

Abstract
Purpose:
Members of the ciliary neurotrophic factor (CNTF) family of cytokines have been shown to influence neuronal differentiation during retinal development and enhance cell survival in various retinal degeneration models. However, the cellular mechanism of CNTF signaling and the target cell types for CNTF in the developing retina remain unidentified. The purpose of this study is to characterize expression patterns of proteins involved in cytokine signal transduction in the mouse retina, thus to assess the potential responsiveness of different retinal cell types to CNTF-like cytokine signals.
Methods:
The expression profiles of various cytokine signal transduction components, including receptor subunits CNTF receptor alpha (CNTFRα) and gp130, intracellular protein kinases, Jak2 and Tyk2, as well as latent transcription factors, STAT1 and STAT3, were determined by immunohistochemical staining of mouse retinal sections derived from different postnatal stages. In addition, the distribution of ERK was studied by immunofluorescent staining.
Results:
In the neonatal retina, intense staining signals for gp130, CNTFRα, Jak2, Tyk2, STAT1, and STAT3 were present in the differentiated ganglion cell layer and the developing inner plexiform layer of the mouse retina. Detectable staining signals were also observed in the ventricular zone of the early postnatal mouse retina. From P5 to P10, cytokine signaling molecules also accumulated in the developing outer plexiform layer. In the adult retina, cytokine signaling components examined were localized to the ganglion cell layer, the inner nuclear layer, and the two plexiform layers. In addition, regions corresponding to the inner and/or outer segments of the photoreceptor cells showed positive staining for cytokine signaling components. In contrast, the ERK2 protein kinase was found throughout the neonatal retina. In the mature retina, ERK2 was concentrated in the ganglion cells and the inner plexiform layer, while a lesser expression of ERK2 was detected in the inner nuclear layer, the outer plexiform layers, and the outer nuclear layer.
Conclusions:
In the neonatal mouse retina, signaling components of the Jak-STAT pathway and ERK2 are differentially expressed. All cytokine signaling components included in this study are expressed in the differentiated inner retina as well as in cells occupying the ventricular zone, suggesting that both postmitotic neurons and proliferative progenitors may directly respond to CNTF-like cytokines during postnatal development. The distribution of cytokine signaling pathway components in the adult mouse retina is consistent with previous findings that ganglion cells and Müller glia are the primary target cell types for CNTF.
Development of the vertebrate retina from an undifferentiated neural epithelium to a mature neural network is regulated by both cell-intrinsic factors and cell-extrinsic cues [1–4]. Ciliary neurotrophic factor (CNTF) has been shown to be a potent growth factor capable of influencing retinal cell differentiation. During the peak period of rod photoreceptor cell production in rodents, exposure of retinal tissues to exogenous CNTF results in inhibition of rhodopsin expression and enhancement of bipolar cell marker expression [5,6]. In contrast to rodents, CNTF promotes the differentiation of a subclass of cone photoreceptors expressing green opsin in the developing chick retina [7–9]. In addition to its strong effects on neuronal differentiation, CNTF promotes the long-term survival and axon outgrowth of retinal ganglion cells [10–13] and prevents photoreceptor cell degeneration caused by mutations and light-induced damages in various rodent models [14–20].
The CNTF subfamily of cytokines, including leukemia inhibitory factor (LIF), interleukin-6, oncostatin M, and cardiotrophin-1, share two common transmembrane receptor components: gp130 and LIFRβ [21]. In addition, CNTF also requires a specific receptor subunit, CNTFRα, that is anchored to the cell membrane by a glycosylphosphatidylinositol (GPI) linkage [22,23]. The binding of CNTF to its tripartite receptor induces dimerization of gp130 and LIFRβ, which in turn activates receptor-associated Jak protein tyrosine kinases [24]. In vitro, CNTF can activate three known Jak kinases: Jak1, Jak2, and Tyk2 [25]. Activation of Jak kinases causes phosphorylation of latent cytoplasmic transcription factors of the signal transducer and activator of transcription (STAT) family [25]. Among the seven STAT proteins identified in mammals [26], CNTF family of cytokines selectively phosphorylates STAT1 and STAT3 [27–29]. The tyrosine-phosphorylated forms of STAT dimerize and translocate into the nucleus to regulate gene transcription [27–30]. In addition to the Jak-STAT signaling pathway, the CNTF subfamily of cytokines can also activate the extracellular signal-regulated kinase (ERK, also known as the p42/44 mitogen-activated protein (MAP) kinase) [31] and the phosphatidylinositol 3 kinase-Akt pathways [32].
Despite the known effects of CNTF on retinal differentiation and cell survival, the cellular mechanisms of CNTF signaling are not well understood. In the adult rodent retina, strong neural protective effects of CNTF have been observed for photoreceptor cells, however, CNTF-induced activation of STAT3 and ERK has been detected only in ganglion cells and Müller cells [33,34], suggesting that CNTF may not directly act upon photoreceptor cells. In the postnatal developing retina, which consists of proliferating progenitor cells and postmitotic neurons, the strong inhibitory effect of CNTF on rod photoreceptor differentiation may be caused either by direct cytokine action upon photoreceptor precursors or by secondary signals released by postmitotic neurons that have received CNTF signals. To further elucidate the cellular mechanisms of cytokine signaling during retinal differentiation, it is necessary to determine which cytokine signaling components are present in the retina and whether cytokine signaling components are expressed in a dynamic pattern during development and maturation of the retina. Here, we report the expression patterns of several key proteins mediating cytokine signal transduction in the postnatal mouse retina. The distribution of these signaling components indicates that both progenitor cells and postmitotic neurons are potential targets for CNTF signaling during early postnatal retinal development.
METHODS
Animals:
CD 1 mice were purchased from Charles River (Wilmington, MA). Immediately after being enucleated, the eyes were submerged in 4% paraformaldelhyde (PFA) in PBS at 4°C and the cornea and lens were removed. The resulting eye cups were fixed further in 4% PFA/PBS for 3–4 h at 4°C followed by cryoprotecting in 30% sucrose/PBS and embedding in OCT. Tissue sections of 16 μm thickness were cut with a cryostat. Tissue sections derived from different postnatal ages were collected and arranged on glass slides so that each slide contained sections from different ages. Mouse pups within 24 h of birth were considered as postnatal day 0 (P0) animals, and the rest were staged accordingly. The ages of mature mice used ranged from P24 to P36, and no differences in protein expression were found within this time period. A minimum of three eyes from a given age was analyzed.
Immunostaining:
The following affinity-purified primary antibodies and the corresponding antigen peptides were purchased from Santa Cruz Biotechnology (Santa Cruz, CA): rabbit polyclonal antibodies against Jak2 (C-20), Tyk2 (C-20), STAT1 (E-23), STAT3 (K-15), gp130 (C-20), ERK2 (C-14), and a goat polyclonal antibody against CNTFRα (R-20). The following secondary antibodies were used in this study: biotinylated goat anti-rabbit IgG (Vector Laboratories), biotinylated horse anti-goat IgG (Vector Laboratories, Burlingame, CA), and Alexa 594-conjugated goat anti-rabbit IgG (Molecular Probes, Eugene, OR).
Cryosections were postfixed with 4% PFA/PBS at room temperature for 10 min and washed with PBS 4 times for 10 min each. To inhibit endogenous peroxidases, sections were incubated with 0.3% H2O2 in water for 30 min at room temperature and followed by PBS washes. Sections were then incubated with a blocking solution containing 10% FCS, 2% goat serum (for rabbit antibodies) or 2% horse serum (for the goat antibody), and 0.1% Triton X-100 in DMEM for 1 h at room temperature or overnight at 4°C to block non-specific binding sites. Primary antibodies were diluted using the corresponding blocking solutions at the following ratios: Jak2 (1:200), Tyk2 (1:200), STAT1 (1:400), STAT3 (1:400), gp130 (1:400), ERK2 (1:200), and CNTFRα (1:50). To test if a given antigen peptide can block the binding sites of the corresponding antibody, the peptides were diluted to 5–10 μg/ml in a blocking solution containing the corresponding primary antibody and incubated at room temperature for 2 h prior to applying to sections. Incubations with primary antibodies or primary antibodies preincubated with antigen peptides were performed in parallel at 4°C overnight. Primary antibody incubation is followed by extensive washes with PBS containing 0.1% Tween 20 (PBT). Sections were subsequently incubated with Alexa 594-conjugated goat anti-rabbit IgG (1:200), biotinylated goat anti-rabbit IgG (1:200), or biotinylated horse anti-goat IgG (1:200) diluted into corresponding blocking solutions for 1 h at room temperature. Following washes with PBT, horseradish peroxidase immunohistochemistry was performed using the Elite ABC kit (Vector Laboratories) and 3’, 3’-Diaminobenzidine (DAB) as chromogen at room temperature for 20 min. Slides were washed extensively with H2O, and during the last wash, treated for 5 min with 1 μg/ml 4’−6-diamidino-2-phenylindole (DAPI). Slides were mounted with Gel Mount (Biomeda, Foster City, CA). Nomarski or fluorescent images were viewed and photographed with a Nikon E800 microscope equipped with a SPOT II digital camera. Fluorescent images were recorded with the same exposure times for Alexa 594 and DAPI, respectively.
Background peroxidase staining was detected in retinal blood vessels, which was abolished with the pretreatment of H2O2 (data not shown). Parallel staining performed without primary antibodies and with secondary antibodies alone showed no staining signals. Peptide preincubation experiments showed that for a given antibody only preincubation with the antigen peptide but not nonspecific peptides resulted in the loss of staining signals.
RESULTS
Expression of gp130 and CNTFRα in the retina:
To examine the distribution of the cytokine receptor subunit gp130, affinity purified rabbit polyclonal antibodies were used to stain mouse retinal sections (Figure 1). At P1, predominant gp130 expression was detected in the ganglion cell layer and the differentiating inner plexiform layer (Figure 1A). However, a lower intensity staining signal of gp130 protein was present in the ventricular zone of the mouse retina at P1 (Figure 1A). This pattern of expression persisted throughout the first postnatal week. At P7, the staining signals increased in the inner half of the inner nuclear layer and also appeared in the outer plexiform layer (Figure 1D). By P10, the outer plexiform layer became intensely stained compared to younger retinas (Figure 1E). In the mature retina (P36), the strongest staining signals for gp130 were detected in the ganglion cell layer, and intense staining signals were also present in a subset of cells in the inner half of the inner nuclear layer, and in both plexiform layers (Figure 1G). Interestingly, gp130 immunostaining signals were also present in both the outer and inner segments of mature photoreceptors (Figure 1G). Preincubation of the anti-gp130 antibody with the corresponding peptide antigen abolished the staining signals at P1 (Figure 1B) and other stages (data not shown). Staining sections with the secondary antibody alone resulted in no staining signals (Figure 1H, other data not shown).
Figure 1.
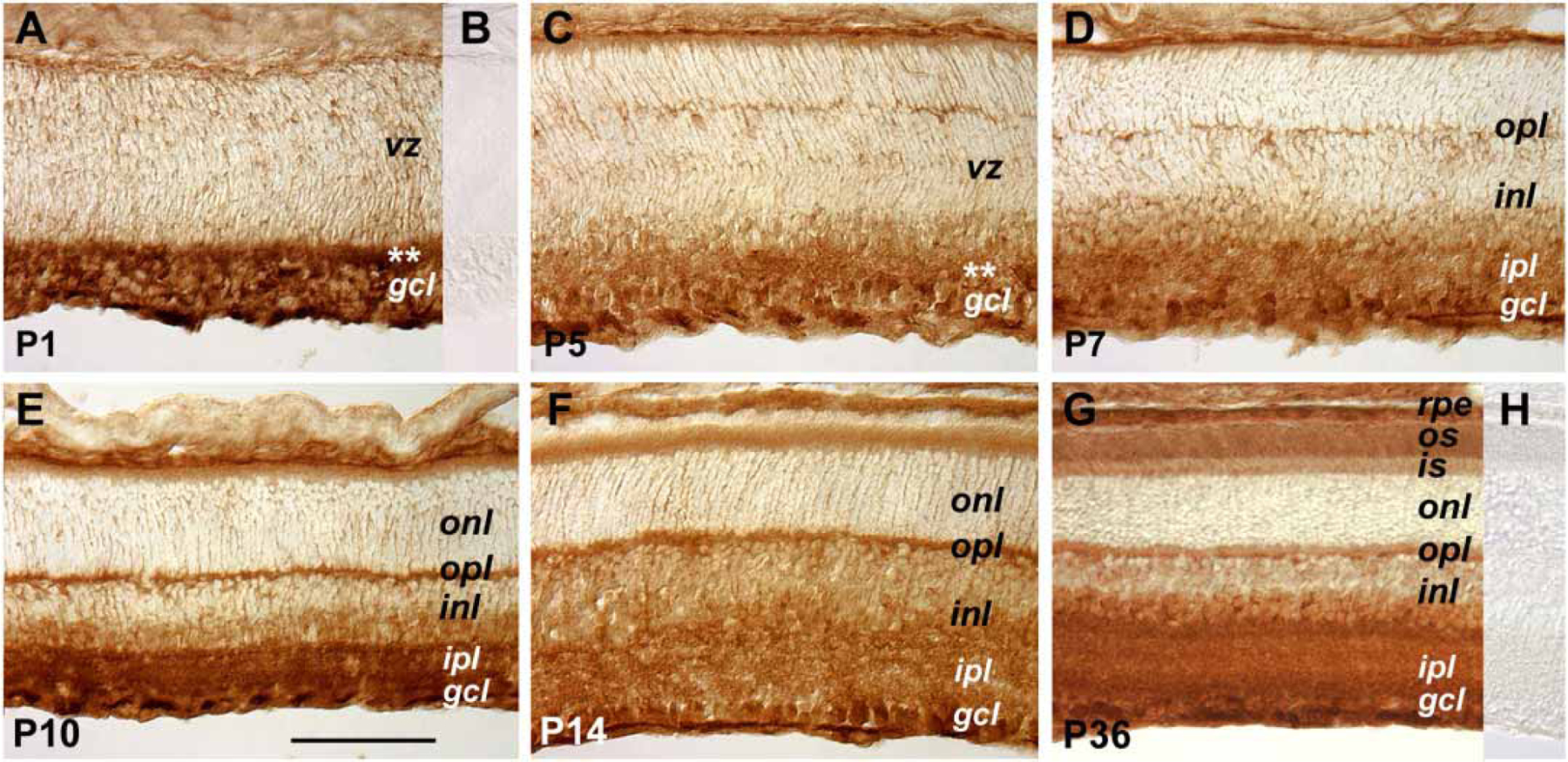
Expression patterns of the gp130 receptor. Cryosections of retinas from different postnatal ages immunohistochemically stained for gp130 are shown. Panels B and H were derived from the same developmental stages as panels A and G, respectively. The primary antibodies used to stain the section shown in panel B were preincubated with the antigen peptide. The staining of panel H was performed without primary antibodies. The asterisks in panels A and C indicate the position of the developing inner plexiform layer at P1 and P5, respectively. The scale bar shown in panel E represents 100 μm for all panels. The meanings of the abbreviations referred to in this figure are: gcl (ganglion cell layer), inl (inner nuclear layer), ipl (inner plexiform layer), is (inner segment), onl (outer nuclear layer), opl (outer plexiform layer), os (outer segment), rpe (retinal pigmented epithelium), vz (ventricular zone).
Expression of the CNTFRα protein was characterized by immunohistochemical staining using an affinity purified goat polyclonal antibody (Figure 2). At P1, most CNTFRα staining signals were found in the inner retina, where the ganglion cells and the differentiating inner plexiform layer were located (Figure 2A). Similar staining in the inner retina was maintained at P5 (Figure 2B,C) and P7 (Figure 2D). In addition, detectable staining, which appeared as clustered punctate signals, persisted in the ventricular zone throughout the first postnatal week (Figure 2A–D). By P10, the outer plexiform layer became positively stained for CNTFRα (Figure 2E). From P10 on, the most intense staining was found in the apical surface of the developing photoreceptor cell layer (Figure 2E,F). In the mature retina (P24), CNTFRα staining signals were localized to the ganglion cell layer, the two plexiform layers, and the inner nuclear layer (Figure 2G,H). Furthermore, prominent staining signals were detected in the inner and outer segments of photoreceptors (Figure 2H), and among them a subset of the outer segments was more intensely stained than the rest.
Figure 2.
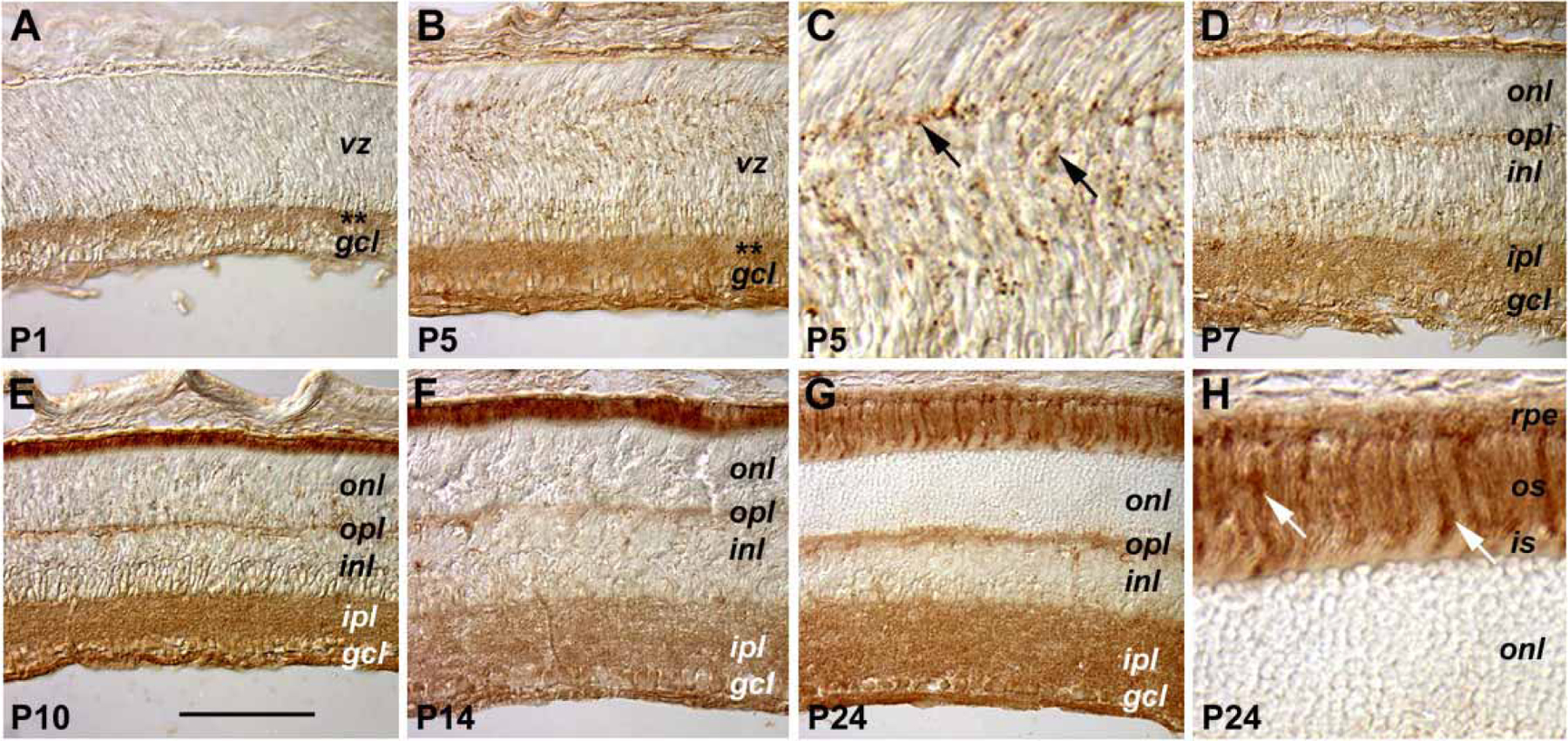
Expression patterns of the CNTFRα protein. Cryosections of retinas from different postnatal ages immunohistochemically stained for CNTFRα are shown. The asterisks in panels A and B indicate the position of the developing inner plexiform layer at P1 and P5, respectively. The scale bar shown in panel E represents 100 μm for panels A, B, D, E, F, and G. Panels C and H are 2.5 fold enlargement of portions of panels B and G, respectively. Arrows in panel C indicate the punctate staining signals in the ventricular zone of the P5 retina. Arrows in panel H point to heavily stained outer segments of a subset of photoreceptors at P24. The meanings of the abbreviations referred to in this figure are: gcl (ganglion cell layer), inl (inner nuclear layer), ipl (inner plexiform layer), is (inner segment), onl (outer nuclear layer), opl (outer plexiform layer), os (outer segment), rpe (retinal pigmented epithelium), vz (ventricular zone).
Distribution of Tyk2 and Jak2 protein kinases in the retina:
The expression patterns of the Jak1 protein kinase in the mouse retina have been reported previously [35]. To characterize the expression profiles of the other two Jak tyrosine kinases involved in mediating cytokine signals, we performed immunohistochemistry using specific antibodies against Tyk2 and Jak2. At P1, similar protein expression patterns were observed for Tyk2 (Figure 3A) and Jak2 (Figure 3D) with intense staining in the inner retina and lesser staining in the ventricular zone. At P5, sparsely distributed cells adjacent to the emerging outer plexiform layer showed strong staining signals for Tyk2 (Figure 3B) and Jak2 (Figure 3E). These cells were likely postmitotic horizontal cells. By three weeks of age, both kinases were present in the two plexiform layers as well as in the ganglion cell layer and the inner nuclear layer (Figure 3C,F). In addition, staining signals for both Tyk2 and Jak2 were also present in the inner segments of the mature photoreceptors. Similar protein expression patterns of Tyk2 and Jak2 were found in the postnatal rat retina, and staining signals for each antibody were specifically blocked by preincubation with their corresponding antigen peptides (data not shown).
Figure 3.
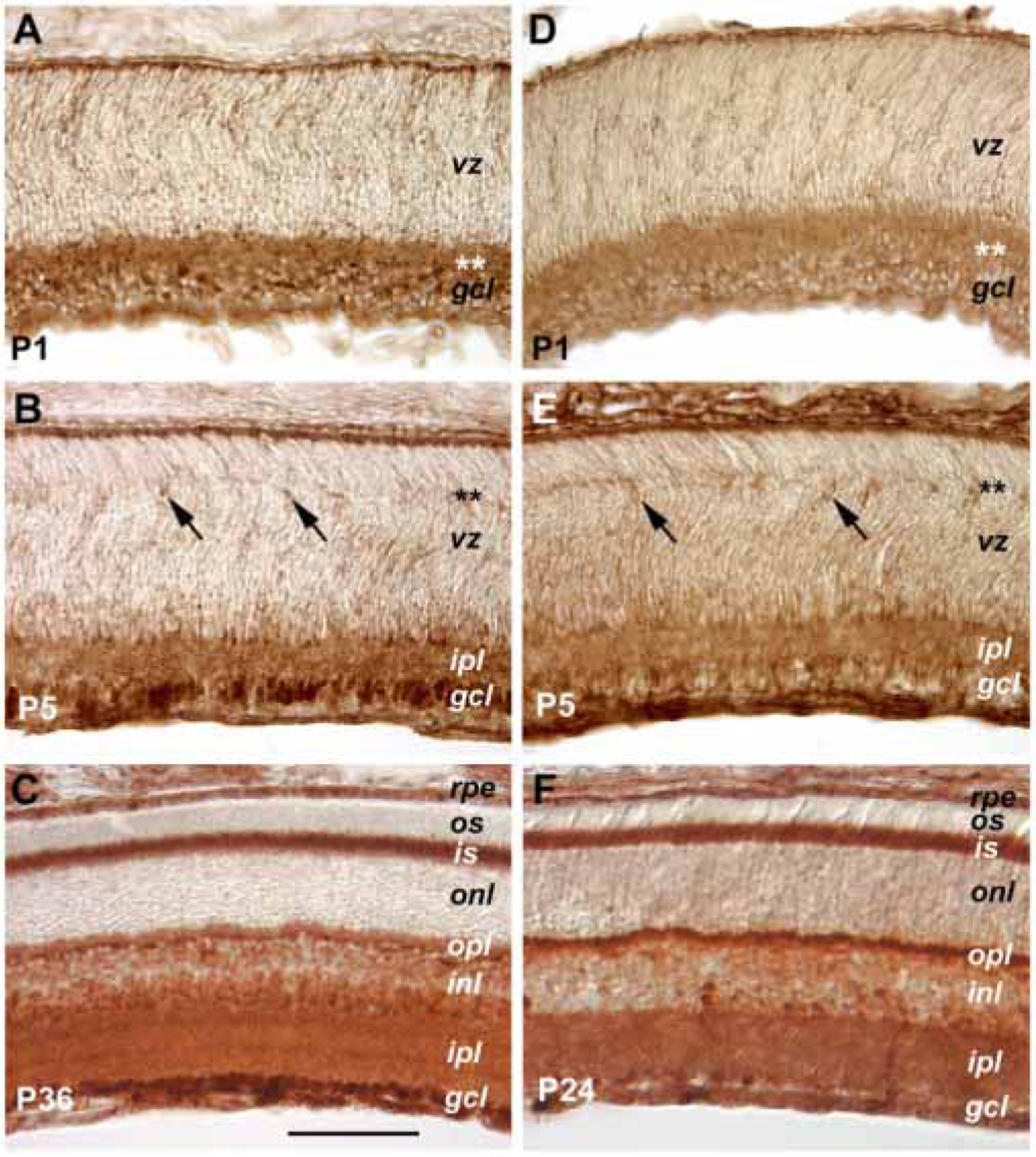
Expression of Tyk2 and Jak2 protein kinases. Cryosections of retinas from three postnatal ages immunohistochemically stained for Tyk2 (A, B, C) and Jak2 (D, E, F) are shown. The asterisks indicate the positions of the developing inner plexiform layer at P1 (A, D) and the emerging outer plexiform layer at P5 (B, E). The scale bar shown in panel C represents 100 μm for all panels. Arrows in B and E point to positively stained cells located adjacent to the forming outer plexiform layer. The meanings of the abbreviations referred to in this figure are: gcl (ganglion cell layer), inl (inner nuclear layer), ipl (inner plexiform layer), is (inner segment), onl (outer nuclear layer), opl (outer plexiform layer), os (outer segment), rpe (retinal pigmented epithelium), vz (ventricular zone).
Patterns of STAT1 and STAT3 expression:
Since both STAT1 and STAT3 transduce signals of the CNTF subfamily of cytokines, their expression patterns were examined by immunohistochemistry using specific antibodies against these two proteins. At P1, distributions of STAT1 and STAT3 mimicked those of the Jak family of kinases with predominant expression in the inner retina and lower levels in the ventricular zone (Figure 4A,G). At P5, STAT3 staining delineated the emerging outer plexiform layer and the adjacent presumptive horizontal cells (Figure 4I). By P14, both of the plexiform layers, the ganglion cell layer, and the inner nuclear layer expressed STAT1 (Figure 4D) and STAT3 (Figure 4J). Furthermore, the inner segments of the more mature photoreceptor cells also exhibited STAT1 (Figure 4D,E) and STAT3 (Figure 4J,K) staining signals. The immunostaining signals of STAT1 and STAT3 were abolished by incubation with corresponding blocking peptides prior to staining (Figure 4B,F,H,L).
Figure 4.
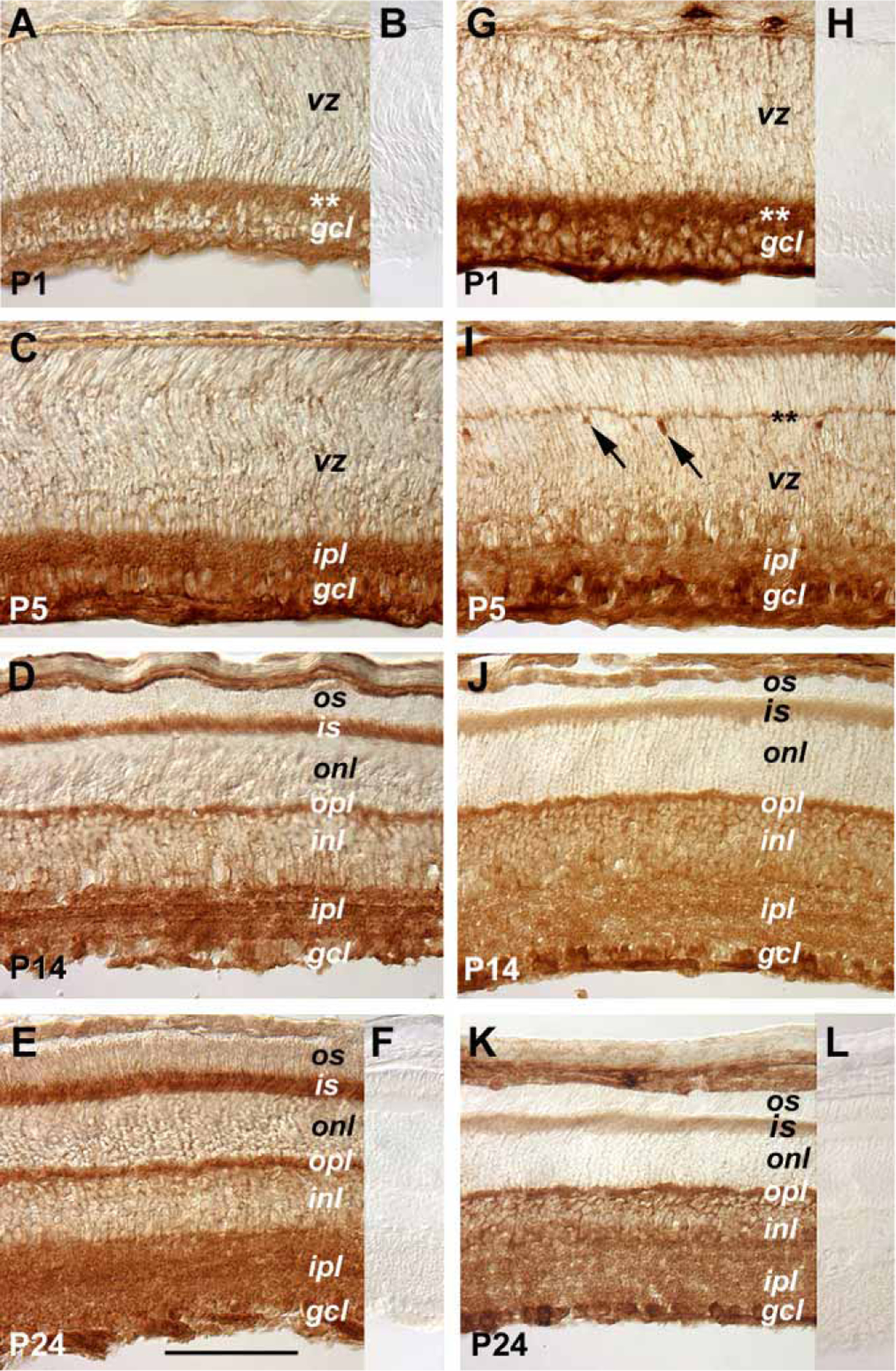
Expression of STAT1 and STAT3 signal transducers. Cryosections of retinas from distinct postnatal ages immunohistochemically stained for STAT1 (A, C, D, E) and STAT3 (G, I, J, K) are shown. Panels B, F, H, and L show staining results of primary antibodies preincubated with the antigen peptides for the corresponding sections shown in A, E, G, K, respectively. The asterisks indicate the positions of the developing inner plexiform layer at P1 (A, G) and the emerging outer plexiform layer at P5 (I). The scale bar shown in panel E represents 100 μm for all panels. Arrows in I point to strongly stained cells located adjacent to the forming outer plexiform layer. The meanings of the abbreviations referred to in this figure are: gcl (ganglion cell layer), inl (inner nuclear layer), ipl (inner plexiform layer), is (inner segment), onl (outer nuclear layer), opl (outer plexiform layer), os (outer segment), rpe (retinal pigmented epithelium), vz (ventricular zone).
Expression of the ERK kinase:
Since CNTF can activate the p42/44 ERK as well as the Jak-STAT signaling pathways, the distribution of the ERK protein kinase was characterized by immunofluorescent staining (Figure 5). At P0, ERK2 protein staining signal was distributed throughout the retina in both the ventricular zone occupied by progenitor cells and the inner retina occupied by postmitotic neurons (Figure 5A,D). By P5, there was still broad distribution of ERK2 in the retina, however, a decrease of ERK2 expression was detected in the developing outer nuclear layer (Figure 5B,E). In the mature retina (P36), robust staining of ERK2 was observed in the ganglion cell layer and the inner plexiform layer, whereas detectable staining of ERK2 was present in the inner nuclear layer, outer nuclear layer, and outer plexiform layer (Figure 5C,F). In the absence of the anti-ERK2 antibody, no signals were detected after incubation with the secondary antibody (Figure 5G–L), indicating that the weak staining signals seen in the outer nuclear layer of the mature retina were indeed due to the presence of the ERK protein.
Figure 5.
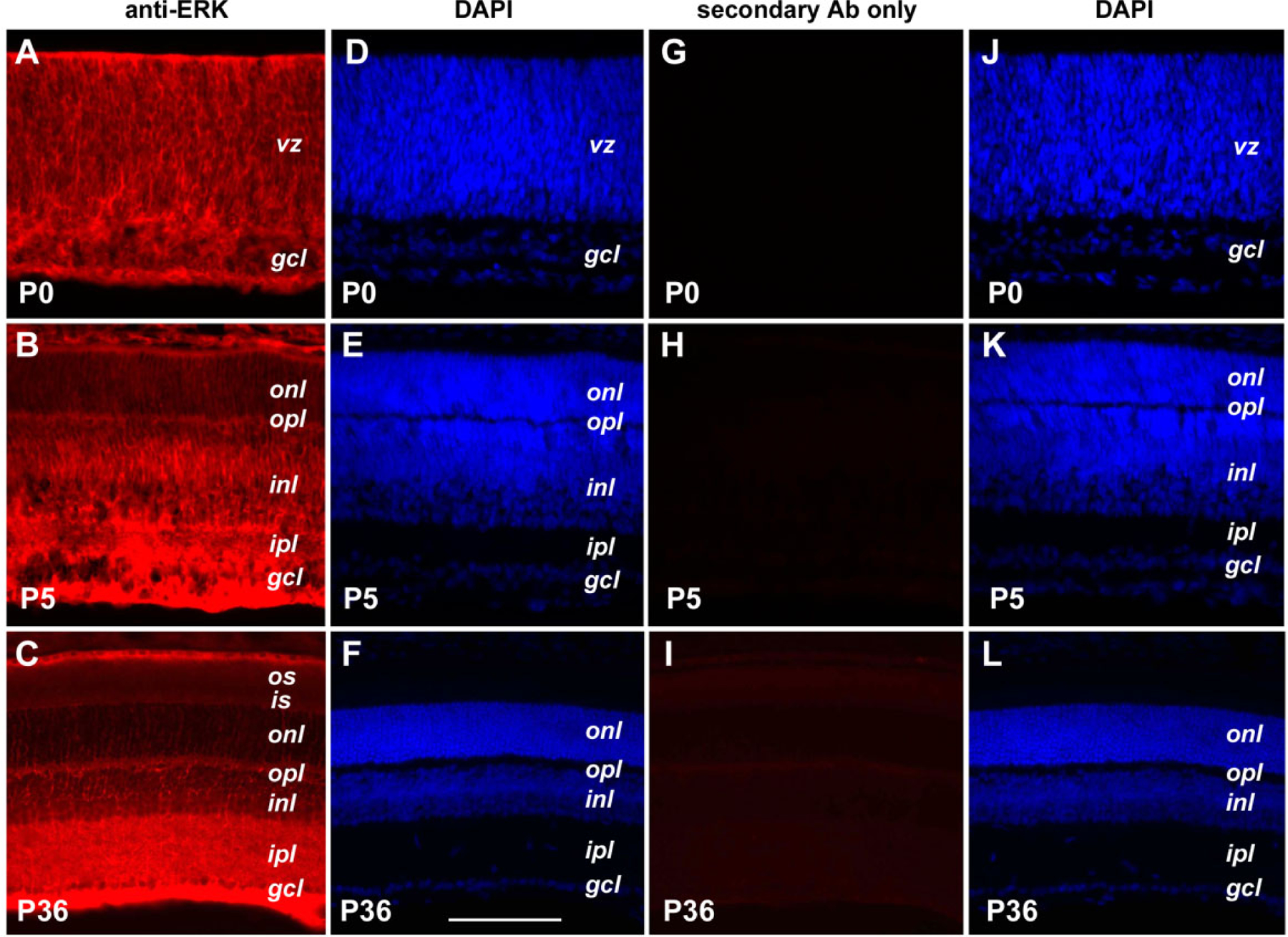
Expression of the ERK2 protein kinase. Cryosections of retinas from three postnatal ages are shown. Panels A, B, and C show sections immunofluorescently stained for ERK2. Panels D, E, and F show DAPI staining images of the same sections shown in A, C, and E, respectively. Panels G, H, and I show sections stained for secondary antibody alone; panels J, K, and L show their corresponding DAPI images. The scale bar in panel F represents 100 μm for all panels. The meanings of the abbreviations referred to in this figure are: gcl (ganglion cell layer), inl (inner nuclear layer), ipl (inner plexiform layer), is (inner segment), onl (outer nuclear layer), opl (outer plexiform layer), os (outer segment), rpe (retinal pigmented epithelium), vz (ventricular zone).
DISCUSSION
In this study, we analyzed the expression patterns of several types of cytokine signaling components as an initial step towards characterizing cytokine responsive cells in the postnatal mouse retina. Our results demonstrate that the Jak-STAT signaling pathway components, including receptor subunits gp130 and CNTFRα, intracellular protein kinases Jak2 and Tyk2, and latent signal transducing factors STAT1 and STAT3, exhibit similar expression profiles during early postnatal retinal development, whereas ERK has distinct expression patterns in the neonatal retina.
In the newborn retina, the Jak-STAT signaling pathway components that we examined are predominantly present in the inner retina, where differentiated ganglion cells and postmitotic amacrine cells reside. This distribution pattern indicates that postmitotic neurons located in the inner retina are likely to be the primary target cells for CNTF-like cytokines at this stage. Our results also revealed an increased expression of cytokine signaling components in the differentiating amacrine and horizontal cell populations within the first postnatal week. In the mature retina, the inner nuclear layer exhibits a lower expression of cytokine signaling components compared to the ganglion cell layer, while the inner and outer plexiform layers are heavily stained. These patterns of expression are consistent with the previously reported CNTF-induced signaling events in ganglion cells and Müller glia at the adult stage [33,34]. The distribution patterns of the Jak-STAT signaling components suggest that cytokine responsiveness among retinal cells may change during postnatal retinal differentiation.
The CNTF subfamily of cytokines can affect retinal differentiation during the postnatal period [5,6]. In particular, it has been demonstrated that postmitotic rod photoreceptor precursors can be influenced by CNTF treatment to suppress rhodopsin expression and turn on bipolar cell markers [5,36]. However, it remains undetermined if the observed cytokine effects are due to the direct signaling of cytokines to postmitotic rod precursors, or are instead due to indirect signaling from factors produced by differentiated neurons in response to cytokine treatments. Here, we demonstrate that, although predominant expression of cytokine signaling components has been found in the inner retina containing postmitotic neurons, a low yet significant expression of cytokine signaling molecules are present within the ventricular zone, which is occupied by proliferating progenitor cells and postmitotic rod and cone precursors. The expression of necessary cytokine signaling components in the ventricular zone raises the possibility that retinal progenitor cells as well as undifferentiated photoreceptor precursors may also directly respond to cytokine signals.
CNTFRα is a specific cytokine receptor component required for the binding and signal transduction of CNTF [22]. The expression patterns of CNTFRα have not been previously characterized in the developing mouse retina. Our immunohistochemical staining results indicate that CNTFRα protein is present in the postnatal inner retina accompanied by punctate staining signals in the ventricular zone. Thus, CNTFRα may mediate CNTF signals to postmitotic neurons and progenitor cells. Interestingly, in the mature mouse retina, the CNTFRα protein is relatively abundant in the inner and outer segments of photoreceptor cells in comparison to the ganglion cell layer, the inner and outer nuclear layer, and the two plexiform layers. This finding is consistent with the reported expression patterns of the chicken CNTFRα protein [37], which was found in all retinal cell types including the photoreceptor cells. Recent immunocytochemical studies in the mature retina of two other mammalian species have yielded strikingly similar staining patterns as we have observed in mouse. In the adult canine retina, an antibody raised against the chicken CNTFRα gives rise to strong staining signals in the inner segments of photoreceptors [38], while in the mature rat retina a commercial antibody, which is different from the one used in this study, revealed distribution of CNTFRα in the outer segments of photoreceptor cells [39]. Therefore, the expression of CNTFRα protein in the mature vertebrate retina appears to be conserved.
CNTF can activate all three members of the Jak kinase family in vitro. While Jak2-deficient mice die at embryonic day 12 [40,41] and Jak1 null mice die perinatally with defective CNTF signaling [42], Tyk2 knock out mice did not show developmental defects [43,44]. In the mouse retina, Jak1 [35], Jak2, and Tyk2 are expressed in overlapping patterns, which may result in a functional redundancy. Further studies to selectively or combinatorially eliminate their activity are necessary to delineate their individual functions in the retina. Our study also shows that STAT1 and STAT3 are expressed in similar patterns during the critical postnatal developmental period of the retina. These results complement a recent report describing the expression of STAT factors in the embryonic and adult mouse retina [45]. The individual requirements for STAT1 and STAT3 in retinal differentiation remain undetermined at presence. STAT1 null mice exhibit defects in interferon-meditated anti-viral immunity [46,47], and the potential retinal phenotypes have not been reported. Targeted disruption of STAT3 causes early embryonic lethality [48], therefore conditional disruption of the STAT3 gene in the retina is necessary to address its role in cytokine signaling and to determine if STAT1 can replace STAT3 functionally.
The cellular mechanism of CNTF signaling in the mature retina is not well understood. It has been demonstrated previously that intravitreal injection of CNTF or its analog in the adult eye results in STAT3 and ERK phosphorylation in the ganglion cells and Müller cells but not in photoreceptor cells [33,34]. Intriguingly, we have detected the expression of several cytokine signaling components in the inner or outer segments of the adult mouse photoreceptor cells by immunocytochemistry. Among these, gp130 and CNTFRα are prominently expressed in the outer segments of photoreceptors. It is unlikely that cytokine signaling proteins detected in the outer segments are due to expression by Müller glial processes, which terminate at the position of the inner segment. Therefore, it is plausible that the lack of cytokine signaling response in the mature photoreceptors maybe due to the existence of negative regulatory mechanisms for cytokine signaling in the mature retina [49]. Alternatively, cytokine signaling pathways can be directly activated in mature photoreceptors upon direct access to cytokine signals.
In contrast to expression patterns of the Jak-STAT signaling pathway components, ERK2 in the neonatal retina is ubiquitously distributed, indicating that both proliferating progenitors and postmitotic retinal neurons have access to this major signaling component when exposed to extracellular cues including cytokines. As the retina undergoes further differentiation, ERK2 expression decreases in the outer nuclear layer. It remains to be determined, however, if the Jak-STAT and the ERK signaling pathways are differentially activated by cytokines among various cell types during postnatal retinal development.
ACKNOWLEDGEMENTS
This work was in part supported by grants from the Research to Prevent Blindness Foundation, the March of Dimes Birth Defect Foundation, the Karl Kirchgessner Foundation, and the National Institute of Health (EY012270, EY014440) to Xian-Jie Yang. Kun Do Rhee is supported by a NIH predoctoral fellowship (T32 EY07026).
REFERENCES
- 1.Cepko CL, Austin CP, Yang X, Alexiades M, Ezzeddine D. Cell fate determination in the vertebrate retina. Proc Natl Acad Sci U S A 1996; 93:589–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Harris WA. Cellular diversification in the vertebrate retina. Curr Opin Genet Dev 1997; 7:651–8. [DOI] [PubMed] [Google Scholar]
- 3.Reh TA, Levine EM. Multipotential stem cells and progenitors in the vertebrate retina. J Neurobiol 1998; 36:206–20. [PubMed] [Google Scholar]
- 4.Livesey FJ, Cepko CL. Vertebrate neural cell-fate determination: lessons from the retina. Nat Rev Neurosci 2001; 2:109–18. [DOI] [PubMed] [Google Scholar]
- 5.Ezzeddine ZD, Yang X, DeChiara T, Yancopoulos G, Cepko CL. Postmitotic cells fated to become rod photoreceptors can be respecified by CNTF treatment of the retina. Development 1997; 124:1055–67. [DOI] [PubMed] [Google Scholar]
- 6.Neophytou C, Vernallis AB, Smith A, Raff MC. Muller-cell-derived leukaemia inhibitory factor arrests rod photoreceptor differentiation at a postmitotic pre-rod stage of development. Development 1997; 124:2345–54. [DOI] [PubMed] [Google Scholar]
- 7.Fuhrmann S, Kirsch M, Hofmann HD. Ciliary neurotrophic factor promotes chick photoreceptor development in vitro. Development 1995; 121:2695–706. [DOI] [PubMed] [Google Scholar]
- 8.Kirsch M, Fuhrmann S, Wiese A, Hofmann HD. CNTF exerts opposite effects on in vitro development of rat and chick photoreceptors. Neuroreport 1996; 7:697–700. [DOI] [PubMed] [Google Scholar]
- 9.Xie HQ, Adler R. Green cone opsin and rhodopsin regulation by CNTF and staurosporine in cultured chick photoreceptors. Invest Ophthalmol Vis Sci 2000; 41:4317–23. [PubMed] [Google Scholar]
- 10.Meyer-Franke A, Kaplan MR, Pfrieger FW, Barres BA. Characterization of the signaling interactions that promote the survival and growth of developing retinal ganglion cells in culture. Neuron 1995; 15:805–19. [DOI] [PubMed] [Google Scholar]
- 11.Cui Q, Lu Q, So KF, Yip HK. CNTF, not other trophic factors, promotes axonal regeneration of axotomized retinal ganglion cells in adult hamsters. Invest Ophthalmol Vis Sci 1999; 40:760–6. [PubMed] [Google Scholar]
- 12.Jo SA, Wang E, Benowitz LI. Ciliary neurotrophic factor is and axogenesis factor for retinal ganglion cells. Neuroscience 1999; 89:579–91. [DOI] [PubMed] [Google Scholar]
- 13.Weise J, Isenmann S, Klocker N, Kugler S, Hirsch S, Gravel C, Bahr M. Adenovirus-mediated expression of ciliary neurotrophic factor (CNTF) rescues axotomized rat retinal ganglion cells but does not support axonal regeneration in vivo. Neurobiol Dis 2000; 7:212–23. [DOI] [PubMed] [Google Scholar]
- 14.LaVail MM, Unoki K, Yasumura D, Matthes MT, Yancopoulos GD, Steinberg RH. Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc Natl Acad Sci U S A 1992; 89:11249–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.LaVail MM, Yasumura D, Matthes MT, Lau-Villacorta C, Unoki K, Sung CH, Steinberg RH. Protection of mouse photoreceptors by survival factors in retinal degenerations. Invest Ophthalmol Vis Sci 1998; 39:592–602. [PubMed] [Google Scholar]
- 16.Cayouette M, Behn D, Sendtner M, Lachapelle P, Gravel C. Intraocular gene transfer of ciliary neurotrophic factor prevents death and increases responsiveness of rod photoreceptors in the retinal degeneration slow mouse. J Neurosci 1998; 18:9282–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chong NH, Alexander RA, Waters L, Barnett KC, Bird AC, Luthert PJ. Repeated injections of a ciliary neurotrophic factor analogue leading to long-term photoreceptor survival in hereditary retinal degeneration. Invest Ophthalmol Vis Sci 1999; 40:1298–305. [PubMed] [Google Scholar]
- 18.Liang FQ, Dejneka NS, Cohen DR, Krasnoperova NV, Lem J, Maguire AM, Dudus L, Fisher KJ, Bennett J. AAV-mediated delivery of ciliary neurotrophic factor prolongs photoreceptor survival in the rhodopsin knockout mouse. Mol Ther 2001; 3:241–8. [DOI] [PubMed] [Google Scholar]
- 19.Bok D, Yasumura D, Matthes MT, Ruiz A, Duncan JL, Chappelow AV, Zolutukhin S, Hauswirth W, LaVail MM. Effects of adeno-associated virus-vectored ciliary neurotrophic factor on retinal structure and function in mice with a P216L rds/peripherin mutation. Exp Eye Res 2002; 74:719–35. [DOI] [PubMed] [Google Scholar]
- 20.Tao W, Wen R, Goddard MB, Sherman SD, O’Rourke PJ, Stabila PF, Bell WJ, Dean BJ, Kauper KA, Budz VA, Tsiaras WG, Acland GM, Pearce-Kelling S, Laties AM, Aguirre GD. Encapsulated cell-based delivery of CNTF reduces photoreceptor degeneration in animal models of retinitis pigmentosa. Invest Ophthalmol Vis Sci 2002; 43:3292–8. [PubMed] [Google Scholar]
- 21.Ip NY. The neurotrophins and neuropoietic cytokines: two families of growth factors acting on neural and hematopoietic cells. Ann N Y Acad Sci 1998; 840:97–106. [DOI] [PubMed] [Google Scholar]
- 22.Ip NY, McClain J, Barrezueta NX, Aldrich TH, Pan L, Li Y, Wiegand SJ, Friedman B, Davis S, Yancopoulos GD. The alpha component of the CNTF receptor is required for signaling and defines potential CNTF targets in the adult and during development. Neuron 1993; 10:89–102. [DOI] [PubMed] [Google Scholar]
- 23.Davis S, Aldrich TH, Stahl N, Pan L, Taga T, Kishimoto T, Ip NY, Yancopoulos GD. LIFR beta and gp130 as heterodimerizing signal transducers of the tripartite CNTF receptor. Science 1993; 260:1805–8. [DOI] [PubMed] [Google Scholar]
- 24.Ihle JN, Kerr IM. Jaks and Stats in signaling by the cytokine receptor superfamily. Trends Genet 1995; 11:69–74. [DOI] [PubMed] [Google Scholar]
- 25.Stahl N, Yancopoulos GD. The tripartite CNTF receptor complex: activation and signaling involves components shared with other cytokines. J Neurobiol 1994; 25:1454–66. [DOI] [PubMed] [Google Scholar]
- 26.Darnell JE Jr. STATs and gene regulation. Science 1997; 277:1630–5. [DOI] [PubMed] [Google Scholar]
- 27.Bonni A, Frank DA, Schindler C, Greenberg ME. Characterization of a pathway for ciliary neurotrophic factor signaling to the nucleus. Science 1993; 262:1575–9. [DOI] [PubMed] [Google Scholar]
- 28.Akira S, Nishio Y, Inoue M, Wang XJ, Wei S, Matsusaka T, Yoshida K, Sudo T, Naruto M, Kishimoto T. Molecular cloning of APRF, a novel IFN-stimulated gene factor 3 p91-related transcription factor involved in the gp130-mediated signaling pathway. Cell 1994; 77:63–71. [DOI] [PubMed] [Google Scholar]
- 29.Rajan P, Symes AJ, Fink JS. STAT proteins are activated by ciliary neurotrophic factor in cells of central nervous system origin. J Neurosci Res 1996; 43:403–11. [DOI] [PubMed] [Google Scholar]
- 30.Darnell JE Jr, Kerr IM, Stark GR. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994; 264:1415–21. [DOI] [PubMed] [Google Scholar]
- 31.Boulton TG, Stahl N, Yancopoulos GD. Ciliary neurotrophic factor/leukemia inhibitory factor/interleukin 6/oncostatin M family of cytokines induces tyrosine phosphorylation of a common set of proteins overlapping those induced by other cytokines and growth factors. J Biol Chem 1994; 269:11648–55. [PubMed] [Google Scholar]
- 32.Oh H, Fujio Y, Kunisada K, Hirota H, Matsui H, Kishimoto T, Yamauchi-Takihara K. Activation of phosphatidylinositol 3-kinase through glycoprotein 130 induces protein kinase B and p70 S6 kinase phosphorylation in cardiac myocytes. J Biol Chem 1998; 273:9703–10. [DOI] [PubMed] [Google Scholar]
- 33.Peterson WM, Wang Q, Tzekova R, Wiegand SJ. Ciliary neurotrophic factor and stress stimuli activate the Jak-STAT pathway in retinal neurons and glia. J Neurosci 2000; 20:4081–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wahlin KJ, Campochiaro PA, Zack DJ, Adler R. Neurotrophic factors cause activation of intracellular signaling pathways in Muller cells and other cells of the inner retina, but not photoreceptors. Invest Ophthalmol Vis Sci 2000; 41:927–36. [PubMed] [Google Scholar]
- 35.Yang X, Chung D, Cepko CL. Molecular cloning of the murine JAK1 protein tyrosine kinase and its expression in the mouse central nervous system. J Neurosci 1993; 13:3006–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Schulz-Key S, Hofmann HD, Beisenherz-Huss C, Barbisch C, Kirsch M. Ciliary neurotrophic factor as a transient negative regulator of rod development in rat retina. Invest Ophthalmol Vis Sci 2002; 43:3099–108. [PubMed] [Google Scholar]
- 37.Fuhrmann S, Kirsch M, Heller S, Rohrer H, Hofmann HD. Differential regulation of ciliary neurotrophic factor receptor-alpha expression in all major neuronal cell classes during development of the chick retina. J Comp Neurol 1998; 400:244–54. [PubMed] [Google Scholar]
- 38.Beltran WA, Zhang Q, Kijas JW, Gu D, Rohrer H, Jordan JA, Aguirre GD. Cloning, mapping, and retinal expression of the canine ciliary neurotrophic factor receptor alpha (CNTFRalpha). Invest Ophthalmol Vis Sci 2003; 44:3642–9. [DOI] [PubMed] [Google Scholar]
- 39.Valter K, Bisti S, Stone J. Location of CNTFRalpha on outer segments: evidence of the site of action of CNTF in rat retina. Brain Res 2003; 985:169–175. [DOI] [PubMed] [Google Scholar]
- 40.Neubauer H, Cumano A, Muller M, Wu H, Huffstadt U, Pfeffer K. Jak2 deficiency defines an essential developmental checkpoint in definitive hematopoiesis. Cell 1998; 93:397–409. [DOI] [PubMed] [Google Scholar]
- 41.Parganas E, Wang D, Stravopodis D, Topham DJ, Marine JC, Teglund S, Vanin EF, Bodner S, Colamonici OR, van Deursen JM, Grosveld G, Ihle JN. Jak2 is essential for signaling through a variety of cytokine receptors. Cell 1998; 93:385–95. [DOI] [PubMed] [Google Scholar]
- 42.Rodig SJ, Meraz MA, White JM, Lampe PA, Riley JK, Arthur CD, King KL, Sheehan KC, Yin L, Pennica D, Johnson EM Jr, Schreiber RD. Disruption of the Jak1 gene demonstrates obligatory and nonredundant roles of the Jaks in cytokine-induced biologic responses. Cell 1998; 93:373–83. [DOI] [PubMed] [Google Scholar]
- 43.Shimoda K, Kato K, Aoki K, Matsuda T, Miyamoto A, Shibamori M, Yamashita M, Numata A, Takase K, Kobayashi S, Shibata S, Asano Y, Gondo H, Sekiguchi K, Nakayama K, Nakayama T, Okamura T, Okamura S, Niho Y, Nakayama K. Tyk2 plays a restricted role in IFN alpha signaling, although it is required for IL-12-mediated T cell function. Immunity 2000; 13:561–71. [DOI] [PubMed] [Google Scholar]
- 44.Karaghiosoff M, Neubauer H, Lassnig C, Kovarik P, Schindler H, Pircher H, McCoy B, Bogdan C, Decker T, Brem G, Pfeffer K, Muller M. Partial impairment of cytokine responses in Tyk2-deficinet mice. Immunity 2000; 13:549–60. [DOI] [PubMed] [Google Scholar]
- 45.Zhang SS, Wei JY, Li C, Barnstable CJ, Fu XY. Expression and activation of STAT proteins during mouse retina development. Exp Eye Res 2003; 76:421–31. [DOI] [PubMed] [Google Scholar]
- 46.Durbin JE, Hackenmiller R, Simon MC, Levy DE. Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 1996; 84:443–50. [DOI] [PubMed] [Google Scholar]
- 47.Meraz MA, White JM, Sheehan KC, Bach EA, Rodig SJ, Dighe AS, Kaplan DH, Riley JK, Greenlund AC, Campbell D, Carver-Moore K, DuBois RN, Clark R, Aguet M, Schreiber RD. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell 1996; 84:431–42. [DOI] [PubMed] [Google Scholar]
- 48.Takeda K, Noguchi K, Shi W, Tanaka T, Matsumoto M, Yoshida N, Kishimoto T, Akira S. Targeted disruption of the mouse Stat3 gene leads to early embryonic lethality. Proc Natl Acad Sci U S A 1997; 94:3801–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.O’Shea JJ, Gadina M, Schreiber RD. Cytokine signaling in 2002: new surprises in the Jak/Stat pathway. Cell 2002; 109:S121–31. [DOI] [PubMed] [Google Scholar]