

Abstract
OBJECTIVE:
Nociceptin (N/OFQ) is an anti-stress neuropeptide transmitter in the brain that counteracts corticotrophin-releasing factor (CRF)-mediated stress and anxiety symptoms during drug and alcohol withdrawal. It also inhibits the release of a wide array of neurotransmitters including dopamine and glutamate, which allows for it to block the rewarding properties of cocaine. Chronic cocaine administration in rodents has been shown to decrease N/OFQ and increase nociceptive opioid peptide receptors (NOP) in the nucleus accumbens. No previous studies have reported on the in vivo status of NOP in chronic cocaine-abusing humans.
METHODS:
[11C]NOP-1A and PET were used to measure in vivo NOP binding in 24 cocaine use disorder (CUD) and 25 healthy control subjects matched for age, sex, and smoking status. CUD subjects with no comorbid psychiatric or medical disorders were scanned following two weeks of outpatient monitored abstinence. [11C]NOP-1A distribution volume (VT) were measured with kinetic analysis using the arterial input function in brain regions that mediate reward and stress behaviors. CUD subjects were also followed for 12-weeks after the PET scans to document relapse and relate it to VT.
RESULTS:
We observed a significant increase in [11C]NOP-1A VT in CUD compared to healthy controls. This increase, which was generalized across all the regions of interest (~10%), was most prominent in the midbrain, ventral striatum and cerebellum. However, increased VT in these regions did not predict relapse.
CONCLUSION:
Increased NOP in CUD suggests an adaptive response to decreased N/OFQ, or increased CRF transmission, or both. Future studies should examine the interactions between CRF and NOP to elucidate their role in negative reinforcement and relapse. NOP agonist medications to enhance N/OFQ should be explored as a therapeutic to treat CUD.
Keywords: [11C]NOP-1A, positron emission tomography (PET), cocaine use disorders nociceptin/orphanin FQ peptide receptors (NOP)
INTRODUCTION
The endogenous neuropeptide nociceptin (N/OFQ) and its target nociceptive opioid peptide receptor (NOP) are structurally similar to dynorphin A and its target kappa opioid receptors (1, 2). However, N/OFQ has no affinity for the mu, kappa, or delta opioid receptors; and endogenous opioids such as endorphins, enkephalins and dynorphin do not bind to NOP. N/OFQ is an anti-stress/resilience mediating neuropeptide in the brain that counteracts stress and anxiety, which is primarily mediated by corticotrophin releasing factor (CRF), norepinephrine, orexin, vasopressin and dynorphin (3). N/OFQ blocks stress-induced analgesia, anorexia, and anxious behaviors in rodents (4–6). N/OFQ knockout mice show increased anxiety-like behaviors compared to wild-type mice (7). N/OFQ also blocks the rewarding properties of cocaine in behavioral paradigms including the acquisition of conditioned place preference (8, 9). In agreement with this, increases in the rewarding properties of cocaine have been observed in NOP knock-out mice (10). Studies have also shown that N/OFQ can reverse the psychomotor sensitization response that develops following chronic exposure to cocaine (considered a rodent model for drug craving) (9, 11, 12). N/OFQ also reduces cocaine-induced dopamine release in the nucleus accumbens (12–14). Two previous studies in rodents have evaluated the effects of chronic cocaine on N/OFQ and NOP. In the first report that used both radioimmunoassay and immuno-autoradiographic techniques, animals chronically administered cocaine showed decreased N/OFQ in the nucleus accumbens, substantia nigra/ventral tegmental area, and medial (but not lateral) caudate-putamen compared to controls (15). In a replication of this observation the expression of N/OFQ precursor protein (pro-N/OFQ) was found to be decreased in the nucleus accumbens and the lateral (but not medial) caudate-putamen in chronic cocaine rodents (16). NOP gene expression (mRNA) examined in the same chronic cocaine animals were significantly increased in the nucleus accumbens (~ 50%) but decreased in the caudate-putamen (~ 50–100%) relative to controls. In summary, several basic investigations suggest that N/OFQ and NOP alterations modulate drug-induced reward in cocaine use disorders (CUD). They also suggest a role for N/OFQ in balancing the functions of the stress-mediating neuropeptides such as CRF that promote negative reinforcement and lead to relapse (3).
No previous in vivo studies have characterized the N/OFQ system in chronic cocaine abusing humans due to the lack of a radiotracer to image NOP. [11C]NOP-1A, first radiolabeled and validated by the NIMH Molecular Imaging Branch, has been successfully used to measure NOP in humans (17–19). Here, we used [11C]NOP-1A and PET to measure the in vivo binding to NOP in 24 recently abstinent subjects with CUD and 25 healthy comparison subjects (HC). Following the [11C]NOP-1A PET scan, CUD subjects were enrolled into a follow-up protocol in which they were monitored three times per week for 12-weeks using a contingency management approach. The objective of this follow-up was to document relapse and relate it with NOP. We hypothesized an increase in [11C]NOP-1A binding in CUD compared to HC in brain regions that mediate reward and stress behaviors. Such a finding would support the notion that NOP is upregulated in response to decreased N/OFQ in CUD.
MATERIALS AND METHODS
Human subjects
The University of Pittsburgh Human Research Protection Office and Radioactive Drug Research Committee approved the study. All subjects provided written informed consent. CUD subjects interested in abstaining from cocaine were recruited through advertisements displayed at addiction clinics, buses, newspapers and websites. Subjects were also recruited via a research registry administered by the University of Pittsburgh (Pitt+Me).
Study criteria for CUD subjects were [1] males or females between 18 and 50 years old; [2] fulfill DSM-5 criteria for CUD as assessed by the Structured Clinical Interview for DSM-5 (20) (SCID-5); [3] no other current psychiatric or drug/alcohol use disorder diagnosis except CUD (tobacco use disorder and recreational cannabis/alcohol use were not exclusionary); [4] no medical or neurological illness; [5] not currently taking any medical or psychotropic medications; [6] not currently pregnant; [7] no recent research or occupational radioactivity exposure; and [8] no contraindications for magnetic resonance imaging (MRI). Study criteria for healthy controls were: [1] males of females between 18 and 50 years old; [2] absence of present and past psychiatric and addictive disorders as assessed by SCID-5; and criteria 4 to 8 above.
Demographic, substance use and family history of addiction data were all recorded as recommended by NIDA in the Substance abuse and addiction assessments core PhenX Toolkit (https://www.phenxtoolkit.org). Clinical assessments performed included the Substance Use Inventory, Tiffany Cocaine Craving Questionnaire (CCQ), Fagerstrom Test for Nicotine Dependence (FTND), Perceived Stress Scale (PSS), Hamilton Anxiety Rating Scale (HAM-A), and Barratt Simplified Measure of Social Status (BSMSS). CUD subjects were monitored three times per week in the outpatient setting for abstinence from cocaine for a minimum of ten days before the PET scans. CUD subjects were only scanned if a urine drug screen confirmed the absence of all drugs of abuse (including cannabis) on scan day. Subjects were not allowed to use any nicotine products following arrival to the PET facility, i.e., their last cigarette was a minimum of 2 hours prior to the PET scan. Female subjects were scanned irrespective of menstrual cycle phase or hormonal contraceptive use
PET image acquisition and analysis
Before PET imaging, a magnetization prepared rapid gradient echo structural MRI scan was obtained using a Siemens 3T Trio scanner for region determination. [11C]NOP-1A PET imaging sessions were conducted with the Siemens Biograph64 mCT scanner using methods previously described (17, 21–24). Following a low-dose CT scan of the brain that was acquired for attenuation correction, subjects received an intravenous bolus injection of [11C]NOP-1A, and emission data was collected for 70 minutes. Metabolite-corrected arterial input function and plasma free fraction (fP) measurements were performed in all subjects. PET data were reconstructed by filtered back projection using the camera’s built-in software. The image analysis software PMOD, version 3.802 (PMOD Technologies LLC, Zurich, Switzerland) was used to conduct frame-to-frame motion correction for head movement. The MR-PET image alignment was performed using a normalized mutual information algorithm. Regions of interest were generated for each subject using the built-in brain parcellation work-flow within PMOD’s PNEURO Tool (25). Region generation was based off the AAL-VOIs atlas (26, 27). Regions of interest included the amygdala, hippocampus, midbrain, cerebellum, striatum (ventral striatum, caudate and putamen) and prefrontal cortex (specifically the dorsolateral, orbital, medial, and anterior cingulate) subdivisions. These regions that have been a focus of N/OFQ and NOP basic investigations are the same regions we examined in a prior study in alcoholics (24). All regions generated by the brain parcellation tool were visually inspected and adjusted as deemed necessary by an image analyst trained in manual region drawing. Regional volumes and time activity curves were also generated in PMOD. Derivation of [11C]NOP-1A volume of distribution expressed relative to total plasma concentration (VT) in the regions of interest were performed using a two-tissue compartment kinetic analysis using the arterial input function implemented in MATLAB (21, 22, 28). VT, which includes both the receptor-bound specific and non-specific binding was used as the outcome measure (as opposed to the binding potential relative to non-displaceable uptake, BPND) because there is no brain region in the brain that is devoid of specific binding to NOP (23).
Relapse monitoring protocol for CUD subjects
In order to document relapse, CUD subjects were enrolled in a 12-week follow-up protocol after the [11C]NOP-1A PET scan. This follow-up protocol was modeled after the contingency management protocol used in a previous [11C]raclopride-amphetamine PET study (29, 30). Briefly, CUD met with the research team three times per week for urine drug screens (UDS) and earned voucher points on an escalating schedule for each negative UDS. Subjects earned bonus points for every three-consecutive cocaine-free UDS (one week of abstinence). Missed appointments reset the voucher points to a value that was lower by 10 points. Subjects had the potential to earn a maximum of $1197.00 for providing cocaine-free UDS on all of the scheduled monitoring visits. The money earned was disbursed to them on a weekly basis via a debit card. Subjects were terminated from the follow-up protocol: for testing positive for cocaine three times (i.e., were allowed only three distinct relapses); for missing three consecutive scheduled appointments (i.e., were lost to follow-up for a week); and for initiating any medications that influence N/OFQ (for example, buprenorphine, etc.). Subjects were also monitored once a week for depression and suicidal ideation, and de-briefed on their week’s progress concerning abstinence.
Statistical analysis
All statistical analyses were conducted using IBM SPSS v.25. Normality of data was examined using Kolmgorov-Smirnov and Shapiro-Wilkes tests. Group demographic and baseline scan parameter (such as injected dose, mass, plasma clearance) comparisons were performed with unpaired t-tests. The primary analyses examined overall group differences in [11C]NOP-1A VT with a linear mixed model analysis (LMM) performed with regions of interest as a repeated measure and diagnostic group (CUD vs. HC) as the fixed factor. The effect of sex and smoking status on VT and diagnostic group were subsequently examined in a second level LMM analyses by including them as fixed factors in the model. Post-hoc unpaired t-tests in the individual regions of interest were also conducted. A two-tailed probability value of p ≤ 0.05 was selected as the significance level for the primary analyses (LMM that included all the regions of interest). A Benjamini-Hochberg false discovery rate (FDR) correction with α = 0.05 was applied to correct for multiple comparisons in the individual regions of interest in the post-hoc analyses.
Group comparison of regional VT in CUD who abstained, relapsed and drop-out were evaluated using a LMM (as described previously). Studies in CUD have previously shown that early treatment response and motivation as reflected by clinic attendance in the first 2–3 weeks predicts sustained abstinence in CUD subjects’ during 12-weeks of contingency management treatment (29, 31). Based on this we also used a LMM to compare VT in CUD subjects who relapsed and/or dropped-out within the first 2 weeks (out of a 12-week follow-up) to that in those who did not relapse and/or continued in the protocol past 2 weeks. The amount of voucher money earned, which reflects abstinence was used as a secondary outcome measure in a Pearson product moment correlation (only regions that survived FDR correction were used for all correlational analyses). Lastly, a Cox proportional hazards model was used to determine the association between time to relapse and regional VT in the CUD group. Time to relapse was defined as the time between the date of [11C]NOP-1A PET scan and the date of the first relapse to cocaine (positive cocaine UDS) over the 12-week follow-up period. Data were censored if the subjects did not experience relapse by the end of the follow-up period, including those lost to follow-up. Models were run with and without adjustment for potential confounding factors (such as sex, PSS, CCQ). The relationship between regional VT and other clinical measures of stress (PSS), anxiety (HAM-A), craving (CCQ) and severity (cocaine use duration in years, frequency in days and amount spent in dollars) of cocaine abuse in CUD were assessed by Pearson product moment correlation. A two-tailed probability value of p ≤ 0.05 was selected as the significance level for all the analyses that involved clinical measures.
RESULTS
Twenty-four individuals with CUD were matched with 25 HC on age, sex, ethnicity and nicotine status as closely as possible. Table 1 lists the demographics variables and clinical characteristics of the study sample.
Table 1.
Demographic and clinical characteristics of cocaine use disorder and healthy control subjects
| Cocaine use disorder | Healthy controls | |
|---|---|---|
| n = 24 | n = 25 | |
| Age | 40 ± 9 | 37 ± 9 |
| Sex | ||
| Females | 10 | 11 |
| Males | 14 | 14 |
| Ethnicity | ||
| African American | 10 | 5 |
| Caucasian | 12 | 18 |
| Asian | 0 | 1 |
| Hispanic | 1 | 0 |
| More than one race | 1 | 1 |
| Barratt Simplified Measure of Social Status | ||
| Education | 14 ± 3 | 15 ± 4 |
| Occupation | 21 ± 10 | 27 ± 4* |
| Total | 33 ± 13 | 42 ± 12* |
| Tobacco use | 16 | 13 |
| > 10 cigarettes per day | 7 | 4 |
| Cannabis use (recreational) | 4 | - |
| Hamilton Anxiety Rating Scale (range 0 to 56) | 8.5 ± 7.5 | 2.1 ± 3.7* |
| Perceived Stress Scale (range 0 to 40) | 15.2 ± 7.8 | 5.5 ± 5.3* |
| Cocaine Craving Questionnaire (range 0 to 70) | 26 ± 15 | |
| Age of first use of cocaine (years) | 23 ± 8 | |
| Duration of cocaine use (years) | 15 ± 9 | |
| Cocaine use frequency per week (days) | 2 ± 2 | |
| Amount spent on cocaine per week (dollars) | 126 ± 64 | |
| 12-week follow-up protocol to monitor for relapse following PET scan | ||
| Final Outcome | ||
| Abstinent | 6 | |
| Relapsed (cocaine +) | 9 | |
| Drop-out | 9 | |
| Amount of money earned in vouchers (dollars) | 415 ± 476 | |
| Time to an event, i.e., first relapse or drop-out or completion (days) | 35 ± 34 | |
| Time to first relapse (days) | 13 ± 14 | |
| Duration in follow-up protocol (days) | 43 ± 35 |
p < 0.05, unpaired t-tests for numerical variables and Chi-square test for categorical variables.
[11C]NOP-1A scan parameters
Table 2 shows no significant group differences between CUD and HC in any of the baseline scan parameters. Consistent with that reported previously, [11C]NOP-1A plasma free fraction (fP) measurements were not reliable because the tracer showed relatively high retention to the filter in the saline buffer solution condition (24). No significant group differences in the regions of interest volume determined from MRI scans were noted (data not shown).
Table 2.
[11C]NOP-1A scan parameters
| Scan Parameter | Cocaine use disorder n = 24 |
Healthy controls n=25 |
|---|---|---|
| Injected dose (mCi) | 12.4 ± 0.7 | 12.3 ± 1.0 |
| Specific Activity (Ci/mmoles) | 2169 ± 975 | 2230 ± 558 |
| Injected mass (μg) | 2.8 ± 1.0 | 2.5 ± 0.6 |
| Plasma free fraction (fP, %) † | 14.5% ± 3.2% | 13.2% ± 2.3% |
| Plasma free fraction buffer (fP, %) † | 71.0% ± 15.7% | 76.1% 11.5% |
| Clearance (L/h) | 151.8 ± 38.9 | 155.1 ± 38.1 |
Values are mean ± SD in cocaine use disorder and healthy control subjects
SD - Standard deviation
Data was only available from n=23 CUD and 22 HC due to problems with sample processing.
In vivo binding of [11C]NOP-1A (VT)
Regional [11C]NOP-1A VT was significantly higher in CUD compared to HC (LMM, effect of diagnosis, F (1, 47) = 4.39, p = 0.042; effect of region, F (10, 470) = 520.20, p < 0.001; region * diagnosis interaction, F (10, 470) = 1.45, p = 0.156).
[11C]NOP-1A VT in the regions of interest were on average 7 to 12% lower in females compared to males (Figure 1). The inclusion of sex as a factor in the LMM did not alter the significance of the above described result (effect of diagnosis, F (1, 45) = 4.65, p = 0.037; effect of sex, F (1, 45) = 4.45, p = 0.041; sex * diagnosis interaction, F (1, 45) = 0.25, p = 0.622). However, the effect of diagnosis fell short of significance when tobacco use status was included as an additional factor in the LMM (effect of diagnosis, F=3.88, df=1, 43, p=0.055; effect of smoking, F=0.03, df=1, 43, p=0.86; smoking * diagnosis interaction, F=0.11, df=1, 43, p=0.74). Excluding the four CUD subjects who tested positive for cannabis during the outpatient monitoring prior to PET did not change the results (data from n = 20 CUD and n = 25 HC; LMM, effect of diagnosis, F (1, 43) = 5.65, p =0.022; effect of region, F (10, 430) = 500.02, p < 0.001; region * diagnosis interaction, F (10, 430 = 1.97, p = 0.035).
Figure 1.
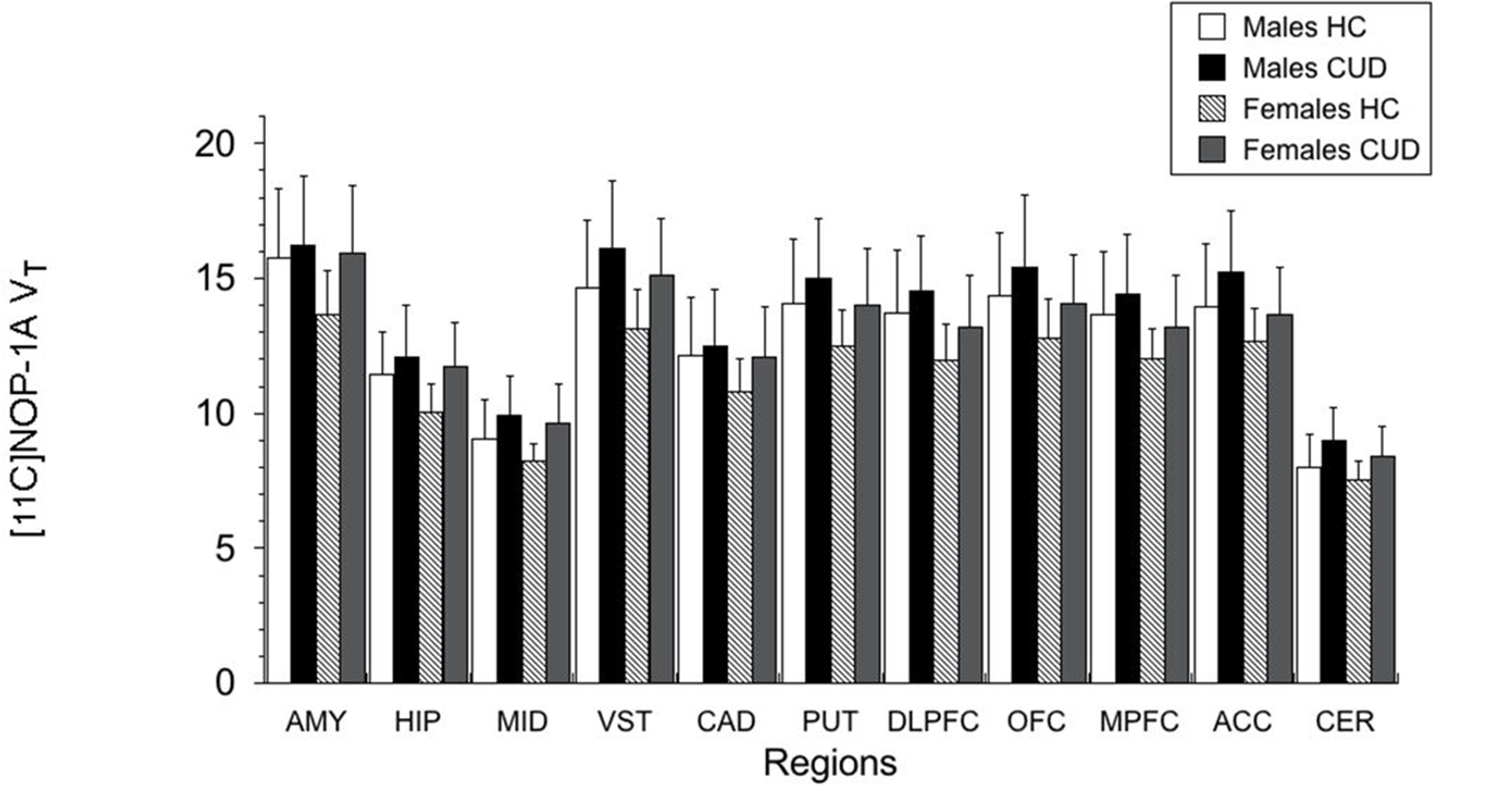
Female subjects when compared to male subjects had significantly lower VT values in the regions of interest. Sex-based differences were not present with respect to the increase in VT in CUD relative to HC (sex * diagnosis interaction, p = 0.62). AMY: amygdala, HIP: hippocampus, MID: midbrain, VST: ventral striatum, CAD: caudate, PUT: putamen, DLPFC: dorsolateral prefrontal cortex, OFC: orbital frontal cortex, MPFC: medial prefrontal cortex, ACC: anterior cingulate cortex, CER: cerebellum.
Unpaired t-tests conducted at the level of the individual regions of interest were significant in four of the eleven regions of interest (see Table 3). The comparisons in the ventral striatum and cerebellum survived the FDR correction.
Table 3.
[11C]NOP-1A volume of distribution expressed relative to total plasma concentration (VT) in regions of interest
| Cocaine use disorder | Healthy controls | Uncorrected p-values | |
|---|---|---|---|
| Regions of interest | n = 24 | n = 25 | |
| Amygdala | 16.10 ± 2.48 | 14.86 ± 2.40 | 0.084 |
| Hippocampus | 11.93 ± 1.76 | 10.85 ± 1.53 | 0.027 |
| Midbrain | 9.81 ± 1.40 | 8.72 ± 1.22 | 0.0056 |
| Ventral Striatum | 15.70 ± 2.35 | 13.99 ± 2.21 | 0.0118* |
| Caudate | 12.32 ± 1.99 | 11.56 ± 1.90 | 0.177 |
| Putamen | 14.60 ± 2.18 | 13.37 ± 2.13 | 0.051 |
| Dorsolateral Prefrontal Cortex | 13.97 ± 2.09 | 12.98 ± 2.11 | 0.105 |
| Orbital Frontal Cortex | 14.85 ± 2.40 | 13.68 ± 2.15 | 0.078 |
| Medial Prefrontal Cortex | 13.93 ± 2.14 | 12.96 ± 2.05 | 0.115 |
| Anterior Cingulate Cortex | 14.57 ± 2.18 | 13.39 ± 1.99 | 0.053 |
| Cerebellum | 8.73 ± 1.21 | 7.81 ± 1.03 | 0.0062* |
Values are mean ± SD in cocaine use disorder and healthy control subjects
SD - Standard deviation
Significant following multiple hypotheses correction with FDR
Relationship between VT and relapse to cocaine during follow up
There were no significant differences in VT between CUD who abstained vs. relapsed vs. drop-out during the follow-up period (LMM, effect of final outcome status, F (2, 21) = 0.13, p = 0.88; effect of region, F (10, 210) = 230.65, p < 0.001; region * diagnosis interaction, F (20, 210) = 0.49, p = 0.97, see Figure 2). Stratification of the CUD subjects based on whether they relapsed and/or dropped-out within the first two weeks (n = 9 CUD < 2-weeks; n = 15 > 2 weeks) also revealed no group differences in [11C]NOP-1A VT (LMM, effect of duration, F (1, 22) = 1.24, p = 0.28; effect of region, F (10, 220) = 223.81, p < 0.001; time * region interaction, F (10, 220) = 0.65, p = 0.77. There were also no significant correlations between VT and the amount of voucher money earned during follow-up (midbrain, r = 0.19, p = 0.36; ventral striatum, r = 0.18, p = 0.41; and cerebellum r = 0.18, p = 0.40). Lastly, VT in the regions of interest were also not predictive of time to relapse to cocaine (midbrain, Exp (B) = 1.0, p = 0.99; ventral striatum, Exp (B) = 0.97, p = 0.78; and cerebellum, Exp (B) = 1.02, p = 0.95). These results were unchanged irrespective of whether the models were adjusted for potential confounding factors such as sex, PSS and CCQ (data not shown).
Figure 2.
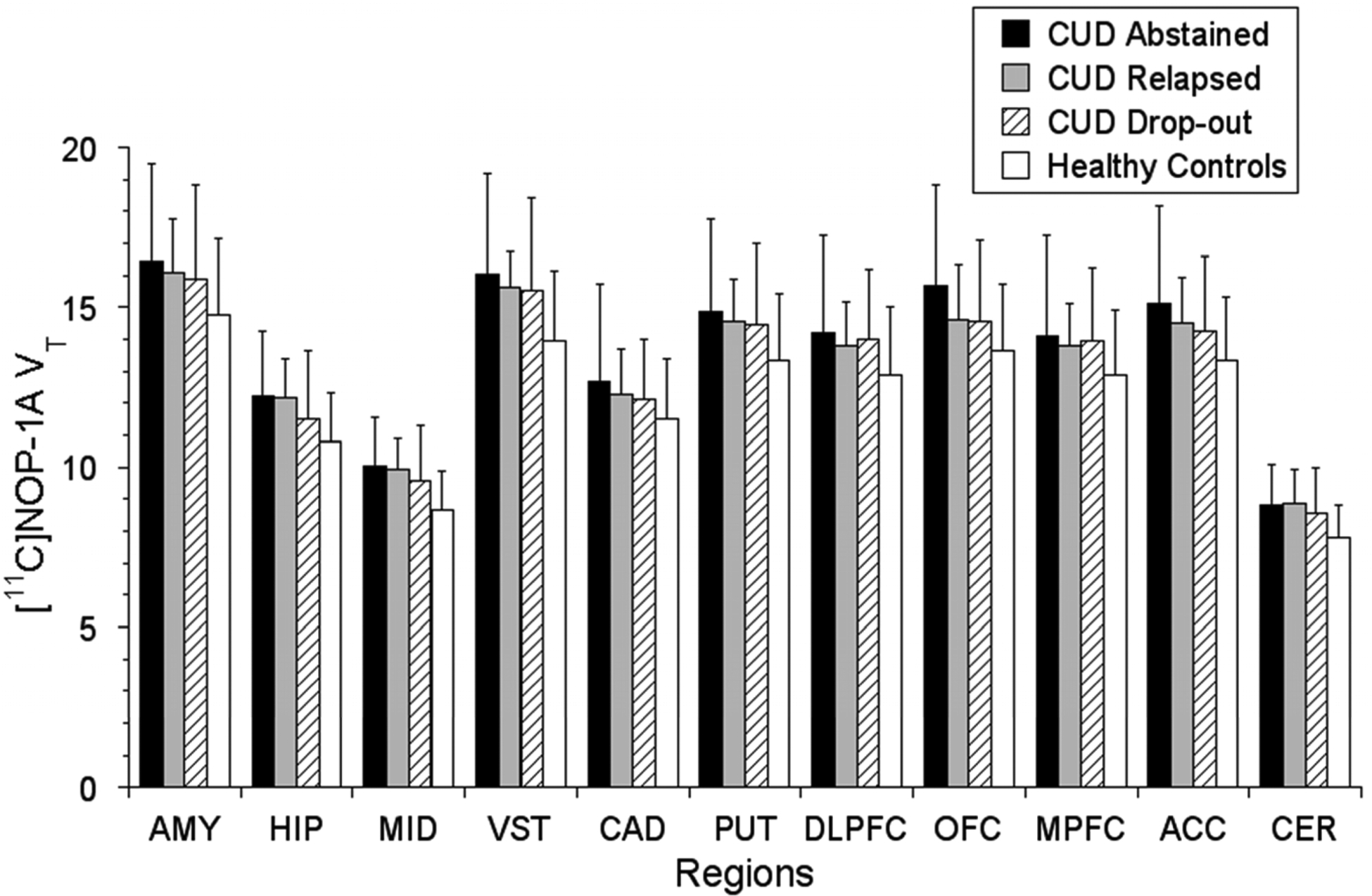
CUD subjects who abstained (n = 6), relapsed (n = 9) and drop-out (n = 9) during the 12-week follow-up period demonstrated no significant differences in VT. Also, included in the figure for a visual contrast is baseline VT data from healthy controls (n = 25). Regions shown are AMY: amygdala, HIP: hippocampus, MID: midbrain, VST: ventral striatum, CAD: caudate, PUT: putamen, DLPFC: dorsolateral prefrontal cortex, OFC: orbital frontal cortex, MPFC: medial prefrontal cortex, ACC: anterior cingulate cortex, CER: cerebellum.
Higher levels of perceived stress on PSS were predictive of less time to relapse (Exp (B) = 1.08, p = 0.05). No such relationship was observed between cocaine cravings (CCQ) and time to relapse (Exp (B) = 1.00, p = 0.88).
Relationships between VT and clinical measures
No significant relationships were observed between regional [11C]NOP-1A VT and any of the other clinical measures (including cocaine use frequency, duration and amount of money spent, perceived stress, anxiety and cocaine craving) in CUD.
Relationship between VT and age
The negative relationship between age and VT fell short of significance in the medial prefrontal (r = −0.39, p = 0.06) and dorsolateral prefrontal cortex (r = −0.36, p = 0.08), but not other regions (data not shown, p > 0.1) in healthy controls. --REMOVE LINE ABOUT MULTIPLE CORRECTION-- No such relationship between age and VT were noted in any of the regions in CUD.
DISCUSSION
In this [11C]NOP-1A PET study, we found that: (1) cocaine abuse in humans is associated with a generalized ~10% increase in binding to NOP in brain regions that mediate reward and stress behaviors (the four regions that were significant included the midbrain, ventral striatum hippocampus and cerebellum); (2) females have less NOP binding compared to males; and (3) increased stress during cocaine withdrawal predicts an early relapse. An important negative result is the lack of a relationship between VT and relapse.
Increased [11C]NOP-1A binding in cocaine users suggests a compensatory upregulation of NOP in response to decreased N/OFQ in the brain. This is supported by studies in chronic cocaine rodents that have shown decreases in N/OFQ in the nucleus accumbens, caudate-putamen and substantia nigra/ventral tegmental area (15, 16). However, NOP’s regulatory response to decreased N/OFQ in these rodents has been shown to be variable at the level of the striatum, i.e., increased in the nucleus accumbens (ventral striatum) and decreased in the caudate-putamen(16). The PET data in CUD, which showed an increase in NOP in the ventral striatum and putamen and no change in the caudate, is only partially consistent with these rodent data. The lack of an increase in NOP in the caudate, which is observed in both the humans and rodent studies, is still noteworthy because it might relate to the fact that the caudate is the only striatal subdivision in which dopamine transmission is not blunted in CUD (32). Studies examining the relationship between NOP VT and amphetamine-induced dopamine release in CUD might clarify this relationship. Behavioral paradigms that mimic chronic stress in rodents consistently show upregulation of NOP in the limbic-brain regions (33–36). Thus, increased NOP in CUD might also reflect an adaptive response to balance the effects of CRF, which increases stress and promotes relapse. Supportive of this is a study in which an upregulation of NOP in the bed nucleus of stria terminalis and the amygdala were noted following increases in CRF (33, 37). Similar interactions between NOP and other stress-promoting neuropeptides such as dynorphin have also been described in chronic cocaine rodents (16). Future imaging studies with [11C]NOP-1A investigating NOP-CRF and NOP-dynorphin interactions in CUD should be considered because they might clarify the interplay between stress and anti-stress neuropeptides in negative reinforcement and relapse. N/OFQ stimulates NOP to inhibit calcium channels and activate potassium channels (38). This allows for N/OFQ to inhibit the release of multiple neurotransmitters including dopamine, serotonin, acetylcholine, glutamate, and GABA (39). Increased N/OFQ has been shown to inhibit cocaine-induced dopamine release in the nucleus accumbens (12–14). This mechanism has therapeutic potential because it can decrease the cocaine reward experience. It is likely that N/OFQ directly modulates midbrain dopamine neurons to impact cocaine-induced dopamine release because 50–90% of tyrosine hydroxylase positive cells (TH +) in the ventral tegmental area and substantia nigra express NOP (40, 41). However, an effect for pre-synaptic NOP on dopamine terminals cannot be ruled out because administration of N/OFQ into the nucleus accumbens also blocks cocaine-induced dopamine release (13). Increased N/OFQ has also been shown to decrease glutamate transmission in the reward- and stress-mediating brain regions such as the cortex, midbrain, amygdala and cerebellum (42–44). This mechanism also has therapeutic potential because increased glutamate in CUD have been linked to relapse and reinstatement (45). These mechanistic investigations and this PET study support clinical trials with a NOP agonist to increase N/OFQ transmission in CUD. The inability to link NOP VT with relapse in this study cannot be viewed as less supportive of this conceptual model because it is not possible to quantitate endogenous N/OFQ levels from a baseline [11C]NOP-1A scan. [11C]NOP-1A imaging paradigms that measure the regulatory response of NOP to stress/CRF might be more successful in predicting relapse in CUD.
Sex-based differences have been reported for stress-mediating neuropeptides and their receptors including CRF and kappa-opioid receptors (46, 47). To our knowledge, decreased NOP in females relative to males is the first report of sex-based differences in an anti-stress system in the brain. This unreported observation was also present as a trend level finding in a separate set of fifteen healthy controls (5 females and 10 males; eleven regions of interest; LMM, effect of gender on VT, p = 0.055, region, p < 0.001, gender * region interaction, p = 0.012) in our previous [11C]NOP-1A alcohol study (19). Studies to understand the clinical relevance of decreased NOP in females are necessary because it may explain previously described behavioral and physiological phenomena, such as females reporting greater subjective levels of stress, showing increased sensitivity to CRF, etc. It may also provide clues as to the reasons why females are at higher risk for certain chronic stress disorders such as depression, anxiety, chronic pain, etc. Sex had no effect on the increased NOP in CUD, i.e., there was no sex * diagnosis interaction. However, the effect size of the difference in [11C]NOP-1A VT in CUD Vs. controls was much larger in females compared to males (Table 4). This is interesting as chronic cocaine rodent and human studies consistently report more negative affect, withdrawal symptoms and stress-induced relapse/reinstatement in females compared to males (48). It is tempting to speculate greater decreases in resilience-mediating N/OFQ and/or increases in stress-mediating CRF (both of which have been shown to upregulate NOP receptors in basic investigations) as reasons for more unpleasant symptoms and subsequent relapse in chronic cocaine abusing females. Future [11C]NOP-1A PET studies should focus on these sex differences, and also control for the fluctuations in sex hormones associated with menstrual cycle and hormonal contraceptive use.
Table 4.
Effect size of the difference in [11C]NOP-1A VT in cocaine use disorder vs. healthy controls by sex
| Regions of interest | Males (d) |
Female (d) |
Effect size ratio (female/male) |
|---|---|---|---|
| Amygdala | 0.19 | 1.10 | 5.8 |
| Hippocampus | 0.37 | 1.27 | 3.4 |
| Midbrain | 0.62 | 1.33 | 2.1 |
| Ventral Striatum | 0.62 | 1.12 | 1.8 |
| Caudate | 0.16 | 0.85 | 5.3 |
| Putamen | 0.44 | 0.89 | 2.0 |
| Dorsolateral Prefrontal Cortex | 0.38 | 0.73 | 1.9 |
| Orbital Frontal Cortex | 0.45 | 0.77 | 1.7 |
| Medial Prefrontal Cortex | 0.35 | 0.77 | 2.2 |
| Anterior Cingulate Cortex | 0.57 | 0.70 | 1.2 |
| Cerebellum | 0.81 | 0.96 | 1.2 |
d is the Cohen’s effect size maximum likelihood estimation of the difference between cocaine use disorder and healthy controls
The role of negative reinforcement in promoting stress/anxiety, and leading to relapse to drugs/alcohol is well established in rodent models of addiction (3). Consistent with this model, we found that increased stress during cocaine withdrawal is predictive of less time to relapse. However, we found no relationship between perceived stress/anxiety and the anti-stress/resilience conferring NOP (VT). Subjects were scanned after they abstained from cocaine for a minimum of ten days, which may have limited the ability to detect such a relationship. PET studies focusing on the neurochemistry of withdrawal are warranted because the behavioral symptoms experienced during this period predicts relapse.
We were unable to exclude differences in [11C]NOP-1A non-specific binding (VND) and plasma free fraction (fP) as contributors to VT. Reassuringly, prior [11C]NOP-1A blocking studies in humans suggest 50–75% of VT represents specific binding to NOP (18). In summary, we demonstrated a significant increase in NOP in CUD. The data showed a trend towards regionally-selective increases in NOP that were convincingly evidenced in the midbrain, ventral striatum and cerebellum. Increased NOP in CUD may be indicative of an adaptive response to decreased N/OFQ, or increased CRF, or both. The clinical development of NOP agonists to enhance N/OFQ transmission should be explored as a treatment for CUD.
ACKNOWLEDGEMENTS.
The project described above was funded by Award Numbers R01DA026472 from the National Institute on Drug Abuse (NIDA). Recruitment of subjects were in part supported by a research registry (Pitt+Me), which is funded by the National Institutes of Health (NIH) Clinical and Translational Science Award (CTSA) program, Award Number UL1 TR001857.
The content is solely the responsibility of the authors and does not necessarily represent the official views of NIDA or the NIH.
Footnotes
DISCLOSURES
None
REFERENCES
- 1.Reinscheid RK, Nothacker HP, Bourson A, Ardati A, Henningsen RA, Bunzow JR, Grandy DK, Langen H, Monsma FJ Jr., Civelli O. Orphanin FQ: a neuropeptide that activates an opioidlike G protein-coupled receptor. Science. 1995;270:792–794. [DOI] [PubMed] [Google Scholar]
- 2.Meunier JC, Mollereau C, Toll L, Suaudeau C, Moisand C, Alvinerie P, Butour JL, Guillemot JC, Ferrara P, Monsarrat B, et al. Isolation and structure of the endogenous agonist of opioid receptor-like ORL1 receptor. Nature. 1995;377:532–535. [DOI] [PubMed] [Google Scholar]
- 3.Koob GF. A role for brain stress systems in addiction. Neuron. 2008;59:11–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mogil JS, Grisel JE, Reinscheid RK, Civelli O, Belknap JK, Grandy DK. Orphanin FQ is a functional anti-opioid peptide. Neuroscience. 1996;75:333–337. [DOI] [PubMed] [Google Scholar]
- 5.Jenck F, Moreau JL, Martin JR, Kilpatrick GJ, Reinscheid RK, Monsma FJ Jr., Nothacker HP, Civelli O. Orphanin FQ acts as an anxiolytic to attenuate behavioral responses to stress. Proc Natl Acad Sci U S A. 1997;94:14854–14858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ciccocioppo R, Biondini M, Antonelli L, Wichmann J, Jenck F, Massi M. Reversal of stress- and CRF-induced anorexia in rats by the synthetic nociceptin/orphanin FQ receptor agonist, Ro 64–6198. Psychopharmacology (Berl). 2002;161:113–119. [DOI] [PubMed] [Google Scholar]
- 7.Koster A, Montkowski A, Schulz S, Stube EM, Knaudt K, Jenck F, Moreau JL, Nothacker HP, Civelli O, Reinscheid RK. Targeted disruption of the orphanin FQ/nociceptin gene increases stress susceptibility and impairs stress adaptation in mice. Proc Natl Acad Sci U S A. 1999;96:10444–10449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sakoori K, Murphy NP. Central administration of nociceptin/orphanin FQ blocks the acquisition of conditioned place preference to morphine and cocaine, but not conditioned place aversion to naloxone in mice. Psychopharmacology (Berl). 2004;172:129–136. [DOI] [PubMed] [Google Scholar]
- 9.Bebawy D, Marquez P, Samboul S, Parikh D, Hamid A, Lutfy K. Orphanin FQ/nociceptin not only blocks but also reverses behavioral adaptive changes induced by repeated cocaine in mice. Biol Psychiatry. 2010;68:223–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Marquez P, Nguyen AT, Hamid A, Lutfy K. The endogenous OFQ/N/ORL-1 receptor system regulates the rewarding effects of acute cocaine. Neuropharmacology. 2008;54:564–568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lutfy K, Khaliq I, Carroll FI, Maidment NT. Orphanin FQ/nociceptin blocks cocaine-induced behavioral sensitization in rats. Psychopharmacology (Berl). 2002;164:168–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lutfy K, Do T, Maidment NT. Orphanin FQ/nociceptin attenuates motor stimulation and changes in nucleus accumbens extracellular dopamine induced by cocaine in rats. Psychopharmacology (Berl). 2001;154:1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vazquez-DeRose J, Stauber G, Khroyan TV, Xie XS, Zaveri NT, Toll L. Retrodialysis of N/OFQ into the nucleus accumbens shell blocks cocaine-induced increases in extracellular dopamine and locomotor activity. Eur J Pharmacol. 2013;699:200–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Murphy NP, Ly HT, Maidment NT. Intracerebroventricular orphanin FQ/nociceptin suppresses dopamine release in the nucleus accumbens of anaesthetized rats. Neuroscience. 1996;75:1–4. [DOI] [PubMed] [Google Scholar]
- 15.Romualdi P, Di Benedetto M, D’Addario C, Collins SL, Wade D, Candeletti S, Izenwasser S. Chronic cocaine produces decreases in N/OFQ peptide levels in select rat brain regions. J Mol Neurosci. 2007;31:159–164. [DOI] [PubMed] [Google Scholar]
- 16.Caputi FF, Di Benedetto M, Carretta D, Bastias del Carmen Candia S, D’Addario C, Cavina C, Candeletti S, Romualdi P. Dynorphin/KOP and nociceptin/NOP gene expression and epigenetic changes by cocaine in rat striatum and nucleus accumbens. Prog Neuropsychopharmacol Biol Psychiatry. 2014;49:36–46. [DOI] [PubMed] [Google Scholar]
- 17.Pike VW, Rash KS, Chen Z, Pedregal C, Statnick MA, Kimura Y, Hong J, Zoghbi SS, Fujita M, Toledo MA, Diaz N, Gackenheimer SL, Tauscher JT, Barth VN, Innis RB. Synthesis and evaluation of radioligands for imaging brain nociceptin/orphanin FQ peptide (NOP) receptors with positron emission tomography. J Med Chem. 2011;54:2687–2700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Raddad E, Chappell A, Meyer J, Wilson A, Ruegg CE, Tauscher J, Statnick MA, Barth V, Zhang X, Verfaille SJ. Occupancy of Nociceptin/Orphanin FQ Peptide Receptors by the Antagonist LY2940094 in Rats and Healthy Human Subjects. Drug Metab Dispos. 2016;44:1536–1542. [DOI] [PubMed] [Google Scholar]
- 19.Narendran R, Ciccocioppo R, Lopresti B, Paris J, Himes ML, Mason NS. Nociceptin Receptors in Alcohol Use Disorders: A Positron Emission Tomography Study Using [(11)C]NOP-1A. Biol Psychiatry. 2018;84:708–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.First M, Williams JBW, Karg RS, Spitzer RL: Structured Clinical Interview for DSM-5—Research Version (SCID-5 for DSM-5, Research Version; SCID-5-RV). Arlington, VA, American Psychiatric Association; 2015; 2015. [Google Scholar]
- 21.Lohith TG, Zoghbi SS, Morse CL, Araneta MD, Barth VN, Goebl NA, Tauscher JT, Pike VW, Innis RB, Fujita M. Retest imaging of [11C]NOP-1A binding to nociceptin/orphanin FQ peptide (NOP) receptors in the brain of healthy humans. Neuroimage. 2014;87:89–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lohith TG, Zoghbi SS, Morse CL, Araneta MF, Barth VN, Goebl NA, Tauscher JT, Pike VW, Innis RB, Fujita M. Brain and whole-body imaging of nociceptin/orphanin FQ peptide receptor in humans using the PET ligand 11C-NOP-1A. J Nucl Med. 2012;53:385–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kimura Y, Fujita M, Hong J, Lohith TG, Gladding RL, Zoghbi SS, Tauscher JA, Goebl N, Rash KS, Chen Z, Pedregal C, Barth VN, Pike VW, Innis RB. Brain and whole-body imaging in rhesus monkeys of 11C-NOP-1A, a promising PET radioligand for nociceptin/orphanin FQ peptide receptors. J Nucl Med. 2011;52:1638–1645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Narendran R, Ciccocioppo R, Lopresti B, Paris J, Himes ML, Mason NS. Nociceptin Receptors in Alcohol Use Disorders: A Positron Emission Tomography Study Using [11C]NOP-1A. Biol Psychiatry. 2017;In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Douaud GGV, Ribeiro MJ, Lethimonnier F, Maroy R, Verny C, Krystkowiak P, Damier P, Bachoud-Levi AC, Hantraye P et al. Distribution of grey matter atrophy in Huntington’s disease patients: a combined ROI-based and voxel-based morphometric study. Neuroimage. 2006;32:1562–1575. [DOI] [PubMed] [Google Scholar]
- 26.Tzourio-Mazoyer BL N, Papathanassiou D, Crivello F, Étard O, Delcroix N, Mazoyer B, and Joliot M. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage. 2002;15:273–289. [DOI] [PubMed] [Google Scholar]
- 27.Collins DZ AP ; Kollokian V ; Sled JG ; Kabani NJ ; Holmes CJ ; Evans AC. Design and construction of a realistic digital brain phantom. IEEE Transactions on Medical Imaging. 1998;17:463–468. [DOI] [PubMed] [Google Scholar]
- 28.Innis RB, Cunningham VJ, Delforge J, Fujita M, Gjedde A, Gunn RN, Holden J, Houle S, Huang SC, Ichise M, Iida H, Ito H, Kimura Y, Koeppe RA, Knudsen GM, Knuuti J, Lammertsma AA, Laruelle M, Logan J, Maguire RP, Mintun MA, Morris ED, Parsey R, Price JC, Slifstein M, Sossi V, Suhara T, Votaw JR, Wong DF, Carson RE. Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab. 2007;27:1533–1539. [DOI] [PubMed] [Google Scholar]
- 29.Martinez D, Carpenter KM, Liu F, Slifstein M, Broft A, Friedman AC, Kumar D, Van Heertum R, Kleber HD, Nunes E. Imaging dopamine transmission in cocaine dependence: link between neurochemistry and response to treatment. The American journal of psychiatry. 2011;168:634–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Higgins ST, Wong CJ, Badger GJ, Ogden DE, Dantona RL. Contingent reinforcement increases cocaine abstinence during outpatient treatment and 1 year of follow-up. J Consult Clin Psychol. 2000;68:64–72. [DOI] [PubMed] [Google Scholar]
- 31.Luo SX, Martinez D, Carpenter KM, Slifstein M, Nunes EV. Multimodal predictive modeling of individual treatment outcome in cocaine dependence with combined neuroimaging and behavioral predictors. Drug Alcohol Depend. 2014;143:29–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Martinez D, Narendran R, Foltin R, S M, Hwang D-R, Broft A, Huang Y, Cooper T, Fischman M, Kleber H, Laruelle M. Amphetamine-induced dopamine release is markedly blunted in cocaine dependent subjects and predictive of the choice to self administer cocaine. Am J Psychiatry. 2007;164:622–629. [DOI] [PubMed] [Google Scholar]
- 33.Ciccocioppo R, de Guglielmo G, Hansson AC, Ubaldi M, Kallupi M, Cruz MT, Oleata CS, Heilig M, Roberto M. Restraint stress alters nociceptin/orphanin FQ and CRF systems in the rat central amygdala: significance for anxiety-like behaviors. J Neurosci. 2014;34:363–372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Green MK, Devine DP. Nociceptin/orphanin FQ and NOP receptor gene regulation after acute or repeated social defeat stress. Neuropeptides. 2009;43:507–514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Reiss D, Wolter-Sutter A, Krezel W, Ouagazzal AM. Effects of social crowding on emotionality and expression of hippocampal nociceptin/orphanin FQ system transcripts in mice. Behav Brain Res. 2007;184:167–173. [DOI] [PubMed] [Google Scholar]
- 36.Zhang Y, Simpson-Durand CD, Standifer KM. Nociceptin/orphanin FQ peptide receptor antagonist JTC-801 reverses pain and anxiety symptoms in a rat model of post-traumatic stress disorder. Br J Pharmacol. 2015;172:571–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rodi D, Zucchini S, Simonato M, Cifani C, Massi M, Polidori C. Functional antagonism between nociceptin/orphanin FQ (N/OFQ) and corticotropin-releasing factor (CRF) in the rat brain: evidence for involvement of the bed nucleus of the stria terminalis. Psychopharmacology (Berl). 2008;196:523–531. [DOI] [PubMed] [Google Scholar]
- 38.Toll L, Bruchas MR, Calo G, Cox BM, Zaveri NT. Nociceptin/Orphanin FQ Receptor Structure, Signaling, Ligands, Functions, and Interactions with Opioid Systems. Pharmacol Rev. 2016;68:419–457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Schlicker E, Morari M. Nociceptin/orphanin FQ and neurotransmitter release in the central nervous system. Peptides. 2000;21:1023–1029. [DOI] [PubMed] [Google Scholar]
- 40.Maidment NT, Chen Y, Tan AM, Murphy NP, Leslie FM. Rat ventral midbrain dopamine neurons express the orphanin FQ/nociceptin receptor ORL-1. Neuroreport. 2002;13:1137–1140. [DOI] [PubMed] [Google Scholar]
- 41.Norton CS, Neal CR, Kumar S, Akil H, Watson SJ. Nociceptin/orphanin FQ and opioid receptor-like receptor mRNA expression in dopamine systems. J Comp Neurol. 2002;444:358–368. [DOI] [PubMed] [Google Scholar]
- 42.Kallupi M, Varodayan FP, Oleata CS, Correia D, Luu G, Roberto M. Nociceptin/Orphanin FQ Decreases Glutamate Transmission and Blocks Ethanol-Induced Effects in the Central Amygdala of Naive and Ethanol-Dependent Rats. Neuropsychopharmacology. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nicol B, Lambert DG, Rowbotham DJ, Okuda-Ashitaka E, Ito S, Smart D, McKnight AT. Nocistatin reverses nociceptin inhibition of glutamate release from rat brain slices. Eur J Pharmacol. 1998;356:R1–3. [DOI] [PubMed] [Google Scholar]
- 44.Nicol B, Lambert DG, Rowbotham DJ, Smart D, McKnight AT. Nociceptin induced inhibition of K+ evoked glutamate release from rat cerebrocortical slices. Br J Pharmacol. 1996;119:1081–1083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Spencer S, Scofield M, Kalivas PW. The good and bad news about glutamate in drug addiction. J Psychopharmacol. 2016;30:1095–1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Vijay A, Wang S, Worhunsky P, Zheng MQ, Nabulsi N, Ropchan J, Krishnan-Sarin S, Huang Y, Morris ED. PET imaging reveals sex differences in kappa opioid receptor availability in humans, in vivo. Am J Nucl Med Mol Imaging. 2016;6:205–214. [PMC free article] [PubMed] [Google Scholar]
- 47.Veldhuis JD, Sharma A, Roelfsema F. Age-dependent and gender-dependent regulation of hypothalamic-adrenocorticotropic-adrenal axis. Endocrinol Metab Clin North Am. 2013;42:201–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Becker JB, Koob GF. Sex Differences in Animal Models: Focus on Addiction. Pharmacol Rev. 2016;68:242–263. [DOI] [PMC free article] [PubMed] [Google Scholar]