

Abstract
Interleukin 33 (IL33) is a cytokine found in the extracellular space (mature IL33) or in the cell nucleus (full-length IL33). Nuclear accumulation of IL33 has been reported in intestinal epithelial cells (IEC) during intestinal inflammation and cancer, but a functional role for this nuclear form remains unclear. To study the role of nuclear IL33 in IEC, we generated transgenic mice expressing full-length IL33 in the intestinal epithelium (Vfl33 mice). Expression of full-length IL33 in the epithelium resulted in accumulation of IL33 protein in the nucleus and secretion of IL33. Over-expression of full-length IL33 by IEC did not promote gut inflammation, but induced expression of genes in the IEC and lamina propria lymphocytes (LPL) that correlated negatively with genes expressed in inflammatory bowel diseases (IBD). Because the IL33 receptor ST2 is expressed by IEC, there was the potential that both the mature and full-length forms could mediate this effect. To specifically interrogate the transcriptional role of nuclear IL33, we intercrossed the Vfl33 mice with ST2- deficient mice. ST2 deficiency completely abrogated the transcriptional effects elicited by IL33 expression, suggesting that the transcriptional effects of IL33 on IEC are mediated by its mature, not its nuclear form.
Keywords: IL33, Intestinal epithelial cells, Inflammation, Inflammatory bowel disease, RNA-Seq
1. Introduction
Interleukin 33 (IL33), a member of the IL-1 family of cytokines [1], was originally described as a nuclear protein from human high endothelial venules [2]. Subsequent studies showed that IL33 acts as a cytokine, binding a heterodimeric receptor complex consisting of the ST2 receptor (ST2L) and the IL-1R accessory protein. The expression of this heterodimeric receptor has been detected on a variety of inflammatory cells [3,4], including eosinophils, basophils, macrophages, T helper 2 cells (Th2 cells), regulatory T cells, NK cells, B cells and group 2 innate lymphoid cells (ILC2) [5–7]. IL33 plays a role in the host defense against infection and has been reported to be involved in the pathogenesis of a wide range of diseases [8].
In the gastrointestinal tract, IL33 is normally expressed by stromal and immune cells, and IL33 protein has been detected in the nuclei of such cells [9,10]. IL33 is not normally expressed by epithelial cells, but recent evidence suggests that it can function as a novel epithelial “alarmin”[11], because it can be released as a danger signal by damaged, stressed, or necrotic cells to alert the immune system of a local threat. Epithelial expression of IL33 has been reported in samples from patients with ulcerative colitis [9,10,12–15] and cancer [16].
IL33 is believed to be a dual-function protein, functioning as conventional cytokine via its extracellular form (mature IL33) or as a transcriptional regulator via its nuclear form (full-length IL33). Although the molecular mechanism of release and processing of IL33 are not yet clear [17], it appears that the full-length IL33 released from injured or necrotic cells is biologically active [11,18–20], and this bioactivity can be transiently increased several-fold by limited proteolysis of the N-terminal domain (mature IL33) in inflamed tissue [21,22] before bioactivity is lost by destruction or oxidization of the C-terminal core tetrahedron structure [23]. The N-terminal domain of full-length IL33 is necessary for nuclear translocation, but it is unclear where it binds to the chromatin and whether it directly regulates gene expression in the intestinal epithelial cells (IEC). In this study, we investigate the biological properties of the full-length IL33, focusing on its transcriptional properties.
2. Materials and methods
2.1. Mouse strains
C57BL/6 mice were purchased from The Jackson laboratory (Bar Harbor, ME). ST2−/− mice were generated in our laboratory as described by He et al [16]. Mice were maintained under specific pathogen-free conditions. All experiments involving animals were performed following guidelines of the Animal Care and Use Committee of the Icahn School of Medicine at Mount Sinai
2.2. Ethics approval
All animal experiments in this study were approved by the Institutional Animal Care and Use Committee of Icahn School of Medicine at Mount Sinai, and were performed in accordance with the approved guidelines for animal experimentation at the Icahn School of Medicine at Mount Sinai (IACUC-2015–0004).
2.3. Generation of transgenic mice expressing IL33 in the intestinal epithelium
The cDNA of IL33 full-length form was cloned into a pBS-Villin vector that contained a 9 kb segment of the mouse villin promoter [24]. The pBS-Villin/IL33 plasmid was verified by sequencing, and the transgene was isolated from the plasmid by restriction enzyme digestion and gel purification. To generate transgenic mice, the transgene was microinjected into C57BL/6 mouse eggs. Identification of the transgenic Vfl33 mice was done by PCR amplification using the following primers: 5′- ggctgtgatagcacacagga-3′ and 5′ttcgcctgcggtgctgctgaac −3′.
2.4. Enzyme-linked immunosorbent assay
Small pieces of small intestine or colon (5 mm of mid-part) were isolated, rinsed in PBS, weighed, and cultured overnight in 12-well tissue culture plates (Costar) in 1000 μl complete DMEM at 37 °C in an atmosphere containing 5% CO2. After centrifugation to pellet debris, culture supernatants were transferred to fresh tubes and stored at −80 °C. IL33 was quantified in the supernatant of intestinal explant cultures from Vfl33 and WT mice by enzyme-linked immunosorbent assay (ELISA) according to standard manufacturer’s recommendations (eBioscience) and the results were normalized to the weight of the intestinal explant.
2.5. Reverse-transcription polymerase chain reaction
Total RNA from tissues cells was extracted using the RNeasy mini Kit (Qiagen) according to the manufacturer’s instructions. Complementary DNA (cDNA) was generated with Superscript III (Invitrogen). Quantitative PCR was performed using SYBR Green Dye (Roche) on the 7500 Real Time System (Applied Biosystems) machine. Thermal cycling conditions used were as follows: 50 °C for 2 min and 95 °C for 10 min, 40 cycles of 95 °C for 15 s, 60 °C for 1 min, followed by dissociation stage. Results were normalized to the housekeeping gene Ubiquitin. Relative expression levels were calculated as 2(Ct(Ubiquitin)-Ct(gene)). Primers were designed using Primer3Plus software [26].
2.6. Histology and immunofluorescence staining
Tissues were dissected, fixed in 10% phosphate-buffered formalin, and then processed for paraffin sections. Five-micrometer sections were stained with hematoxylin and eosin (H&E) for histological analyses. For immunofluorescence staining, five-micrometer sections were dewaxed by immersion in xylene (twice for 5 min each time) and hydrated by serial immersion in 100%, 90%, 80%, and 70% ethanol and PBS. Antigen retrieval was performed by microwaving sections for 20 min in Target Retrieval Solution (DAKO). Sections were washed with PBS (twice for 10 min each time), and blocking buffer (10% BSA in TBS) was added for 1 h. Sections were incubated with primary antibody in blocking buffer overnight at 4 °C. After washing, conjugated secondary Abs were added and then incubated for 35 min. Cell nuclei were stained using 4′,6-Diamidino-2-Phenylindole (DAPI). The slides were next washed and mounted with Fluoromount-G (Southern Biotech). Images were captured using a Nikon fluorescence microscope. Colocalization was performed with ImageJ and the colocalization finder plug-in.
2.7. Western blot analysis
Intestine were opened longitudinally and thoroughly washed in PBS and then homogenized in ice-cold lysis buffer (100 mM Tris-HCl, pH 6.8, 4% SDS, 20% glycerol, 200 mM β-mercaptoethanol, 1 mM phenylmethylsulfonyl fluoride and 1 μg/mL aprotinin). Lysates were then centrifuged at 12 000g for 15 min to remove insoluble cell debris. Protein content was quantified using the Bio-Rad protein assay (Bio-Rad) and 15 μg of protein was separated by SDS-PAGE and transferred onto polyvinylidene difluoride membranes. The membrane was blocked for 1 h in buffer (TBS, 5% milk, 0.1% Tween 20) and then incubated with the primary antibody (Rat Anti-Mouse IL-33 Monoclonal Antibody) (Catalog # MAB3626, R&D Systems) in dilution buffer (TBS, 5% bovine serum albumin, 0.1% Tween 20) overnight at 4 °C. The membrane was then washed three times with wash buffer (TBS, 0.1% Tween 20), incubated with Rat IgG HRP-conjugated Antibody (Catalog # HAF005, R&D Systems) and visualized with the enhanced chemiluminescent detection system (Amersham Biosciences).
2.8. Isolation of IEC and LPL
Intestines were opened longitudinally and thoroughly washed in PBS. The intestine was then incubated in 30 ml PBS containing 1 mM dithiothreitol (DTT) on room temperature for 15 min. The intestine was then removed and briefly washed in PBS and incubated in 25 ml PBS containing 5.2 mM ethylenediaminetetraacetic acid (EDTA) at 4 °C at 200 RPM for 30 min. The cells were then subjected to 30 s vigorous shaking and the tissue removed. The cells were then centrifuged at 1000g for 5 min at 4 °C, washed in PBS containing 10% FBS and spun for a further 5 min at 4 °C at 1000g. These cells constituted the IEC population. To isolate lamina propria lymphocytes (LPL), the remaining tissues were performed as described before [16].
2.9. Cell sorting
Cell pellets were first pre-incubated with anti-mouse CD16/CD32 for blockade of Fc γ receptors, then were washed and incubated for 30 min with fluorescent conjugated antibodies against CD45 and EpCAM in a total volume of 500 μl PBS containing 2 mM EDTA and 2% (vol/vol) fetal bovine serum. DAPI (Invitrogen) was used to distinguish live cells from dead cells during cell sorting. Stained IECs (DAPI−CD45−Epcam+) and LPL (DAPI−CD45+) were purified with a MoFlo Astrios cell sorter (DakoCytomation). Cells were >95% pure after sorting.
2.10. Microarray analysis
Total RNA from the sorted intestinal CD45+ cells from WT and Vfl33 mice was extract using RNeasy Micro Kit (Qiagen). Microarrays were done and analyzed as described before [27,28]. In order to analyses the pathways that the differentially expressed genes are involved in, KEGG pathway enrichment analyses were performed using ClueGo [29,30]. A cut-off of 0.4 was set for kappa score and terms including at least 3 genes were retrieved.
2.11. RNA-seq
Following cell sorting into Trizol LS reagent, samples were shipped on dry ice to the Center for Functional Genomics and the Microarray & HT Sequencing Core Facility at the University at Albany (Rensselaer). Total RNA from sorted cells (3 −9×105 cells) was extracted using the RNeasy micro Kit (Qiagen) with an on-column DNAse digestion step included according to the manufacturer’s instructions. RNA quality was assessed using the Nanodrop (Thermo Scientific) and Bioanalyzer Total RNA Pico assay (Agilent). Total RNA with a RNA integrity number (RIN) value of 7 or greater was deemed of good quality to perform the subsequent protocols. 100 pg of total RNA was oligo-dT primed using the SMART-Seq v4 Ultra Low Input RNA Kit (Clontech) and resulting the cDNA was amplified using 15 cycles of PCR. The double stranded cDNA (dscDNA) was purified using AMPure XP magnetic beads and assessed for quality using the Qubit dsDNA HS assay and an Agilent Bioanalyzer high sensitivity dscDNA chip (expected size ~600 bp–9000 bp). The Illumina Nextera XT kit was used for library preparation wherein 125 pg dscDNA was fragmented and adaptor sequences added to the ends of fragments following which 12 cycles of PCR amplification was performed. The DNA library was purified using AMPure XP magnetic beads and final library assessed using Qubit dsDNA HS assay for concentration and an Agilent Bioanalyzer high sensitivity DNA assay for size (expected range ~ 600–740 bp). Library quantitation was also done using a NEBNext Library Quant kit for Illumina. Each library was then diluted to 4 nM, pooled and denatured as per standard Illumina protocols to generate a denatured 20 pM pool. A single end 75 bp sequencing was performed on the Illumina Nextseq 500 by loading 1.8 pM library with 5% PhiX on to a 75 cycle high output flow cell. The RNAseq data was checked for quality using the Illumina FastQC algorithm on Basespace.
2.12. Transcriptome analysis
RNA-Seq data from IECs was mapped to the mouse reference genome (UCSC/mm10) using Tophat version 2.1.0[31]. Gene-level sequence counts were extracted for all annotated protein-coding genes using htseq-count version 0.6.1 [32] by taking the strict intersection between reads and the transcript models associated with each gene. Raw count data were filtered to remove low expressed genes with less than five counts in any sample. Differentially expressed genes between groups were analyzed using Bioconductor EdgeR package version 3.10.2 Bioconductor/R [33,34]. Statistically significant differentially expressed genes between groups (Q < 0.05) were selected in genewise log-likelihood ratio tests that were corrected for multiple testing by Benjamini and Hochberg FDR.
2.13. NextBio
Meta-analysis was conducted by NextBio (www.nextbio.com) [35]. The gene list (Vfl33 vs WT) from LPL Microarray results and IEC RNA-seq results were used as input to query a collection of individual biosets in NextBio database. The NextBio application “disease atlas” was used. The association score based on statistical significance across different diseases (100 to the most significant inside the diseases).
2.14. Statistics
Differences between groups were analyzed with nonparametric Mann-Whitney test. For the comparison of more than two groups a one-way ANOVA followed by a Bonferroni multiple comparison tests were performed. All statistical analyses were performed using GraphPad Prism 5 software.
3. Results
3.1. Generation of transgenic mice expressing nuclear IL33 in the epithelium
To investigate the function of full-length IL33 in epithelial cells, we cloned the cDNA encoding the full-length form of IL33 downstream of the villin promoter (Fig. 1A). The transgene was injected into mouse eggs and 3 transgenic lines were derived. These animals are referred to as Vfl33 mice (Fig. 1A). The Vfl33 mice were healthy and reproduced normally. To examine IL33 expression and select a line for further studies, we extracted RNA from the small and large intestine of control and transgenic mice. As expected, we detected increased expression of IL33 mRNA in the small intestine and large intestine in the Vfl33 transgenic mice compared with their littermate control WT mice (Fig. 1B). To examine expression of IL33 protein, we performed ELISA in the gut extracts and found that IL33 production in the supernatant was elevated in the intestine of transgenic mice compared to WT mice (Fig. 1C). Finally, we examined the cellular expression of IL33. Because we expressed the full-length form of IL33, we expected that it should be located in the nucleus rather than the cytoplasm. Immunostaining of intestinal sections showed that IL33 immunoreactivity was indeed detected in the nucleus of transgenic, but not control intestinal epithelial cells (Fig. 1D). Similar qualitative results were obtained by analysis of animals in the 3 transgenic lines and line 2 was selected for further analysis.
Fig. 1.

Generation of transgenic mice expressing full-length IL33 in the gut epithelium. (A) Scheme for generation of Vfl33 mice. A transgene encoding the IL33 full-length form under the control of the murine villin promoter (9kb) was used to generate Vfl33 mice. (B) Relative expression levels of IL33 mRNA were analyzed by qPCR in the small intestine (SI) and colon of wild-type (WT) and Vfl33 mice. Data were normalized to the expression levels of the Ubiquitin transcript. Means ± s.e.m., n = 6 per group. ***P < 0.001, one-way ANOVA. (C) Enzyme linked immunosorbent assay of IL33 in the gut explants from WT and Vfl33 mice. Data were normalized to the weight of the intestine explant. Means ± s.e.m., n =4 per group. ***P < 0.001, one-way ANOVA. (D) Immunofluorescence staining for IL33 (red) in the gut of WT and Vfl33 mice. Notice that transgenic expression of IL33 in the nucleus of intestinal epithelial cells (IEC) in Vfl33 mice. IEC stained with an anti-Epcam antibody (green). Scale bars, 50 μm. Inset shows higher magnification of the boxed area. (E) Western blot analysis of IL33 expression in the gut of WT and Vfl33 mice. The Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) protein levels were used as protein loading controls. The expected molecular sizes for the full-length and mature forms of IL33 are 37 and 18 kDa, respectively.
Although the molecular mechanism of release and processing of IL33 are not yet clear [17], the studies done so far show that the full-length IL33 released from injured or necrotic cells is biologically active [11,18–20], and that its bioactivity can be transiently increased several-fold by limited proteolysis of the N-terminal domain (mature-IL33) in the tissue [21,22]. We next examined whether mature IL33 could be detected in the intestine of Vfl33 mice. To do so, we performed Western blot to analyze the different molecular species of IL33 in the intestine. We found that the predominant form of IL33 in the extracts was the full-length form (37kD band), but that a 18kD (mature IL33) was also present (Fig. 1E). Together the results indicate that the transgenic IL33 was correctly targeted to IEC of both small and large intestine, and that it accumulated in the nucleus. In addition they suggest that the processed IL33 mature form was also produced, although at lower concentration.
3.2. Over-expression of full-length IL33 by IEC does not promote intestinal inflammation
To examine the potential impact of epithelial-specific IL33 on the intestinal inflammation, we performed histological analysis of the gut in Vfl33 mice. We found that there was no inflammation in the gut of transgenic mice (Fig. 2A). Consistent with this, FACS analysis of LPL showed that there were no differences in the total number of CD45+ cells between Vfl33 and WT mice (Fig. 2B). The results suggested that over-expression of full-length IL33 by IEC did not promote gut inflammation.
Fig. 2.
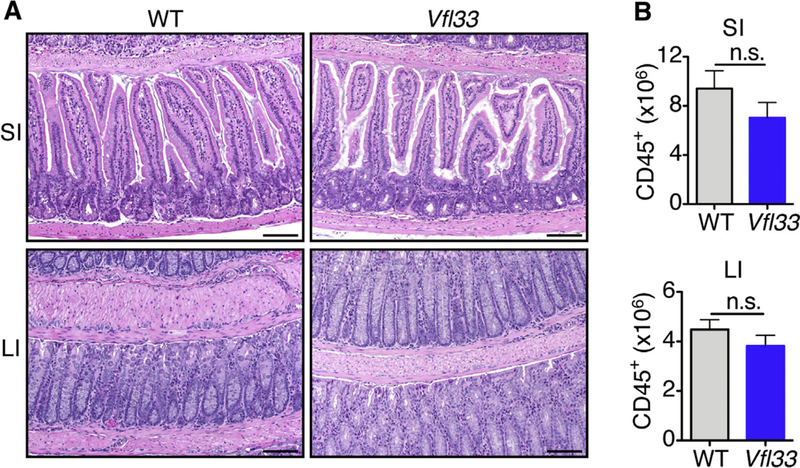
Over-expression of full-length IL33 by IEC does not promote gut inflammation. (A) Representative H&E-stained intestinal sections of Vfl33 mice. Scale bars, 50 μm. (B) Total CD45+ cells in the gut of Vfl33 and WT mice (n = 6 mice/group). N.s, non significant; by Mann-Whitney test.
3.3. Epithelial-derived IL33 triggers gene expression in lamina propria leukocytes
To examine if IL33 expression affected intestinal immune cells, we sorted lamina propria CD45+ cells from both groups to perform microarray analysis. The results show that 105 genes were upregulated and 58 genes were downregulated in Vfl33 mice compared to WT mice (Fig. 3A &3B). It has been reported that IL33 can enhance the primary differentiation of CD4+ Th1, Th2, and Treg [36]. We found that expression of full-length IL33 was associated with increased Th2 immune response in the gut, consistent with a signature of up-regulated Th2 transcription factor (Gata3) and Th2 cytokines (Il4 and Il13) by quantitative PCR (q-PCR) (Fig. 3C). In addition, expression of retnla and retnlb, which are induced in Th2 environment [37], was also up-regulated in the gut of Vfl33 mice (Fig. 3C).
Fig. 3.

Epithelial-derived IL33 triggers gene expression in lamina propria leukocytes. (A-B) Leukocytes (CD45+) isolated from Vfl33 and WT LPL were analyzed by microarray (n =4–5/group). (A) Quantile-normalized expression values were analyzed using a paired design and filtered for Q < 0.05 and −1.2 > fold change > 1.2. Z score–normalized data were subjected to hierarchical clustering. (B) Plot of logFC (log fold change) versus mean expression of all detected transcripts (gray) and significant genes (red). (C) Quantitative PCR analysis of the selected Th2 immune response associated genes in the gut of Vfl33 and WT mice (n =5–7/group). Data were normalized to the expression levels of the Ubiquitin transcript. Means ± s.e.m., *P < 0.05, **P < 0.01, Mann-Whitney test. (D) The pathway enriched among total 163 differential expressed genes in the Vfl33 versus WT LPL by KEGG analysis. (E) The mRNA expression profile of Vfl33 LPL vs WT LPL correlated with various digestive system diseases by using NextBio meta-analysis program. The correlated diseases are shown with the corresponding significant scores as well as the number of biosets within each disease. Columns are colored according to correlation with query: positive correlation (red) and negative correlation (blue).
Pathway analysis of differentially expressed mRNAs is designed to provide insight into the cell pathways associated with these genes. Pathway analysis of 163 differential expressed genes in CD45+ cells of Vfl33 mice showed that “inflammatory bowel disease” was the top pathway (Fig. 3D), suggesting that epithelium expressed IL33 could have a role in the IBD. In addition, we used the gene list (Vfl33 vs WT) from the LPL microarray as input to query a collection of individual biosets in NextBio database. The NextBio application “disease atlas”, which focuses on digestive system disease, was used for analysis. We found that differentially expressed Vfl33 genes negatively correlated with genes expressed in inflammatory bowel disease, Crohn’s disease, enteritis of small intestine and colitis (Fig. 3E). Together, the pathway analyses and the NextBio meta-analysis suggest that epithelium-derived IL33 regulates gene expression, and that many of these differentially expressed genes negatively correlate with genes expressed in IBD.
3.4. Full-length IL33 regulates gene expression in the epithelium
To investigate the function of IL33 in the epithelium, we sorted intestinal epithelial cells from Vfl33 mice and WT mice, extracted mRNA and preformed RNA sequencing. The results showed that 103 genes were upregulated and 52 genes were downregulated in transgenic IEC compared to controls (Fig. 4A). Since the IL33 regulated genes in the lamina propria leukocytes were associated with IBD, we asked next if the differentially expressed genes of the intestinal epithelial cells also correlated with IBD. To do so we filtered the differentially expressed genes in the IEC RNA-seq results using the disease category locator of the NextBio software. The results indicated that the differentially expressed genes shared high similarity with genes associated with digestive system disease (Fig. 4B). Notably, differentially expressed genes were negatively correlated with intestinal inflammatory diseases such as enteritis of small intestine, IBD, colitis and Crohn’s disease (Fig. 4B). Taken together, the transcriptomic analyses of both epithelial cells and leukocytes suggest a protective role of epithelial-derived IL33 in intestinal inflammation.
Fig. 4.

Differential expression of epithelial genes triggered by IL33 correlates with a subset of genes involved in diseases of the digestive system. (A) Vfl33 and WT IEC were analyzed by RNA-Seq/edgeR using a paired design (n = 3–4/group). Plot of logFC (log fold change) versus logCPM (log counts per million) of all detected transcripts. Points are colored according to expression status: non-significant genes (gray) and significant genes (155 genes; Q < 0.05 and −2 > fold change > 2; red). (B) NextBio meta-analysis showed that the mRNA expression profile of Vfl33 IEC vs WT IEC negatively correlated with various digestive system diseases. The correlated diseases are shown with the corresponding significant scores as well as the number of biosets within each disease. Columns are colored according to correlation with query: positive correlation (red) and negative correlation (blue).
3.5. IL33–induced gene regulation in IEC is not dependent on its nuclear localization
IL33 is hypothesized to be a dual-function protein, functioning as a conventional cytokine and/or as a transcriptional regulator. To investigate if the nuclear form of IL33 directly regulated transcription, we expressed the IL33 full-length form and simultaneously deleted expression of its receptor ST2. To do so, we crossed IL33 transgenic mice (Vfl33) with ST2 deficient mice (ST2−/−)[16] to generate Vfl33 ST2−/− mice. First, we confirmed that shutdown the IL33/ST2 signaling did not change the nuclear location of full–length IL33 in Vfl33 ST2−/− mice (Fig. 5A). We then sorted IEC from WT, Vfl33, ST2−/− and Vfl33 ST2−/− mice, extracted RNA and preformed RNA-seq. To our surprise, WT, ST2−/− and Vfl33 ST2−/− clustered together, and apart from Vfl33 by principal component analysis (Fig. 5B). Further analysis shown that with exception of IL33, there were no differences in gene expression between IEC from Vfl33 ST2−/− and those from WT mice (Fig. 5C). In addition, there were no differences in gene expression between IEC of WT and ST2−/− mice (Fig. 5B). As shown in Fig. 5D, none of the differentially expressed genes between Vfl33 and WT mice were differentially expressed in Vfl33 ST2−/− mice. Thus, the RNA-seq analyses do not support a role of nuclear IL33 in the regulation of gene expression in the IEC. All together, the results suggested that IL33 regulates gene expression in the epithelium via its interaction with epithelial-expressed ST2.
Fig. 5.

Genetic deletion of ST2 in Vfl33 mice abrogates IL33 induced gene expression in IEC. (A) Immunofluorescence staining for IL33 (red) in the intestine of Vfl33 ST2−/− mice. Scale bars, 50 μm. (B) Principal component analysis of RNAseq expression data from all biological replicates of IEC from WT, Vfl33, ST2−/− and Vfl33 ST2−/− (n = 3–4/group). (C) Vfl33 ST2−/− and WT IEC were analyzed by RNA-Seq/edgeR using a paired design (n =3–4/group). Plot of logFC (log fold change) versus logCPM (log counts per million) of all detected transcripts. Points are colored according to expression status: non-significant genes (gray) and significant gene (Il33 gene; Q < 0.05; red). (D) Heat map depiction of all differentially expressed genes between WT and Vfl33 in all groups (WT, Vfl33, ST2−/− and Vfl33 ST2−/−). Red indicates increased and green indicates decreased expression in Vfl33 IEC compared with WT IEC.
4. Discussion
IL33 has been detected in the nucleus of epithelial cells in intestinal inflammation and cancer, but whether this nuclear form of IL33 contributes to transcriptional regulation, is a matter of debate. In this study, we generated transgenic mice to investigate the role of IL33 on IEC (Fig. 1). We found that the bulk of the IL33 protein present in intestinal extracts of these animals corresponded to the full-length form, but also detected the mature, processed form. Immunostaining experiments documented the nuclear accumulation of IL33 in the IEC. Comparison of gene expression profiles of sorted IEC from WT and Vfl33 mice showed a marked difference, clearly supporting a role for IL33 in regulating IEC gene expression (Fig. 3A). The precise effect of epithelial-derived IL33 to the gut inflammatory conditions has remained unclear. Experimental data obtained using different animal models of intestinal inflammation have produced conflicting results [38,39], with IL33 having both pro-inflammatory and anti-inflammatory effects. Previous studies done by our group have shown that expression of mature IL33 by IEC does not promote gut inflammation [16], suggesting that epithelium derived mature IL33 does not display pro-inflammatory properties. In line with this observation, over-expression of full-length IL33 by IEC reported here also does not promote gut inflammation (Fig. 2). Meta-analysis of genes differentially expressed in Vfl33 mice in the IEC negatively correlated with intestinal inflammatory diseases such as enteritis of small intestine, IBD, colitis and Crohn’s disease (Fig. 4), suggesting a protective role of epithelial-derived IL33 in the intestinal inflammation.
We suggest that the expression of IL33 by IEC, contributes a protective effect on the acute inflammation that ensues from damage of the gut epithelium. Therefore, IL33 production may serve as a counterregulatory response to inflammation. The protective effects could derive from a direct cytokine role of IL33 on IEC, leading to expression of genes that are negatively associated with inflammation as described above. This protective ability may not be only related to the effects of IL33 signaling on IEC. Release of the mature IL33 could also affect immune cells. Indeed, our microarray analyses indicate that epithelium-derived IL33 differentially regulated expression of several genes in lamina propria leukocytes. One of the most upregulated genes in lamina propria leukocytes was the IL33 receptor ST2. Previous work from our lab has shown that epithelial-derived IL33 induces expansion of ST2+ Treg cells in the intestine [16]. Tregs induced by IL33 signaling could ameliorate inflammation [40]. Other genes to prominently expressed by the Vfl33 leukocytes were those involved in the Th2 response in the gut (Fig. 3). Indeed, induction of Th2 responses may be of benefit when mucosal inflammation is mediated through Th1 or Th17 pathways [41]. These results add to the clinical evidence that administration of helminths as a therapeutic option in IBD, as helminths may exert their anti-inflammatory effect through the induction of specific Th2 cytokines [42,43]. Epithelial-derived IL33 could directly act on Th17 cells to help them to acquire immunosuppressive phenotype [44]. In addition, IL33 induced M2 macrophage has been reported in the contribution of attenuation of colitis [45,46]. Th2 cytokines also appear to influence resolution of inflammation by inducing polarization of macrophages to M2 macrophages [45]. Finally, the genes regulated in lamina propria leukocytes by IL33 negatively correlated with genes associated with intestinal inflammatory diseases (Fig. 3).
Most of the studies on the function of the nuclear function of IL33 have been performed in endothelial cells [47]. It has been shown that nuclear IL33 can bind to the acidic pocket formed by histones H2A and H2B [48,49], and to bind to the transcriptional repressor histone methyltransferase [50], but it is unclear if this physical association has functional properties. Some studies have reported that IL33 can affect nuclear factor-kB (NF-kB) activity in a gene dependent manner. Binding of IL33 to the NF-kB p65 subunit in the nucleus reduces p65-triggered gene expression to dampen the production of proinflammatory cytokines [51]. However, others have reported that nuclear IL33 could bind to the promoter region of p65, positively regulating its transcriptional activity in endothelial cells [52]. Thus, while there is evidence for binding of IL33 to nuclear proteins in endothelial cells, its ability to regulate gene transcription remains controversial.
Here we provide direct evidence that IL33 can regulate gene expression in the epithelium. Under these conditions IL33 could affect gene regulation by acting directly in the nucleus, or by autocrine regulation via ST2, which is expressed by epithelial cells [14], or both. To discriminate among these possibilities, we forced IL33 expression in epithelial cells and ablated expression of ST2 in all cells, including epithelial cells. Surprisingly, the only gene to be upregulated in these IEC in these conditions was the transgenic full-length, nuclear form of IL33. The results indicate that IL33 induced transcriptional changes via autocrine activation of ST2 and suggest that there are no transcriptional activities associated with its nuclear form in IEC (Fig. 5). We cannot formally rule out that the nuclear form of IL33 controls microRNA transcription, because our methods for RNA preparation did not enrich for microRNAs. The impact of any increased miRNA expression, however, would be in the levels of mRNA transcripts, but no changes were detected when the transcriptomes of WT and Vfl33 ST2−/− IEC were compared. We would thus posit that there are no transcriptional activities associated with the nuclear form of IL33 in IEC. Our transcriptional studies in IEC corroborate proteomic studies done by Girard’s group using endothelial cells [53]. Using RNA silencing strategies they showed that in endothelial cells IL33 acts as a cytokine, not as a nuclear factor, regulating gene expression [53]. The actual function of the nuclear form may be related to control the availability of the mature form in circulation. Elegant studies done by Bessa et al. [54] show that in the absence of nuclear localization, IL33 is released into the circulation leading to widespread non-resolving inflammation that culminates in the death of the animal [54]. Therefore, the main purpose of IL33 nuclear localization and chromatin association may be the regulation of its potent extracellular (mature IL33) cytokine activity [53], not control of transcription, as demonstrated here.
Some of cytokines in the IL1 family must undergo enzymatic processing, involving cleavage of the inactive full-length form into a functional cytokine [55]. Unlike IL1β and IL18, IL33 has biological activity even as a full-length form, which is similar to another IL1 family member IL1α [56,57]. Mature form of IL33 has significantly increased biological activity compared to its full-length form [21,22]. However, keeping IL33 in the nuclear of producing cells is essential for avoiding unwanted cytokine activity [58]. The mechanism of regulation through nuclear localization/sequestration of the protein is not unique to IL33, nuclear IL1α also can act as a sink to prevent inflammation [59]. Thus, the evolutionary conserved nuclear localization of the IL-1 family of precursor proteins may be important for prevention of inflammation. The mechanisms that regulate IL33 secretion from the cell and the factors that regulate its expression remain unanswered questions in IL33 biology.
5. Conclusion
In summary, expression of full-length IL33 in the epithelium resulted in accumulation of IL33 protein in the nucleus and secretion of IL33. Accordingly, expression of full-length IL33 in the epithelium promoted expression of genes in the neighboring lamina propria leukocytes and in epithelial cells. The gene program activated by IL33 in these cells suggests that this molecule has a role in resolution of the inflammatory response. The transcriptional program elicited by expression of full length IL33 was promoted by its mature, processed, form via binding to its receptor ST2, not by its nuclear form in intestinal epithelial cells.
Acknowledgments
We thank all members of the Lira Lab for their support. We thank Claudia Canasto-Chibuque for colony maintenance, Chao Yang for Western Blot and Dr. Kevin Kelley and the Mouse Genetics Shared Research Facility for assistance in generation of transgenic mice.
Funding
This work was supported in part by the SUCCESS (Sinai Ulcerative Colitis Clinical, Experimental and System Studies) grant from the Bacchetta Foundation. Funds for the generation of the Vfl33 mice were provided by Janssen Pharmaceuticals. Lili Chen received a Research Fellowship Award (327362) from the Crohn’s & Colitis Foundation of America (CCFA). The funding body had no influence on the study design, data collection, analysis and interpretation, on the writing and the decision to submit the article for publication.
Footnotes
Conflict of interest statement
The authors declare no competing financial interests.
References
- [1].Schmitz J, Owyang A, Oldham E, Song YL, Murphy E, McClanahan TK, Zurawski G, Moshrefi M, Qin JZ, Li XX, Gorman DM, Bazan JF, Kastelein RA, IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines, Immunity 23 (5) (2005) 479–490. [DOI] [PubMed] [Google Scholar]
- [2].Baekkevold ES, Roussigne M, Yamanaka T, Johansen FE, Jahnsen FL, Amalric F, Brandtzaeg P, Erard M, Haraldsen G, Girard JP, Molecular characterization of NF-HEV, a nuclear factor preferentially expressed in human high endothelial venules, Am. J. Pathol 163 (1) (2003) 69–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Chackerian AA, Oldham ER, Murphy EE, Schmitz J, Pflanz S, Kastelein RA, IL-1 receptor accessory protein and ST2 comprise the IL-33 receptor complex, J. Immunol 179 (4) (2007) 2551–2555. [DOI] [PubMed] [Google Scholar]
- [4].Greenfeder SA, Nunes P, Kwee L, Labow M, Chizzonite RA, Ju G, Molecular cloning and characterization of a second subunit of the interleukin 1 receptor complex, J. Biol. Chem 270 (23) (1995) 13757–13765. [DOI] [PubMed] [Google Scholar]
- [5].Ohno T, Morita H, Arae K, Matsumoto K, Nakae S, Interleukin-33 in allergy, Allergy 67 (10) (2012) 1203–1214. [DOI] [PubMed] [Google Scholar]
- [6].Mirchandani AS, Salmond RJ, Liew FY, Interleukin-33 and the function of innate lymphoid cells, Trends Immunol. 33 (8) (2012) 389–396. [DOI] [PubMed] [Google Scholar]
- [7].Lu J, Kang J, Zhang C, Zhang X, The role of IL-33/ST2L signals in the immune cells, Immunol. Lett 164 (1) (2015) 11–17. [DOI] [PubMed] [Google Scholar]
- [8].Liew FY, Pitman NI, McInnes IB, Disease-associated functions of IL-33: the new kid in the IL-1 family, Nat. Rev. Immunol 10 (2) (2010) 103–110. [DOI] [PubMed] [Google Scholar]
- [9].Pastorelli L, Garg RR, Hoang SB, Spina L, Mattioli B, Scarpa M, Fiocchi C, Vecchi M, Pizarro TT, Epithelial-derived IL-33 and its receptor ST2 are dysregulated in ulcerative colitis and in experimental Th1/Th2 driven enteritis, Proc. Natl. Acad. Sci. USA 107 (17) (2010) 8017–8022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Seidelin JB, Bjerrum JT, Coskun M, Widjaya B, Vainer B, Nielsen OH, IL-33 is upregulated in colonocytes of ulcerative colitis, Immunol. Lett 128 (1) (2010) 80–85. [DOI] [PubMed] [Google Scholar]
- [11].Luthi AU, Cullen SP, McNeela EA, Duriez PJ, Afonina IS, Sheridan C, Brumatti G, Taylor RC, Kersse K, Vandenabeele P, Lavelle EC, Martin SJ, Suppression of Interleukin-33 Bioactivity through Proteolysis by Apoptotic Caspases, Immunity 31 (1) (2009) 84–98. [DOI] [PubMed] [Google Scholar]
- [12].Beltran CJ, Nunez LE, Diaz-Jimenez D, Farfan N, Candia E, Heine C, Lopez F, Gonzalez MJ, Quera R, Hermoso MA, Characterization of the novel ST2/IL-33 system in patients with inflammatory bowel disease, Inflamm. Bowel Dis. 16 (7) (2010) 1097–1107. [DOI] [PubMed] [Google Scholar]
- [13].Kobori A, Yagi Y, Imaeda H, Ban H, Bamba S, Tsujikawa T, Saito Y, Fujiyama Y, Andoh A, Interleukin-33 expression is specifically enhanced in inflamed mucosa of ulcerative colitis, J. Gastroenterol 45 (10) (2010) 999–1007. [DOI] [PubMed] [Google Scholar]
- [14].Sedhom MAK, Pichery M, Murdoch JR, Foligne B, Ortega N, Normand S, Mertz K, Sanmugalingam D, Brault L, Grandjean T, Lefrancais E, Fallon PG, Quesniaux V, Peyrin-Biroulet L, Cathomas G, Junt T, Chamaillard M, Girard JP, Ryffel B, Neutralisation of the interleukin-33/ST2 pathway ameliorates experimental colitis through enhancement of mucosal healing in mice, Gut 62 (12) (2013) 1714–1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Sponheim J, Pollheimer J, Olsen T, Balogh J, Hammarstrom C, Loos T, Kasprzycka M, Sorensen DR, Nilsen HR, Kuchler AM, Vatn MH, Haraldsen G, Inflammatory bowel disease-associated interleukin-33 is preferentially expressed in ulceration-associated myofibroblasts, Am. J. Pathol 177 (6) (2010) 2804–2815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].He Z, Chen L, Souto FO, Canasto-Chibuque C, Bongers G, Deshpande M, Harpaz N, Ko HM, Kelley K, Furtado GC, Lira SA, Epithelial-derived IL-33 promotes intestinal tumorigenesis in Apc Min/+ mice, Sci. Rep 7 (1) (2017) 5520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Martin NT, Martin MU, Interleukin 33 is a guardian of barriers and a local alarmin, Nat. Immunol 17 (2) (2016) 122–131. [DOI] [PubMed] [Google Scholar]
- [18].Hong J, Bae S, Jhun H, Lee S, Choi J, Kang T, Kwak A, Hong K, Kim E, Jo S, Kim S, Identification of constitutively active interleukin 33 (IL-33) splice variant, J. Biol. Chem 286 (22) (2011) 20078–20086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Cayrol C, Girard JP, The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1, Proc. Natl. Acad. Sci. USA 106 (22) (2009) 9021–9026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Ali S, Nguyen DQ, Falk W, Martin MU, Caspase 3 inactivates biologically active full length interleukin-33 as a classical cytokine but does not prohibit nuclear translocation, Biochem. Biophys. Res. Commun 391 (3) (2010) 1512–1516. [DOI] [PubMed] [Google Scholar]
- [21].Lefrancais E, Duval A, Mirey E, Roga S, Espinosa E, Cayrol C, Girard JP, Central domain of IL-33 is cleaved by mast cell proteases for potent activation of group-2 innate lymphoid cells, Proc. Natl. Acad. Sci. USA 111 (43) (2014) 15502–15507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Lefrancais E, Roga S, Gautier V, Gonzalez-de-Peredo A, Monsarrat B, Girard JP, Cayrol C, IL-33 is processed into mature bioactive forms by neutrophil elastase and cathepsin G, Proc. Natl. Acad. Sci. USA 109 (5) (2012) 1673–1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Lefrancais E, Cayrol C, Mechanisms of IL-33 processing and secretion: differences and similarities between IL-1 family members, Eur. Cytokine Netw 23 (4) (2012) 120–127. [DOI] [PubMed] [Google Scholar]
- [24].Pinto D, Robine S, Jaisser F, El Marjou F, Louvard D, Regulatory sequences of the mouse villin gene that efficiently drive transgenic expression in immature and differentiated epithelial cells of small and large intestines, J. Biol. Chem 274 (10) (1999) 6476–6482. [DOI] [PubMed] [Google Scholar]
- [25].reference [Google Scholar]
- [26].Untergasser A, Nijveen H, Rao X, Bisseling T, Geurts R, Leunissen JA, Primer3Plus, an enhanced web interface to Primer3, Nucleic acids research 35(Web Server issue) (2007) W71–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Bongers G, Pacer ME, Geraldino TH, Chen L, He Z, Hashimoto D, Furtado GC, Ochando J, Kelley KA, Clemente JC, Merad M, van Bakel H, Lira SA, Interplay of host microbiota, genetic perturbations, and inflammation promotes local development of intestinal neoplasms in mice, J. Exp. Med 211 (3) (2014) 457–472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Furtado GC, Pacer ME, Bongers G, Benezech C, He Z, Chen L, Berin MC, Kollias G, Caamano JH, Lira SA, TNFalpha-dependent development of lymphoid tissue in the absence of RORgammat(+) lymphoid tissue inducer cells, Mucosal Immunol. 7 (3) (2014) 602–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Bindea G, Mlecnik B, Hackl H, Charoentong P, Tosolini M, Kirilovsky A, Fridman WH, Pages F, Trajanoski Z, Galon J, ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks, Bioinformatics 25 (8) (2009) 1091–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T, Cytoscape: a software environment for integrated models of biomolecular interaction networks, Genome Res. 13 (11) (2003) 2498–2504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Trapnell C, Pachter L, Salzberg SL, TopHat: discovering splice junctions with RNA-Seq, Bioinformatics 25 (9) (2009) 1105–1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Anders S, Pyl PT, Huber W, HTSeq–a Python framework to work with highthroughput sequencing data, Bioinformatics 31 (2) (2015) 166–169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].McCarthy DJ, Chen Y, Smyth GK, Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation, Nucleic Acids Res. 40 (10) (2012) 4288–4297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Robinson MD, McCarthy DJ, Smyth GK, edgeR: a Bioconductor package for differential expression analysis of digital gene expression data, Bioinformatics 26 (1) (2010) 139–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Wellman AS, Metukuri MR, Kazgan N, Xu XJ, Xu Q, Ren NSX, Czopik A, Shanahan MT, Kang A, Chen W, Azcarate-Peril MA, Gulati AS, Fargo DC, Guarente L, Li XL, Intestinal epithelial sirtuin 1 regulates intestinal inflammation during aging in mice by altering the intestinal microbiota, Gastroenterology 153 (3) (2017) 772–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Peine M, Marek RM, Lohning M, IL-33 in T cell differentiation, function, and immune homeostasis, Trends Immunol. 37 (5) (2016) 321–333. [DOI] [PubMed] [Google Scholar]
- [37].Pesce JT, Ramalingam TR, Wilson MS, Mentink-Kane MM, Thompson RW, Cheever AW, Urban JF Jr., Wynn TA, Retnla (relmalpha/fizz1) suppresses helminth-induced Th2-type immunity, PLoS Pathog. 5 (4) (2009) e1000393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Pastorelli L, De Salvo C, Vecchi M, Pizarro TT, The role of IL-33 in gut mucosal inflammation, Mediat. Inflamm 2013 (2013) 608187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Nunes T, Bernardazzi C, de Souza HS, Interleukin-33 and inflammatory bowel diseases: lessons from human studies, Mediat. Inflamm 2014 (2014) 423957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Schiering C, Krausgruber T, Chomka A, Frohlich A, Adelmann K, Wohlfert EA, Pott J, Griseri T, Bollrath J, Hegazy AN, Harrison OJ, Owens BMJ, Lohning M, Belkaid Y, Fallon PG, Powrie F, The alarmin IL-33 promotes regulatory T-cell function in the intestine, Nature 513 (7519) (2014) 564–568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Bamias G, Cominelli F, Role of type 2 immunity in intestinal inflammation, Curr. Opin. Gastroenterol 31 (6) (2015) 471–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Moreels TG, Pelckmans PA, Gastrointestinal parasites: potential therapy for refractory inflammatory bowel diseases, Inflamm. Bowel Dis 11 (2) (2005) 178–184. [DOI] [PubMed] [Google Scholar]
- [43].Taghipour N, Aghdaei HA, Haghighi A, Mossafa N, Tabaei SJ, RostamiNejad M, Potential treatment of inflammatory bowel disease: a review of helminths therapy, Gastroenterol. Hepatol. Bed Bench 7 (1) (2014) 9–16. [PMC free article] [PubMed] [Google Scholar]
- [44].Pascual-Reguant A, Bayat Sarmadi J, Baumann C, Noster R, Cirera-Salinas D, Curato C, Pelczar P, Huber S, Zielinski CE, Lohning M, Hauser AE, Esplugues E, TH17 cells express ST2 and are controlled by the alarmin IL-33 in the small intestine, Mucosal Immunol. 10 (6) (2017) 1431–1442. [DOI] [PubMed] [Google Scholar]
- [45].Seo DH, Che X, Kwak MS, Kim S, Kim JH, Ma HW, Kim DH, Kim TI, Kim WH, Kim SW, Cheon JH, Interleukin-33 regulates intestinal inflammation by modulating macrophages in inflammatory bowel disease, Sci. Rep 7 (1) (2017) 851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Tu L, Chen J, Xu D, Xie Z, Yu B, Tao Y, Shi G, Duan L, IL-33-induced alternatively activated macrophage attenuates the development of TNBS-induced colitis, Oncotarget 8 (17) (2017) 27704–27714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Hodzic Z, Schill EM, Bolock AM, Good M, IL-33 and the intestine: The good, the bad, and the inflammatory, Cytokine 100 (2017) 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Roussel L, Erard M, Cayrol C, Girard JP, Molecular mimicry between IL-33 and KSHV for attachment to chromatin through the H2A–H2B acidic pocket, EMBO Rep. 9 (10) (2008) 1006–1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Carriere V, Roussel L, Ortega N, Lacorre DA, Americh L, Aguilar L, Bouche G, Girard JP, IL-33, the IL-1-like cytokine ligand for ST2 receptor, is a chromatinassociated nuclear factor in vivo, Proc. Natl. Acad. Sci. USA 104 (1) (2007) 282–287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Shao D, Perros F, Caramori G, Meng C, Dormuller P, Chou PC, Church C, Papi A, Casolari P, Welsh D, Peacock A, Humbert M, Adcock IM, Wort SJ, Nuclear IL-33 regulates soluble ST2 receptor and IL-6 expression in primary human arterial endothelial cells and is decreased in idiopathic pulmonary arterial hypertension, Biochem. Biophys. Res. Commun 451 (1) (2014) 8–14. [DOI] [PubMed] [Google Scholar]
- [51].Ali S, Mohs A, Thomas M, Klare J, Ross R, Schmitz ML, Martin MU, The dual function cytokine IL-33 interacts with the transcription factor NF-kappaB to dampen NF-kappaB-stimulated gene transcription, J. Immunol 187 (4) (2011) 1609–1616. [DOI] [PubMed] [Google Scholar]
- [52].Choi YS, Park JA, Kim J, Rho SS, Park H, Kim YM, Kwon YG, Nuclear IL-33 is a transcriptional regulator of NF-kappaB p65 and induces endothelial cell activation, Biochem. Biophys. Res. Commun 421 (2) (2012) 305–311. [DOI] [PubMed] [Google Scholar]
- [53].Gautier V, Cayrol C, Farache D, Roga S, Monsarrat B, Burlet-Schiltz O, Gonzalez de Peredo A, Girard JP, Extracellular IL-33 cytokine, but not endogenous nuclear IL-33, regulates protein expression in endothelial cells, Sci. Rep 6 (2016) 34255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Bessa J, Meyer CA, de Vera Mudry MC, Schlicht S, Smith SH, Iglesias A, Cote-Sierra J, Altered subcellular localization of IL-33 leads to non-resolving lethal inflammation, J. Autoimmun 55 (2014) 33–41. [DOI] [PubMed] [Google Scholar]
- [55].Dinarello CA, Immunological and inflammatory functions of the interleukin-1 family, Annu. Rev. Immunol 27 (2009) 519–550. [DOI] [PubMed] [Google Scholar]
- [56].Malik A, Kanneganti TD, Function and regulation of IL-1alpha in inflammatory diseases and cancer, Immunol. Rev 281 (1) (2018) 124–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Di Paolo NC, Shayakhmetov DM, Interleukin 1alpha and the inflammatory process, Nat. Immunol 17 (8) (2016) 906–913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Cayrol C, Girard JP, Interleukin-33 (IL-33): a nuclear cytokine from the IL-1 family, Immunol. Rev 281 (1) (2018) 154–168. [DOI] [PubMed] [Google Scholar]
- [59].Dinarello CA, Overview of the IL-1 family in innate inflammation and acquired immunity, Immunol. Rev 281 (1) (2018) 8–27. [DOI] [PMC free article] [PubMed] [Google Scholar]