

Abstract
Bone is a dynamic organ that undergoes constant remodeling, an energetically costly process by which old bone is replaced and localized bone defects are repaired to renew the skeleton over time, thereby maintaining skeletal health. This review provides a general overview of bone’s main players (bone lining cells, osteocytes, osteoclasts, reversal cells, and osteoblasts) that participate in bone remodeling. Placing emphasis on the family of extracellular matrix metalloproteinases (MMPs), we describe how: (i) Convergence of multiple protease families (including MMPs and cysteine proteinases) ensures complexity and robustness of the bone remodeling process, (ii) Enzymatic activity of MMPs affects bone physiology at the molecular and cellular levels and (iii) Either overexpression or deficiency/insufficiency of individual MMPs impairs healthy bone remodeling and systemic metabolism. Today, it is generally accepted that proteolytic activity is required for the degradation of bone tissue in osteoarthritis and osteoporosis. However, it is increasingly evident that inactivating mutations in MMP genes can also lead to bone pathology including osteolysis and metabolic abnormalities such as delayed growth. We argue that there remains a need to rethink the role played by proteases in bone physiology and pathology.
Keywords: bone, remodeling, metabolism, matrix metalloproteinase, deficiency, underactivity
Introduction
Bone is a hard, dense, rigid form of highly specialized connective tissue making up the skeleton of vertebrates. Bone protects internal organs, supports body structures, and aids in locomotion (Maffioli and Derosa, 2015). In addition, bone provides an environment for hematopoiesis (i.e., formation and development of blood cells) in the bone marrow, and acts as a homeostatic reservoir of calcium, phosphorus, insulin-like growth factors, transforming growth factor-β, and cytokines. Bone buffers the blood against drastic pH changes, thus detoxifying the circulation from heavy metals (Rauner et al., 2012). Bone develops by intramembranous ossification (e.g., bone of the clavicle, some skull bones), endochondral ossification (e.g., the appendicular and axial skeleton) or pseudo-metamorphic ossification (Rauner et al., 2012).
Bone remodeling is a complex process involving the sequential resorption of bone tissue and deposition of new bone at the same site (Kerschan-Schindl and Ebenbichler, 2012). Together with bone structure, geometry, size, and density, remodeling determines bone’s overall mechanical properties (e.g., the strength) (Mosekilde et al., 1993; Jiang et al., 1997; Ikeda et al., 2003; Shahnazari et al., 2009) as well as enables the repair of damaged bone and the adaption of bone to changing biomechanical forces (Kerschan-Schindl and Ebenbichler, 2012).
We review here the prevailing view of the bone remodeling process with an emphasis on well-accepted and newly emerging roles played by matrix metalloproteinases (MMPs) and cysteine proteinases in this process. Finally, we review the increasing number of instances in which inactivating mutations in MMP genes are found to lead to bone pathology including osteolysis and metabolic abnormalities such as delayed growth.
General Overview on the Cycle of Bone Remodeling
The bone remodeling process consists of four distinct consecutive phases spanning over 3–6 months (Datta et al., 2008).
The first phase of bone remodeling is known as the ‘activation phase’ and can be triggered by mechanical and nutritional stress on the bone as well as by hormones (e.g., parathyroid hormone, estrogen) (Parra-Torres et al., 2013). As described in Table 1, terminally differentiated osteocyte cell is a key player in the activation phase (Rauner et al., 2012; Parra-Torres et al., 2013).
TABLE 1.
Osteocytes and the activation phase of bone remodeling.
| Origin of osteocytes | Differentiation stages: (i) From mature osteoblasts to preosteocyte type I with dendritic projections formation; (ii) from preosteocyte type I to preosteocyte type II with cytoskeleton rearrangement; (iii) from preosteocyte type II to preosteocyte type III (mature osteocyte trapped within the mineralized bone matrix) with canaliculae formation (Hirao et al., 2007; Paiva and Granjeiro, 2017). |
| Important factors involved in osteocytogenesis | (i) Pre-osteoblasts (Stro1, CD29, CD105, CD166); (ii) Osteoblast (Cbfa1 and osterix for differentiation, alkaline phosphase and collagen for the production of osteoid, osteocalcin, casein kinase II); (iii) Osteoid osteocyte (Phex and MEPE for regulation of biomineralization and mineral metabolism, E11/gp38 and MMP-14 for dendrite/canaliculi formation, destrin for cytoskeleton rearrangement); (iv) Mineralizing osteocyte (DMP1 for regulation of biomineralization and mineral metabolism, CapG for cytoskeleton regulation); (v) Mature osteocyte (sclerostin, FGF23 for regulation of renal phosphate excretion, ORP150 for preserving viability in a hypoxic environment) (Bonewald, 2011). Other factors include TGF-β (D’Angelo et al., 2001; Karsdal et al., 2002), MMP-2/MMP-13/MMP-14 proteolytic axis (Barthelemi et al., 2012), Cx43, Dkk-1, Fetuin A, RANKL, MCS-F, and osteoprotegerin (Chen et al., 2018). |
| Key signaling events involved in osteocytogenesis | Osteocalcin, ALP, and other genes specific for osteoblast differentiation gradually downregulate (Paiva and Granjeiro, 2017). At the same time, different genes specific for osteocyte differentiation upregulate (such as CD44 [Hughes et al., 1994], E11/gp38 [Zhang et al., 2006], Phex [Ruchon et al., 2000; Westbroek et al., 2002], Fimbrin [Tanaka-Kamioka et al., 1998], MEPE [Rowe et al., 2004], DMP1 [Feng et al., 2006; Toyosawa et al., 2012], sclerostin [Poole et al., 2005; Balemans et al., 2008], ORP150 [Gao et al., 2010], and FGF23 [Liu et al., 2006]). Transcription factors involved in the process of osteoblast/osteocyte transition are ATF-4, whose expression is regulated by JNK, and members of the AP-1 group (Matsuguchi et al., 2009). |
| Role of osteocytes | (i) Maintain physical connections with each other, and also other players (osteoclasts, osteoblasts) of the bone remodeling cycle through a widespread network of tiny channels called canaliculi (Civitelli, 2008). (ii) May remodel the perilacunar matrix (e.g., during lactation) by expressing cathepsin K and acid phosphatase. (iii) Regulate bone remodeling by expressing M-CSF and RANKL (stimulate osteoclast formation and activity) as well as NO and OPG (inhibit osteoclast formation and activity). Also, osteocytes control bone formation by secreting activators (e.g., NO, ATP, PEG2) and inhibitors (e.g., sFRP1, DKK1, sclerostin) of the Wnt signaling pathway. (iv) Source of factors (e.g., sclerostin) and regulators (e.g., FGF-23, DMP-1, Phex, MEPE) of phosphate metabolism. (v) Manage the bone’s reservoir of calcium. (vi) Function as mechanosensory cells (Bonewald, 2011; Dallas et al., 2013; Bellido, 2014). |
| Molecular mechanism that underlies the function of osteocytes as mechanosensory cells | Osteocytes are good mechanosensors (i.e., they detect changes of mechanical stimuli) in bone tissue which serve to sense and respond to alterations produced when a bone is mechanically loaded. Such alterations may be physical deformation of the bone matrix, fluid flow shear stress generated by variations in canalicular fluid flow and electrical streaming potentials (Bonewald and Mundy, 1990; Mundy, 1993; Manolagas, 2000; Miyauchi et al., 2000; Bonewald and Johnson, 2008; Datta et al., 2008; Parra-Torres et al., 2013; Takemura et al., 2019). Mechanical strain signal is converted into a cellular response (i.e., biochemical signals) with the participation of membrane proteins (such as CD44, connexins, integrins, and ion channels) and downstream mediators of intracellular signaling (such as guanine regulatory proteins, mitogen activated protein kinase, cyclic adenosine monophosphate, inositol triphosphate, and intracellular calcium) (Rawlinson et al., 1996; Burger and Klein-Nulend, 1999; Mikuni-Takagaki, 1999; Miyauchi et al., 2000; Gu et al., 2001; Alford et al., 2003; Kapur et al., 2003; Plotkin et al., 2005; Rubin et al., 2006; Miyauchi et al., 2006). On the other hand, bone remodeling is also controlled by upregulation of RANKL and sclerostin in response to a decrease in mechanical signals (Parra-Torres et al., 2013). The precise signaling biochemical pathways (e.g., Wnt/β-catenin) and regulatory mechanisms that may mediate adaptive responses activated by mechanical loading and unloading in bone remain to be completely delineated (Dallas et al., 2013; Parra-Torres et al., 2013). |
| Other consequences of osteocyte activities on bone remodeling | Retraction of the bone lining cells (elongated mature osteoblasts) on the endosteal surface (which is a thin layer of cell-rich connective tissue), and also digestion of the underlying collagenous membrane by collagenases (Murray et al., 1995; Karsdal et al., 2001; Datta et al., 2008; Kerschan-Schindl and Ebenbichler, 2012). |
MMP, matrix metalloproteinase; TGF, transforming growth factor; RANKL, receptor activator of nuclear factor kappa B ligand; MAPK, mitogen-activated protein kinase; TIMP, tissue inhibitor of metalloproteinase; ALP, alkaline phosphatase; Phex, phosphate-regulating endopeptidase homolog X-linked; MEPE, matrix extracellular phosphoglycoprotein; Cx, connexin; Dkk, Dickkopf WNT signaling pathway inhibitor; Phex, phosphate-regulating endopeptidase homolog X-linked; DMP, dentin matrix acidic phosphoprotein precursor; M-CSF, Macrophage colony-stimulating factor; ORP, oxygen regulated protein; FGF, fibroblast growth factor; ATF, activating transcription factor; JNK, c-Jun N-terminal kinase; AP, activator protein; CD29, integrin beta-1; CD105, endoglin; CD166, activated leukocyte cell adhesion molecule; CapG, capping actin protein, gelsolin like; ORP150, 150-kDa oxygen-regulated protein; OPG; osteoprotegerin, ATP, adenosine triphosphate; NO, nitric oxide; sFRP1, secreted frizzled-related protein 1; DKK1, Dickkopf WNT signaling pathway inhibitor 1; PEG2, prostaglandin E2; Wnt, Wingless-type MMTV integration site family.
The second phase lasts 8–10 days (Teitelbaum, 2007) and is called the ‘bone resorption phase’ – a process by which large multinucleated osteoclast cells break down old bone organic matrix impregnated with minerals (e.g., calcium phosphate nanocrystals), as described in Table 2.
TABLE 2.
Osteoclasts and the bone resorption phase.
| Origin of osteoclasts | Differentiation stages: Hematopoietic stem cell precursors differentiate into monocyte and macrophage, and then they fuse into end-differentiated multinucleated (bone resorbing) cells (Tanaka et al., 1993; Quinn et al., 1998; Roodman, 1999; Udagawa et al., 1999; Holmbeck and Szabov, 2006; Bar-Shavit, 2007; Bruzzaniti and Baron, 2006). Osteocyte apoptosis is thought to contribute to the recruitment of osteoclast precursors by diminishing the secretion of osteocyte-derived factors (e.g., TGF-β) that have inhibitory effect on osteoclast formation (Heino et al., 2002; Aguirre et al., 2006). |
| Main factors involved in osteoclastogenesis | Osteoblasts, osteocytes, RANKL, M-CSF, OPG, TNF, ILs, mineralized bone particles containing osteocalcin, DC-STAMP, OC-STAMP (Tanaka et al., 1993; Roach, 1994; Wiebe et al., 1996; Kotake et al., 1999; Udagawa et al., 1999; Marie, 2003; Miyamoto, 2006; Kim et al., 2011; Hienz et al., 2015; Plotkin and Bruzzaniti, 2019). |
| Key signaling events involved in osteoclastogenesis | After the induction of PU.1, the stem cell precursor is determined to the osteoclastic lineage (Tondravi et al., 1997). Then, cell proliferation is induced following expression and activation of c-fms by the precursor. RANK is subsequently expressed and activated by RANKL, after which RANK interacts with the TRAF family members (e.g., TRAF2, TRAF6) and lead to downstream activation of MAP kinases and NF-kβ. This process is aided by co-signaling from other receptors (such as TREM2, OSCAR, DAP 12, and FcRγ) (Koga et al., 2004; Mocsai et al., 2004). The interaction between immunoreceptors (e.g., TREM2, OSCAR) and FcRγ/FcRc adapters activates Syk kinases, leading to PLCγ activation. Ca(II), which is mobilized from the intracellular stores, activates calcineurin, resulting in dephosphorylation of NFATc1. Moreover, the activation of calcineurin involves the activation of phospholipase-Cγ and Tec kinases (Mocsai et al., 2004; Faccio et al., 2005; Wada et al., 2005). In general, most signaling pathways (MAPKs, NF-κB, AP-1, Ca(II), Src/PI3K/AKt) which are activated in the osteoclast converge to induce the activity of NFATc1 (Gori et al., 2000; Ishida et al., 2002; Takayanagi et al., 2002; Matsuo et al., 2004; Paiva and Granjeiro, 2017; Plotkin and Bruzzaniti, 2019; Zheng et al., 2019). Upon translocation to the nucleus, NFATc1 acts together with c-fos to promote the expression of key osteoclast genes. Some of the osteoclast differentiation genes to which NFATc1 binds directly are OSCAR (Kim Y. et al., 2005), cathepsin K (Matsumoto et al., 2004), calcitonin receptor (Matsuo et al., 2004), integrin β3 (Crotti et al., 2006, 2008), MMP-9 (Sundaram et al., 2007), and TRAP (Matsuo et al., 2004; Paiva and Granjeiro, 2017). Of note, another factor which controls NFATc1 is OPG, which functions as a decoy receptor for RANKL, thus inhibiting the differentiation of osteoclasts (Lacey et al., 1998). Osteoclastogenesis is regulated by the RANKL/OPG balance. Opposing effects on RANK during osteoclast differentiation is exerted by LGR4 which signals through G-protein or Wnt signaling pathways (Luo et al., 2016). Cytokines which inhibit RANK signaling on osteoclasts are IL-10, IFNs (α, β), and GM-CSF. |
| Mechanisms that underlie the action of osteoclasts | During initiation of the resorption phase, the mature osteoclasts (1-2% of bone cells) attach to the bone surface via αvβ3, αvβ5, α2β1, and αvβ1 integrins (Vaananen and Horton, 1995; Datta et al., 2008; Rauner et al., 2012; Plotkin and Bruzzaniti, 2019). At the bone/osteoclast surface, a ruffled border which is entirely surrounded by a sealing zone is formed, thereby creating an isolated resorption (Howship’s) lacuna (i.e., scalloped erosion) (Miyauchi et al., 1991; Mimura et al., 1994; Teitelbaum, 2000; Teitelbaum and Ross, 2003). Osteoclasts dissolve mineral (hydroxyapatite) and organic components (e.g., type I collagen) of the bone matrix in the resorption lacuna (Teitelbaum et al., 1995; Rauner et al., 2012). This resorption process is mediated by the secretion of hydrogen ions, to acidify the resorption compartment beneath osteoclasts and dissolve hydroxyapatite crystals (Blair et al., 1989; Teti et al., 1989). Hydrogen ions, supplied by the reaction of water and carbon dioxide and catalyzed by carbonic anhydrase II, are transported into the resorption lacuna by ATPases located in the ruffled border of osteoclasts (Baron, 1989; Mattsson et al., 1994; Li et al., 1999; Bruzzaniti and Baron, 2006; Hienz et al., 2015). Hydrochloric acid formed with chloride ions pumped into the resorption lacuna dissolves the mineralized bone matrix (Silver et al., 1988; Plotkin and Bruzzaniti, 2019). In addition, lysosomal enzymes (e.g., cathepsin K), bone-derived collagenases, and other proteinases (e.g., tartrate-resistant acid phosphatase) act in concert to mediate the resorption process (Bord et al., 1996; Gelb et al., 1996; Saftig et al., 1998; Boyle et al., 2003; Teitelbaum, 2007; Hienz et al., 2015). Osteoclast-mediated bone resorption, which takes a few (2-4) weeks during each remodeling cycle, results in Howship’s lacuna on the surface of trabecular bone and cylindrical Haversian canals in cortical bone (Bruzzaniti and Baron, 2006; Teitelbaum, 2007; Hienz et al., 2015). After one resorption lacuna is completed, the osteoclast cells die by apoptosis (Plotkin and Bruzzaniti, 2019) or move along the bone surface to resume resorption. This phase lasts approximately 8-10 days (Teitelbaum, 2007). |
| Systemic and local factors that stimulate bone resorption | Osteocytes as the major source of RANKL; thyroid hormones; PTH/PTHrP; calcitriol; glucocorticoids; growth factors (FGF, PDGF, EGF); TNF-α; colony-stimulating factors (M-CSF, GM-CSF); IL-1, -6, -7, -8, -11, -15, -17; PGE1, 2, 12; PGH2 (MacDonald, 1986; Dempster et al., 1993; Raisz, 1993; Kawaguchi et al., 1994, 1995; Nash et al., 1994; Holt et al., 1996; Lanske et al., 1999; Roodman, 1999; Lam et al., 2000; Compston, 2001; Ragab et al., 2002; Sher et al., 2004; Eijken et al., 2005; Dai et al., 2006; Zhang et al., 2008; Kini and Nandeesh, 2012; Rauner et al., 2012; Parra-Torres et al., 2013; Paiva and Granjeiro, 2017; Hachemi et al., 2018; Bellido and Gallant, 2019). |
RANK, receptor activator of nuclear factor kappa B; RANKL, receptor activator of nuclear factor kappa B ligand; GM-CSF, granulocyte-macrophage colony-stimulating factor; OPG, osteoprotegerin; TNF, tumor necrosis factor; IL, interleukin; DC-STAMP, dendritic-cell specific transmembrane protein; OC-STAMP, osteoclast stimulatory transmembrane protein; NFkB, nuclear-factor kappa B; TRAF6, TNF receptor-associated factor 6; TREM2, triggering receptor expressed on myeloid cells-2; OSCAR, osteoclast-associated receptor; DAP, DNAX-activating protein; FcRγ, Fc common receptor γ chain; FcRc, soluble Fc receptor from a group C streptococcus; Syk, spleen tyrosine kinase; PLC, phospholipase C; NFATc1, nuclear factor of activated T cell cytoplasmic 1; Tec, tyrosine protein kynase; AP, activator protein; Src, steroid receptor coactivator; PI3K, phosphatidylinositol 3-phosphate kinase; TRAP, tartrate-resistant acid phosphatase; LGR, leucine-rich repeat-containing G protein-coupled receptor; IFN, interferon; PTH, parathyroid hormone; PTHrP, PTH-related protein; FGF, fibroblast growth factor; PDGF, platelet-derived growth factor; EGF epidermal growth factor; M-CSF, macrophage colony-stimulating factor; PGE, prostaglandin E; PGH, prostaglandin H.
The third ‘reversal’ phase connects osteoclastic bone tissue resorption and osteoblastic bone tissue formation (Delaisse, 2014) and lasts 7–14 days (Pettit et al., 2008; Hienz et al., 2015). After departure of the osteoclast from a cavity in bones undergoing resorption, which is a resorptive lacuna known as the Howship’s lacuna, bone lining cells occupy the Howship’s lacuna and clean it (Everts et al., 2002). The cleaning process occurs by enwrapping and digesting non-mineralized collagenous proteins protruding from the bone surface left by osteoclasts. This cleaning process is a requirement for the subsequent deposition of a first layer of collagen along the Howship’s lacuna (Everts et al., 2002). Four types of osteoclast-derived coupling factors stimulate bone formation during the reversal phase: (i) Matrix-derived factors including transforming growth factor-β, bone morphogenetic protein-2, platelet-derived growth factor, and insulin-like growth factors, which are released during bone tissue resorption, (ii) Osteoclast-secreted factors, including cardiotrophin-1, sphingosine-1-phosphate, collagen triple helix repeat containing 1, and complement factor 3a, (iii) Osteoclast membrane-bound factors such as EphrinB2 and Semaphorin D, and (iv) Structural changes brought about by the osteoclast on the bone tissue surface (Sims and Martin, 2014). Reversal cells originating from pre-osteoblast cells (Andersen et al., 2013) colonize the osteoclast-eroded surface and respond to osteoclast-derived messages and coupling factors along with fibroblast-like cells covering the surface of bone (known as bone lining cells), osteoblast precursors, and canopy cells (Delaisse, 2014; Sims and Martin, 2014; Lassen et al., 2017; Pirapaharan et al., 2019).
The fourth phase of the bone remodeling cycle is ‘formation,’ when mononucleate osteoblast cells synthesize new bone organic matrix formed by collagen fibers and non-collagenous proteins (e.g., bone sialoprotein, osteopontin, osteocalcin, proteoglycans) that later becomes surrounded and impregnated with mineral deposit mainly in the form of calcium hydroxyapatite. A summary of osteoblastogenesis, the roles played by osteoblasts during this last phase, and the fate of osteoblasts is described in Table 3.
TABLE 3.
Osteoblasts and the bone formation phase.
| Origin of osteoblasts | Differentiation stages: (i) From stem cell to mesenchymal (adult) stem cell; (ii) from mesenchymal stem cell to preosteoblast (immature); (iii) from preosteoblast to mature osteoblast (Datta et al., 2008). |
| Key factors involved in osteoblastogenesis | Hormones (such as PTH, glucocorticoids, estrogen, leptin, 1,25-dihy-droxyvitamin D3) (Datta et al., 2008; Mohanakrishnan et al., 2018; Arumugam et al., 2019; Plotkin and Bruzzaniti, 2019), growth factors (such as EGF, TGF-β, IGF) (Datta et al., 2008; Canalis, 2009; Plotkin and Bruzzaniti, 2019), local factors (such as the family of intracellular glycoproteins known as BMPs -2, -4, -6, -7) (Shore et al., 2006; Wutzl et al., 2010), members of the Wnt family in a paracrine/autocrine fashion (Bodine and Komm, 2006), Sonic and Indian hedgehogs (Maeda et al., 2007; Guan et al., 2009), cell-to-cell communication through receptors (such as Notch, Ephrin-Ephrin) and connexins (e.g., Cx43) (Plotkin and Bruzzaniti, 2019). |
| Key signaling events involved in the canonical Wnt/β-catenin pathway | Wnt proteins bind to FZD receptor and its co-receptor (e.g., LRP4, LRP5, LRP6). CK1α then phosphorylates Dvl and in turn the complex Dvl-Frat1-axin-LRP5/6-FZD is formed. These events result in GSK3β inhibition, thereby avoiding modification (degradation, phosphorylation) of β-catenin. The stable β-catenin is then translocated to the nucleus to activate transcription factors (e.g., TCF, LEF), thus inducing the transcription of Wnt target genes (e.g., osteoprotegerin) (Datta et al., 2008; Plotkin and Bruzzaniti, 2019). Wnt signaling is regulated by a variety of molecules at the levels of extracellular inhibition of Wnt ligands or LRP4/5/6, co-receptors, intracellular signaling, and transcription (Gong et al., 2001; Boyden et al., 2002; Tian et al., 2003; Logan and Nusse, 2004; Semenov et al., 2005; Datta et al., 2008; Chen et al., 2019). Besides the canonical Wnt/β-catenin pathway, Wnt ligands can also activate other different signaling cascades (such as the Wnt-Ca(II), planar cell polarity, and protein kinase A pathways). |
| Key signaling events involved in the BMPs pathway | BMPs (e.g., BMP-2, BMP-7) as well as other signaling pathways (e.g., members of the Wnt pathway, TGFβ1, Indian hedgehog, notch, ephrin [Huang et al., 2007; Datta et al., 2008; Rauner et al., 2012; Plotkin and Bruzzaniti, 2019]) converge to regulate the expression of runx2 and others (such as β-catenin [Krishnan et al., 2006], osterix [Kim et al., 2006], msx2 [Liu et al., 1999; Satokata et al., 2000], NFATc1 [Koga et al., 2005], ATF4 [Tozum et al., 2004], Dlx3/5/6 [Harris et al., 2003], FGFR3, FGFs [e.g., -2, -9, -18], Phex, NFAT2, ALP) (Paiva and Granjeiro, 2017). Specifically, the expression of runx2, which is the master transcription factor, is controlled by Twist and menin-1, TAZ, and post-translational modifications. Also, Runx2 phosphorylation under TGF-β1 stimulation occurs at three serine amino acids (Arumugam et al., 2018). Osterix may interact with NFAT2, which participates in regulating gene transcription (e.g., for osteopontin, osteoclacin, osteonectin) (Paiva and Granjeiro, 2017). |
| Roles played by osteoblasts | Once osteoclasts have created a resorption cavity and detached from the bone surface, osteoblasts move into the cavity to initiate bone formation (Datta et al., 2008). Osteoblasts synthesize and lay down new unmineralized bone matrix (osteoid), which is subsequently mineralized (e.g., forming hydroxyapatite) over a period of about 20 days. Osteoblasts also synthesize and secrete the bone matrix proteins osteopontin, osteocalcin, bone sialoprotein, proteoglycans, and alkaline phosphatase (Baron, 1989; Roach, 1994; Ducy et al., 2000; Datta et al., 2008; Hienz et al., 2015). Why is the synthesis of non-collagen proteins necessary? These non-collagenous bone matrix proteins help to coordinate matrix mineralization and are essential for cellular adhesion (such as chemoattractant activity by osteocalcin), and regulation of cell activity (such as the osteopontin- and osteonectin-displayed cell activities) during coupling of bone resorption and formation (Robey, 1989; Raynal et al., 1996; Hienz et al., 2015). There is another function of osteoblasts that is worth highlighting. Osteoblasts also inhibit the ability of osteoclasts to degrade osseous tissue (Datta et al., 2008). |
| Stimulators of osteoblast functions | The increased formation of osteoid to build bone is stimulated by hormones (such as the pituitary-secreted growth hormone, sex hormones [estrogens and androgens], and thyroid hormone) (Kini and Nandeesh, 2012). Other factors that have stimulating effect on bone formation are insulin, vitamin D metabolites, IGF-I, IGF-II, TGF-β, BMP-2, BMP-4, BMP-6, BMP-7, IL-13, IFN, and OPG (Baylink et al., 1993; Cohick and Clemmons, 1993; Fraher, 1993; Rosen and Donahue, 1998; Yamaguchi et al., 2000; Canalis et al., 2003; Lovibond et al., 2003; Datta et al., 2008; Tang et al., 2009; Ruan et al., 2010; Kini and Nandeesh, 2012; Xian et al., 2012; Hienz et al., 2015). |
| Osteoblast fate | Bone-forming osteoblasts become encased in the mineralized matrix surrounding them, turning into osteocytes that gradually stop synthesizing osteoid (i.e., the newly formed unmineralized organic bone matrix) (Datta et al., 2008; Rauner et al., 2012). Osteocytes are evenly distributed throughout the bone matrix which enables contact with osteoblasts and vasculature (Kamioka et al., 2001; Plotkin et al., 2002; Zhao et al., 2002; Plotkin et al., 2008). Osteocytes not only facilitate mechanosensation as described in Table 1, but also control bone structure (amount and quality) through mineralization inhibitors such as dentin matrix protein-1, fetuin-A, and Wnt inhibitor (Poole et al., 2005; Feng et al., 2006; Coen et al., 2009; Liu et al., 2009; Rauner et al., 2012). Although it was thought that osteocytes remain inactive until the next bone remodeling cycle (Mikuni-Takagaki, 1999; Kamioka et al., 2001; Zhao et al., 2002; Knothe-Tate et al., 2004; Datta et al., 2008), it is now accepted that osteocytes constantly remodel the surrounding extracellular matrix (Yee et al., 2019). Another fate of osteoblasts is to become bone lining cells, which cover the freshly formed endosteal bone surface thus forming a physical barrier to avoid the process of osteoclast adhesion and bone resorption. |
PTH, parathyroid hormone; EGF, epidermal growth factor; TGF transforming growth factor, IGF, insulin-like growth factor; BMP, bone morphogenetic protein; LRP, low-density lipoprotein receptor-related protein; FZD, seven-span transmembrane receptor protein Frizzled; CK, casein kinase; Dvl, disheveled; GSK, glycogen synthase kinase; TCF, T cell factor; LEF, lymphoid enhancer binding factor; runx2, runt-related transcription factor 2; osterix, Sp7 transcription factor; msx2, homeobox factor; NFAT, nuclear factor of activated T cells; ATF, activating transcription factor; Dlx, distal-less homeobox; FGF, fibroblast growth factor; FGFR, fibroblast growth factor receptor; Phex, phosphate-regulating neutral endopeptidase; ALP, alkaline phosphatase; TAZ, tafazzin; IL, interleukin; IFN, interferon; OPG; osteoprotegerin; Wnt, Wingless-type MMTV integration site family.
While bone formation surpasses resorption during childhood, bone formation and resorption are in balance during young adulthood. However, an unbalanced bone loss occurs with aging (Datta et al., 2008; Rauner et al., 2012; Brandi and Piscitelli, 2013) and could predispose an individual to skeletal disorders including: (i) inflammatory bone loss in periodontal disease, (ii) arthritis (stimulation of bone resorption and inhibition of bone formation by prostaglandins and cytokines), (iii) osteoporosis (bone resorption outpaces bone formation), (iv) hyperparathyroidism and hyperthyroidism (greatly increased rate of bone resorption and formation), (v) Paget’s disease (increased and abnormal [shape, weakness, and brittleness] bone formation), (vi) osteomalacia (delayed/defficient bone mineralization), and (vi) osteopetrosis (failure of osteoclasts to resorb bone) (Roodman et al., 1992; Delmas, 1995; Gallagher, 1997; Mills and Frausto, 1997; Raisz, 1997; Charles and Key, 1998; Schneider et al., 1998; Siris, 1998; Kini and Nandeesh, 2012).
Matrix Metalloproteinases: Modulators of Bone Remodeling
Matrix metalloproteinases are a family of at least 24 highly homologous, multi-domain enzymes (Figure 1) with the capacity to degrade virtually all extracellular matrix components including collagen, aggrecan, elastin, and fibronectin (Lu et al., 2011; Fernandez-Patron et al., 2016).
FIGURE 1.
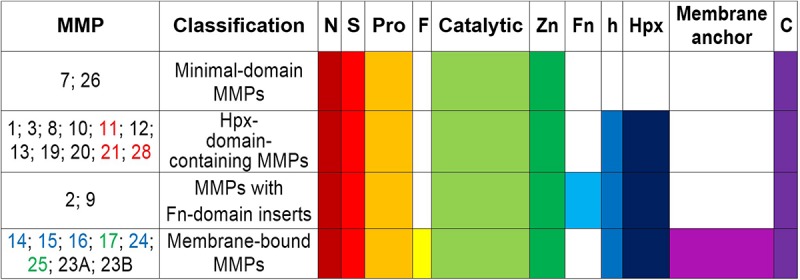
Schematic structure and classification of matrix metalloproteinases. S, amino-terminal signal sequence; Pro, pro-peptide; Zn(II)-binding site; h, hinge region; Hpx, hemopexin; FN, collagen-binding type II repeats of fibronectin; F, furin; MT-MMPs, membrane-type MMPs; N, N-terminus; C, C- terminus. MMPs -11, -21, and -28 (all in red) contain a Furin-like cleavage domain. MMPs -17 and -25 (both in green) contain a glycophosphatidyl inositol-anchoring sequence. MMPs -14, -15, -16, and -24 (all in blue) comprise a transmembrane domain with a cytosolic tail. MMP23A and MMP23B lack the signal peptide, the cysteine-switch motif and the hemopexin-like domain, but they contain a unique cysteine-rich domain, an immunoglobulin-like domain and an N-terminal type II transmembrane domain (Velasco et al., 1999).
All MMP family members are synthesized as catalytically inactive (latent) pro-enzymes (pro-MMPs) that contain a: signal N-terminal peptide sequence (∼20 amino acids), pro-peptide domain (∼80 amino acids), catalytic domain (approximately 160 amino acids), hinge (linker peptide) region of variable length (10–30 amino acids), and a hemopexin-like C-terminal domain (Hpx) (∼210 amino acids). The smallest MMPs (MMP-7 and MMP-26) lack the hinge and hemopexin domains, and therefore exhibit a reduced affinity for gelatin. MMP-23 has unique domains (such as the cysteine array, IgG-like domain, interleukin-1 type II receptor-like domains) instead of the hemopexin domain (Massova et al., 1998; Pei et al., 2000; Bode and Maskos, 2003; Visse and Nagase, 2003; Nagase et al., 2006; Piccard et al., 2007; Lopez-Otin et al., 2009; Bonnans et al., 2014; Vandooren et al., 2014; Vandenbroucke and Libert, 2014; Cui et al., 2017). The amino-terminal signal peptide targets the pro-MMPs to the rough endoplasmic reticulum, whereas the C-terminus harbors a cysteine residue and a furin cleavage site (PRCGXPD), both of which are important for conversion into the mature, active enzyme (Bonnans et al., 2014). Presence of an intact pro-peptide accounts for the latency of pro-MMPs, which can be overriden through the activation of a “cysteine-switch” mechanism (Van Wart and Birkedal-Hansen, 1990). The pro-peptide contains a cysteine residue that prevents catalytic activity when it is coordinated with a Zn(II)-ion in the catalytic domain (Springman et al., 1990; Van Wart and Birkedal-Hansen, 1990). The cysteine-Zn(II) interaction can be disrupted by alkylating compounds such as the organomercurial 4-aminophenylmercuric acetate as well as by serine proteases and other MMPs such as membrane-type MMPs, which act at the cell surface to which they anchor through their transmembrane domain/short cytoplasmic tail or by glycosylphosphatidylinositol linkage (Bonnans et al., 2014). MMP autolysis is another mechanism of activation mediated by allosteric perturbation of the inactive proenzyme (Springman et al., 1990; Van Wart and Birkedal-Hansen, 1990; Pei and Weiss, 1995; Pei et al., 2000; Meng et al., 2016). The catalytic domain harbors the Zn(II)-binding motif HEXXHXXGXXH, a catalytic Zn(II), a structural Zn(II), specific pockets related to specificity (S1, S2,…Sn and S1′, S2′,…Sn′) and coordinated Ca(II) ions which confer stabilization. The catalytic Zn(II) is coordinated by three histidine residues (Bode and Maskos, 2003; Bonnans et al., 2014; Vandenbroucke and Libert, 2014). The hinge domain is flexible and mediates interactions with substrates, cell-surface proteins, and tissue inhibitors (Cui et al., 2017; Liu and Khalil, 2017). The hemopexin domain modulates substrate recognition and specificity, binding to cell-surface receptors and inhibitors, activation of MMPs, and cellular MMP internalization for degradation (Visse and Nagase, 2003; Nagase et al., 2006; Piccard et al., 2007).
Matrix metalloproteinases expression and activity are tightly regulated at various levels: gene transcription, translation and secretion of the inactive enzyme precursor, proteolytic activation of the zymogen, spatial localization, interaction with specific extracellular matrix proteins, and inhibition by endogenous inhibitors (such as tissue inhibitors of MMPs [TIMPs 1-4], α2-macroglobulin, and human fibrinogen) (Sottrup-Jensen, 1989; Overall et al., 1991; Kusano et al., 1998; Zeng et al., 1998; Sternlicht and Werb, 2001; Han et al., 2003; Greenlee et al., 2007; Clark et al., 2008; Fanjul-Fernandez et al., 2010; Hadler-Olsen et al., 2011; Arpino et al., 2015; Sarker et al., 2019). Despite their similar names, TIMPs 1-4 exhibit large differences in their primary sequence, tissue expression, transcriptional regulation and in their inhibitory spectrum (Brew et al., 2000). In bone, TIMP-2 and TIMP-3, unlike TIMP-1, are effective inhibitors of the membrane-type MMPs (e.g., MMP-14), while TIMP-3 displays the broadest inhibitory actions of all TIMPs against metalloproteinases. Unlike TIMP-1, -2, and -4, which are soluble, TIMP-3 has basic amino acid residues in its C- and N-termini through which TIMP-3 attaches to heparan and chondroitin sulfate in the extracellular matrix and inhibits both MMPs and members of ‘a disintegrin and metalloproteinase’ (ADAM) and ‘a disintegrin and metalloproteinase with thrombospondin domains’ (ADAMTS) family including ADAM-17 and ADAMTS-4/-5 (Porter et al., 2005; Javaheri et al., 2016). Deficiency of tissue inhibitors (TIMP-1, -2, or -4) has minor impact on bone phenotype. However, both Timp3 deficiency and transgenic overexpression alters craniofacial bones of endochondral and intramembranous origins in mice, while the growth plates appear normal in these mice (Javaheri et al., 2016). Paradoxically, mice deficient in RECK (an MMP inhibitor anchored on the cell membrane with inhibitory actions against MMP-2, -9, and -14 and ADAM-10) die in utero displaying a perturbed extracellular matrix organization (Javaheri et al., 2016).
These observations suggest that bone remodeling may not be solely defined by the balance/imbalance between MMPs and TIMPs. Rather, other molecules expressed and released in the settings of bone physiology and pathology such as RECK (Paiva and Granjeiro, 2014) and some acute phase reactants (alpha 2-macroglobulin, fibrinogen) may regulate/dysregulate MMP activity in inflammatory conditions thus perturbing the normal bone remodeling process (Cook et al., 2018; Sarker et al., 2019). A consequence implied by the latter notion is that MMPs, ADAMs and ADAMTS molecules may be released from bone or non-bone tissues to influence bone remodeling through autocrine and paracrine actions. In other words, MMPs likely circulate bound to non-classical inhibitors (such as acute phase reactants) being recruited to sites of active bone remodeling, where local substrates act as chemoattractants and local activators (other proteases, reactive oxygen species) activate them.
The aforementioned levels of regulation effectively dissociate MMP expression from MMP activity (e.g., since overexpression of endogenous MMP inhibitors would effectively reduce MMP activity). Current biochemical techniques for assessing MMP activity are non-reliable. However, as research requires a proxy, MMP expression is often used as a surrogate (albeit incorrectly) for MMP activity. There remains an urgent need for highly sensitive, specific, and robust methods for assessing the activity potential of individual MMPs such that therapeutic strategies can be designed to specifically reduce the activity of overactive MMPs (i.e., those whose activity levels are above baseline) or to increase the activity of underactive MMPs (i.e., those whose activity levels are below baseline).
Roles of MMPs Associated to Bone Development and Remodeling
The biochemical actions of MMPs are intimately linked to their cells of origin. Table 4 describes cell-specific roles of MMPs in physiological bone remodeling. Osteoclast-mediated bone resorption in calvaria and long bones requires normal enzymatic activity of MMPs and cysteine proteinases such as cathepsin K whose deficiency impairs bone remodeling (Everts et al., 1999; Delaisse et al., 2003). This is evidenced in osteoclasts from patients with pycnodysostosis (an osteopetrosis-like bone disease related to loss-of-function mutations in the cathepsin K gene) and osteoclasts from cathepsin K-deficient mice which are unable to efficiently digest organic bone matrix, resulting in large, mineral-free areas of bone matrix (Everts et al., 1998, 2009). Cysteine proteinases synthesized and used by the different osteoclasts for bone matrix digestion (Everts et al., 2006) can degrade intramembranous bones as well as osteoclast-derived MMPs (Everts et al., 2009). Cysteine proteinases are secreted to act in the low pH environments formed by osteoclasts in the resorption sites, with MMPs degrading the rest of the bone matrix when the pH increases (Everts et al., 1998) as well as contributing to the digestion of fibrillar, non-mineralized collagen in Howship’s lacunae abandoned by osteoclast cells (Everts et al., 2002). These complementary and overlapping contributions of the MMP and cysteine proteinase families make the process of bone tissue remodeling both complex and robust.
TABLE 4.
Specific roles of MMPs under physiological conditions in bone remodeling.
| Entity | MMP | Role | References |
| Cartilage and bone cells | Network of multiple MMPs (mainly widely expressed MMP-2, -7, -9, -12, -13, -14, -16) | Maintain bone and cartilage health by their normal proteolytic activity. | Everts et al., 1992; Meikle et al., 1992; Mattot et al., 1995; Apte et al., 1997; Johansson et al., 1997; Bord et al., 1998; Jimenez et al., 1999; Filanti et al., 2000 |
| Control bone tissue remodeling at the levels of osteocyte viability and activities, osteoclast recruitment and function, bone matrix solubilization, coupling of bone resorption and formation, osteoblast recruitment and survival, cell-extracellular matrix interaction, and cell–cell interaction. | Blavier and Delaisse, 1995; Bord et al., 1998; Engsig et al., 2000; Hou et al., 2004; Inada et al., 2004; Karsdal et al., 2004; Holmbeck et al., 2005; Kasper et al., 2007; Manduca et al., 2009; Lu et al., 2010; Ortega et al., 2010; Tang et al., 2012; Madsen et al., 2013; Lozito et al., 2014; Almalki and Agrawal, 2016 | ||
| Regulate the bioavailability of soluble RANKL, thereby promoting the formation of multinucleated osteoclast cells, acquisition of osteoclast-specific differentiation markers, binding of osteoclasts to bone surfaces, promotion of osteoclast survival, and stimulation of bone resorption. | Bellido et al., 2019 | ||
| Mesenchymal stem cells | Network of multiple MMPs, tissue inhibitors of MMPs and RECK | (i) Modulates the commitment and differentiation of mesenchymal stem cells. (ii) Impacts osteoblastic migration, spreading, and differentiation. |
Kasper et al., 2007; Lu et al., 2010; Lozito and Tuan, 2011; Egea et al., 2012; Almalki and Agrawal, 2016; Mahl et al., 2016 |
| MMP-16 | Controls mesenchymal stem cells viability. | Paiva and Granjeiro, 2017 | |
| MMP-2 and MMP-9 | Promote the directional migration of bone marrow mesenchymal stem cells. | Lv et al., 2017 | |
| Osteocytes | MMP-2, MMP-13 and MMP-14 | Modulate the formation of the osteocyte canalicular network. | Barthelemi et al., 2012 |
| MMP-13 | Regulates the remodeling of the osteocyte lacunar-canalicular network in mid-cortical bone matrix, which is critical for the active maintenance of bone quality (matrix composition, organization, fracture resistance). | Tang et al., 2012; Alliston, 2014 | |
| MMP-14 | Essential for cell adhesion, invasion, and cell-cell communication events. | Hughes et al., 1994; Paiva and Granjeiro, 2017 | |
| Osteoclasts | MMP-9 | Participates in cell recruitment (by generating collagen-derived endostatin which prevents osteoclast chemotaxis), survival (e.g., by activating pro-TNF-α), adhesion (e.g., by cleaving intercellular adhesion molecule-1), as well as in degradation of cytokines important to osteoclastogenesis such as IL-1β. | Gearing et al., 1995; Ito et al., 1996; Ferreras et al., 2000; Fiore et al., 2002 |
| MMP-12 | Modulates the interaction between osteoclasts and bone matrix through multiple mechanisms including: (i) cleavage of osteopontin, vitronectin, bone sialoprotein and osteonectin, (ii) activation of TNF-α, (iii) generation of endostatin from collagen, and (iv) digestion of urokinase-type plasminogen activator receptor/uPAR. | Koolwijk et al., 2001; Hou et al., 2004; Paiva and Granjeiro, 2017 | |
| MMP-14 | Sheds CD14 receptor to impinge on osteoclast adhesion and migration as well as being involved in monocyte/macrophage fusion (e.g., by modulating the Rac1 pathway). | Kajita et al., 2001; Vivinus-Nebot et al., 2004; Gonzalo et al., 2010 | |
| The CD44/MMP-9/MMP-14 axis | Mediates pro-MMP-9 activation on the osteoclast membrane thereby modulating osteoclast migration in bone tissue resorption. | Chellaiah and Ma, 2013 | |
| MMP-14 and MMP-7 | Promote RANKL availability, which implicates the RANK/RANKL/osteoprotegerin axis in osteoclast maturation and activation. | Lynch et al., 2005; Hikita et al., 2006; Aiken and Khokha, 2010 | |
| Bone matrix | MMP-1, -2, -8, -9, -13, -14, and -15 | Necessary for extracellular matrix turnover. | Paiva and Granjeiro, 2017 |
| MMPs -2, -3, -7, -9, -12, -14 | Cleave and regulate bone matrix-associated non-collagenous proteins (such as osteonectin, vitronectin, osteopontin, bone sialoprotein) as well as cell membrane- and matrix-anchored latent growth factors. | Sasaki et al., 1997; Agnihotri et al., 2001; Sage et al., 2003; Lindsey et al., 2015 | |
| MMP-14 | The collagen fragments produced by MMP-14 are endocytosed via uPARAP/Endo180 for total lysosomal degradation. | Lafleur et al., 2006; Lee et al., 2006; Messaritou et al., 2009 | |
| Osteoblasts | MMP-2 | Critical for osteoblast differentiation and survival. | Paiva and Granjeiro, 2017 |
| MMP-14 | Serves to preserve osteoblast survival once osteoblasts have stopped the synthesis of new bone matrix, thus aiding in the transition from osteoblasts to osteocytes. | Karsdal et al., 2004 | |
| Bone remodeling | MMPs from osteoblasts and bone lining cells | Preceding osteoclast adhesion and resorption, MMPs participate in the cleavage of organic matrix (such as cathepsin-cleaved collagen and non-collagenous proteins). | Holliday et al., 1997; Stahle-Backdahl et al., 1997; Yamagiwa et al., 1999; Paiva and Granjeiro, 2017 |
| MMP-13 | Active in regulating bone mass through osteoblasts, and forming osteocyte canalicular network. | Page-McCaw et al., 2007; Barthelemi et al., 2012 | |
| MMP-14/CD44 | Activates Pro-MMP-9 on osteoclast membrane surface during osteoclast recruitment, adhesion, resorption and migration. | Paiva and Granjeiro, 2017 |
MMPs, matrix metalloproteinases; RECK, reversion-inducing cysteine-rich protein with Kazal motifs; TNF, tumor necrosis factor; IL, interleukin; Rac1, Ras-related C3 botulinum toxin substrate 1 pathway; RANK, receptor activator of nuclear factor kappa B; RANKL, RANK ligand; uPARAP/Endo180, endocytic collagen receptor of collagen and collagen fragments for degradation in the lysosomes.
The involvement of MMPs in bone remodeling has become clear with the aid of animal models such as MMP-deficient mice, which show a variety of bone abnormalities (Table 5). Impaired bone tissue remodeling in Mmp2–/– mice (Table 5, row 2) is characterized by a reduced number of osteoblasts and osteoclasts, disruption of the canicular network exacerbating osteocyte death, disruption of the MMP-2-osteopontin-bone sialoprotein axis, and promotion of osteolysis (Martignetti et al., 2001; Inoue et al., 2006; Mosig et al., 2007; Malaponte et al., 2016). MMP-9-deficient mice show alterations in cartilage-bone replacement during endochondral ossification (Vu et al., 1998) (Table 5, row 3). This phenotype may be explained by an inefficient degradation of the cartilage matrix, which leads to a diminished bioavailability of extracellular matrix-derived vascular endothelial growth factor and consequently effects osteoclasts and endothelial cells movement into the cartilage (Ortega et al., 2010). Bone tissue modeling and remodeling processes are altered in MMP-13 deficient mice (Table 5, row 4) (Inada et al., 2004; Stickens et al., 2004; Ortega et al., 2005). MMP-14 deficiency (Table 5, row 5), which is associated with high lethality, results in the most drastic skeletal phenotype among MMP-deficient mice (Holmbeck et al., 1999; Zhou et al., 2000). Double gene-deficient mice lacking at least one MMP gene have been engineered and their bone phenotype have been studied. For instance, double-knockout mice lacking MMP-2 and uPARAP/Endo180 (endocytic receptor of collagen and collagen fragments for degradation in the lysosomes) show reduced bone mineral density, short long bones, and poor trabecular bone quality (Madsen et al., 2013). MMP-8 and MMP-13 double-deficient mice have abnormal growth plate as well as augmented metaphyseal trabecular bone mineral density (Inada et al., 2001, 2002; Stickens et al., 2004). Double knockout mice lacking MMP-9 and MMP-13 exhibit expanded growth plates, disorganized hypertrophic chondrocyte zone, increased number of end-differentiated hypertrophic cells, and delayed formation of the bone marrow cavity (Kennedy et al., 2005; Paiva and Granjeiro, 2014). The bone phenotype of mice with a double knockout for MMP-14 and MMP-2 reassembles that of MMP-14-deficient mice (Oh et al., 2004). MMP-14 and MMP-16 double-knockout mice develop a bone phenotype that affects ossification (intramembranous and endochondral) and is characterized by severe irregularities, including (i) high mortality associated to developmental defects, (ii) noticeable craniofacial malformations such as cleft palate, thinner cranial vault bones, deficiently developed parietal, as well as frontal and nasal bones, (iii) altered growth plate, and (iv) cortical bone shortening (Paiva and Granjeiro, 2014). MMP-14 and uPARAP/Endo180 double-knockout mice die soon after birth (Wagenaar-Miller et al., 2007). As listed in Table 6, MMP activity contributes to numerous bone pathologies including arthritis, osteoporosis, osteonecrosis, periodontitis, sinonasal osteitis, degenerated lumbar disk tissues, and bone cancer metastasis (Aiken and Khokha, 2010; Koskinen et al., 2011; Mittal et al., 2016; Rose and Kooyman, 2016; Lazarus et al., 2017; Paiva and Granjeiro, 2017; Tauro and Lynch, 2018; Zhang et al., 2018). The roles played by MMPs in these pathologies are influenced by non-matrix proteins such as TIMPs, transforming growth factor, vascular endothelial growth factor, bone morphogenic proteins, activated protein C, and the Wnt [Wingless-type MMTV integration site family]/β-catenin (Table 7).
TABLE 5.
Selected skeletal phenotypes associated to MMP deficiency in mice.
| Genotype | Phenotype | References |
| Mmp2–/– | MMP-2 knockout (vs. wild-type) mice show: (i) craniofacial defects (such as shorter and broader snouts, hypertelorism, smaller jaws, dome-shaped and taller skulls), (ii) severe arthritis and joint contractures (even in young mice) with articular cartilage destruction and erosion of the underlying bone surface, (iii) joint pathology with increased cellular infiltration and proteoglycan depletion in antigen-induced arthritis, (iv) diminished bone integrity (such as long bones with osteopenia, fractured tibiae), (v) anomalous bone development (e.g., reduced number of long bones, decreased femur and tibia length in adult mice, calvarial bones with a greater [48%] thickness by 55 weeks of age, trabecular bone with fewer osteocytes), (vi) progressive decrease in bone mineral density and increase in bone porosity (characterized by e.g., low trabecular connectivity density, reduced mineral-collagen relation, thinner diaphyseal cortex, less nanoindentation modulus), (vii) increased number of empty lacunae as the mice aged (e.g., about 3-fold by 55 weeks of age), (viii) loss of the canalicular network architecture in calvariae and slighter in long bones, and (ix) presumably expression of bone sialoprotein (which increases osteoblast differentiation and activity) and osteopontin (which increases osteoclast activity). | Inoue et al., 2006; Mosig et al., 2007; Lieu et al., 2011; Nyman et al., 2011; Madsen et al., 2013 |
| Mmp9–/– | MMP-9 knockout (vs. wild-type) mice show: (i) long bones (e.g., metatarsals) with increased (e.g., 4-8-fold for 3 weeks old mice) hypertrophic (cartilage) zones, (ii) 10% shorter long bones, which is the only remaining phenotype in older MMP-9 deficient mice, (iii) irregularly shaped bone spicules, (iv) delayed endochondral ossification, (v) expanded zone of hypertrophic chondrocytes in the growth plate, (vi) reduced vascular invasion into the hypertrophic cartilage, (vii) slowed apoptosis of hypertrophic chondrocytes, (viii) impaired osteoclast/condroclast recruitment, (ix) anomalous growth in trabecular bone mass, and (x) improved connectivity density of the tibia trabeculae. This phenotype eventually resolve, resulting in correction of bone growth defects after approximately 4 weeks of age. | Vu et al., 1998; Ortega et al., 2003; Nyman et al., 2011; Kojima et al., 2013 |
| Mmp13–/– | Mmp13–/– (vs. Mmp13±) mice embryos show: (i) progressive changes in the embryonic growth plates (e.g., increased length which persisted in adults), (ii) delayed endochondral ossification, (iii) augmented metaphyseal trabecular bone mass as the mice aged (e.g., 3 months old), (iv) diminished resistance to fracture in long bones, (v) delay in fracture repair, (vi) defective vascular penetration and chondroclast attraction to the fracture callus, (vii) noticeable expression of collagen type X, osteopontin, and VEGF by hypertrophic chondrocytes. | Inada et al., 2001; Inada et al., 2002; Inada et al., 2004; Stickens et al., 2004; Kosaki et al., 2007; Tang et al., 2012; Singh et al., 2013 |
| Mmp14–/– | MMP-14 knockout (vs. wild-type) mice show: (i) progressive disturbances (e.g., smaller body size and weight, very high postnatal mortality), possibly caused by deprived feeding and therefore malnutrition, (ii) craniofacial dysmorphism in surviving mice (e.g., short snout, hypertelorism, dome-shaped skull, orbital protrusions, unclosed cranial sutures), (iii) incomplete cartilage remodeling, (iv) impaired formation of secondary ossification centers in the epiphyses, (v) ankylosis resulting from joints with arthritis and other factors (e.g., greater vascularity of the ligaments and tendons, overgrowth of hypercellular and wrongly vascularized synovial tissue), (vi) augmented bone resorption, (vii) osteopenia, (viii) osteoporosis, (ix) dwarfism, (x) mesenchymal stem cells commitment to chondrogenesis and adipogenesis instead of osteogenesis. | Holmbeck et al., 1999; Zhou et al., 2000; Holmbeck et al., 2003 |
| Mmp16–/– | MMP-16 knockout (vs. wild-type) mice show shorter size associated with reduced viability of mesenchymal cells in bone tissues. | Shi et al., 2008; Loffek et al., 2011 |
MMP, matrix metalloproteinase; VEGF, vascular endothelial growth factor.
TABLE 6.
Involvement of MMPs in bone pathologies.
| MMP | Reported involvement | References |
| MMP-1 | Abundant in the diaphysis and metaphyses of long bones being upregulated in arthritis. | Gack et al., 1995; Wu et al., 2008; Rose and Kooyman, 2016 |
| MMP-2 | Required for maintenance of bone mineral density and strength and in bone development (e.g., by affecting intramembranous and endochondral ossification); however, deregulated MMP-2 expression is observed in the settings of metabolic syndrome, osteoporosis, osteonecrosis of the jaws, ligamentum flavum degeneration in lumbar spinal canal stenosis, as well as in bone pre-metastatic niche formation. | Duerr et al., 2004; Suh et al., 2004; Tester et al., 2004; Durie et al., 2005; Lynch, 2011; Fernandez-Patron et al., 2016; Rose and Kooyman, 2016; Sugimoto et al., 2018 |
| MMP-3 | Overexpressed in osteoarthritis (in cartilage and the synovium) and also acts on primary tumor growth. | Okada et al., 1992; Tester et al., 2004; Lynch, 2011; Paiva and Granjeiro, 2017 |
| MMP-8 | Modulates human dentin and remodeling, but its deregulation may exacerbate periodontitis although it may be protective against inflammatory arthritis. | Sulkala et al., 2007; Cox et al., 2010; Mauramo et al., 2018 |
| MMP-9 | Participates in chondrocyte biology; specific processes in which the enzyme is involved are apoptosis of hypertrophic chondrocytes present in utero, bone development (e.g., by being highly active to angiogenesis in the growth plate), strength and toughness of bone, as well as the regulation of gene pathways responsible for osteoclastogenesis). In turn, MMP-9 overexpression contributes to sinonasal osteitis, rheumatoid arthritis, and degenerated lumbar disk tissues. | Vu et al., 1998; Liang et al., 2016; Mittal et al., 2016; Ahrens et al., 1996; Li et al., 2017 |
| Osteoporotic bone (vs. normal bone) tissues express higher MMP-9 levels. | Zhao et al., 1997 | |
| Involved in secondary (metastatic) breast cancer in the bone (e.g., by promoting angiogenesis, regulating VEGF bioavailability, contributing to bone remodeling) or prostate cancer (e.g., by influencing bone osteoblastic and osteoclastic activity). | Bergers et al., 2000; Mannello et al., 2005; Pego et al., 2018 | |
| MMP-13 | Required for bone development; it participates in the transition from cartilage to bone at the growth plates of long bones and in the remodeling of bone spicules. In turn, MMP-13-mediated degradation of articular cartilage exacerbates osteoarthritis. | Inada et al., 2004; Stickens et al., 2004; Page-McCaw et al., 2007; Holmbeck et al., 1999; Mittal et al., 2016; Rose and Kooyman, 2016 |
| In linking osteoarthritis to metabolic syndrome, the presence of adiponectin positively correlates with the presence of membrane-expressed PGE synthase and MMP-13. | Francin et al., 2014 | |
| Overexpressed in congenital spondyloepiphyseal dysplasia which results in early development of osteoarthritis. | Rose and Kooyman, 2016 | |
| In addition to typical bone collagen matrix degradation, MMP-13 regulates bone resorption in periodontal disease through osteoclast differentiation (by inactivating galectin-3, an inhibitor of osteoclastogenesis) and osteoclast activation (by activating osteoclast-secreted pro-MMP-9 and favoring RANKL and TGF-β1 signaling). | Nannuru et al., 2010; Pivetta et al., 2011; Cavalla et al., 2017 | |
| In breast cancer resulting from bone metastasis, MMP-13 deregulation may alter osteoblast morphology and bone resorption through differentiation of pre-osteoclasts, osteoclast activation, and osteolysis. | Stickens et al., 2004; Page-McCaw et al., 2007; Shah et al., 2012 | |
| MMP-14 | Contributes to bone development (endochondral and intramembranous ossification) and remodeling. Extracellular matrix remodeling by MMP-14 influences cell shape inducing the formation of a complex between MMP-14 and beta1-integrin, which activates the Rho/GTPase cascade leading to nuclear translocation of YAP and TAZ – this series of signaling events is necessary for mesenchymal stem cells commitment during development. Palmitoylation (i.e., addition of 16-carbon palmitate to proteins) enables MMP-14 to anchor to cell membrane. This post-translational modification of MMP-14 has a major impact on bone development and bone tissue metabolism likely through influencing MMP-14 correct membrane localization and also decreasing the expression of osteocalcin and vascular endothelial growth factor in osteoblasts and chondrocytes. In turn, MMP-14 is critical for osteoclast resorption thus contributing to the pathogenesis of osteoporosis. | Holmbeck et al., 1999, 2003; Zhou et al., 2000; Liao et al., 2004; Hienz et al., 2015; Tang et al., 2013; Paiva and Granjeiro, 2014; Song et al., 2014 |
| Involved in bone cancer metastasis acting alongside MMP-1 and MMP-11. | McGowan and Duffy, 2008; Rowe and Weiss, 2009; Paiva and Granjeiro, 2017 | |
| MMP-3 and MMP-9 | Contribute to cartilage endplate degeneration. | Zhang et al., 2018 |
| MMP-2, MMP-9, and MMP-13 | In experimental glucocorticoid-induced osteoporosis and osteocytic osteolysis, these three enzymes are upregulated in the trabecular bone of the metaphysis whereas MMP-2 and MMP-13 are expressed in the cortical bone diaphysis. | Sun et al., 2016 |
VEGF, vascular endothelial growth factor; PGE, prostaglandin E synthase; RANKL, receptor activator of NF-kappa B ligand; TGF transforming growth factor; MMP, matrix metalloproteinase; YAP, Yes-association protein; TAZ, transcriptional coactivator with PDZ-binding motif.
TABLE 7.
Interactions of MMPs with other proteins in bone development/remodeling.
| Protein | Effect on the partner | Effect on bone development/remodeling |
| TIMPs | Inhibit all MMPs | Control bone resorption and formation (Bord et al., 1999; Sobue et al., 2001; Huang et al., 2002; Geoffroy et al., 2004; Haeusler et al., 2005; Sahebjam et al., 2007; Shen et al., 2010; Miller et al., 2017). |
| MMP-2/MMP-9 | Control TGF-β (bioavailability and bioactivity) | Decrease the mechanical properties (modulus, hardness) of mice bones, when TGF-β signaling is augmented (Dallas et al., 1995; Balooch et al., 2005; Nyman et al., 2011) |
| MMP-9 | Regulates VEGF (bioavailability and bioactivity) | Exerts chemotactic action on osteoclasts, which affects osteoclast recruitment during bone resorption (Bergers et al., 2000; Engsig et al., 2000; Ortega et al., 2010). |
| MMP-14 | Activates TGF-β | Helps to preserve the survival of osteoblasts and their differentiation into osteocytes (Karsdal et al., 2002). |
| TGF-β | Upregulates MMP-13 | Promotes bone resorption associated to changes in osteoblast morphology (Karsdal et al., 2001). |
| BMPs | Regulates MMP-2 | Obstructs tissue remodeling and regeneration in Poecilia latipinna (Rajaram et al., 2016). |
| Regulates MMP-9 | Impairs bone remodeling (e.g., augmented bone mass during early development) and chondrocyte commitment (e.g., in the mouse C3H10T1/2 stem cell line) (Kamiya et al., 2008; Choi et al., 2009; Rajaram et al., 2016). | |
| Wnt/β-catenin | Regulates MMP-2 | Affects bone development (cartilage formation, endochondral ossification, growth plate organization, chondrocyte function) (Tamamura et al., 2005). |
| Upregulates MMP-9 | Modulates cartilage degradation and bone resorption (Tamamura et al., 2005). | |
| Regulates MMP-13 | Modulates cartilage vascularization (Tamamura et al., 2005; Nakashima and Tamura, 2006; Chen et al., 2008; Papathanasiou et al., 2012). | |
| aPC | Upregulates MMP-2 activity | Suppresses cartilage and bone degradation as well as pro-inflammatory signaling in rheumathoid arthritis patients (Nguyen et al., 2000; Buisson-Legendre et al., 2004; Xue et al., 2007). |
| Downregulates MMP-9 activity | Suppresses cartilage pro-inflammatory signaling as well as cartilage and bone degradation in rheumathoid arthritis patients (Xue et al., 2007; Xue et al., 2014). |
TIMP, tissue inhibitor of MMPs; MMP, matrix metalloproteinase; TGF-β, transforming growth factor; VEGF, vascular endothelial growth factor; BMP, bone morphogenic protein; aPC, activated protein C.
MMPs as Sheddases
Beyond the direct degradation of extracellular matrix substrates (e.g., collagen), MMP-mediated cleavage of substrates can lead to the release (shedding) into the extracellular matrix of soluble fragments of cell membrane-anchored receptor ligands. This extracellular event enables ligand-mediated activation of cognate receptors and elicits downstream intracellular signal transduction cascades which modify gene transcription and, ultimately, cell behavior. A prominent example pertinent to osteoblasts is the release of RANKL, which is the ligand of receptor activator of nuclear factor kappa B (RANK), by MMP-14. This MMP-14/RANKL/RANK/signal transduction axis regulates osteoblastogenesis and osteoclastogenesis, making MMP-14 crucial for normal bone formation (Bonfil et al., 2007; Thiolloy et al., 2009; Sabbota et al., 2010; Bonfil and Cher, 2011). The ligand shedding activity of MMPs influences the propensity to cancer metastasis and bone disease. For instance, MMP-14-mediated shedding of RANKL and downstream activation of RANK in the left supraclavicular lymph node cells of the prostate stimulates the non-receptor tyrosine kinase, SRC, to effectively increase the migration of prostate tumor cells which can metastasize to bone (Sabbota et al., 2010). Similarly, osteoclast-derived MMP-7 solubilizes osteoblast-bound RANKL whose release into the tumor-bone microenvironment promotes osteoclast activation in bone metastatic sites contributing to prostate and mammary tumor-induced osteolysis (Lynch et al., 2005; Thiolloy et al., 2009).
MMP-Generated Neoepitopes
The proteolytic action of MMPs on extracellular matrix macromolecules can result in the exposure of neo-epitopes (i.e., unique bioactive MMP-generated fragments). Compared to healthy subject controls, patients with ankylosing spondylitis (which is a form of arthritis that causes inflammation of the vertebrae) show significantly higher levels of different neo-epitopes such as C1M, C2M, C3M, C4M, C5M, C6M, and C7M from collagen type I, II, III, IV, V, VI, and VII (Veidal et al., 2012; Genovese and Karsdal, 2016). Some of these neo-epitopes have been combined (e.g., C2M, C3M, and C6M) for diagnostic purposes (Bay-Jensen et al., 2012). IPEN341-342FFGV is an MMP cleavage site which could be useful as diagnostic and prognostic makers for osteoarthritis (Bay-Jensen et al., 2011). Similarly, other MMP-generated neo-epitopes derived from collagen type II (e.g., C2C, C2M, C-terminal telopeptide of type II collagen (CTX-II), and TIINE) hold biomarker potential for osteoarthritis (Karsdal et al., 2010; Qvist et al., 2010; Karsdal et al., 2011).
Over-Overexpression of MMPs
Over-expression of MMPs is frequently reported in arthritis (Burrage et al., 2006; Tokito and Jougasaki, 2016). Collagenolytic MMPs (such as MMP-1, -2, -8, -13, and -14) are expressed in the arthritic joint and likely participate in the degradation of cartilage type II collagen, while MMP-3, -7, and -9 can degrade aggrecan leading to joint destruction (Puliti et al., 2012; Tokito and Jougasaki, 2016). Such a pathological mechanism has been proposed for MMP-3 and MMP-13 in degenerative joint disease in the elderly (Neuhold et al., 2001; Troeberg and Nagase, 2012; Jackson et al., 2014; Pap and Korb-Pap, 2015). Other contributions to osteoarthritis from activities related to MMP-3 include MMP-3-mediated activation of MMP-1 and MMP-13 (Mancini and di Battista, 2006; Tokito and Jougasaki, 2016). In rheumatoid arthritis, MMP-14 is greatly expressed in fibroblast-like synoviocytes and macrophages, and it could be an effector to cartilage destruction (Pap et al., 2000; Sabeh et al., 2010). MMP-1 and MMP-3 likely participate in cartilage destruction in rheumatoid arthritis and osteoarthritis (Burrage et al., 2006; Fiedorczyk et al., 2006; Tokito and Jougasaki, 2016). As a result, MMP overexpression could be therapeutically targeted in arthritis (Tokito and Jougasaki, 2016). Whether reducing MMP expression (or activity) levels provides a clinical benefit is unclear. In experimental models, many synthetic MMP inhibitors have shown positive effects (Ishikawa et al., 2005). At the clinical level, however, all efforts with MMP inhibitors to block the damaging activity of MMPs in arthritis and other non-neoplastic conditions were regrettably unsuccessful (Burrage et al., 2006; Tokito and Jougasaki, 2016). Reasons for these failures include: (i) deficient clinical trial designs (Burrage et al., 2006), (ii) unwanted characteristics of MMP inhibitors (side effects including musculoskeletal pain, low oral bioavailability, short in vivo half-lives, and lack of selectivity [Iyer et al., 2012; Fields, 2015; Tokito and Jougasaki, 2016]), (iii) inability of MMP inhibitors to infiltrate the cartilage/bone/synovial interface (Burrage et al., 2006), (iv) neglect of the highly complex functions served by MMPs in physiological and disease states (Iyer et al., 2012; Li et al., 2013; Sawicki, 2013) and (v) broad tissue distribution and substrate promiscuity exhibited by MMPs and their substrates (Burrage et al., 2006; Tokito and Jougasaki, 2016). To date, there remains a need for highly selective MMP inhibitors and for better information on the disease-specific substrates, which could be therapeutically targeted as shown by recent studies with MMP-13 in osteoarthritis (Li et al., 2011) as well as for more efficient and reliable techniques to sensitively measure condition-specific MMP activity potential (not just MMP expression levels).
MMP Gene Polymorphism
A nucleotide polymorphism, by which an additional guanine creates an ETS transcription factor binding site (5′-GGA-3′) at position 1607 in the promoter sequence of the MMP-1 gene, has been related to bone mineral density (BMD) (Rutter et al., 1998). This polymorphism is associated with increased transcription of the MMP-1 gene and elevated MMP-1 activity. Among 819 postmenopausal Japanese women, BMD (e.g., D50, D100) for the distal radius had a lower value in women with the GG/GG genotype (47.9%) than in those with other (e.g., G/GG [41.9%], G/G [10.3%], G/G + G/GG [52.1%]) genotypes. A -1562C3 thymine polymorphism in the MMP-9 gene has been related to BMD in a population-based study (1114 Japanese men and 1087 women). It seems that the T allele (e.g., in men with CT or TT genotypes) of MMP-9, which shows greater transcriptional activity than the C allele (e.g., in men with CC genotype), is linked to decreased bone mass, and has a predominant effect on BMD (Zhang et al., 1999; Yamada et al., 2004). A single nucleotide polymorphism rs17576 may be involved in the pathogenesis of lumbar disk herniation (Jing et al., 2018); while the G allele of rs17576 appears to correlate with more severe stages of disk degeneration.
MMP Deficiency and Insufficiency in Humans
Having discussed the roles of MMPs under physiological and pathological conditions, we will next discuss how their deficiency and insufficiency relates to bone metabolic abnormalities.
MMP-2 gene deficiency leads to a rare human skeletal disorder1, which was first reported in consanguineous Saudi Arabian families, and is characterized by severe bone alterations (Martignetti et al., 2001). Osteolytic and metabolic changes linked to MMP-2 deficiency affect tarsal, carpal, and phalangeal bones, cause severe arthropathy, osteoporosis, fibrous nodules, distinctive craniofacial defects such as exophthalmos, brachycephaly, and flattened nasal bridges and dwarfism (Al-Aqeel et al., 2000; Al-Mayouf et al., 2000; Al-Aqeel, 2005; Mosig et al., 2007; Page-McCaw et al., 2007; Castberg et al., 2013). This complex syndrome is currently categorized as a form of Torg syndrome and results from homoallelic mutations in the gene for MMP-2 located at 16q12-21 (Martignetti et al., 2001; Liang et al., 2016). A Tyr codon in the MMP-2 prodomain is replaced with the Y244X stop codon and an Arg is replaced with a His (R101H) in the cysteine-containing domain (PRCGNPD substituted by PHCGNPD). The R101H mutation is suggested to perturb coordination of Cys102 to the catalytic Zn(II) domain, consequently activating intracellular pro-MMP-2 and leading to its auto-degradation (Kennedy et al., 2005; Krane and Inada, 2008). A homoallelic missense mutation in the catalytic Zn(II) domain (E404K) has been revealed in Winchester syndrome (another variant of multicentric osteolysis) (Zankl et al., 2005). These rare Torg and Winchester arthritic syndromes together with others (such as multicentric osteolysis with nodulosis and arthropathy [known as MONA]) belong to a general family of hereditary autosomal dominant and recessive skeletal disorders with progressive bone loss and joint destruction (Al-Mayouf et al., 2000; Martignetti et al., 2001; Al-Aqeel, 2005; Zankl et al., 2005; Rouzier et al., 2006; Mosig et al., 2007; Tuysuz et al., 2009).
Similar to MMP-2, a homozygous dominant mutation (Ser substituted by Phe [F56S]) in the pro-region domain of MMP-13 also results in a bone development disorder known as spondyloepimetaphyseal dysplasia-Missouri type (Kennedy et al., 2005)2. This disorder, which appears to spontaneously resolve by adolescence, is characterized by anomalous modeling of long bones, mild defects in epiphysis, moderate to severe changes in the metaphysis morphology, pear-shaped vertebrae, femoral and tibial bowing, genu varum deformities, and osteoarthritis. While the biochemical mechanisms linking MMP-13 to these bone abnormalities remain unclear, the phenotype of MMP-13 deficiency could be due to a late exit of chondrocyte cells from the growth plate (Kennedy et al., 2005).
MMP-14 is widely considered one of the physiological activators of MMP-2 as it converts pro-MMP-2 into mature MMP-2 at the cell surface (Fernandez-Patron et al., 2016). An MMP-14 homoallelic mutation (T > R replacement in the signal peptide domain) destabilizes the interaction (e.g., recognition and binding) of the MMP-14 signal peptide with the signal recognition particle complex, thus affecting MMP-14 targeting to the plasma membrane (Evans et al., 2012). This MMP-14 homoallelic mutation causes an apparent deficiency of biochemically active MMP-14 at the cell membrane which impairs pro-MMP-2 activation and causes a condition of MMP-2 activity deficiency with Winchester syndrome (Evans et al., 2012)3.
A missense homozygous mutation (g.16250T > A, which replaces His226 of the Zn(II) catalytic domain with Gln [p.H226Q]), in the MMP20 gene disrupts the metal-binding site and prevents MMP-20 proteolytic activity regarding enamel matrix proteins (Ozdemir et al., 2005)4. This mutation may lead to autosomal-recessive hypomaturation amelogenesis imperfecta, a group of inherited heterogeneous diseases that alter enamel development (amount, composition, structure) in humans (Kim J.W. et al., 2005). Another mutation in the intron 6 splice acceptor (g.30561A > T) that causes this disease is specifically characterized by pigmented teeth with a mottled and rough surface (Kim J.W. et al., 2005).
Partial loss of MMP activity or impaired MMP secretion can lead to MMP activity insufficiency. A pervasive cause of MMP insufficiency can be medications with such MMP inhibitory actions including: (i) Statins (200 million prescriptions in the United States/year; 14 million prescriptions for lovastatin alone in 2014)5 which can cause myositis and rhabdomyolysis (Luan et al., 2003; Thompson et al., 2003). (ii) Doxycycline (7 million prescriptions in 2014)5 with side-effects including joint inflammation in humans and cardiac inflammation in mice (Berry et al., 2015). (iii) Therapeutic antibodies against MMPs and MMP inhibitor drugs for treating patients with rheumatoid arthritis, severely active Crohn’s disease, and cystic fibrosis6. If these antibodies reduce MMP activity below baseline levels, they would cause MMP insufficiency with unpredictable consequences. Pharmacological MMP-inhibitors in Phase 3 clinical trials conducted during 1997 and 1998 in patients with advanced cancers led to an as of yet poorly understood, very severe inflammatory musculoskeletal syndrome (Zucker et al., 2000; Coussens et al., 2002). Another common cause of MMP insufficiency could be the pathological elevation of endogenous MMP inhibitors (e.g., tissue inhibitors of MMPs, α-2-macroglobulin, RECK) (Mott et al., 2000; Oh et al., 2001; Nagase et al., 2006; Klein and Bischoff, 2011). In addition, there is fibrinogen, an acute phase reactant in arthritis, which our laboratory discovered recently to inhibit MMP-2 in a cohort of rheumatoid arthritis patients (Sarker et al., 2019).
Summary
In summary, bone lining cells, osteocytes, osteoclasts, reversal cells, and osteoblasts are responsible for constant bone tissue remodeling (Figure 2). The activation of this multicellular unit and the intense communication between the bone cells is tightly regulated by mechanical stimuli, apoptosis, as well as systemic and local factors such as hormones and cytokines including RANKL, CSF-M, IL-3, and IL-6. Proteases of the MMP and cysteine proteinase families converge in the modulation of bone remodeling. Whereas proteolytic activity has long been thought to be required for the degradation of bone tissue in osteoarthritis and osteoporosis, inactivating mutations in MMP genes can also lead to bone pathology including osteolysis and metabolic abnormalities such as delayed growth. Thus, there remains a need to rethink the role played by proteases in bone physiology and pathology. More specific information related to bone remodeling and presumed pathways by which proteases, in particular MMPs, contribute to bone tissue remodeling in health and disease is provided in previous excellent reviews (Kini and Nandeesh, 2012; Rauner et al., 2012; Hienz et al., 2015; Liang et al., 2016; Mittal et al., 2016; Franco et al., 2017; Paiva and Granjeiro, 2017; Tauro and Lynch, 2018; Plotkin and Bruzzaniti, 2019).
FIGURE 2.
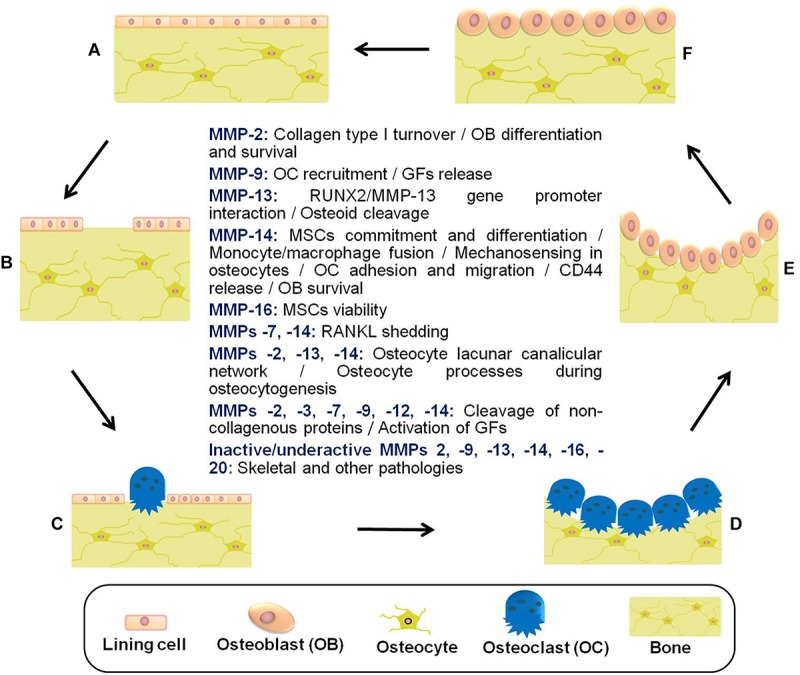
Schematic representation of the bone remodeling cycle with emphasis on the manifold roles played by matrix metalloproteinases. (A) Osteocytes detect mechanical stress or respond to biochemical stimuli. (B) Lining cells of the endosteal bone surface retract and proteases (e.g., MMPs) remove bone underlying membrane. (C) Osteoclasts are attracted and fused to become activated. (D) The underlying bone is digested by active multinucleated osteoclasts. (E) Osteoblasts are recruited to the bone resorption cavity. (F) New osteoid is formed by osteoblasts, and then mineralized (Datta et al., 2008; Fernandez-Patron et al., 2016; Paiva and Granjeiro, 2017; Cook et al., 2018). Other pathologies related to inactive/underactive MMPs are excessive inflammation, cardiovascular disorders, and metabolic dysregulation. MMP underactivity could also result from undesired side effects of common medications with MMP inhibitory actions (e.g., statins) (Cook et al., 2018). MSCs, mesenchymal stem cells; GFs, growth factors; RUNX2, runt-related transcription factor 2; RANKL, receptor activator of NF-kappa B ligand.
Author Contributions
EH and CF-P worked together on the conception, design, edition, revision, and approval of review manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank MSc. Angela Sosa for help with Figure 2 illustration.
Funding. CF-P was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant.
References
- Agnihotri R., Crawford H. C., Haro H., Matrisian L. M., Havrda M. C., Liaw L. (2001). Osteopontin, a novel substrate for matrix metalloproteinase-3 (Stromelysin-1) and matrix metalloproteinase-7 (Matrilysin). J. Biol. Chem. 276 28261–28267. 10.1074/jbc.m103608200 [DOI] [PubMed] [Google Scholar]
- Aguirre J. I., Plotkin L. I., Stewart S. A., Weinstein R. S., Parfitt A. M., Manolagas S. C., et al. (2006). Osteocyte apoptosis is induced by weightlessness in mice and precedes osteoclast recruitment and bone loss. J. Bone Miner. Res. 21 605–615. 10.1359/jbmr.060107 [DOI] [PubMed] [Google Scholar]
- Ahrens D., Koch A. E., Pope R. M., Stein-Picarella M., Niedbala M. J. (1996). Expression of matrix metalloproteinase 9 (96-kd gelatinase B) in human rheumatoid arthritis. Arthritis Rheum. 39 1576–1587. 10.1002/art.1780390919 [DOI] [PubMed] [Google Scholar]
- Aiken A., Khokha R. (2010). Unraveling metalloproteinase function in skeletal biology and disease using genetically altered mice. Biochim. Biophys. Acta 1803 121–132. 10.1016/j.bbamcr.2009.07.002 [DOI] [PubMed] [Google Scholar]
- Al-Aqeel A., Al Sewairi W., Edress B., Gorlin R. J., Desnick R. J., Martignetti J. A. (2000). Inherited multicentric osteolysis with arthritis: a variant resembling Torg syndrome in a Saudi family. Am. J. Med. Genet. 93 11–18. [DOI] [PubMed] [Google Scholar]
- Al-Aqeel A. I. (2005). Al-Aqeel Sewairi syndrome, a new autosomal recessive disorder with multicentric osteolysis, nodulosis and arthropathy. The first genetic defect of matrix metalloproteinase 2 gene. Saudi Med. J. 26 24–30. [PubMed] [Google Scholar]
- Alford A. I., Jacobs C. R., Donahue H. J. (2003). Oscillating fluid flow regulates gap junction communication in osteocytic MLO-Y4 cells by an ERK1/2 MAP kinase-dependent mechanism. Bone 33 64–70. 10.1016/s8756-3282(03)00167-4 [DOI] [PubMed] [Google Scholar]
- Alliston T. (2014). Biological regulation of bone quality. Curr. Osteoporos. Rep. 12 366–375. 10.1007/s11914-014-0213-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almalki S. G., Agrawal D. K. (2016). Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell. Res. Ther. 7 1–12. 10.1186/s13287-016-0393-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al-Mayouf S. M., Majeed M., Hugosson C., Bahabri S. (2000). New form of idiopathic osteolysis: nodulosis, arthropathy and osteolysis (NAO) syndrome. Am. J. Med. Genet. 93 5–10. [DOI] [PubMed] [Google Scholar]
- Andersen T. L., Abdelgawad M. E., Kristensen H. B., Hauge E. M., Rolighed L., Bollerslev J., et al. (2013). Understanding coupling between bone resorption and formation: are reversal cells the missing link? Am. J. Pathol. 183 235–246. 10.1016/j.ajpath.2013.03.006 [DOI] [PubMed] [Google Scholar]
- Apte S. S., Fukai N., Beier D. R., Olsen B. R. (1997). The matrix metalloproteinase-14 (MMP-14) gene is structurally distinct from other MMP genes and is co-expressed with the TIMP-2 gene during mouse embryogenesis. J. Biol. Chem. 272 25511–25517. 10.1074/jbc.272.41.25511 [DOI] [PubMed] [Google Scholar]
- Arpino V., Brock M., Gill S. E. (2015). The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. 4 247–254. 10.1016/j.matbio.2015.03.005 [DOI] [PubMed] [Google Scholar]
- Arumugam B., Vairamani M., Partridge N. C., Selvamurugan N. (2018). Characterization of Runx2 phosphorylation sites required for TGF-β1-mediated stimulation of matrix metalloproteinase-13 expression in osteoblastic cells. J. Cell Physiol. 233 1082–1094. 10.1002/jcp.25964 [DOI] [PubMed] [Google Scholar]
- Arumugam B., Vishal M., Shreya S., Malavika D., Rajpriya V., He Z., et al. (2019). Parathyroid hormone-stimulation of Runx2 during osteoblast differentiation via the regulation of lnc-SUPT3H-1:16 (RUNX2-AS1:32) and miR-6797-5p. Biochimie 158 43–52. 10.1016/j.biochi.2018.12.006 [DOI] [PubMed] [Google Scholar]
- Balemans W., Piters E., Cleiren E., Ai M., Van Wesenbeeck L., Warman M. L., et al. (2008). The binding between sclerostin and LRP5 is altered by DKK1 and by high-bone mass LRP5 mutations. Calcif. Tissue Int. 82 445–453. 10.1007/s00223-008-9130-9 [DOI] [PubMed] [Google Scholar]
- Balooch G., Balooch M., Nalla R. K., Schilling S., Filvaroff E. H., Marshall G. W., et al. (2005). TGF-beta regulates the mechanical properties and composition of bone matrix. Proc. Natl. Acad. Sci. U.S.A. 102 18813–18818. 10.1073/pnas.0507417102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baron R. (1989). Molecular mechanisms of bone resorption by the osteoclast. Anat. Rec. 224 317–324. 10.1002/ar.1092240220 [DOI] [PubMed] [Google Scholar]
- Bar-Shavit Z. (2007). Theosteoclast: amultinucleated, hematopoieticorigin, bone-resorbing osteoimmune cell. J. Cell Biochem. 102 1130–1139. 10.1002/jcb.21553 [DOI] [PubMed] [Google Scholar]
- Barthelemi S., Robinet J., Garnotel R., Antonicelli F., Schittly E., Hornebeck W., et al. (2012). Mechanical forces-induced human osteoblasts differentiation involves MMP-2/MMP-13/MT1-MMP proteolytic cascade. J. Cell Biochem. 113 760–772. 10.1002/jcb.23401 [DOI] [PubMed] [Google Scholar]
- Bay-Jensen A. C., Leeming D. J., Kleyer A., Veidal S. S., Schett G., Karsdal M. A. (2012). Ankylosing spondylitis is characterized by an increased turnover of several different metalloproteinase-derived collagen species: a cross-sectional study. Rheumatol. Int. 32 3565–3572. 10.1007/s00296-011-2237-8 [DOI] [PubMed] [Google Scholar]
- Bay-Jensen A. C., Liu Q., Byrjalsen I., Li Y., Wang J., Pedersen C., et al. (2011). Enzyme-linked immunosorbent assay (ELISAs) for metalloproteinase derived type II collagen neoepitope, CIIM–increased serum CIIM in subjects with severe radiographic osteoarthritis. Clin. Biochem. 44 423–429. 10.1016/j.clinbiochem.2011.01.001 [DOI] [PubMed] [Google Scholar]
- Baylink D. J., Finkelman R. D., Mohan S. (1993). Growth factors to stimulate bone formation. J. Bone. Miner. Res. 8 565–572. [DOI] [PubMed] [Google Scholar]
- Bellido T. (2014). Osteocyte-driven bone remodeling. Calcif. Tissue Int. 94 25–34. 10.1007/s00223-013-9774-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellido T., Gallant K. M. H. (2019). “Hormonal effects on bone cells,” in Basic and Applied Bone Biology, eds Burr D. B., Allen M. R., (Cambridge, MA: Academic Press; ), 299–314. 10.1016/b978-0-12-416015-6.00015-0 [DOI] [Google Scholar]
- Bellido T., Plotkin L. I., Bruzzaniti A. (2019). “Bone cells,” in Basic and Applied Bone Biology, eds Burr D. B., Allen M. R., (Cambridge, MA: Academic Press; ), 37–55. [Google Scholar]
- Bergers G., Brekken R., McMahon G., Vu T. H., Itoh T., Tamaki K., et al. (2000). Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell. Biol. 2 737–744. 10.1038/35036374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry E., Hernandez-Anzaldo S., Ghomashchi F., Lehner R., Murakami M., Gelb M. H., et al. (2015). Matrix metalloproteinase-2 negatively regulates cardiac secreted phospholipase A2 to modulate inflammation and fever. J. Am. Heart. Assoc. 4 1–22. 10.1161/JAHA.115.001868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair H. C., Teitelbaum S. L., Ghiselli R., Gluck S. (1989). Osteoclastic bone resorption by a polarized vacuolar proton pump. Science 245 855–857. 10.1126/science.2528207 [DOI] [PubMed] [Google Scholar]
- Blavier L., Delaisse J. M. (1995). Matrix metalloproteinases are obligatory for the migration of preosteoclasts to the developing marrow cavity of primitive long bones. J. Cell Sci. 108 3649–3659. [DOI] [PubMed] [Google Scholar]
- Bode W., Maskos K. (2003). Structural basis of the matrix metalloproteinases and their physiological inhibitors, the tissue inhibitors of metalloproteinases. Biol. Chem. 384 863–872. [DOI] [PubMed] [Google Scholar]
- Bodine P. V., Komm B. S. (2006). Wnt signaling and osteoblastogenesis. Rev. Endocr. Metab. Disord. 7 33–39. 10.1007/s11154-006-9002-4 [DOI] [PubMed] [Google Scholar]
- Bonewald L. F. (2011). The amazing osteocyte. J. Bone Miner. Res. 26 229–238. 10.1002/jbmr.320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonewald L. F., Johnson M. L. (2008). Osteocytes, mechanosensing and Wnt signaling. Bone 42 606–615. 10.1016/j.bone.2007.12.224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonewald L. F., Mundy G. R. (1990). Role of transforming growth factor-beta in bone remodeling. Clin. Orthop. Relat. Res. 250 261–276. [PubMed] [Google Scholar]
- Bonfil R. D., Cher M. L. (2011). The role of proteolytic enzymes in metastatic bone disease. IBMS BoneKEy 8 16–36. 10.1138/20110487 [DOI] [Google Scholar]
- Bonfil R. D., Dong Z., Trindade Filho J. C., Sabbota A., Osenkowski P., Nabha S., et al. (2007). Prostate cancer-associated membrane type 1-matrix metalloproteinase: a pivotal role in bone response and intraosseous tumor growth. Am. J. Pathol. 170 2100–2111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnans C., Chou J., Werb Z. (2014). Remodelling the extracellular matrix in development and disease. Nature Rev. Mol. Cell. Biol. 15 786–801. 10.1038/nrm3904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bord S., Horner A., Beeton C. A., Hembry R. M., Compston J. E. (1999). Tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) distribution in normal and pathological human bone. Bone 24 229–235. 10.1016/s8756-3282(98)00174-4 [DOI] [PubMed] [Google Scholar]
- Bord S., Horner A., Hembry R. M., Compston J. E. (1998). Stromelysin-1 (MMP-3) and stromelysin-2 (MMP-10) expression in developing human bone: potential roles in skeletal development. Bone 23 7–12. 10.1016/s8756-3282(98)00064-7 [DOI] [PubMed] [Google Scholar]
- Bord S., Horner A., Hembry R. M., Reynolds J. J., Compston J. E. (1996). Production of collagenase by human osteoblasts and osteoclasts in vivo. Bone 19 35–40. 10.1016/8756-3282(96)00106-8 [DOI] [PubMed] [Google Scholar]
- Boyden L. M., Mao J., Belsky J., Mitzner L., Farhi A., Mitnick M. A., et al. (2002). High bone density due to a mutation in LDL receptor related protein 5. N. Engl. J. Med. 346 1513–1521. 10.1056/nejmoa013444 [DOI] [PubMed] [Google Scholar]
- Boyle W. J., Simonet W. S., Lacey D. L. (2003). Osteoclast differentiation and activation. Nature 423 337–342. 10.1038/nature01658 [DOI] [PubMed] [Google Scholar]
- Brandi M. L., Piscitelli P. (2013). “Epidemiology of osteoporosis and fragility fractures,” in Osteoporosis and Bone Densitometry Measurements, ed. Guglielmi G., (Berlin: Springer-Verlag; ), 1–4. 10.1007/174_2012_747 [DOI] [Google Scholar]
- Brew K., Dinakarpandian D., Nagase H. (2000). Tissue inhibitors of metalloproteinases: evolution, structure and function. Biochim. Biophys. Acta 1477 267–283. 10.1016/s0167-4838(99)00279-4 [DOI] [PubMed] [Google Scholar]
- Bruzzaniti A., Baron R. (2006). Molecular regulation of osteoclast activity. Rev. Endocr. Metab. Disord. 7 123–139. 10.1007/s11154-006-9009-x [DOI] [PubMed] [Google Scholar]
- Buisson-Legendre N., Smith S., March L., Jackson C. (2004). Elevation of activated protein C in synovial joints in rheumatoid arthritis and its correlation with matrix metalloproteinase 2. Arthritis Rheum. 50 2151–2156. 10.1002/art.20313 [DOI] [PubMed] [Google Scholar]
- Burger E. H., Klein-Nulend J. (1999). Mechanotransduction in bone-role of the laculocanalicular network. FASEB 13 S101–S112. [PubMed] [Google Scholar]
- Burrage P. S., Mix K. S., Brinckerhoff C. E. (2006). Matrix metalloproteinases: role in arthritis. Front. Biosci. 11:529–543. [DOI] [PubMed] [Google Scholar]
- Canalis E. (2009). Growth factor control of bone mass. J. Cell Biochem. 108 769–777. 10.1002/jcb.22322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canalis E., Economides A. N., Gazzerro E. (2003). Bone morphogenetic proteins, their antagonists, and the skeleton. Endocr. Rev. 24 218–235. 10.1210/er.2002-0023 [DOI] [PubMed] [Google Scholar]
- Castberg F. C., Kjaergaard S., Mosig R. A., Lobl M., Martignetti C., Martignetti J. A., et al. (2013). Multicentric osteolysis with nodulosis and arthropathy (MONA) with cardiac malformation, mimicking polyarticular juvenile idiopathic arthritis: case report and literature review. Eur. J. Pediatr. 172 1657–1663. 10.1007/s00431-013-2102-8 [DOI] [PubMed] [Google Scholar]
- Cavalla F., Hernandez-Rios P., Sorsa T., Biguetti C., Hernandez M. (2017). Matrix metalloproteinases as regulators of periodontal inflammation. Int. J. Mol. Sci. 18:440. 10.3390/ijms18020440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charles J. M., Key L. L. (1998). Developmental spectrum of children with congenital osteopetrosis. J. Pediatr. 132 371–374. 10.1016/s0022-3476(98)70467-6 [DOI] [PubMed] [Google Scholar]
- Chellaiah M. A., Ma T. (2013). Membrane localization of membrane type 1 matrix metalloproteinase by CD44 regulates the activation of pro-matrix metalloproteinase 9 in osteoclasts. Biomed. Res. Int. 2013:302392. 10.1155/2013/302392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen D., Xie R., Shu B., Landay A. L., Wei C., Reiser J., et al. (2019). Wnt signaling in bone, kidney, intestine, and adipose tissue and interorgan interaction in aging. Ann. N. Y. Acad. Sci. 1442 48–60. 10.1111/nyas.13945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen M., Zhu M., Awad H., Li T. F., Sheu T. J., Boyce B. F., et al. (2008). Inhibition of beta-catenin signaling causes defects in postnatal cartilage development. J. Cell Sci. 121 1455–1465. 10.1242/jcs.020362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Wang L., Zhao K., Wang H. (2018). Osteocytogenesis: roles of physicochemical factors, collagen cleavage, and exogenous molecules. Tissue Eng. Part B Rev. 24 215–225. 10.1089/ten.teb.2017.0378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi Y. A., Kang S. S., Jin E. J. (2009). BMP-2 treatment of C3H10T1/2 mesenchymal cells blocks MMP-9 activity during chondrocyte commitment. Cell Biol. Int. 33 887–892. 10.1016/j.cellbi.2009.04.020 [DOI] [PubMed] [Google Scholar]
- Civitelli R. (2008). Cell-cell communication in the osteoblast/osteocyte lineage. Arch. Biochem. Biophys. 473 188–192. 10.1016/j.abb.2008.04.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark I. M., Swingler T. E., Sampieri C. L., Edwards D. R. (2008). The regulation of matrix metalloproteinases and their inhibitors. Int. J. Biochem. Cell Biol. 40 1362–1378. 10.1016/j.biocel.2007.12.006 [DOI] [PubMed] [Google Scholar]
- Coen G., Ballanti P., Silvestrini G., Mantella D., Manni M., Di Giulio S., et al. (2009). Immunohistochemical localization and mRNA expression of matrix Glaprotein and fetuin-A in bone biopsies of hemodialysis patients. Virchows Arch. 454 263–271. 10.1007/s00428-008-0724-4 [DOI] [PubMed] [Google Scholar]
- Cohick W. S., Clemmons D. R. (1993). The insulin-like growth factors. Annu. Rev. Physiol. 55 131–153. [DOI] [PubMed] [Google Scholar]
- Compston J. E. (2001). Sex steroids and bone. Physiol. Rev. 81 419–447. [DOI] [PubMed] [Google Scholar]
- Cook R., Sarker H., Fernandez-Patron C. (2018). Pathologies of matrix metalloproteinase-2 underactivity: A perspective on a neglected condition. Can. J. Physiol. Pharmacol. 97 1–7. 10.1139/cjpp-2018-0525 [DOI] [PubMed] [Google Scholar]
- Coussens L. M., Fingleton B., Matrisian L. M. (2002). Matrix metalloproteinase inhibitors and cancer: trials and tribulations. Science 295 2387–2392. 10.1126/science.1067100 [DOI] [PubMed] [Google Scholar]
- Cox J. H., Starr A. E., Kappelhoff R., Yan R., Roberts C. R., Overall C. M. (2010). Matrix metalloproteinase 8 deficiency in mice exacerbates inflammatory arthritis through delayed neutrophil apoptosis and reduced caspase 11 expression. Arthritis Rheum. 62 3645–3655. 10.1002/art.27757 [DOI] [PubMed] [Google Scholar]
- Crotti T. N., Flannery M., Walsh N. C., Fleming J. D., Goldring S. R., McHugh K. P. (2006). NFATc1 regulation of the human beta3 integrin promoter in osteoclast differentiation. Gene 372 92–102. 10.1016/j.gene.2005.12.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crotti T. N., Sharma S. M., Fleming J. D., Flannery M. R., Ostrowski M. C., Goldring S. R., et al. (2008). PU.1 and NFATc1 mediate osteoclastic induction of the mouse β3 integrin promoter. J. Cell Physiol. 215 636–644. 10.1002/jcp.21344 [DOI] [PubMed] [Google Scholar]
- Cui N., Hu M., Khalil R. A. (2017). Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 147 1–73. 10.1016/bs.pmbts.2017.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai J. C., He P., Chen X., Greenfield E. M. (2006). TNF alpha and PTH utilize distinct mechanisms to induce IL-6 and RANKL expression with markedly different kinetics. Bone 38 509–520. 10.1016/j.bone.2005.10.007 [DOI] [PubMed] [Google Scholar]
- Dallas S. L., Miyazono K., Skerry T. M., Mundy G. R., Bonewald L. F. (1995). Dual role for the latent transforming growth factor-beta binding protein in storage of latent TGF-beta in the extracellular matrix and as a structural matrix protein. J. Cell. Biol. 131 539–549. 10.1083/jcb.131.2.539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallas S. L., Prideaux M., Bonewald L. F. (2013). The osteocyte: an endocrine cell and more. Endocr. Rev. 34 658–690. 10.1210/er.2012-1026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Angelo M., Billings P. C., Pacifici M., Leboy P. S., Kirsch T. (2001). Authentic matrix vesicles contain active metalloproteases (MMP): a role for matrix vesicle-associated MMP-13 in activation of transforming growth factor-beta. J. Biol. Chem. 276 11347–11353. 10.1074/jbc.m009725200 [DOI] [PubMed] [Google Scholar]
- Datta H. K., Ng W. F., Walker J. A., Tuck S. P., Varanasi S. S. (2008). The cell biology of bone metabolism. J. Clin. Pathol. 61 577–587. 10.1136/jcp.2007.048868 [DOI] [PubMed] [Google Scholar]
- Delaisse J. M. (2014). The reversal phase of the bone-remodeling cycle: cellular prerequisites for coupling resorption and formation. Bonekey Rep. 3:561. 10.1038/bonekey.2014.56 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delaisse J. M., Andersen T. L., Engsig M. T., Henriksen K., Troen T., Blavier L. (2003). Matrix metalloproteinases (MMP) and cathepsin K contribute differently to osteoclastic activities. Microsc. Res. Tech. 61 504–513. 10.1002/jemt.10374 [DOI] [PubMed] [Google Scholar]
- Delmas P. D. (1995). “Biochemical markers for the assessment of bone turnover,” in Osteoporosis: Etiology, Diagnosis, and Management, 2nd Edn, eds Riggs B., Melton L. J., (Philadelphia: Lippincott-Raven; ), 319–333. [Google Scholar]
- Dempster D. W., Cosman F., Parisien M., Shen V., Lindsay R. (1993). Anabolic actions of parathyroid hormone on bone. Endocr. Rev. 14 690–709. 10.1210/er.14.6.690 [DOI] [PubMed] [Google Scholar]
- Ducy P., Schinke T., Karsenty G. (2000). The osteoblast: a sophisticated fibroblast under central surveillance. Science 289 1501–1504. 10.1126/science.289.5484.1501 [DOI] [PubMed] [Google Scholar]
- Duerr S., Stremme S., Soeder S., Bau B., Aigner T. (2004). MMP-2/gelatinase A is a gene product of human adult articular chondrocytes and is increased in osteoarthritic cartilage. Clin. Exp. Rheumatol. 22 603–608. [PubMed] [Google Scholar]
- Durie B. G. M., Katz M., Crowley J. (2005). Osteonecrosis of the jaw and bisphosphonates. N. Engl. J. Med. 353 99–102. [DOI] [PubMed] [Google Scholar]
- Egea V., Zahler S., Rieth N., Neth P., Popp T., Kehe K., et al. (2012). Tissue inhibitor of metalloproteinase-1 (TIMP-1) regulates mesenchymal stem cells through let-7f microRNA and Wnt/β-catenin signaling. Proc. Natl. Acad. Sci. U.S.A. 109 E309–E316. 10.1073/pnas.1115083109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eijken M., Hewison M., Cooper M. S., de Jong F. H., Chiba H., Stewart P. M., et al. (2005). 11beta-Hydroxysteroid dehydrogenase expression and glucocorticoid synthesis are directed by a molecular switch during osteoblast differentiation. Mol. Endocrinol. 19 621–631. 10.1210/me.2004-0212 [DOI] [PubMed] [Google Scholar]
- Engsig M. T., Chen Q. J., Vu T. H., Pedersen A. C., Therkidsen B., Lund L. R., et al. (2000). Matrix metalloproteinase 9 and vascular endothelial growth factor are essential for osteoclast recruitment into developing long bones. J. Cell Biol. 151 879–889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans B. R., Mosig R. A., Lobl M., Martignetti C. R., Camacho C., Grum-Tokars V., et al. (2012). Mutation of membrane type-1 metalloproteinase, MT1-MMP, causes the multicentric osteolysis and arthritis disease winchester syndrome. Am. J. Hum. Genet. 91 572–576. 10.1016/j.ajhg.2012.07.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everts V., de Vries T. J., Helfrich M. H. (2009). Osteoclast heterogeneity: lessons from osteopetrosis and inflammatory conditions. Biochim. Biophys. Acta 1792 757–765. 10.1016/j.bbadis.2009.05.004 [DOI] [PubMed] [Google Scholar]
- Everts V., Delaissé J. M., Korper W., Beertsen W. (1998). Cysteine proteinases and matrix metalloproteinases play distinct roles in the subosteoclastic resorption zone. J. Bone Miner. Res. 13 1420–1430. 10.1359/jbmr.1998.13.9.1420 [DOI] [PubMed] [Google Scholar]
- Everts V., Delaissé J. M., Korper W., Jansen D. C., Tigchelaar-Gutter W., Saftig P., et al. (2002). The bone lining cell: its role in cleaning Howship’s lacunae and initiating bone formation. J. Bone Miner. Res. 17 77–90. 10.1359/jbmr.2002.17.1.77 [DOI] [PubMed] [Google Scholar]
- Everts V., Delaisse J. M., Korper W., Niehof A., Vaes G., Beertsen W. (1992). Degradation of collagen in the bone-resorbing compartment underlying the osteoclast involves both cysteine-proteinases and matrix metalloproteinases. J. Cell Physiol. 150 221–231. 10.1002/jcp.1041500202 [DOI] [PubMed] [Google Scholar]
- Everts V., Korper W., Hoeben K. A., Jansen I. D., Bromme D., Cleutjens K. B., et al. (2006). Osteoclastic bone degradation and the role of different cysteine proteinases and matrix metalloproteinases: differences between calvaria and long bone. J. Bone Miner. Res. 21 1399–1408. 10.1359/jbmr.060614 [DOI] [PubMed] [Google Scholar]
- Everts V., Korper W., Jansen D. C., Steinfort J., Lammerse I., Heera S., et al. (1999). Functional heterogeneity of osteoclasts: matrix metalloproteinases participate in osteoclastic resorption of calvarial bone but not in resorption of long bone. FASEB J. 1999 1219–1230. 10.1096/fasebj.13.10.1219 [DOI] [PubMed] [Google Scholar]
- Faccio R., Teitelbaum S. L., Fujikawa K., Chappel J., Zallone A., Tybulewicz V. L., et al. (2005). Vav3 regulates osteoclast function and bone mass. Nat. Med. 11 284–290. 10.1038/nm1194 [DOI] [PubMed] [Google Scholar]
- Fanjul-Fernandez M., Folgueras A. R., Cabrera S., Lopez-Otin C. (2010). Matrix metalloproteinases: evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. 1803 3–19. 10.1016/j.bbamcr.2009.07.004 [DOI] [PubMed] [Google Scholar]
- Feng J. Q., Ward L. M., Liu S., Lu Y., Xie Y., Yuan B., et al. (2006). Loss of DMP1 causes rickets and osteomalacia and identifies a role for osteocytes in mineral metabolism. Nat. Genet. 38 1310–1315. 10.1038/ng1905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Patron C., Kassiri Z., Leung D. (2016). Modulation of systemic metabolism by MMP-2: from MMP-2 deficiency in mice to MMP-2 deficiency in patients. Compr. Physiol. 6 1935–1949. 10.1002/cphy.c160010 [DOI] [PubMed] [Google Scholar]
- Ferreras M., Felbor U., Lenhard T., Olsen B. R., Delaisse J. M. (2000). Generation and degradation of human endostatin proteins by various proteinases. FEBS Lett. 486 247–251. 10.1016/s0014-5793(00)02249-3 [DOI] [PubMed] [Google Scholar]
- Fiedorczyk M., Klimiuk P. A., Sierakowski S., Gindzienska-Sieskiewicz E., Chwiecko J. (2006). Serum matrix metalloproteinases and tissue inhibitors of metalloproteinases in patients with early rheumatoid arthritis. J. Rheumatol. 33 1523–1529. [PubMed] [Google Scholar]
- Fields G. B. (2015). New strategies for targeting matrix metalloproteinases. Matrix Biol. 4 239–246. 10.1016/j.matbio.2015.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filanti C., Dickson G. R., Di Martino D., Ulivi V., Sanguineti C., Romano P., et al. (2000). The expression of metalloproteinase-2, -9, and -14 and of tissue inhibitors-1 and -2 is developmentally modulated during osteogenesis in vitro, the mature osteoblastic phenotype expressing metalloproteinase-14. J. Bone Miner. Res. 15 2154–2168. 10.1359/jbmr.2000.15.11.2154 [DOI] [PubMed] [Google Scholar]
- Fiore E., Fusco C., Romero P., Stamenkovic I. (2002). Matrix metalloproteinase 9 (MMP-9/gelatinase B) proteolytically cleaves ICAM-1 and participates in tumor cell resistance to natural killer cell-mediated cytotoxicity. Oncogene 21 5213–5223. 10.1038/sj.onc.1205684 [DOI] [PubMed] [Google Scholar]
- Fraher L. (1993). Biochemical markers of bone turnover. Clin. Biochem. 26 431–432. [DOI] [PubMed] [Google Scholar]
- Francin P. J., Abot A., Guillaume C., Moulin D., Bianchi A., Gegout-Pottie P., et al. (2014). Association between adiponectin and cartilage degradation in human osteoarthritis. Osteoarthr. Cartil. 22 519–526. 10.1016/j.joca.2014.01.002 [DOI] [PubMed] [Google Scholar]
- Franco C., Patricia H. R., Timo S., Claudia B., Marcela H. (2017). Matrix metalloproteinases as regulators of periodontal inflammation. Int. J. Mol. Sci. 18:E440. 10.3390/ijms18020440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gack S., Vallon R., Schmidt J., Grigoriadis A., Tuckermann J., Schenkel J., et al. (1995). Expression of interstitial collagenase during skeletal development of the mouse is restricted to osteoblast-like cells and hypertrophic chondrocytes. Cell Growth Differ. 6 759–767. [PubMed] [Google Scholar]
- Gallagher S. K. (1997). Biochemical markers of bone metabolism as they relate to osteoporosis. MLO 29:50. [Google Scholar]
- Gao Y. Y., Liu B. Q., Du Z. X., Zhang H. Y., Niu X. F., Wang H. Q. (2010). Implication of oxygen regulated protein 150 (ORP150) in apoptosis induced by proteasome inhibitors in human thyroid cancer cells. J. Clin. Endocrinol. Metab. 95 E319–E326. 10.1210/jc.2010-1043 [DOI] [PubMed] [Google Scholar]
- Gearing A. J. H., Beckett P., Christodoulou M., Churchill M., Clements J. M., Crimmin M., et al. (1995). Matrix metalloproteinases and processing of pro-TNF-alpha. J. Leukoc. Biol. 57 774–777. [DOI] [PubMed] [Google Scholar]
- Gelb B. D., Shi G. P., Chapman H. A., Desnick R. J. (1996). Pycnodysostosis, a lysosomal disease caused by cathepsin K deficiency. Science 273 1236–1238. 10.1126/science.273.5279.1236 [DOI] [PubMed] [Google Scholar]
- Genovese F., Karsdal M. A. (2016). Protein degradation fragments as diagnostic and prognostic biomarkers of connective tissue diseases: understanding the extracellular matrix message and implication for current and future serological biomarkers. Expert Rev. Proteomics 13 213–225. 10.1586/14789450.2016.1134327 [DOI] [PubMed] [Google Scholar]
- Geoffroy V., Marty-Morieux C., Le Goupil N., Clement-Lacroix P., Terraz C., Frain M., et al. (2004). In vivo inhibition of osteoblastic metalloproteinases leads to increased trabecular bone mass. J. Bone Miner. Res. 19 811–822. 10.1359/jbmr.040119 [DOI] [PubMed] [Google Scholar]
- Gong Y., Slee R. B., Fukai N., Rawadi G., Roman-Roman S., Reginato A. M., et al. (2001). LDL receptor-related protein 5 (LRP5) affects bone accrual and eye development. Cell 107 513–523. [DOI] [PubMed] [Google Scholar]
- Gonzalo P., Guadamillas M. C., Hernandez-Riquer M. V., Pollan A., Grande-García A., Bartolome R. A., et al. (2010). MT1-MMP is required for myeloid cell fusion via regulation of Rac1 signaling. Dev. Cell 18 77–89. 10.1016/j.devcel.2009.11.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gori F., Hofbauer L. C., Dunstan C. R., Spelsberg T. C., Khosla S., Riggs B. L. (2000). The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology 141 4768–4776. 10.1210/endo.141.12.7840 [DOI] [PubMed] [Google Scholar]
- Greenlee K. J., Werb Z., Kheradmand F. (2007). Matrix metalloproteinases in lung: multiple, multifarious, and multifaceted. Physiol. Rev. 87 69–98. 10.1152/physrev.00022.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Y., Preston M. R., El Haj A. J., Howl J. D., Publicover S. J. (2001). Three types of K+ currents in murine osteocyte-like cells (MLO-Y4). Bone 28 29–37. 10.1016/s8756-3282(00)00439-7 [DOI] [PubMed] [Google Scholar]
- Guan C. C., Yan M., Jiang X. Q., Zhang P., Zhang X. L., Li J., et al. (2009). Sonic hedgehog alleviates the inhibitory effects of high glucose on the osteoblastic differentiation of bone marrow stromal cells. Bone 45 1146–1152. 10.1016/j.bone.2009.08.009 [DOI] [PubMed] [Google Scholar]
- Hachemi Y., Rapp A. E., Picke A. K., Weidinger G., Ignatius A., Tuckermann J. (2018). Molecular mechanisms of glucocorticoids on skeleton and bone regeneration after fracture. J. Mol. Endocrinol. 61 R75–R90. 10.1530/JME-18-0024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadler-Olsen E., Fadnes B., Sylte I., Uhlin-Hansen L., Winberg J. O. (2011). Regulation of matrix metalloproteinase activity in health and disease. FEBS J. 278 28–45. 10.1111/j.1742-4658.2010.07920.x [DOI] [PubMed] [Google Scholar]
- Haeusler G., Walter I., Helmreich M., Egerbacher M. (2005). Localization of matrix metalloproteinases, (MMPs) their tissue inhibitors, and vascular endothelial growth factor (VEGF) in growth plates of children and adolescents indicates a role for MMPs in human postnatal growth and skeletal maturation. Calcif. Tissue Int. 76 326–335. 10.1007/s00223-004-0161-6 [DOI] [PubMed] [Google Scholar]
- Han X., Boyd P. J., Colgan S., Madri J. A., Haas T. L. (2003). Transcriptional up-regulation of endothelial cell matrix metalloproteinase-2 in response to extracellular cues involves GATA-2. J. Biol. Chem. 278 47785–47791. 10.1074/jbc.m309482200 [DOI] [PubMed] [Google Scholar]
- Harris S. E., Guo D., Harris M. A., Krishnaswamy A., Lichtler A. (2003). Transcriptional regulation of BMP-2 activated genes in osteoblasts using gene expression microarray analysis: role of Dlx2 and Dlx5 transcription factors. Front. Biosci. 8:s1249–s1265. [DOI] [PubMed] [Google Scholar]
- Heino T. J., Hentunen T. A., Vaananen H. K. (2002). Osteocytes inhibit osteoclastic bone resorption through transforming growth factor-beta: enhancement by estrogen. J. Cell. Biochem. 85 185–197. 10.1002/jcb.10109 [DOI] [PubMed] [Google Scholar]
- Hienz S. A., Paliwal S., Ivanovski S. (2015). Mechanisms of bone resorption in periodontitis. J. Immunol. Res. 2015:615486. 10.1155/2015/615486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hikita A., Yana I., Wakeyama H., Nakamura M., Kadono Y., Oshima Y., et al. (2006). Negative regulation of osteoclastogenesis by ectodomain shedding of receptor activator of NF-kappa B ligand. J. Biol. Chem. 281 36846–36855. 10.1074/jbc.m606656200 [DOI] [PubMed] [Google Scholar]
- Hirao M., Hashimoto J., Yamasaki N., Ando W., Tsuboi H., Myoui A., et al. (2007). Oxygen tension is an important mediator of the transformation of osteoblasts to osteocytes. J. Bone Miner. Metab. 25 266–276. 10.1007/s00774-007-0765-9 [DOI] [PubMed] [Google Scholar]
- Holliday L. S., Welgus H. G., Fliszar C. J., Veith G. M., Jeffrey J. J., Gluck S. L. (1997). Initiation of osteoclast bone resorption by interstitial collagenase. J. Biol. Chem. 272 22053–22058. 10.1074/jbc.272.35.22053 [DOI] [PubMed] [Google Scholar]
- Holmbeck K., Bianco P., Caterina J., Yamada S., Kromer M., Kuznetsov S. A., et al. (1999). MT1-MMP-deficient mice develop dwarfism, osteopenia, arthritis, and connective tissue disease due to inadequate collagen turnover. Cell 99 81–92. 10.1016/s0092-8674(00)80064-1 [DOI] [PubMed] [Google Scholar]
- Holmbeck K., Bianco P., Chrysovergis K., Yamada S., Birkedal-Hansen H. J. (2003). MT1-MMP-dependent, apoptotic remodeling of unmineralized cartilage: a critical process in skeletal growth. Cell Biol. 163 661–671. 10.1083/jcb.200307061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmbeck K., Bianco P., Pidoux I., Inoue S., Billinghurst R. C., Wu W., et al. (2005). The metalloproteinase MT1-MMP is required for normal development and maintenance of osteocyte processes in bone. J. Cell Sci. 118 147–156. 10.1242/jcs.01581 [DOI] [PubMed] [Google Scholar]
- Holmbeck K., Szabov L. (2006). Aspects of extracellular matrix remodeling in development and disease. Birth Defects Res. C Embryo. Today 78 11–23. 10.1002/bdrc.20064 [DOI] [PubMed] [Google Scholar]
- Holt I., Davie M. W., Marshall M. J. (1996). Osteoclasts are not the main source of interleukin-6 in mouse parietal bone. Bone 18 221–226. 10.1016/8756-3282(95)00482-3 [DOI] [PubMed] [Google Scholar]
- Hou P., Troen T., Ovejero M. C., Kirkegaard T., Andersen T. L., Byrjalsen I., et al. (2004). Matrix metalloproteinase-12 (MMP-12) in osteoclasts: new lesson on the involvement of MMPs in bone resorption. Bone 34 37–47. 10.1016/j.bone.2003.08.011 [DOI] [PubMed] [Google Scholar]
- Huang W., Li W. Q., Dehnade F., Zafarullah M. (2002). Tissue inhibitor of metalloproteinases-4 (TIMP-4) gene expression is increased in human osteoarthritic femoral head cartilage. J. Cell. Biochem. 85 295–303. 10.1002/jcb.10138 [DOI] [PubMed] [Google Scholar]
- Huang W., Yang S., Shao J., Li Y. P. (2007). Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. 12:3068–3092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes D. E., Salter D. M., Simpson R. (1994). CD44 expression in human bone: a novel marker of osteocytic differentiation. J. Bone Miner. Res. 9 39–44. 10.1002/jbmr.5650090106 [DOI] [PubMed] [Google Scholar]
- Ikeda S., Morishita Y., Tsutsumi H., Ito M., Shiraishi A., Arita S., et al. (2003). Reductions in bone turnover, mineral, and structure associated with mechanical properties of lumbar vertebra and femur in glucocorticoid-treated growing minipigs. Bone 33 779–787. 10.1016/s8756-3282(03)00263-1 [DOI] [PubMed] [Google Scholar]
- Inada M., Wang Y., Byrne M. H., Miyaura C., Krane S. M. (2001). Mice with null mutation in collagenase-3 (Matrix Metalloproteinase [MMP]-13) exhibit altered bone remodeling and increased bone mass. J. Bone Miner. Res. 16:S149. [Google Scholar]
- Inada M., Wang Y., Byrne M. H., Miyaura C., Krane S. M. (2002). Loss of function of matrix metalloproteinase-13 (MMP-13) affects collagen accumulation and bone formation. J. Bone Miner. Res. 16:S171. [Google Scholar]
- Inada M., Wang Y., Byrne M. H., Rahman M. U., Miyaura C., Lopez-Otin C., et al. (2004). Critical roles for collagenase-3 (Mmp13) in development of growth plate cartilage and in endochondral ossification. Proc. Natl. Acad. Sci. U.S.A. 101 17192–17197. 10.1073/pnas.0407788101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoue K., Mikuni-Takagaki Y., Oikawa K., Itoh T., Inada M., Noguchi T., et al. (2006). A crucial role for matrix metalloproteinase 2 in osteocytic canalicular formation and bone metabolism. J. Biol. Chem. 281 33814–33824. 10.1074/jbc.m607290200 [DOI] [PubMed] [Google Scholar]
- Ishida N., Hayashi K., Hoshijima M., Ogawa T., Koga S., Miyatake Y., et al. (2002). Large scale gene expression analysis of osteoclastogenesis in vitro and elucidation of NFAT2 as a key regulator. J. Biol. Chem. 277 41147–41156. 10.1074/jbc.m205063200 [DOI] [PubMed] [Google Scholar]
- Ishikawa T., Nishigaki F., Miyata S., Hirayama Y., Minoura K., Imanishi J., et al. (2005). Prevention of progressive joint destruction in collagen-induced arthritis in rats by a novel matrix metalloproteinase inhibitor, FR255031. Br. J. Pharmacol. 144 133–143. 10.1038/sj.bjp.0706054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito A., Mukaiyama A., Itoh Y., Nagase H., Thogersen I. B., Enghild J. J., et al. (1996). Degradation of interleukin 1 beta by matrix metalloproteinases. J. Biol. Chem. 271 14657–14660. [DOI] [PubMed] [Google Scholar]
- Iyer R. P., Patterson N. L., Fields G. B., Lindsey M. L. (2012). The history of matrix metalloproteinases: milestones, myths, and misperceptions. Am. J. Physiol. Heart Circ. Physiol. 303 H919–H930. 10.1152/ajpheart.00577.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson M. T., Moradi B., Smith M. M., Jackson C. J., Little C. B. (2014). Activation of matrix metalloproteinases 2, 9, and 13 by activated protein C in human osteoarthritic cartilage chondrocytes. Arthritis Rheumatol. 66 1525–1536. 10.1002/art.38401 [DOI] [PubMed] [Google Scholar]
- Javaheri B., Hopkinson M., Poulet B., Pollard A. S., Shefelbine S. J., Chang Y. M., et al. (2016). Deficiency and also transgenic overexpression of TIMP-3 both lead to compromised bone mass and architecture in vivo. PLoS One 11:e0159657. 10.1371/journal.pone.0159657 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang Y. B., Zhao J., Genant H. K., Dequeker J., Geusens P. (1997). Long-term changes in bone mineral and biomechanical properties of vertebrae and femur in aging, dietary calcium restricted and/or estrogen-deprived/-replaced rats. J. Bone Miner. Res. 12 820–831. 10.1359/jbmr.1997.12.5.820 [DOI] [PubMed] [Google Scholar]
- Jimenez M. J., Balbin M., Lopez J. M., Alvarez J., Komori T., Lopez-Otın C. (1999). Collagenase 3 is a target of Cbfa1, a transcription factor of the runt gene family involved in bone formation. Mol. Cell. Biol. 19 4431–4442. 10.1128/mcb.19.6.4431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jing R., Liu Y., Guo P., Ni T., Gao X., Mei R., et al. (2018). Evaluation of common variants in matrix metalloproteinase-9 gene with lumbar disc herniation in han chinese population. Genet. Test Mol. Biomarkers 22 622–629. 10.1089/gtmb.2018.0080 [DOI] [PubMed] [Google Scholar]
- Johansson N., Saarialho-Kere U., Airola K., Herva R., Nissinen L., Westermarck J., et al. (1997). Collagenase-3 (MMP-13) is expressed by hypertrophic chondrocytes, periosteal cells, and osteoblasts during human fetal bone development. Dev. Dyn. 208 387–397. [DOI] [PubMed] [Google Scholar]
- Kajita M., Itoh Y., Chiba T., Mori H., Okada A., Kinoh H., et al. (2001). Membrane-type 1 matrix metalloproteinase cleaves CD44 and promotes cell migration. J. Cell Biol. 153 893–904. 10.1083/jcb.153.5.893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamioka H., Honjo T., Takano-Yamamoto T. (2001). A three-dimensional distribution of osteocyte processes revealed by the combination of confocal laser scanning microscopy and differential interference contrast microscopy. Bone 28 145–149. 10.1016/s8756-3282(00)00421-x [DOI] [PubMed] [Google Scholar]
- Kamiya N., Ye L., Kobayashi T., Mochida Y., Yamauchi M., Kronenberg H. M., et al. (2008). BMP signaling negatively regulates bone mass through sclerostin by inhibiting the canonical Wnt pathway. Development 135 3801–3811. 10.1242/dev.025825 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapur S., Baylink D. J., Lau K. H. (2003). Fluid flow shear stress stimulates human osteoblast proliferation and differentiation through multiple interacting and competing signal transduction pathways. Bone 32 241–251. 10.1016/s8756-3282(02)00979-1 [DOI] [PubMed] [Google Scholar]
- Karsdal M. A., Andersen T. A., Bonewald L., Christiansen C. (2004). Matrix metalloproteinases (MMPs) safeguard osteoblasts from apoptosis during transdifferentiation into osteocytes: MT1-MMP maintains osteocyte viability. DNA Cell Biol. 23 155–165. 10.1089/104454904322964751 [DOI] [PubMed] [Google Scholar]
- Karsdal M. A., Fjording M. S., Foged N. T., Delaisse J. M., Lochter A. (2001). Transforming growth factor-β-induced osteoblast elongation regulates osteoclastic bone resorption through a p38 mitogen-activated protein kinase- and matrix metalloproteinase-dependent pathway. J. Biol. Chem. 276 39350–39358. 10.1074/jbc.m008738200 [DOI] [PubMed] [Google Scholar]
- Karsdal M. A., Henriksen K., Leeming D. J., Woodworth T., Vassiliadis E., Bay-Jensen A. C. (2010). Novel combinations of Post-Translational Modification (PTM) neo-epitopes provide tissue-specific biochemical markers–are they the cause or the consequence of the disease? Clin. Biochem. 43 793–804. 10.1016/j.clinbiochem.2010.03.015 [DOI] [PubMed] [Google Scholar]
- Karsdal M. A., Larsen L., Engsig M. T., Lou H., Ferreras M., Lochter A., et al. (2002). Matrix metalloproteinase-dependent activation of latent transforming growth factor-β controls the conversion of osteoblasts into osteocytes by blocking osteoblast apoptosis. J. Biol. Chem. 277 44061–44067. 10.1074/jbc.m207205200 [DOI] [PubMed] [Google Scholar]
- Karsdal M. A., Woodworth T., Henriksen K., Maksymowych W. P., Genant H., Vergnaud P., et al. (2011). Biochemical markers of ongoing joint damage in rheumatoid arthritis - current and future applications, limitations and opportunities. Arthritis Res. Ther. 13:215. 10.1186/ar3280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasper G., Glaeser J. D., Geissler S., Ode A., Tuischer J., Matziolis G., et al. (2007). Matrix metalloprotease activity is an essential link between mechanical stimulus and mesenchymal stem cell behavior. Stem Cells 25 1985–1994. 10.1634/stemcells.2006-0676 [DOI] [PubMed] [Google Scholar]
- Kawaguchi H., Pilbeam C. C., Raisz L. G. (1994). Anabolic effects of 3,3’,5-triiodothyronine and triiodothyroacetic acid in cultured neonatal mouse parietal bones. Endocrinology 135 971–976. 10.1210/endo.135.3.7520864 [DOI] [PubMed] [Google Scholar]
- Kawaguchi H., Pilbean C. C., Harrison J. R., Raisz L. G. (1995). The role of prostaglandins in the regulation of bone metabolism. Clin. Orthop. 313 36–46. [PubMed] [Google Scholar]
- Kennedy A. M., Inada M., Krane S. M., Christie P. T., Harding B., Lopez-Otin C., et al. (2005). MMP13 mutation causes spondyloepimetaphyseal dysplasia. Missouri type (SEMDMO). J. Clin. Invest. 115 2832–2842. 10.1172/jci22900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerschan-Schindl K., Ebenbichler G. (2012). “Osteoimmunological aspects of biomechanics,” in Principles of Osteoimmunology, Molecular Mechanisms and Clinical Applications, ed. Pietschmann P., (New York, NY: SpringerWienNewYork; ), 97–107. [Google Scholar]
- Kim H. J., Zhao H., Kitaura H., Bhattacharyya S., Brewer J. A., Muglia L. J., et al. (2006). Glucocorticoids suppress bone formation via the osteoclast. J. Clin. Invest. 116 2152–2160. 10.1172/jci28084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J. W., Simmer J. P., Hart T. C., Hart P. S., Ramaswami M. D., Bartlett J. D., et al. (2005). MP-20 mutation in autosomal recessive pigmented hypomaturation amelogenesis imperfecta. J. Med. Genet. 42 271–275. 10.1136/jmg.2004.024505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim M. H., Park M., Baek S. H., Kim H. J., Kim S. H. (2011). Molecules and signaling pathways involved in the expression of OC-STAMP during osteoclastogenesis. Amino Acids 40 1447–1459. 10.1007/s00726-010-0755-4 [DOI] [PubMed] [Google Scholar]
- Kim Y., Sato K., Asagiri M., Morita I., Soma K., Takayanagi H. (2005). Contribution of nuclear factor of activated T cells c1 to the transcriptional control of immunoreceptor osteoclast-associated receptor but not triggering receptor expressed by myeloid cells-2 during osteoclastogenesis. J. Biol. Chem. 280 32905–32913. 10.1074/jbc.m505820200 [DOI] [PubMed] [Google Scholar]
- Kini U., Nandeesh B. N. (2012). “Physiology of bone formation, remodeling, and metabolism,” in Radionuclide and Hybrid Bone Imaging, eds Fogelman I., Gnanasegaran G., Van der Wall H., (Berlin: Springer-Verlag; ), 29–57. 10.1007/978-3-642-02400-9_2 [DOI] [Google Scholar]
- Klein T., Bischoff R. (2011). Physiology and pathophysiology of matrix metalloproteases. Amino Acids 41 271–290. 10.1007/s00726-010-0689-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knothe-Tate M. L., Adamson J. R., Tami A. E., Bauer T. W. (2004). The osteocyte. Int. J. Biochem. Cell Biol. 36 1–8. [DOI] [PubMed] [Google Scholar]
- Koga T., Inui M., Inoue K., Kim S., Suematsu A., Kobayashi E., et al. (2004). Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature 428 758–763. 10.1038/nature02444 [DOI] [PubMed] [Google Scholar]
- Koga T., Matsui Y., Asagiri M., Kodama T., de Crombrugghe B., Nakashima K., et al. (2005). NFAT and Osterix cooperatively regulate bone formation. Nat. Med. 11 880–885. 10.1038/nm1270 [DOI] [PubMed] [Google Scholar]
- Kojima T., Hasegawa T., de Freitas P. H., Yamamoto T., Sasaki M., Horiuchi K., et al. (2013). Histochemical aspects of the vascular invasion at the erosion zone of the epiphyseal cartilage in MMP-9-deficient mice. Biomed. Res. 34 119–128. 10.2220/biomedres.34.119 [DOI] [PubMed] [Google Scholar]
- Koolwijk P., Sidenius N., Peters E., Sier C. F., Hanemaaijer R., Blasi F., et al. (2001). Proteolysis of the urokinase-type plasminogen activator receptor by metalloproteinase-12: implication for angiogenesis in fibrin matrices. Blood 97 3123–3131. 10.1182/blood.v97.10.3123 [DOI] [PubMed] [Google Scholar]
- Kosaki N., Takaishi H., Kamekura S., Kimura T., Okada Y., Minqi L., et al. (2007). Impaired bone fracture healing in matrix metalloproteinase-13 deficient mice. Biochem. Biophys. Res. Commun. 354 846–851. 10.1016/j.bbrc.2006.12.234 [DOI] [PubMed] [Google Scholar]
- Koskinen A., Vuolteenaho K., Nieminen R., Moilanen T., Moilanen E. (2011). Leptin enhances MMP-1, MMP-3 and MMP-13 production in human osteoarthritic cartilage and correlates with MMP-1 and MMP-3 in synovial fluid from OA patients. Clin. Exp. Rheumatol. 29 57–64. [PubMed] [Google Scholar]
- Kotake S., Udagawa N., Takahashi N., Matsuzaki K., Itoh K., Ishiyama S., et al. (1999). IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J. Clin. Invest. 103 1345–1352. 10.1172/jci5703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krane S. M., Inada M. (2008). Matrix metalloproteinases and bone. Bone 43 7–18. 10.1016/j.bone.2008.03.020 [DOI] [PubMed] [Google Scholar]
- Krishnan V., Bryant H. U., Macdougald O. A. (2006). Regulation of bone mass by Wnt signaling. J. Clin. Invest. 116 1202–1209. 10.1172/jci28551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kusano K., Miyaura C., Inada M., Tamura T., Ito A., Nagase H., et al. (1998). Regulation of matrix metalloproteinases (MMP-2, -3, -9, and -13) by interleukin-1 and interleukin-6 in mouse calvaria: association of MMP induction with bone resorption. Endocrinology 139 1338–1345. 10.1210/endo.139.3.5818 [DOI] [PubMed] [Google Scholar]
- Lacey D. L., Timms E., Tan H. L., Kelley M. J., Dunstan C. R., Burgess T., et al. (1998). Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 93 165–176. 10.1016/s0092-8674(00)81569-x [DOI] [PubMed] [Google Scholar]
- Lafleur M. A., Mercuri F. A., Ruangpanit N., Seiki M., Sato H., Thompson E. W. (2006). Type I collagen abrogates the clathrin-mediated internalization of membrane type 1 matrix metalloproteinase (MT1-MMP) via the MT1-MMP hemopexin domain. J. Biol. Chem. 281 6826–6840. 10.1074/jbc.m513084200 [DOI] [PubMed] [Google Scholar]
- Lam J., Takeshita S., Barker J. E., Kanagawa O., Ross F. P., Teitelbaum S. L. (2000). TNF-alpha induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANK ligand. J. Clin. Invest. 106 1481–1488. 10.1172/jci11176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanske B., Amling M., Neff L., Guiducci J., Baron R., Kronenberg H. M. (1999). Ablation of the PTHrP gene or the PTH/PTHrP receptor gene leads to distinct abnormalities in bone development. J. Clin. Invest. 104 399–407. 10.1172/jci6629 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lassen N. E., Andersen T. L., Pløen G. G., Søe K., Hauge E. M., Harving S., et al. (2017). Coupling of bone resorption and formation in real time: new knowledge gained from human haversian BMUs. J. Bone Miner. Res. 32 1395–1405. 10.1002/jbmr.3091 [DOI] [PubMed] [Google Scholar]
- Lazarus S., Tseng H. W., Lawrence F., Woodruff M. A., Duncan E. L., Pettit A. R. (2017). Characterization of normal murine carpal bone development prompts re-evaluation of pathologic osteolysis as the cause of human carpal-tarsal osteolysis disorders. Am. J. Pathol. 187 1923–1934. 10.1016/j.ajpath.2017.05.007 [DOI] [PubMed] [Google Scholar]
- Lee H., Overall C. M., McCulloch C. A., Sodek J. (2006). A critical role for the membrane-type 1 matrix metalloproteinase in collagen phagocytosis. Mol. Biol. Cell. 17 4812–4826. 10.1091/mbc.e06-06-0486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li G., Liu D., Zhang Y., Qian Y., Zhang H., Guo S., et al. (2013). Celastrol inhibits lipopolysaccharide-stimulated rheumatoid fibroblast-like synoviocyte invasion through suppression of TLR4/NF-κB-mediated matrix metalloproteinase-9 expression. PLoS One 8:e008905. 10.1371/journal.pone.0068905 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Li N. G., Shi Z. H., Tang Y. P., Wang Z. J., Song S. L., Qian L. H., et al. (2011). New hope for the treatment of osteoarthritis through selective inhibition of MMP-13. Curr. Med. Chem. 18 977–1001. 10.2174/092986711794940905 [DOI] [PubMed] [Google Scholar]
- Li P. B., Tang W. J., Wang K., Zou K., Che B. (2017). Expressions of IL-1α and MMP-9 in degenerated lumbar disc tissues and their clinical significance. Eur. Rev. Med. Pharmacol. Sci. 21 4007–4013. [PubMed] [Google Scholar]
- Li Y. P., Chen W., Liang Y., Li E., Stashenko P. (1999). Atp6i-deficient mice exhibit severe osteopetrosis due to loss of osteoclast-mediated extracellular acidification. Nat. Genet. 23 447–451. 10.1038/70563 [DOI] [PubMed] [Google Scholar]
- Liang H. P. H., Xu J., Xue M., Jackson C. J. (2016). Matrix metalloproteinases in bone development and pathology: current knowledge and potential clinical utility. Metalloproteinases Med. 3 93–102. 10.2147/mnm.s92187 [DOI] [Google Scholar]
- Liao E. Y., Liao H. J., Guo L. J., Zhou H. D., Wu X. P., Dai R. C., et al. (2004). Membrane-type matrix metalloproteinase-1 (MT1-MMP) is down-regulated in estrogen-deficient rat osteoblast in vivo. J. Endocrinol. Invest. 27 1–5. 10.1007/bf03350902 [DOI] [PubMed] [Google Scholar]
- Lieu S., Hansen E., Dedini R., Behonick D., Werb Z., Miclau T., et al. (2011). Impaired remodeling phase of fracture repair in the absence of matrix metalloproteinase-2. Dis. Model Mech. 4 203–211. 10.1242/dmm.006304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsey M. L., Zouein F. A., Tian Y., Iyer R. P., De Castro Bras L. E. (2015). Osteopontin is proteolytically processed by matrix metalloproteinase 9. Can. J. Physiol. Pharmacol. 293 879–886. 10.1139/cjpp-2015-0019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J., Khalil R. A. (2017). Matrix metalloproteinase inhibitors as investigational and therapeutic tools in unrestrained tissue remodeling and pathological disorders. Prog. Mol. Biol. Transl. Sci. 148 355–420. 10.1016/bs.pmbts.2017.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S., Tang W., Fang J., Ren J., Li H., Xiao Z., et al. (2009). Novel regulators of Fgf23 expression and mineralization in Hyp bone. Mol. Endocrinol. 23 1505–1518. 10.1210/me.2009-0085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S., Zhou J., Tang W., Jiang X., Rowe D. W., Quarles D. L. (2006). Pathogenic role of Fgf23 in Hyp mice. Am. J. Physiol. Endocrinol. Metab. 291 E38–E49. 10.1210/me.2009-0085 [DOI] [PubMed] [Google Scholar]
- Liu Y. H., Tang Z., Kundu R. K., Wu L., Luo W., Zhu D., et al. (1999). Msx2 gene dosage influences the number of proliferative osteogenic cells in growth centers of the developing murine skull: a possible mechanism for MSX2-mediated craniosynostosis in humans. Dev. Biol. 205 260–274. 10.1006/dbio.1998.9114 [DOI] [PubMed] [Google Scholar]
- Loffek S., Schilling O., Franzke C. W. (2011). Series “matrix metalloproteinases in lung health and disease”: biological role of matrix metalloproteinases: a critical balance. Eur. Respir. J. 38 191–208. 10.1183/09031936.00146510 [DOI] [PubMed] [Google Scholar]
- Logan C. Y., Nusse R. (2004). The Wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 20 781–810. [DOI] [PubMed] [Google Scholar]
- Lopez-Otin C., Palavalli L. H., Samuels Y. (2009). Protective roles of matrix metalloproteinases: from mouse models to human cancer. Cell Cycle 8 3657–3662. 10.4161/cc.8.22.9956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovibond A. C., Haque S. J., Chambers T. J., Fox S. W. (2003). TGF-beta-induced SOCS3 expression augments TNF-alpha-induced osteoclast formation. Biochem. Biophys. Res. Commun. 309 762–767. 10.1016/j.bbrc.2003.08.068 [DOI] [PubMed] [Google Scholar]
- Lozito T. P., Jackson W. M., Nesti L. J., Tuan R. S. (2014). Human mesenchymal stem cells generate a distinct pericellular zone of MMP activities via binding of MMPs and secretion of high levels of TIMPs. Matrix Biol. 34 132–143. 10.1016/j.matbio.2013.10.003 [DOI] [PubMed] [Google Scholar]
- Lozito T. P., Tuan R. S. (2011). Mesenchymal stem cells inhibit both endogenous and exogenous MMPs via secreted TIMPs. J. Cell Physiol. 226 385–396. 10.1002/jcp.22344 [DOI] [PubMed] [Google Scholar]
- Lu C., Li X. Y., Hu Y., Rowe R. G., Weiss S. J. (2010). MT1-MMP controls human mesenchymal stem cell trafficking and differentiation. Blood 115 221–229. 10.1182/blood-2009-06-228494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu P., Takai K., Weaver V. M., Werb Z. (2011). Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 3:a005058. 10.1101/cshperspect.a005058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luan Z., Chase A. J., Newby A. C. (2003). Statins inhibit secretion of metalloproteinases-1, -2, -3, and -9 from vascular smooth muscle cells and macrophages. Arterioscler. Thromb. Vasc. Biol. 23 769–775. 10.1161/01.atv.0000068646.76823.ae [DOI] [PubMed] [Google Scholar]
- Luo J., Yang Z., Ma Y., Yue Z., Lin H., Qu G., et al. (2016). LGR4 is a receptor for RANKL and negatively regulates osteoclast differentiation and bone resorption. Nat. Med. 22 539–546. 10.1038/nm.4076 [DOI] [PubMed] [Google Scholar]
- Lv C., Yang S., Chen X., Zhu X., Lin W., Wang L., et al. (2017). MicroRNA-21 promotes bone mesenchymal stem cells migration in vitro by activating PI3K/Akt/MMPs pathway. J. Clin. Neurosci. 46 156–162. 10.1016/j.jocn.2017.07.040 [DOI] [PubMed] [Google Scholar]
- Lynch C. C. (2011). Matrix metalloproteinases as master regulators of the vicious cycle of bone metastasis. Bone 48 44–53. 10.1016/j.bone.2010.06.007 [DOI] [PubMed] [Google Scholar]
- Lynch C. C., Hikosaka A., Acuff H. B., Martin M. D., Kawai N., Singh R. K., et al. (2005). MMP-7 promotes prostate cancer-induced osteolysis via the solubilization of RANKL. Cancer Cell 7 485–496. 10.1016/j.ccr.2005.04.013 [DOI] [PubMed] [Google Scholar]
- MacDonald B. R. (1986). Parathyroid hormone, prostaglandins and bone resorption. World Rev. Nutr. Diet 47 163–201. 10.1159/000412334 [DOI] [PubMed] [Google Scholar]
- Madsen D. H., Jürgensen H. J., Ingvarsen S., Melander M. C., Albrechtsen R., Hald A., et al. (2013). Differential actions of the endocytic collagen receptor uPARAP/Endo180 and the collagenase MMP-2 in bone homeostasis. PLoS One 8:e71261. 10.1371/journal.pone.0071261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeda Y., Nakamura E., Nguyen M. T., Suva L. J., Swain F. L., Razzaque M. S., et al. (2007). Indian Hedgehog produced by postnatal chondrocytes is essential for maintaining a growth plate and trabecular bone. Proc. Natl. Acad. Sci. U.S.A. 104 6382–6387. 10.1073/pnas.0608449104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maffioli P., Derosa G. (2015). “Overview of biochemical markers of bone metabolism,” in Bone Disease, Biomarkers in Disease: Methods, Discoveries and Applications, ed. Preedy V. R., (Dordrecht: Springer Science+Business Media; ), 1–19. 10.1007/978-94-007-7745-3_24-1 [DOI] [Google Scholar]
- Mahl C., Egea V., Megens R. T. A., Pitsch T., Santovito D., Weber C., et al. (2016). RECK (reversion-inducing cysteine-rich protein with Kazal motifs) regulates migration, differentiation and Wnt/β-catenin signaling in human mesenchymal stem cells. Cell Mol. Life Sci. 73 1489–1501. 10.1007/s00018-015-2054-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malaponte G., Hafsi S., Polesel J., Castellano G., Spessotto P., Guarneri C., et al. (2016). Tumor microenvironment in diffuse large B-cell lymphoma: matrix metalloproteinases activation is mediated by osteopontin overexpression. Biochim. Biophys. Acta 1863 483–489. 10.1016/j.bbamcr.2015.09.018 [DOI] [PubMed] [Google Scholar]
- Mancini A., di Battista J. A. (2006). Transcriptional regulation of matrix metalloprotease gene expression in health and disease. Front. Biosci. 11:423–446. [DOI] [PubMed] [Google Scholar]
- Manduca P., Castagnino A., Lombardini D., Marchisio S., Soldano S., Ulivi V., et al. (2009). Role of MT1-MMP in the osteogenic differentiation. Bone 44 251–265. 10.1016/j.bone.2008.10.046 [DOI] [PubMed] [Google Scholar]
- Mannello F., Tonti G., Papa S. (2005). Matrix metalloproteinase inhibitors as anticancer therapeutics. Curr. Cancer Drug Targets 5 285–298. 10.2174/1568009054064615 [DOI] [PubMed] [Google Scholar]
- Manolagas S. C. (2000). Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr. Rev. 21 115–137. 10.1210/edrv.21.2.0395 [DOI] [PubMed] [Google Scholar]
- Marie P. J. (2003). Fibroblast growth factor signaling controlling osteoblast differentiation. Gene 316 23–32. 10.1016/s0378-1119(03)00748-0 [DOI] [PubMed] [Google Scholar]
- Martignetti J. A., Aqeel A. A., Sewairi W. A., Boumah C. E., Kambouris M., Mayouf S. A., et al. (2001). Mutation of the matrix metalloproteinase 2 gene (MMP2) causes a multicentric osteolysis and arthritis syndrome. Nat. Genet. 28 261–265. 10.1038/90100 [DOI] [PubMed] [Google Scholar]
- Massova I., Kotra L. P., Fridman R., Mobashery S. (1998). Matrix metalloproteinases: structures, evolution, and diversification. FASEB J. 12 1075–1095. 10.1096/fasebj.12.12.1075 [DOI] [PubMed] [Google Scholar]
- Matsuguchi T., Chiba N., Bandow K., Kakimoto K., Masuda A., Ohnishi T. (2009). JNK activity is essential for Atf4 expression and late-stage osteoblast differentiation. J. Bone Miner. Res. 24 398–410. 10.1359/jbmr.081107 [DOI] [PubMed] [Google Scholar]
- Matsumoto M., Kogawa M., Wada S., Takayanagi H., Tsujimoto M., Katayama S., et al. (2004). Essential role of p38 mitogen-activated protein kinase in cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1. J. Biol. Chem. 279 45969–45979. 10.1074/jbc.m408795200 [DOI] [PubMed] [Google Scholar]
- Matsuo K., Galson D. L., Zhao C., Peng L., Laplace C., Wang K. Z., et al. (2004). Nuclear factor of activated T-cells (NFAT) rescues osteoclastogenesis in precursors lacking c-Fos. J. Biol. Chem. 279 26475–26480. 10.1074/jbc.m313973200 [DOI] [PubMed] [Google Scholar]
- Mattot V., Raes M. B., Henriet P., Eeckhout Y., Stehelin D., Vandenbunder B., et al. (1995). Expression of interstitial collagenase is restricted to skeletal tissue during mouse embryogenesis. J. Cell Sci. 108 529–535. [DOI] [PubMed] [Google Scholar]
- Mattsson J. P., Schlesinger P. H., Keeling D. J., Teitelbaum S. L., Stone D. K., Xie X. S. (1994). Isolation and reconstitution of a vacuolar-type proton pump of osteoclast membranes. J. Biol. Chem. 269 24979–24982. [PubMed] [Google Scholar]
- Mauramo M., Ramseier A. M., Mauramo E., Buser A., Tervahartiala T., Sorsa T., et al. (2018). Associations of oral fluid MMP-8 with periodontitis in Swiss adult subjects. Oral Dis. 24 449–455. 10.1111/odi.12769 [DOI] [PubMed] [Google Scholar]
- McGowan P. M., Duffy M. J. (2008). Matrix metalloproteinase expression and outcome in patients with breast cancer: analysis of a published database. Ann. Oncol. 19 1566–1572. 10.1093/annonc/mdn180 [DOI] [PubMed] [Google Scholar]
- Meikle M. C., Bord S., Hembry R. M., Compston J., Croucher P. I., Reynolds J. J. (1992). Human osteoblasts in culture synthesize collagenase and other matrix metalloproteinases in response to osteotropic hormones and cytokines. J. Cell Sci. 103 1093–1099. [DOI] [PubMed] [Google Scholar]
- Meng F., Yang H., Aitha M., George S., Tierney D. L., Crowder M. W. (2016). Biochemical and spectroscopic characterization of the catalytic domain of MMP16 (cdMMP16). J. Biol. Inorg. Chem. 21 523–535. 10.1007/s00775-016-1362-y [DOI] [PubMed] [Google Scholar]
- Messaritou G., East L., Roghi C., Isacke C. M., Yarwood H. (2009). Membrane type-1 matrix metalloproteinase activity is regulated by the endocytic collagen receptor Endo180. J. Cell Sci. 122 4042–4048. 10.1242/jcs.044305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikuni-Takagaki Y. (1999). Mechanical responses and signal transduction pathways in stretched osteocytes. J. Bone Miner. Metab. 17 57–60. 10.1007/s007740050065 [DOI] [PubMed] [Google Scholar]
- Miller B., Spevak L., Lukashova L., Javaheri B., Pitsillides A. A., Boskey A., et al. (2017). Altered bone mechanics, architecture and composition in the skeleton of TIMP-3-deficient mice. Calcif. Tissue Int. 100 631–640. 10.1007/s00223-017-0248-5 [DOI] [PubMed] [Google Scholar]
- Mills B. G., Frausto A. (1997). Cytokines expressed in multinucleated cells: Paget’s disease and giant cell tumors versus normal bone. Calcif. Tissue Int. 61 16–21. 10.1007/s002239900285 [DOI] [PubMed] [Google Scholar]
- Mimura H., Cao X., Ross F. P., Chiba M., Teitelbaum S. L. (1994). 1,25-Dihydroxyvitamin D3 transcriptionally activates the beta 3-integrin subunit gene in avian osteoclast precursors. Endocrinology 134 1061–1066. 10.1210/endo.134.3.8119143 [DOI] [PubMed] [Google Scholar]
- Mittal R., Patel A. P., Debs L. H., Nguyen D., Patel K., Grati M., et al. (2016). Intricate functions of matrix metalloproteinases in physiological and pathological conditions. J. Cell Physiol. 231 2599–2621. 10.1002/jcp.25430 [DOI] [PubMed] [Google Scholar]
- Miyamoto T. (2006). The dendritic cell-specific transmembrane protein DC-STAMP is essential for osteoclast fusion and osteoclast bone-resorbing activity. Mod. Rheumatol. 16 341–342. 10.3109/s10165-006-0524-0 [DOI] [PubMed] [Google Scholar]
- Miyauchi A., Alvarez J., Greenfield E. M., Teti A., Grano M., Colucci S., et al. (1991). Recognition of osteopontin and related peptides by an alpha v beta 3 integrin stimulates immediate cell signals in osteoclasts. J. Biol. Chem. 266 20369–20374. [PubMed] [Google Scholar]
- Miyauchi A., Gotoh M., Kamioka H., Notoya K., Sekiya H., Takagi Y., et al. (2006). AlphaVbeta3 integrin ligands enhance volume-sensitive calcium influx in mechanically stretched osteocytes. J Bone Miner. Metab. 24 498–504. 10.1007/s00774-006-0716-x [DOI] [PubMed] [Google Scholar]
- Miyauchi A., Notoya K., Mikuni-Takagaki Y., Takagi Y., Goto M., Miki Y., et al. (2000). Parathyroid hormone-activated volume-sensitive calcium influx pathways in mechanically located osteocytes. J. Biol. Chem. 275 3335–3342. 10.1074/jbc.275.5.3335 [DOI] [PubMed] [Google Scholar]
- Mocsai A., Humphrey M. B., Van Ziffle J. A., Hu Y., Burghardt A., Spusta S. C., et al. (2004). The immunomodulatory adapter proteins DAP12 and Fc receptor gamma-chain (FcRgamma) regulate development of functional osteoclasts through the Syk tyrosine kinase. Proc. Natl. Acad. Sci. U.S.A. 101 6158–6163. 10.1073/pnas.0401602101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohanakrishnan V., Balasubramanian A., Mahalingam G., Partridge N. C., Ramachandran I., Selvamurugan N. (2018). Parathyroid hormone-induced down-regulation of miR-532-5p for matrix metalloproteinase-13 expression in rat osteoblasts. J. Cell Biochem. 119 6181–6193. 10.1002/jcb.26827 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mosekilde L., Danielsen D. D., Knudsen U. B. (1993). The effect of aging and ovariectomy on the vertebral bone mass and biomechanical properties of mature rats. Bone 14 1–6. 10.1016/8756-3282(93)90248-9 [DOI] [PubMed] [Google Scholar]
- Mosig R. A., Dowling O., DiFeo A., Ramirez M. C., Parker I. C., Abe E., et al. (2007). Loss of MMP-2 disrupts skeletal and craniofacial development and results in decreased bone mineralization, joint erosion and defects in osteoblast and osteoclast growth. Hum. Mol. Genet. 16 1113–1123. 10.1093/hmg/ddm060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mott J. D., Thomas C. L., Rosenbach M. T., Takahara K., Greenspan D. S., Banda M. J. (2000). Post-translational proteolytic processing of procollagen C-terminal proteinase enhancer releases a metalloproteinase inhibitor. J. Biol. Chem. 275 1384–1390. 10.1074/jbc.275.2.1384 [DOI] [PubMed] [Google Scholar]
- Mundy G. (1993). Cytokines and growth factors in the regulation of bone remodeling. J. Bone Miner. Res. 8 S505–S510. [DOI] [PubMed] [Google Scholar]
- Murray E. J. B., Tram K. K.-T., Spencer M. J., Tidball J. G., Murray S. S., Lee D. B. N. (1995). PTH mediated osteoblast retraction: possible participation of the calpain pathway. Miner. Electrolyte Metab. 21 184–188. [PubMed] [Google Scholar]
- Nagase H., Visse R., Murphy G. (2006). Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 69 562–573. 10.1016/j.cardiores.2005.12.002 [DOI] [PubMed] [Google Scholar]
- Nakashima A., Tamura M. (2006). Regulation of matrix metalloproteinase-13 and tissue inhibitor of matrix metalloproteinase-1 gene expression by WNT3A and bone morphogenetic protein-2 in osteoblastic differentiation. Front. Biosci. 11:1667–1678. [DOI] [PubMed] [Google Scholar]
- Nannuru K. C., Futakuchi M., Varney M. L., Vincent T. M., Marcusson E. G., Singh R. K. (2010). Matrix metalloproteinase (MMP)-13 regulates mammary tumor-induced osteolysis by activating MMP9 and transforming growth factor-beta signaling at the tumor-bone interface. Cancer Res. 70 3494–3504. 10.1158/0008-5472.CAN-09-3251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nash T. J., Howlett C. R., Martin C., Steele J., Johnson K. A., Hicklin D. J. (1994). Effects of platelet-derived growth factor on tibial osteotomies in rabbits. Bone 15 203–208. 10.1016/8756-3282(94)90709-9 [DOI] [PubMed] [Google Scholar]
- Neuhold L. A., Killar L., Zhao W., Sung M. L., Warner L., Kulik J., et al. (2001). Postnatal expression in hyaline cartilage of constitutively active human collagenase-13 (MMP-13) induces osteoarthritis in mice. J. Clin. Investig. 107 35–44. 10.1172/jci10564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen M., Arkell J., Jackson C. J. (2000). Activated protein C directly activates human endothelial gelatinase A. J. Biol. Chem. 275 9095–9098. 10.1074/jbc.275.13.9095 [DOI] [PubMed] [Google Scholar]
- Nyman J. S., Lynch C. C., Perrien D. S., Thiolloy S., O’Quinn E. C., Patil C. A., et al. (2011). Differential effects between the loss of MMP-2 and MMP-9 on structural and tissue-level properties of bone. J. Bone. Miner. Res. 26 1252–1260. 10.1002/jbmr.326 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh J., Takahashi R., Adachi E., Kondo S., Kuratomi S., Noma A., et al. (2004). Mutations in two matrix metalloproteinase genes, MMP-2 and MT1-MMP, are synthetic lethal in mice. Oncogene 23 5041–5048. 10.1038/sj.onc.1207688 [DOI] [PubMed] [Google Scholar]
- Oh J., Takahashi R., Kondo S., Mizoguchi A., Adachi E., Sasahara R. M., et al. (2001). The membrane-anchored MMP inhibitor RECK is a key regulator of extracellular matrix integrity and angiogenesis. Cell 107 789–800. 10.1016/s0092-8674(01)00597-9 [DOI] [PubMed] [Google Scholar]
- Okada M., Shinmei Y., Tanaka O., Naka K., Kimura A., Nakanishi I., et al. (1992). Localization of matrix metalloproteinase 3 (stromelysin) in osteoarthritic cartilage and synovium. Lab. Invest. 66 680–690. [PubMed] [Google Scholar]
- Ortega N., Behonick D., Dominique S., Werb Z. (2003). How proteases regulate bone morphogenesis. Ann. N. Y. Acad. Sci. 995 109–116. 10.1111/j.1749-6632.2003.tb03214.x [DOI] [PubMed] [Google Scholar]
- Ortega N., Behonick D. J., Colnot C., Cooper D. N., Werb Z. (2005). Galectin-3 is a downstream regulator of matrix metalloproteinase-9 function during endochondral bone formation. Mol. Biol. Cell 16 3028–3039. 10.1091/mbc.e04-12-1119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortega N., Wang K., Ferrara N., Werb Z., Vu T. H. (2010). Complementary interplay between matrix metalloproteinase-9, vascular endothelial growth factor and osteoclast function drives endochondral bone formation. Dis. Model Mech. 3 224–235. 10.1242/dmm.004226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overall C. M., Wrana J. L., Sodek J. (1991). Transcriptional and posttranscriptional regulation of 72-kDa gelatinase type-IV collagenase by transforming growth factor-beta-1 in human fibroblasts: comparisons with collagenase and tissue inhibitor of matrix metalloproteinase gene-expression. J. Biol. Chem. 266 14064–14071. [PubMed] [Google Scholar]
- Ozdemir D., Hart P. S., Ryul O. H., Choi S. J., Ozdemir-Karatas M., Firatli E., et al. (2005). MMP20 active-site mutation in hypomaturation amelogenesis imperfecta. J. Dent. Res. 84 1031–1035. 10.1177/154405910508401112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page-McCaw A., Ewald A. J., Werb Z. (2007). Matrix metalloproteinases and the regulation of tissue remodeling. Nat. Rev. Mol. Cell Biol. 8 221–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paiva K. B. S., Granjeiro J. M. (2014). Bone tissue remodeling and development: focus on matrix metalloproteinase functions. Arch. Biochem. Biophys. 561 74–87. 10.1016/j.abb.2014.07.034 [DOI] [PubMed] [Google Scholar]
- Paiva K. B. S., Granjeiro J. M. (2017). Matrix metalloproteinases in bone resorption, remodeling, and repair. Prog. Mol. Biol. Transl. Sci. 148 203–303. 10.1016/bs.pmbts.2017.05.001 [DOI] [PubMed] [Google Scholar]
- Pap T., Korb-Pap A. (2015). Cartilage damage in osteoarthritis and rheumatoid arthritis-two unequal siblings. Nat. Rev. Rheumatol. 11 606–615. 10.1038/nrrheum.2015.95 [DOI] [PubMed] [Google Scholar]
- Pap T., Shigeyama Y., Kuchen S., Fernihough J. K., Simmen B., Gay R. E., et al. (2000). Differential expression pattern of membrane-type matrix metalloproteinases in rheumatoid arthritis. Arthritis Rheum. 43 1226–1232. [DOI] [PubMed] [Google Scholar]
- Papathanasiou I., Malizos K. N., Tsezou A. (2012). Bone morphogenetic protein-2-induced Wnt/beta-catenin signaling pathway activation through enhanced low-density-lipoprotein receptor-related protein 5 catabolic activity contributes to hypertrophy in osteoarthritic chondrocytes. Arthritis Res. Ther. 14:R82. 10.1186/ar3805 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parra-Torres A. Y., Valdés-Flores M., Orozco L., Velázquez-Cruz R. (2013). “Molecular aspects of bone remodeling,” in Topics in Osteoporosis, ed. Flores M. V., (London: IntechOpen; ). [Google Scholar]
- Pego E. R., Fernandez I., Nuñez M. J. (2018). Molecular basis of the effect of MMP-9 on the prostate bone metastasis: a review. Urol. Oncol. 36 272–282. 10.1016/j.urolonc.2018.03.009 [DOI] [PubMed] [Google Scholar]
- Pei D., Kang T., Qi H. (2000). Cysteine array matrix metalloproteinase (CA-MMP)/MMP-23 is a type II transmembrane matrix metalloproteinase regulated by a single cleavage for both secretion and activation. J. Biol. Chem. 275 33988–33997. 10.1074/jbc.m006493200 [DOI] [PubMed] [Google Scholar]
- Pei D., Weiss S. J. (1995). Furin-dependent intracellular activation of the human stromelysin-3 zymogen. Nature 375 244–247. 10.1038/375244a0 [DOI] [PubMed] [Google Scholar]
- Pettit A. R., Chang M. K., Hume D. A., Raggatt L. G. (2008). Osteal macrophages: a new twist on coupling during bone dynamics. Bone 43 976–982. 10.1016/j.bone.2008.08.128 [DOI] [PubMed] [Google Scholar]
- Piccard H., Van den Steen P. E., Opdenakker G. (2007). Hemopexin domains as multifunctional liganding modules in matrix metalloproteinases and other proteins. J. Leukoc. Biol. 81 870–892. 10.1189/jlb.1006629 [DOI] [PubMed] [Google Scholar]
- Pirapaharan D. C., Olesen J. B., Andersen T. L., Christensen S. B., Kjærsgaard-Andersen P., Delaisse J. M., et al. (2019). Catabolic activity of osteoblast lineage cells contributes to osteoclastic bone resorption in vitro. J. Cell Sci. 15 132. 10.1242/jcs.229351 [DOI] [PubMed] [Google Scholar]
- Pivetta E., Scapolan M., Pecolo M., Wassermann B., Abu-Rumeileh I., Balestreri L., et al. (2011). MMP-13 stimulates osteoclast differentiation and activation in tumour breast bone metastases. Breast Cancer Res. 13:R105. 10.1186/bcr3047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plotkin L. I., Bruzzaniti A. (2019). Molecular signaling in bone cells: regulation of cell differentiation and survival. Adv. Protein Chem. Struc. Biol. 116 237–281. 10.1016/bs.apcsb.2019.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plotkin L. I., Lezcano V., Thostenson J., Weinstein R. S., Manolagas S. C., Bellido T. (2008). Connexin 43 is required for the anti-apoptotic effect of bisphosphonates on osteocytes and osteoblasts in vivo. J. Bone Miner. Res. 23 1712–1721. 10.1359/jbmr.080617 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plotkin L. I., Manolagas S. C., Bellido T. (2002). Transduction of cell survival signals by connexin-43 hemichannels. J. Biol. Chem. 277 8648–8657. 10.1074/jbc.m108625200 [DOI] [PubMed] [Google Scholar]
- Plotkin L. I., Mathov I., Aguirre J. I., Parfitt A. M., Mangolas S. C., Bellido T. (2005). Mechanical stimulation prevents osteocyte apoptosis: requirement of integrins, Src kinases, and ERKs. Am. J. Physiol. Cell Physiol. 289 C633–C643. [DOI] [PubMed] [Google Scholar]
- Poole K. E., van Bezooijen R. L., Loveridge N., Hamersma H., Papapoulos S. E., Lowik C. W., et al. (2005). Sclerostin is a delayed secreted product of osteocytes that inhibits bone formation. FASEB J. 19 1842–1844. 10.1096/fj.05-4221fje [DOI] [PubMed] [Google Scholar]
- Porter S., Clark I. M., Kevorkian L., Edward D. R. (2005). The ADAMTS metalloproteinases. Biochem. J. 386 15–27. 10.1042/bj20040424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Puliti M., Momi S., Falcinelli E., Gresele P., Bistoni F., Tissi L. (2012). Contribution of matrix metalloproteinase 2 to joint destruction in group B Streptococcus-induced murine arthritis. Arthritis Rheum. 64 1089–1097. 10.1002/art.33450 [DOI] [PubMed] [Google Scholar]
- Quinn J. M. W., Neale S., Fujikawa Y., McGee J. D., Athanasou N. A. (1998). Human osteoclast formation from blood monocytes, peritoneal macrophages, and bone marrow cells. Calcif. Tissue Int. 62 527–531. 10.1007/s002239900473 [DOI] [PubMed] [Google Scholar]
- Qvist P., Christiansen C., Karsdal M. A., Madsen S. H., Sondergaard B. C., Bay-Jensen A. C. (2010). Application of biochemical markers in development of drugs for treatment of osteoarthritis. Biomarkers 15 1–19. 10.3109/13547500903295873 [DOI] [PubMed] [Google Scholar]
- Ragab A. A., Nalepka J. L., Bi Y., Greenfield E. M. (2002). Cytokines synergistically induce osteoclast differentiation: support by immortalized or normal calvarial cells. Am. J. Physiol. 283 C679–C687. [DOI] [PubMed] [Google Scholar]
- Raisz L. G. (1993). Bone cell biology: new approaches and unanswered questions. J. Bone Miner. Res. 8 457–465. [DOI] [PubMed] [Google Scholar]
- Raisz L. G. (1997). The osteoporosis revolution. Ann. Intern. Med. 126 458–462. [DOI] [PubMed] [Google Scholar]
- Rajaram S., Murawala H., Buch P., Patel S., Balakrishnan S. (2016). Inhibition of BMP signaling reduces MMP-2 and MMP-9 expression and obstructs wound healing in regenerating fin of teleost fish Poecilia latipinna. Fish Physiol. Biochem. 42 787–794. 10.1007/s10695-015-0175-1 [DOI] [PubMed] [Google Scholar]
- Rauner M., Stein N., Hofbauer L. C. (2012). “Basics of bone biology,” in Principles of Osteoimmunology, Molecular Mechanisms and Clinical Applications, ed. Pietschmann P., (New York, NY: SpringerWienNewYork; ), 1–26. 10.1007/978-3-7091-0520-7_1 [DOI] [Google Scholar]
- Rawlinson S. C., Pitsillides A. A., Lanyon L. E. (1996). Involvement of different ion channels in osteoblasts’and osteocytes’ early responses to mechanical strain. Bone 19 609–614. 10.1016/s8756-3282(96)00260-8 [DOI] [PubMed] [Google Scholar]
- Raynal C. P., Delmas D., Chenu C. (1996). Bone sialoprotein stimulates in vitro bone resorption. Endocrinology 137 2347–2354. 10.1210/endo.137.6.8641185 [DOI] [PubMed] [Google Scholar]
- Roach H. I. (1994). Why does bone matrix contain non-collagenous proteins? The possible roles of osteocalcin, osteonectin, osteopontin and bone sialoprotein in bone mineralisation and resorption. Cell Biol. Int. 18 617–628. 10.1006/cbir.1994.1088 [DOI] [PubMed] [Google Scholar]
- Robey P. G. (1989). The biochemistry of bone. Endocrinol. Metab. Clin. North Am. 18 858–902. [PubMed] [Google Scholar]
- Roodman G. D. (1999). Cell biology of the osteoclast. Exp. Hematol. 27 1229–1241. 10.1016/s0301-472x(99)00061-2 [DOI] [PubMed] [Google Scholar]
- Roodman G. D., Kurihara N., Ohsaki Y., Kukita A., Hosking D., Demulder A., et al. (1992). Interleukin-6: a potential autocrine/paracrine agent in Paget’s disease of bone. J. Clin. Invest. 89 46–52. 10.1172/jci115584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose B. J., Kooyman D. L. (2016). A tale of two joints: the role of matrix metalloproteases in cartilage biology. Dis. Markers. 2016 4895050. 10.1155/2016/4895050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen C. J., Donahue L. R. (1998). Insulin-like growth factors and bone-the osteoporosis connection revisited. Proc. Soc. Exp. Biol. Med. 219 1–7. 10.3181/00379727-219-44310 [DOI] [PubMed] [Google Scholar]
- Rouzier C., Vanatka R., Bannwarth S., Philip N., Coussement A., Paquis-Flucklinger V., et al. (2006). A novel homozygous MMP2 mutation in a family with Winchester syndrome. Clin. Genet. 69 271–276. 10.1111/j.1399-0004.2006.00584.x [DOI] [PubMed] [Google Scholar]
- Rowe P. S. N., Kumagai Y., Gutierrez G., Garrett I. R., Blacher R., Rosen D., et al. (2004). MEPE has the properties of an osteoblastic phosphatonin and minhibin. Bone 34 303–319. 10.1016/j.bone.2003.10.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowe R. G., Weiss S. (2009). Navigating ECM barriers at the invasive front: the cancer cell-stroma interface. Annu. Rev. Cell Dev. Biol. 25 567–595. 10.1146/annurev.cellbio.24.110707.175315 [DOI] [PubMed] [Google Scholar]
- Ruan M., Pederson L., Bradley E. W., Bamberger A. M., Oursler M. J. (2010). Transforming growth factor-{beta} coordinately induces suppressor of cytokine signaling 3 and leukemia inhibitory factor to suppress osteoclast apoptosis. Endocrinology 151 1713–1722. 10.1210/en.2009-0813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubin J., Rubin C., Jacobs C. R. (2006). Molecular pathways mediating mechanical signalling in bone. Gene 367 1–16. 10.1016/j.gene.2005.10.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruchon A. F., Tenenhouse H. S., Marcinkiewicz M., Siegfried G., Aubin J. E., DesGroseillers L., et al. (2000). Developmental expression and tissue distribution of Phex protein: effect of the Hyp mutation and relationship to bone markers. J. Bone Miner. Res. 15 1440–1450. 10.1359/jbmr.2000.15.8.1440 [DOI] [PubMed] [Google Scholar]
- Rutter J. L., Mitchell T. I., Buttice G., Meyers J., Gusella J. F., Ozelius L. J., et al. (1998). A single nucleotide polymorphism in the matrix metalloproteinase-1 promoter creates an Ets binding site and augments transcription. Cancer Res. 58 5321–5325. [PubMed] [Google Scholar]
- Sabbota A. L., Kim H. R., Zhe X., Fridman R., Bonfil R. D., Cher M. L. (2010). Shedding of RANKL by tumor-associated MT1-MMP activates Src-dependent prostate cancer cell migration. Cancer Res 70 5558–5566. 10.1158/0008-5472.CAN-09-4416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sabeh F., Fox D., Weiss S. J. (2010). Membrane-type I matrix metalloproteinase-dependent regulation of rheumatoid arthritis synoviocyte function. J. Immunol. 184 6396–6406. 10.4049/jimmunol.0904068 [DOI] [PubMed] [Google Scholar]
- Saftig P., Hunziker E., Wehmeyer O., Jones S., Boyde A., Rommerskirch W., et al. (1998). Impaired osteoclastic bone resorption leads to osteopetrosis in cathepsin-K-deficient mice. Proc. Natl. Acad. Sci. U.S.A. 95 13453–13458. 10.1073/pnas.95.23.13453 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sage E. H., Reed M., Funk S. E., Truong T., Steadele M., Puolakkainen P., et al. (2003). Cleavage of the matricellular protein SPARC by matrix metalloproteinase 3 produces polypeptides that influence angiogenesis. J. Biol. Chem. 278 37849–37857. 10.1074/jbc.m302946200 [DOI] [PubMed] [Google Scholar]
- Sahebjam S., Khokha R., Mort J. S. (2007). Increased collagen and aggrecan degradation with age in the joints of Timp3(-/-) mice. Arthritis Rheum. 56 905–909. 10.1002/art.22427 [DOI] [PubMed] [Google Scholar]
- Sarker H., Hardy E., Haimour A., Maksymowych W. P., Botto L. D., Fernandez-Patron C. (2019). Identification of fibrinogen as a natural inhibitor of MMP-2. Sci. Rep. 9:4340. 10.1038/s41598-019-40983-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sasaki T., Göhring W., Mann K., Maurer P., Hohenester E., Knäuper V., et al. (1997). Limited cleavage of extracellular matrix protein BM-40 by matrix metalloproteinases increases its affinity for collagens. J. Biol. Chem. 272 9237–9243. 10.1074/jbc.272.14.9237 [DOI] [PubMed] [Google Scholar]
- Satokata I., Ma L., Ohshima H., Bei M., Woo I., Nishizawa K., et al. (2000). Msx2 deficiency in mice causes pleiotropic defects in bone growth and ectodermal organ formation. Nat. Genet. 24 391–395. 10.1038/74231 [DOI] [PubMed] [Google Scholar]
- Sawicki G. (2013). Intracellular regulation of matrix metalloproteinase-2 activity: new strategies in treatment and protection of heart subjected to oxidative stress. Scientifica 2013:130451. 10.1155/2013/130451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider G. B., Key L. L., Popoff S. N. (1998). Osteopetrosis. Therapeutic strategies. Endocrinologist 8 409–417. 10.1097/00019616-199811000-00004 [DOI] [Google Scholar]
- Semenov M., Tamai K., He X. (2005). SOST is a ligand for LRP5/LRP6 and a Wnt signaling inhibitor. J. Biol. Chem. 280 26770–26775. 10.1074/jbc.m504308200 [DOI] [PubMed] [Google Scholar]
- Shah M., Huang D., Blick T., Connor A., Reiter L. A., Hardink J. R., et al. (2012). An MMP13-selective inhibitor delays primary tumor growth and the onset of tumor-associated osteolytic lesions in experimental models of breast cancer. PLoS One 7:e29615. 10.1371/journal.pone.0029615 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahnazari M., Martin B. R., Legette L. L., Lachcik P. J., Welch J., Weaver C. M. (2009). Diet calcium levelbut not calcium supplement particle size affects bone density and mechanical properties in ovariectomized rats. J. Nutr. 139 1308–1314. 10.3945/jn.108.101071 [DOI] [PubMed] [Google Scholar]
- Shen Y., Winkler I. G., Barbier V., Sims N. A., Hendy J., Levesque J. P. (2010). Tissue inhibitor of metalloproteinase-3 (TIMP-3) regulates hematopoiesis and bone formation in vivo. PLoS One 5:e13086. 10.1371/journal.pone.0013086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sher L. B., Woitge H. W., Adams D. J., Gronowicz G. A., Krozowski Z., Harrison J. R., et al. (2004). Transgenic expression of 11beta-hydroxysteroid dehydrogenase type 2 in osteoblasts reveals an anabolic role for endogenous glucocorticoids in bone. Endocrinology 145 922–929. 10.1210/en.2003-0655 [DOI] [PubMed] [Google Scholar]
- Shi J., Son M. Y., Yamada S., Szabova L., Kahan S., Chrysovergis K., et al. (2008). Membrane-type MMPs enable extracellular matrix permissiveness and mesenchymal cell proliferation during embryogenesis. Dev. Biol. 313 196–209. 10.1016/j.ydbio.2007.10.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shore E. M., Xu M., Feldman G. J., Fenstermacher D. A., Cho T. J., Choi I. H., et al. (2006). A recurrent mutation in the BMP type I receptor ACVR1 causes inherited and sporadic fibrodysplasia ossificans progressiva. Nat. Genet. 38 525–527. 10.1038/ng1783 [DOI] [PubMed] [Google Scholar]
- Silver I. A., Murrills R. J., Etherington D. J. (1988). Microelectrode studies on the acid microenvironment beneath adherent macrophages and osteoclasts. Exp. Cell. Res. 175 266–276. 10.1016/0014-4827(88)90191-7 [DOI] [PubMed] [Google Scholar]
- Sims N. A., Martin T. J. (2014). Coupling the activities of bone formation and resorption: a multitude of signals within the basic multicellular unit. Bonekey Rep. 3:481. 10.1038/bonekey.2013.215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A., Rajasekaran N., Hartenstein B., Szabowski S., Gajda M., Angel P., et al. (2013). Collagenase-3 (MMP-13) deficiency protects C57BL/6 mice from antibody-induced arthritis. Arthritis Res. Ther. 15:R222. 10.1186/ar4423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siris E. S. (1998). Paget’s disease of bone. J. Bone Miner. Res. 13 1061–1065. [DOI] [PubMed] [Google Scholar]
- Sobue T., Hakeda Y., Kobayashi Y., Hayakawa H., Yamashita K., Aoki T., et al. (2001). Tissue inhibitor of metalloproteinases 1 and 2 directly stimulate the bone-resorbing activity of isolated mature osteoclasts. J. Bone Miner. Res. 16 2205–2214. 10.1359/jbmr.2001.16.12.2205 [DOI] [PubMed] [Google Scholar]
- Song I. W., Li W. R., Chen L. Y., Shen L. F., Liu K. M., Yen J. J., et al. (2014). Palmitoyl acyltransferase, Zdhhc13, facilitates bone mass acquisition by regulating postnatal epiphyseal development and endochondral ossification: a mouse model. PLoS One 9:e92194. 10.1371/journal.pone.0092194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sottrup-Jensen L. (1989). Alpha-macroglobulins: structure, shape, and mechanism of proteinase complex formation. J. Biol. Chem. 264 11539–11542. [PubMed] [Google Scholar]
- Springman E. B., Angleton E. L., Birkedal-Hansen H., Van Wart H. E. (1990). Multiple modes of activation of latent human fibroblast collagenase: evidence for the role of a Cys73 active-site zinc complex in latency and a “cysteine switch” mechanism for activation. Proc. Natl. Acad. Sci. U.S.A. 87 364–368. 10.1073/pnas.87.1.364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stahle-Backdahl M., Sandstedt B., Bruce K., Lindahl A., Jimenez M. G., Vega J. A., et al. (1997). Collagenase-3 (MMP-13) is expressed during human fetal ossification and re-expressed in postnatal bone remodeling and in rheumatoid arthritis. Lab. Invest. 76 717–728. [PubMed] [Google Scholar]
- Sternlicht M. D., Werb Z. (2001). How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell. Dev. Biol. 17 463–516. 10.1146/annurev.cellbio.17.1.463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stickens D., Behonick D. J., Ortega N., Heyer B., Hartenstein B., Yu Y., et al. (2004). Altered endochondral bone development in matrix metalloproteinase 13-deficient mice. Development 131 5883–5895. 10.1242/dev.01461 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugimoto K., Nakamura T., Tokunaga T., Uehara Y., Okada T., Taniwaki T., et al. (2018). Matrix metalloproteinase promotes elastic fiber degradation in ligamentum flavum degeneration. PLoS One 13:e0200872. 10.1371/journal.pone.0200872 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh W. K., Wang S. X., Jheon A. H., Moreno L., Yoshinaga S. K., Ganss B., et al. (2004). The immune regulatory protein B7-H3 promotes osteoblast differentiation and bone mineralization. Proc. Natl. Acad. Sci. U.S.A. 101 12969–12973. 10.1073/pnas.0405259101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulkala M., Tervahartiala T., Sorsa T., Larmas M., Salo T., Tjaderhane L. (2007). Matrix metalloproteinase-8 (MMP-8) is the major collagenase in human dentin. Arch. Oral Biol. 52 121–127. 10.1016/j.archoralbio.2006.08.009 [DOI] [PubMed] [Google Scholar]
- Sun B., Sun J., Han X., Liu H., Li J., Du J., et al. (2016). Immunolocalization of MMP 2, 9 and 13 in prednisolone induced osteoporosis in mice. Histol. Histopathol. 31 647–656. 10.14670/HH-11-702 [DOI] [PubMed] [Google Scholar]
- Sundaram K., Nishimura R., Senn J., Youssef R. F., London S. D., Reddy S. V. (2007). RANK ligand signaling modulates the matrix metalloproteinase-9 gene expression during osteoclast differentiation. Exp. Cell Res. 313 168–178. 10.1016/j.yexcr.2006.10.001 [DOI] [PubMed] [Google Scholar]
- Takayanagi H., Kim S., Koga T., Nishina H., Isshiki M., Yoshida H., et al. (2002). Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 3 889–901. 10.1016/s1534-5807(02)00369-6 [DOI] [PubMed] [Google Scholar]
- Takemura Y., Moriyama Y., Ayukawa Y., Kurata K., Rakhmatia Y. D., Koyano K. (2019). Mechanical loading induced osteocyte apoptosis and connexin 43 expression in three-dimensional cell culture and dental implant model. J. Biomed. Mater. Res. A 107 815–827. 10.1002/jbm.a.36597 [DOI] [PubMed] [Google Scholar]
- Tamamura Y., Otani T., Kanatani N., Koyama E., Kitagaki J., Komori T., et al. (2005). Developmental regulation of Wnt/beta-catenin signals is required for growth plate assembly, cartilage integrity, and endochondral ossification. J. Biol. Chem. 280 19185–19195. 10.1074/jbc.m414275200 [DOI] [PubMed] [Google Scholar]
- Tanaka S., Takahashi N., Udagawa N., Tamura T., Akatsu T., Stanley E. R., et al. (1993). Macrophage colony-stimulating factor is indispensable for both proliferation and differentiation of osteoclast progenitors. J. Clin. Invest. 91 257–263. 10.1172/jci116179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka-Kamioka K., Kamioka H., Ris H., Lim S. S. (1998). Osteocyte shape is dependent on actin filaments and osteocyte processes are unique actin-rich projections. J. Bone Miner. Res. 13 1555–1568. 10.1359/jbmr.1998.13.10.1555 [DOI] [PubMed] [Google Scholar]
- Tang S. Y., Herber R. P., Ho S. P., Alliston T. (2012). Matrix metalloproteinase-13 is required for osteocytic perilacunar remodeling and maintains bone fracture resistance. J. Bone Miner. Res. 27 1936–1950. 10.1002/jbmr.1646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang Y., Rowe R. G., Botvinick E. L., Kurup A., Putnam A. J., Seiki M., et al. (2013). MT1-MMP-dependent control of skeletal stem cell commitment via a β1-integrin/YAP/TAZ signaling axis. Dev. Cell 25 402–416. 10.1016/j.devcel.2013.04.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang Y., Wu X., Lei W., Pang L., Wan C., Shi Z., et al. (2009). TGF-beta1-induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nat. Med. 15 757–765. 10.1038/nm.1979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tauro M., Lynch C. C. (2018). Cutting to the Chase: How matrix metalloproteinase- 2 activity controls breast-cancer-to-bone metastasis. Cancers 10:E185. 10.3390/cancers10060185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teitelbaum S. L. (2000). Bone resorption by osteoclasts. Science 289 1504–1508. 10.1126/science.289.5484.1504 [DOI] [PubMed] [Google Scholar]
- Teitelbaum S. L. (2007). Osteoclasts: what do they do and how do they do it? Am. J. Pathol. 170 427–435. 10.2353/ajpath.2007.060834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teitelbaum S. L., Abu-Amer Y., Ross F. P. (1995). Molecular mechanisms of bone resorption. J. Cell Biochem. 59 1–10. [DOI] [PubMed] [Google Scholar]
- Teitelbaum S. L., Ross F. P. (2003). Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 4 638–649. 10.1038/nrg1122 [DOI] [PubMed] [Google Scholar]
- Tester A. M., Waltham M., Oh S. J., Bae S. N., Bills M. M., Walker E. C., et al. (2004). Pro-matrix metalloproteinase-2 transfection increases orthotopic primary growth and experimental metastasis of MDA-MB-231 human breast cancer cells in nude mice. Cancer Res. 64 652–658. 10.1158/0008-5472.can-0384-2 [DOI] [PubMed] [Google Scholar]
- Teti A., Blair H. C., Schlesinger P., Grano M., Zambonin-Zallone A., Kahn A. J., et al. (1989). Extracellular protons acidify osteoclasts, reduce cytosolic calcium, and promote expression of cell-matrix attachment structures. J. Clin. Invest. 84 773–780. 10.1172/jci114235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thiolloy S., Halpern J., Holt G. E., Schwartz H. S., Mundy G. R., Matrisian L. M., et al. (2009). Osteoclast-derived matrix metalloproteinase-7, but not matrix metalloproteinase-9, contributes to tumor-induced osteolysis. Cancer Res. 69 6747–6755. 10.1158/0008-5472.CAN-08-3949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson P. D., Clarkson P., Karas R. H. (2003). Statin-associated myopathy. JAMA 289 1681–1690. [DOI] [PubMed] [Google Scholar]
- Tian E., Zhan F., Walker R., Rasmussen E., Ma Y., Barlogie B., et al. (2003). The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N. Engl. J. Med. 349 2483–2494. 10.1056/nejmoa030847 [DOI] [PubMed] [Google Scholar]
- Tokito A., Jougasaki M. (2016). Matrix metalloproteinases in non-neoplastic disorders. Int. J. Mol. Sci. 17:1178. 10.3390/ijms17071178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tondravi M. M., McKercher S. R., Anderson K., Erdmann J. M., Quiroz M., Maki R., et al. (1997). Osteopetrosis in mice lacking haematopoietic transcription factor PU.1. Nature 386 81–84. 10.1038/386081a0 [DOI] [PubMed] [Google Scholar]
- Toyosawa S., Oya K., Sato S., Ishida K. (2012). Osteocyte and DMP1. Clin. Calcium 22 713–720. [PubMed] [Google Scholar]
- Tozum T. F., Oppenlander M. E., Koh-Paige A. J., Robins D. M., McCauley L. K. (2004). Effects of sex steroid receptor specificity in the regulation of skeletal metabolism. Calcif. Tissue Int. 75 60–70. 10.1007/s00223-004-0119-8 [DOI] [PubMed] [Google Scholar]
- Troeberg L., Nagase H. (2012). Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta 1824 133–145. 10.1016/j.bbapap.2011.06.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuysuz B., Mosig R., Altun G., Sancak S., Glucksman M. J., Martignetti J. A. (2009). A novel matrix metalloproteinase 2 (MMP2) terminal hemopexin domain mutation in a family with multicentric osteolysis with nodulosis and arthritis with cardiac defects. Eur. J. Hum. Genet. 17 565–572. 10.1038/ejhg.2008.204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Udagawa N., Takahashi N., Jimi E., Matsuzaki K., Tsurukai T., Itoh K., et al. (1999). Osteoblasts/stromal cells stimulate osteoclast activation through expression of osteoclast differentiation factor/RANKL but not macrophage colony-stimulating factor: receptor activator of NF-kappa B ligand. Bone 25 517–523. 10.1016/s8756-3282(99)00210-0 [DOI] [PubMed] [Google Scholar]
- Vaananen H. K., Horton M. (1995). The osteoclast clear zone is a specialized cell-extracellular matrix adhesion structure. J. Cell Sci. 108 2729–2732. [DOI] [PubMed] [Google Scholar]
- Van Wart H. E., Birkedal-Hansen H. (1990). The cysteine switch: a principle of regulation of metalloproteinase activity with potential applicability to the entire matrix metalloproteinase gene family. Proc. Natl. Acad. Sci. U.S.A. 87 5578–5582. 10.1073/pnas.87.14.5578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenbroucke R. E., Libert C. (2014). Is there new hope for therapeutic matrix metalloproteinase inhibition? Nature Rev. Drug Discov. 13 904–927. 10.1038/nrd4390 [DOI] [PubMed] [Google Scholar]
- Vandooren J., Van Damme J., Opdenakker G. (2014). On the structure and functions of gelatinase B/matrix metalloproteinase-9 in neuroinflammation. Prog. Brain Res. 214 193–206. 10.1016/B978-0-444-63486-3.00009-8 [DOI] [PubMed] [Google Scholar]
- Veidal S. S., Larsen D. V., Chen X., Sun S., Zheng Q., Bay-Jensen A. C., et al. (2012). MMP mediated type V collagen degradation (C5M) is elevated in ankylosing spondylitis. Clin. Biochem. 45 541–546. 10.1016/j.clinbiochem.2012.02.007 [DOI] [PubMed] [Google Scholar]
- Velasco G., Pendas A. M., Fueyo A., Knäuper V., Murphy G., Lopez-Otın C. (1999). Cloning and characterization of human MMP-23, a new matrix metalloproteinase predominantly expressed in reproductive tissues and lacking conserved domains in other family members. J. Biol. Chem. 274 4570–4576. 10.1074/jbc.274.8.4570 [DOI] [PubMed] [Google Scholar]
- Visse R., Nagase H. (2003). Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circ. Res. 92 827–839. 10.1161/01.res.0000070112.80711.3d [DOI] [PubMed] [Google Scholar]
- Vivinus-Nebot M., Rousselle P., Breittmayer J. P., Cenciarini C., Berrih-Aknin S., Spong S., et al. (2004). Mature human thymocytes migrate on laminin-5 with activation of metalloproteinase-14 and cleavage of CD44. J. Immunol. 172 1397–1406. 10.4049/jimmunol.172.3.1397 [DOI] [PubMed] [Google Scholar]
- Vu T. H., Shipley J. M., Bergers G., Berger J. E., Helms J. A., Hanahan D., et al. (1998). MMP-9/gelatinase B is a key regulator of growth plate angiogenesis and apoptosis of hypetrophic chondrocytes. Cell 93 411–422. 10.1016/s0092-8674(00)81169-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada T., Nakashima T., Oliveira-dos-Santos A. J., Gasser J., Hara H., Schett G., et al. (2005). The molecular scaffold Gab2 is a crucial component of RANK signaling and osteoclastogenesis. Nat Med. 11 394–399. 10.1038/nm1203 [DOI] [PubMed] [Google Scholar]
- Wagenaar-Miller R. A., Engelholm L. H., Gavard J., Yamada S. S., Gutkind J. S., Behrendt N., et al. (2007). Complementary roles of intracellular and pericellular collagen degradation pathways in vivo. Mol. Cell. Biol. 27 6309–6322. 10.1128/mcb.00291-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westbroek I., De Rooij K. E., Nijweide P. J. (2002). Osteocyte-specific monoclonal antibody MAb OB7.3 is directed against Phex protein. J. Bone Miner. Res. 17 845–853. 10.1359/jbmr.2002.17.5.845 [DOI] [PubMed] [Google Scholar]
- Wiebe S. H., Hafezi M., Sandhu H. S., Sims S. M., Dixon S. J. (1996). Osteoclast activation in inflammatory periodontal diseases. Oral Dis. 2 167–180. 10.1111/j.1601-0825.1996.tb00218.x [DOI] [PubMed] [Google Scholar]
- Wu H., Du J., Zheng Q. (2008). Expression of MMP-1 in cartilage and synovium of experimentally induced rabbit ACLT traumatic osteoarthritis: immunohistochemical study. Rheumatol. Int. 29 31–36. 10.1007/s00296-008-0636-2 [DOI] [PubMed] [Google Scholar]
- Wutzl A., Rauner M., Seemann R., Millesi W., Krepler P., Pietschmann P., et al. (2010). Bone morphogenetic proteins 2, 5, and 6 in combination stimulate osteoblasts but not osteoclasts in vitro. J. Orthop. Res. 28 1431–1439. 10.1002/jor.21144 [DOI] [PubMed] [Google Scholar]
- Xian L., Wu X., Pang L., Lou M., Rosen C. J., Qiu T., et al. (2012). Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells. Nat. Med. 18 1095–1101. 10.1038/nm.2793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xue M., March L., Sambrook P. N., Jackson C. J. (2007). Differential regulation of matrix metalloproteinase 2 and matrix metalloproteinase 9 by activated protein C: relevance to inflammation in rheumatoid arthritis. Arthritis Rheum. 56 2864–2874. 10.1002/art.22844 [DOI] [PubMed] [Google Scholar]
- Xue M., McKelvey K., Shen K., Minhas N., March L., Park S. Y., et al. (2014). Endogenous MMP-9 and not MMP-2 promotes rheumatoid synovial fibroblast survival, inflammation and cartilage degradation. Rheumatology 53 2270–2279. 10.1093/rheumatology/keu254 [DOI] [PubMed] [Google Scholar]
- Yamada Y., Ando F., Niino N., Shimokata H. (2004). Association of a polymorphism of the matrix metalloproteinase-9 gene with bone mineral density in japanese men. Metabolism 53 135–137. 10.1016/j.metabol.2003.09.003 [DOI] [PubMed] [Google Scholar]
- Yamagiwa H., Tokunaga K., Hayami T., Hatano H., Uchida M., Endo N., et al. (1999). Expression of metalloproteinase-13 (collagenase-3) is induced during fracture healing in mice. Bone 25 197–203. 10.1016/s8756-3282(99)00157-x [DOI] [PubMed] [Google Scholar]
- Yamaguchi A., Komori T., Suda T. (2000). Regulation of osteoblast differentiation mediated by bone morphogenetic proteins, hedgehogs, and Cbfa1. Endocr. Rev. 21 393–411. 10.1210/edrv.21.4.0403 [DOI] [PubMed] [Google Scholar]
- Yee C. S., Schurman C. A., White C. R., Alliston T. (2019). Investigating osteocytic perilacunar/canalicular remodeling. Curr. Osteoporos Rep. 17 157–168. 10.1007/s11914-019-00514-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zankl A., Bonafe L., Calcaterra V., Di Rocco M., Superti-Furga A. (2005). Winchester syndrome caused by a homozygous mutation affecting the active site of matrix metalloproteinase 2. Clin. Genet. 67 261–266. 10.1111/j.1399-0004.2004.00402.x [DOI] [PubMed] [Google Scholar]
- Zeng Y., Rosborough R. C., Li Y., Gupta A. R., Bennett J. (1998). Temporal and spatial regulation of gene expression mediated by the promoter for the human tissue inhibitor of metalloproteinases-3 (TIMP-3)-encoding gene. Dev. Dyn. 211 228–237. [DOI] [PubMed] [Google Scholar]
- Zhang B., Henney A., Eriksson P., Hamsten A., Watkins H., Ye S. (1999). Genetic variation at the matrix metalloproteinase-9 locus on chromosome 20q12.2-13.1. Hum. Genet. 105 418–423. 10.1007/s004399900167 [DOI] [PubMed] [Google Scholar]
- Zhang J. F., Wang G. L., Zhou Z. J., Fang X. Q., Chen S., Fan S. W. (2018). Expression of matrix metalloproteinases, tissue inhibitors of metalloproteinases, and interleukins in vertebral cartilage endplate. Orthop. Surg. 10 306–311. 10.1111/os.12409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K., Barragan-Adjemian C., Ye L., Kotha S., Dallas M., Lu Y., et al. (2006). E11/gp38 selective expression in osteocytes: regulation by mechanical strain and role in dendrite elongation. Mol. Cell Biol. 26 4539–4552. 10.1128/mcb.02120-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W., Yang N., Shi X. M. (2008). Regulation of mesenchymal stem cell osteogenic differentiation by glucocorticoid-induced leucine zipper (GILZ). J. Biol. Chem. 283 4723–4729. 10.1074/jbc.m704147200 [DOI] [PubMed] [Google Scholar]
- Zhao H., Cai G., Du J., Xia Z., Wang L., Zhu T. (1997). Expression of matrix metalloproteinase-9 mRNA in osteoporotic bone tissues. J. Tongji. Med. Univ. 17 28–31. 10.1007/bf02887998 [DOI] [PubMed] [Google Scholar]
- Zhao S., Zhang Y. K., Harris S., Ahuja S. S., Bonewald L. F. (2002). MLO-Y4 osteocyte-like cells support osteoclast formation and activation. J. Bone Miner. Res. 17 2068–2079. 10.1359/jbmr.2002.17.11.2068 [DOI] [PubMed] [Google Scholar]
- Zheng H., Liu J., Tycksen E., Nunley R., McAlinden A. (2019). MicroRNA-181a/b-1 over-expression enhances osteogenesis by modulating PTEN/PI3K/AKT signaling and mitochondrial metabolism. Bone 123 92–102. 10.1016/j.bone.2019.03.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Z., Apte S. S., Soininen R., Cao R., Baaklini G. Y., Rauser R. W., et al. (2000). Impaired endochondral ossification and angiogenesis in mice deficient in membrane type matrix metalloproteinase I. Proc. Natl. Acad. Sci. U.S.A. 97 4052–4057. 10.1073/pnas.060037197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zucker S., Cao J., Chen W. T. (2000). Critical appraisal of the use of matrix metalloproteinase inhibitors in cancer treatment. Oncogene 19 6642–6650. 10.1038/sj.onc.1204097 [DOI] [PubMed] [Google Scholar]