

Abstract
Intracellular parasites of the genus Eimeria are described as tissue/host‐specific. Phylogenetic classification of rodent Eimeria suggested that some species have a broader host range than previously assumed. We explore whether Eimeria spp. infecting house mice are misclassified by the most widely used molecular markers due to a lack of resolution, or whether, instead, these parasite species are indeed infecting multiple host species.
With the commonly used markers (18S/COI), we recovered monophyletic clades of E. falciformis and E. vermiformis from Mus that included E. apionodes identified in other rodent host species (Apodemus spp., Myodes glareolus, and Microtus arvalis). A lack of internal resolution in these clades could suggest the existence of a species complex with a wide host range infecting murid and cricetid rodents. We question, however, the power of COI and 18S markers to provide adequate resolution for assessing host specificity. In addition to the rarely used marker ORF470 from the apicoplast genome, we present multilocus genotyping as an alternative approach. Phylogenetic analysis of 35 nuclear markers differentiated E. falciformis from house mice from isolates from Apodemus hosts. Isolates of E. vermiformis from Mus are still found in clusters interspersed with non‐Mus isolates, even with this high‐resolution data.
In conclusion, we show that species‐level resolution should not be assumed for COI and 18S markers in coccidia. Host–parasite cospeciation at shallow phylogenetic nodes, as well as contemporary coccidian host ranges more generally, is still open questions that need to be addressed using novel genetic markers with higher resolution.
Keywords: 18S, COI, Eimeria, multilocus sequence typing, phylogenetics, rodents
Eimeria is described as tissue and host‐specific. For species infecting rodents, however, a broader host range has been reported based on phylogenetic inference with 18S and COI markers. We developed a multilocus genotyping approach to increase the resolution for particular parasite groups and show that the approach is able to differentiate a genetic cluster with specialized host usage in a case unresolved by 18S and COI. In another case, the approach recovers a cluster of isolates interleaved with regards to their host usage, and question whether failure to resolve parasite clusters with specialist host ranges should be interpreted as evidence for host generalism.

1. INTRODUCTION
Coccidians of the genus Eimeria have been described as monoxenous, intracellular parasites (Becker, 1934; Long & Joyner, 1984; Marquardt, 1981). Two different characteristics extensively used to delineate Eimeria species are their assumed high degree of host and tissue specificity. It is not clear, however, whether host specificity is the same for Eimeria species infecting hosts in different clades. Eimeria species of rodents show a degree of host specificity (Ball & Lewis, 1984; De Vos, 1970; Duszynski, 2011; Wilber, Duszynski, Upton, Seville, & Corliss, 1998), but individual isolates can experimentally infect different species and even genera of rodents (Levine & Ivens, 1988; Upton, McAllister, Brillhart, Duszynski, & Wash, 1992).
Descriptions of Eimeria species are based on the size and shape of sporulated oocysts and their internal structures. The life cycles of a few species have additionally been studied and data on their dynamics (e.g., the patent period, the time before oocysts are shed in feces) are available (Duszynski, Eastham, & Yates, 1982; Hnida, Wilson, & Duszynski, 1998; Lainson & Shaw, 1990; Levine & Ivens, 1965; Mesfin & Bellamy, 1978; Todd & Hammond, 1968; Todd & Lepp, 1971; Turner, Penzhorn, & Getz, 2016; Wash, Duszynski, & Yates, 1985). For field studies, the morphology of sporulated oocysts alone is considered insufficient to infer species identity because of inadequate reference descriptions (MacPherson & Gajadhar, 1993; Tenter et al., 2002).
Genetic markers from nuclear (nu) and mitochondrial (mt) genomes, and less frequently of the apicoplast (ap) genome, have been used to complement morphological taxonomy with phylogenetic analyses (Hnida & Duszynski, 1999a, 1999b; Kvičerová, Mikeš, & Hypša, 2011; Ogedengbe, Ogedengbe, Hafeez, & Barta, 2015; Zhao & Duszynski, 2001a). Based on the assumption of host specificity of individual Eimeria species, phylogenetic analysis of nuclear small subunit ribosomal (18S) rDNA and cytochrome c oxidase I (COI) fragments supports predominant host–parasite cospeciation (Ogedengbe, El‐Sherry, Ogedengbe, Chapman, & Barta, 2018). Species infecting rodents, however, are found in two separate clades, generating marked discrepancy between parasite and host phylogeny at deeper nodes (Kvičerová & Hypša, 2013). At shallow nodes of the phylogeny for rodent coccidians, cases of host generalism have been suggested (Mácová et al., 2018). Host specificity of Eimeria species infecting rodents is not as undisputed as in other hosts such as poultry (Barta et al., 1997) or rabbits (Kvičerová, Pakandl, & Hypša, 2008). Kvičerová and Hypša (2013) suggested that adaptation rather than cospeciation is shaping rodent Eimeria cophylogenies. Mácová et al. (2018) added that host ecology and distribution may favor host‐switches among closely related rodent species. A high specificity of E. apionodes naturally infecting Apodemus flavicollis was originally suggested based on failed attempts to experimentally infect other rodents: Myodes (Clethrionomys) glareolus, Microtus arvalis, or Mus musculus (Pellérdy, 1954). It is, however, unclear if this result holds for the multiple isolates that have been assigned as E. apionodes.
We studied wild populations of Mus musculus and other rodents to assess the diversity of Eimeria isolates at shallow depth of phylogenetic relationships. We test host specificity based on phylogenetic analysis using established markers (nu 18S, mt COI, and ap ORF470). We question in how far these markers are polymorphic enough to resolve between genetic clusters with different host usage (and whether a negative result for genetic differentiation therefore suggests cases of generalism). We develop and apply multilocus sequence typing to disentangle relationships unresolved by 18S and COI markers.
2. MATERIAL AND METHODS
2.1. Origin of samples
DNA was extracted from the colon content or gastrointestinal tissue of house mice (Mus musculus) infected with Eimeria. These samples came from rodents captured in farms and private properties in the German federal states of Mecklenburg‐Vorpommern, Bavaria, and Brandenburg (capture permit No. 2347/35/2014) and in Bohemia (Czech Republic) between 2014 and 2017 (Jarquín‐Díaz et al., 2019). Additionally, DNA from gastrointestinal tract and tissue or feces of Apodemus spp. from different regions in Europe (including areas overlapping with those sampled for house mice) were also included (Mácová et al., 2018) (Figure 1) (Data S1).
Figure 1.
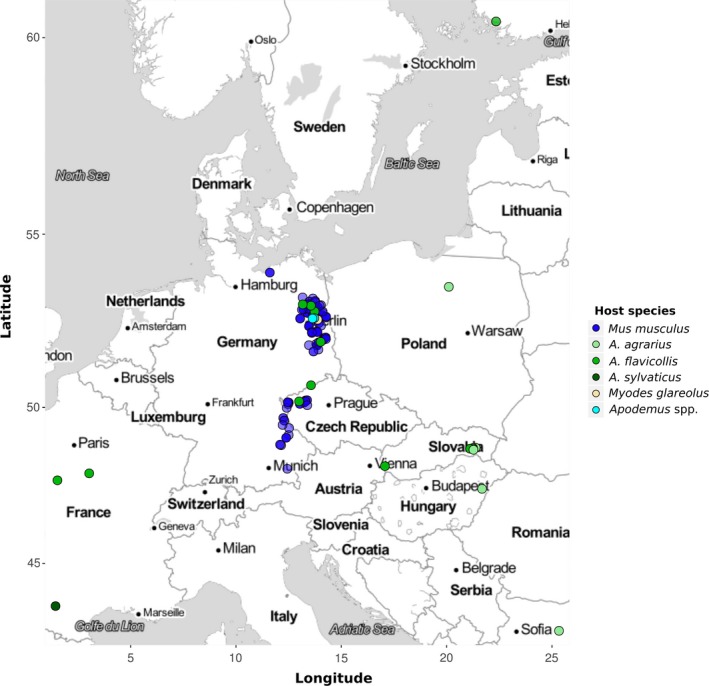
Location of rodent samples. Mus musculus samples were collected from the German federal states of Mecklenburg‐Vorpommern, Bavaria, and Brandenburg and in Bohemia (Czech Republic). Non‐Mus samples were collected from different countries within Europe. Color in the points indicates the host species
2.2. Host identification
Rodents were first identified visually based on their morphology. Identification of Mus musculus at the subspecies‐level was confirmed based on a set of previously described markers (Ďureje, Macholán, Baird, & Piálek, 2012). In order to confirm the species of non‐Mus rodents, a fragment of cytochrome b (~900 bp) was amplified from host DNA. PCRs were performed according to the protocols described by Reutter, Petit, Brünner, and Vogel (2003) for Apodemus spp., Abramson, Rodchenkova, and Kostygov (2009) (primers UCBO_F/LM_R), and Jaarola and Searle (2002) (primers L14641M/H15408M) for rodents belonging to the subfamily Arvicolinae (Myodes spp. and Microtus spp.).
2.3. PCR amplification (nu 18S rDNA, mt COI, and ap ORF470)
For phylogenetic analysis, nuclear small subunit ribosomal DNA (18S; ~1,500 bp), a fragment of the mitochondrial cytochrome c oxidase subunit I (COI; ~800 bp) gene and apicoplast ORF470 (~800 bp) were amplified using primers previously reported by Kvičerová et al. (2008), Ogedengbe, Hanner, and Barta (2011) and Zhao and Duszynski (2001b), respectively.
When COI failed to amplify with this protocol, an alternative pair of primers was used: Eim_COI_M_F (ATGTCACTNTCTCCAACCTCAGT) and Eim_COI_M_R (GAGCAACATCAANAGCAGTGT). These primers amplify a ~700 bp fragment of COI and were designed based on the mitochondrial genome of E. falciformis (CM008276.1) (Heitlinger, Spork, Lucius, & Dieterich, 2014; Jarquín‐Díaz et al., 2019).
PCRs were carried out in a Labcycler (SensoQuest GmbH, Göttingen, Germany) using 0.025 U/µL of DreamTaqTMDNA Polymerase (Thermo Scientific, Waltham, USA), 1X DreamTaq Buffer, 0.5 mM dNTP Mix, 0.25 µM from each primer, and 1–20 ng/µl of DNA template in 25 µl reaction. A concentration of 0.25 mM dNTP mix and a supplementation with 0.5 mMMgCl2 was used for the ap ORF470 amplification. The thermocycling protocol consisted of 95°C initial denaturation (4 min) followed by 35 cycles of 92°C denaturation (45 s), annealing at 52°C (30 s/Eim_COI); 53°C (45 s/18S); 55°C (30 s/COI); 50°C (45 s/ORF470); 72°C extension 90 s (18S/ORF470), 20 s (COI/Eim_COI), and a final extension at 72°C (10 min). DNA from oocysts of E. falciformis BayerHaberkorn1970 and DNA from colon content of a noninfected laboratory (NMRI) mouse were used as positive and negative controls, respectively.
All PCR products from nu 18S, mt COI, and ap ORF470 of the expected size were purified using the SAP‐Exo Kit (Jena Bioscience GmbH, Jena, Germany) and sequenced in both directions by LGC Genomics (Berlin, Germany). Quality assessment and sequence assembly was performed in Geneious v6.1.8. All sequences were submitted to the NCBI GenBank database (Accession numbers: nu 18S rDNA [MH751925‐MH752036, MK246860‐MK246868, and MK625202‐MK625210];mt COI [MH777467‐MH777593, MH755302‐MH755324, MK257106‐MK257114, and MK631866‐MK631868] and ap ORF470 [MH755325‐MH755450, MK257115‐MK257125, and MK631869‐MK631884]).
2.4. Phylogenetic analysis and inference of intraspecific genetic diversity
Datasets for each gene and a concatenated alignment (nu 18S, mt COI, and ap ORF470) were created adding closely related reference sequences available in the GenBank (Data S2).
Protein‐coding sequences (mt COI and ap ORF470) were aligned by translation using the Multiple Align algorithm and translation frame 1 with the genetic code for “mold protozoan mitochondrial,” 18S sequences were aligned using MUSCLE (Edgar, 2004), both through Geneious v6.1.8.
Phylogenetic trees for all datasets were constructed using maximum likelihood (ML) and Bayesian inference (BI) methods, implemented in PhyML v3.0 (Guindon et al., 2010) and MrBayes v3.2.6 (Huelsenbeck & Ronquist, 2001; Ronquist et al., 2012), respectively. Sequence evolution models most appropriate for each dataset were determined in JModelTest v2.1.10 (Posada, 2008). For ML trees, a bootstrap analysis with 1,000 replicates was performed, whereas MCMC for BI was run with two cold and two hot chains for 1,000,000 generations or until the split freq value was below 0.05. The concatenated dataset was analyzed using partitions and locus‐specific models. Trees were visualized with FigTree v1.4.2 (Rambaut, 2012). A haplotype network of mt COI sequences was inferred using a codon‐based alignment trimmed to 500 bp available for all isolates. Haplotype frequencies were calculated and a network was constructed with the R package “pegas” v0.11 (Paradis et al., 2018).
2.5. Multimarker genotyping PCR and high‐throughput sequencing
Samples positive for E. falciformis and E. vermiformis from Mus musculus and Eimeria spp. from Apodemus with indistinguishable 18S and COI sequences were used for a multimarker amplification using the microfluidics PCR system Fluidigm Access Array 48 x 48 (Fluidigm, San Francisco, California, USA). We used target‐specific primers (Data S3) that were designed based on the genome of E. falciformis (Heitlinger et al., 2014) to amplify exons of nuclear genes (Data S4) and coding and noncoding regions from the apicoplast genome (Data S5). Library preparation was performed according to the protocol Access Array Barcode Library for Illumina Sequencers (single direction indexing) as described by the manufacturer (Fluidigm, San Francisco, California, USA). The library was purified using Agencourt AMPure XP Reagent beads (Beckman Coulter Life Sciences, Krefeld, Germany). Quality and integrity of the library was confirmed using the Agilent 2200 Tape Station with D1000 ScreenTapes (Agilent Technologies). Sequences were generated at the Berlin Center for Genomics in Biodiversity Research (BeGenDiv) on the Illumina MiSeq platform (Illumina) in two runs, one using “v3 chemistry” with 600 cycles, the other “v2 chemistry” with 500 cycles. All sequencing raw data can be accessed through the BioProject PRJNA548431 in the NCBI Short Read Archive (SRA).
2.6. Bioinformatic analysis of multilocus sequence typing
Screening and trimming of sequencing reads was performed using the package dada2 v1.2.1 (Callahan et al., 2016). All reads were trimmed to 245 bases, while allowing a maximum of 4 expected errors (maxEE). Sorting and assignment to amplicons was performed with the package MultiAmplicon v0.1 (Heitlinger, 2019) and the most abundant sequence was recorded for each marker in each sample (recording but disregarding minority sequence in nonclonal infection for further analysis; see Data S6). Sequences were aligned using the function “AlignSeqs” from the package DECIPHER v2.10.0 (Wright, 2016) and nontarget sequences were excluded from alignments if >20% divergence was observed with other sequences (such as in cases off‐target amplification of mostly bacterial sequences). Alignments were controlled for the absence of insertions/deletions (indels) that distort the open reading frame. Prevalent multiple‐of‐3‐mere indels corresponding to homopolymeric amino acid repeats (HAARs; Heitlinger et al., 2014) of diverse length were coded as missing data due to their unclear model of evolution. The function “dudi.pcr” from the packages ade4 v1.7‐13 (Dray & Dufour, 2007) and adegenet v2.1.1 (Jombart, 2008) was used to visualize genetic distances between samples based on all markers. The code for this pipeline is available at https://github.com/VictorHJD/AA_Eimeria_Genotyping.
The alignments of the concatenated sequences were then exported. The number of informative sites was summarized using the tool DIVEIN (Deng et al., 2010) and phylogenetic trees were computed by Bayesian inference in MrBayes v3.2.6 (Huelsenbeck & Ronquist, 2001; Ronquist et al., 2012). A partitioned model was implemented to estimate the tree considering each gene separately. The analysis was performed with two runs, with 1,000,000 generations leading to a split frequency value below 0.05, and 200,000 generations were discarded as burn‐in when estimating posterior probability. Additionally, maximum likelihood trees were inferred with 1,000 bootstrap replicates in PhyML v3.0 (Guindon et al., 2010).
The topology of ML and BI trees was compared and summarized into a consensus tree with minimum clade frequency threshold of 0.95 using the program SumTrees v4.3.0 (Sukumaran & Holder, 2010).
3. RESULTS
3.1. Established markers do not recover clades corresponding to species with different host usage
We performed phylogenetic analyses using nuclear, mitochondrial, and apicoplast markers to assess the clustering of our sequences into groups of previously described species.
We inferred a phylogenetic tree of nu 18S based on 215 sequences (509–1,795 bp). Of these, 111 from parasites in house mice (M. musculus) (3 from ileum tissue, 16 from cecum tissue, and 92 from colon content) and 18 from parasites in non‐Mus rodents were generated in the present study (3 from ileum tissue, 3 from cecum tissue, 3 from colon content, and 9 from feces). To test for host specificity of house mouse Eimeria, we included reference sequences from related Eimeria species described in murid and cricetid rodents. Isosporasp. sequences identified in Talpa europaea moles were used as an outgroup. Both ML and BI rooted trees shared the general topology (Figure 2).
Figure 2.

Phylogenetic trees inferred from nuclear small ribosomal subunit (18S rDNA). Phylogenetic tree based on 18S rDNA sequences. Numbers in the branches represent Bayesian posterior probability and bootstrap value. The three collapsed groups cluster Eimeria sequences from Mus musculus of this study. Reference sequences from other rodents were included. The scale bar represents sequence divergence. Hosts for closely related sequences of E. falciformis and E. vermiformis are indicated in the expanded form of the group. * represents sequences generated in the present study. Tissue of origin is indicated in brackets. Sequences in bold were included in the multimarker phylogenetic inference
The sequences derived from Mus musculus samples clustered in three well‐supported monophyletic groups: one comprising reference sequences of E. falciformis (E. falciformis group), another of E. ferrisi (E. ferrisi group), and the third of E. vermiformis (E. vermiformis group). All three groups, however, included sequences of Eimeria from other cricetid and murid hosts without showing internal substructure linked to the observed host species infected (host usage) (Figure 2).
The phylogenetic tree of mt COI was based on 233 sequences (381–804 bp), 149 of which were obtained from Eimeria infecting house mice (3 from ileum, 16 from cecum tissue, and 130 from colon content) and 12 from non‐Mus rodents in our study (2 from ileum, 1 from cecum, 6 from colon content, and 3 from feces) (Figure 1). Similar to 18S, COI sequences derived from house mice clustered in three monophyletic groups including reference sequences of E. falciformis (n = 26), E. ferrisi (n = 109), and E. vermiformis (n = 13). Groups of E. falciformis and E. vermiformis also include sequences derived from Eimeria isolates of common voles (Mi. arvalis), bank voles (My. glareolus), short‐tailed voles (Mi. agrestis), yellow‐necked mice (A. flavicollis), or wood mice (A. sylvaticus). In addition to our isolates from M. musculus, the E. ferrisi groups contain sequences of E. burdai and E. nafuko, species described from sub‐Saharan mole rats (Heliophobius argenteocinereus). Again, the clades do not show further substructure indicative of host usage (Figure 3).
Figure 3.
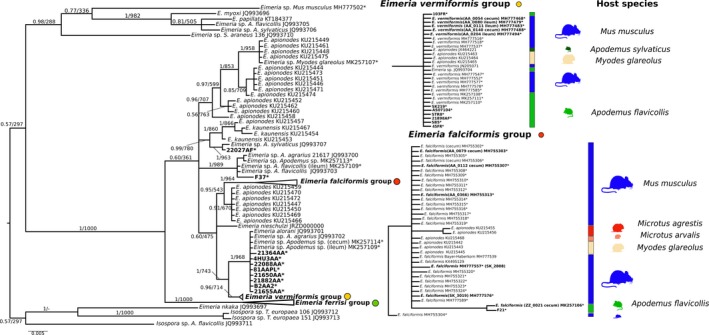
Phylogenetic trees inferred from mitochondrial cytochrome c oxidase (COI) sequences. Phylogenetic tree based on COI. Numbers in the branches represent Bayesian posterior probability and bootstrap value. The three collapsed groups cluster Eimeria sequences from Mus musculus of this study. Reference sequences from other rodents were included. The scale bar represents sequence divergence. Hosts for closely related sequences of E. falciformis and E. vermiformis are indicated in the expanded form of the group. * represents sequences generated in the present study. Tissue of origin is indicated in brackets. Sequences in bold were included in the multimarker phylogenetic inference
A phylogenetic tree of ORF470 was based on 172 sequences (Figure 4) and showed a similar topology to the COI and 18S trees. Sequences derived from Eimeriaisolates from Mus musculus (n = 125) also clustered into the same three groups. For this marker, the number of sequences available in databases from other cricetid and murid rodents is very limited, and none of the available sequences clustered within the highly supported “species clusters” of our isolates. In contrast to nu 18S and mt COI, our newly generated sequences from isolates detected in A. flavicollis and A. sylvaticus formed separate clusters that were basal to the E. falciformis group (n = 3), and outside of the E. vermiformis group (n = 4) (Figure 4).
Figure 4.
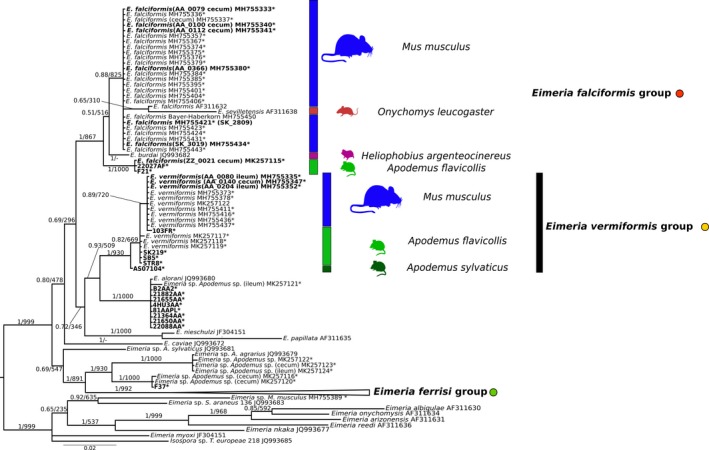
Phylogenetic trees inferred from apicoplast open reading frame 470 (ORF470) sequences. Phylogenetic tree based on ORF470 sequences. Numbers in the branches represent Bayesian posterior probability and bootstrap value. The three collapsed groups cluster Eimeria sequences from Mus musculus of this study. Reference sequences from other rodents were included. The scale bar represents sequence divergence. Hosts for closely related sequences of E. falciformis and E. vermiformis are indicated in the expanded form of the group. * represents sequences generated in the present study. Tissue of origin is indicated in brackets. Sequences in bold were included in the multimarker phylogenetic inference
To combine all available information into a single phylogenetic analysis, we used a concatenated alignment. In the tree constructed from this alignment (Data S7), the clusters from E. vermiformis and E. ferrisi observed in the individual phylogenies with 18S, COI, and ORF470 were confirmed in the concatenated tree. Sequences from the E. falciformis group were found in an unresolved basal position with E. apionodes isolates derived from Myodes sp. and Microtus sp. This result probably indicates conflicting signals for different markers and missing data.
3.2. Low genetic diversity of mt COI in rodent Eimeria isolates
With the aim to estimate the genetic diversity of isolates of Eimeria from different rodent hosts, we constructed a haplotype network (Figure 5) from 161 COI sequences obtained in this study combined with 59 previously published sequences (alignment of 459 bp without gaps). The network comprised 20 different haplotypes with up to 14 polymorphic nucleotide sites among them. The network confirms the lack of genetic differentiation of E. falciformis and E. vermiformis from some isolates described as E. apionodes in non‐Mus hosts using sequences of COI.
Figure 5.
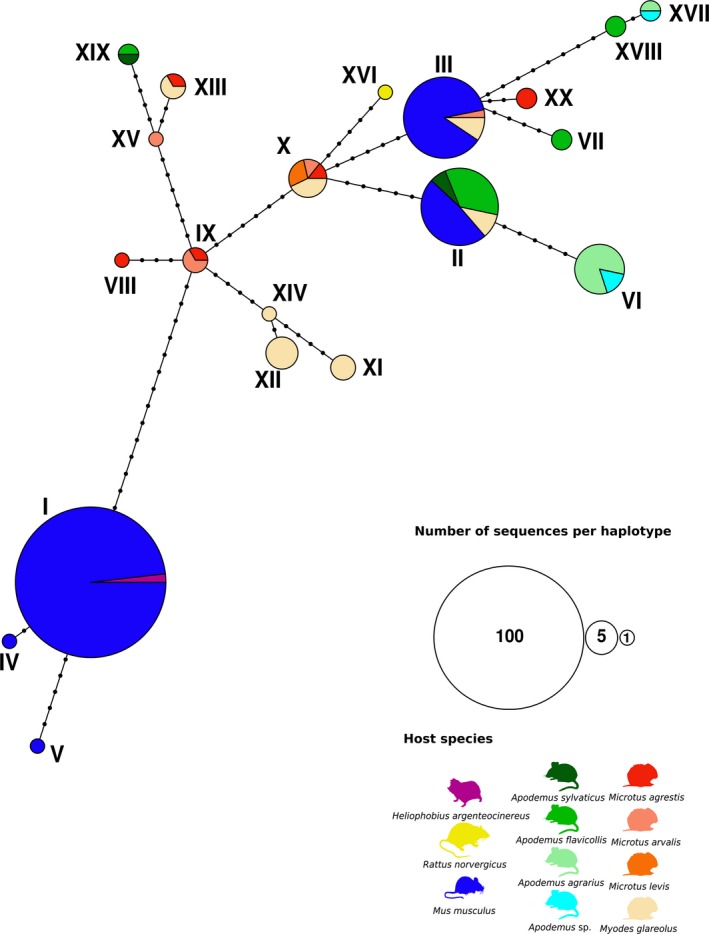
Statistical parsimony network of Eimeria spp. haplotypes for COI sequences. Network based on a 459 bp region of the gene coding for the mitochondrial cytochrome c oxidase from Eimeria isolates detected in rodents (Mus musculus, Apodemus flavicollis, A. sylvaticus, A. agrarius) caught in Europe. Previously published sequences from different species of Eimeria infecting cricetid and murid rodents were also included. Coloring of each haplotype is based on the host species from the Eimeria isolate. Every haplotype is marked with a consecutive number and its size indicates the number of sequences included on it. Each node represents a mutational step between two haplotypes
3.3. Multilocus genotyping
To determine whether markers with a higher resolution could distinguish host usage patterns for the “rodent parasite models” E. falciformis and E. vermiformis, we designed a multilocus sequence typing approach. Thirty‐five markers targeting exons in the nuclear genome (Data S4) and 5 regions of the apicoplast genome were amplified for 19 samples from Apodemus spp. hosts, 12 samples from house mice and corresponding regions from the reference genome of E. falciformis and E. vermiformis were included. All the isolates used correspond to Eimeria species with different morphology (Data S10 and S11).
A multivariate analysis identified three clusters of isolates for the nuclear markers: one group included the laboratory isolate of E. vermiformis, another the isolate of E. falciformis, and a third group only contained Eimeria isolates from Apodemusagrarius (Figure 6b). This result was corroborated by phylogenetic analysis of SNPs (2019 informative alignment columns). We excluded prevalent indels from this analysis. Indels in protein‐coding genes (all “in‐frame” with a length divisible by three) correspond to homopolymeric amino acid repeats (HAARs) and are expected in protein‐coding genes of Eimeria spp. (Heitlinger et al., 2014; Reid et al., 2014) (Data S8). Three clades were recovered in this tree (Figure 6a): Despite the apparently morphological differences (Data S10 and S11), the laboratory isolate of E. vermiformis from Mus musculus was indistinguishable from a field isolate from Apodemus flavicollis. Other isolates from house mouse and A. flavicollis and A. sylvaticus clustered in an unresolved internal relationship with house mouse E. vermiformis isolates (Group II). We note that four of five sequences for E. vermiformis from house mice were amplified from ileum tissue, the primary location of infection with this species (in contrast, E. falciformis infects primarily the cecum; Jarquín‐Díaz et al., 2019). A second clade recovered by nuclear multilocus analysis contained E. falciformis from house mice. This clade showed a well‐supported substructure in which 7 house mouse field isolates grouped with the laboratory isolate BayerHaberkorn1970 but were separated from 4 Eimeria isolates from A. flavicollis (Group III). This substructure agrees with the morphological difference previously observed based on the presence of polar granule in E. falciformis and its absence in E. apionodes (Data S10 and S11).
Figure 6.
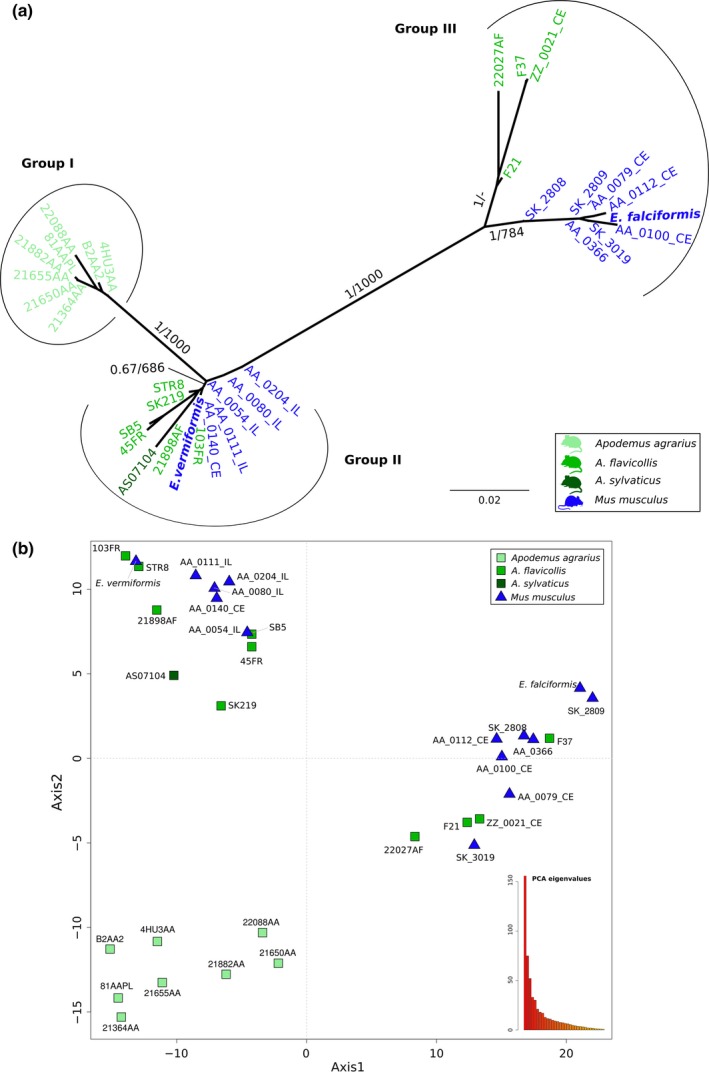
Nuclear multilocus genotyping of Eimeria isolates from Mus musculus and Apodemus. (a) The phylogenetic tree was estimated with a multimarker dataset formed with 35 nuclear markers from 31 Eimeria isolates derived from wild Mus musculus and three species of Apodemus (A. agrarius, A. sylvaticus, A. flavicollis). Eimeria falciformis and E. vermiformis sequences were included as reference. The scale bar represents sequence divergence. Color represents the host of origin for the isolates. Bootstrap support values and Bayesian posterior probabilities are shown on branches. (b) Principal component analysis based on single nucleotide polymorphisms (SNPs) from the same Eimeria isolates. Samples form three clusters. Shape indicates the genus of host and colors the species. Eigenvalues of the dimensions are shown in an insert to visualize the proportion of variance explained by the axes
Analyses based on apicoplast markers (both multivariate clustering and phylogenetic analyses; Figure 7) identified similar groups: a well‐separated cluster with isolates from A. agrarius, a cluster containing E. vermiformis, and another containing E. falciformis isolates. Some differences between the apicoplast and nuclear markers were obvious, though. Eimeria isolates from M. musculus (AA_0054_IL, AA_0080_IL, AA_0111_IL, and AA_0112_CE) were less similar to the E. vermiformis group, leading to a multivariate clustering between the E. falciformis and E. vermiformis groups (Figure 5b). This was recovered in a phylogenetic tree as isolates appeared at the end of a long branch in the E. vermiformis group (Figure 7a). In an analysis of apicoplast markers, the E. falciformis isolates from Mus were not differentiated from those from A. flavicollis. Inspection of phylogenetic trees for individual markers (Data S12) highlighted problems with the apicoplast dataset: samples that had been previously reported as coinfected with E. ferrisi (AA_0080_IL, AA_0111_IL, and AA_0112_CE), showed an aberrant clustering for different markers. Samples AA_0080_IL and AA_0111_IL clustered in the group of E. falciformis with Ap12, while AA_0112_CE clustered with Ap5, in disagreement with the consensus species trees for other markers. We conclude that for these samples E. ferrisi or even E. falciformis apicoplast sequences were likely amplified and recovered as the majority sequence.
Figure 7.
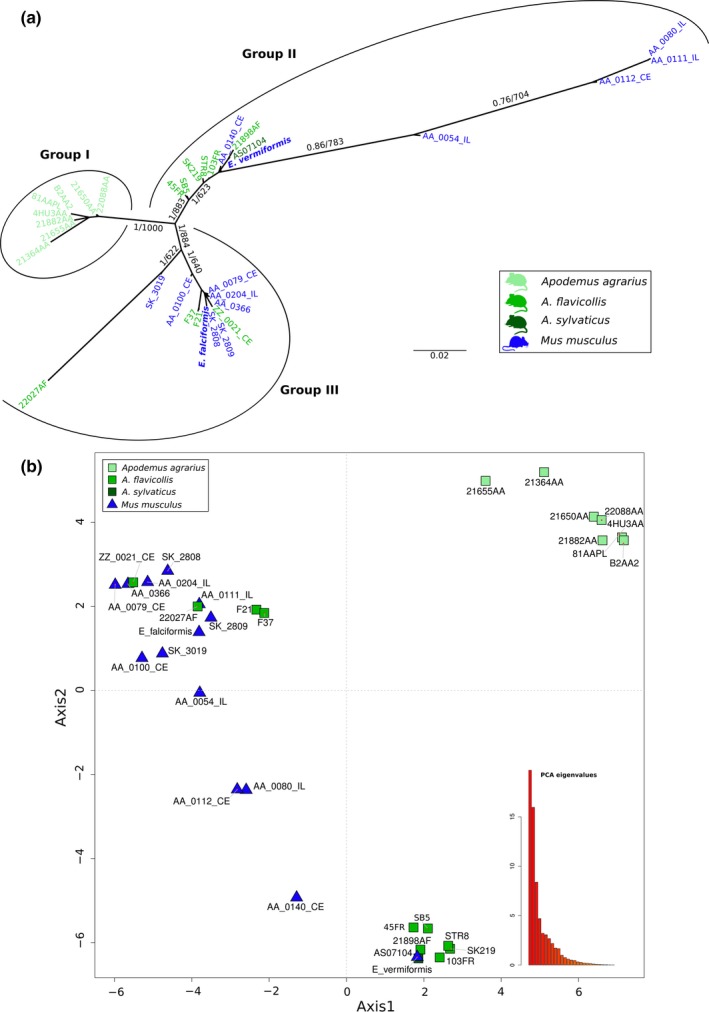
Apicoplast multilocus genotyping of Eimeria isolates from Mus musculus and Apodemus. (a) The phylogenetic tree was estimated with a multimarker dataset formed with 5 apicoplast markers from 31 Eimeria wild isolates derived from Mus musculus and three species of Apodemus (A. agrarius, A. sylvaticus, A. flavicollis). Eimeria falciformis and E. vermiformis sequences were included as reference. The scale bar represents the sequence divergence. Color represents the host of origin for the isolates. Bootstrap support values and Bayesian posterior probabilities are shown on branches. (b) Principal component analysis based on single nucleotide polymorphisms (SNPs) from the same Eimeria isolates. Samples form three clusters based on the similarities for all the SNPs. Shape indicates the genus of host and colors the species. Eigenvalues of the dimensions are shown in an insert to visualize the proportion of variance explained by the axes
4. DISCUSSION
We studied whether coccidia with different host usage can be distinguished with currently used molecular markers, using the example of Eimeria species in house mice and related rodents. We found that commonly used phylogenetic markers, nu 18S rDNA and mt COI, are not sufficiently variable to differentiate parasite isolates that would be regarded as separate species based on host usage. The relatively rarely used marker ap ORF470 from the apicoplast genome seems to provide slightly better resolution. We developed a multilocus genotyping approach to show that E. falciformis from the house mouse can likely be distinguished from related isolates from other hosts based on nuclear markers. In contrast, even with this high‐resolution approach E. vermiformis from house mice and isolates from other host species were found in a nested and unresolved cluster.
Phylogenies derived from each of the analyzed markers (esp. 18S) confirmed the topology of rodent Eimeria species observed before at deeper nodes of the phylogeny (Kvičerová et al., 2008; Ogedengbe et al., 2018; Zhao & Duszynski, 2001a). At the tips of the phylogeny, 18S sequences of E. falciformis and E. vermiformis isolates clustered with isolates from hosts of different genera or even families (Figure 2). This result was expected to some extent, as phylogenetic analyses with 18S sequences usually fail to separate closely related parasites isolated from closely related hosts (Ogedengbe et al., 2018).
Previous studies described COI as a universal barcode variable enough to resolve relationships between coccidians, including Eimeria (Ogedengbe et al., 2011, 2018). We therefore expected to differentiate our house mouse isolates from species found in other hosts using COI. Neither phylogenetic (Figure 3) nor haplotype inference (Figure 5), however, supported differentiation of E. falciformis and E. vermiformis from some of the isolates described as E. apionodes. Many of the COI sequences were even identical for isolates from different hosts. Limited resolution of COI outside of metazoans has been reported before (Meyer & Paulay, 2005). Rodent hosts of Eimeria, in the families Muridae (Mus, Rattus, Apodemus) and Cricetidae (Myodes, Microtus), diverged around 25 million years ago (Churakov et al., 2010; Steppan, Adkins, & Anderson, 2004) and it seems possible that COI of coccidia evolves at such slow rates that it fails to differentiate Eimeria species with similar divergence. We stress that for rodent coccidia, COI should not be assumed to resolve bona fide species with different host usage.
The potential of the apicoplast marker ORF470 to distinguish rodent Eimeria species has been highlighted before (Ogedengbe et al., 2015; Zhao & Duszynski, 2001b), but few studies have followed the recommendation to use this marker. Consequently, few database sequences are available. Phylogenetic analysis of these sequences (Figure 4) separates our three species clusters well and shows hints of internal structure separating E. apionodes derived from A. flavicollis from house mouse isolates. Our work increases the number of sequences available for ORF470 and supports its use as a marker for discrimination of Eimeria species.
To distinguish E. falciformis, E. vermiformis (from house mice), and E. apionodes (from Apodemus spp.), we established and used a multilocus sequence typing protocol. Our multilocus approach supports a differentiation of E. falciformis (infecting the house mouse; Eimer, 1870; Haberkorn, 1970) from E. apionodes (infecting A. flavicolis; Pellérdy, 1954). The same approach was unable to distinguish M. musculus‐derived E. vermiformis isolates from one “E. apionodes” isolate from A. flavicollis (Figures 2, 3, 4, 6 and 7). This suggests a broad host range of genetically indistinguishable Eimeria isolates which have been assigned to paraphyletic species E. apionodes and E. vermiformis.
Multilocus genotyping using apicoplast markers showed some discrepancies with the nuclear analysis. These discrepancies can be attributed to double infections previously discovered in those particular isolates (Jarquín‐Díaz et al., 2019). Compared to the nuclear genome, the apicoplast genome is present in much higher copy numbers (Heitlinger et al., 2014). This, combined with more conserved primer binding sites, can lead to amplification of nontarget sequences such as those of the prevalent E. ferrisi (Jarquín‐Díaz et al., 2019) creating artificial “chimeric” isolates in case of double infections.
We use our system also as a test case whether the commonly used markers (18S, COI) provide enough resolution to assess parasite specificity. We conclude that unresolved genetic clusters and monomorphic haplotypes currently identified via 18S and COI genotyping should not be assumed to indicate parasite species with generalist host usage. Novel nuclear markers are needed in addition to ORF470 to analyze host species specificity of rodent Eimeria. Care must be taken to avoid potential artifacts introduced by double infection and mixed amplification.
Whether other Eimeria species from different rodent hosts are indeed phylogenetically distinguishable species (or whether genetically differentiable clusters show different host usage) is still an open question. This question needs to be addressed more broadly with markers providing higher resolution than 18S or COI. This question is highly relevant as hypotheses, assumptions, and predictions concerning host–parasite interactions from evolutionary (Adamson & Caira, 1994; Combes, 2001; Poulin, Krasnov, & Mouillot, 2011; Schmid‐Hempel, 2011), ecological (Fenton & Brockhurst, 2008; Forbes, Muma, & Smith, 2002; Kassen, 2002), and mechanistic (Rathore et al., 2003) perspectives depend on the placement of parasite species in the specialist–generalist continuum (Schmid‐Hempel, 2011).
CONFLICT OF INTEREST
None declared.
AUTHOR CONTRIBUTIONS
VHJD and EH designed the project and obtained funding. VHJD, AM, TRS, JJ, KB, ST, and JK obtained data, VHJD, AB, and EH designed the analysis, VHJD, AB, and EH performed the analysis. VHJD, EH, and JK interpreted the results. VHJD and EH wrote the manuscript with contributions from all other authors. EH supervised the project.
Supporting information
ACKNOWLEDGMENTS
We thank Jaroslav Piálek and his team (Institute of Vertebrate Biology, AS CR, Brno, Department of Population Biology in Studenec) for help with catching and genotyping of house mice. Deborah Dymke and Julia Murata assisted the processing of samples in the Heitlinger group. We acknowledge the support by the German Research Foundation (DFG) and the Open Access Publication Fund of Humboldt University Berlin. We thank Susan Mbedi and Sarah Sparmann from the Berlin Center for Genomics in Biodiversity Research (BeGenDiv) for their technical guidance during the library preparation and Daniel P. Benesh for language revision and proofreading of the manuscript.
Jarquín‐Díaz VH, Balard A, Mácová A, et al. Generalist Eimeria species in rodents: Multilocus analyses indicate inadequate resolution of established markers. Ecol Evol. 2020;10:1378–1389. 10.1002/ece3.5992
Funding information
This work was supported by the German Foundation of Scientific Research (DFG) [grant number: 285969495/HE 7320/2‐1 to EH] and the German Academic Exchange Service (DAAD) [grant number: 57214224, scholarship to VHJD] and the Research Training Group 2046 "Parasite Infections: From Experimental Models to Natural Systems" (GRK2046) [associated student VHJD]. AM and JK were supported by the Czech Science Foundation [grant number: 17‐19831S].
DATA AVAILABILITY STATEMENT
DNA sequences: nu 18S rDNA [MH751925‐MH752036, MK246860‐MK246868 and MK625202‐MK625210]; mt COI [MH777467‐MH777593, MH755302‐MH755324, MK257106‐MK257114, and MK631866‐MK631868], and ap ORF470 [MH755325‐MH755450, MK257115‐MK257125, and MK631869‐MK631884]).
All sequencing raw data can be accessed through the BioProject PRJNA548431 in the NCBI Short Read Archive (SRA).
The code for the pipeline used in the multilocus type analysis is available on github at https://github.com/VictorHJD/AA_Eimeria_Genotyping.
REFERENCES
- Abramson, N. I. , Rodchenkova, E. N. , & Kostygov, A. Y. (2009). Genetic variation and phylogeography of the bank vole (Clethrionomys glareolus, Arvicolinae, Rodentia) in Russia with special reference to the introgression of the mtDNA of a closely related species, red‐backed vole (Cl. Rutilus). Russian Journal of Genetics, 45(5), 533 10.1134/S1022795409050044 [DOI] [PubMed] [Google Scholar]
- Adamson, M. L. , & Caira, J. N. (1994). Evolutionary factors influencing the nature of parasite specificity. Parasitology, 109(S1), S85–S95. 10.1017/S0031182000085103 [DOI] [PubMed] [Google Scholar]
- Ball, S. J. , & Lewis, D. C. (1984). Eimeviu (Protozoa: Coccidia) in wild populations of some British rodents. Journal of Zoology, 202(3), 373–381. [Google Scholar]
- Barta, J. R. , Martin, D. S. , Liberator, P. A. , Dashkevicz, M. , Anderson, J. W. , Feighner, S. D. , … Profous‐Juchelka, H. (1997). Phylogenetic relationships among eight Eimeria species infecting domestic fowl inferred using complete small subunit ribosomal DNA sequences. The Journal of Parasitology, 83(2), 262–271. 10.2307/3284453 [DOI] [PubMed] [Google Scholar]
- Becker, E. R. (1934). Coccidia and coccidiosis of domesticated, game and laboratory animals and of man. Berlin; Verlag Paul Parey: Budapest, Hungary: Akademiai Kiado. [Google Scholar]
- Callahan, B. J. , McMurdie, P. J. , Rosen, M. J. , Han, A. W. , Johnson, A. J. A. , & Holmes, S. P. (2016). DADA2: High‐resolution sample inference from Illumina amplicon data. Nature Methods, 13(7), 581 10.1038/nmeth.3869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churakov, G. , Sadasivuni, M. K. , Rosenbloom, K. R. , Huchon, D. , Brosius, J. , & Schmitz, J. (2010). Rodent evolution: Back to the root. Molecular Biology and Evolution, 27(6), 1315–1326. 10.1093/molbev/msq019 [DOI] [PubMed] [Google Scholar]
- Combes, C. (2001). Parasitism: The ecology and evolution of intimate interactions. Chicago, IL: University of Chicago Press. [Google Scholar]
- De Vos, A. J. (1970). Studies on the host range of Eimeria chinchillae de Vos & van der Westhuizen, 1968. The Onderstepoort Journal of Veterinary Research, 37(1), 29–36. [PubMed] [Google Scholar]
- Deng, W. , Maust, B. , Nickle*, D. , Learn**, G. , Liu, Y. I. , Heath, L. , … Mullins, J. (2010). DIVEIN: A web server to analyze phylogenies, sequence divergence, diversity, and informative sites. BioTechniques, 48(5), 405–408. 10.2144/000113370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dray, S. , & Dufour, A. B. (2007). The ade4 package: Implementing the duality diagram for ecologists. Journal of Statistical Software, 22(4), 1–20. [Google Scholar]
- Ďureje, Ľ. , Macholán, M. , Baird, S. J. , & Piálek, J. (2012). The mouse hybrid zone in Central Europe: From morphology to molecules. Folia Zoologica, 61(3–4), 308–319. 10.25225/fozo.v61.i3.a13.2012 [DOI] [Google Scholar]
- Duszynski, D. W. (Ed.). (2011). Eimeria e LS (pp. 1192–1196). Chichester, UK: John Wiley & Sons, Ltd; 10.1002/9780470015902.a0001962.pub2 [DOI] [Google Scholar]
- Duszynski, D. W. , Eastham, G. , & Yates, T. L. (1982). Eimeria from jumping mice (Zapus spp.): A new species and genetic and geographic features of Z. hudsonius luteus . The Journal of Parasitology, 68(6), 1146–1148. [Google Scholar]
- Edgar, R. C. (2004). MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32(5), 1792–1797. 10.1093/nar/gkh340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eimer, T. (1870). Ueber Die Ei‐Oder Kugelförmigen Sogenannten Psorospermien Der Wirbelthiere: Ein Beitrag Zur Entwicklungsgeschichte Der Gregarinen Und Zur Kenntniss Dieser Parasiten Als Krankheitsursache. A. Stuber. [Google Scholar]
- Fenton, A. , & Brockhurst, M. A. (2008). The role of specialist parasites in structuring host communities. Ecological Research, 23(5), 795–804. 10.1007/s11284-007-0440-6 [DOI] [Google Scholar]
- Forbes, M. R. , Muma, K. E. , & Smith, B. P. (2002). Diffuse coevolution: Constraints on a generalist parasite favor use of a dead end host. Ecography, 25(3), 345–351. 10.1034/j.1600-0587.2002.250311.x [DOI] [Google Scholar]
- Guindon, S. , Dufayard, J. F. , Lefort, V. , Anisimova, M. , Hordijk, W. , & Gascuel, O. (2010). New algorithms and methods to estimate maximum‐likelihood phylogenies: Assessing the performance of PhyML 3.0. Systematic Biology, 59(3), 307–321. 10.1093/sysbio/syq010 [DOI] [PubMed] [Google Scholar]
- Haberkorn, A. (1970). Die Entwicklung von Eimeria falciformis (Eimer 1870) in der weißen Maus (Mus musculus). Zeitschrift Für Parasitenkunde, 34(1), 49–67. [Google Scholar]
- Heitlinger, E. (2019). MultiAmpliconv0.1. Retrieved from: https://derele.github.io/MultiAmplicon/index.html
- Heitlinger, E. , Spork, S. , Lucius, R. , & Dieterich, C. (2014). The genome of Eimeria falciformis‐reduction and specialization in a single host apicomplexan parasite. BMC Genomics, 15(1), 696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hnida, J. A. , & Duszynski, D. W. (1999a). Taxonomy and systematics of some Eimeria species of murid rodents as determined by the ITS1 region of the ribosomal gene complex. Parasitology, 119(4), 349–357. [DOI] [PubMed] [Google Scholar]
- Hnida, J. A. , & Duszynski, D. W. (1999b). Taxonomy and phylogeny of some Eimeria (Apicomplexa: Eimeriidae) species of rodents as determined by polymerase chain reaction/restriction‐fragment‐length polymorphism analysis of 18s rDNA. Parasitology Research, 85(11), 887–894. [DOI] [PubMed] [Google Scholar]
- Hnida, J. A. , Wilson, W. D. , & Duszynski, D. W. (1998). A New Eimeria Species (Apicomplexa: Eimeriidae) Infecting Onychomys Species (Rodentia: Muridae) in New Mexico and Arizona. The Journal of Parasitology, 84(6), 1207– 10.2307/3284675 [DOI] [PubMed] [Google Scholar]
- Huelsenbeck, J. P. , & Ronquist, F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics, 17(8), 754–755. 10.1093/bioinformatics/17.8.754 [DOI] [PubMed] [Google Scholar]
- Jaarola, M. , & Searle, J. B. (2002). Phylogeography of field voles (Microtus agrestis) in Eurasia inferred from mitochondrial DNA sequences. Molecular Ecology, 11(12), 2613–2621. 10.1046/j.1365-294X.2002.01639.x [DOI] [PubMed] [Google Scholar]
- Jarquín‐Díaz, V. H. , Balard, A. , Jost, J. , Kraft, J. , Dikmen, M. N. , Jana, K. , & Heitlinger, E. (2019). Detection and quantification of house mouse Eimeria at the species level–challenges and solutions for the assessment of Coccidia in wildlife. International Journal for Parasitology: Parasites and Wildlife, 10, 29–40. 10.1016/j.ijppaw.2019.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jombart, T. (2008). adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics, 24(11), 1403–1405. 10.1093/bioinformatics/btn129 [DOI] [PubMed] [Google Scholar]
- Kassen, R. (2002). The experimental evolution of specialists, generalists, and the maintenance of diversity. Journal of Evolutionary Biology, 15(2), 173–190. 10.1046/j.1420-9101.2002.00377.x [DOI] [Google Scholar]
- Kvičerová, J. , & Hypša, V. (2013). Host‐parasite incongruences in rodent Eimeria suggest significant role of adaptation rather than cophylogeny in maintenance of host specificity. PLoS ONE, 8(7), e63601 10.1371/journal.pone.0063601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- KvičerovÁ, J. , Mikeš, V. , & Hypša, V. (2011). Third lineage of rodent eimerians: Morphology, phylogeny and re‐description of Eimeria myoxi (Apicomplexa: Eimeriidae) from Eliomys quercinus (Rodentia: Gliridae). Parasitology, 138(10), 1217–1223. [DOI] [PubMed] [Google Scholar]
- Kvičerová, J. , Pakandl, M. , & Hypša, V. (2008). Phylogenetic relationships among Eimeria spp. (Apicomplexa, Eimeriidae) infecting rabbits: Evolutionary significance of biological and morphological features. Parasitology, 135(4), 443–452. [DOI] [PubMed] [Google Scholar]
- Lainson, R. , & Shaw, J. J. (1990). Coccidia of Brazilian mammals: Eimeria corticulata n. sp. (Apicomplexa: Eimeriidae) from the anteater Tamandua tetradactyla (Xenarthra: Myrmecophagidae) and Eimeria zygodontomyis n. sp. from the cane mouse Zygodontomys lasiurus (Rodentia: Cricetidae). The Journal of Protozoology, 37(1), 51–54. [DOI] [PubMed] [Google Scholar]
- Levine, N. D. , & Ivens, V. (1965). The coccidian parasites (Protozoa, Sporozoa) of rodents 33. Illinois Biological Monographs, 33. [Google Scholar]
- Levine, N. D. , & Ivens, V. (1988). Cross transmission of Eimeria spp. (Protozoa, Apicomplexa) of rodents—a review. The Journal of Protozoology, 35(3), 434–437. [DOI] [PubMed] [Google Scholar]
- Long, P. L. , & Joyner, L. P. (1984). Problems in the identification of species of Eimeria . The Journal of Protozoology, 31(4), 535–541. 10.1111/j.1550-7408.1984.tb05498.x [DOI] [PubMed] [Google Scholar]
- Mácová, A. , Hoblíková, A. , Hypša, V. , Stanko, M. , Martinů, J. , & Kvičerová, J. (2018). Mysteries of host switching: Diversification and host specificity in rodent‐coccidia associations. Molecular Phylogenetics and Evolution, 127, 179–189. 10.1016/j.ympev.2018.05.009 [DOI] [PubMed] [Google Scholar]
- MacPherson, J. M. , & Gajadhar, A. A. (1993). Differentiation of seven Eimeria species by random amplified polymorphic DNA. Veterinary Parasitology, 45(3–4), 257–266. 10.1016/0304-4017(93)90080-7 [DOI] [PubMed] [Google Scholar]
- Marquardt, W. C. (1981). Host and site specificity in the coccidia: A perspective 1. The Journal of Protozoology, 28(2), 243–244. 10.1111/j.1550-7408.1981.tb02841.x [DOI] [Google Scholar]
- Mesfin, G. M. , & Bellamy, J. E. (1978). The life cycle of Eimeria falciformis var. pragensis (Sporozoa: Coccidia) in the mouse, Mus musculus. The Journal of Parasitology, 64(4), 696–705. [PubMed] [Google Scholar]
- Meyer, C. P. , & Paulay, G. (2005). DNA barcoding: Error rates based on comprehensive sampling. PLoS Biology, 3(12), e422 10.1371/journal.pbio.0030422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogedengbe, J. D. , Hanner, R. H. , & Barta, J. R. (2011). DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina, Apicomplexa, Alveolata). International Journal for Parasitology, 41(8), 843–850. 10.1016/j.ijpara.2011.03.007 [DOI] [PubMed] [Google Scholar]
- Ogedengbe, J. D. , Ogedengbe, M. E. , Hafeez, M. A. , & Barta, J. R. (2015). Molecular phylogenetics of eimeriid coccidia (Eimeriidae, Eimeriorina, Apicomplexa, Alveolata): A preliminary multi‐gene and multi‐genome approach. Parasitology Research, 114(11), 4149–4160. 10.1007/s00436-015-4646-1 [DOI] [PubMed] [Google Scholar]
- Ogedengbe, M. E. , El‐Sherry, S. , Ogedengbe, J. D. , Chapman, H. D. , & Barta, J. R. (2018). Phylogenies based on combined mitochondrial and nuclear sequences conflict with morphologically defined genera in the eimeriid coccidia (Apicomplexa). International Journal for Parasitology, 48(1), 59–69. 10.1016/j.ijpara.2017.07.008 [DOI] [PubMed] [Google Scholar]
- Paradis, E. , Jombart, T. , Brian, K. , Schliep, K. , Potts, A. , Winter, D. , & Kamvar, Z. N. (2018). Population and evolutionary genetics analysis system. R Package Version 11, 1. [Google Scholar]
- Pellérdy, L. (1954). Zur Kenntnis der Coccidien aus Apodemus flavicollis . Acta Veterinaria Academiae Scientarum Hungaricae, 4, 187–191. [Google Scholar]
- Posada, D. (2008). jModelTest: Phylogenetic model averaging. Molecular Biology and Evolution, 25(7), 1253–1256. 10.1093/molbev/msn083 [DOI] [PubMed] [Google Scholar]
- Poulin, R. , Krasnov, B. R. , & Mouillot, D. (2011). Host specificity in phylogenetic and geographic space. Trends in Parasitology, 27(8), 355–361. 10.1016/j.pt.2011.05.003 [DOI] [PubMed] [Google Scholar]
- Rambaut, A. (2012). FigTree v1.4. Available at: http://tree.bio.ed.ac.uk/software/figtree/ (Accessed October 2017). [Google Scholar]
- Rathore, D. , Hrstka, S. C. , Sacci, J. B. , De la Vega, P. , Linhardt, R. J. , Kumar, S. , & McCutchan, T. F. (2003). Molecular mechanism of host specificity in Plasmodium falciparum infection role of circumsporozoite protein. Journal of Biological Chemistry, 278(42), 40905–40910. [DOI] [PubMed] [Google Scholar]
- Reid, A. J. , Blake, D. P. , Ansari, H. R. , Billington, K. , Browne, H. P. , Bryant, J. , … Pain, A. (2014). Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Research, 24(10), 1676–1685. 10.1101/gr.168955.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reutter, B. A. , Petit, E. , Brünner, H. , & Vogel, P. (2003). Cytochrome b haplotype divergences in West European Apodemus. Mammalian Biology‐Zeitschrift Für Säugetierkunde, 68(3), 153–164. 10.1078/1616-5047-00077 [DOI] [Google Scholar]
- Ronquist, F. , Teslenko, M. , Van Der Mark, P. , Ayres, D. L. , Darling, A. , Höhna, S. , … Huelsenbeck, J. P. (2012). MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61(3), 539–542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid Hempel, P. (2011). Evolutionary parasitology the integrated study of infections, immunology, ecology, and genetics (No. 574.5249 S2). Reprinted. New York: Oxford University Press. [Google Scholar]
- Steppan, S. J. , Adkins, R. M. , & Anderson, J. (2004). Phylogeny and divergence‐date estimates of rapid radiations in muroid rodents based on multiple nuclear genes. Systematic Biology, 53(4), 533–553. 10.1080/10635150490468701 [DOI] [PubMed] [Google Scholar]
- Sukumaran, J. , & Holder, M. T. (2010). DendroPy: A Python library for phylogenetic computing. Bioinformatics, 26(12), 1569–1571. 10.1093/bioinformatics/btq228 [DOI] [PubMed] [Google Scholar]
- Tenter, A. M. , Barta, J. R. , Beveridge, I. , Duszynski, D. W. , Mehlhorn, H. , Morrison, D. A. , … Conrad, P. A. (2002). The conceptual basis for a new classification of the coccidia. International Journal for Parasitology, 32(5), 595–616. 10.1016/S0020-7519(02)00021-8 [DOI] [PubMed] [Google Scholar]
- Todd, K. S. Jr , & Hammond, D. M. (1968). Life cycle and host specificity of Eimeria callospermophili Henry, 1932 from the Uinta ground squirrel Spermophilus armatus. The Journal of Protozoology, 15(1), 1–8. [DOI] [PubMed] [Google Scholar]
- Todd, K. S. Jr , & Lepp, D. L. (1971). The life cycle of Eimeria vermiformis Ernst, Chobotar and Hammond, 1971 in the Mouse Mus musculus 1. The Journal of Protozoology, 18(2), 332–337. [DOI] [PubMed] [Google Scholar]
- Turner, W. C. , Penzhorn, B. L. , & Getz, W. M. (2016). Description of 3 new species of Eimeria (Apicomplexa: Eimeriidae) from Springbok (Antidorcas marsupialis) in Namibia. Comparative Parasitology, 83(2), 202–212. [Google Scholar]
- Upton, S. J. , McAllister, C. T. , Brillhart, D. B. , Duszynski, D. W. , & Wash, C. D. (1992). Cross‐transmission studies with Eimeria arizonensis‐like oocysts (Apicomplexa) in New World rodents of the genera Baiomys, Neotoma, Onychomys, Peromyscus, and Reithrodontomys (Muridae). The Journal of Parasitology, 78, 406–413. 10.2307/3283636 [DOI] [PubMed] [Google Scholar]
- Wash, C. D. , Duszynski, D. W. , & Yates, T. L. (1985). Eimerians from different karyotypes of the Japanese wood mouse (Apodemus spp.), with descriptions of two new species and a redescription of Eimeria montgomeryae Lewis and Ball, 1983. The Journal of Parasitology, 71, 808–814. [PubMed] [Google Scholar]
- Wilber, P. G. , Duszynski, D. W. , Upton, S. J. , Seville, R. S. , & Corliss, J. O. (1998). A revision of the taxonomy and nomenclature of the Eimeria spp. (Apicomplexa: Eimeriidae) from rodents in the Tribe Marmotini (Sciuridae). Systematic Parasitology, 39(2), 113–135. [Google Scholar]
- Wright, E. S. (2016). Using DECIPHER v2. 0 to analyze big biological sequence data in R. R Journal, 8(1), 352–359. [Google Scholar]
- Zhao, X. , & Duszynski, D. (2001a). Molecular phylogenies suggest the oocyst residuum can be used to distinguish two independent lineages of Eimeria spp in rodents. Parasitology Research, 87(8), 638–643. [DOI] [PubMed] [Google Scholar]
- Zhao, X. , & Duszynski, D. W. (2001b). Phylogenetic relationships among rodent Eimeria species determined by plastid ORF470 and nuclear 18S rDNA sequences. International Journal for Parasitology, 31(7), 715–719. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
DNA sequences: nu 18S rDNA [MH751925‐MH752036, MK246860‐MK246868 and MK625202‐MK625210]; mt COI [MH777467‐MH777593, MH755302‐MH755324, MK257106‐MK257114, and MK631866‐MK631868], and ap ORF470 [MH755325‐MH755450, MK257115‐MK257125, and MK631869‐MK631884]).
All sequencing raw data can be accessed through the BioProject PRJNA548431 in the NCBI Short Read Archive (SRA).
The code for the pipeline used in the multilocus type analysis is available on github at https://github.com/VictorHJD/AA_Eimeria_Genotyping.