

Abstract
Background and Purpose
Human malignant hyperthermia (MH) syndrome is induced by volatile anaesthetics and involves increased levels of cystathionine β‐synthase (CBS)‐derived H2S within skeletal muscle. This increase contributes to skeletal muscle hypercontractility. Kv7 channels, expressed in skeletal muscle, may be a molecular target for H2S. Here, we have investigated the role of Kv7 channels in MH.
Experimental Approach
Skeletal muscle biopsies were obtained from MH‐susceptible (MHS) and MH‐negative (MHN) patients. Immunohistochemistry, RT‐PCR, Western blot, and in vitro contracture test (IVCT) were carried out. Development and characterization of primary human skeletal muscle cells (PHSKMC) and evaluation of cell membrane potential were also performed. The persulfidation state of Kv7 channels and polysulfide levels were measured.
Key Results
Kv7 channels were similarly expressed in MHN and MHS biopsies. The IVCT revealed an anomalous contractility of MHS biopsies following exposure to the Kv7 channel opener retigabine. Incubation of negative biopsies with NaHS, prior to retigabine addition, led to an MHS‐like positive response. MHS‐derived PHSKMC challenged with retigabine showed a paradoxical depolarizing effect, compared with the canonical hyperpolarizing effect. CBS expression and activity were increased in MHS biopsies, resulting in a major polysulfide bioavailability. Persulfidation of Kv7.4 channels was significantly higher in MHS than in MHN biopsies.
Conclusions and Implications
In skeletal muscle of MHS patients, CBS‐derived H2S induced persulfidation of Kv7 channels. This post‐translational modification switches the hyperpolarizing activity into depolarizing. This mechanism can contribute to the pathological skeletal muscle hypercontractility typical of MH syndrome.
Linked Articles
This article is part of a themed section on Hydrogen Sulfide in Biology & Medicine. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v177.4/issuetoc
Abbreviations
- 3‐MST
3‐mercaptopyruvate sulfurtransferase
- BKCa
big conductance calcium‐activated K channels
- CBS
cystathionine β‐synthase
- CSE
cystathionine γ‐lyase
- H2Sn
polysulfides
- IVCT
in vitro contracture test
- KATP
ATP‐sensitive potassium channels
- MH
malignant hyperthermia
- MHN
malignant hyperthermia negative
- MHS
malignant hyperthermia susceptible
- PHSKMC
primary human skeletal muscle cells
- SKM
skeletal muscle
1. What is already known
An increase in CBS‐derived H2S within SKM occurs in human malignant hyperthermia.
This contributes to SKM hypercontractility, one of the features of MH.
2. What this study adds
Augmented levels of CBS‐derived H2S drive persulfidation of Kv7 channels.
This post‐translational modification overturns the Kv7 activity.
3. What is the clinical significance
The paradoxical Kv7 channel activity participates in the pathological hypercontractility of SKM.
Evaluation of H2S content and Kv7 persulfidation could help to diagnose malignant hyperthermia.
1. INTRODUCTION
Malignant hyperthermia (MH) is an inherited pharmacogenetic disorder of skeletal muscle (SKM) characterized by a massive and uncontrolled release of calcium within SKM triggered by volatile inhalation anaesthetics (http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2401, http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2505, http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=7175, http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=7296, and http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=7156), depolarizing muscle agents (http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=4004), or extreme physical activity in a hot environment (Hopkins et al., 2015; Brislin & Theroux, 2013; Rosenberg, Pollock, Schiemann, Bulger, & Stowell, 2015). In susceptible subjects, MH syndrome appears anytime during anaesthesia or early post‐operative period, and if it is not rapidly recognized and promptly controlled, it can be fatal. It is characterized by tachycardia and muscle rigidity, followed by metabolic acidosis, rhabdomyolysis and a rapid increase of body temperature. The sustained muscle contraction causes a rapid depletion of ATP, oxygen consumption, and heat production, with body temperatures reaching 43–44°C. All these events lead to cyanosis, hypermetabolic crisis, cardiac and/or renal dysfunction and disseminated intravascular coagulation (Hopkins, 2011; Kim, 2012). The reported incidence of MH has been estimated between 1:5,000 and 1:50,000 exposures to anaesthetics with a mortality rate around 10%. Unfortunately, the morbidity rate is much higher, around 35% (Kim, 2012; Stratman, Flynn, & Hatton, 2009). Genetic studies have identified several loci within the human genome that are linked to MH susceptibility, among them MH susceptible (MHS)1, associated with http://www.guidetopharmacology.org/GRAC/ObjectDisplayForward?objectId=747 gene http://www.guidetopharmacology.org/GRAC/ObjectDisplayForward?objectId=747) mutations on chromosome 19q13.1, and MHS3, associated with CACNA2D1 gene mutations on chromosome 1q32, which codes for the α2∂ subunit of http://www.guidetopharmacology.org/GRAC/FamilyIntroductionForward?familyId=80 (Anderson‐Pompa et al., 2008; Rosenberg & Rueffert, 2011). To date, more than 300 different mutations of RyR1 gene have been associated to MH susceptibility. However, much less than 50% of susceptible subjects display RyR1 gene mutations (Jurkatt‐Rott, McCarthy, & Lehmann‐Horn, 2000; Girard et al., 2002). For these reasons, the gold standard for MH diagnosis relies on a bioassay defined “in vitro contracture test” (IVCT; Hopkins et al., 2015). IVCT is an invasive diagnostic procedure that requires a biopsy of the vastus group of quadriceps muscle mounted in an organ bath in order to assess the SKM contractility in response to two different triggers: halothane and http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=407. According to the European group protocol for investigation of MH susceptibility, the diagnosis depends on the increased muscle contracture observed, which unveils the defect of excitation–contraction coupling mechanism. It is important to note that the majority of susceptible subjects have a healthy phenotype and it is, therefore, impossible to diagnose MH susceptibility without the IVCT procedure.
Hydrogen sulfide (http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=9532 2S) is produced in several tissues and organ systems and in animals and humans by both enzymic (the major portion) and non‐enzymic pathways. In mammals, H2S biosynthesis is controlled by a variety of enzymes, widely expressed throughout the body (Szabo & Papapetropoulos, 2017; Wallace & Wang, 2015). The best characterized H2S‐generating enzyme is http://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=279#1443, which is the main source of H2S in the CNS (Abe & Kimura, 1996; Miles & Kraus, 2004), although CBS is abundantly expressed also in peripheral tissues (Bucci et al., 2014; Fiorucci, Distrutti, Cirino, & Wallace, 2006; Hosoki, Matsuki, & Kimura, 1997; Szabo et al., 2013; Vellecco et al., 2016; Zhang, Pan, Yang, Xiao, & Yu, 2013). http://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=279#1444) is the main H2S‐generating enzyme in the vasculature and has also been found in many other parts of the body (Bucci et al., 2012; Cirino, Vellecco, & Bucci, 2017; Fiorucci et al., 2006; Vellecco et al., 2016; Zhao, Zhang, Lu, & Wang, 2001). CBS and CSE are involved in different steps of the transsulfuration pathway, although both enzymes need the amino acid http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=4782 as substrate and require pyridoxal‐5‐phosphate as a co‐factor (Kabil & Banerjee, 2014; Kimura, 2014). http://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=279#1446 is the third H2S‐generating enzyme, and it is pyridoxal‐5‐phosphate independent and uses 3‐mercaptopyruvate as substrate. 3‐MST is expressed in vasculature and in smooth muscle component of several organs (Modis, Panopoulos, Coletta, Papapetropoulos, & Szabo, 2013; Shibuya, Mikami, Kimura, Nagahara, & Kimura, 2009; Vellecco et al., 2016). The presence of hydrogen sulfide (H2S) signalling in SKM has been recently identified, and its role in SKM function is currently under investigation (see Vellecco, Armogida, & Bucci, 2018). We and others have shown that SKM expresses the three constitutive enzymes responsible for H2S biosynthesis in rats (Chen et al., 2010; Du, Li, Yang, Li, & Jin, 2013) and humans (Islam, Polhemus, Donnarumma, Brewster, & Lefer, 2015; Vellecco et al., 2016) that actively generate H2S. In this context, we have recently demonstrated that in MHS subjects, there is an increase in CBS‐derived H2S within the SKM that contributes to MH‐related hypercontractility (Vellecco et al., 2016). Among the possible molecular targets of H2S, the voltage‐gated potassium channels Kv7.1–Kv7.5 (or http://www.guidetopharmacology.org/GRAC/FamilyIntroductionForward?familyId=81) have been identified in several cell types regulating the resting membrane potential and electrical excitability (Di Cesare Mannelli et al., 2017; Martelli et al., 2013; Miceli, Soldovieri, Martire, & Taglialatela, 2008). In particular, in SKM cells, Kv7 channels promote myogenesis as well as preventing statin‐induced toxicity (Iannotti et al., 2010; Roura‐Ferrer, Solé, Martínez‐Mármol, Villalonga, & Felipe, 2008). Notably, among Kv7 channels, the http://www.guidetopharmacology.org/GRAC/ObjectDisplayForward?objectId=563 channels are the most highly expressed, in both murine and human SKM cells (Iannotti et al., 2014; Iannotti, Barrese, Formisano, Miceli, & Taglialatela, 2013).
However, the mechanism by which H2S affects the activity of Kv7 channels remains unknown. Therefore, in order to better define the molecular basis of H2S‐related MH hypercontractility, this study investigated the involvement of Kv7 channels in the MH syndrome.
2. METHODS
2.1. Human skeletal muscle biopsies
This research has been carried out in accordance with the Declaration of Helsinki (2013) of the World Medical Association. The Ethical Committee of the Institution (Cardarelli Hospital Centre for the Study of Malignant Hyperthermia) approved the study and its protocols (4/13 prot. 358). The subjects have given written informed consent. The Cardarelli Hospital Center for the Study of Malignant Hyperthermia opened in December 1989. In these 29 years (1989–2018), 2,650 patients have been identified . An accurate anamnesis has been carried out in order to identify potential muscular disorders, problems occurring during previous exposure to general anaesthetics, and sudden neonatal death. Contextually, a clinical assessment revealing muscle skeletal alterations, such as cifoscoliosis, club foot, and flying scapulas, has been performed together with clinical evaluation of squint, cryptorchidism, palpebral ptosis, inguinal, and/or dischernia. As eligible subjects, 930 patients agreed to a muscle biopsy inorder to carry out the IVCT test. Of these patients, 67% have been diagnosed as MH negative (MHN; negative diagnosis; 623 subjects) and 15% diagnosed as MH suceptible(MHS; susceptibility diagnosis; 136). During the last three years (2015–2018), 150 IVCT tests were performed and 65% were diagnosed as MHN diagnosed (98 subjects) and 15% as MHS (23 subjects). The present study has been performed on 20 MHS and on 50 MHN biopsies not required for IVCT, that is, after diagnosis has been made (male and female with age >20 years) over 18 months from December 2016 to June 2018. All the surgical procedures have been performed at the Cardarelli Hospital.
2.2. Development and full characterization of primary human skeletal muscle cell culture
Sterile muscle pieces from MHN and MHS biopsies were used. Briefly, pieces were cut in bundles, and each bundle was transferred to a sterile cell culture insert within a six‐well plate containing DMEM (4.5 g·L−1 glucose) supplemented with 20% FBS, 7‐mM HEPES, 4 μg·ml−1 insulin, 1% (vol/vol) antibiotics (penicillin/streptomycin and gentamycin), 2‐mM l‐glutamine, and 1% (vol/vol) amphotericin B according to Censier, Urwyler, Zorzato, & Treves, (1998), with some modifications. After 8–10 days, satellite cells could be clearly seen growing attached to the insert surrounding the biopsy fragment. The culture insert was then removed, and after 15 days, the medium was changed in DMEM without amphotericin B. Cells were kept in an incubator at 37°C under 5% CO2, until confluence was reached. For the evaluation of cell membrane potential, the cells were exposed to differentiation medium, DMEM supplemented with 10% horse serum for 3 days in order to promote the fusion of myogenic satellite cells into myotubes.
2.3. Evaluation of cell membrane potential in response to potassium channel modulation in primary human skeletal muscle cells
The hyperpolarizing and the depolarizing effects have been evaluated on MHN‐derived and MHS‐derived primary human SKM cells (PHSKMC) by a fluorimetric method (Martelli et al., 2013). Briefly, PHSKMC have been seeded onto a 96‐well black plate, clear bottom, at density of 20 × 103 per well, and after 24 hr, the culture medium has been replaced, and cells have been incubated for 72 hr with the differentiation medium. After 72 hr, the differentiation medium was removed, and cells incubated for 1 hr with an appropriate buffer (HEPES 20 mM, NaCl 120 mM, KCl 2 mM, CaCl2 .2H2O 2 mM, MgCl2 .6H2O 1 mM, and glucose 5 mM, pH 7.4, at room temperature) containing the bisoxonol dye bis‐(1,3‐dibutylbarbituric acid) DiBac4(3) at 2.5 μM. This membrane potential‐sensitive dye DiBac4(3) allows a non‐electrophysiological measurement of cell membrane potential; in fact, this lipophylic and negatively charged oxonol dye shuffles between cellular and extracellular fluids in a membrane potential‐dependent manner (following the Nernst equation), thus allowing to assess changes in membrane potential by means of fluorimetric recording. An increase of fluorescence, corresponding to an inward flow of the dye, reflects a membrane depolarization, whereas a decrease in fluorescence, due to an outward flow of the dye, is linked to membrane hyperpolarization. The fluorimetric recording is carried out at excitation and emission wavelengths of 488 and 520 nm respectively. After the assessment of baseline fluorescence, when required by the experimental protocol, an excess of a well‐known H2S donor, http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=6278 3 mM, or its vehicle was incubated with the PHSKM cells for 20 min before the addition of the potassium channel modulators. We used the KATP activator http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2413 (10 μM) and the KATP blocker http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2414 (10 μM); the BKCa activator http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=4272 (10 μM) and the BKCa blocker http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=4218 (100 nM); the Kv7 activator http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2601 (50 μM) and the Kv7 blocker http://www.guidetopharmacology.org/GRAC/LigandDisplayForward?ligandId=2596 (10 μM). The relative fluorescence decrease or increase, linked to hyperpolarizing or depolarizing effects of the potassium channel modulators, was calculated as follows:
where F 0 is the basal fluorescence before the addition of the tested compounds and F t is the fluorescence at time t after their administration. Results are expressed as the means (±SEM) of fluorescence values recorded for hyperpolarizing or depolarizing compounds during the 35 min of the experiment. The experiments (n = 5) were carried out using six replicates.
2.4. Immunohistochemistry
The antibody‐based procedures used in this study comply with the recommendations made by the British Journal of Pharmacology (Alexander et al., 2018). Bundles of SKM from MHN or MHS subjects were fixed in neutral buffered formalin and then embedded in paraffin. Sections (4 μm) were stained with haematoxylin and eosin and analysed under light microscopy. MHN‐derived and MHS‐derived PHSKMC were rinsed twice with ice‐cold PBS and centrifuged at 200× g for 5 min. Then the cell population was smeared, fixed using standard fixative for cytology (citofix), and processed by a modular automated tissue processor (Leica ASP300S). Once embedded in a paraffin block, cells were cut into 3 μm sections, using a microtome, floated in a 40°C water bath containing distilled water, and mounted onto STARFROST slides. The slides were analysed by using a fully automated staining instrument Benchmark Ultra Ventana coupled to the ultraView Universal DAB Detection Kit (Ventana). In details, for each slide, Ventana labels have been printed, which include a unique barcode containing all protocol information. The slides have been loaded onto the BenchMark carousel and read by the Ventana Barcode Reader. The automated BenchMark system puts the slides through a series of user‐defined de‐paraffinization and antigen retrieval steps before starting with the antibody staining. The primary antibodies, such as anti‐desmin (Ventana), anti‐myogenin (Ventana), and SKM‐specific α‐actin (Ventana), were applied automatically in a pre‐diluted dispenser. The antibody for Kv7.4 channels (KCNQ4; 1:100; Elabscience Houston, Texas) was manually titrated onto the slide. For this antibody, a specific protocol was carried out: de‐paraffinization at 72°C, antigenic unmasking at 95°C for 8 min, enzymic digestion with protease‐2 for 4 min, incubation with primary antibody for 40 min, and post‐counterstain with haematoxylin for 8 min. The images were taken with an Olympus DP21 video camera connected to an Olympus BX43 microscope using Olympus Stream image analysis software.
2.5. Western blotting
Muscle biopsies obtained from MHN and MHS patients and MHN‐PHSKMC and MHS‐PHSKMC were homogenized in RIPA buffer as previously described (Vellecco et al., 2016). Briefly, denatured proteins (40 μg) were separated on 10% SDS polyacrylamide gels and transferred to PVDF membranes. Membranes were blocked in PBS containing 0.1% vol/vol Tween‐20 and 3% wt/vol non‐fat dry milk for 30 min, followed by overnight incubation at 4°C with rabbit polyclonal Kv7.4 antibody (1:1,000). Membranes have been extensively washed in PBS containing 0.1% vol/vol Tween‐20 prior to incubation with HRP‐conjugated secondary antibody for 2 hr. Following incubation, membranes have been washed and developed using ImageQuant‐400 (GE Healthcare, USA). The target protein band intensity has been normalized relative to the intensity of the housekeeping protein GAPDH (1:5,000, Sigma‐Aldrich, Milan, Italy).
2.6. RNA purification and quantitative real‐time PCR
Total RNA was isolated from human samples, using the TRI Reagent (Sigma‐Aldrich, Milan, Italy), according to the manufacturer's instructions, followed by spectrophotometric quantization as previously described (Panza et al., 2015). Final preparation of RNA was considered DNA‐free and protein‐free if the ratio between readings at 260/280 nm was ≥1.7. Isolated mRNA was reverse transcribed by use of iScript Reverse Transcription Supermix for RTqPCR (Bio‐Rad, Milan, Italy). The quantitative real‐time PCR was carried out in CFX384 real‐time PCR detection system (Bio‐Rad, Milan, Italy) with specific primers by the use of SYBR Green master mix kit (Bio‐Rad, Milan, Italy). Samples were amplified simultaneously in triplicate in one‐assay run with a non‐template control blank for each primer pair to control for contamination or primer dimers formation, and the ct value for each experimental group was determined. The housekeeping gene (ribosomal protein S16) was used as an internal control to normalize the ct values, using the 2−ΔCt formula. Primers sequence (5′–>3′) hKv7.1 fw: GCGTCTCCATCTACAGCACG; hKv7.1 rev: GAAGTGGTAAACGAAGCATTTCC; hKv7.2 fw: ATCCTGGAAATCGTGACTATCGT; hKv7.2 rev: TTCCGGGCAAACTTGAGCC; hKv7.3 fw: AGAGGTGGATGCACAAGGAGA; hKv7.3 rev: CTTCCCACGTTTTGGGTGTC; hKv7.4 fw: GTGATGATCGTGGTTTTCGGC; hKv7.4 rev: CGATGAAGTCGATGACACAGAA; hKv7.5 fw: CAGGGGCCAGAGTATTAAGAGC; and hKv7.5 rev: CTTCTGCACTTTGGTGGGACT.
2.7. In vitro contracture test
The procedure has been performed accordingly to the “European Malignant Hyperthermia Group (EMHG) guidelines for investigation of malignant hyperthermia susceptibility” (http://www.emhg.org). Briefly, the muscular biopsy of the vastus group of the quadriceps muscle was taken under regional anaesthesia, under a femoral nerve block with 2% carbocaine. A 3‐ to 5‐cm incision was made, 4–5 muscular bundles of 15‐ to 25‐mm length, and 2‐ to 3‐mm thickness have been dissected within 15 min and quickly transferred to Krebs–Ringer solution with the following composition: NaCl 118.1 mM, KCl 3.4 mM, MgSO4 0.8 mM, KH2PO4 1.2 mM, glucose 11.1 mM, NaHCO3 25.0 mM, and CaCl2 2.5 mM; pH 7.4. The muscle samples were transported at room temperature from the operating theatre to the laboratory within 30 min. Once in the laboratory, the muscle bundles in Krebs‐Ringer were oxygenated with carbogen (95% oxygen and 5% carbon dioxide mixture), which continued for the duration of the whole procedure. The time from biopsy to the end of the tests did not exceed 5 hr. Each bundle was tied on both ends and mounted vertically in the experimental tissue bath (15‐ml volume) at 37°C so that the lower end was fixed and the upper end was connected to an isometric transducer (Isometric Transducer 1–10 g, Ugo Basile) with a resting tension of 0.2 g (2 mN). The length between sutures was measured and defined as the initial length. After leaving the tissue for 5 min at initial length, an electrical stimulus was applied (Stimulator LI 12006, Ugo Basile) with a 1‐ to 2‐ms supramaximal stimulus at a frequency of 0.2 Hz. During electrical stimulation, the muscle was stretched slowly to 150 ± 10% of initial length; this new length was considered to be the optimal length. The muscle was allowed to stabilize at the optimal length for at least 15 min and until baseline force did not vary more than 0.2 g (2 mN) within a 10‐min period. The signal was amplified, continuously recorded, and analysed by PowerLab System. After a 20‐min equilibration, two different protocols were applied. One requires the addition of progressive concentrations of caffeine (0.5, 1.0, 1.5, 2.0, 3.0, and 4.0 mM) to the isolated organ bath; the other one requires the addition of progressive concentrations of halothane (0.5%, 1%, 2%, and 3%) in the isolated organ bath. An increase in resting tension of at least 0.2 g (2 mN) has been considered significant and allows a positive diagnosis to be made.
2.8. Malignant hyperthermia susceptibility diagnosis
According to EMHG guidelines (Hopkins et al., 2015), a positive test and therefore a diagnosis depends on the increased resting tension of the tested muscle specimen: The threshold value for susceptibility is established as an increase of ≥0.2 g above the lowest resting tensions, as follows (Figure S1):
MHS hc (formerly MHS, MH‐susceptible muscle): a caffeine threshold at caffeine concentration of 2.0 mM or less and a halothane threshold concentration at 0.44 mM or less.
MHN (MH non‐susceptible muscle): A caffeine threshold at a caffeine concentration of 3 mM or more and halothane threshold concentration above 0.44 mM.
MHS h (formerly MHE, MH equivocal muscle): The suffix has been added to indicate an abnormal response to halothane only.
MHS c (formerly MHE, MH equivocal muscle): The suffix has been added to indicate an abnormal response to caffeine only.
2.9. In vitro study
The MHN and MHS biopsies not required for IVCT, that is, after diagnosis was made, were mounted in an isolated organ bath connected to an isometric transducer (Isometric Transducer 1–10 g, Ugo Basile) with a resting tension of 0.2 g. The muscle was allowed to stabilize for at least 15 min. The signal was amplified, continuously recorded, and analysed by PowerLab System (ADInstruments Lab Chart 8.0). To evaluate the possible involvement of Kv7 channels in muscle functionality, both MHN and MHS bundles have been exposed to retigabine (1 mM), and the changes in resting tension were carefully observed for 15 min. In another set of experiments, MHN tissues were incubated for 5 min with NaHS (3 mM) or vehicle (dH2O), and then retigabine (1 mM) was added to the organ bath The changes in bundle resting tension were similarly monitored for 15 min.
2.10. Protein persulfidation
Persulfidation has been detected using a modified biotin switch assay (Bibli, Hu, et al., 2018). In brief, samples were precipitated with 20% trichloroacetic acid and stored at −80°C for 24 hr. Precipitates were washed with 10% and 5% trichloroacetic acid and then centrifuged (16,000× g, 30 min, 4°C) before being resuspended in HENS buffer (250‐mM HEPES–NaOH, 1‐mM EDTA, 0.1‐mM neocuproine, 100‐μM deferoxamine, and 2.5% SDS) containing 20mM methanethiosulfonate and protease and phosphatase inhibitors. Acetone precipitation was performed, and pellets were resuspended in 300‐μl qPerS‐SID lysis buffer (6‐M urea, 100‐mM NaCl, 2% SDS, 5‐mM EDTA, and 200‐mM Tris, pH 8.2, 50‐mM iodoacetyl–PEG2–biotin, and 2.5‐mM dimedone), sonicated, and incubated for 2 hr at room temperature in the dark. Lysates (500 μg) were precipitated with acetone, and protein pellets were resuspended in 50‐μl Tris–HCl (50 mM, pH 8.5) containing guanidinium chloride (GdmCl 6 mM) and incubated at 95°C for 5 min. A negative control was generated for each sample by adding DTT (1 mM) during biotin cross‐linking. Ten per cent of the sample was immediately boiled at 95°C by addition of 3% SDS, 1% β‐mercaptoethanol, 8‐M urea, and 0.005% bromophenol blue and used for the identification of the levels of Kv7.4 channels among the samples (input). Ninety per cent of the samples was used for biotin immunoprecipitation overnight (4°C) using a high‐capacity streptavidin resin. Elution was performed by addition of 3% SDS, 1% β‐mercaptoethanol, 8‐M urea, and 0.005% bromophenol blue in PBS for 15 min at room temperature followed by 15 min at 95°C. Persulfidated proteins were then detected by SDS‐PAGE and Western blotting with a specific antibody against Kv7.4 channels.
2.11. Measurement of H2S and polysulfides, and assay of CBS enzymic activity
The tissue polysulfides were measured based on the Sulfane Sulfur Probe 4 (SSP4) selective reaction, as previously described (Bibli, Luck, et al., 2018). This method traps selectively H2S, and derivatives such as H2S‐related sulfane sulfur compounds, including persulfides (R–S–SH) and polysulfides (R–Sn–SH or R–S–Sn–S–R), and subsequently, the concentrations are calculated from the LC/MS/MS results. In brief, 1 mg of human tissue was lysed in ice‐cold RIPA buffer (50‐mM Tris–HCl, pH 7.5, 150‐mM NaCl, 25‐mM NaF, 10‐mM Na4P2O7, 1% Triton X‐100, and 0.5% sodium deoxycholate) in the absence of SDS but in the presence of protease and phosphatase inhibitors. Protein (0.5 mg) was used, and substrates of CBS (we only assayed for CBS as it is the only enzyme up‐regulated), that is, l‐homocysteine (100 μM) and l‐cysteine (100 μM) along with pyridoxal phosphate (10 μM) and SSP4 (10 μM), were added to the samples. Samples were then incubated for 60 min at 37°C. Thereafter, the tissue supernatant was collected, and floating cells were removed by centrifugation (16,000× g, 10 min, 4°C). The specific products of the reaction of H2S with SPP4 were quantified by LC/MS/MS using a 1290 Infinity UHPLC system (Agilent Technologies Deutschland GmbH, Waldbronn, Germany) and a C18 Phenomenex Kinetex (150 × 2.1 mm) column, protected by a Phenomenex C18 guard cartridge. For the quantification of H2Sn production, standard curves for fluorescein and SSP4 have been generated in RIPA buffer.
2.12. Data and statistical analysis
The data and statistical analysis comply with the recommendations of the British Journal of Pharmacology on experimental design and analysis in pharmacology (Curtis et al., 2018). The data have been expressed as mean ± SEM. The number of replicates has been at least (n) of five per group except for three experiments performed on human MHS sample (Figures 2 and 4b,c). This is due to the low incidence of the syndrome (being a rare disease) coupled to the difficulty of obtaining human tissue from MHS subjects which was not required for diagnosis. For these figures, statistical analysis of the dataset was not undertaken. The comparisons between groups were performed using Student's t test or one‐way ANOVA followed by Bonferroni or Dunnett's post hoc tests, as appropriate. Post hoc tests were performed when ANOVAs indicated that a significant difference existed between the groups. Data have been considered statistically significant when a value of P < .05 has been achieved. GraphPad Prism 5 (GraphPad Software Inc., San Diego, USA) has been employed for data analysis.
Figure 2.
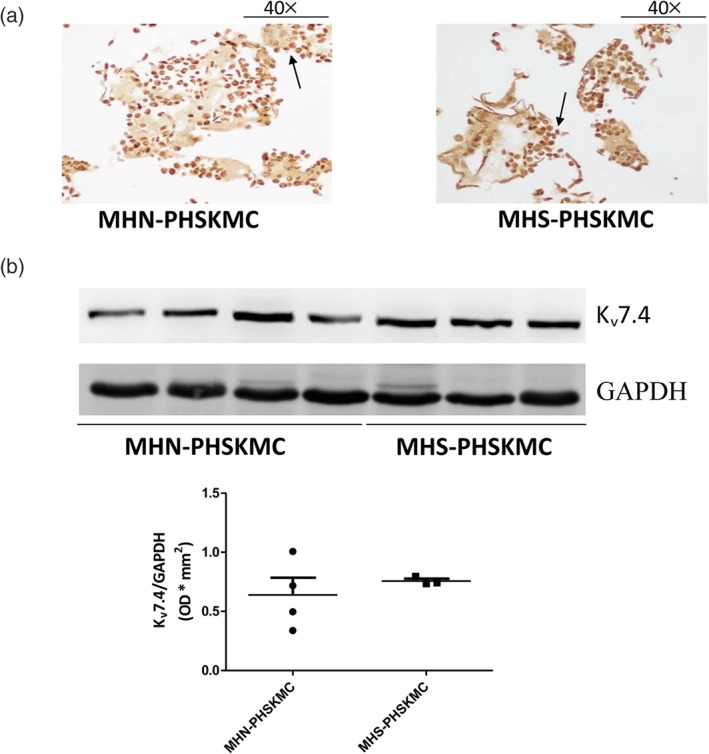
Primary human skeletal muscle cells (PHSKMC) derived from malignant hyperthermia‐negative (MHN; negative patients) and malignant hyperthermia‐susceptible (MHS; susceptible patients) biopsies do not reveal any difference in Kv7.4 channel expression. (a) Immunohistochemical staining of PHSKMC derived from both MHN and MHS shows a cytoplasmic localization of Kv7.4 channels (indicated with black arrows, 40× magnification), n = 6 for MHN‐derived PHSKMC and n = 3 for MHS‐derived PHSKMC. (b) Western blotting reveals no difference in Kv7.4 channel expression between PHSKMC derived from both MHN and MHS biopsies, n = 4 for MHN and n = 3 for MHS. Data shown are the individual values with means SEM
Figure 4.
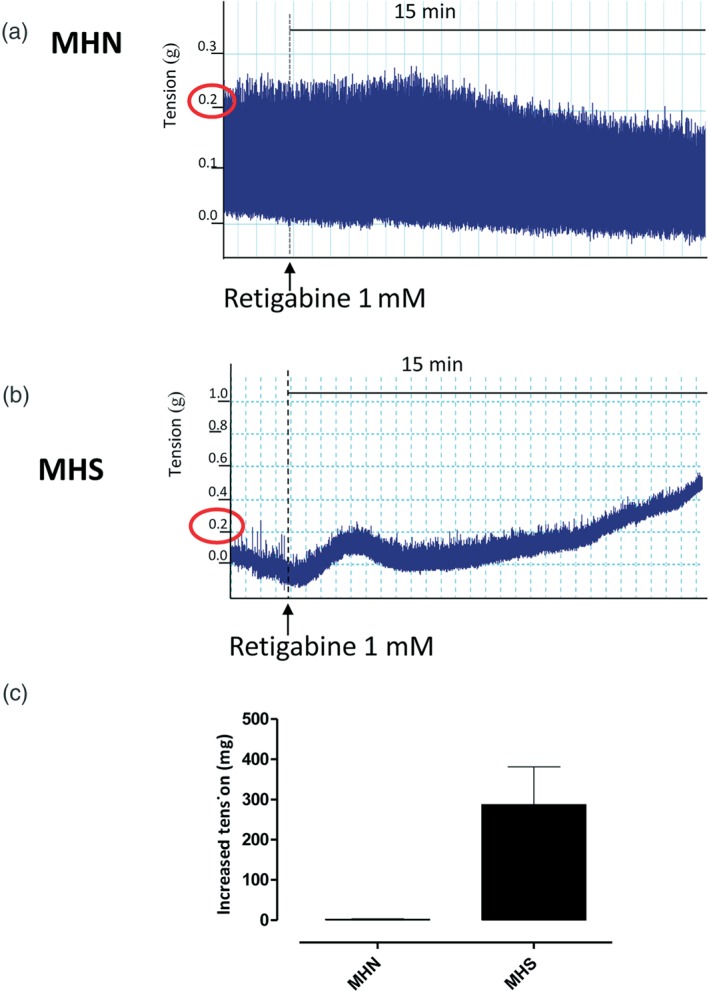
Retigabine triggers a contractile response in malignant hyperthermia‐susceptible (MHS; susceptible subjects) but not in malignant hyperthermia‐negative (MHN; negative subjects) biopsies. (a) Representative trace of skeletal muscle bundle obtained from MHN subjects incubated with retigabine (1 mM) for 15 min. (b) Representative trace of skeletal muscle bundle obtained from MHS subjects incubated with retigabine for 15 min. Retigabine increases the contracture in MHS but not in MHN bundles, n = 5 for MHN biopsies and n = 4 for MHS biopsies. (c) Effect of retigabine incubation, expressed as an increased tension, in MHN and MHS biopsies. The value enclosed in red circleindicates the established threshold for susceptibility diagnosis
2.13. Materials
All reagents for Krebs solution preparation were purchased from Carlo Erba Reagents (Milan, Italy). Caffeine, Tris–HCl, Triton‐X, sodium deoxycholate, sodium chloride, EDTA, protease inhibitor cocktail, sodium fluoride, DMSO, bisoxonol dye bis‐(1,3‐dibutylbarbituric acid) DiBac4(3), cromakalim, glibenclamide, NS1619, iberiotoxin, and all reagents for culture cells were purchased from Sigma‐Aldrich (Milan, Italy). All the drugs were dissolved in distilled H2O, except for DiBac4(3), cromakalim, glibenclamide, NS1619, and iberiotoxin, which were dissolved in DMSO. Retigabine and XE991 have been obtained from Tocris (Bristol, UK) and dissolved in DMSO. Iodoacetyl–PEG2–biotin and high‐affinity streptavidin resin were purchased from Thermo Scientific (Heidelberg, Germany). SSP4 was purchased from Dojindo (GERBU Biotechnik GmbH, Heidelberg, Germany).
2.14. Nomenclature of targets and ligands
Key protein targets and ligands in this article are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from IUPHAR/BPS Guide to PHARMACOLOGY (Harding et al., 2018), and are permanently archived in the Concise Guide to PHARMACOLOGY 2017/18 (Alexander, Fabbro et al., 2017; Alexander, Striessnig, et al., 2017).
3. RESULTS
3.1. Primary human skeletal muscle cells derived from MHN (negative patients) and MHS (susceptible patients) biopsies do not reveal any difference in morphology and in Kv7.4 channel expression
First, as shown in Figure 1a,b, no difference in morphological structure and three‐dimensional organization of the fibres was found between MHN and MHS biopsies. In PHSKMC derived from MHS and MHN biopsies, immunohistochemistry analysis reveals a strong positivity of SKM‐specific α‐actin, desmin, and myogenin as SKM markers (over 80% of cellular population) confirming the SKM phenotype (Figure 1c,d,e). Immunohistochemical techniques also revealed a clear cytoplasmic expression of Kv7.4 channels, the main subtype expressed in SKM (Iannotti et al., 2013), in PHSKMC of both MHN‐ and MHS‐derived biopsies (Figure 2a). Western blot analysis confirmed the presence of Kv7.4 channels that is equally expressed in both MHN‐PHSKMC and MHS‐PHSKMC (Figure 2b).
Figure 1.
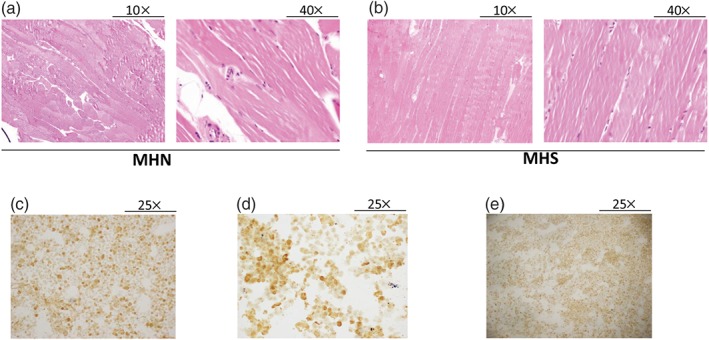
Full characterization of primary human skeletal muscle cells derived from malignant hyperthermia‐negative (MHN; negative patients) and malignant hyperthermia‐susceptible (MHS; susceptible patients) biopsies. (a, b) Haematoxylin and eosin staining of muscle fibres derived from MHN and MHS subjects shows no significant difference between the two groups. Left panel, 10× magnification; right panel, 40× magnification. The pictures are representative of n = 6 for each group. (c–e) Immunohistochemical staining of primary human skeletal muscle cells reveals a strong positivity to actin‐specific muscle, desmin, and myogenin, respectively (25× magnification). The photographs are representative of n = 6 for each group
3.2. In PHSKMC derived from MHS biopsies (susceptible subjects), activation of Kv7 channels induces a paradoxical depolarizing response
In order to obtain a general pattern of the potassium channel activity in these cells, the membrane‐hyperpolarizing or membrane‐depolarizing effects of openers and blockers of different subtypes of K+ channels were assayed. As expected, in MHN‐derived PHSKMC (cells derived from negative biopsies), all the K+ channel activators tested evoked a hyperpolarizing effect, even though with different intensity, while all the K+ channel blockers induced a depolarizing effect (Figure 3a). Interestingly, in MHS‐derived PHSKMC (cells derived from susceptible subjects), this pattern was very different (Figure 3b). In particular, the hyperpolarizing effects of both cromakalim and NS1619 were significantly amplified. Surprisingly, the effects of retigabine were completely reversed in MHS‐derived cells, as this Kv7.4 channel activator evoked a paradoxical depolarizing effect. However, the depolarizing profile of the K+ channel blockers in MHS‐derived cells was conserved. Glibenclamide induced a modest depolarization, while both iberiotoxin and XE991 evoked a very clear depolarizing effect.
Figure 3.
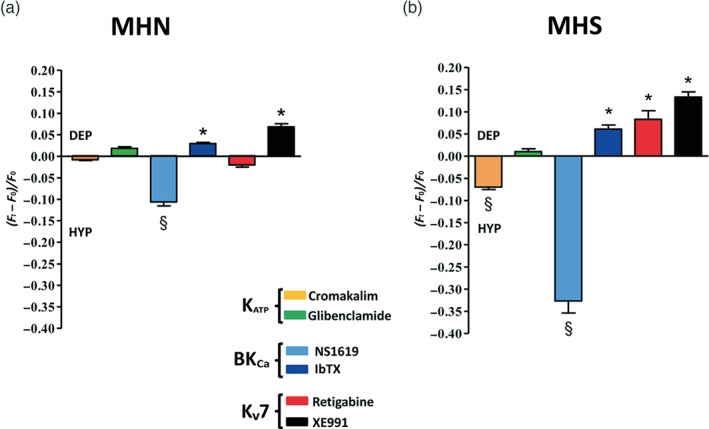
Activation of Kv7 channels induces a paradoxical depolarizing (DEP) response in primary human skeletal muscle cells (PHSKMC) derived from malignant hyperthermia‐susceptible (MHS) biopsies (susceptible subjects). (a) Evaluation of activators and blockers of KATP, BKCa, and Kv7 channels on the membrane potential of malignant hyperthermia‐negative (MHN)‐derived PHSKMC. As expected, the K channel activators evoked a hyperpolarizing (HYP) effect, while the K channel blockers induced a DEP effect. (b) Evaluation of activators and blockers of KATP, BKCa, and Kv7 potassium channels on the membrane potential of MHS‐derived PHSKMC reveals a significant reversal of the response to the Kv7 channel opener, retigabine, compared with that observed in PHSKMC from MHN patients. Values shown are the mean (± SEM; n = 5) fluorescence ratios recorded for HYP or DEP compounds during the experiment. Each experiment was carried out with six replicates. The asterisks indicate a significant depolarizing difference from the effect evoked by vehicle (*P < .05). The symbol § indicates a significant hyperpolarizing difference from the effect evoked by vehicle (§ P < .05).
3.3. Retigabine triggers a contractile response in samples from MHS (susceptible subjects) but not in those from MHN (negative subjects)
To confirm the physiopathological relevance of the unexpected depolarizing activity of Kv7 channels observed in MHS‐derived PHSKMC following retigabine stimulation, for the whole tissue, rather than an aspect of the cell culture, we exposed both MHN and MHS bundles to retigabine, using the IVCT procedure. As shown in Figure 4a, in MHN biopsies (negative patients), retigabine, at concentration of 1 mM, did not affect tissue viability or changes in baseline tension within the assay time, that is, 15 min. Conversely, exposure of MHS biopsies (susceptible patients) to retigabine (1 mM) increased baseline tension by more than 0.2 g, mimicking the IVCT response elicited by the triggers used in IVCT in susceptible patients (Figure 4b). Therefore, the phenomenon observed in PHSKMC implies a key role for Kv7 channels located on SKM.
3.4. H2S triggers an MHS‐like response to retigabine in both MHN samples and in MHN‐derived PHSKMC
To evaluate if an augmented H2S content could influence the contractility of SKM biopsies to retigabine, MHN tissues (negative patients) were incubated with NaHS (3 mM for 5 min) or vehicle prior to addition of retigabine (1 mM, Figure 5a,b). Under these experimental conditions, NaHS pretreatment leads to an increase of muscle contracture following retigabine exposure, similar in onset and shape, to a typical MHS response, showing more than 0.2‐g increase from baseline tension. When MHN samples were exposed to vehicle (H2O), used as a control for NaHS, and challenged with retigabine, no change in baseline tension was recorded (Figure 5a). To confirm that augmented levels of H2S are causative of the paradoxical depolarizing activity observed in MHS‐derived cells, MHN‐PHSKMC were incubated with NaHS and then exposed to retigabine or XE991, selective Kv7.4 channel activator or blocker respectively. As shown in Figure 5d, in these conditions, retigabine exerts a paradoxical depolarizing activity with an analogue trend inversion on MHS‐derived cells.
Figure 5.
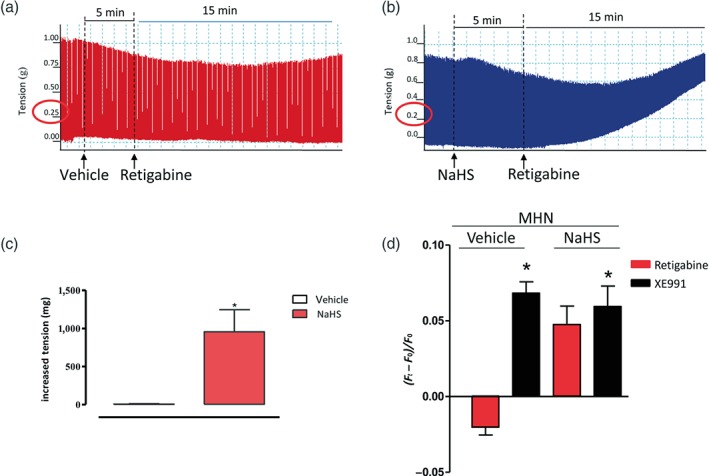
H2S triggers a malignant hyperthermia‐susceptible (MHS)‐like response to retigabine in both malignant hyperthermia‐negative (MHN) biopsies and MHN‐derived PHSKMC. (a) Representative trace of skeletal muscle bundle obtained from MHN subjects incubated for 5 min with vehicle (dH2O) followed by retigabine stimulation (1 mM). No increase in tension was observed. (b) Representative trace of skeletal muscle bundle obtained from MHN subjects incubated for 5 min with NaHS (3 mM) followed by retigabine stimulation (1 mM). The increase in tension achieved was similar to that for MHS samples, that is, >0.2 g above the resting tension. (c) Effect of NaHS (3 mM) or vehicle (dH2O) incubation in MHN biopsies followed by retigabine stimulation. Values shown are the mean (± SEM; n = 5) increases in tension. *P < .05, significantly different from vehicle. (d) Evaluation of retigabine and XE991, selective Kv7 potassium channel opener and blocker, respectively, on the membrane potential of MHN‐PHSKMC incubated with NaHS. Retigabine exerts a paradoxical depolarizing activity in cells pretreated with NaHS with a similar trend observed in MHS‐derived cells. No difference was detected between the two groups, following XE991. Values shown are the mean (± SEM; n = 5) fluorescence ratios recorded during the experiment. Each experiment was carried out with six replicates. *P < .05, significantly different from vehicle
3.5. Kv7 channels are persulfidated in MHS biopsies
To evaluate whether Kv7 channels could be differently expressed in MHN (negative patients) and MHS (susceptible patients) biopsies, real‐time PCR and western blot analysis for Kv7.4 channels protein were performed. As shown in Figure 6a,b, there were no significant differences in Kv7 mRNA (a) and protein (b) expression between MHN and MHS biopsies. Moreover, no differences were detected between MHN and MHS biopsies in the expression of all Kv7.4 channel subtypes (Figure S2). On the other hand, the persulfidation study clearly shows that elevated signals for Kv7.4 channels were detected in the MHS samples, in contrast to those in MHN samples. In detail, the Kv7.4 channel protein contains only one highly nucleophilic cysteine (Cys87) based on the prediction software Dianna 1.1. Therefore, potential persulfidation of the Kv7.4 channels was addressed based on a modified biotin switch assay, as previously described. The signal was lost in the presence of a reducing agent DTT, confirming that the detected signal is due to persulfidation (Figure 6c). These data are in accordance with the higher levels of CBS‐derived polysulfides (H2Sn) detected in MHS biopsies compared with those in the MHN biopsies (Figure 6d).
Figure 6.
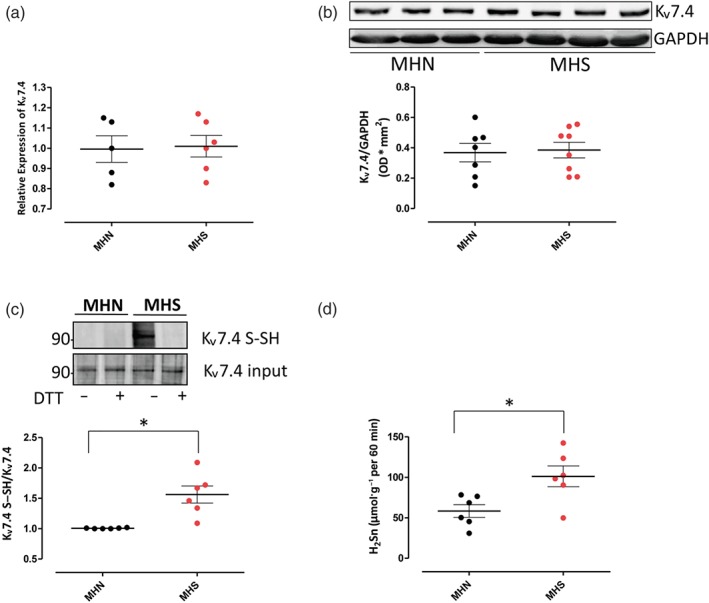
Kv7.4 channels are persulfidated in malignant hyperthermia‐susceptible (MHS) biopsies. (a) The mRNA levels of Kv7.4 are not significantly different between malignant hyperthermia‐negative (MHN) and MHS biopsies, n = 5 for MHN and n = 6 for MHS. (b) Western blotting reveals no significant difference between MHN and MHS biopsies, n = 7 for MHN and n = 8 for MHS. (c) Persulfidation analysis shows that only in MHS biopsies Kv7.4 S–SH is detected, and no band has been observed in MHN biopsies. DTT, a reducing agent, has been used to eliminate the modification prior to detection. Data shown are the individual values with means ± SEM from n = 6 experiments in each group. *P < .05, significantly different as indicated. (d) Evaluation of CBS‐derived polysulfides (H2Sn) levels in tissue homogenates reveals a significant increase in MHS samples compared with MHN samples. Data shown are the individual values with means ± SEM; n = 6 for each group. *P < .05, significantly different as indicated
4. DISCUSSION
In this study, we have investigated the involvement of the voltage‐gated Kv7 channels in susceptibility to develop MH. The whole study has been performed on human biopsies of vastus muscle obtained from MH negative and MH susceptible patients, undergoing the IVCT procedure, along with primary cell cultures derived from these biopsies. This study stems from the original finding that SKM of MHS subjects displays an augmented level of H2S and that such anomalous H2S content is due to the overexpression of CBS. The increase in H2S contributes to the SKM hypercontractility (Vellecco et al., 2016). Different subtypes of potassium channels are known to be molecular targets for H2S, such as KATP channels (Jiang, Tang, Cao, Wu, & Wang, 2010; Lowicka & Beltowski, 2007) and small‐ and intermediate‐conductance calcium‐activated K channels (SKCa and IKCa channels, respectively; Li et al., 2010; Mustafa et al., 2011; Tang et al., 2013; Telezhkin et al., 2010). Recently, Kv7 channels have been added as a new target for H2S, and their activation accounts for H2S‐induced vasorelaxation (Hedegaard et al., 2014; Hedegaard et al., 2016; Martelli et al., 2013) and H2S‐induced protection against neuropathic pain (Di Cesare Mannelli et al., 2017). It is known that Kv7 channels contribute to resting membrane potential and electrical excitation control of SKM (Miceli et al., 2008; Roura‐Ferrer et al., 2008) and that the subtype Kv7.4 channel has an essential role in the induction and maintenance of differentiated state in skeletal myotubes in murine C2C12 myoblasts (Iannotti et al., 2010). In addition, exposure of human aortic smooth muscle cells to H2S donors induces membrane hyperpolarization that is reversed by the Kv7 channel blocker XE‐991, suggesting that interactions between H2S and Kv7 channels have a physiological relevance to smooth muscle function (Martelli et al., 2013). All this evidence indicates the Kv7 channels as potential players in the MH syndrome. As it is difficult to obtain human samples from individuals susceptible to this rare syndrome, we have developed and characterized primary cell cultures (PHSKMC), derived from both MHN and MHS biopsies. No significant change, in terms of cell morphology and Kv7 expression, was detected between MHN‐derived PHSKMC and MHS‐derived PHSKMC. However, when we evaluated the membrane‐hyperpolarizing or depolarizing effects, exerted by openers or blockers of KATP, BKCa, and Kv7 channel subtypes on MHS‐derived PHSKMC, an anomalous activity of K+ channels was disclosed. Indeed, the pattern of response obtained from MHN‐derived PHSKMC shows the expected activity of the K+ channels: The openers cromakalim, NS1619, and retigabine induce hyperpolarization, while the blockers glibenclamide, NS1619, and XE991 induce depolarization. When the same experiment has been performed in MHS‐derived PHSKMC, the pattern of results was markedly different: The signals induced by the K+ channel opener retigabine (Kv7 channel opener) was switched to depolarizing activity, while cromakalim (KATP opener) and NS1619 (BKCa opener) displayed an enhanced hyperpolarizing activity. This behaviour implies, most likely, a compensatory effect to counterbalance the reversed activity of Kv7 channels. On the basis of the above observations, in SKM cells of MHS individuals, Kv7 activation would be expected to induce muscle contraction instead of muscle relaxation. The finding that, although in MHS biopsies Kv7.4 channel activity is strongly perturbed by persulfidation, no difference was detected between MHN and MHS SKM morphology implies an “alternative” pattern of differentiation of MHS SKM in order to maintain the physiological status of the tissue.
Next, we challenged MHS bundles with retigabine using the IVCT procedures. The presence of a consistent significant contracting effect induced by retigabine per se in MHS biopsies (susceptible patients) confirms a switch of the physiological response following Kv7 activation. In particular, retigabine‐induced increase in baseline tension exceeds 0.2 g; this value represents the threshold used in IVCT, following addition of triggers, to diagnose MH susceptibility. In contrast, no change in baseline tension was observed when retigabine, as well as IVCT triggers, were added to MHN bundles. It is important to highlight that the concentration ranges used are in line with IVCT guidelines that aim to achieve better reproducibility of the response for fulfilment of the quality criteria with consequent patient safety (Hopkins et al., 2015). We have previously shown that incubation of MHN biopsies with NaHS, followed by addition of the triggers used in IVCT (caffeine and halothane), leads to an increase of muscle contracture that is typical of MHS biopsies (over 0.2 g from baseline tension; Vellecco et al., 2016). Following the same protocol, MHN (negative patients) biopsies exposed to NaHS prior to challenge with retigabine display an increase in baseline tension. The shape and magnitude of the contracture elicited are similar to those obtained in MHS (susceptible patients) challenged with retigabine only. This finding implies that the high levels of H2S, present locally in the SKM of MHS susceptible patients (Vellecco et al., 2016), affect Kv7 activity. Indeed, H2S incubation can induce, in MHN samples, a switch of the response to retigabine into the MHS‐like response. To obtain further evidence on the mechanisms so far described, we used MHN‐derived PHSKMC incubated with NaHS or vehicle (H2O) and then stimulated with retigabine. The results clearly show that in cells pretreated with NaHS, retigabine evokes a depolarizing activity. These results obtained from PHSKMC clearly reproduce the effect observed, using the IVCT diagnostic procedure.
The data presented here, taken together with our previous study (Vellecco et al., 2016), suggest that in patients with MH, the augmented levels of CBS‐derived H2S, within the SKM, induce an anomalous behaviour of Kv7 channels reversing its normal, physiological hyperpolarizing activity into depolarization. This molecular event induces a hypercontractility of SKM that, most likely, contributes to muscle rigidity, one of the early clinical signs of MH syndrome. However, how H2S can affect Kv7 channels is still to be addressed. Changes in Kv7 mRNA expression and protein content are not involved, as demonstrated by the RT‐PCR and Western blot analysis performed on both MHN and MHS biopsies, confirming the data obtained by immunohistochemistry.
In the recent relevant reports, one broad post‐translational effect of H2S that affects biological function is the sulfhydration or persulfidation of target proteins (Paul & Snyder, 2012; Zhang, Du, Tang, Huang, & Jin, 2017). Several proteins that participate in a variety of physiological and/or pathological processes, such as cell proliferation/survival, mitochondrial bioenergetics, vascular reactivity and atherogenesis, and oxidative stress and inflammation are recognized as targets for sulfhydration (Bibli, Hu, et al., 2018; Jiang et al., 2010; Módis et al., 2016; Mustafa et al., 2009; Mustafa et al., 2011; Zhao, Li, Wu, Lai, & Yang, 2014). In this context, Kv7 channels have been recently proposed to be redox sensitive. In particular, highly nucleophilic cysteine residues can be oxidized by hydrogen peroxide to sulfenic (SOH) acid, to sulfinic (SO) acid, and finally to sulfonic (SO2) acid (Sahoo et al., 2014). Depending on the extent of oxidation, such modified cysteines will affect the channel activity. Therefore, we have explored the possibility that CBS‐derived H2S or H2Sn could induce post‐translational modifications affecting Kv7 channel function. The data clearly show that, only in MHS (susceptible patients) biopsies, the Kv7.4 channels were persulfidated. This evidence is compatible with the finding that MHS samples exhibit increased CBS activity in the presence of its substrates, as demonstrated by higher H2Sn bioavailability.
In conclusion, we have demonstrated that in SKM of MHS patients, CBS‐derived H2S drives persulfidation of Kv7 channels. This post‐translational event markedly changed the Kv7 channel signalling, switching its normal hyperpolarizing activity into a depolarizing effect. This molecular event translates in a pathological hypercontractility of SKM, typical of the MH syndrome. Therefore, Kv7 channels can be added to the target proteins for persulfidation–sulfhydration. This experimental finding may have a clinical utility as it may be possible to use, as the diagnostic marker, the CBS content and/or CBS‐derived polysulfides, using needle biopsies rather than a 2‐ to 3‐cm length of vastus muscle. In addition, retigabine could be used as trigger in the IVCT procedure to identify a subcluster of patients where Kv7 channels play a major role. Further studies will be necessary to confirm if retigabine could substitute for halothane and caffeine in the IVCT diagnostic test.
CONFLICT OF INTEREST
The authors declare no conflicts of interest.
AUTHOR CONTRIBUTIONS
V.V., M.V., C.C., and G.d.D. developed and characterized the primary human skeletal muscle cells (PHSKMC). A.M., V.C., and V.C. performed the cell membrane potential experiments on PHSKMC and analysed the data. A.M., M.V., E.M.B., and M.M. performed the in vitro studies and analysed the data. O.L.M. and E.P. performed the Western blot and RT‐PCR experiments. I.S.B. and I.F. performed the persulfidation experiments and analysed the data. The study has been conceived and designed by M.B. and G.C. M.B. and V.V. coordinated all the data, and M.B. and G.C. wrote and revised the manuscript.
DECLARATION OF TRANSPARENCY AND SCIENTIFIC RIGOUR
This Declaration acknowledges that this paper adheres to the principles for transparent reporting and scientific rigour of preclinical research as stated in the BJP guidelines for https://bpspubs.onlinelibrary.wiley.com/doi/full/10.1111/bph.14207, and https://bpspubs.onlinelibrary.wiley.com/doi/full/10.1111/bph.14208, and as recommended by funding agencies, publishers, and other organizations engaged with supporting research.
Supporting information
Figure S1. IVCT diagnosis for MHN and MHS subjects
(A) MHN diagnosis after caffeine exposure: after 20 minutes of muscle bundle stabilization, caffeine has been added to the organ bath with a flow of 5 ml/min at progressive concentration of 0.5, 1.0, 1.5, 2.0, 3.0, 4.0 (6 min of contact for each concentration). The absence of any contracture from the resting tension provides an MHN diagnosis for caffeine. (B) MHN diagnosis after halothane exposure: after 20 minutes of stabilization, the muscle bundle has been exposed to progressive halothane concentration of 0.5%, 1%, 2% and 3% (5 min of contact for each concentration). The absence of any contracture from the resting tension provides an MHN diagnosis for halothane. (C) MHS diagnosis after caffeine exposure: after 20 minutes of muscle bundle stabilization, caffeine has been added to the organ bath with a flow of 5 ml/min at progressive concentration of 0.5, 1.0, 1.5, 2.0, 3.0, 4.0 (6 min of contact for each concentration). The progressive increase in resting tension above 0.2 g (2mN) provides an MHS diagnosis for caffeine. (D) MHS diagnosis after halothane exposure: after 20 minutes of stabilization, the muscle bundle has been exposed to progressive halothane concentration of 0.5%, 1%, 2% and 3% (5 min of contact for each concentration). The progressive increase in resting tension ≥0.2 g (2mN) provides a MHS diagnosis for halothane.
Figure S2. RT‐PCR of the different subtype of Kv7 channels in MHN and MHS muscle biopsies mRNA levels of Kv7.1, Kv7.2, Kv7.3, Kv7.5 are not significantly different between MHN and MHS biopsies, n = 5 for MHN and n = 6 for MHS.
ACKNOWLEDGEMENTS
We thank Dr Carlo Petroccione, MD Plastic Surgeon, U.O.C Centro Grandi Ustionati—Chirurgia Plastica ricostruttiva, and Dr Romolo Villani, Head of U.O.C. Terapia Intensiva Grandi Ustionati (T.I.G.U.) Azienda Ospedaliera di Rilievo Nazionale “A. Cardarelli,” for the access to the “malignant hyperthermia‐susceptible subjects” database and for the availability of human biopsies. This work was supported by PRIN by the Italian MIUR (Ministry of Instruction, University and Research) in the programme “Project of Relevant National Interest” (PRIN, 2015), the Goethe University (Nachwuchsforschern Grant 2018 to I.S.B.), and the Deutsche Forschungsgemeinschaft (SFB 815/A6 to I.F. and Exzellenzcluster 147 “Cardio‐Pulmonary Systems”).
Vellecco V, Martelli A, Bibli IS, et al. Anomalous Kv7 channel activity in human malignant hyperthermia syndrome unmasks a key role for H2S and persulfidation in skeletal muscle. Br J Pharmacol. 2020;177:810–823. 10.1111/bph.14700
Valentina Vellecco and Alma Martelli equally contributed to the manuscript.
REFERENCES
- Abe, K. , & Kimura, H. (1996). The possible role of hydrogen sulfide as an endogenous neuromodulator. The Journal of Neuroscience, 16, 1066–1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Fabbro, D. , Kelly, E. , Marrion, N. V. , Peters, J. A. , Faccenda, E. , … CGTP Collaborators (2017). THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Enzymes. British Journal of Pharmacology, 174, S272–S359. 10.1111/bph.13877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Roberts, R. E. , Broughton, B. R. S. , Sobey, C. G. , George, C. H. , Stanford, S. C. , … Ahluwalia, A. (2018). Goals and practicalities of immunoblotting and immunohistochemistry: A guide for submission to the British Journal of Pharmacology. British Journal of Pharmacology, 175, 407–411. 10.1111/bph.14112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander, S. P. H. , Striessnig, J. , Kelly, E. , Marrion, N. V. , Peters, J. A. , Faccenda, E. , … CGTP Collaborators (2017). THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Voltage‐gated ion channels. British Journal of Pharmacology, 174, S160–S194. 10.1111/bph.13884 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson‐Pompa, K. , Foster, A. , Parker, L. , Wilks, L. , Cheek, D. J. , Mill, T. D. ,…. Mc Sherry, G. (2008). Genetics and susceptibility to malignant hyperthermia. Critical Care Nurse, 28, 32–36. [PubMed] [Google Scholar]
- Bibli, S. I. , Hu, J. , Sigala, F. , Wittig, I. , Heidler, J. , Zukunft, S. , … Fleming, I. (2018). Cystathionine γ lyase sulfhydrates the RNA binding protein HuR to preserve endothelial cell function and delay atherogenesis. Circulation, 139, 101–114. 10.1161/CIRCULATIONAHA.118.034757 [DOI] [PubMed] [Google Scholar]
- Bibli, S. I. , Luck, B. , Zukunft, S. , Wittig, J. , Chen, W. , Xian, M. , … Fleming, I. (2018). A selective and sensitive method for quantification of endogenous polysulfide production in biological samples. Redox Biology, 18, 295–304. 10.1016/j.redox.2018.07.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brislin, R. P. , & Theroux, M. C. (2013). Core myopathies and malignant hyperthermia susceptibility: A review. Paediatric Anaesthesia, 23, 834–841. [DOI] [PubMed] [Google Scholar]
- Bucci, M. , Papapetropoulos, A. , Vellecco, V. , Zhou, Z. , Zaid, A. , Giannogonas, P. , … Cirino, G. (2012). cGMP‐dependent protein kinase contributes to hydrogen sulfide‐stimulated vasorelaxation. PLoS ONE, 7, e53319 10.1371/journal.pone.0053319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bucci, M. , Vellecco, V. , Cantalupo, A. , Brancaleone, V. , Zhou, Z. , Evangelista, S. , … Cirino, G. (2014). Hydrogen sulfide accounts for the peripheral vascular effects of zofenopril independently of ACE inhibition. Cardiovascular Research, 102, 138–147. 10.1093/cvr/cvu026 [DOI] [PubMed] [Google Scholar]
- Censier, K. , Urwyler, A. , Zorzato, F. , & Treves, S. (1998). Intracellular calcium homeostasis in human primary muscle cells from malignant hyperthermia‐susceptible and normal individuals. Effect of overexpression of recombinant wild‐type and Arg163Cys mutated ryanodine receptors. The Journal of Clinical Investigation, 101, 1233–12342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, N. C. , Yang, F. , Capecci, L. M. , Gu, Z. Y. , Schafer, A. I. , Durante, W. , … Wang, H. (2010). Regulation of homocysteine metabolism and methylation in human and mouse tissues. The FASEB Journal, 24, 2804–2817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cirino, G. , Vellecco, V. , & Bucci, M. (2017). Nitric oxide–hydrogen sulfide: The gas paradigm of the vascular system. British Journal of Pharmacology, 174, 4021–4031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis, M. J. , Ashton, J. C. , Moon, L. D. F. , Ahluwalia, A. (2018). Clarification of the basis for the selection of requirements for publication in the British Journal of Pharmacology. Br J Pharmacol, 175(18), 3633–3635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Cesare Mannelli, L. , Lucarini, E. , Micheli, L. , Mosca, I. , Ambrosino, P. , Soldovieri, M. V. , … Ghelardini, C. (2017). Effects of natural and synthetic isothiocyanate‐based H2S‐releasers against chemotherapy‐induced neuropathic pain: Role of Kv7 potassium channels. Neuropharmacology, 121, 49–59. [DOI] [PubMed] [Google Scholar]
- Du, J. , Li, W. , Yang, C. , Li, Q. , & Jin, H. (2013). Hydrogen sulfide is endogenously generated in rat skeletal muscle and exerts a protective effect against oxidative stress. Chinese Medical Journal, 126, 930–936. [PubMed] [Google Scholar]
- Fiorucci, S. , Distrutti, E. , Cirino, G. , & Wallace, J. L. (2006). The emerging roles of hydrogen sulfide in the gastrointestinal tract and liver. Gastroenterology, 131, 259–271. [DOI] [PubMed] [Google Scholar]
- Girard, T. , Treves, S. , Censier, K. , Mueller, C. R. , Zorzat, F. , & Urwyler, A. (2002). Phenotyping malignant hyperthermia susceptibility by measuring halothane‐induced changes in myoplasmic calcium concentration in cultured human skeletal muscle cells. British Journal of Anaesthesia, 89, 571–579. [DOI] [PubMed] [Google Scholar]
- Harding, S. D. , Sharman, J. L. , Faccenda, E. , Southan, C. , Pawson, A. J. , Ireland, S. , … NC‐IUPHAR (2018). The IUPHAR/BPS Guide to PHARMACOLOGY in 2018: Updates and expansion to encompass the new guide to IMMUNOPHARMACOLOGY. Nucleic Acids Research, 46, D1091–D1106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedegaard, E. R. , Gouliaev, A. , Winther, A. K. , Arcanjo, D. D. , Aalling, M. , Renaltan, N. S. , … Simonsen, U. (2016). Involvement of potassium channels and calcium‐independent mechanisms in hydrogen sulfide‐induced relaxation of rat mesenteric small arteries. The Journal of Pharmacology and Experimental Therapeutics, 356, 53–63. 10.1124/jpet.115.227017 [DOI] [PubMed] [Google Scholar]
- Hedegaard, E. R. , Nielsen, B. D. , Kun, A. , Hughes, A. D. , Krøigaard, C. , Mogensen, S. , … Simonsen, U. (2014). KV7 channels are involved in hypoxia‐induced vasodilatation of porcine coronary arteries. British Journal of Pharmacology, 171, 69–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hopkins, P. M. (2011). Malignant hyperthermia: Pharmacology of triggering. British Journal of Anaesthesia, 107, 48–56. [DOI] [PubMed] [Google Scholar]
- Hopkins, P. M. , Rüffert, H. , Snoeck, M. M. , Girard, T. , Glahn, K. P. E. , Ellis, F. R. , … European Malignant Hyperthermia Group (2015). European Malignant Hyperthermia Group guidelines for investigation of malignant hyperthermia susceptibility. British Journal of Anaesthesia, 115, 531–539. [DOI] [PubMed] [Google Scholar]
- Hosoki, R. , Matsuki, N. , & Kimura, H. (1997). The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochemical and Biophysical Research Communications, 237, 527–531. [DOI] [PubMed] [Google Scholar]
- Iannotti, F. A. , Barrese, V. , Formisano, L. , Miceli, F. , & Taglialatela, M. (2013). Specification of skeletal muscle differentiation by repressor element‐1 silencing transcription factor (REST)‐regulated Kv7.4 potassium channels. Molecular Biology of the Cell, 24, 274–284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iannotti, F. A. , Panza, E. , Barrese, V. , Viggiano, D. , Soldovieri, M. V. , & Taglialatela, M. (2010). Expression, localization, and pharmacological role of Kv7 potassium channels in skeletal muscle proliferation, differentiation, and survival after myotoxic insults. The Journal of Pharmacology and Experimental Therapeutics, 332(3), 811–820. [DOI] [PubMed] [Google Scholar]
- Iannotti, F. A. , Silvestri, C. , Mazzarella, E. , Martella, A. , Calvigioni, D. , Piscitelli, F. , … Di Marzo, V. (2014. Jun 17). The endocannabinoid 2‐AG controls skeletal muscle cell differentiation via CB1 receptor‐dependent inhibition of Kv7 channels. Proceedings of the National Academy of Sciences of the United States of America, 111(24), E2472–E2481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Islam, K. N. , Polhemus, D. J. , Donnarumma, E. , Brewster, L. P. , & Lefer, D. J. (2015). Hydrogen sulfide levels and nuclear factor‐erythroid 2‐related factor 2 (NRF2) activity are attenuated in the setting of critical limb ischemia (CLI). Journal of the American Heart Association, 4, e001986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang, B. , Tang, G. , Cao, K. , Wu, L. , & Wang, R. (2010). Molecular mechanism for H2S‐induced activation of KATP channels. Antioxidants & Redox Signaling, 12, 1167–1178. [DOI] [PubMed] [Google Scholar]
- Jurkatt‐Rott, K. , McCarthy, T. , & Lehmann‐Horn, F. (2000). Genetics and pathogenesis of malignant hyperthermia. Muscle & Nerve, 23, 4–17. [DOI] [PubMed] [Google Scholar]
- Kabil, O. , & Banerjee, R. (2014). Enzimology of H2S biogenesis, decay and signaling. Antioxidants & Redox Signaling, 20, 770–782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, D. C. (2012). Malignant hyperthermia. Korean Journal of Anesthesiology, 63, 391–401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura, H. (2014). Production and physiological effect of hydrogen sulfide. Antioxidants & Redox Signaling, 20, 783–793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Q. , Sun, B. , Wang, X. , Jin, Z. , Zhou, Y. , & Dong, L. (2010). A crucial role for hydrogen sulfide in oxygen sensing via modulating large conductance calcium‐activated potassium channels. Antioxidants & Redox Signaling, 12, 1179–1189. [DOI] [PubMed] [Google Scholar]
- Lowicka, E. , & Beltowski, J. (2007). Hydrogen sulfide (H2S)—The third gas of interest for pharmacologists. Pharmacological Reports, 59, 4–24. [PubMed] [Google Scholar]
- Martelli, A. , Testai, L. , Breschi, M. C. , Lawson, K. , McKay, N. G. , Miceli, F. , … Calderone, V. (2013). Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacological Research, 70, 27–34. 10.1016/j.phrs.2012.12.005 [DOI] [PubMed] [Google Scholar]
- Miceli, F. , Soldovieri, M. V. , Martire, M. , & Taglialatela, M. (2008). Molecular pharmacology and therapeutic potential of neuronal Kv7‐modulating drugs. Current Opinion in Pharmacology, 8, 65–74. [DOI] [PubMed] [Google Scholar]
- Miles, E. W. , & Kraus, J. P. (2004). Cystathionine β‐synthase: Structure, function, regulation, and location of homocystinuria‐causing mutations. The Journal of Biological Chemistry, 279, 29871–29874. [DOI] [PubMed] [Google Scholar]
- Módis, K. , Ju, Y. , Ahmad, A. , Untereiner, A. A. , Altaany, Z. , Wu, L. , … Wang, R. (2016). S‐Sulfhydration of ATP synthase by hydrogen sulfide stimulates mitochondrial bioenergetics. Pharmacological Research, 113, 116–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Modis, K. , Panopoulos, P. , Coletta, C. , Papapetropoulos, A. , & Szabo, C. (2013). Hydrogen sulfide mediated stimulation of mitochondrial electron transport involves inhibition of the mitochondrial phosphodiesterase 2A, elevation of cAMP and activation of protein kinase A. Biochemical Pharmacology, 86, 1311–1319. [DOI] [PubMed] [Google Scholar]
- Mustafa, A. K. , Gadalla, M. M. , Sen, N. , Kim, S. , Mu, W. , Gazi, S. K. , … Snyder, S. H. (2009). H2S signals through protein S‐sulfhydration. Science Signaling, 2(96), ra72 10.1126/scisignal.2000464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustafa, A. K. , Sikka, G. , Gazi, S. K. , Steppan, J. , Jung, S. M. , Bhunia, A. K. , … Snyder, S. H. (2011). Hydrogen sulfide as endothelium‐derived hyperpolarizing factor sulfhydrates potassium channels. Circulation Research, 109, 1259–1268. 10.1161/CIRCRESAHA.111.240242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panza, E. , De Cicco, P. , Armogida, C. , Scognamiglio, G. , Gigantino, V. , Botti, G. , … Ianaro, A. (2015). Role of the cystathionine γ lyase/hydrogen sulfide pathway in human melanoma progression. Pigment Cell & Melanoma, Research, 28, 61–72. 10.1111/pcmr.12312 [DOI] [PubMed] [Google Scholar]
- Paul, B. D. , & Snyder, S. H. (2012). H2S signaling through protein sulfhydration and beyond. Nature Reviews. Molecular Cell Biology, 13, 499–507. [DOI] [PubMed] [Google Scholar]
- Rosenberg, H. , Pollock, N. , Schiemann, A. , Bulger, T. , & Stowell, K. (2015). Malignant hyperthermia: A review. Orphanet Journal of Rare Diseases, 2, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenberg, H. , & Rueffert, H. (2011). Clinical utility gene card for malignant hyperthermia. European Journal of Human Genetics, 19 10.1038/ejhg.2010.248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roura‐Ferrer, M. , Solé, L. , Martínez‐Mármol, R. , Villalonga, N. , & Felipe, A. (2008). Skeletal muscle Kv7 (KCNQ) channels in myoblast differentiation and proliferation. Biochemical and Biophysical Research Communications, 369, 1094–1097. [DOI] [PubMed] [Google Scholar]
- Sahoo, N. , Hoshi, T. , & Heinemann, S. H. (2014). Oxidative modulation of voltage‐gated potassium channels. Antioxidants & Redox Signaling, 21(6), 933–952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibuya, N. , Mikami, Y. , Kimura, Y. , Nagahara, N. , & Kimura, H. (2009). Vascular endothelium expresses 3‐mercaptopyruvate sulfurtransferase and produces hydrogen sulfide. Journal of Biochemistry, 146, 623–626. [DOI] [PubMed] [Google Scholar]
- Stratman, R. C. , Flynn, D. , & Hatton, K. W. (2009). Malignant hyperthermia: A pharmacogenetics disorder. Orthopedics, 32, 835–838. [DOI] [PubMed] [Google Scholar]
- Szabo, C. , Coletta, C. , Chao, C. , Modis, K. , Szczesny, B. , Papapetropoulos, A. , & Hellmich, M. R. (2013). Tumor derived hydrogen sulfide, produced by cystathionine‐β‐synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer. Proceedings of the National Academy of Sciences of the United States of America, 110, 12474–12479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabo, C. , & Papapetropoulos, A. (2017). International Union of Basic and Clinical Pharmacology. CII: Pharmacological modulation of H2S levels: H2S donors and H2S biosynthesis inhibitors. Pharmacological Reviews, 69, 497–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang, G. , Yang, G. , Jiang, B. , Ju, Y. , Wu, L. , & Wang, R. (2013). H₂S is an endothelium‐derived hyperpolarizing factor. Antioxidants & Redox Signaling, 19, 1634–1646. [DOI] [PubMed] [Google Scholar]
- Telezhkin, V. , Brazier, S. P. , Cayzac, S. H. , Wilkinson, W. J. , Riccardi, D. , & Kemp, P. J. (2010). Mechanism of inhibition by hydrogen sulfide of native and recombinant BKCa channels. Respiratory Physiology & Neurobiology, 172, 169–178. [DOI] [PubMed] [Google Scholar]
- Vellecco, V. , Armogida, C. , & Bucci, M. (2018). Hydrogen sulfide pathway and skeletal muscle: An introductory review. British Journal of Pharmacology, 175, 3090–3099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vellecco, V. , Mancini, A. , Ianaro, A. , Calderone, V. , Attanasio, C. , Cantalupo, A. , … Bucci, M. (2016). Cystathionine β‐synthase‐derived hydrogen sulfide is involved in human malignant hyperthermia. Clinical Science (Lond), 130, 35–44. [DOI] [PubMed] [Google Scholar]
- Wallace, J. L. , & Wang, R. (2015). Hydrogen sulfide‐based therapeutics: Exploiting a unique but ubiquitous gasotransmitter. Nature Reviews Drug Discovery, 14, 329–345. [DOI] [PubMed] [Google Scholar]
- Zhang, D. , Du, J. , Tang, C. , Huang, Y. , & Jin, H. (2017). H2S‐induced sulfhydration: Biological function and detection methodology. Frontiers in Pharmacology, 8, 608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, L. , Pan, C. , Yang, B. , Xiao, Y. , & Yu, B. (2013). Enhanced expression of cystathionine β‐synthase and cystathionine γ‐lyase during acute cholecystitis‐induced gallbladder inflammation. PLoS ONE, 8, e82711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, K. , Li, S. , Wu, L. , Lai, C. , & Yang, G. (2014). Hydrogen sulfide represses androgen receptor transactivation by targeting at the second zinc finger module. The Journal of Biological Chemistry, 289, 20824–20835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, W. , Zhang, J. , Lu, Y. , & Wang, R. (2001). The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. The EMBO Journal, 20, 6008–6016. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. IVCT diagnosis for MHN and MHS subjects
(A) MHN diagnosis after caffeine exposure: after 20 minutes of muscle bundle stabilization, caffeine has been added to the organ bath with a flow of 5 ml/min at progressive concentration of 0.5, 1.0, 1.5, 2.0, 3.0, 4.0 (6 min of contact for each concentration). The absence of any contracture from the resting tension provides an MHN diagnosis for caffeine. (B) MHN diagnosis after halothane exposure: after 20 minutes of stabilization, the muscle bundle has been exposed to progressive halothane concentration of 0.5%, 1%, 2% and 3% (5 min of contact for each concentration). The absence of any contracture from the resting tension provides an MHN diagnosis for halothane. (C) MHS diagnosis after caffeine exposure: after 20 minutes of muscle bundle stabilization, caffeine has been added to the organ bath with a flow of 5 ml/min at progressive concentration of 0.5, 1.0, 1.5, 2.0, 3.0, 4.0 (6 min of contact for each concentration). The progressive increase in resting tension above 0.2 g (2mN) provides an MHS diagnosis for caffeine. (D) MHS diagnosis after halothane exposure: after 20 minutes of stabilization, the muscle bundle has been exposed to progressive halothane concentration of 0.5%, 1%, 2% and 3% (5 min of contact for each concentration). The progressive increase in resting tension ≥0.2 g (2mN) provides a MHS diagnosis for halothane.
Figure S2. RT‐PCR of the different subtype of Kv7 channels in MHN and MHS muscle biopsies mRNA levels of Kv7.1, Kv7.2, Kv7.3, Kv7.5 are not significantly different between MHN and MHS biopsies, n = 5 for MHN and n = 6 for MHS.