

Abstract
Gliomas are the most common neoplasm of the human central nervous system. Glioblastoma multiforme (GBM) is one of the most serious types of gliomas. Although considerable progress has been made in the development of cancer therapeutic agents, several antineoplastic drugs fail to penetrate the blood-brain barrier (BBB), resulting in a low survival rate of glioma patients. Recent studies have revealed that the traditional Chinese medicine Buxus microphylla contains the main active component Cyclovirobuxine D (CVB-D), which can cross the BBB with a novel delivery system. However, it remains unclear whether CVB-D exerts anticancer effects against GBM and low-grade glioma (LGG). The aim of the present study was to explore the feasibility of CVB-D as a new effective agent in the treatment of GBM and LGG. The ability of CVB-D to inhibit GBM and LGG cell proliferation was detected by CCK8 assay. Flow cytometry was used to detect cell cycle progression and apoptosis induction by Annexin V-FITC/PI assay. The expression levels of the apoptosis-associated proteins, namely cleaved caspase-3 and Bax/Bcl-2, were detected by western blot analysis. The mitochondrial membrane potential (ΔΨm) was detected by Rh123 dyed fluorescence micrograph. Hoechst staining was used to observe the morphological characteristics of the apoptotic cells. The scratch test was used to evaluate the migration of GBM and LGG cells. The results indicated that CVB-D reduced cell viability of T98G and Hs683 cells. Flow cytometry demonstrated that CVB-D-treated cells were arrested at the S phase of their cell cycle. The expression levels of the apoptosis-associated proteins were increased in CVB-D-treated cells. Rh123 and Hoechst staining indicated morphological changes and mitochondrial membrane potential changes of the cells undergoing apoptosis. The data confirmed that CVB-D inhibited cell proliferation by arresting the cell cycle of GBM and LLG cells and that it promoted the induction of cell apoptosis by altering the mitochondrial membrane potential. The findings of the present study indicate the potential value of CVB-D in the treatment of glioma.
Keywords: Cyclovirobuxine D, glioma, apoptosis, cell cycle, migration, proliferation, traditional Chinese medicine
Introduction
Gliomas are the most common neoplasm of the central nervous system in humans. Glioblastoma multiforme (GBM) is one of the most serious types of gliomas. Although significant progress has been made in the development of novel therapies for the treatment of this disease, the survival rate of glioma patients remains considerably low. The median survival time in patients with newly diagnosed GBM is frequently less than 15 months (1,2). Approximately 77,000 new glioblastoma cases are diagnosed annually in the United States and Europe (3). The 5-year survival rate of patients is ~5% (4). Despite maximal tumor resection, high-dose radiation and temozolomide (TMZ) chemotherapy that are currently used in clinical treatment, the prognosis of GBM has not improved significantly. The majority of low-grade glioma (LGG) cases will develop into GBM with conventional therapy. Therefore, it is urgent to explore a new effective and feasible treatment for GBM and LGG.
Cyclovirobuxine D (CVB-D) is the main active component of the traditional Chinese medicine Buxus microphylla and has demonstrated a definitive therapeutic effect on various cardiovascular diseases (5,6). The mechanism of action of CVB-D has been recently studied (7–13). CVB-D [molecular formula: C26-H46-N2-O; molecular weight: 402.662; chemical name: 9,19-cyclopregnan-16-ol,4,4,14-trimethyl-3,20-bis(methylamino)-,(3β,5α,16α,20S)-] is a triterpenoid alkaloid (Fig. 1) extracted from the traditional Chinese medicine Buxus microphylla. This extract has also been investigated for its antitumor effect against breast cancer (13). It is interesting to note that CVB-D can cross the blood-brain barrier (BBB), suggesting that it can have higher efficacy against GBM and LGG treatment than other anti-cancer compounds (14). However, whether and how CVB-D affects GBM and LGG growth remains unknown. In view of these findings, the aim of the present study was to investigate the effects of CVB-D on human GBM and LGG cells.
Figure 1.
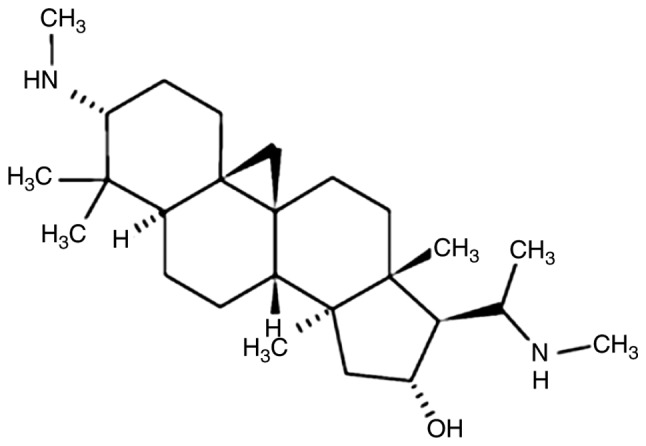
Chemical structure of CVB-D. CVB-D, Cyclovirobuxine D.
Materials and methods
Materials
CVB-D (C26H46N2O, FW 402.66, purity 66; YuanYe Biotechnology Co., Ltd.), the Rhodamine 123 (Rh123) kit (BB-41051-1), the Annexin V-FITC/PI apoptosis test kit (BB-4101-2) and the cell cycle test kit (BB-4104-2) were purchased from Shanghai BestBio Biotechnology Co., Ltd. Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular Technologies, Inc. Fetal bovine serum (100 kU/l) and penicillin and streptomycin (100 mg/l) were purchased from Gibco-BRL Invitrogen; Thermo Fisher Scientific, Inc. The BCA Protein Assay Kit (P0010), and the reagent kits for the protein measurements were purchased from Beyotime Institute of Biotechnology. Bax (cat no. ab69643), Bcl-2 (cat. no. ab32124), caspase-3 (cat. no. ab32351) and cleaved caspase-3 (cat. no. 9664S), beta-actin (cat. no. 4970S) antibodies were purchased from Cell Signaling Technology and Abcam, Inc. Horseradish peroxidase-conjugated anti-rabbit (cat. no. ab44171) and anti-mouse (cat. no. ab21172) antibodies were obtained from Bioworld Technology, Inc. Immobilon western chemiluminescent HRP Substrate and PVDF membranes were obtained from EMD Millipore. The Hoechst 33342 staining kit (C1028) was purchased from Beyotime Institute of Biotechnology.
Cell culture and cell lines
The GBM cell line T98G and the LGG cell line Hs683 were obtained from ATCC. The cells were cultured in high glucose DMEM medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C under an atmosphere of 5% CO2 and 95% air. In addition, high glucose DMEM supplemented with 2% FBS was used in the scratch testing. The medium (half of the total volume) was changed every 3 days during the culture period.
Cell viability assay and colony formation ability
Cell proliferation of T98G and Hs683 cells was monitored by CCK-8 assay. The cells were plated in a 96-well plate at a concentration of 1×106 cells/ml. The cells were pretreated with various concentrations of CVB-D (0, 15, 30, 60, 120 and 240 µmol/l) for 24, 48 and 72 h. Almost all cells reduced cell viability when treated with 240 µmol/l for 72 h, which was the highest concentration and longest time-scale threshold in the experiment. Each group included 6 replicates and was assessed in 3 different experiments. The 96-well plate was washed with PBS (0.01 M) and 90 µl of serum-free medium was mixed with 10 µl CCK-8 solution and added to the wells. The plate was incubated at 37°C without light. Following incubation for 1–2 h, the absorbance was monitored at 450 nm using a microplate reader (Tecan Group, Ltd.). The colony formation ability of T98G and Hs683 cells was analyzed following CVB-D treatment (0, 5, 10, 20, 40 and 80 µmol/l). It was determined that few cells floated when treated with 80 µmol/l, which the highest threshold concentration for 24 h, and had no effect on colony formation results. The cells were plated in 6-well plates with 500 cells/well. The cells were pretreated with various concentrations of CVB-D (0, 5, 10, 20, 40 and 80 µmol/l) for 24 h. Subsequently, the medium was changed in the culture for the next 10 days. Finally, the cells were permeabilized with 4% polyformaldehyde (10 min at room temperature) and stained with crystal violet (10 min, at room temperature, 0.05% w/v). The software GraphPad Prism 8 (GraphPad Software, Inc.) was used to perform statistical analysis of cell viability results.
Capture of mitochondrial membrane potential (MMP or ΔΨm)
MMP was assessed using fluorescent Rh123 dye using 6 different concentrations of CVB-D (0, 40, 80, 120, 160 and 200 µmol/l) for 6 h. Few cells floated and an evident trend was captured when cells were treated with 200 µmol/l CVB-D for 6 h, thus, this was determined as our highest concentration and time-scale threshold for the Rh123 staining experiment. Rh123 was added to the cultures according to the manufacturer's instructions. The images were captured by a flow cytometry (Beckman Coulter). The change in low staining of FITC+ cells corresponded to the MMP change of the cells. The software GraphPad Prism 8 was used to perform statistical analysis of the results.
Hoechst staining
A total of 6 different concentrations CVB-D (0, 40, 80, 120, 160 and 200 µmol/l) were added to the cells in 6-well plates for 6 h. Few cells floated and an evident trend was captured when cells were treated with 200 µmol/l CVB-D for 6 h, thus, this was determined as our highest concentration and time-scale threshold for the Hoechst 33342 staining experiment. Hoechst 33342 was added to the cultures according to the manufacturer's instructions. The images were captured by a fluorescent microscope (Zeiss GmbH). The change in brightness corresponded to the percentage of apoptotic cells. The software Image-Pro Plus 6.0 (Media Cybernetics, Inc.) was used to measure the mean fluorescence intensity (MFI) of figures.
Scratch testing
A total of 6 different concentrations CVB-D (0, 20, 40, 60, 80 and 100 µmol/l) were added to the cells in a 6-well plate with 2% FBS-DMEM culture medium for 12 h after the scratch wound was made. Few cells float and an evident trend was captured when cells were treated with 100 µmol/l CVB-D for 12 h, thus, this was determined as the highest concentration and time-scale threshold for the scratch experiment. A high concentration and a long culture duration increase the number of cells floating which affects the experimental results. Subsequently, the images were captured by a bright field view of the microscope (Zeiss GmbH). The distances between the the two edges of the scratched cells corresponded to the migration of GBM and LGG cells. The software ImageJ (version 1.50i; National Institutes of Health) was used to do assess the distance between the edges.
Flow cytometric assessment for apoptosis
T98G and Hs683 cells were seeded in 6-well plates and pretreated with 0, 80, 120, 160, 200 and 240 µmol/l CVB-D for 24 h. The highest concentration and longest time-scale were determined by cell viability experiment. Cells had <50% viability when treated with 240 µmol/l CVB-D for 24 h and an evident trend of apoptosis was captured. The induction of apoptosis was assessed by Annexin V-FITC/PI apoptosis test. The cell cycle analysis was performed by seeding cells in a 6-well plate and pretreating them with 0, 80, 160 and 240 µmol/l CVB-D for 24 h. The highest concentration and longest time-scale were determined by cell viability experiment. Cell had <50% viability when treated with 240 µmol/l CVB-D for 24 h and evident trend of the cell cycle was captured. The two tests were carried out in accordance with the manufacturer's instructions. The ModFit software (version 4.1) was used for cell cycle analysis. The CytExpert software (version 2.0) was used for detection of apoptosis and cell cycle progression. The software GraphPad Prism 8 was used to perform statistical analysis of the results.
Western blot analysis
All cells were subcultured in 6-well plates. The cells were incubated initially for 24 h and subsequently treated with 0, 50, 100 and 200 µmol/l CVB-D for an additional 24 h. Since cell lysis had evident reduced protein collection when treated with 240 µmol/l CVB-D for 24 h, the highest concentration and longest time-scale was determined as 200 µmol/l. The concentration ratio 2:1 was used for grouping and an obvious trend was captured in the western blot experiment. The cells were subsequently washed with ice-cold PBS and lysed in RIPA buffer with 1X PMSF (Beyotime Institute of Biotechnology). The protein concentration was measured with the BCA method. SDS-polyacrylamide gels (10%) were used for electrophoresis. Following electrophoresis, the proteins were transferred to PVDF membranes and blocked in 5% skimmed milk in TBST for 1 h. The membranes were probed with primary antibodies for β-actin, Bax, Bcl-2, caspase-3 and cleaved caspase-3 (all 1:1,000; all from Cell Signaling Technology, Inc.) at 4°C overnight. The membranes were subsequently washed and incubated with secondary antibodies for 1 h [1:10,000, goat anti-mouse IgG (H+L) HRP, product code ab21172; 1:10,000, goat anti-rabbit IgG (H+L) HRP, product code ab44171] according to the source of primary antibodies. The membranes were subsequently visualized by chemiluminescent ECL reagent (EMD Millipore). The software ImageJ (version 1.50i) was used for densitometric analysis of protein expression and statistical analysis by SPSS 22 software (IBM Corp.).
Statistical analysis
The unpaired Student's t-test was used to determine the differences between the two groups. The data were presented as the mean ± standard error of mean (SEM). One way ANOVA followed by Bonferroni's post hoc test were used to perform multi-group data comparisons using GraphPad Prism 8 and SPSS 22 software. P<0.05 was used to indicate a statistically significant difference.
Results
CVB-D reduces cell viability and colony formation ability of GBM and LGG cells
Crystal violet staining indicated that the number of colonies of CVB-D-treated T98G and Hs683 cells were markedly decreased compared with those noted in untreated cells (Fig. 2). Subsequently, various concentrations of CVB-D were used to treat the cells. Following incubation with 0, 15, 30, 60, 120 and 240 µmol/l CVB-D for 24, 48 and 72 h, the viability of the cells was assessed using a CCK-8 kit. The results indicated that the cell viability was reduced in a concentration- and time-dependent manner (Fig. 3). All dosing groups exhibited statistical significant differences compared with the control group (data not shown).
Figure 2.

Colony formation abilities of T98G and Hs683 cells treated with CVB-D. CVB-D, Cyclovirobuxine D.
Figure 3.

CVB-D reduces cell viability of T98G and Hs683 cells. Each experiment involved three replicates. CVB-D, Cyclovirobuxine D.
CVB-D arrests cells at the S and G0/G1 phase
The cell cycle plays an important role in cancer cell proliferation. Therefore, the cell cycle of CVB-D pretreated GBM cells was assessed by flow cytometry (Fig. 4). Subsequently, treatment of GBM cells with different concentrations of CVB-D arrested the cells at the S phase. Concomitantly, the number of cells in the other two populations was decreased. LGG cells were arrested at the G0/G1 phase according to the concentration of CVB-D used. In addition, the number of cells in the other two populations were both decreased. These phenomena indicated that the effects of CVB-D on the cell cycle were concentration-dependent, which may be used as a strategy for tumor therapy.
Figure 4.

Cell cycle of CVB-D-treated glioma cells. (A) Cell cycle analysis of CVB-D treated T98G and Hs683 cells as detected by flow cytometry (0, 80, 160 and 240 µmol/l). (B) Statistical analysis of the percentage of cells at the G0/G1, S and G2/M phases of the T98G and Hs683 cell cycle. Each group was compared with the 0 µmol/l group with regard to the S phase. **P<0.01, ***P<0.001, ****P<0.0001. CVB-D, Cyclovirobuxine D.
CVB-D induces cell apoptosis by a mitochondrial-dependent pathway
CVB-D pretreated GBM and LGG cells were stained by PI and Annexin V-FITC dyes. The results indicated that the surviving cells were PI−/Annexin V−. PI−/Annexin V+ and PI+/Annexin V+ cells represented the two phases of cell apoptosis. Early and late apoptotic cells were labeled as PI−/Annexin V+ and PI+/Annexin V+, respectively. The number of surviving cells was decreased following an increase in the treatment of the cells by CVB-D (Fig. 5). Subsequently, Hoechst staining was used for the observation of the morphological characteristics of the apoptotic GBM and LGG cells. Each Hoechst staining group was monitored by fluorescence microscopy (Fig. 6). Concomitantly, Rh123 staining was used to confirm the induction of cell apoptosis by the mitochondrial-dependent pathway (Fig. 7).
Figure 5.

CVB-D induces apoptosis of T98G and Hs683 cells. (A) Apoptotic analysis was performed by flow cytometry. (B) Statistical analysis of the percentages of apoptotic cells following treatment with various concentrations of CVB-D. Each treatment dosing group was compared with the control group. *P<0.05, ***P<0.001, ****P<0.0001. Survival: PI−/Annexin V−; early apoptosis: PI−/Annexin V+; late apoptosis: PI+/Annexin V+. Each experiment involved three replicates. CVB-D, Cyclovirobuxine D.
Figure 6.

Hoechst staining is used to observe the morphological characteristics of T98G and Hs683 cells. The results indicated that the percentage of hyperchromatic nuclei was increased as the concentration of CVB-D increased. The MFI was assessed by Image-Pro Plus 6.0, and statistically analyzed by GraphPad Prism 8. *P<0.05. CVB-D, Cyclovirobuxine D; MFI, mean fluorescence intensity.
Figure 7.

Rh123 staining of T98G and Hs683 cells following various concentrations of CVB-D pretreatment. The staining analysis was performed by flow cytometry. The results indicated that the hyperchromatic cells were increased as the concentration of CVB-D increased. The change in low staining of FITC+ cells corresponded to the MMP change of the cells (MMP or ΔΨm). Statistical analysis of low staining of FITC+ cells was performed using SPSS 22. *P<0.05. CVB-D, Cyclovirobuxine D; Rh123, Rhodamine 123.
CVB-D significantly inhibits tumor cell migration
Initially, various concentrations of CVB-D (0, 20, 40, 60, 80 and 100 µmol/l) were used for 12 h pretreatment of the cells and cell migration was assessed by light microscopy. The results indicated that CVB-D significantly inhibited tumor cell migration of GBM and LGG cells. Significant differences were noted in all the CVB-D groups compared with the control samples (Fig. 8).
Figure 8.

Scratch test of T98G and Hs683 cells. The scratch test was conducted at 12 h and the results indicated that tumor cell migration was significantly inhibited following a concentration increase of CVB-D. Distances between the edges of the cells was analyzed by SPSS 22. *P<0.05. CVB-D, Cyclovirobuxine D.
CVB-D induces the expression levels of apoptosis-associated proteins in GBM and LGG cells
The expression levels of the proteins associated with cell apoptosis were examined by western blotting. The results indicated that the expression levels of Bax and cleaved caspase-3 were increased. In contrast to these findings, the expression levels of caspase-3 and Bcl-2 were decreased in CVB-D-pretreated T98G and Hs683 cells in a concentration-dependent manner. These results indicated that the apoptotic process was regulated by increased expression in the levels of the apoptosis-associated proteins. The ratios of Bax/Bcl-2 and cleaved caspase-3 were increased suggesting the induction of apoptosis in T98G and Hs683 cells (Fig. 9).
Figure 9.

Various concentrations of CVB-D induce the expression of apoptosis-associated proteins in T98G and Hs683 cells. The cells were pretreated with various concentrations of CVB-D for 6 h and the protein bands were detected by western blotting. β-actin was used as the control protein. The software ImageJ (version 1.50i) was used for densitometric analysis of the protein expression and statistical analysis was performed using SPSS 22 software. *P<0.05. CVB-D, Cyclovirobuxine D.
Discussion
Human brain glioblastoma is a malignant tumor, which usually recurs from the surgical resection area and is characterized by the tumor infiltration or the transfer of tumor cells along the white matter fiber bundle. The main treatment strategy involves the inhibition of glioblastoma cell migration. Previous studies have revealed that its average incidence rate is 3.19/100,000 in the population, whereas the 5-year survival rate of GBM patients is estimated to ~5% (4,15). Conventional surgery and radiotherapy/chemotherapy do not effectively prolong the survival of patients with GBM. These patients do not often survive from postoperative tumor recurrence and their median survival rate is estimated to ~15 months (15). Therefore, it is important to identify and develop new effective strategies for the treatment of GBM.
CVB-D is the main active component of the traditional Chinese medicine Buxus microphylla, which has been used to treat cardiovascular diseases (12). One of the main obstacles in the treatment of brain tumors is the inability of the chemotherapeutic drugs to cross the BBB. Therefore, although several novel drugs have been successfully applied in the treatment of various types of tumors their efficacy is considerably low in brain tumors, notably in GBM and LGG. The antitumor effect of CVB-D has been investigated in breast and gastric cancers (13,16). However, it is uncertain whether it is effective in the treatment of GBM and LGG. Previous studies have revealed that cell apoptosis can be induced by a variety of drugs and physical and chemical factors (17–19). Moreover, a previous study demonstrated that CVD-B exhibited antitumor effects via the Akt/mTOR pathway (13), whereas it has also been revealed that the Akt/mTOR signaling pathway is associated with various physiological processes including cell growth, proliferation and apoptosis (20–22). A recent study developed a novel drug delivery system for CVB-D, which can in theory enhance the efficacy of CVB-D therapy (14).
CVB-D affects the expression of the family members of cysteine-containing aspartate-specific proteases (caspases), which contain several proteins that play a key role in the cellular process of apoptosis. Caspase-3 is one of the most important members of this protein family, which has been reported to be the key executor of cell apoptosis. Upon activation of caspase enzymes by the external apoptotic signals, the apoptotic cascade is activated. Subsequently, the apoptosis-signaling pathway of the cells is activated by the interaction of caspases with several other proteases. In the present study, it was demonstrated that CVB-D induced apoptosis via upregulation of apoptosis-associated proteins. Future studies may address the investigation of the gene expression profile of CVB-D-induced apoptotic cells.
The data reported in the present study revealed that the GBM and LGG cell migration and cell cycle progression were inhibited following treatment of the cells with a low concentration of CVB-D. This strategy can be applied as a potential adjuvant therapy in GBM or LGG patients that are treated with temozolamide.
In addition, CVB-D could regulate calcium levels inside the cells (23). Previous studies have suggested that calcium levels play an important role in tumor growth, apoptosis and differentiation (24–26). The hypothesis that CVB-D can promote tumor apoptosis by affecting calcium ion channels can be further explored in future studies and can aid the investigation of the antitumor activity of this compound.
The present study, to the best of our knowledge, is the first to reveal the antitumor activity of CVB-D in human GBM and LGG. The results demonstrated that CVB-D inhibited cell proliferation and tumor migration, while it induced apoptosis via the mitochondrial-dependent pathway. CVB-D is a main component of the traditional Chinese medicine Buxus microphylla and exhibits potent antitumor effects against GBM and LGG. This evidence can offer insight in the treatment of this type of disease.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science Foundation of China (grant. no. 31360258); Special fund for science and technology development of Guangdong Province (no. 2016A020215036); natural Science Foundation of Guangdong Province (nos. 2015A030313077, 2015A030313047 and 2017A030310192); project of Educational Commission of Guangdong Province (2018GkQNCX085); Science and Technology Program of Jiangmen (2019E021).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
LZ, KG and JX designed the study. LZ performed the experiments and HT wrote the manuscript. HT, FW, SO, TW, YF helped to performed the experiments and collected the data, HT and FW participated in the statistical analysis. All authors read and approved the final manuscript. All authors have read and approved the final manuscript and agree to be accountable for all aspects of the research in ensuring that the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Imperato JP, Paleologos NA, Vick NA. Effects of treatment on long-term survivors with malignant astrocytomas. Ann Neurol. 1990;28:818–822. doi: 10.1002/ana.410280614. [DOI] [PubMed] [Google Scholar]
- 2.Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352:987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 3.Ashok AR, Pouratian N, Sherman J, Ahmed G, Shaffrey ME. Advances in brain tumor surgery. Neurol Clin. 2007;25:975–1003. doi: 10.1016/j.ncl.2007.07.006. [DOI] [PubMed] [Google Scholar]
- 4.Shan P, Mao RB, Xu JM, Li JX. The beneficial effects of cyclovirobuxine D (CVBD) in coronary heart disease. A double blind analysis of 110 cases. J Tradit Chin Med. 1984;4:15–19. [PubMed] [Google Scholar]
- 5.Grossini E, Battaqlia A, Brunelleschi S, Mary DA, Molinari C, Viano I, Vacca G. Coronary effects of cyclovirobuxine D in anesthetized pigs and in isolated porcine coronary arteries. Life Sci. 1999;65:59–65. doi: 10.1016/S0024-3205(99)00278-7. [DOI] [PubMed] [Google Scholar]
- 6.Yu B, Fang TH, Lü GH, Xu HQ, Lu JF. Beneficial effect of cyclovirobuxine D on heart failure rats following myocardial infarction. Fitoterapia. 2011;82:868–877. doi: 10.1016/j.fitote.2011.04.016. [DOI] [PubMed] [Google Scholar]
- 7.Grossini E, Avanzi G, Gallicchio M, Molinari C, Vacca G, Bellomo G. Regulation of Ca2+ Movements by cyclovirobuxine D in ECV304 endothelial cells. Pharmacol Res. 2005;52:154–161. doi: 10.1016/j.phrs.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 8.Chen QW, Shan HL, Sun HL, Wang H, Yang BF. Effects of cyclovirobuxine D on intracellular Ca2+ and L-type Ca2+ current in rat ventricular cardiomyocytes. Yao Xue Xue Bao. 2004;39:500–503. (In Chinese) [PubMed] [Google Scholar]
- 9.Hu D, Liu X, Wang Y, Chen S. Cyclovirobuxine D ameliorates acute myocardial ischemia by K(ATP)channel opening, nitric oxide release and anti-thrombosis. Eur J Pharmacol. 2007;569:103–109. doi: 10.1016/j.ejphar.2007.04.038. [DOI] [PubMed] [Google Scholar]
- 10.Yue Y, Liu R, Liu J, Dong Q, Fan J. Experimental and theoretical investigation on the interaction between cyclovirobuxine D and human serum albumin. Spectrochim Acta A Mol. Biomol. Spectrosc. 2014;128:552–558. doi: 10.1016/j.saa.2014.03.007. [DOI] [PubMed] [Google Scholar]
- 11.Guo Q, Guo J, Yang R, Peng H, Zhao J, Li L, Peng S. Cyclovirobuxine D attenuates doxorubicin-induced cardiomyopathy by suppression of oxidative damage and mitochondrial biogenesis impairment. Oxid Med Cell Longev. 2015;2015:151972. doi: 10.1155/2015/151972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lu J, Sun D, Gao S, Gao Y, Ye J, Liu P. Cyclovirobuxine D induces autophagy-associated cell death via the Akt/mTOR pathway in MCF-7 human breast cancer cells. J Pharmacol Sci. 2014;125:74–82. doi: 10.1254/jphs.14013FP. [DOI] [PubMed] [Google Scholar]
- 13.Burri SH, Gondi V, Brown PD, Mehta MP. The evolving role of tumor treating fields in managing glioblastoma: Guide for Oncologists. Am J Clin Oncol. 2018;41:191–196. doi: 10.1097/COC.0000000000000395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Thakkar JP, Dolecek TA, Horbinski C, Ostrom QT, Lightner DD, Barnholtz-Sloan JS, Villano JL. Epidemiologic and molecular prognostic review of glioblastoma. Cancer Epidemiol Biomarkers Prev. 2014;23:1985–1996. doi: 10.1158/1055-9965.EPI-14-0275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu J, Tan Z, Chen J, Dong C. Cyclovirobuxine D inhibits cell proliferation and induces mitochondria-mediated apoptosis in human gastric cancer cells. Molecules. 2015;20:20659–20668. doi: 10.3390/molecules201119729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhao W, Li X, Wang J, Wang C, Jia Y, Yuan S, Huang Y, Shi Y, Tong Z. Decreasing eukaryotic initiation factor 3C (EIF3C) suppresses proliferation and stimulates apoptosis in breast cancer cell lines through mammalian target of rapamycin (mTOR) pathway. Med Sci Monit. 2017;23:4182–4191. doi: 10.12659/MSM.906389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lin J, Feng J, Yang H, Yan Z, Li Q, Wei L, Lai Z, Jin Y, Peng J. Scutellaria barbata D. Don inhibits 5-fluorouracil resistance in colorectal cancer by regulating PI3K/AKT pathway. Oncol Rep. 2017;38:2293–2300. doi: 10.3892/or.2017.5892. [DOI] [PubMed] [Google Scholar]
- 18.Tan J, Shen W, Shi W, Chen X, Sun D, Xu C, Yan Q, Cheng H, Lai Y, Ji H. ONTD induces growth arrest and apoptosis of human hepatoma Bel-7402 cells though a peroxisome proliferator-activated receptor γ-dependent pathway. Toxicol In Vitro. 2017;45:44–53. doi: 10.1016/j.tiv.2017.08.012. [DOI] [PubMed] [Google Scholar]
- 19.Chen S, Fisher RC, Sign S, Molina LA, Shenoy AK, Lopez MC, Baker HV, Koomen JM, Chen Y, Gittleman H, et al. Inhibition of PI3K/Akt/mTOR signaling in PI3KR2-overexpressing colon cancer stem cells reduces tumor growth due to apoptosis. Oncotarget. 2016;8:50476–50488. doi: 10.18632/oncotarget.9919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Amini-Farsani Z, Sangtarash MH, Shamsara M, Teimori H. MiR-221/222 promote chemoresistance to cisplatin in ovarian cancer cells by targeting PTEN/PI3K/AKT signaling pathway. Cytotechnology. 2018;70:203–213. doi: 10.1007/s10616-017-0134-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zahed Panah M, Nikbakht M, Sajjadi SM, Rostami S, Norooznezhad AH, Kamranzadeh Fumani H, Ghavamzadeh A, Mohammadi S. Anti-apoptotic effects of osteopontin via the up-regulation of AKT/mTOR/β-catenin loop in acute myeloid leukemia cells. Int J Hematol Oncol Stem Cell Res. 2017;11:148–157. [PMC free article] [PubMed] [Google Scholar]
- 22.Yu B, Ruan M, Zhou L, Xu L, Fang T. Influence of cyclovirobuxine D on intracellular [Ca2+] regulation and the expression of the calcium cycling proteins in rat myocytes. Fitoterapia. 2012;83:1653–1665. doi: 10.1016/j.fitote.2012.09.019. [DOI] [PubMed] [Google Scholar]
- 23.Chen YF, Chen YT, Chiu WT, Shen MR. Remodeling of calcium signaling in tumor progression. J Biomed Sci. 2013;20:23. doi: 10.1186/1423-0127-20-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stewart TA, Yapa KT, Monteith GR. Altered calcium signaling in cancer cells. Biochim Biophys Acta. 2015;1848:2502–2511. doi: 10.1016/j.bbamem.2014.08.016. [DOI] [PubMed] [Google Scholar]
- 25.Macià A, Herreros J, Martí RM, Cantí C. Calcium channel expression and applicability as targeted therapies in melanoma. Biomed Res Int. 2015;2015:587135. doi: 10.1155/2015/587135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wei H, Liu T, Jiang N, Zhou K, Yang K, Ning W, Yu Y. A novel delivery system of Cyclovirobuxine D for brain targeting: Angiopep-conjugated polysorbate 80-coated liposomes via intranasal administration. J Biomed Nanotechnol. 2018;14:1252–1262. doi: 10.1166/jbn.2018.2581. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.