

Abstract
Expanding the network of protected areas is a core strategy for conserving biodiversity in the face of climate change. Here, we explore the impacts on reserve network cost and configuration associated with planning for climate change in the USA using networks that prioritize areas projected to be climatically suitable for 1460 species both today and into the future, climatic refugia and areas likely to facilitate climate-driven species movements. For 14% of the species, networks of sites selected solely to protect areas currently climatically suitable failed to provide climatically suitable habitat in the future. Protecting sites climatically suitable for species today and in the future significantly changed the distribution of priority sites across the USA—increasing relative protection in the northeast, northwest and central USA. Protecting areas projected to retain their climatic suitability for species cost 59% more than solely protecting currently suitable areas. Including all climatic refugia and 20% of areas that facilitate climate-driven movements increased the cost by another 18%. Our results indicate that protecting some types of climatic refugia may be a relatively inexpensive adaptation strategy. Moreover, although addressing climate change in conservation plans will have significant implications for the configuration of networks, the increased cost of doing so may be relatively modest.
This article is part of the theme issue ‘Climate change and ecosystems: threats, opportunities and solutions’.
Keywords: climate change, protected areas, conservation planning, refugia, connectivity
1. Introduction
Many species are shifting their distributions in ways that are consistent with predictions based on recent changes in climate [1–4]. As climate change progresses, climate-induced range shifts are expected to increase [5,6]. These shifts have significant ramifications for what is arguably one of the most effective conservation strategies—protecting areas from development and other land-use conversions. As species’ ranges shift, areas that have been protected to provide habitat for those species may cease to contain suitable climatic conditions [7–9].
Several approaches to augmenting existing protected area networks to better conserve biodiversity in the face of climate change have been proposed [10–13]. Two of the most recommended involve growing the network of protected areas, by increasing either the size of existing reserves or the number of reserves [10]. The former could allow species to track suitable climatic conditions within existing protected areas. Increasing the number of reserves would provide additional places for species to go as climates change—increasing the chances that a species could track suitable climatic conditions.
Yet, to be most effective, reserve network expansions must be implemented strategically. A number of approaches to addressing species needs in an expanded reserve network have been proposed [14]. One such approach that has recently received much attention is protecting so-called refugia [15–17]. There is ample evidence of historical climatic macrorefugia [18,19], regions of a continent that facilitate the persistence of species in the face of large-scale, sustained, climatic change [20]. As a result, researchers have developed several approaches for identifying potential macrorefugia for modern climate change, including areas where specific species are projected to persist in the future [21], where climate velocities are low [17], where little climatic change is expected [22], where environmental diversity is high [17], and areas that retain increasingly rare climatic conditions [23].
In addition to expanding the reserve network, another often-recommended strategy for addressing climate impacts on biodiversity involves increasing the connectivity of the landscape [10]. By making it easier for plants and animals to move from place to place, increasing connectivity may allow more species to track suitable climatic conditions, particularly in human-dominated landscapes. The benefits of a larger network will be limited if species are unable to move from their current locations to suitable areas in the future. Although the idea of increasing connectivity to protect biodiversity is not new, few studies have developed connectivity-planning approaches that directly address climate-driven movements [24,25].
Despite the wealth of suggestions for addressing climate change in conservation planning, relatively few studies have tested the impact of incorporating these approaches into the conservation-planning process [26]—and even fewer have explored the effects of more than one approach at a time [27]. Here, we explore the effects of addressing climate change on both the configuration and the cost of a national protected area network using three such approaches. We selected sites to augment current protected areas in the USA using different combinations of areas projected to be currently climatically suitable for species, areas projected to be climatically suitable for species in the future, potential climate refugia and areas important for increasing connectivity to facilitate climate-driven movements. We compared both the distributions and estimated relative costs of the resulting new networks.
2. Methods
(a). Networks
To explore the effect of applying each of the three approaches to addressing climate change on network cost and configuration, we selected five sets of networks to augment the current protected areas in the USA to meet five different goals (table 1). First, we selected sites to protect currently climatically suitable areas for plants and vertebrates. Second, we selected sets of sites that protected species-specific refugia. These networks protected sites that were projected to be climatically suitable for species both today and into the future. Third, we selected networks that protected currently climatically suitable areas for species and areas of potential climatic macrorefugia—locations that retain increasingly rare climatic conditions. Fourth, we selected networks that protected currently climatically suitable areas for species and areas identified as providing connectivity to facilitate climate-driven species movements (hereafter climate corridors). Finally, we selected networks that protected species-specific refugia, climatic refugia and climate corridors (i.e. combining the second, third and fourth goals).
Table 1.
Description of five types of reserve networks selected to augment current protected areas using different approaches to address climate change.
| network | targets | approach |
|---|---|---|
| current species | current species distributions | protect areas projected to be suitable for species today |
| species refugia | species-specific refugia | protect areas projected to be suitable for species today and into the future |
| climatic refugia | current species distributions and climatic refugia | protect areas projected to be currently suitable for species and climate analogues with shrinking geographical footprints |
| connectivity | current species distributions and climate corridors | protect areas projected to be currently suitable for species and areas with corridors designed to facilitate climate-driven movements |
| all | species-specific refugia, climatic refugia and climate corridors | protect areas projected to be currently suitable for species, climate analogues with shrinking geographical footprints, and corridors designed to facilitate climate-driven movements |
(b). Sites
We developed a grid of 78 412 10 km × 10 km cells to represent potential sites to include in an expanded reserve network across the contiguous USA. We considered sites with at least 50% of their area in protected status as protected. To identify protected areas, we used the Conservation Biology Institute's protected areas database [28], including all protected areas with Gap Analysis Project (GAP) status 1 or 2 (electronic supplementary material, figure S1). Lands with GAP status 1 or 2 are protected from conversion from a natural state and have a mandated management plan to maintain a natural state.
To estimate planning unit-specific average costs of land acquisition, we used the layer of county-specific land prices for crops, pasture, forest and range (which includes both grasslands and shrublands) described in Withey et al. [29] (electronic supplementary material, figure S2). For each planning unit, we calculated a weighted average cost, based on the current land cover (National Land Cover Database 2001 land cover grid, 2011 edition [30]) within counties overlapped by the planning unit. The land price estimates incorporated the present value of future returns owing to development [29], but we also assumed that urban land cover would not be targeted for conservation. Network costs were estimated assuming 1000 ha of land would be protected at each site selected to be included in a network.
(c). Current and projected future climatic suitability
We inferred current climatic suitability with species distribution models (SDMs) using presence-only occurrence data from the Global Biodiversity Information Facility [31]. We identified 2237 species of plants, birds, mammals, reptiles and amphibians with distributions that overlapped the study area and with at least 25 occurrence records. SDMs were constructed with MaxEnt, with pseudo-absences randomly selected from locations within an area 1.5 times larger than the extent of the occurrence data. A taxon-specific prior based on target-group sampling was used to account for sampling bias [32]. Model output was discretized to predictions of presence/absence using a threshold that maximized the sum of model sensitivity and specificity.
To identify species-specific refugia, we projected future climatic suitability using the projections from three global climate models (GCMs; INM-CM4 [33], MIROC5 [34] and GFDL-CM3 [35]), run for RCP 8.5, for the period 2061–2080 using data from the WorldClim database [36]. Projections from three GCMs were selected to represent mild, moderate and considerable change, respectively. We chose RCP 8.5 to explore the higher end of projected changes. Thus, our resulting estimates should be interpreted as higher bounds of spatial differences and costs. To be considered species-specific refugia, sites had to be projected to be climatically suitable across all three climate-change scenarios and current climatic conditions.
We removed the most widespread species from the initial set of 2237 modelled species, using only those with distributions covering less than one-third of the area of the country. Removing widespread species decreased the processing time without significantly changing the solutions (e.g. costs of reserve networks with the full set of species were within 0.1% of those with the reduced set). This produced a dataset of 1460 species including 500 plants, 301 birds, 256 mammals, 206 reptiles and 197 amphibians.
(d). Connectivity
We defined climate corridors using an existing connectivity analysis conducted for the contiguous USA [37] (electronic supplementary material, figure S3). The analysis involved identifying pathways that connected warmer to cooler areas that were less impacted by human activities. These pathways both avoided human impacts on the landscape and the crossing of steep climatic gradients, thus producing routes that avoided passing through areas likely to be climatically unsuitable [38]. The routes were based solely on existing climatic gradients using climate data mapped at 1 km resolution [39]. The level of human impact on the landscape was based on two existing layers that incorporated a combination of land-use, roads, road use, population density, coastal access, railroads, navigable rivers and night-time lighting [40,41]. To include climate corridors in selected protected area networks, we mapped these routes to the 10 km grid cells and calculated the length of the routes intersecting each grid cell. To ensure that we captured contiguous corridors, we identified all pairs of cells sharing a common route and prioritized the clustering of these cells in the site-selection process.
(e). Climatic refugia
We used a map of climatic macrorefugia that identified locations with future climatic conditions that are rare relative to their historical distribution (electronic supplementary material, figure S4). Specifically, for each location on the landscape, this approach found all other locations with analogous climatic conditions within a specified distance. These macrorefugia are locations for which the number of climate analogues declines in the future, indicating that those climatic conditions become increasingly rare [23]. We used a binary, composite map that included macrorefugia identified using eight different combinations of dispersal constraint (i.e. analogue search radius) and climate sensitivity (i.e. analogue threshold value). Refugia were identified using the same three GCMs used to project species-specific climatic suitability, RCP 8.5 and a 30-year average for the end of century (2071–2100). These data were originally mapped at 1 km resolution and resampled to a 10 km resolution using a majority rule algorithm. We selected all locations consistently identified as refugia by all three GCMs.
(f). Site selection
All of the protected area networks we selected were designed to increase the protection of species across the USA. For the reserve networks selected to protect areas of current climatic suitability for species or species-specific refugia, we selected sets of protected areas to include a total of 50, 100, 200, 400 and 800 sites for each species, respectively. Including a range of targets for protection allowed us to explore how differences in costs scaled with target size and avoided the arbitrary selection of a specific target. For species with fewer than the target number of sites with climatic suitability, we required all suitable sites to be protected.
We explored the impact of including 20%, 40%, 60%, 80% and 100% of the modelled climatic refugia in the reserve networks. To ensure that the refugia were distributed as equally across the country as possible, we required representation at each percentage within each level-2 ecoregion [42]—a total of 12 of 21 ecoregions contained refugia. Likewise, we explored the effects of including 20%, 40%, 60%, 80% and 100% of the total length of all identified corridors in the reserve networks. Again, we required representation at each of these levels within each ecoregion. Because the corridors were more widely spread than the refugia, this involved protecting corridors in 21 ecoregions. To account for the fact that corridors often spanned more than one site, we forced the reserve-selection algorithm to prioritize sites that shared a common corridor.
All site selection was done with the conservation planning software Marxan [43]. Marxan uses simulated annealing to find sets of sites that protect the conservation elements—species, refugia, corridors, in the present study—while minimizing the total cost of the reserve network. For each type of reserve network, we completed 100 Marxan runs with 750 million iterations each. To capture continuous sections of corridors, we used the boundary length modifier in Marxan. The boundary length modifier allows the user to adjust the cost of shared boundaries between sites. When shared boundary costs are high, the algorithm favours sets of sites that are clumped. We labelled only the boundaries between sites that shared corridors as shared boundaries and used the boundary length modifier to produce reserve networks that captured corridors as they traversed sites. We explored the impacts of using a boundary length modifier that ranged from 0 to 1000 to select a suitable value for the two sets of networks involving corridors.
3. Results
We found that reserve networks designed solely to protect areas currently climatically suitable for species failed to protect 14% of the species at the specified target levels based on projected future climatic suitability. These tended to be species with smaller distributions. Accounting for climate change in protected area networks substantially altered the distribution of selected sites (figure 1). Each of the four approaches to addressing climate change required more sites than did the additional protection of currently climatically suitable areas. For example, to protect species-specific refugia, several sites were added across the country, particularly in the northeastern, northwestern and central USA (figure 2a). In addition, the importance of sites selected to protect currently climatically suitable locations often changed once future climatic suitability was considered. Only 45% of sites selected based on current suitability were selected with the same frequency in networks designed to also capture future suitability (grey areas in figure 2a).
Figure 1.

Maps of the relative importance of sites selected to protect (a) current species distributions, (b) species-specific refugia, (c) current species distributions and climatic refugia, (d) current species distributions and corridors designed to facilitate climate-driven species movements, and (e) species-specific refugia, climatic refugia and corridors designed to facilitate climate-driven species movements. Values in the maps represent the number of times a site was included in 100 reserve networks designed to include their respective targets while minimizing total cost of the reserve network.
Figure 2.
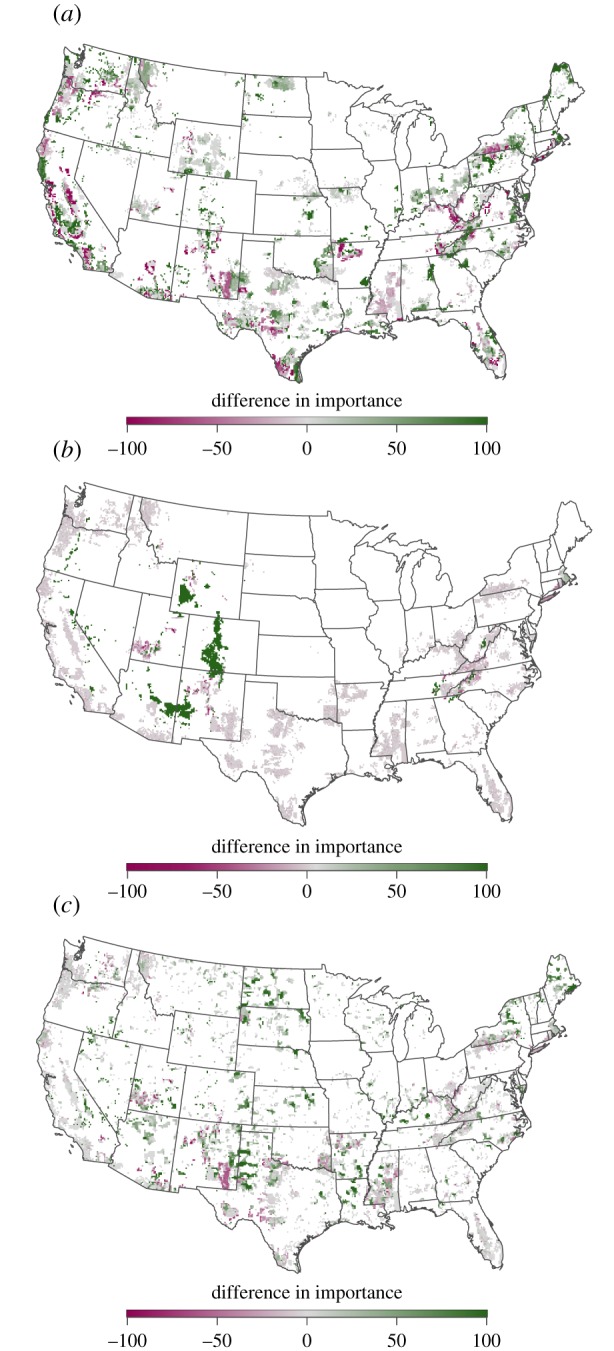
The difference in the relative importance of sites for (a) protecting species-specific refugia and current species distributions (figure 1b,a), (b) species current distributions and climatic refugia and solely protecting current species distributions (figure 1c,a), and (c) current species distributions and corridors designed to facilitate climate-driven species movements and solely protecting current species distributions (figure 1d,a). Green areas are more important for addressing climate change and pink areas are important for protecting current species distributions but are less important for addressing climate change.
Protecting species-specific refugia—areas projected to be climatically suitable for species today and in the future—would cost 59% more than protecting species' current distributions alone (figure 3). This relative increase in cost grew with increased levels of protection (figure 4). The cost differential occurs largely because once both current and future species locations are prioritized, the number of site options which can satisfy that requirement is greatly reduced (compared to networks that only prioritize currently suitable areas). On average, there were 66% fewer sites projected to be climatically suitable, both currently and into the future, for a species than there were sites projected to be currently suitable.
Figure 3.

Average costs of 100 reserve networks selected based on each of five different approaches to addressing climate change in the conservation-planning process (see table 1 for explanations of approaches). Error bars span the minimum to the maximum costs across the 100 networks.
Figure 4.

Average costs of 100 reserve networks selected to protect sites harbouring current species distributions and networks selected to protect species-specific refugia (sites projected to be climatically suitable for species today and into the future) in different number of sites (i.e. each species had to be protected in 50, 100, 200, 400 and 800 sites). Error bars span the minimum to the maximum costs across the 100 networks.
Only solutions requiring protection of at least 40% of climatic refugia had a discernable effect on site selection because 26% of the refugia were already captured in the existing reserve network. Reserve networks requiring protection of 100% of refugia added several sites to the networks, the largest concentrations being in the southern Rocky Mountains and the Colorado Plateau in the southwestern USA (figures 1c and 2b). Costs were only slightly higher for including 40%, 60%, 80% and even 100% of the refugia (figure 5). Protecting all of the modelled refugia increased the cost of the reserve networks designed to solely protect sites currently suitable for species by 16% (figure 3).
Figure 5.

Average costs of 100 reserve networks that provided different levels of coverage for climatic refugia and corridors designed to facilitate climate-driven species movements. Error bars span the minimum to the maximum costs across the 100 networks.
Including 20% of the total length of corridors designed to facilitate climate-driven movements resulted in numerous additions to the reserve network and resulted in a more evenly distributed set of sites throughout the USA (figures 1d and 2c). Including these sites for corridors increased the cost of the reserve networks by 17% (figure 3). Protecting additional corridors increased network costs exponentially, with a doubling of costs at somewhere between 60% and 80% of total length of corridors (figure 5b).
4. Discussion
Species will respond to modern climate change by persisting in place, moving to track suitable climates, or going extinct. Expansion of protected area networks in and of itself may allow more species to avoid extinction. However, without explicitly incorporating climate change considerations, such efforts are unlikely to protect all species effectively in the future. In our analysis, expanding the contiguous United States network based solely on species’ current climatic suitability failed to protect climatically suitable areas in the future for 14% of species. Our results also indicate that incorporating both current and future climatically suitable locations would increase the costs by 59%. Further, adding 100% of the climatic refugia and 20% of the modelled climate corridors would increase the cost by an additional 18%. However, requiring higher levels of protection (e.g. additional sites with climatic suitability for each species) or more than approximately 40% of the modelled connectivity resulted in significant cost multipliers. It is important to note that our cost estimates are based on a relatively arbitrary decision to protect 1000 ha at each site and that they do not account for differences in land ownership. For example, one might want to protect more or less land at a given site depending on species-specific needs or logistical constraints and it might be less expensive to protect land already in the public domain. Thus, we caution against reading too much into the dollar costs themselves and focus on the relative costs here.
Climatic refugia identify locations most likely to facilitate species persistence despite broad-scale climatic changes [15]. As the climate continues to change, many of the climatic conditions in which species currently exist are projected to disappear or become more scarce [23]. Fortunately, many of the climatic conditions with shrinking footprints in the USA are already protected. This protection is in part a consequence of the historical conservation of many higher elevation sites across the country—a pattern that has generally been seen as problematic for efficiently protecting biodiversity [44]. In the face of climate change, the higher elevation bias in the distribution of protected areas might prove to be beneficial—in part, by protecting climatic refugia. Our results indicate that capturing this particular type of climatic refugia will be relatively inexpensive, making the protection of rare-climate refugia a relatively low-cost adaptation strategy. That said, there are other types of climatic refugia that are more widely distributed, including both macrorefugia at broad scales [17,21] and microrefugia at finer scales [45]. Conserving these other refugia would probably be more expensive.
For species that need to move to track suitable climates, the permeability of the landscape will be critical. Theoretically, by protecting species in locations that are projected to be climatically suitable today and into the future, the networks we designed should reduce the need for climate-driven movements within the network. Nonetheless, individuals will need to move to climatic refugia and to the new protected areas from other parts of the landscape. By including climate corridors, our goal was to increase the connectivity of the landscape and not necessarily the connectivity of the reserve network itself. Doing the latter would involve requiring the aggregation of sites and/or the selection of corridors between sites [27]. Although connecting the network would probably improve the ability of species to track suitable climates, without directly accounting for climate-driven movements they might not do so. Ideally, a network will both protect species where they are today and where they need to be in the future and connect the routes they will need to take to track suitable climates throughout the network.
The reserve-selection process required us to make a number of relatively subjective decisions. We explored the implications of some of these decisions by including multiple target levels for species protection, refugia and corridor lengths. In addition, we explored a set of scenarios in which we used projected future climate suitability for species from any of the GCMs (instead of requiring suitability across all three) as well as refugia identified using any of the three GCM projections (instead of requiring areas to have been identified as refugia based on all three GCM projections). As one would expect, given the additional flexibility in the solution space, the resulting networks were more similar to the historical networks than the results presented here and the costs of the resulting networks were lower than the networks for species-specific refugia and climatic refugia presented here. Nonetheless, the general patterns and conclusions drawn from these alternative analyses are similar (electronic supplementary material, figure S5).
Although there are some similarities, the reserve networks identified in the present study differ substantially from many of the areas identified in other recent prioritizations of lands in the USA for conservation [17,29,46]. These differences are driven by multiple factors. For example, some priority mapping studies use very different prioritization approaches [29,46]; others do not use any type of biodiversity data [17]; some do not address climate change [29]; and others address climate change but avoid the use of climate-change or climate-impact projections [46]. In addition, of these studies, ours, to our knowledge, is the only one that used projected current or future climatic suitability of individual species. It is important to note that we used a single species distribution modelling approach to project climatic suitability—using a different type of model would possibly alter the spatial patterns of the resulting networks. To that end, one should view these maps not as static blueprints of lands to be protected but as guidance on how and to where conservation priorities will probably need to shift to address climate change. Maps such as these that incorporate climate projections and multiple datasets need to be regularly updated as new projections and data become available and as additional lands are protected.
As our results indicate, protected area networks that are not designed to address climate change will probably fail to protect all species into the future. Incorporating climate change into the conservation-planning process will geographically shift priorities and increase costs. The additional cost, however, might be relatively modest compared to the baseline costs of expanding reserve networks without addressing climate change.
Supplementary Material
Acknowledgements
We thank P. Kareiva and S. Polasky for valuable discussions.
Data accessibility
The data are uploaded to the Dryad Digital Repository: https://doi.org/10.5061/dryad.jm63xsj6d [47].
Authors' contributions
J.J.L. conceived of the study. J.L.M., J.C.W., D.S.R. and H.P.P. helped design the analyses. J.J.L., J.L.M., J.C.W., D.S.R. and C.R.R. conducted the analyses and all authors contributed to the writing of the paper.
Competing interests
We declare we have no competing interests.
Funding
This work has been funded in part by the David and Lucile Packard Foundation (grant no. 2011-36858), the National Science Foundation (grant no. CNH-0814366) and the Wilburforce Foundation through the funding of AdaptWest.
References
- 1.Chen IC, Hill JK, Ohlemueller R, Roy DB, Thomas CD. 2011. Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026. ( 10.1126/science.1206432) [DOI] [PubMed] [Google Scholar]
- 2.Pinsky ML, Worm B, Fogarty MJ, Sarmiento JL, Levin SA. 2013. Marine taxa track local climate velocities. Science 341, 1239–1242. ( 10.1126/science.1239352) [DOI] [PubMed] [Google Scholar]
- 3.Moritz C, Patton JL, Conroy CJ, Parra JL, White GC, Beissinger SR. 2008. Impact of a century of climate change of small-mammal communities in Yosemite National Park, USA. Science 322, 261–264. ( 10.1126/science.1163428) [DOI] [PubMed] [Google Scholar]
- 4.Parmesan C, Yohe G. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42. ( 10.1038/nature01286) [DOI] [PubMed] [Google Scholar]
- 5.Dyderski MK, Paź S, Frelich LE, Jagodziński AM. 2018. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 24, 1150–1163. ( 10.1111/gcb.13925) [DOI] [PubMed] [Google Scholar]
- 6.García Molinos J, et al. 2016. Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Change 6, 83–88. ( 10.1038/nclimate2769) [DOI] [Google Scholar]
- 7.Hole DG, Willis SG, Pain DJ, Fishpool LD, Butchart SHM, Collingham YC, Rahbek C, Huntley B. 2009. Projected impacts of climate change on a continent-wide protected area network. Ecol. Lett. 12, 420–431. ( 10.1111/j.1461-0248.2009.01297.x) [DOI] [PubMed] [Google Scholar]
- 8.Araújo MB, Cabeza M, Thuiller W, Hannah L, Williams PH. 2004. Would climate change drive species out of reserves? An assessment of existing reserve-selection methods. Glob. Change Biol. 10, 1618–1626. ( 10.1111/j.1365-2486.2004.00828.x) [DOI] [Google Scholar]
- 9.Hannah L, Midgley GF, Andelman S, Araujo MB, Hughes G, Martinez-Meyer E, Pearson RG, Williams P. 2007. Protected area needs in a changing climate. Front. Ecol. Environ. 5, 131–138. ( 10.1890/1540-9295(2007)5[131:PANIAC]2.0.CO;2) [DOI] [Google Scholar]
- 10.Heller NE, Zavaleta ES. 2009. Biodiversity management in the face of climate change: a review of 22 years of recommendations. Biol. Conserv. 142, 14–32. ( 10.1016/j.biocon.2008.10.006) [DOI] [Google Scholar]
- 11.Lawler JJ. 2009. Climate change adaptation strategies for resource management and conservation planning. Ann. NY Acad. Sci. 1162, 79–98. ( 10.1111/j.1749-6632.2009.04147.x) [DOI] [PubMed] [Google Scholar]
- 12.Mawdsley JR, O'Malley R, Ojima DS. 2009. A review of climate-change adaptation strategies for wildlife management and biodiversity conservation. Conserv. Biol. 23, 1080–1089. ( 10.1111/j.1523-1739.2009.01264.x) [DOI] [PubMed] [Google Scholar]
- 13.Carvalho SB, Brito JC, Crespo EG, Watts ME, Possingham HP. 2011. Conservation planning under climate change: toward accounting for uncertainty in predicted species distributions to increase confidence in conservation investments in space and time. Biol. Conserv. 144, 2020–2030. ( 10.1016/j.biocon.2011.04.024) [DOI] [Google Scholar]
- 14.Groves CR, et al. 2012. Incorporating climate change into systematic conservation planning. Biodivers. Conserv. 21, 1651–1671. ( 10.1007/s10531-012-0269-3) [DOI] [Google Scholar]
- 15.Keppel G, Van Niel KP, Wardell-Johnson GW, Yates CJ, Byrne M, Mucina L, Schut AGT, Hopper SD, Franklin SE.. 2012. Refugia: identifying and understanding safe havens for biodiversity under climate change. Glob. Ecol. Biogeogr. 21, 393–404. ( 10.1111/j.1466-8238.2011.00686.x) [DOI] [Google Scholar]
- 16.Morelli TL, et al. 2016. Managing climate change refugia for climate adaptation. PLoS ONE 11, e0159909 ( 10.1371/journal.pone.0159909) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Carroll C, Roberts DR, Michalak JL, Lawler JJ, Nielsen SE, Stralberg D, Hamann A, McRae BH, Wang T. 2017. Scale-dependent complementarity of climatic velocity and environmental diversity for identifying priority areas for conservation under climate change. Glob. Change Biol. 23, 4508–4520. ( 10.1111/gcb.13679) [DOI] [PubMed] [Google Scholar]
- 18.Byrne M. 2008. Evidence for multiple refugia at different time scales during Pleistocene climatic oscillations in southern Australia inferred from phylogeography. Quat. Sci. Rev. 27, 2576–2585. ( 10.1016/j.quascirev.2008.08.032) [DOI] [Google Scholar]
- 19.Tzedakis PC, Lawson IT, Frogley MR, Hewitt GM, Preece RC. 2002. Buffered tree population changes in a Quaternary refugium: evolutionary implications. Science 297, 2044–2047. ( 10.1126/science.1073083) [DOI] [PubMed] [Google Scholar]
- 20.Keppel G, Mokany K, Wardell-Johnson GW, Phillips BL, Welbergen JA, Reside AE. 2015. The capacity of refugia for conservation planning under climate change. Front. Ecol. Environ. 13, 106–112. ( 10.1890/140055) [DOI] [Google Scholar]
- 21.Stralberg D, Carroll C, Pedlar JH, Wilsey CB, McKenney DW, Nielsen SE. 2018. Macrorefugia for North American trees and songbirds: climatic limiting factors and multi-scale topographic influences. Glob. Ecol. Biogeogr. 27, 690–703. ( 10.1111/geb.12731) [DOI] [Google Scholar]
- 22.Belote RT, Carroll C, Martinuzzi S, Michalak J, Williams JW, Williamson MA, Aplet GH. 2018. Assessing agreement among alternative climate change projections to inform conservation recommendations in the contiguous United States. Sci. Rep. 8, 9441 ( 10.1038/s41598-018-27721-6) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Michalak JL, Lawler JJ, Roberts DR, Carroll C. 2018. Distribution and protection of climatic refugia in North America. Conserv. Biol. 32, 1414–1425. ( 10.1111/cobi.13130) [DOI] [PubMed] [Google Scholar]
- 24.Littlefield CE, Krosby M, Michalak JL, Lawler JJ. 2019 Connectivity for species on the move: supporting climate-driven range shifts. Front. Ecol. Environ. 17, 270–278. ( 10.1002/fee.2043) [DOI] [Google Scholar]
- 25.Williams PH, Hannah L, Andelman SJ, Midgley GF, Araújo MB, Hughes G, Manne L, Martinez-Meyer E, Pearson RG. 2005. Planning for climate change: identifying minimum-dispersal corridors for the Cape Proteaceae. Conserv. Biol. 19, 1063–1074. ( 10.1111/j.1523-1739.2005.00080.x) [DOI] [Google Scholar]
- 26.Jones KR, Watson JEM, Possingham HP, Klein CJ. 2016. Incorporating climate change into spatial conservation prioritisation: a review. Biol. Conserv. 194, 121–130. ( 10.1016/j.biocon.2015.12.008) [DOI] [Google Scholar]
- 27.Game ET, Lipsett-Moore G, Saxon E, Peterson N, Sheppard S. 2011. Incorporating climate change adaptation into national conservation assessments. Glob. Change Biol. 17, 3150–3160. ( 10.1111/j.1365-2486.2011.02457.x) [DOI] [Google Scholar]
- 28.CBI (The Conservation Biology Institute). 2010. PAD-US 1.1 (CBI edition). Corvallis, OR: CBI. [Google Scholar]
- 29.Withey JC, et al. 2012. Maximizing return on conservation investment in the conterminous USA. Ecol. Lett. 15, 1249–1256. ( 10.1111/j.1461-0248.2012.01847.x) [DOI] [PubMed] [Google Scholar]
- 30.Homer C, et al. 2007. Completion of the 2001 national land cover database for the conterminous United States. Photogramm. Eng. Remote Sens. 73, 337–341. [Google Scholar]
- 31.GBIF.org. 2016. Global biodiversity information facility occurrence download. See gbif.org.
- 32.Phillips SJ, Dudík M, Elith J, Graham CH, Lehmann A, Leathwick J, Ferrier S. 2009. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 19, 181–197. ( 10.1890/07-2153.1) [DOI] [PubMed] [Google Scholar]
- 33.Donner LJ, et al. 2011. The dynamical core, physical parameterizations, and basic simulation characteristics of the atmospheric component AM3 of the GFDL global coupled model CM3. J. Clim. 24, 3484–3519. ( 10.1175/2011JCLI3955.1) [DOI] [Google Scholar]
- 34.Volodin EM, Dianskii NA, Gusev AV. 2010. Simulating present-day climate with the INMCM4.0 coupled model of the atmospheric and oceanic general circulations. Izv. Atmospheric Ocean. Phys. 46, 414–431. ( 10.1134/S000143381004002X) [DOI] [Google Scholar]
- 35.Watanabe M, et al. 2010. Improved climate simulation by MIROC5: mean states, variability, and climate sensitivity. J. Clim. 23, 6312–6335. ( 10.1175/2010JCLI3679.1) [DOI] [Google Scholar]
- 36.Fick SE, Hijmans RJ. 2017. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. ( 10.1002/joc.5086) [DOI] [Google Scholar]
- 37.McGuire JL, Lawler JJ, McRae BH, Nuñez TA, Theobald DM. 2016. Achieving climate connectivity in a fragmented landscape. Proc. Natl Acad. Sci. USA 113, 7195–7200. ( 10.1073/pnas.1602817113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nuñez TA, Lawler JJ, Mcrae BH, Pierce DJ, Krosby MB, Kavanagh DM, Singleton PH, Tewksbury JJ. 2013. Connectivity planning to address climate change. Conserv. Biol. 27, 407–416. ( 10.1111/cobi.12014) [DOI] [PubMed] [Google Scholar]
- 39.Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978. ( 10.1002/joc.1276) [DOI] [Google Scholar]
- 40.Theobald DM. 2013. A general model to quantify ecological integrity for landscape assessments and US application. Landsc. Ecol. 28, 1859–1874. ( 10.1007/s10980-013-9941-6) [DOI] [Google Scholar]
- 41.Wildlife Conservation Society (WCS), Center for International Earth Science Information, Network (CIESIN). 2005. Last of the Wild Project, Version 2. 2005 (LWP-2): Global Human Influence Index (HII) Dataset (Geographic). See sedac.ciesin.columbia.edu/data/set/wildareas-v2-human-influence-index-geographic (accessed 12 June 2012).
- 42.Omernik JM, Griffith GE. 2014. Ecoregions of the conterminous United States: evolution of a hierarchical spatial framework. Environ. Manage. 54, 1249–1266. ( 10.1007/s00267-014-0364-1) [DOI] [PubMed] [Google Scholar]
- 43.Ball IR, Possingham HP, Watts M. 2009. Marxan and relatives: software for spatial conservation prioritisation. In Spatial conservation prioritisation: quantitative methods and computational tools (eds A Moilanen, KA Wilson, HP Possingham), pp. 185–195. Oxford, UK: Oxford University Press. [Google Scholar]
- 44.Scott JM, Davis FW, McGhie RG, Wright RG, Groves C, Estes J. 2001. Nature reserves: do they capture the full range of America's biological diversity? Ecol. Appl. 11, 999–1007. ( 10.1890/1051-0761(2001)011[0999:NRDTCT]2.0.CO;2) [DOI] [Google Scholar]
- 45.Dobrowski SZ. 2011. A climatic basis for microrefugia: the influence of terrain on climate. Glob. Change Biol. 17, 1022–1035. ( 10.1111/j.1365-2486.2010.02263.x) [DOI] [Google Scholar]
- 46.Belote RT, Dietz MS, Jenkins CN, McKinley PS, Irwin GH, Fullman TJ, Leppi JC, Aplet GH. 2017. Wild, connected, and diverse: building a more resilient system of protected areas. Ecol. Appl. 27, 1050–1056. ( 10.1002/eap.1527) [DOI] [PubMed] [Google Scholar]
- 47.Lawler JJ, Rinnan DS, Michalak JL, Withey JC, Randels CR, Possingham HP.2020. Data from: Planning for climate change through additions to a national protected area network: implications for cost and configuration. Dryad Digital Repository. ( ) [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Lawler JJ, Rinnan DS, Michalak JL, Withey JC, Randels CR, Possingham HP.2020. Data from: Planning for climate change through additions to a national protected area network: implications for cost and configuration. Dryad Digital Repository. ( ) [DOI] [PMC free article] [PubMed]
Supplementary Materials
Data Availability Statement
The data are uploaded to the Dryad Digital Repository: https://doi.org/10.5061/dryad.jm63xsj6d [47].