

Abstract
Freshness, although it is a concept difficult to define in wines, can be understood as a combination of different circumstances. Organolepticwise, bluish red, floral and fruity, more acidic and full-bodied wines, are perceived as younger and fresher by consumers. In traditional winemaking processes, these attributes are hard to boost if no other technology or biotechnology is involved. In this regard, the right selection of yeast strains plays an important role in meeting these parameters and obtaining wines with fresher profiles. Another approach in getting fresh wines is through the use of novel non-thermal technologies during winemaking. Herein, the contributions of non-Saccharomyces yeasts and emerging technologies to these parameters are reviewed and discussed.
Keywords: non-Saccharomyces, wine, yeast metabolites, biopolymers, sensory quality, freshness
1. Introduction
Wine freshness is a complex concept comprising mouth, smell and visual perceptions [1]. In this regard, the freshness of the wine could be defined by the combination of a fruity aroma reminiscent of the grape variety, moderate ethanol content and high acidity. Therefore, it is not only a question of achieving a refreshing flavour in the wine, but also of preserving the typicity of the grape variety. The freshness of wine can be reached from different microbiological perspectives, for instance, by using different non-Saccharomyces yeasts that produce organic acids [2]. It can also be reached through the ability of microorganisms, yeasts and bacteria, that synthesise and subsequently free enzymes capable of releasing volatile thiols (e.g., 3-mercaptohexan-1-ol and 3-mercaptohexyl acetate) or terpenes (e.g., geraniol, linalool) [3], thus contributing to the fresh aroma of the wine. Similarly, the freshness of the wine could also be correlated to some extent with increased production of acetate esters [4].
Fruity and floral aroma are responsible for wine aromatic freshness and they are strongly connected with the production of acetate esters from higher alcohols or short chain fatty acids ethanolic esters during fermentation [5]. Selected Saccharomyces strains can help to improve the concentrations of these compounds, but especially several non-Saccharomyces species are able to increase them significantly during fermentation. Among them, Torulaspora delbrueckii (Td) [6], Wickerhamomyces anomalus (Wa) [7], Metschnikowia pulcherrima (Mp) [8], Hanseniaspora vineae (Hv) and Hanseniaspora/Kloeckera spp. [9], Lachancea thermotolerans [2,10], or Candida stellata (Cs) [11] have proven their effectiveness. Some of them also have the ability to release varietal aroma from precursors such as glycosylated terpenes or bonded thiols by means of β-glucosidase [12] or C-S-lyase activities [13]. Many of these yeast species are commercialised as dry or frozen products, while others will be available in the near future. Moreover, the number of available selected strains of these species is progressively growing in the market.
Concerning the sense of taste, freshness is strongly connected with acidity. Acid wines are perceived as more refreshing than low acidity-high pH wines which frequently show heavy and winey profiles. Acidity can be improved by a biological way during fermentation. Some yeasts are able to produce malic acid and lactic acid affecting wine’s pH, but also some other acids in lower concentrations such as succinic acid, pyruvic acid, etc., without significant repercussions to the pH values. The production of malic acid is quite typical in many strains of Saccharomyces cerevisiae (Sc), however, the production is low, usually below 1 g/L [14]. Malic acid is unstable because it can be metabolised by lactic acid bacteria and its technical utility is quite low. Conversely, lactic acid is an interesting option because some non-Saccharomyces yeasts as Lachancea thermotolerans (Lt) can produce it in high concentration [2], even in oenological conditions at a variable range of pH [15]. Moreover, this acid is stable along the winemaking processes and its sensory perception is good [1] and, unlike general belief, it can be described as citric fruit acidity without dairy notes [15].
Freshness can also be related to colour. Red-brown hues make consumers perceive wines as oxidised, therefore colour hue and intensity influence the sensory profile. Yeast can affect colour in several aspects [16]: (i) production of organic acids with repercussion on pH increases the colour of anthocyanins by hyperchromic effect, but it also affects colour stability [2], (ii) the release of yeast metabolites or the expression of hydroxycinnamate decarboxylase activities promote the formation of stable pigments like vitisin or vinylphenolic adducts, respectively [17,18], (iii) the enhancement of polymeric pigments formation [19,20], (iv) the removal of anthocyanins by cell walls adsorption during fermentation [21,22,23], and (v) the production of reductive compounds as glutathione (GSH) with protective effects on wine anthocyanins [2]. The use of some non-Saccharomyces species has proven effective in colour protection through pH modification or the formation of stable pyranoanthocyanins or polymeric pigments.
Wine structurewise, there is a release of yeast polysaccharides during fermentation and also during ageing on lees (AOL). AOL is an interesting technique that involves contact between the lees and the wine after fermentation. In addition, AOL protects the fruity aroma due to its reductive properties [24]. The use of non-Saccharomyces is a current biotechnology to increase the polysaccharide content and to speed up the AOL process [25,26]. Moreover, emerging technologies such as ultrasounds [27,28,29] or ultra-high pressure homogenization [30] facilitate the release of yeast cell wall polysaccharides during AOL. Nonetheless, the use of AOL counteracts freshness by modulating the acidity.
The classification of some non-Saccharomyces used in wine biotechnology can be done by molecular techniques according to their phylogenetic relations. The classification of yeast species and genera was formerly assigned from phenotypical analysis in function of morphology of vegetative cells and sexual states, and physiological responses in growth media and fermentation essays; at present, this classification is given by gene sequencing [31] or genotype, and this is the reason why yeasts species with similar phenotype are grouped in different clusters/branches of phylogenetic trees (Figure 1).
Figure 1.

Phylogenetic relationships among wine yeast species based on analysis of D1/D2 LSU rRNA gene sequences. The evolutionary history was inferred by using the Maximum Likelihood method based on the Tamura-Nei model in MEGA7. GenBank accession numbers follow strain numbers: Saccharomyces cerevisiae NRRL Y-12632/AY048154; Metschnikowia pulcherrima NRRL Y-7111/U45736 Metschnikowia fructicola B-4(1)/EU441890; Lachancea thermotolerans CBS 2803/KY108273; Torulaspora delbrueckii NRRL Y-866/U72156; Wickerhamomyces anomalus NRRL Y-366/U74592; Pichia kluyveri NRRL Y-11519/U75727; Hanseniaspora uvarum NRRL Y-1614/U84229; Hanseniaspora opuntiae CBS 8733/AJ512453; Hanseniaspora vineae NRRL Y-17529/U84224; Hanseniaspora osmophila NRRL Y-1613/U84228; Hanseniaspora guilliermondii NRRL Y-1625/U84230; Schizosaccharomyces pombe NRRL Y-12796/AY048171; Starmerella bombicola 16-D-2/KF935227; Starmerella bacillaris CBS 1713/KY109779.
One of the main drawbacks in the oenological use of non-Saccharomyces is the difficulty in reaching suitable implantations in musts with native microflora. Thus, their metabolome expression is limited and so is the release of metabolites and the production of enzymatic activities with sensory repercussion. Emerging non-thermal technologies open the door to eliminate the wild microbiome from grapes allowing the suitable implantation of these non-Saccharomyces yeasts [32]. Many of these techniques such as high hydrostatic pressures, ultra-high pressure homogenization, β-irradiation, pulsed electric fields, etc., also have concomitant advantages as the enhancement of the extraction of volatile compounds and polyphenols from grapes and the inactivation of oxidative enzymes allowing the reduction in sulphites content.
2. Influence of Non-Saccharomyces Yeasts on Wine Aroma
The formation of fruity or floral aroma during fermentation is an important contribution of non-Saccharomyces yeasts to wine freshness. Many esters with sensory impact are formed as a consequence of the amino acids catabolism by the Ehrlich transamination pathway (Figure 2). The higher alcohols synthesised in this process can be esterified with acetic acid to produce low threshold aromatic esters (e.g., 2-phenylethyl acetate, isoamyl acetate, isobutyl acetate), most of them with floral or fruity descriptors (e.g., rose petals, banana, pear). Moreover, several acids and short chain fatty acids (e.g., lactic, acetic, butyric) are also formed during the yeast metabolism. Some of them produce fruity or sweet profiles (e.g., strawberry, toffee), enhancing freshness and reducing the neutral and winey profile of the conventional S. cerevisiae strains.
Figure 2.

Metabolic pathways involved in the formation of floral and fruity esters in yeasts. Production of acetate esters by Ehrlich catabolism of amino acids and production of fatty acids ethyl esters (TA: transaminase, KDC: 2-keto acid decarboxylase, ADH: alcohol dehydrogenase, Atf: acyltransferase). Formation of fatty acid ethyl esters by acylation with acyl-CoA (PDC: pyruvate decarboxylase, ADH: alcohol dehydrogenase, Eeb1/Eht1: ethyl ester biosynthesis/ethanol hexanoyl transferase).
The production of aromatic compounds by yeasts has been previously reviewed in several key studies that highlight the role of the production of acetate esters from higher alcohols, fatty acid ethyl esters (Figure 2) and the enzymatic activities that release terpenes and thiolic compounds from cysteine-derivatives [5,33,34].
Regarding the H. vineae species, the main esters with sensory repercussion from its exometabolome are benzyl acetate, 2-phenylethyl acetate, ethyl lactate and 3-methylbutyl acetate [35]. Among them, the first two esters are released in higher concentrations (2–7×) than S. cerevisiae; by comparison, ethyl lactate is usually produced below the sensory threshold. Formation de novo of benzyl acetate and 2-phenylethyl acetate from sugars by the chorismate-prephenate pathway has been reported ([36], Figure 3). It has been observed that nitrogen nutrition affected the formation of benzyl and 2-phenylethyl acetates. High doses of diammonium phosphate (100–250 mg/L) inhibited the production of these phenylpropanoid compounds and negatively affected the global balance of aromatic esters [37]. Benzyl alcohol, precursor of benzyl acetate, can be obtained de novo without using grape precursors by 11 different H. vineae strains at concentrations 20–200 fold higher than with S. cerevisiae strains [36].
Figure 3.

Metabolic pathways involved in the de novo synthesis from sugars of floral esters by H. vineae following the chorismate-prephenate-(S)-mandelate/phenylalanine pathway (ChM: chorismate mutase, PrD: prephenate dehydratase, hmaS: hydroxymandelate synthase, hmO: hydroxymandelate oxidase, bDC: benzoylformate decarboxylase, aDH: benzyl alcohol dehydrogenase; aAT: aromatic aminotransferase; Atf: acyltransferase). Adapted from [36,38,39].
Currently, Torulaspora delbrueckii is a non-Saccharomyces yeast with great oenological potential due to its good fermentation performance with low levels of ethanol, higher alcohols, volatile fatty acids and volatile acidity, and its notable contribution to the aromatic profile of the wine [6,40]. Some authors have found a synergistic effect between T. delbrueckii and S. cerevisiae when fermenting in co-inoculation with a high proportion of the non-Saccharomyces yeast, leading to enhanced aromatic properties in the wines [40,41]. In addition, certain volatile compounds contributed by T. delbrueckii have been positively related to the fruity character of the wine. For instance, linalyl acetate, with citrus aroma descriptor, far exceeded its odour threshold (0.05 mg/L) in white Muscat fermentations by T. delbrueckii [40]. Similarly, sequential fermentation between T. delbrueckii and S. cerevisiae resulted in higher concentrations of 3-mercaptohexan-1-ol (grape fruit and passion fruit descriptors [42]) and its acetate (boxwood and passion fruit descriptors [43]) [41]. 3-Ethoxy-1-propanol is another interesting volatile compound that is produced specifically by T. delbrueckii and can contribute to the fruity character of wine with a descriptor of blackcurrant aroma [44,45]. However, in most cases it is not possible to state with complete certainty that a non-Saccharomyces yeast is responsible for specific changes in the volatile profile (always in the same way), as other important factors may be involved such as the strain variability factor or the ratio of dominance against other fermentative yeasts, especially in comparison with Saccharomyces cerevisiae [6]. Therefore, it is of great interest to know those species or strains that tend to preserve the fresh-fruit aromatic intensity coming from the grape variety, as well as the working conditions under which these effects are enhanced.
Pichia kluivery is another yeast species capable of releasing varietal aromas. Among these aroma precursors are thiol-type varietal aromas: 4-methyl-4-mercaptopentan-2-one (4M4MP-boxwood, broom), 3-mercaptohexanol (3MH-grapefruit), 3-mercaptohexyl acetate (3MHA-passion fruit) [42]. P. kluivery produces thiol aromas in a concentration similar to S. cerevisiae strains in single fermentations, but it is able to enhance the production of such compounds in co-inoculations with S. cerevisiae strains [46]. The production of 3MH increases from 625 ng/L to 3000 ng/L while the production of 3MHA increases from 500 ng/L to 1700 ng/L from single to co-inoculation. respectively.
Metschnikowia pulcherrima, like the aforementioned non-Saccharomyces yeast species, are also able to influence the aroma profile of wines and, therefore, to have an impact in the sensory quality of wines [47]. M. pulcherrima contributes to the volatile fraction with the production of aroma compounds due to its β-glucosidase activity [48]. In addition, some M. pulcherrima strains were also reported to have β-xylosidase activity [48,49] which increases the enzymatic activity during fermentation. In this regard, M. pulcherrima seems prone to releasing more monoterpenols and 2-phenyl ethanol than other yeast species such as H. guilliermondii [48]. Linalool, geraniol and nerol are among these monoterpenol compounds.
The following table summarises some of these metabolites mainly contributed by non-Saccharomyces yeasts (Table 1).
Table 1.
Main metabolites of non-Saccharomyces yeasts, sensory repercussion and technical impact.
| Non-Saccharomyces Species | Metabolite/ Biopolymer |
Structure | Sensory Repercussion | Technical Impact | Sensory Olfactive Threshold (µg/L)1 * If usually above This Value |
Reference |
|---|---|---|---|---|---|---|
| Hanseniaspora/Kloeckera | 2-Phenylethyl acetate |
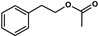
|
Floral, rose petals hints | Enhance floral notes x2-10 compared to S. cerevisiae |
250 * | [35,37] |
| Mannans | Cell wall polysaccharides, mannoproteins | Increased mouthfeel, even perceptible after fermentation | [50] | |||
| Hanseniaspora vineae | Benzyl acetate |
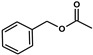
|
Floral jasmine aroma | Floral | 2 * | [35,51] |
| Lachancea thermotolerans | 2-Phenylethyl acetate |
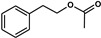
|
Floral, rose petals hints | 10–50 mg/L | 250 * | [15] |
| Ethyl lactate |
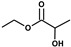
|
Strawberry, toffee | >40 mg/L High sensory threshold |
150,000 | [15] | |
| Lactic acid |
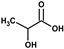
|
Citric acidity | 0.3–16 g/L Up to 0.5 pH reductions in oenological conditions Slight sugar depletion with some alcohol reduction |
[15,52] | ||
| Metschnikowia pulcherrima | 2-Phenylethanol |
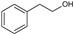
|
Rose-like odour | >30 mg/L | 14,000 * | [48] |
| Monoterpens (e.g., linalool) |
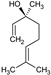
|
Floral | Increase varietal aromas by hydrolysing glucoside terpenes | 25 * | [48] | |
| Pichia kluyveri | Mercaptohexanol (3-MH) |
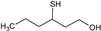
|
Grapefruit, passion fruit | Fruity smell: > 625 ng/L single fermentation to 3000 ng/L co-inoculation | 0.060 * | [46] |
| Mercaptohexyl acetate (3-MHA) |
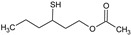
|
Grapefruit, passion fruit | Fruity smell: > 500 ng/L single fermentation to 1700 ng/L co-inoculation | 0.004 * | [46] | |
| Schizosaccharomyces pombe | Pyruvate |
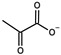
|
Stable pigments, colour stability Precursor for vitisin A type compounds: 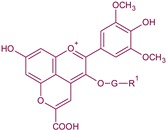
|
Enhance the formation of vitisin A derivatives Some strains also vinylphenolic pyranoanthocyanins |
[53] | |
| Cell wall polysaccharides, mannoproteins | Better wine structure, softening of the astringency | Increased mouthfeel | ||||
| Torulaspora delbrueckii | 2-Phenylethyl acetate |
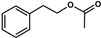
|
Flower, honey | 1.2-2x compared to S. cerevisiae & S. uvarum | 250 * | [42,44] |
| Ethyl hexanoate |
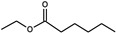
|
Apple | Fruity smell | 62 * | [44] | |
| 3-Ethoxy-1-propanol |
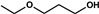
|
Black currant, solvent | Black fruity smell | [44] | ||
| Wickerhamomyces anomalus | 2-phenylethyl acetate |
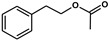
|
Flower, honey | 250 * | [42] | |
| Isoamyl acetate |
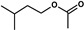
|
Banana | Enhance fruitiness | 30 * | [42] | |
| Ethyl acetate |
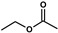
|
Fruity at low concentration | Fruity smell at low concentration Enhance complexity |
12,300 * | [54] |
3. Acidity and pH Control
The effect of L. thermotolerans in wine freshness is especially significant because of the influence on pH by the production of lactic acid; some strains are capable of producing more than 16 g/L [52]. Recent reviews have highlighted the oenological relevance of this species [2,10]. The phenotypic and genotypic variability of strains from many origins worldwide have been studied [55,56]. The formation of lactic acid is derived from pyruvate in the glycolytic metabolism of sugars. The enzyme lactate dehydrogenase is involved in the reduction of pyruvate to lactic acid and it is a typical feature found in many strains of L. thermotolerans [15]. This acidification happens at the beginning of the alcoholic fermentation, 3–5 days of fermentation [2], with a significant effect when the population exceeds 6-log CFU/mL. Effective pH reductions have been reported in mixed fermentations with inoculation ratio 7-log:3-log (Lt:Sc) [57]. The early acidification helps L. thermotolerans to be competitive even when high wild populations are present in the must during fermentation. Nitrogen contents also affect the production of lactic acid; YAN values closed to 200 mg/L improve lactic acid production [2]. Strong effects in pH have been observed in Tempranillo red wines with initial pH ca. 4; lactic acid levels exceeded 6 g/L when specific L. thermotolerans strain was used and the pH value decreased around 0.5 units [15]. L. thermotolerans has been described as low volatile acidity producer, ranging most of the strains 0.3-0.5 mg/L [15,57,58], and also as an interesting bio-tool to control the acetic acid production [59]. Since lactic acid production can come from sugar depletion, there could be a slight reduction of alcoholic degree (0.3–0.5% vol.) [15]. Current literature describes fermentative performances of around 8–10% vol. [56,57], however, this can be improved with appropriate strain selection. Single pure fermentations with L. thermotolerans, isolated and selected in ongoing research works, might be possible since these strains are able to yield 12–13% vol. ethanol.
Lactic acid can be described as citric fruit acidity without providing the wines with dairy notes. The perceived effect is the enhancement of the freshness in mouth. Dairy notes resembling compounds, such as diacetyl or acetoin, are produced at similar concentration when used in sequential inoculations than in S. cerevisiae pure fermentations [15].
Even when non-Saccharomyces are described as low sulphite-resistant, many L. thermotolerans strains have the capacity to grow and ferment in the presence of sulphites. Some strains can ferment in the presence of conventional concentrations of 50–60 mg/L of total SO2 with a slight reduction in the final alcoholic degree. It is also possible to ferment at slower rate under 18 °C with 70 mg/L of total SO2 and 15 mg/L of free SO2 which resembles winemaking conditions.
4. Non-Saccharomyces-Mediated Formation of Stable Pyranoanthocyanin and Polymeric Pigments
Different yeast genera may contribute to the formation of stable pigments either during the fermentation or during the ageing period. In this regard, anthocyanins extracted from the grape’s skins are transformed through condensation reactions into less sensitive pigments against pH variations in wines, addition of SO2 to improve the wine’s stability and temperature fluctuations during the product’s shelf life. Among the species known for having this activity are S. pombe due to an increase of vitisin A formation [60]; T. delbrueckii and M. pulcherrima that favour the production of oligomeric fractions in sequential fermentations [19]; L. thermotolerans that is able to increase the amount of lactic acid during must fermentation [2], which pH reduction improves the colour intensity of wines and its stability by increasing the molecular SO2. It also adsorbs less pigments on the cell wall which produces higher concentration of final total pigments in sequential fermentations [20].
Maloalcoholic fermentation (MAF) produces the metabolisation of malic acid into ethanol. The malic enzyme (ME) of S. pombe has 15-folds higher affinity by malic acid than the ME of S. cerevisiae [61]. This higher efficiency is due to the presence of the specific malate permease transporter in S. pombe (mae1p) facilitating the entrance of malate into the cytosolic media, and also because the ME is located in the cytosol conversely to S. cerevisiae in which this pathway is produced in the mitochondrion [61]. MAF of S. pombe increases the release of extracellular pyruvate facilitating the formation of stable vitisin A-type derivatives [62] by chemical condensation with grape anthocyanins [17] (Figure 4). The higher the extracellular release of pyruvate, the higher the formation of stable pyranoanthocyanins. Pyranoanthocyanins are more stable pigments under oenological conditions not only because of their lower sensibility to SO2 bleaching, but also to their lesser hypochromic pH effect [63].
Figure 4.

Maloalcoholic fermentation by Schizosaccharomyces pombe and its influence in the subsequent formation of stable vitisin A-type pigments by chemical condensation with grape anthocyanins (adapted from [17,61]).
Besides pyruvic acid, acetaldehyde is another fermentative metabolite playing an important role in the formation of stable pigments in red wines. Such is the case of all the pyranoanthocynin-type pigments [18,64], and the oligomers condensed through ethyl bridges [65] (Figure 5). The formation of ethyl linked oligomers happens preferentially with (-)-epicatechin rather than with (+)-catechin in acidic conditions [66]. This mechanism is explained from the slower rate in which the condensation with the latter flavanol takes place. Nonetheless, the stability of oligomeric pigments varies with the absence/presence of acetaldehyde linkages and the configuration of the pigment. The configuration of anthocyanin-flavanol ethyl linked dimers is more stable than the configuration of flavanol-flavanol ethyl linked dimers [67], but both are apparently less stable than the dimers formed from direct condensation of anthocyanin-flavanol moieties [68] which takes place slower over time in aged wines.
Figure 5.

Example of red wine pigments and their occurrence during the different winemaking stages. From top to bottom the pigments shown in the figure are: malvidin-3-glucoside, malvidin-3-glucoside pyruvic acid (vitisin A), malvidin-3-glucoside-4-vinyl (vitisin B), malvidin-3-glucoside 4-vinylcatechol (pinotin A), dimer malvidin-3-glucoside-catechin, dimer malvidin-3-glucoside-ethyl-catechin, malvidin-3-glucoside-pyruvic acid vinyl catechin (portisin type A) and malvidin-3-glucoside pyruvic acid vinyl phenol (portisin type B). Yeasts species contributing to the formation of particular pigments are also indicated.
Pinotins are pigments produced from the reaction of anthocyanins or pyranoanthocyanins with hydroxycinnamic acids or 4-vinyl phenols [69]; in this last case, the yeast strains with positive hydroxycinnamate decarboxylase activity can produce the intermediate compound that will condense via nucleophilic attack at position C4 of the anthocyanin structure [70]. Other pigments, mainly formed during the ageing period, comprise the previous formation of vitisins for further condensation with vinyl-flavan-3-ols and/or hydroxycinnamic acids or 4-vinyl phenols [71]; these reactions would produce the so called portisin-type pigments with bluish tonality due to an absorption at 570 nm [72] of the electromagnetic spectrum.
In the same way that yeast strains promote the formation of pigments, emerging preservation technologies for grape berries/must processing and winemaking could improve the colour in wines (Figure 5). Such is the case of the high hydrostatic pressure (HHP) that increases total anthocyanins extraction and the formation of pyranoanthocyanins [73], especially the amount of vitisin A; the use of pulsed electric fields (PEF) or the ultrasounds (US) that have shown to produce an increase in anthocyanin extraction from cell wall structures after suffering membrane disruption [74,75]; the micro-oxygenation (MO) could also have an impact in the colour of red wines through the formation of vitisins or direct condensation of anthocyanin and tannins where oxygen and reactive species may play an important role [76].
Other information related to the nature and bioavailability of anthocyanins, their properties and industrial applications as well as the formation of pyranoanthocyanins and polymeric pigments has been recently published in reviews [77,78,79,80].
5. Commercially Available Non-Saccharomyces Yeasts
The use of non-Saccharomyces yeasts has grown progressively from the initial applications of T. delbrueckii to enhance aromatic profile with two widely spread strains BiodivaTM Td291 (Lallemand, Blagnac Cedex, France) and PreludeTM (CHR Hansen, Hoersholm, Denmark), and also from the use of S. pombe reticulated in alginate beads for demalication (Proenol), L. thermotolerans (formerly Kluyveromyces thermotolerans) and P. kluyveri (CHR Hansen). Non-Saccharomyces yeasts are frequently offered mixed with S. cerevisiae to ensure the full depletion of the sugars and achieve wine dryness. Main commercial non-Saccharomyces yeast species were described by Morata and Suarez [81], and are shown in an updated list in Table 2.
Table 2.
Non-Saccharomyces species commercially available and main applications in wine production.
| Non-Saccharomyces Species Commercially Available | Brand, Producer, Year, Format | Sensory Repercussion | Fermentative Performance (% vol. Ethanol) Recommended Inoculation Dose |
Application. Requirements. | Reference |
|---|---|---|---|---|---|
| Hanseniaspora vineae | OENOBRANDS Launch 2021 Dry yeast |
Enhance production of fruity and floral esters De novo formation of floral esters from sugars Increased body, softness and roundness |
10% | Low SO2 Nutrition: thiamine and yeast extract |
http://www.oenobrands.com/en/our-innovation |
| Lachancea thermotolerans | CONCERTOTM CHr HANSEN 2012 Dry yeast |
Red and black fruit integration Round mouthfeel Soft acidification Low volatile acidity, H2S and SO2 |
10% 25 g/HL |
Red/White wines from warm areas | https://www.chr-hansen.com/ |
| LEVEL2 LAKTIA™ LALLEMAND 2018 Dry yeast |
Enhancement of aromatic complexity, freshness and acidity Lactic acid production Low volatile acidity High glycerol production |
<10% 25 g/HL |
High nitrogen requirements Free SO2 must be <15 mg/L |
https://www.lallemandwine.com/ | |
| Metschnikowia fructicola | Gaïa™ LALLEMAND 2016 Dry yeast |
Improvement of the sensory expression Preservation of varietal character |
Low to no fermentative power 7–25 g/HL |
Reduce the use of SO2 and volatile acidity Active K2. Biological control Resistant to low pH and 50 mg/L SO2 |
https://www.lallemandwine.com/ |
| Metschnikowia pulcherrima | LEVEL2 FLAVIA® MP346 LALLEMAND 2013 Dry yeast |
Release of varietal aromas | 9% 25 g/HL |
Specific enzymatic activity helping in releasing varietal aromas (terpenes and thiols) Free SO2 must be <15 mg/L |
https://www.lallemandwine.com/ |
| LEVULIA® PULCHERRIMA AEB Dry yeast |
High production of 2-phenyl and isoamyl acetates and terpenes Low volatile acidity |
11.5% 20–50 g/HL |
Release of varietal aromas (terpenes) | https://www.aeb-group.com/ | |
| Pichia kluyveri | FROOTZEN® CHr HANSEN 2010 Frozen yeast |
Enhancement of volatile thiols Blackcurrant, liquorice, black pepper, menthol aromas |
4–5% 1 bag/10kL |
Enhance fruitiness | https://www.chr-hansen.com/ |
| Torulaspora delbrueckii | PRELUDETM CHr HANSEN 2009 Frozen yeast |
Enhance production of medium chain fatty acid esters Increased body, softness and roundness |
9% 25 g/HL |
Enhance fruitiness Promote malolactic fermentation by depletion of medium chain fatty acids |
https://www.chr-hansen.com/ |
| LEVEL2 BIODIVA™ TD291 LALLEMAND 2009 Dry yeast |
Aromatic complexity, ester production Low volatile acidity |
10% 25 g/HL |
Tolerance to osmotic pressure. Adapted for fermenting late harvest and ice wines Free SO2 must be <15 mg/L |
https://www.lallemandwine.com/ | |
| ZYMAFLORE® Alpha LAFFORT 2010 Dry yeast |
Aromatic complexity and good mouthfeel Suitable for making expressive and full-bodied wines Revelation of thiol-type varietal aromas (3MH, 3MHA) |
10% 25 g/HL |
Medium nitrogen requirements Low volatile acidity, volatile phenols and H2S |
https://laffort.com/en/ | |
|
Torulaspora delbrueckii + Saccharomyces spp. |
Oenoferm® wild & pure F3 Erbslöh Dry yeast |
Enhanced mono terpenes and formation of fruity esters Support the ripe and exotic fruit aroma Full-bodied wines |
- 20–40 g/HL |
Moderate to high nitrogen requirement High alcohol tolerance |
https://erbsloeh.com/en/ |
|
Torulaspora delbrueckii +
Metschnikowia pulcherrima |
ZYMAFLORE® ÉGIDE LAFFORT 2017 Dry yeast |
Organoleptic neutrality and control the microflora | 10% 2–5 g/HL |
Bioprotection of grapes and juices Restriction of the growth of indigenous flora SO2 reduction |
https://laffort.com/en/ |
| Schizosaccharomyces pombe | ProMalic®
PROENOL Dry beads |
Wine deacidification | 100g/HL | Maloalcoholic fermentation Free SO2 must be <14 mg/L |
https://www.proenol.com/ |
Ternary inoculations have been proposed at industrial level by Chr. Hansen in the commercial blend of three stains, S. cerevisiae (60%), T. delbrueckii (20%) and L. thermotolerans (60%:20%:20%) known as Viniflora® Melody™. This approach is really interesting to improve freshness and complexity. The use of two non-Saccharomyces species enhances the floral notes and fruitiness and S. cerevisiae guarantees the suitable end of fermentation. Moreover, when an acidifier yeast is used, such as L. thermotolerans, the wine freshness is also enhanced.
6. Non-Saccharomyces and Off-Flavour Production
Undesired aroma compounds could be produced during fermentation by non-Saccharomyces yeasts (Table 3). These volatile compounds comprise large amounts of volatile acidity, sulphur compounds, excessive fusel alcohol aromas, etc. Among yeast strains known as sulphydric acid producers (H2S), as in the case of T. delbrueckii strains [82], some M. pulcherrima isolates have been studied due to their low ability to produce such volatile compound [83]. The production of H2S has been studied in 9 strains of T. delbrueckii, all them ranked 3–4 in a 0 to 5 scale showing stronger production of this defective compound compared with S. cerevisiae and other oenological non-Saccharomyces yeasts [57]. Other odd smells related to the metabolism of M. pulcherrima are produced by aliphatic carboxylic acids such as hexanoic acid and octanoic acid [84]. The aroma associated to these compounds resembles fatty and cheese for the first acid, and rancid or harsh for the second one.
Table 3.
Non-Saccharomyces yeast species involved in the production of undesired volatile compounds in wines.
| Non-Saccharomyces species | Metabolite | Structure | Off-Smell/Off-Flavour | Technical Impact. Sensory Threshold (µg/L) 1 |
Reference |
|---|---|---|---|---|---|
| Hanseniaspora/Kloeckera | Acetic acid |
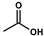
|
Vinegar taste | Many species/strains >0.6 g/L of volatile acidity 300,000 |
[85] |
| Ethyl acetate |
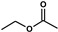
|
Solvent smell, nail varnish | Many species/strains >100 mg/L 12,300 |
[85] | |
| Lachancea thermotolerans | Lactic acid |
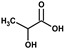
|
Some strains can produce excessive sour taste | Some strains >7 g/L; maximum described 16 g/L | [15,52] |
| Metschnikowia pulcherrima | Acetic acid |
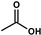
|
Vinegar taste | >0.7 g/L 300,000 |
[86] |
| Ethyl acetate |
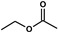
|
Solvent smell, nail varnish | >200 mg/L 12,300 |
[84] | |
| Hexanoic acid |
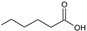
|
Fatty, cheese | >1.1 mg/L 420 |
[84] | |
| Octanoic acid |
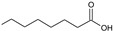
|
Rancid, harsh | >1.2 mg/L 500 |
[84] | |
| Schizosaccharomyces pombe | Acetic acid |
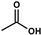
|
Vinegar taste | Many strains >1 g/L of volatile acidity 300,000 |
[62,86] |
| Torulaspora delbrueckii | Sulphydric acid |
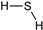
|
Reductive smell, egg rot smell | Impact depending on the intensity 1.1 |
[57,82] |
| Wickerhamomyces anomalus | Ethyl acetate |
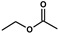
|
Solvent smell, nail varnish | ≈100 mg/L 12,300 |
[85,87] |
| Acetic acid |
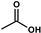
|
Vinegar taste | 0.02 g/L 300,000 |
[86,87,88] |
Apiculate yeasts with high prevalence in grape microbiome are usually described as high producers of volatile acidity. When the production is measured in pure fermentation of 11 strains of different species, some strains of H. osmophila, H. valbiensys and H. uvarum generated 0.6-0.8 mg/L acetic acid. However, other strains of these species, and also H. vineae, produced 0.4-0.5 mg/L [85]. Therefore, specific selection can help to obtain strains with suitable levels of volatile acidity. Many Hanseniaspora species frequently show high values of ethyl acetate ranging from 50 to >300 mg/L [85].
7. Biological Control of Indigenous Yeasts Producing Defective Off-Flavours
Biological control by yeasts is a powerful bio-tool to control spoilage indigenous populations which produces off-flavours and metabolites affecting wine quality and freshness. Several antimicrobial activities have been described in yeasts such as the production of pulcherrimin, killer factors, etc. (Table 4).
Table 4.
Non-Saccharomyces yeast species involved in the production of biocontrol agents.
| Non-Saccharomyces Species | Metabolite | Antimicrobial Effect | Technical Impact | Reference |
|---|---|---|---|---|
| Hanseniaspora uvarum | trans-Cinnamaldehyde | Inhibition of mycelium growth and conidia germination | Biocontrol of Botrytis cinerea | [90,91] |
| Metschnikowia pulcherrima | Pulcherrimin | Iron depletion | Biological control Effective inhibitory activity against several yeasts: Candida tropicalis, Candida albicans, Brettanomyces/Dekkera, Hanseniaspora and Pichia genera; and some fungi: Botrytis cinerea, Penicillium spp., Alternaria spp. and Monilia spp. | [8,93,94,95,96,97,98] |
| Killer factor | Membrane permeabilization | Biological control | [57] | |
| Metschnikowia fructicola | Killer factor: active K2 | Increase of membrane permeability | Biological control of apiculate yeasts: K. apiculate/H. uvarum Reduction of volatile acidity |
www.lallemandwine.com/ |
| Wickerhamomyces anomalus | Exo-β-1,3 glucanase | Wall-lytic enzymes | Inhibition of Botrytis cinerea | [92] |
| Pikt killer toxin | Mycocins that control apiculate wine yeasts | Biological control of Dekkera/Brettanomyces spp. | [92,99] | |
| Torulaspora delbrueckii | Kbarr-1 killer toxin | Toxicity against sensitive strains of S. cerevisiae | Better implantation and domination of the fermentation on the native S. cerevisiae of the must | [45] |
Metschnikowia fructicola is used and commercialised for biological control of apiculate yeasts (K. apiculate/H. uvarum) reducing the formation of volatile acidity at the early stages of fermentation. M. fructicola produces Killer factor K2 which increases membrane permeability, that reduces respiration activity and lowers intracellular ATP content decreasing cell viability [89]. Several volatile compounds produced by Hanseniaspora uvarum have been described as effective inhibitors against the development of Botrytis cinerea [90] and, among them, trans-cinnamaldehyde showed the stronger inhibition of mycelium growth also avoiding conidia germination [91].
The species Wickerhamomyces anomalus has both properties conferred, biocontrol agent and antimicrobial agent due to its activity against moulds in diverse environments and to the production of killer toxins vs. spoilage yeast [92]. In the last decade, a Torulaspora delbrueckii killer strain was successfully isolated and its fermentative performance was assessed against S. cerevisiae [45]. This killer strain prevailed over S. cerevisiae, but only under conditions of must sterility or with high inoculum population. This biocontrol tool is interesting to ensure the metabolic prevalence of non-Saccharomyces yeasts versus the native must yeasts.
8. Emerging Technologies to Improve the Implantation of Non-Saccharomyces
A suitable expression of metabolites during fermentation, and therefore a significant sensory effect, depends on the implantation of the non-Saccharomyces yeasts used. The influence of the wild initial population, usually around 4-log CFU/mL of yeasts and 2-log CFU/mL of bacteria, is a determining factor for the successful implantation of the desired selected yeast. Especially when the non-Saccharomyces to implant is a weak fermenter (M. pulcherrima, H. vineae, L. thermotolerans, etc.) or it has slow fermentative kinetics (e.g., S. pombe). The reduction or elimination of the wild microbiota in the grape is a way to promote the implantation of non-Saccharomyces species, and therefore, to better express their metabolome producing a significant impact in the sensory profile.
The use of non-thermal technologies provides a way to eliminate or control the indigenous population. At the same time, the sensory profile of the grape is preserved and thermal degradation of pigments and aromatic compounds or undesired oxidations are avoided. Some of these technologies can be highly effective in the control of microorganisms, and at the same time extremely gentle with the sensory components [32]. Most of the technologies affect the structure of skin cells facilitating the extraction of tannins, anthocyanins and aroma compounds. The use of High Hydrostatic Pressure (HHP), Ultra High Pressure Homogenization (UHPH), Pulsed Electric Fields (PEF), Ultrasound (US), β-irradiation (βi), Pulsed Light (PL) and Ozonation, favour the control of indigenous yeasts in grape and musts promoting the implantation of weak yeast starters of non-Saccharomyces.
HHP has demonstrated a high efficiency eliminating yeast in crushed grapes reaching 4-log inactivation [100]; however, lactic acid bacteria populations were partially reduced too, and only 1-log remained viable even at 550 MPa-10 min. Several non-Saccharomyces: S. pombe, T. delbrueckii, M. pulcherrima and L. thermotolerans, showed better implantation when grapes are processed at 400 MPa-10min [73]. In addition, HHP increases the extraction of phenols, especially anthocyanins, facilitating faster macerations and wines with higher colour intensity [100,101,102].
UHPH is also a highly efficient technology to eliminate indigenous microorganisms from musts and, conversely to HHP, it is also extremely efficient against bacteria. Initial populations of 6-log CFU/mL yeast and 4-log CFU/mL aerobic and lactic acid bacteria were undetected in 1 mL after continuous processing of the must at 300 MPa [103]. Additionally, UHPH can inactivate oxidative enzymes favouring the reduction of SO2 levels [30].
PEF produces cell wall poration causing microbial inactivation [104], and it also promotes the extraction of phenolic compounds from the cell wall of grape’s skins, especially anthocyanins. The pre-fermentative use of 10 kV/cm increased anthocyanin content and wine colour intensity, and 5 kV/cm affected total polyphenol extraction [105]. PEF technology can be used continuously at industrial scale to facilitate maceration [75,106,107]. The use of PEFs to control microbial populations requires higher intensities, frequently >30 kV/cm [108].
Ultrasound irradiation is also an interesting technique to increase the extraction of phenolic compounds from grapes. However, the antimicrobial efficiency is reduced and it is associated to the thermal effect which produces degradation of sensory quality.
PL is another highly effective non-thermal technology with the ability to destroy vegetative and sporulate forms of microorganisms. The application can be done in a continuous way over the destemmed grape when is selected in a sorting table. The use of this technology can help reducing SO2 levels and improving the implantation of non-Saccharomyces, as well as performing yeast-bacteria co-inoculations [109].
9. Cell Wall Polysaccharides from Non-Saccharomyces Yeasts
The ageing on lees (AOL) process consists in a long contact of lees with wine during the ageing period. The lees are formed essentially by yeast biomass. This technique is traditionally used in the elaboration of sparkling wines after the second fermentation in the bottle [110]. During the AOL, the yeast autolysis is produced. This phenomenon involves the decomposition of the cell membranes and the release of intracellular compounds to the wine. In addition, the cell wall is degraded by the action of enzymes produced by the dead yeast [111]; this includes the mannoproteins, the major cell wall polysaccharide. A major drawback of this technique is the time needed for the autolysis to be completed and the mannoproteins to be released which, in most cases, is of several months under conventional oenological conditions [25,26,112]. Research studies using different techniques to speed up this process include ultrasounds [28,29] and pulsed electric fields [113,114,115]. In this way, the time required for the AOL is shorten.
The different compounds released during yeast autolysis have a direct organoleptic repercussion in the wine aged. The amino acids and peptides act as flavour precursors and malolactic fermentation promoters [116]. A lipid fraction is also released to the wine during AOL; these compounds seem to have an impact on the foaming properties [117]. Regarding the wall polysaccharides, the mannoproteins work against tartaric and protein precipitations [118]. Polysaccharides decrease the astringency perception [119] and increase the body of the wine aged. Finally, the mannoproteins provide a higher monomeric anthocyanins stabilization [120].
The AOL could also impact the freshness perception of wines in terms of acidity and volatile fraction. The AOL seems not to have a direct influence on the acidity of aged wines. A decrease of tartaric acid after AOL has been observed [121], nonetheless, no significant differences were detected in the rest of the acids and titratable acidity. Other authors showed only minor differences in total acidity after 180 days of AOL in Chardonnay white wines [122].
The repercussion of the AOL on the aromatic fraction has been also studied. It has been observed that the AOL significantly increases the volatile compounds in wines from Bombino bianco grapes [123], and it was also observed that after long AOL (30 months), the wines from different grape varieties do not change their varietal aromatic characteristics [124]. In this regard, other aromatic compounds related to freshness perception such as volatile thiols, terpenes, acetate esters of higher alcohols or ethyl esters of short chain fatty acids, have been under study. Volatile thiols (4-methyl-4-mercaptopentanone and 3-mercaptohexanol) were analysed in wines after eight months of AOL in oak barrels [125]. The concentration of these compounds was larger in AOL wines. Therefore, it existed a protective effect on typical fruity aromas found in young wines. This effect could be explained by the increase of the sulphur tripeptide glutathione that play a major role in protecting volatile thiols during the aging of bottled white wines [125]. Terpenoid compounds, important contributors to varietal aroma of wines because of their low perception threshold and their relation with floral odour, increased their content in wines after ten months aged on lees [126]; α-terpineol, E-nerolidol and Z-nerolidol, as an example, significantly increased in Airén variety after contact with lees during ageing [124]. This effect might be possible due to the release of β-glucosidases during the yeast autolysis. Esters, compounds responsible in large part for the fresh and fruity aroma of wine, experienced a significant increase after the AOL of Chardonnay wines [126]. This increase was observed in Airén wines as well [124]. Nonetheless, besides that increase in the concentration of esters during ageing, some of these compounds could be absorbed by the yeast lees after long periods [127].
The nature of the cell wall of the different yeast species is different, therefore the use of non-Saccharomyces in AOL technique will result in wines with different chemical and organoleptic composition. The use of Torulaspora delbrueckii and other yeasts as Saccharomycodes ludwigii and Zygosaccharomyces bailii, both considered spoilage yeasts, can increase the polysaccharides content in wines, especially when spoilage yeasts are used [128]. Other yeast species such as Schizosaccharomyces pombe and Saccharomycodes ludwigii can also modify the polysaccharides content in wine [25]; an increase of ten-folds, in comparison to S. cerevisiae, were observed in wines after only 28 days of ageing when using both species, S. pombe and S. ludwigii; despite these observation, better results were obtained with S. pombe, S. ludwigii and D. bruxellensis compared to the control Saccharomyces strain [26].
10. Conclusions
If the 20th century was the time of S. cerevisiae, then the 21st is the time of non-Saccharomyces yeasts. The applications and commercial relevance of these species is becoming increasingly important to develop new opportunities to improve wine’s quality. Today’s research lines are mainly focused on the use of non-Saccharomyces yeasts to enhance sensory quality, including wine aroma, colour and structure. To describe wine’s quality and stability, is to talk about molecules with sensory repercussion or microbiological and physicochemical properties that are highly influenced by pH variations. The natural production of organic acids by some non-Saccharomyces yeasts helps to get safer and more stable wines even during barrel and bottle ageing. Moreover, the biocontrol properties of these yeasts enables the safe production of wines by the reduction of the content of wine preservatives such as sulphites. In this regard, the use of emerging non-thermal physical technologies would also become the basis for the new oenology with low use of chemical additives and a more controlled fermentative microbiota.
Funding
Ministerio de Ciencia, Innovación y Universidades project: RTI2018-096626-B-I00 and European Regional Development Fund (ERDF), through the National Smart Growth Operational Programme FEDER INTERCONECTA EXP-00111498/ITC-20181125, project: FRESHWINES.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Morata A., Loira I., Del Fresno J.M., Escott C., Bañuelos M.A., Tesfaye W., González C., Palomero F., Suárez-Lepe J.A. Strategies to Improve the Freshness in Wines from Warm Areas. In: Morata A., Iris L., editors. Advances in Grape and Wine Biotechnology. InTech; London, UK: 2019. [Google Scholar]
- 2.Morata A., Loira I., Tesfaye W., Bañuelos M.A., González C., Suárez Lepe J.A. Lachancea thermotolerans applications in wine technology. Fermentation. 2018;4:53. doi: 10.3390/fermentation4030053. [DOI] [Google Scholar]
- 3.Delfini C., Cocito C., Bonino M., Schellino R., Gaia P., Baiocchi C. Definitive evidence for the actual contribution of yeast in the transformation of neutral precursors of grape aromas. J. Agric. Food Chem. 2001;49:5397–5408. doi: 10.1021/jf010613a. [DOI] [PubMed] [Google Scholar]
- 4.Jackson R. Wine Science: Principles and Applications. 3rd ed. Academic Press; London, UK: 2008. p. 751. [Google Scholar]
- 5.Dzialo M.C., Park R., Steensels J., Lievens B., Verstrepen K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017;41:S95–S128. doi: 10.1093/femsre/fux031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ramírez M., Velázquez R. The Yeast Torulaspora delbrueckii: An Interesting But Difficult-To-Use Tool for Winemaking. Fermentation. 2018;4:94. doi: 10.3390/fermentation4040094. [DOI] [Google Scholar]
- 7.Padilla B., Gil J., Manzanares P. Challenges of the Non-Conventional Yeast Wickerhamomyces anomalus in Winemaking. Fermentation. 2018;4:68. doi: 10.3390/fermentation4030068. [DOI] [Google Scholar]
- 8.Morata A., Loira I., Escott C., del Fresno J.M., Bañuelos M.A., Suárez-Lepe J.A. Applications of Metschnikowia pulcherrima in Wine Biotechnology. Fermentation. 2019;5:63. doi: 10.3390/fermentation5030063. [DOI] [Google Scholar]
- 9.Martin V., Valera M., Medina K., Boido E., Carrau F. Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review. Fermentation. 2018;4:76. doi: 10.3390/fermentation4030076. [DOI] [Google Scholar]
- 10.Porter T.J., Divol B., Setati M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019;119:378–389. doi: 10.1016/j.foodres.2019.02.003. [DOI] [PubMed] [Google Scholar]
- 11.García M., Esteve-Zarzoso B., Cabellos J., Arroyo T. Advances in the Study of Candida stellata. Fermentation. 2018;4:74. doi: 10.3390/fermentation4030074. [DOI] [Google Scholar]
- 12.Rosi I., Vinella M., Domizio P. Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 1994;77:519–527. doi: 10.1111/j.1365-2672.1994.tb04396.x. [DOI] [PubMed] [Google Scholar]
- 13.Zott K., Thibon C., Bely M., Lonvaud-Funel A., Dubourdieu D., Masneuf-Pomarede I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011;151:210–215. doi: 10.1016/j.ijfoodmicro.2011.08.026. [DOI] [PubMed] [Google Scholar]
- 14.Zelle R.M., De Hulster E., Van Winden W.A., De Waard P., Dijkema C., Winkler A.A., Geertman J.M.A., Van Dijken J.P., Pronk J.T., Van Maris A.J.A. Malic acid production by Saccharomyces cerevisiae: Engineering of pyruvate carboxylation, oxaloacetate reduction, and malate export. Appl. Environ. Microbiol. 2008;74:2766–2777. doi: 10.1128/AEM.02591-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Morata A., Bañuelos M.A., Vaquero C., Loira I., Cuerda R., Palomero F., González C., Suárez-Lepe J.A., Wang J., Han S., et al. Lachancea thermotolerans as a tool to improve pH in red wines from warm regions. Eur. Food Res. Technol. 2019;245:885–894. doi: 10.1007/s00217-019-03229-9. [DOI] [Google Scholar]
- 16.Morata A., Loira I., Suárez Lepe J.A. Influence of Yeasts in Wine Colour. In: Morata A., editor. Grape and Wine Biotechnology. InTech; London, UK: 2016. pp. 285–305. [Google Scholar]
- 17.Morata A., Gómez-Cordovés M.C., Colomo B., Suárez J.A. Pyruvic Acid and Acetaldehyde Production by Different Strains of Sacchromyces cerevisiae: Relationship with Vitisin A and B Formation in Red Wines. J. Agric. Food Chem. 2003;51:7402–7409. doi: 10.1021/jf0304167. [DOI] [PubMed] [Google Scholar]
- 18.Morata A., Gómez-Cordovés M.C., Calderón F., Suárez-Lepe J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006;106:123–129. doi: 10.1016/j.ijfoodmicro.2005.05.019. [DOI] [PubMed] [Google Scholar]
- 19.Escott C., Morata A., Loira I., Tesfaye W., Suarez-Lepe J.A. Characterization of polymeric pigments and pyranoanthocyanins formed in microfermentations of non-Saccharomyces yeasts. J. Appl. Microbiol. 2016;121:1346–1356. doi: 10.1111/jam.13255. [DOI] [PubMed] [Google Scholar]
- 20.Escott C., Del Fresno J.M., Loira I., Morata A., Tesfaye W., González C., Suárez-Lepe J.A. Formation of polymeric pigments in red wines through sequential fermentation of flavanol-enriched musts with non-Saccharomyces yeasts. Food Chem. 2018;239:975–983. doi: 10.1016/j.foodchem.2017.07.037. [DOI] [PubMed] [Google Scholar]
- 21.Morata A., Gómez-Cordovés M.C., Suberviola J., Bartolomé B., Colomo B., Suárez J.A. Adsorption of Anthocyanins by Yeast Cell Walls during the Fermentation of Red Wines. J. Agric. Food Chem. 2003;51:4084–4088. doi: 10.1021/jf021134u. [DOI] [PubMed] [Google Scholar]
- 22.Caridi A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007;120:167–172. doi: 10.1016/j.ijfoodmicro.2007.08.032. [DOI] [PubMed] [Google Scholar]
- 23.Echeverrigaray S., Scariot F.J., Menegotto M., Delamare A.P.L. Anthocyanin adsorption by Saccharomyces cerevisiae during wine fermentation is associated to the loss of yeast cell wall/membrane integrity. Int. J. Food Microbiol. 2020;314:108383. doi: 10.1016/j.ijfoodmicro.2019.108383. [DOI] [PubMed] [Google Scholar]
- 24.Loira I., Vejarano R., Morata A., Ricardo-Da-Silva J.M., Laureano O., González M.C., Suárez-Lepe J.A. Effect of Saccharomyces strains on the quality of red wines aged on lees. Food Chem. 2013;139:1044–1051. doi: 10.1016/j.foodchem.2013.01.020. [DOI] [PubMed] [Google Scholar]
- 25.Palomero F., Morata A., Benito S., Calderón F., Suárez-Lepe J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009;112:432–441. doi: 10.1016/j.foodchem.2008.05.098. [DOI] [Google Scholar]
- 26.Kulkarni P., Loira I., Morata A., Tesfaye W., González M.C., Suárez-Lepe J.A. Use of Non Saccharomyces yeast strains coupled with ultrasound treatments as a novel technique to accelerate aging over lees of red wines and its repercussion in sensorial parameters. LWT - Food Sci. Technol. 2015;64:1255–1262. doi: 10.1016/j.lwt.2015.07.046. [DOI] [Google Scholar]
- 27.García Martín J.F., Guillemet L., Feng C., Sun D.W. Cell viability and proteins release during ultrasound-assisted yeast lysis of light lees in model wine. Food Chem. 2013;141:934–939. doi: 10.1016/j.foodchem.2013.03.081. [DOI] [PubMed] [Google Scholar]
- 28.Del Fresno J.M., Loira I., Morata A., González C., Suárez-Lepe J.A., Cuerda R. Application of ultrasound to improve lees ageing processes in red wines. Food Chem. 2018;261:157–163. doi: 10.1016/j.foodchem.2018.04.041. [DOI] [PubMed] [Google Scholar]
- 29.Del Fresno J.M., Morata A., Escott C., Loira I., Cuerda R., Suárez-Lepe J.A. Sonication of yeast biomasses to improve the ageing on lees technique in red wines. Molecules. 2019;24:635. doi: 10.3390/molecules24030635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Comuzzo P., Calligaris S. Potential Applications of High Pressure Homogenization in Winemaking: A Review. Beverages. 2019;5:56. doi: 10.3390/beverages5030056. [DOI] [Google Scholar]
- 31.Kurtzman C.P., Robnett C.J. Phylogenetic relationships among yeasts of the “Saccharomyces complex” determined from multigene sequence analyses. FEMS Yeast Res. 2003;3:417–432. doi: 10.1016/S1567-1356(03)00012-6. [DOI] [PubMed] [Google Scholar]
- 32.Morata A., Loira I., Vejarano R., González C., Callejo M.J., Suárez-Lepe J.A. Emerging preservation technologies in grapes for winemaking. Trends Food Sci. Technol. 2017;67:36–43. doi: 10.1016/j.tifs.2017.06.014. [DOI] [Google Scholar]
- 33.Fleet G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003;86:11–22. doi: 10.1016/S0168-1605(03)00245-9. [DOI] [PubMed] [Google Scholar]
- 34.Swiegers J.H., Bartowsky E.J., Henschke P.A., Pretorius I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005;11:139–173. doi: 10.1111/j.1755-0238.2005.tb00285.x. [DOI] [Google Scholar]
- 35.Giorello F., Valera M.J., Martin V., Parada A., Salzman V., Camesasca L., Fariña L., Boido E., Medina K., Dellacassa E., et al. Genomic and Transcriptomic Basis of Hanseniaspora vineae’s Impact on Flavor Diversity and Wine Quality. Appl. E. 2019;85:e01959-18. doi: 10.1128/AEM.01959-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Martin V., Giorello F., Fariña L., Minteguiaga M., Salzman V., Boido E., Aguilar P.S., Gaggero C., Dellacassa E., Mas A., et al. De novo synthesis of benzenoid compounds by the yeast Hanseniaspora vineae increases the flavor diversity of wines. J. Agric. Food Chem. 2016;64:4574–4583. doi: 10.1021/acs.jafc.5b05442. [DOI] [PubMed] [Google Scholar]
- 37.Martin V., Boido E., Giorello F., Mas A., Dellacassa E., Carrau F. Effect of yeast assimilable nitrogen on the synthesis of phenolic aroma compounds by Hanseniaspora vineae strains. Yeast. 2016;33:323–328. doi: 10.1002/yea.3159. [DOI] [PubMed] [Google Scholar]
- 38.Cotton R.G.H., Gibson F. The biosynthesis of phenylalanine and tyrosine; enzymes converting chorismic acid into prephenic acid and their relationships to prephenate dehydratase and prephenate dehydrogenase. Biochim. Biophys. Acta Gen. Subj. 1965;100:76–88. doi: 10.1016/0304-4165(65)90429-0. [DOI] [PubMed] [Google Scholar]
- 39.Pugh S., McKenna R., Halloum I., Nielsen D.R. Engineering Escherichia coli for renewable benzyl alcohol production. Metab. Eng. Commun. 2015;2:39–45. doi: 10.1016/j.meteno.2015.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Marcon A.R., Schwarz L.V., Dutra S.V., Moura S., Agostini F., Delamare A.P.L., Echeverrigaray S. Contribution of a Brazilian Torulaspora delbrueckii isolate and a commercial Saccharomyces cerevisiae to the aroma profile and sensory characteristics of Moscato Branco wines. Aust. J. Grape Wine Res. 2018;24:461–468. doi: 10.1111/ajgw.12347. [DOI] [Google Scholar]
- 41.Renault P., Coulon J., Moine V., Thibon C., Bely M. Enhanced 3-Sulfanylhexan-1-ol Production in Sequential Mixed Fermentation with Torulaspora delbrueckii/Saccharomyces cerevisiae Reveals a Situation of Synergistic Interaction between Two Industrial Strains. Front. Microbiol. 2016;7:293. doi: 10.3389/fmicb.2016.00293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Culleré L., López R., Ferreira V. The Instrumental Analysis of Aroma-Active Compounds for Explaining the Flavor of Red Wines. In: Morata A., editor. Red Wine Technology. Academic Press; London, UK: 2019. pp. 283–307. [Google Scholar]
- 43.Engel K.H., Tressl R. Identification of new sulfur-containing volatiles in yellow passionfruit (Passiflora edulis f. flavicarpa) J. Agric. Food Chem. 1991;39:2249–2252. doi: 10.1021/jf00012a030. [DOI] [Google Scholar]
- 44.Loira I., Vejarano R., Bañuelos M.A., Morata A., Tesfaye W., Uthurry C., Villa A., Cintora I., Suárez-Lepe J.A. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT Food Sci. Technol. 2014;59:915–922. doi: 10.1016/j.lwt.2014.06.019. [DOI] [Google Scholar]
- 45.Velázquez R., Zamora E., Álvarez M.L., Hernández L.M., Ramírez M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015;6:1222. doi: 10.3389/fmicb.2015.01222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Anfang N., Brajkovich M., Goddard M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009;15:1–8. doi: 10.1111/j.1755-0238.2008.00031.x. [DOI] [Google Scholar]
- 47.Ciani M., Maccarelli F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1997;14:199–203. doi: 10.1023/A:1008825928354. [DOI] [Google Scholar]
- 48.Rodríguez M.E., Lopes C., Valles S., Giraudo M.R., Caballero A. Selection and preliminary characterization of β-glycosidases producer Patagonian wild yeasts. Enzyme Microb. Technol. 2007;41:812–820. doi: 10.1016/j.enzmictec.2007.07.004. [DOI] [Google Scholar]
- 49.Manzanares P., Ramón D., Querol A. Screening of non-Saccharomyces wine yeasts for the production of β-D- xylosidase activity. Int. J. Food Microbiol. 1999;46:105–112. doi: 10.1016/S0168-1605(98)00186-X. [DOI] [PubMed] [Google Scholar]
- 50.Spencer J.F.T., Gorin P.A.J. Mannose-containing polysaccharides of the apiculate yeasts Nadsonia, Hanseniaspora, Kloeckera, and Sacharomycodes, and their use as an aid in classification. J. Bacteriol. 1968;96:180–183. doi: 10.1128/jb.96.1.180-183.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang S., Petersen M.A., Liu J., Toldam-Andersen T.B. Influence of pre-fermentation treatments on wine volatile and sensory profile of the new disease tolerant cultivar solaris. Molecules. 2015;20:21609–21625. doi: 10.3390/molecules201219791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Banilas G., Sgouros G., Nisiotou A. Development of microsatellite markers for Lachancea thermotolerans typing and population structure of wine-associated isolates. Microbiol. Res. 2016;193:1–10. doi: 10.1016/j.micres.2016.08.010. [DOI] [PubMed] [Google Scholar]
- 53.Loira I., Morata A., Comuzzo P., Callejo M.J., González C., Calderón F., Suárez-Lepe J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015;76:325–333. doi: 10.1016/j.foodres.2015.06.030. [DOI] [PubMed] [Google Scholar]
- 54.Ferreira V., López R., Cacho J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000;80:1659–1667. doi: 10.1002/1097-0010(20000901)80:11<1659::AID-JSFA693>3.0.CO;2-6. [DOI] [Google Scholar]
- 55.Hranilovic A., Bely M., Masneuf-Pomarede I., Jiranek V., Albertin W. The evolution of Lachancea thermotolerans is driven by geographical determination, anthropisation and flux between different ecosystems. PLoS ONE. 2017;12:1–17. doi: 10.1371/journal.pone.0184652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hranilovic A., Gambetta J.M., Schmidtke L., Boss P.K., Grbin P.R., Masneuf-Pomarede I., Bely M., Albertin W., Jiranek V. Oenological traits of Lachancea thermotolerans show signs of domestication and allopatric differentiation. Sci. Rep. 2018;8:14812–14825. doi: 10.1038/s41598-018-33105-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Comitini F., Gobbi M., Domizio P., Romani C., Lencioni L., Mannazzu I., Ciani M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011;28:873–882. doi: 10.1016/j.fm.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 58.Gobbi M., Comitini F., Domizio P., Romani C., Lencioni L., Mannazzu I., Ciani M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013;33:271–281. doi: 10.1016/j.fm.2012.10.004. [DOI] [PubMed] [Google Scholar]
- 59.Vilela A. Lachancea thermotolerans, the Non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation. 2018;4:56. doi: 10.3390/fermentation4030056. [DOI] [Google Scholar]
- 60.Suárez-Lepe J.A., Palomero F., Benito S., Calderón F., Morata A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012;235:375–383. doi: 10.1007/s00217-012-1785-9. [DOI] [Google Scholar]
- 61.Volschenk H., van Vuuren H.J.J., Viljoen-Bloom M. Malo-ethanolic fermentation in Saccharomyces and Schizosaccharomyces. Curr. Genet. 2003;43:379–391. doi: 10.1007/s00294-003-0411-6. [DOI] [PubMed] [Google Scholar]
- 62.Loira I., Morata A., Palomero F., González C., Suárez-Lepe J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation. 2018;4:70. doi: 10.3390/fermentation4030070. [DOI] [Google Scholar]
- 63.Morata A., Calderón F., González M.C., Gómez-Cordovés M.C., Suárez J.A. Formation of the highly stable pyranoanthocyanins (vitisins A and B) in red wines by the addition of pyruvic acid and acetaldehyde. Food Chem. 2007;100:1144–1152. doi: 10.1016/j.foodchem.2005.11.024. [DOI] [Google Scholar]
- 64.Rentzsch M., Schwarz M., Winterhalter P. Pyranoanthocyanins—An overview on structures, occurrence, and pathways of formation. Trends Food Sci. Technol. 2007;18:526–534. doi: 10.1016/j.tifs.2007.04.014. [DOI] [Google Scholar]
- 65.Eglinton J., Griesser M., Henschke P., Kwiatkowski M., Parker M., Herderich M. Yeast-Mediated Formation of Pigmented Polymers in Red Wine. Red Wine Color. 2004;886:7–21. [Google Scholar]
- 66.Es-Safi N.E., Fulcrand H., Cheynier V., Moutounet M. Competition between (+)-catechin and (-)-epicatechin in acetaldehyde- induced polymerization of flavanols. J. Agric. Food Chem. 1999;47:2088–2095. doi: 10.1021/jf980628h. [DOI] [PubMed] [Google Scholar]
- 67.Es-Safi N.E., Fulcrand H., Cheynier V., Moutounet M. Studies on the acetaldehyde-induced condensation of (-)-epicatechin and malvidin 3-O-glucoside in a model solution system. J. Agric. Food Chem. 1999;47:2096–2102. doi: 10.1021/jf9806309. [DOI] [PubMed] [Google Scholar]
- 68.Dallas C., Ricardo-da-Silva J.M., Laureano O. Products Formed in Model Wine Solutions Involving Anthocyanins, Procyanidin B2, and Acetaldehyde. J. Agric. Food Chem. 1996;44:2402–2407. doi: 10.1021/jf940433j. [DOI] [Google Scholar]
- 69.Quaglieri C., Jourdes M., Waffo-Teguo P., Teissedre P.L. Updated knowledge about pyranoanthocyanins: Impact of oxygen on their contents, and contribution in the winemaking process to overall wine color. Trends Food Sci. Technol. 2017;67:139–149. doi: 10.1016/j.tifs.2017.07.005. [DOI] [Google Scholar]
- 70.Mateus N., Oliveira J., Pissarra J., González-Paramás A.M., Rivas-Gonzalo J.C., Santos-Buelga C., Silva A.M.S., De Freitas V. A new vinylpyranoanthocyanin pigment occurring in aged red wine. Food Chem. 2006;97:689–695. doi: 10.1016/j.foodchem.2005.05.051. [DOI] [Google Scholar]
- 71.Mateus N., Oliveira J., Haettich-Motta M., de Freitas V. New family of bluish pyranoanthocyanins. J. Biomed. Biotechnol. 2004;5:299–305. doi: 10.1155/S1110724304404033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.He F., Liang N.N., Mu L., Pan Q.H., Wang J., Reeves M.J., Duan C.Q. Anthocyanins and their variation in red wines II. Anthocyanin derived pigments and their color evolution. Molecules. 2012;17:1483–1519. doi: 10.3390/molecules17021483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bañuelos M.A., Loira I., Escott C., Del Fresno J.M., Morata A., Sanz P.D., Otero L., Suárez-Lepe J.A. Grape Processing by High Hydrostatic Pressure: Effect on Use of Non-Saccharomyces in Must Fermentation. Food Bioprocess Technol. 2016;9:1769–1778. doi: 10.1007/s11947-016-1760-8. [DOI] [Google Scholar]
- 74.Fava J., Hodara K., Nieto A., Guerrero S., Alzamora S.M., Castro M.A. Structure (micro, ultra, nano), color and mechanical properties of Vitis labrusca L. (grape berry) fruits treated by hydrogen peroxide, UV-C irradiation and ultrasound. Food Res. Int. 2011;44:2938–2948. doi: 10.1016/j.foodres.2011.06.053. [DOI] [Google Scholar]
- 75.Puértolas E., López N., Saldaña G., Álvarez I., Raso J. Evaluation of phenolic extraction during fermentation of red grapes treated by a continuous pulsed electric fields process at pilot-plant scale. J. Food Eng. 2010;98:120–125. doi: 10.1016/j.jfoodeng.2009.12.017. [DOI] [Google Scholar]
- 76.Cano-López M., López-Roca J.M., Pardo-Minguez F., Gómez Plaza E. Oak barrel maturation vs. micro-oxygenation: Effect on the formation of anthocyanin-derived pigments and wine colour. Food Chem. 2010;119:191–195. doi: 10.1016/j.foodchem.2009.06.018. [DOI] [Google Scholar]
- 77.Morata A., López C., Tesfaye W., González C., Escott C. Anthocyanins as natural pigments in beverages. In: Grumezescu A.M., Holban A.M., editors. Value-added ingredients and enrichements of beverages. Woodhead Publishing; London, UK: 2019. pp. 383–428. [Google Scholar]
- 78.Rodriguez-Amaya D.B. Update on natural food pigments—A mini-review on carotenoids, anthocyanins, and betalains. Food Res. Int. 2018;124:200–205. doi: 10.1016/j.foodres.2018.05.028. [DOI] [PubMed] [Google Scholar]
- 79.Braga A.R.C., Murador D.C., de Souza Mesquita L.M., de Rosso V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018;68:31–40. doi: 10.1016/j.jfca.2017.07.031. [DOI] [Google Scholar]
- 80.Morata A., Escott C., Loira I., del Fresno J.M., González C., Suárez-Lepe J.A. Influence of Saccharomyces and non-Saccharomyces Yeasts in the Formation of Pyranoanthocyanins and Polymeric Pigments during Red Wine Making. Molecules. 2019;24:4490. doi: 10.3390/molecules24244490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Morata A., Suárez-Lepe J.A. New Biotechnologies for Wine Fermentation and Ageing. In: V R.R., editor. Advances in Food Biotechnology. John Wiley & Sons Ltd.; Chichester, UK: 2016. pp. 287–302. [Google Scholar]
- 82.Siebert T.E., Solomon M.R., Pollnitz A.P., Jeffery D.W. Selective determination of volatile sulfur compounds in wine by gas chromatography with sulfur chemiluminescence detection. J. Agric. Food Chem. 2010;58:9454–9462. doi: 10.1021/jf102008r. [DOI] [PubMed] [Google Scholar]
- 83.Barbosa C., Lage P., Esteves M., Chambel L., Mendes-Faia A., Mendes-Ferreira A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from Douro Wine Region. Fermentation. 2018;4:8. doi: 10.3390/fermentation4010008. [DOI] [Google Scholar]
- 84.Varela C., Sengler F., Solomon M., Curtin C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016;209:57–64. doi: 10.1016/j.foodchem.2016.04.024. [DOI] [PubMed] [Google Scholar]
- 85.Domizio P., Romani C., Lencioni L., Comitini F., Gobbi M., Mannazzu I., Ciani M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011;147:170–180. doi: 10.1016/j.ijfoodmicro.2011.03.020. [DOI] [PubMed] [Google Scholar]
- 86.Contreras A., Hidalgo C., Schmidt S., Henschke P.A., Curtin C., Varela C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015;205:7–15. doi: 10.1016/j.ijfoodmicro.2015.03.027. [DOI] [PubMed] [Google Scholar]
- 87.Sabel A., Martens S., Petri A., König H., Claus H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2014;64:483–491. doi: 10.1007/s13213-013-0678-x. [DOI] [Google Scholar]
- 88.Fan G., Teng C., Xu D., Fu Z., Minhazul K.A.H.M., Wu Q., Liu P., Yang R., Li X. Enhanced production of ethyl acetate using co-culture of Wickerhamomyces anomalus and Saccharomyces cerevisiae. J. Biosci. Bioeng. 2019;128:564–570. doi: 10.1016/j.jbiosc.2019.05.002. [DOI] [PubMed] [Google Scholar]
- 89.Orentaite I., Poranen M.M., Oksanen H.M., Daugelavicius R., Bamford D.H. K2 killer toxin-induced physiological changes in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016;16:3. doi: 10.1093/femsyr/fow003. [DOI] [PubMed] [Google Scholar]
- 90.Qin X., Xiao H., Cheng X., Zhou H., Si L. Hanseniaspora uvarum prolongs shelf life of strawberry via volatile production. Food Microbiol. 2017;63:205–212. doi: 10.1016/j.fm.2016.11.005. [DOI] [PubMed] [Google Scholar]
- 91.Guo H., Qin X., Wu Y., Yu W., Liu J., Xi Y., Dou G., Wang L., Xiao H. Biocontrol of Gray Mold of Cherry Tomatoes with the Volatile Organic Monomer from Hanseniaspora uvarum, Trans-Cinnamaldehyde. Food Bioprocess Technol. 2019;12:1809–1820. doi: 10.1007/s11947-019-02319-6. [DOI] [Google Scholar]
- 92.Passoth V., Fredlund E., Druvefors U.Ä., Schnürer J. Biotechnology, physiology and genetics of the yeast Pichia anomala. FEMS Yeast Res. 2006;6:3–13. doi: 10.1111/j.1567-1364.2005.00004.x. [DOI] [PubMed] [Google Scholar]
- 93.Csutak O., Vassu T., Sarbu I., Stoica I., Cornea P. Antagonistic activity of three newly isolated yeast strains from the surface of fruits. Food Technol. Biotechnol. 2013;51:70–77. [Google Scholar]
- 94.Janisiewicz W.J., Tworkoski T.J., Kurtzman C.P. Biocontrol potential of Metchnikowia pulcherrima strains against blue mold of apple. Phytopathology. 2001;91:1098–1108. doi: 10.1094/PHYTO.2001.91.11.1098. [DOI] [PubMed] [Google Scholar]
- 95.Kántor A., Hutková J., Petrová J., Hleba L., Kačániová M. Antimicrobial activity of pulcherrimin pigment produced by Metschnikowia pulcherrima against various yeast species. J. Microbiol. Biotechnol. Food Sci. 2015;5:282–285. doi: 10.15414/jmbfs.2015/16.5.3.282-285. [DOI] [Google Scholar]
- 96.Oro L., Ciani M., Comitini F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014;116:1209–1217. doi: 10.1111/jam.12446. [DOI] [PubMed] [Google Scholar]
- 97.Saravanakumar D., Ciavorella A., Spadaro D., Garibaldi A., Gullino M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008;49:121–128. doi: 10.1016/j.postharvbio.2007.11.006. [DOI] [Google Scholar]
- 98.Sipiczki M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006;72:6716–6724. doi: 10.1128/AEM.01275-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Comitini F., Ingeniis De J., Pepe L., Mannazzu I., Ciani M. Pichia anomala and Kluyveromyces wickerhamii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004;238:235–240. doi: 10.1111/j.1574-6968.2004.tb09761.x. [DOI] [PubMed] [Google Scholar]
- 100.Morata A., Loira I., Vejarano R., Bañuelos M.A., Sanz P.D., Otero L., Suárez-Lepe J.A. Grape Processing by High Hydrostatic Pressure: Effect on Microbial Populations, Phenol Extraction and Wine Quality. Food Bioprocess Technol. 2015;8:277–286. doi: 10.1007/s11947-014-1405-8. [DOI] [Google Scholar]
- 101.Corrales M., Toepfl S., Butz P., Knorr D., Tauscher B. Extraction of anthocyanins from grape by-products assisted by ultrasonics, high hydrostatic pressure or pulsed electric fields: A comparison. Innov. Food Sci. Emerg. Technol. 2008;9:85–91. doi: 10.1016/j.ifset.2007.06.002. [DOI] [Google Scholar]
- 102.Corrales M., García A.F., Butz P., Tauscher B. Extraction of anthocyanins from grape skins assisted by high hydrostatic pressure. J. Food Eng. 2009;90:415–421. doi: 10.1016/j.jfoodeng.2008.07.003. [DOI] [Google Scholar]
- 103.Loira I., Morata A., Bañuelos M.A., Puig-Pujol A., Guamis B., González C., Suárez-Lepe J.A. Use of Ultra-High Pressure Homogenization processing in winemaking: Control of microbial populations in grape musts and effects in sensory quality. Innov. Food Sci. Emerg. Technol. 2018;50:50–56. doi: 10.1016/j.ifset.2018.10.005. [DOI] [Google Scholar]
- 104.Saldaña G., Álvarez I., Condón S., Raso J. Microbiological aspects related to the feasibility of PEF technology for food pasteurization. Crit. Rev. Food Sci. Nutr. 2014;54:1415–1426. doi: 10.1080/10408398.2011.638995. [DOI] [PubMed] [Google Scholar]
- 105.López N., Puértolas E., Condón S., Álvarez I., Raso J. Effects of pulsed electric fields on the extraction of phenolic compounds during the fermentation of must of Tempranillo grapes. Innov. Food Sci. Emerg. Technol. 2008;9:477–482. doi: 10.1016/j.ifset.2007.11.001. [DOI] [Google Scholar]
- 106.Puértolas E., López N., Condón S., Álvarez I., Raso J. Potential applications of PEF to improve red wine quality. Trends Food Sci. Technol. 2010;21:247–255. doi: 10.1016/j.tifs.2010.02.002. [DOI] [Google Scholar]
- 107.Puértolas E., Saldaña G., Álvarez I., Raso J. Experimental design approach for the evaluation of anthocyanin content of rosé wines obtained by pulsed electric fields. Influence of temperature and time of maceration. Food Chem. 2011;126:1482–1487. doi: 10.1016/j.foodchem.2010.11.164. [DOI] [Google Scholar]
- 108.Wu Y., Mittal G.S., Griffiths M.W. Effect of pulsed electric field on the inactivation of microorganisms in grape juices with and without antimicrobials. Biosyst. Eng. 2005;90:1–7. doi: 10.1016/j.biosystemseng.2004.07.012. [DOI] [Google Scholar]
- 109.Escott C., Vaquero C., del Fresno J.M., Bañuelos M.A., Loira I., Han S.Y., Bi Y., Morata A., Suárez-Lepe J.A. Pulsed Light Effect in Red Grape Quality and Fermentation. Food Bioprocess Technol. 2017;10:1540–1547. doi: 10.1007/s11947-017-1921-4. [DOI] [Google Scholar]
- 110.Alexandre H., Guilloux-Benatier M. Yeast autolysis in sparkling wine—A review. Aust. J. Grape Wine Res. 2006;12:119–127. doi: 10.1111/j.1755-0238.2006.tb00051.x. [DOI] [Google Scholar]
- 111.Palomero F., Morata A., Benito S., González M.C., Suárez-Lepe J.A. Conventional and enzyme-assisted autolysis during ageing over lees in red wines: Influence on the release of polysaccharides from yeast cell walls and on wine monomeric anthocyanin content. Food Chem. 2007;105:838–846. doi: 10.1016/j.foodchem.2007.01.062. [DOI] [Google Scholar]
- 112.Liu L., Loira I., Morata A., Suárez-Lepe J.A., González M.C., Rauhut D. Shortening the ageing on lees process in wines by using ultrasound and microwave treatments both combined with stirring and abrasion techniques. Eur. Food Res. Technol. 2016;242:559–569. doi: 10.1007/s00217-015-2566-z. [DOI] [Google Scholar]
- 113.Maza M.A., Delso C., Álvarez I., Raso J., Martínez J.M. Effect of pulsed electric fields on mannoproteins release from Saccharomyces cerevisiae during the aging on lees of Caladoc red wine. LWT. 2020;118:108788. doi: 10.1016/j.lwt.2019.108788. [DOI] [PubMed] [Google Scholar]
- 114.Martínez J.M., Delso C., Maza M.A., Álvarez I., Raso J. Pulsed electric fields accelerate release of mannoproteins from Saccharomyces cerevisiae during aging on the lees of Chardonnay wine. Food Res. Int. 2019;116:795–801. doi: 10.1016/j.foodres.2018.09.013. [DOI] [PubMed] [Google Scholar]
- 115.Martínez J.M., Delso C., Aguilar D., Cebrián G., Álvarez I., Raso J. Factors influencing autolysis of Saccharomyces cerevisiae cells induced by pulsed electric fields. Food Microbiol. 2018;73:67–72. doi: 10.1016/j.fm.2017.12.008. [DOI] [PubMed] [Google Scholar]
- 116.Morata A., Calderón F., González M.C., Colomo B., Suárez J.A. Crianza sobre lías, chips y microoxigenación, utilización conjunta en el envejecimiento de vinos tintos. Ventajas del uso de levaduras seleccionadas. Enólogos. 2005;34:1–8. [Google Scholar]
- 117.Gallart M., López-Tamames E., Suberbiola G., Buxaderas S. Influence of Fatty Acids on Wine Foaming. J. Agric. Food Chem. 2002;50:7042–7045. doi: 10.1021/jf0204452. [DOI] [PubMed] [Google Scholar]
- 118.Lubbers S., Charpentier C., Feuillat M., Voilley A. Influence of yeast walls on the behavior of aroma compounds in a model wine. Am. J. Enol. Vitic. 1994;45:29–33. [Google Scholar]
- 119.Del Barrio-Galán R., Pérez-Magariño S., Ortega-Heras M. Effect of the aging on lees and other alternative techniques on the low molecular weight phenols of Tempranillo red wine aged in oak barrels. Anal. Chim. Acta. 2012;732:53–63. doi: 10.1016/j.aca.2011.12.040. [DOI] [PubMed] [Google Scholar]
- 120.Saucier C., Little D., Glories Y. First evidence of acetaldehyde-flavanol condensation products in red wine. Am. J. Enol. Vitic. 1997;3:370–373. [Google Scholar]
- 121.Sartor S., Toaldo I.M., Panceri C.P., Caliari V., Luna A.S., de Gois J.S., Bordignon-Luiz M.T. Changes in organic acids, polyphenolic and elemental composition of rosé sparkling wines treated with mannoproteins during over-lees aging. Food Res. Int. 2019;124:34–42. doi: 10.1016/j.foodres.2018.11.012. [DOI] [PubMed] [Google Scholar]
- 122.González-Marco A., Ancín-Azpilicueta C. Influence of Lees Contact on Evolution of Amines in Chardonnay Wine. J. Food Sci. 2006;71:C544–C548. doi: 10.1111/j.1750-3841.2006.00182.x. [DOI] [Google Scholar]
- 123.Masino F., Montevecchi G., Arfelli G., Antonelli A. Evaluation of the Combined Effects of Enzymatic Treatment and Aging on Lees on the Aroma of Wine from Bombino bianco Grapes. J. Agric. Food Chem. 2008;56:9495–9501. doi: 10.1021/jf8015893. [DOI] [PubMed] [Google Scholar]
- 124.Bueno J.E., Peinado R.A., Medina M., Moreno J. Effect of a short contact time with lees on volatile composition of Airen and Macabeo wines. Biotechnol. Lett. 2006;28:1007–1011. doi: 10.1007/s10529-006-9038-2. [DOI] [PubMed] [Google Scholar]
- 125.Dubourdieu D., Lavigne-Cruege V. The Role of Glutathione on the Aromatic Evolution of Dry White Wine. [(accessed on 7 December 2019)]; Available online: https://www.infowine.com/en/technical_articles/the_role_of_glutathione_on_the_aromatic_evolution_of_dry_white_wine._sc_993.htm#.
- 126.Liberatore M.T., Pati S., Del Nobile M.A., Notte E. La Aroma quality improvement of Chardonnay white wine by fermentation and ageing in barrique on lees. Food Res. Int. 2010;43:996–1002. doi: 10.1016/j.foodres.2010.01.007. [DOI] [Google Scholar]
- 127.Chalier P., Angot B., Delteil D., Doco T., Gunata Z. Interactions between aroma compounds and whole mannoprotein isolated from Saccharomyces cerevisiae strains. Food Chem. 2007;100:22–30. doi: 10.1016/j.foodchem.2005.09.004. [DOI] [Google Scholar]
- 128.Giovani G., Rosi I., Bertuccioli M. Quantification and characterization of cell wall polysaccharides released by non-Saccharomyces yeast strains during alcoholic fermentation. Int. J. Food Microbiol. 2012;160:113–118. doi: 10.1016/j.ijfoodmicro.2012.10.007. [DOI] [PubMed] [Google Scholar]