy many complicated and useful inner organs, as well as the skin. The possession of skin is therefore obviously coupled with the complication of internal structure of the animal organism.
Yet, this does not solve the problem of the true function of skin: possessing skin may be a reason, but also a consequence of evolving higher Eumetazoa. The explanation of the influence of skin on the higher level of complexity of the internal environment of the organism is given by the laws of thermodynamics.
Living organisms are very sophisticated open thermodynamic systems characterized by non-linear and nonequilibrial processes, as well as aniso-tropic and aperiodical structures (3). They have just been defined as a network of inferior negative feedback subordinated to a superior positive feedback (4). The type of a thermodynamic system and the processes taking place in it are to the largest extent determined by its barriers (borders, walls, shields) (5). In the case of a living system, they must fulfil two opposite tasks – to insulate the system from the environment, and to ensure selective transport and exchange of matter, energy, and information between the system and its environment (6). Under these, conditions, one can observe self-organization in the system, coupled with energy dissipation, and characterized by a local decline in entropy, peculiar to what we call the phenomenon of life (3–7).
In the history of life, such borders evolved twice. The cellular membrane appeared rapidly in the beginning, and the skin gradually evolved much later. It clearly belongs to the organism, whereas the status of the skin surface microenvironment, the air between hair shafts or feathers, seems ambigous. Skin is the place where internal and external environments of the organism penetrate each other. From the cybernetic point of view, skin is also a perfect border of the organism: the nervous system is of ectodermal origin, nerves interfere in the development of hair follicles (8, 9), many exteroreceptors are histological and functional parts of skin, even some parts of the skull are of the dermal origin (1).
The true function of skin is clearly to be the thermodynamic barrier of the multicellular living system. Just like the membrane is the reason, and not the consequence of the cellular organisation of living matter, the skin, being a product of histological complication of Eumetazoa, is firstly the reason of the intrinsic complication of Vertebrates.
References
1.Sembrat K. Comparative Histology of Animals. II. Warsaw: Polish Science Publishers; 1981. [Google Scholar]
2.Aldrich HC, Daniel JW, editors. Cell Biology and Physarum and Didynium Cell Biology. II. New York-London: Academic Press; 1982. (A Series of Monographs). [Google Scholar]
3.Prosser V. Place and Role of Physics in Other Sciences. In: Prosser V, et al., editors. Experimental Methods of Biophysics. Praha: Academia; 1989. [Google Scholar]
The question ‘What is the true function of skin?’ seems simple and easy to answer for any dermatologist:
Sustain water and electrolytes in the body
Protect towards physical and chemical damages including light
Immune defense – innate and specific
Tactile organ sensing temperature and damaging insults
A Latin word for skin is ‘integumentum’, meaning ‘a covering, especially of an animal or plant body, a skin, shell, rind, or husk’ (1). ‘Integrity’ is then the function of skin, meaning besides its physical covering ‘unimpaired moral principles; honesty; soundness; the quality of being whole or undivided’ (1).
The essay could stop here. But the posed question has a caveat: ‘true’. What is meant by ‘true’?
You cannot survive without skin. Therefore – posing the question means you will have skin and then – what is it’s ‘true’ function?
It has changed significantly over time. From being our protection to reflecting our integrity, i.e. how we look – to others and ourselves. Physical appearance and looking youthful and energetic are important signals to display. and being able to send those signals may be mandatory for your achievements in society and on the personal level. We still need the four functions as stated above, but clothing and cleansing of skin are significant assistants to normal skin function.
Normal skin including hair and nails are the best investment parts of the human body (2). Its true function today is to make money. I cannot imagine the fortunes made by the cosmetic industry, on hair products, nail polish and emollients, etc. I cannot imagine a better industry – creating so many hundreds of thousands of jobs, bringing joy to the individuals using the products, the business men and women selling the products, the beauty parlours, the industry developing the products and the stock market. And I have no bad feelings about this industry. It makes peoples’ dreams come true. Be beautiful, look beautiful, be young, etc. I have a wonderful person demonstrating this to me every day – my wife.
And there is plenty of potential in the ‘skin industry’: the person who will develop the drug, who can really grow hair on an old man’s balding head will become a billionaire beyond limits. Bill Gates will never get a chance to compete. Who will be the first to develop antiwrinkle creams securing a 20+ year younger look? Again, Bill, you will have stiff competition. I hope those new tycoons will be as wise as Bill Gates and make a Hair and Skin Foundation, where just 10% of the surplus would make a difference for persons in developing countries not having access to sufficient skin health care.
Skin and sex (dermatology and venereology) go together. Just take a 15-minute look at MTV. You don’t need more arguments. Also, a truly vital part of life.
So – what is meant by ‘true function’? Is it the boring physiological facts mentioned earlier, in the essay? Or has ‘skin’ created an industry so dependent upon ‘normal skin’?
Skin is not a remedy for the dermatologist to survive. Skin disease is not a normal function of the skin. But – again – it makes a living – and for many a good living. It is also good business. I don’t need to be specific, but just look at the major dermatological societies and their annual revenues based on congresses and information material.
The ‘true’ function of skin? Another favourite explanation of mine is that skin is an organ, where we can really study ‘biology’ and where we may learn enough to be able to reduce the suffering of our patients or even bring cure: imagine the day we understand atopic eczema or psoriasis and can prevent or ‘cure’ these diseases, which bring so much misery to many of our patients.
In Japan they have an old saying: ‘The truth is depending on the circumstances’ (rin ki ou hen: you can adapt yourself by/to the circumstances) (K. Yamamoto, Tokyo. Personal communication).
Skin is our integumentum. To keep our physical and chemical integrity, to stimulate our image of ourself, our sex life – and our motivation to attend the next skin meeting to learn just a little more.
References
1.The Living Webster Encyclopedic Dictionary of the English Language. Chicago: 1975. [Google Scholar]
Let us first consider the skin epithelium on the one hand, and lung and intestinal epithelia on the other hand. From a topological point of view, they form a continuum. Paradoxically, however, if the function of the latter is considered as exchange, the function of the former is taken as barrier (1). In fact, this barrier function might result from a ‘selfish’ vision, the human being considering himself as permanently attacked by its immediate environment.
This vision of the skin as a barrier contradicts the golden rules of Nature which are: interaction, communication and sharing of finite resources, rules on which rely all organisational networks. These rules indeed apply, in a fractal way, to metabolic pathways, cell–cell interactions, and tissue organisation.
Let’s consider a metabolic pathway: its subcellular compartmentalisation, the enzymes involved, their cofactors and substrates will define the topographical limits, that is to say the ‘envelop’ of this metabolic pathway. Interaction maps, as elaborated by 2-hybrid studies for example (2), now underline and reflect the contact zones between metabolic envelops. Cellular metabolism results from the integration of these contact zones. A striking example of these contact zones is the unexpected contact between glycogen metabolism and Wnt morphogenetic signalling pathway. GSK3, a kinase which inactivates glycogen synthase and thus blocks glycogen synthesis, is also – and surprisingly – a key element in the transduction pathway from Frizzled to Wnt (3), down to β-catenin/LEF-1 and the nucleus, this latter pathway being involved, for example, in hair morphogenesis (4). Interestingly enough, glycogen synthesis is active in the mid part of the human hair follicle but is turned off in these upper and lower parts of the outer root sheath, where reside these pluripotent cells from which the hair follicle is cyclically renewed (5).
On a larger scale, the cell membrane, with its intrinsic and sometimes restricted fluidity (6), receptors and dedicated contact structures, will control tissue organisation. Skin is an excellent paradigm of this concept. Desmosomes control keratinocyte–keratinocyte interactions and by this, the integrity of epidermis. Hemidesmosomes, together with specific trans-membrane protein like α6β4 integrin, in turn control the interaction of the epidermis with the underlying basement membrane and dermis, and by this, the cohesion of skin. If one expands this example to the entire body, one can say that the body general organisation and functions are under the control of and result from harmonious interactions and cross-talks between the different tissues.
If one goes one step further, in a fractal way, one could consider skin no longer as a barrier but rather as an envelop, endowed with interaction and communication functions, aimed at favouring social organisation between individuals.
Isn’t it odd to realise that when one touches other individuals, the feeling of cutaneous contact will be different each time? This contact and the feelings it elicts can easily translate affinity as well as reject, and through that can contribute to social organisation. Isn’t it odd to feel that the one you passionately love insinuate under your own skin? Some one else’s skin can be felt soft and suave, and thus provoke attraction. It can also be felt dry and harsh and provoke rejection. Skin does not lie, it is yes or no. From this point of view, skin certainly has some weight in regulating communication between individuals. Contact lies at the basis of organisation, and skin is one of its most privileged vectors. In my opinion, extreme care should be taken of skin, because it is an often neglected, even rejected, but nevertheless powerful vector of social organisation.
Considering this organiser key role and in order to maintain this vector of social equilibrium, which is constantly altered and assaulted by the environment, skin – and more precisely: epidermis – must be continuously renewed. That would explain why pluripotent cells are spread all over the skin and hair follicles to perpetuate this function. Touch, shake hands, kiss and caress are all kinds of contacts one can compare to the sometimes hesitating and furtive cell-cell contacts which contribute to the sorting phenomenon and tissue segregation. The skin is a precious extension of our brain (and vice versa, by the way). We must thus maintain it, learn how to use it, understand it and respect it, in order to promote and give back a meaning to interindividual contact, and ultimately to apprehend the major role of this organ, too often considered as a defensive barrier rather than a major communication organ: that of a social organizer (7).
Footnotes
1
This essay is my personal and instant point of view, and does not reflect the corporate opinion of the company I am working for.
In this brief commentary I am taking the opportunity to give my personal view of what is the function of skin, with the hope of providing further stimulus for skin-related research. Thus, I will not discuss the most important and obvious functions of this organ for normal homeostasis and protection of the organism, nor the psycho-social function of skin, as a critical conveyor of wellbeing and a determinant of beauty.
Rather, I will dwell on the function of skin as an experimental system that offers unique opportunities to study a number of fundamental questions, ranging from transmission of genetic information, to development and morphogenesis, to growth, differentiation and carcinogenesis.
The power of skin as an experimental system stems from the following obvious facts: 1. the skin is readily, accessible for direct examination and experimentation; 2. its structure is relatively simple, so that interactions between its main cellular components, can be readily studied; 3. its cells can be readily cultured, so that basic information on their growth/differentiation properties can be acquired.
In evaluating the value of skin as an experimental tool, one should ask which conceptually novel insights this system has provided or has the potential to provide, that were not gained by the use of other powerful systems. Thus, studies of pigment-and hair-related traits have long been a benchmark of mouse genetics. Classical dermis/epidermis recombination experiments were the ones pointing to the key role of epithelial–mesenchymal interactions in development and morphogenesis. The notions of multistep carcinogenesis, and genetic susceptibility to cancer, were pioneered by skin chemical carcinogenesis experiments in the mouse. More recently, the study of a human skin tumor, basal cell carcinoma, provided a striking demonstration that developmentally relevant genes identified in Drosophila have mammalian counterparts with a critical involvement in human disease.
The skin, together with the mammary gland, is the only organ from which primary epithelial cells can be cultured in sufficiently large amounts for direct biochemical analysis and genetic manipulation. Studies of primary cells as opposed to established cell lines are necessary in view of the secondary genetic changes which are known to occur upon prolonged cultivation. In addition, growth properties of epithelial cells are substantially different, at times opposite, from those of other better studied cell types, such as fibroblasts or cells of the hematopoietic system. Thus, detailed analysis of the growth/differentiation behavior of primary keratinocytes and underlying regulatory mechanisms has yielded and will continue to yield novel insights into the function of already known molecules, and lead to the discovery of new ones with key cell regulatory functions.
Epithelial cells do not live in isolation but in close coordination with other cells of the same and different types. Thus, the ability of recombining keratinocytes with other relevant cell types for skin reconstitution/in vivo transplantation experiments gives a unique opportunity to dissect the complex interactions involved in organ morphogenesis and homeostasis. An exciting complement to this type of analysis is now provided by the possibility of expressing or deleting specific genes in selective skin compartments, in a topically inducible manner.
The great strength of skin as an experimental system is also its potential weakness. In fact, inherent with the richness of biological and biochemical questions that can be asked, is the continuous risk of being lost in the pursuit of relatively minor issues, rather than addressing truly novel and relevant ones. With this challenge in mind, it can fairly be stated that the main function of skin for a basic cell biologist like myself is that of providing a paradigm for control of organ development, homeostasis and carcinogenesis.
What is the true function of skin? What is an untrue function? Put this way, the question is seen for what it is: more about what we think, than what is.
Sure, our version of events has to correspond with the data, but skin just is. The question, as a problem in reverse optimisation (‘we have the answer, but what was the problem?’) is aimed to force us to review the kinds of stories we like to tell about how things are – and how they got this way. And therein lies its value: a heuristic bootstrap.
Skin is: it got this way through eons of molecular and cellular and species evolution. Success was defined not by vision or plan, but by chance and adaptation to extant circumstances. Of our evolutionary tree, most branches have been pruned –Nature’s little failures – only lots of them. Yet, because there was no vision, nor goal, rather just a subroutine that allows diversity, and then chance and selection, skin is in evolutionary terms merely layer upon layer of biological expediency.
Of course, there will be some modularity of design, just as modularity of protein structure may facilitate evolutionary diversity and success, we may expect to see echoes of such skin design modules, or groupings, reflecting function, such as – to use an example from my own interest – thermoregulation.
Why is human skin so exquisitely tuned to be able to lose heat, and why the sexual dimorphism in this capacity? The answer, we guess, lies several million years ago in Africa. The need for sustained physical activity over long periods of time necessitates the striking efficiency of ecrine glands – capable of excreting more fluid in the pursuit of loss of heat than the kidneys can pass urine. But, contingent on this, and by definition unexpected, was that millions of years later half a dermatologist’s workload would relate to this earlier evolutionary decision. If you want to sweat efficiently, you need to loose body hair. If you loose body hair, you need to protect the interfollicular skin from ultraviolet radiation. The solution chosen was to reroute melanocytes from the follicle into the interfollicular skin and to adapt melanin’s focus from one part of the electromagnetic spectrum to that just a few nanometers shorter. But subsequently, and again contingent on diet and other factors, you need to worry about vitamin D and start to make evolutionary trade offs between sunburning and nutrition. But, for all of this explanation, all we see in the clinic is cancer. The echoes (only) of modularity are there, the layer upon layer of messy evolutionary change is there, but the function, if you can think of it in this way, is a myriad of stories set within a dimension of time where most stories have never been heard by human ears.
Of course the bugs are still in the code. Despite the millions of patches, we are still on a beta testing. And it is exactly this complexity and complication, and the need for time lines to be seen as part of the story, that makes understanding disease so difficult. So don’t expect any psoriasis gene modules: there are just genes. Genes for atopic dermatitis: no, just genes that in an evolutionary context have been associated with disease – or not. Diseases as modules are deep buried or very uncommon.
Here lies the reason why there is an uneasy relation between understanding biology and therapeutic insight. Disease might be complex, but treatment simple, or vice versa. We don’t design therapy by functional modules, rather we pick our pathways, preferably at some higher level of description, and fiddle with them and see what happens. So much of our therapy comes from not understanding at the machine code level, but by manipulating the operating system or playing around with the packages that are available. Like somebody learning a new drawing package we just fiddle around, randomly selecting menus looking for effects, and then build on it. Grand strategies before you have opened the package for the first time are doomed to failure.
So functions may be fine for biology but biology isn’t synonymous with medicine. Engineering should be our mantra: wrenches; hammers; warts and all.