

Abstract
Xylanase and α-amylase enzymes participate in the degradation of organic matter, acting in hemicellulose and starch mineralization, respectively, and are in high demand for industrial use. Mangroves represent a promising source for bioprospecting enzymes due to their unique characteristics, such as fluctuations in oxic/anoxic conditions and salinity. In this context, the present work aimed to bioprospect xylanases from mangrove soil using cultivation-dependent and cultivation-independent methods. Through screening from a metagenomic library, three potentially xylanolytic clones were obtained and sequenced, and reads were assembled into contigs and annotated. The contig MgrBr135 was affiliated with the Planctomycetaceae family and was one of 30 ORFs selected for subcloning that demonstrated only amylase activity. Through the cultivation method, 38 bacterial isolates with xylanolytic activity were isolated. Isolate 11 showed an enzymatic index of 10.9 using the plate assay method. Isolate 39 achieved an enzyme activity of 0.43 U/mL using the colorimetric method with 3,5-dinitrosalicylic acid. Isolate 39 produced xylanase on culture medium with salinity ranging from 1.25 to 5%. Partial 16S rRNA gene sequencing identified isolates in the Bacillus and Paenibacillus genera. The results of this study highlight the importance of mangroves as an enzyme source and show that bacterial groups can be used for starch and hemicellulose degradation.
Electronic supplementary material
The online version of this article (10.1007/s42770-019-00162-7) contains supplementary material, which is available to authorized users.
Keywords: Enzyme, Culture-dependent method, Metagenomic library, Biotechnology
Introduction
Mangroves are tropical ecosystems situated in transitional areas between sea and land that have anaerobic/aerobic sediments and salinity varying from 4 to 30% [1]. These sediments are rich in organic matter but are deficient in nutrients due to a low decomposition rate under oxygen-limiting conditions. Therefore, the microbial community plays an important role in transforming organic matter into sources of nitrogen, phosphorus, carbon, and other nutrients used by plants and organisms [2]. Microorganisms present in mangroves are adapted to diverse environmental characteristics, as there is variation in salinity, nutrients, and oxygen, representing a promising source for bioprospecting microbial enzymes [3].
Xylanases and amylases are enzymes that present industrial interest. Xylanases are glycoside hydrolases (EC 3.2.1.x) that catalyze the hydrolysis of β-1,4-xylosidic linkages of the main chain of xylan, producing xylooligosaccharides. Based on the sequence of the catalytic domains, xylanases have been classified into 6 different glycoside hydrolase (GH) families in the Carbohydrate-Active enZymes (CAZy) database: GH5, GH8, GH10, GH11, GH30, and GH43 [4]. However, xylanases belonging to GH10 and GH11 are the most studied and present differences in substrate specificity and structure [5]. The GH10 xylanases, for example, present less substrate specificity and the GH11 xylanases present a higher specificity to hydrolyze xylan, being recognized as true xylanases [5]. These xylanases are used in industrial processes, such as bioethanol production, animal feed, paper industries, food industries, and bioenergy [6]. Xylanases with special features as halotolerant xylanases received attention because these xylanases can be used in wastewater treatment and industrial processes that require low water potential, such as carbohydrate production and processing of seafood and saline food [7]. Xylan composes hemicellulose, a lignocellulosic material present in organic matter and considered the second most abundant polysaccharide in nature [8].
The 1,4-α-glucan branching enzyme (EC 2.4.1.18) catalyzes the cleavage of α-1,4-glycosidic linkages and the formation of α-1,6-branch points in glycogen and starch substrates. The glucan branching enzymes (GBE) from the Bacteria domain are members of subfamily 9 belonging to the glycoside hydrolase family 13 (GH13) and its secondary structure formed by three domains [9]. The A domain is a central (β/α) barrel domain that presents the catalytic function and it is the most conserved domain [9]. The B domain acts in the enzyme specified, and the C domain is responsible for substrate binding and stabilizes the catalytic domain [9]. Amylolytic enzymes account for 25–33% of worldwide enzyme production and are widely applied in biotechnological processes such as textiles, baking, distilled beverages, beers, glucose syrup production, and dextrin gum preparation [10]. Starch is present in organic matter; found in the roots, stems, and leaves of plants; and used as a glucose storage molecule [11].
The culture-dependent method is a classical strategy for bioprospecting enzymes from environmental samples [12]. However, the necessity to reproduce requirements for microbial growth in laboratory conditions is a limitation of this approach [13]; thus, new strategies for bioprospecting have been driven by culture-independent methods, such as functional screening from metagenomic libraries constructed using environmental DNA [14]. This method favors the discovery of enzymes from uncultivated microorganisms without performing a cultivation step [15]. The metagenomics libraries have permitted the discovery of xylanases [16] and α-amylases [17].
Microbial enzymes have specificity, lower production cost, and structural stability [18]. In addition, thermostable and halotolerant enzymes are required in industrial processes that occur in high temperatures and high salt concentrations; therefore, the bioprospection of enzymes tolerant to these conditions is important [12]. The present study aimed to identify genes and microorganisms from mangrove samples that were able to produce xylanases and amylases, either using approaches dependent and independent of cultivation. Based on previous studies that explored the mangrove sediments as biotechnological source [14, 19–21], the study hypothesized that the conditions found in mangrove areas, mainly the high salinity, could select for organisms harboring genes and enzymes of high industrial interest and these enzymes could be recovered by applying culture-dependent and culture-independent methods.
Materials and methods
Sampling
Sampling was performed in a mangrove located in Bertioga, São Paulo state (latitude 23° 53′ 49″ S and longitude 46° 12′ 28″ W), in November 2010. The mangrove vegetation was composed of Laguncularia racemosa (white mangrove), Avicennia schaueriana (black mangrove), and Rhizophora mangle (red mangrove) [22]. Samples were collected at three different points in triplicate at a depth of 30 cm from the soil surface. Aliquots of 50 g from each soil sample were pooled to generate composite samples and homogenized for bacterial isolation.
Metagenomic fosmid library construction
The metagenomic library of the mangrove soil was constructed using the Cloning-Ready Copy Control pCC2FOS Kit (Epicentre Biotechnologies, Madison, WI, USA). First, the total DNA was extracted from the mangrove sample following the protocol developed by Tsai and Olson [23] with modifications proposed by Vasconcellos et al. [24]. High molecular weight DNA fragments (~ 40 kb) were selected after visualization on a 1% agarose gel by pulsed-field gel electrophoresis (PFGE) at 9 V cm−1, 0.5–0.5 for 5 h, 120° angle, and 12 °C. The DNA fragments were cut from the gel and subjected to the end-repair reaction, insert binding in the pCC2FOS vector, and packing of fosmids in lambda phage, and it was transformed into Escherichia coli EPI300 (Epicentre Biotechnologies, Madison, WI, USA). In total, 12,960 clones with insert sizes ranging between 30 and 40 kb were obtained. For library validation, 30 clones were randomly selected and submitted to restriction analysis.
Metagenomic fosmid library and screening for xylanolytic enzymes
For functional screening, E. coli transformants were inoculated into Petri dishes containing solid medium with xylan (yeast extract 0.5 g L−1; NaNO3 0.5 g L−1; K2HPO4 1.0 g L−1; FeSO4 0.01 g L−1; MgSO4·7H2O 0.5 g L−1; beechwood xylan (Sigma-Aldrich, USA) 2.0 g L−1; and agar 15 g L−1) supplemented with chloramphenicol (12.5 μg/mL) and incubated for 72 h at 37 °C. The xylanase activity of the clones was observed by the presence/absence of hydrolysis halos around the colonies indicated with iodine solution [25].
Extraction of fosmidial DNA and sequencing
The fosmidial DNA of three positive clones for xylanolytic activity named MgrBr18, MgrBr61, and MgrBr135 was extracted using the Qiagen Large-Construct Kit (Qiagen Inc., Hilden, Germany). Sequencing was performed using Ion Torrent (Life Technologies, USA) with the ION Xpress DNA Fragment Library Kit to build the library and ION Xpress™ Template v 2.0 Kit (Life Technologies, Camarillo, USA) and ION Touch “mini robot” (Life Technologies, Camarillo, USA) to sample enrichment. Sample fragments were linked to Ion Sphere Particle (ISP) (metallic beads) and injected in 316 Chip for sequencing. These chips generated up to 100 Mb of sequences (reads) ranging in size from 50 to 200 bp.
Assembly and annotation of contigs
Sequences generated for fosmidial DNA from MgrBr18, MgrBr61, and MgrBr135 were processed for removing low-quality sequences. The assembling positive clone contigs were performed separately by CLC Genomics Workbench Qiagen software (CLC Bio-Qiagen, USA). In the first step, the data generated in the FASTQ format were imported into the CLC software. The sequences were filtered using Create Sequencing QC Report software considering only sequences with Phred > 20. To remove the adapters and barcodes, the sequences were trimmed using Trim Sequences Adapter List. To map and remove the sequences related to the vector (pCC2FOS) and to E. coli (EPI-300), the mapping for reference (Maps Reads to Reference) tool was used. The sequences remaining were used for contig assembling through the “De Novo Assembly” strategy with the following parameters: (i) mismatch cost (3); (ii) interaction cost (3); (iii) deletion cost (3); (iv) length fraction (0.3); and (v) similarity fraction (0.6). After assembly, contigs larger than 20 kb and Phred > 20 were submitted to gene annotation based on the open reading frames (ORFs). After exporting the contigs in CLC Genomics Workbench in FASTA format, these contigs were submitted to the RAST (Rapid Annotation using Subsystem Technology) database (http://rast.nmpdr.org). For a better prediction of genes encoding xylanases, ORF sequences generated and annotated in RAST were compared with the Pfam (Protein Families) database and with the GenBank databases through the BLASTp tool.
Taxonomic and functional annotation of contig MgrBr135
The contig formed with reads of the MgrBr135 clone presented Phred > 20 and size expected in the assembly with approximately 36 kb, which was close to the size of the original insert (40 kb) and was selected for further analyses. To determine the taxonomic affiliation of the insert, nucleotide sequences of the 30 ORFs, generated by the functional annotation, were compared with those present in the GenBank database using the BLASTx program. Based on the functional annotation, one sequence was selected and submitted to the Conserved Domain Database (CDD) software and dBCAN (http://csbl.bmb.uga.edu/dbCAN/) to identify possible catalytic domains, carbohydrate-binding modules, and affiliation with the glycosyl hydrolase family. This sequence was selected for subcloning into the expression vector and to perform the phylogeny study. To study the phylogeny of the ORFs used for subcloning, its amino acid sequence was compared with the GenBank database using the BLASTp tool. The maximum identity obtained was 76%, and sequences with a similarity greater than 74% were used for phylogenetic tree construction. The tree was constructed with the MEGA7 (Molecular Evolutionary Genetics Analysis Version 7.0) program [26]. Sequence alignment was performed with the MUSCLE tool, converted into a distance matrix determined by the JTT matrix–based model, and grouped by the maximum likelihood method. The tree consistency was determined by bootstrap analysis based on 1000 subsamples in the distance matrix.
Subcloning in the heterologous expression vector
ORF 30 was chosen to subcloning due to present similarity with genes that encode hydrolytic enzymes. To amplify the xylanase coding region, the primers, forward 5′-CAGTCTCGAGGAACAAAAAGCAGAAAAAGC-3′ (containing restriction site XhoI) and reverse 5′-TCTCATATCTAGAGCCCCTTCATGTCTGA-3′ (containing restriction site XbaI), were designed. PCR followed the following steps: an initial denaturation at 94 °C for 4 min; 30 cycles of 94 °C for 1 min, 50 °C for 45 s, and 72 °C for 1.5 min; and a final extension at 72 °C for 10 min. The amplified product was purified with the Wizard SV Gel and PCR Clean-Up System Kit (Promega, Madison, WI, USA). Plasmid DNA from the expression vector pBAD/Myc-HisB and PCR product were digested with XhoI (10 U/μL) and XbaI (10 U/μL) enzymes (Thermo Scientific Inc., USA) in microtubes for 4 h at 37 °C. PCR product and digested plasmid were evaluated on 0.8% agarose gel. The marker GeneRuler DNA Ladder 1 kb (Thermo Scientific Inc., USA) and MassRuler Low Range DNA Ladder (Thermo Scientific Inc., USA) were used to determine DNA concentration. Bands at the expected size (2000 bp) were cut from the gel and purified with a QIAEX II Gel Extraction Kit (Qiagen, USA), and 2 μL of the samples was again evaluated on an agarose gel for quality, quantity, and band pattern. The ligation reaction consisted of 50 ng insert, 100 ng/μL pBAD/Myc-HisB, 2 μL T4 DNA ligase buffer (10×) (Invitrogen, USA), 1 U/μL T4 DNA ligase (Invitrogen, USA), and 8.9 μL of sterile deionized water to reach a final volume of 20 μL. The reaction was incubated overnight at 24 °C, and transformation was performed by electroporation of E. coli TOP 10 (Invitrogen, USA). Transformants were cultured on plates with Luria-Bertani (LB) medium: 10 g L−1 tryptone, 5 g L−1 yeast extract, 5 g L−1 NaCl, 2.0 g L−1 xylan, 0.002% of l-arabinose, and 15 g L−1 agar with the antibiotic ampicillin 100 μg/mL for 24 h at 37 °C. To confirm cloning success, the colonies generated were tested by PCR using the same reaction previously described for gene amplification.
Expression assay of the transformants
To determine the expression of the enzymes by the transformants, tests were conducted to detect xylanase and amylase activities. The amylase test was performed based on the identification of a catalytic domain to 1,4-α-glucan branching enzyme (AmyAC_Glg_BE) in the amino acid sequence of the ORF through comparison in the CCD database. For the expression test of xylanase activity, culture medium containing xylan (2.0 g L−1) was used. This medium was applied previously for functional screening of the metagenomic library. For the expression test to amylase activity, medium with starch as the primary carbon source was used (yeast extract 0.5 g L−1; NaNO3 0.5 g L−1; K2HPO4 1.0 g L−1; FeSO4 0.01 g L−1; MgSO4·7H2O 0.5 g L−1; starch (Sigma-Aldrich, USA) 2.0 g L−1; and agar 15 g L−1). Transformant colonies were inoculated with sterilized wooden sticks in Petri dishes containing the culture medium and maintained for 72 h at 37 °C. The plates were stained with an iodine solution and washed with 2 M NaCl to confirm enzyme activity [25].
Isolation, purification, and determination of enzymatic potential of xylanolytic bacteria
To screen the xylanolytic bacteria, a qualitative method was adopted where aliquots (0.1 mL) from serial dilutions (10−2–10−5) of soil and saline water were added in solid medium with xylan as the primary carbon source [27]. The plates were incubated in growth chambers (BOD) at 28 °C for 48 h. The test to confirm xylanolytic activity was performed by the addition of iodine solution to evaluate the degradation halos around the pure colonies [25]. To determine the enzymatic potential through the plate assay method, the thirty-eight isolates and E. coli (negative control) were inoculated with a sterile toothpick in triplicate in solid medium with xylan and incubated for 72 h at 28 °C [28]. The degradation halo was evaluated with the addition of iodine solution and subsequent washes with 5 M NaCl [29]. The enzymatic index (EI) was determined by the ratio between the halo of the degradation mean diameter (dh) and the colony mean diameter (dc). To quantify enzymatic activity, the colorimetric 3,5-dinitrosalicylic acid (DNS) method modified by Miller [30] was used. The isolates and negative control (E. coli) were cultured in triplicate for 4 days under agitation at 28 °C in 15 mL of liquid medium with xylan. To recover crude extracellular enzymes, 2 mL of each sample was centrifuged at 14,000g for 20 min at 4 °C to collect the supernatant that contained the extracellular enzymes [31]. The enzymatic assay was performed according to Samanta et al. [31] and was read at 540-nm length [32] using a standard curve with known xylose concentration. Enzymatic activity was expressed as a unit of enzymatic activity (U) and was defined as the enzyme amount required to release 1 μmol of reducing sugar per minute of reaction in 1 min (1 U/mL) [33]. To determine the enzymatic potential at different salt concentrations, isolate 39, which presented the highest EI, was submitted to enzymatic activity tests with different NaCl concentrations, 1.25%, 2.25%, 3.75%, and 5%, added to the reaction buffer [34], and the same range of NaCl concentrations was used in the culture medium to test enzyme activity under salinity conditions [35].
Molecular identification of the isolates
DNA extraction from 38 isolates was performed using the phenol-chloroform extraction protocol by Sambrook and Russel [36]. Isolate identification was based on partial 16S rRNA gene sequencing using the primers pR1387 (5′-CGGTGTGTACAAGGCCCGGGAACG-3′) and p027 (5′-GAGAGTTTGATCCTGGCTCAG-3′). The amplification reaction was carried out according to Odee et al., and the PCR product was purified for sequencing on an ABI PRISM 3100 genetic analyzer (Thermo Fisher Scientific, USA).
The generated sequences were imported to CLC Genomics Workbench Qiagen software to remove low-quality sequences. Then, sequences were imported to Codoncode Aligner v. 2.0.4 and trimmed using the “clips ends” tool. Only nucleotides with a sequencing quality value ≥ 500 and equivalent to Phred quality > 20 were considered. Valid sequences were used for BLASTn similarity searches in GenBank. Sequences with the greatest similarity to those obtained in the present study were used for inference based on a phylogenetic tree constructed with MEGA7 [26] program. The tree was first constructed by the sequence alignment by MUSCLE, later converted into a distance matrix determined by the Kimura 2-parameter model and grouped by the maximum likelihood method [37]. The consistency of the tree structure was determined by bootstrap analysis based on 1000 subsamples in the distance matrix. Sequence affiliated with E. coli was used as an outgroup sequence for phylogenetic analysis. 16S rRNA sequences were submitted in the NCBI GenBank under accession numbers MH478123- MH478160 (Table 1).
Table 1.
Identification of the similarities among 16S rRNA partial gene sequences of the xylanolytic bacteria isolated from mangrove sediment with sequences in the GenBank database (fragments of approximately 400–1100 bp)
| Isolates | Organism | Ident (%) | Accession number |
|---|---|---|---|
| 2 | Bacillus cereus | 100 | MH478123 |
| 11 | Bacillus megaterium | 99 | MH478124 |
| 30 | Bacillus aquimaris | 100 | MH478125 |
| 48 | Bacillus subtilis | 100 | MH478126 |
| 3 | Bacillus infantis | 100 | MH478127 |
| 12 | Bacillus subtilis | 100 | MH478128 |
| 22 | Bacillus pseudomycoides | 100 | MH478129 |
| 31 | Bacillus cereus | 100 | MH478130 |
| 39 | Bacillus pumilus | 99 | MH478131 |
| 49 | Paenibacillus polymyxa | 99 | MH478132 |
| 4 | Bacillus aquimaris | 100 | MH478133 |
| 13 | Bacillus subtilis | 100 | MH478134 |
| 23 | Bacillus aquimaris | 100 | MH478135 |
| 32 | Bacillus megaterium | 100 | MH478136 |
| 40 | Bacillus pseudomycoides | 100 | MH478137 |
| 50 | Bacillus aquimaris | 100 | MH478138 |
| 14 | Bacillus aquimaris | 100 | MH478139 |
| 25 | Bacillus cereus | 100 | MH478140 |
| 42 | Paenibacillus polymyxa | 99 | MH478141 |
| 51 | Bacillus aquimaris | 99 | MH478142 |
| 7 | Bacillus aquimaris | 99 | MH478143 |
| 15 | Bacillus aquimaris | 100 | MH478144 |
| 26 | Bacillus aquimaris | 99 | MH478145 |
| 34 | Bacillus aquimaris | 99 | MH478146 |
| 43 | Bacillus aquimaris | 100 | MH478147 |
| 8 | Bacillus aquimaris | 100 | MH478148 |
| 16 | Bacillus aquimaris | 100 | MH478149 |
| 27 | Paenibacillus polymyxa | 99 | MH478150 |
| 35 | Bacillus megaterium | 100 | MH478151 |
| 9 | Bacillus pseudomycoides | 100 | MH478152 |
| 18 | Bacillus indicus | 100 | MH478153 |
| 28 | Bacillus megaterium | 100 | MH478154 |
| 36 | Bacillus megaterium | 100 | MH478155 |
| 45 | Bacillus aquimaris | 99 | MH478156 |
| 10 | Bacillus aquimaris | 99 | MH478157 |
| 19 | Bacillus aquimaris | 99 | MH478158 |
| 29 | Bacillus cereus | 100 | MH478159 |
| 37 | Paenibacillus polymyxa | 99 | MH478160 |
Results
Metagenomic fosmid library, screening for xylanolytic enzymes, and annotation of contigs
Three clones (MgrBr18, MgrBr61, and MgrBr135) presenting xylanolytic activity were obtained after functional screening of the metagenomic library. The 316 Chip with 69% sequencing coverage and 2,029,977 reads was used for all evaluated clones with 100 bp as the average size. Most of the sequences presented Phred values between 20 and 30 (Online Resource 1). From all evaluated clones, 34 contigs were obtained for clone MgrBr18, 3 contigs for clone MgrBr61, and 2 contigs for clone MgrBr135. Functional annotation of the contig formed by the MgrBr135 clone reads was prioritized for analysis because this clone presented the best quality, a contig with approximately 36 kb, which was close to the original insert size (40 kb), and it was similar with genes that encode hydrolytic enzymes. In addition, 30 ORFs generated in the RAST server demonstrated a considerable size (1944 nucleotides) that likely contained the coding region of the enzyme of interest.
Taxonomic and functional annotation of contig MgrBr135
The MgrBr135 contig map is presented in Fig. 1, and the functional annotation of the 30 ORFs can be visualized in Table 2. The contig sequence was deposited in the GenBank database (code PRJNA483139), in the BioProject, and SRR7726101 in the Sequence Read Archive (SRA). Of the 30 ORFs, 87% were affiliated with the Planctomycetes phylum, 87% with the Planctomycetia class, 80% with the Planctomycetales order, and 74% with the Planctomycetaceae family (Fig. 2). Through ORF functional analysis, one gene encoding a 1,4-α-glucan branching enzyme composed of 647 amino acids was identified. The sequence of ORF 30 was submitted to comparison with sequences of proteins by the Conserved Domain Database (CDD) and the ORF 30 was identified and confirmed encoding a 1,4-α-glucan branching enzyme (AmyAC_Glg_BE). This enzyme belongs to the glycosyl hydrolase (GH13) family presenting also a carbohydrate-binding domain 48 (CBM 48) as annotated by dbCAM software that based on classification of family and domain from CAZy database (Online Resource 2). Through evaluation based on the phylogenetic tree, a 1,4-α-glucan branching enzyme belonging to Planctomycetes sp. bacteria [38] presented 72.54% of identity with the 1,4-α-glucan branching enzyme isolated in this work (Fig. 3).
Fig. 1.

Map of the contig MgrBr135. The highlighted ORF (red) represents the ORF used in subcloning
Table 2.
Functional comparative annotation of ORFs. The ORF MgrBr135 (accession number SRR7726101) selected to proceed with subcloning is highlighted
| ORF | RAST | Blastp | Pfam |
|---|---|---|---|
| 1 | Regulator protein | Regulator protein | Regulator protein |
| 2 | Cell division protein FtsH | Cell division protein FtsH | Cell division protein FtsH |
| 3 | 1-Deoxy-d-xylulose 5-phosphate reductoisomerase | 1-Deoxy-d-xylulose 5-phosphate reductoisomerase | 1-Deoxy-d-xylulose 5-phosphate reductoisomerase |
| 4 | Intramembrane protease | Probable metalloproteinase | - |
| 5 | UDP-3-O-[3-Hydroxymyristoyl] glucosamine N-acyltransferase | UDP-3-O-(3-Hydroxymyristoyl) glucosamine N-acyltransferase | UDP-3-O-(3-Hydroxymyristoyl) glucosamine N-acyltransferase |
| 6 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 7 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 8 | Acetoacetate metabolism regulatory | Acetoacetate metabolism regulatory | Acetoacetate metabolism regulatory |
| 9 | Ribonucleotide reductase of class II | Ribonucleotide reductase | Ribonucleotide reductase |
| 10 | Ribonucleotide reductase of class II | Ribonucleotide reductase | Ribonucleotide reductase |
| 11 | Ribosomal protein | Ribosomal protein | Ribosomal protein |
| 12 | Ribonuclease | Ribonuclease | Ribonuclease |
| 13 | Hypothetical protein | Hypothetical protein | - |
| 14 | Hypothetical protein | Hypothetical protein | - |
| 15 | Phosphoenolpyruvate protein | Phosphoenolpyruvate protein | Phosphoenolpyruvate protein |
| 16 | Phosphotransferase system, phosphocarrier protein HPr | PTS IIA–like nitrogen regulatory protein PtsN | - |
| 17 | Probable sigma-54 modulation protein | Probable sigma-54 modulation protein | Probable sigma-54 modulation protein |
| 18 | Iron-binding protein | Iron transporter | Iron transporter |
| 19 | Hypothetical protein | Hypothetical protein | - |
| 20 | Serine hydroxymethyltransferase | Glycine hydroxymethyltransferase | Serine hydroxymethyltransferase |
| 21 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 22 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 23 | Hypothetical protein | Hypothetical protein | - |
| 24 | Alpha/beta hydrolase fold | Alpha/beta hydrolase fold protein | Alpha/beta hydrolase fold protein |
| 25 | Hypothetical protein | Hypothetical protein | - |
| 26 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 27 | Radical SAM domain protein | Radical SAM protein | - |
| 28 | Hypothetical protein | Hypothetical protein | Hypothetical protein |
| 29 | Trehalose synthase | Alpha-amylase | Phosphotransferase enzyme family |
| 30 | 1,4-Alpha-glucan branching enzyme | Glycogen branching protein | Alpha amylase |
Fig. 2.

Representation of the taxonomic affiliation of the 30 ORFs generated through the RAST server. Sequences were compared with those present in the GenBank database through the BLASTx program. a Phylum. b Class. c Order. d Family
Fig. 3.

Phylogenetic reconstruction demonstrating the relationships between the metagenomic insert (amino acids) and sequences of the most similar organisms present in the BLASTp database. Phylogeny was determined by distance matrix–based model JTT. Values on the branches indicate the frequency of grouping, as determined by the bootstrap test with 1000 replicates. The obtained α-amylase is indicated with bold letters
Subcloning in an expression vector and the expression of amylase in E. coli transformants
Twenty-four colonies were cultivated in the medium with the antibiotic ampicillin and four of these colonies carried the gene with the expected size of approximately 2000 bp (Online Resource 3). The colonies were inoculated in the culture medium with starch as the primary carbon source and showed a degradation halo (Online Resource 4), and none of the transformants demonstrated enzymatic activity in the medium amended with xylan.
Isolation and enzymatic potential assays
Using targeted microorganism isolation, thirty-eight bacterial isolates were obtained with the ability to consume xylan. Enzymatic indexes ranged from 0 to 10.9, and only isolate 39 showed no degradation halo. Isolate 11 showed the highest value (EI = 10.9) (Fig. 4), and xylanolytic enzyme activity by the DNS method ranged from 0.02 to 0.44 U/mL (Fig. 5). Isolate 39 did not present a degradation halo by the plate assay method but reached the highest enzymatic activity value (0.44 U/mL) determined by the DNS method and was submitted to a test at different salt concentrations. This isolate showed an enzymatic activity reduction (0.33 U/mL) when 3.75% NaCl was added to the reaction buffer (Tukey’s test p > 0.05). Therefore, the enzyme released under such culture conditions can be used in processes using 0 to 2.25% NaCl concentrations. However, the NaCl addition in the culture medium did not alter enzymatic activity (p = 0.35 at p value > 0.05); thus, higher NaCl concentrations could be tested to identify the maximum concentration tolerated by the isolate (Fig. 6).
Fig. 4.
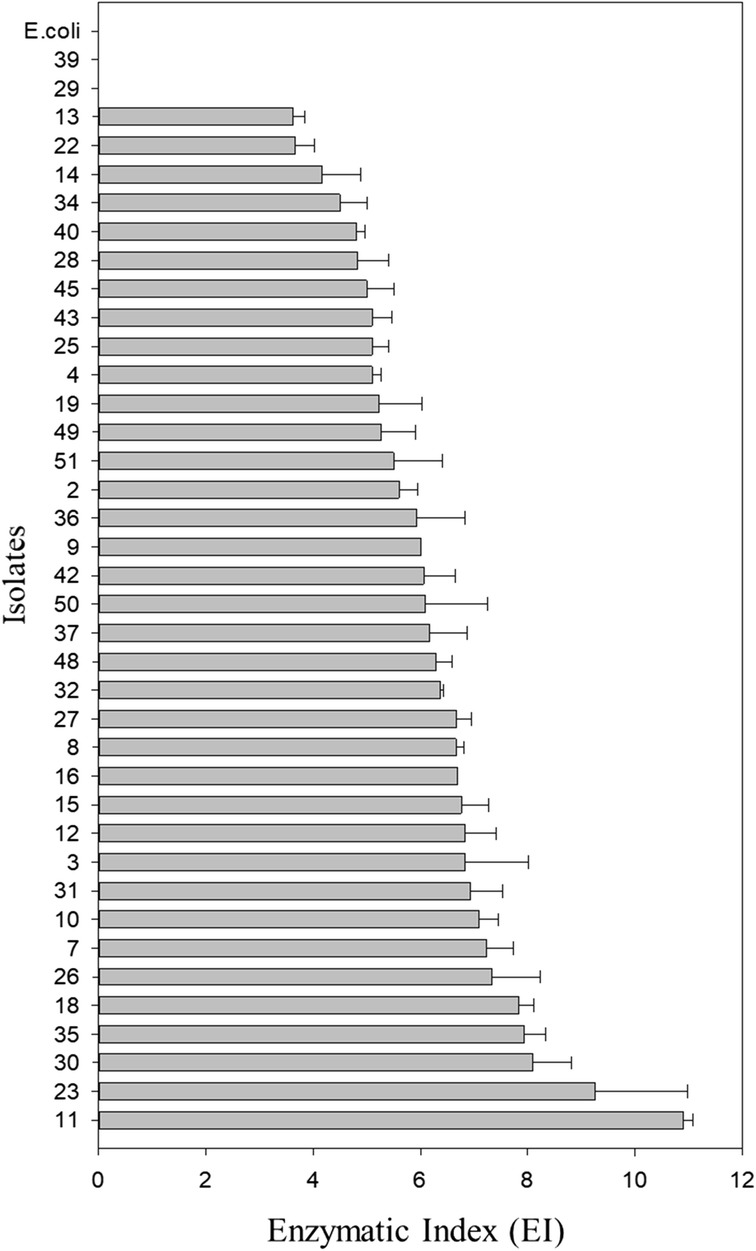
Enzymatic index (EI) of the 38 bacterial isolates from a composite sample of mangrove sediment, an oil spill area, located in the city of Bertioga, São Paulo. The EI was calculated by the ratio of the mean diameter of the degradation halo and the mean diameter of the colony (cm). Error bars indicate the standard deviation above the mean (n = 3)
Fig. 5.
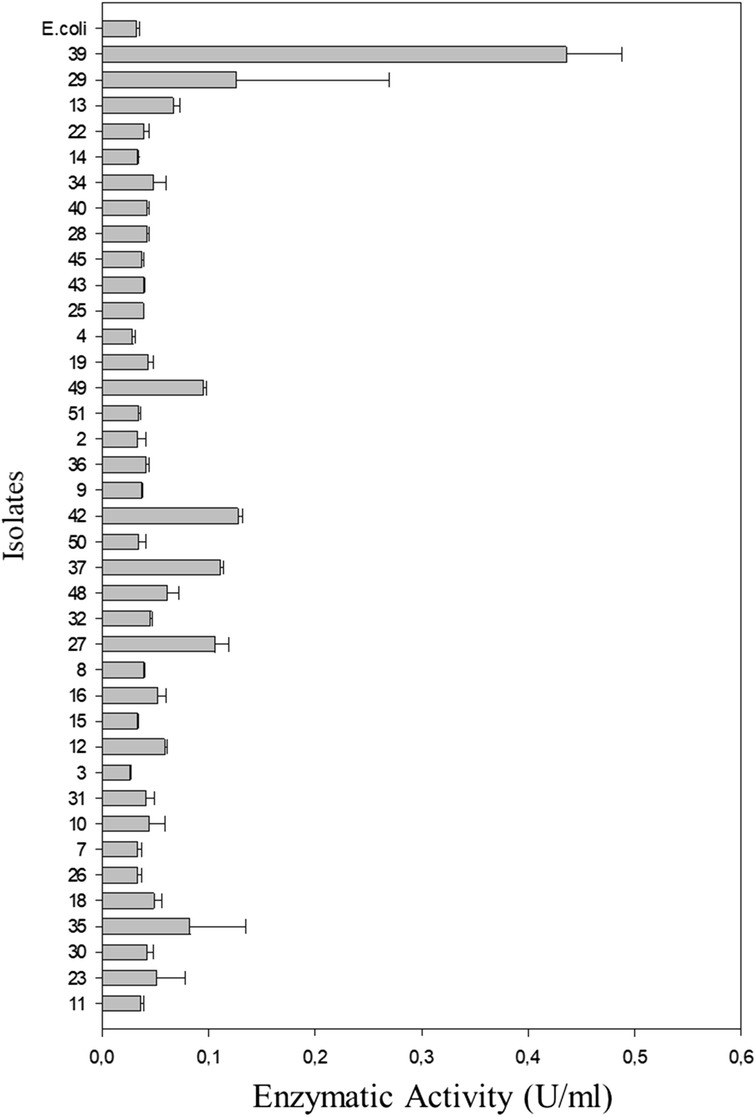
The quantitative xylanolytic activity of 38 bacterial isolates from a composite sample of mangrove sediment located in the city of Bertioga, São Paulo. The activity was measured by the proportion of reducing sugars released by the enzymatic hydrolysis of xylan. The error bars indicate the standard deviation above the mean (n = 3)
Fig. 6.
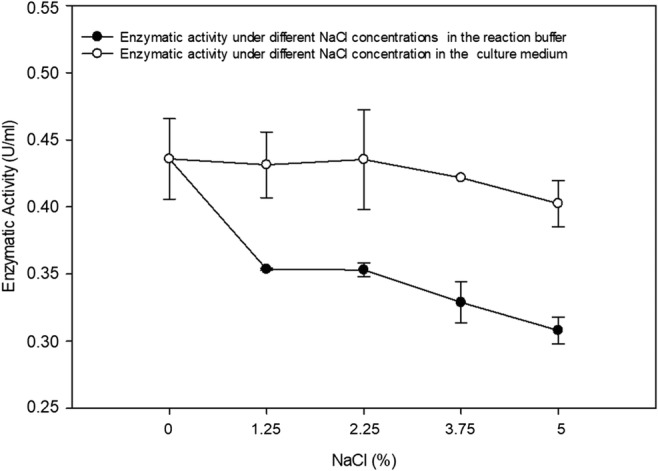
Enzymatic reaction of isolate 39 with different concentrations of NaCl. Growth of isolate 39 at different concentrations of NaCl added to the culture medium and enzymatic activity under addition of the same NaCl concentration range in the reaction buffer
Molecular characterization of the isolates
Phylogenetic reconstruction revealed affiliation of the 38 isolates with the Firmicutes phylum. The Bacillus genus belonging to the Bacillaceae family harbored 89% of these isolates, and 17 isolates were identified that had 99 to 100% identity with the Bacillus aquimaris 16S rRNA gene, and 5 isolates presented the same identity index range with Bacillus megaterium. For the other isolates, 100% identity was demonstrated with 4 isolates similar to B. cereus, 3 isolates similar to B. subtilis, 3 isolates similar to B. pseudomycoides, 1 isolate similar to B. pumilus, 1 isolate similar to B. indicus, and 1 isolate similar to B. infantis. The other 11% of isolates were affiliated with the genus Paenibacillus, classified in the Paenibacillaceae family and showed a 99% identity index with the 16S rRNA of Paenibacillus polymyxa (Fig. 7 and Table 1).
Fig. 7.

Phylogenetic reconstruction demonstrating relationships between isolates analyzed with sequences of the most similar organisms present in the database. Phylogeny was determined by the maximum likelihood method using the distance matrix Kimura 2-parameter model. The Escherichia coli sequence was used as an external group for tree rooting. Values on the branches indicate the frequency of isolate grouping, determined by the bootstrap test with 1000 replicates
Discussion
The advantage of using two different methods to bioprospect microbial enzymes is the ability to access genes belonging to either cultured or uncultured microorganisms [13]. The gene encoding an amylase obtained through functional metagenomics was affiliated with the Planctomycetaceae family, belonging to the Planctomycetes phylum. Most Planctomycetes species have slow growth, making cultivation difficult [39]; thus, the adoption of a culture-independent method is important to allow isolation and subcloning of enzymes belonging to the phylum. This group is involved in nitrogen cycling, participating in the ammonium oxidation process. Planctomycetes have been described as belonging to the mangrove microbial community of the same area used in the present study [22] and are found in other habitats, such as marine water [40], freshwater [41], rhizosphere [42], and soil [43]. The 1,4-α-glucan branching enzyme obtained in this work was most related to an amylase sequence from a genome also belonging to a species of Planctomycetes recovered from an aquifer system [38], and the non-catalytic domain described is designated as a starch-binding domain (SBD) reported to facilitate the binding and insoluble starch [44] degradation. Planctomycetes have been demonstrated to synthesize esterases in mangroves [14] and forest soils [15], and genome analyses have revealed genes that encode cellulases [45] and xylanases [46]. However, studies have not explored amylases produced by Planctomycetes.
Applying a culture-dependent approach, a collection of isolates that produced xylanases were obtained, and all isolates were affiliated with the phylum Firmicutes; this phylum is highly represented by cultured microorganisms described in the literature [47]. The investigation of the microbiota from mangroves in the present work [22], through metagenomics analyses, identified Firmicutes as the second most abundant phylum. Firmicutes play an essential role in organic matter and xenobiotic degradation with a great capacity to promote catabolic reactions of recalcitrant organic compounds [48]. Most isolates obtained in this study were affiliated with the Bacillus genus. This genus plays a fundamental role in biogeochemical cycles and is a group within the Bacteria domain with a great biotechnological importance and industrial application [49]. Numerous hydrolytic enzymes belonging to Bacillus have been characterized, including xylanases [50], cellulases [51], and amylases [52]. Some enzymes are already used on a large scale in bioconversion processes [53]. The other isolates belong to the genus Paenibacillus, and isolates from this genus are capable of hydrolyzing lignocellulosic materials and are often isolated from soil and plant samples [54]. Paenibacillus secrete extracellular enzymes such as xylanase and cellulase, and their enzymatic system has been increasingly studied [55].
Bifunctional enzymes that present two activities were reported to present activities of xylanase-cellulase, xylanase-arabinosidase, xylanase-deacetylase, and xylanase-glucanase [56], but the enzymes that present bifunctional activity to xylanase and amylase were not found in the literature. However, some microorganisms classified as xylanolytic also present amylase activity, such as Gracilibacillus sp. TSCPVG [35], Paenibacillus xylanilyticus sp. nov. [54], and Bacillus amyloliquefaciens US573 [57]. These microorganisms presented in its genomes genes that separately encode amylase and xylanase enzymes. Zhao et al. [58] characterized a clone with 68 ORFs, and through sequence annotation, identified ORF 6 encoding an amylase and a cluster (ORF63 to ORF68) encoding a xylanase. This clone also exhibited amylase and xylanase activity in functional screening. One hypothesis for why an amylase rather than a xylanase was selected in this study is that 11 ORFs deduced as hypothetical proteins were discarded without analysis. Therefore, one of these sequences could contain a gene encoding xylanase since the initial MgrBr135 clone showed a degradation halo in medium with xylan.
Through the xylanase potential assay using the enzymatic index method, isolates with an EI value greater than or equal to 2.0 were obtained. Indexes equivalent to or greater than this value indicate that microorganisms are good extracellular enzyme producers on solid medium and are used as a parameter to compare enzymatic activity [59]. Based on this premise, most isolates had high-production enzyme rates. Eida et al. [60] obtained enzymatic indexes ranging from 0.56 to 2.84 in bacteria present in compost. Bacteria responsible for degrading lignocellulosic material in the digestive tract of the invertebrate Stenochironomus presented EI values from 0 to 3.9 [61]. Singh et al. [62] obtained EI values ranging from 0 to 4.5 in Bacillus, Stenotrophomonas, and Pseudomonas isolates from koala intestines.
Similar results were observed with yeast isolates that did not present the enzymatic index correlated with enzymatic activity [29]. Xylanase yield is governed by some key factors and parameters, such as gene expression level, accessibility to the substrate, pH, temperature, and agitation, and a modification in the culture method may alter enzyme production. The value obtained is higher than that presented by the genus Sugiyamaella, fungi isolated by Sena et al. [63] that achieved 0.05 to 0.29 U/mL activity. Bacteria isolated from Bacillus licheniformis species had xylanolytic activity of 0.44 U/mL when cultivated in minimal medium with xylan added [64]. Other authors reported that enzymatic activity from Bacillus and Streptomyces genera presented higher values than those found in the present study [65]. Sánchez et al. [66] observed the xylanolytic activity of bacteria grown in different salt concentrations and obtained satisfactory xylanase production from Paenibacillus isolates cultivated in 5% sodium chloride. These results suggest that the enzymatic activity of isolate 39 can be stable with the addition of sodium chloride in culture medium and that enzymes produced under these conditions can be tested in reactions with higher NaCl concentrations.
Brazilian mangroves possess a great diversity of microbial hydrolases [14, 21, 67, 68], and the use of different methodologies in this work was effective for accessing and exploring two different enzymes. The culture-dependent method associated with molecular analyses enabled to classify Bacillus and Paenibacillus that possess a xylanolytic capacity, supporting the participation of these bacteria in xylan degradation [14]. This method was more effective to obtain the initial target enzyme, as it allowed the direct isolation of microorganisms that produce xylanases. Through the culture-independent method, the gene that encodes xylanase was not obtained, but this method isolated a gene that encodes an amylase, the 1,4-α-glucan branching enzyme. The amylase accessed through the independent approach is produced by Planctomycetes, a trait newly described for this bacterium in mangroves, indicating its participation in starch degradation. In summary, adopting classical and culture-independent methodologies improved our view of results obtained from previous metagenomics analyses [22] and fosmid library studies [14], supporting the suggestion that the bacterial groups are associated with hydrolysis production. The further use of these resources may confirm the potential of mangrove conditions to modulate enzymes with useful characteristics for particular purposes.
Electronic supplementary material
{kind=link}
The quality parameter determined by Phred value to the sequences belonging to MgrBr18, MgrBr61, and MgrBr135 clones. (PNG 1228 kb)
{kind=link}
Identification of the α-amylase belonging to the family glycosyl hydrolase 13 (GH 13) with the carbohydrate-binding domain 48 (CBM 48) through dbCAN server. (BMP 481 kb)
{kind=link}
The electrophoresis of the amplification products of the 24 transformants to check the subcloning result. Colonies 19 and 20 showed a pronounced amplification. A - GeneRuler DNA Ladder 1 kb. B - MassRuler Low Range DNA Ladder. (PNG 518 kb)
{kind=link}
The of E. coli transformants exhibiting a hydrolysis halo in culture medium with starch. The expression test was performed in triplicate (Figures a, b, and c) with a one negative control (Figure d). (PNG 1434 kb)
Acknowledgments
We thank Danielle Gonçalves Santos for supporting the phylogenetic analyses, the Environmental Microbiology Laboratory of Embrapa Environment for sequencing fosmidial DNA, and the Laboratory of Soil Microbiology ESALQ-USP.
Funding information
We thank the National Council for Scientific and Technological Development (CNPQ) for funding the scientific initiation scholarship of the first author. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflict of interest.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Schaeffer-Novelli Y, Cintrdn-Molero G, Soares MLG, De-Rosa T. Brazilian mangroves. Aquat Ecosyst Health Manag. 2000;3:561–570. doi: 10.1080/14634980008650693. [DOI] [Google Scholar]
- 2.Chen Q, Zhao Q, Li J, et al. Mangrove succession enriches the sediment microbial community in South China. Sci Rep. 2016;6:27468. doi: 10.1038/srep27468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Thatoi H, Behera BC, Mishra RR, Dutta SK. Biodiversity and biotechnological potential of microorganisms from mangrove ecosystems: a review. Ann Microbiol. 2013;63:1–19. doi: 10.1007/s13213-012-0442-7. [DOI] [Google Scholar]
- 4.Paës G, Berrin JG, Beaugrand J. GH11 xylanases: structure/function/properties relationships and applications. Biotechnol Adv. 2012;30:564–592. doi: 10.1016/j.biotechadv.2011.10.003. [DOI] [PubMed] [Google Scholar]
- 5.Sermsathanaswadi J, Baramee S, Tachaapaikoon C, et al. The family 22 carbohydrate-binding module of bifunctional xylanase/β-glucanase Xyn10E from Paenibacillus curdlanolyticus B-6 has an important role in lignocellulose degradation. Enzym Microb Technol. 2017;96:75–84. doi: 10.1016/j.enzmictec.2016.09.015. [DOI] [PubMed] [Google Scholar]
- 6.Hamid Attia, Aftab Muhammad Nauman. Cloning, Purification, and Characterization of Recombinant Thermostable β-Xylanase Tnap_0700 from Thermotoga naphthophila. Applied Biochemistry and Biotechnology. 2019;189(4):1274–1290. doi: 10.1007/s12010-019-03068-0. [DOI] [PubMed] [Google Scholar]
- 7.Hung K-S, Liu S-M, Tzou W-S, et al. Characterization of a novel GH10 thermostable, halophilic xylanase from the marine bacterium Thermoanaerobacterium saccharolyticum NTOU1. Process Biochem. 2011;46:1257–1263. doi: 10.1016/j.procbio.2011.02.009. [DOI] [Google Scholar]
- 8.Chakdar H, Kumar M, Pandiyan K, et al. Bacterial xylanases: biology to biotechnology. 3. Biotech. 2016;6:1–15. doi: 10.1007/s13205-016-0457-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Valk V, Van Der Kaaij RM, Dijkhuizen L. Characterization of the starch-acting MaAmyB enzyme from Microbacterium aurum B8.A representing the novel subfamily GH13-42 with an unusual, multi-domain organization. Sci Rep. 2016;6:2–13. doi: 10.1038/srep36100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sharma A, Satyanarayana T. Microbial acid-stable α-amylases: characteristics, genetic engineering and applications. Process Biochem. 2013;48:201–211. doi: 10.1016/j.procbio.2012.12.018. [DOI] [Google Scholar]
- 11.Zeeman SC, Kossmann J, Smith AM. Starch: its metabolism, evolution, and biotechnological modification in plants. Annu Rev Plant Biol. 2010;61:209–234. doi: 10.1146/annurev-arplant-042809-112301. [DOI] [PubMed] [Google Scholar]
- 12.Nigam P. Microbial enzymes with special characteristics for biotechnological applications. Biomolecules. 2013;3:597–611. doi: 10.3390/biom3030597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Duan CJ, Feng JX. Mining metagenomes for novel cellulase genes. Biotechnol Lett. 2010;32:1765–1775. doi: 10.1007/s10529-010-0356-z. [DOI] [PubMed] [Google Scholar]
- 14.Ottoni JR, Cabral L, de Sousa STP, et al. Functional metagenomics of oil-impacted mangrove sediments reveals high abundance of hydrolases of biotechnological interest. World J Microbiol Biotechnol. 2017;33:1–13. doi: 10.1007/s11274-017-2307-5. [DOI] [PubMed] [Google Scholar]
- 15.Narihiro T, Suzuki A, Yoshimune K, et al. The combination of functional metagenomics and an oil-fed enrichment strategy revealed the phylogenetic diversity of lipolytic bacteria overlooked by the cultivation-based method. Microbes Environ. 2014;29:154–161. doi: 10.1264/jsme2.ME14002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Verma D, Kawarabayasi Y, Miyazaki K, Satyanarayana T. Cloning, expression and characteristics of a novel alkalistable and thermostable xylanase encoding gene (Mxyl) retrieved from compost-soil metagenome. PLoS One. 2013;8:e52459. doi: 10.1371/journal.pone.0052459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu Y, Yu J, Li F, et al. Crystal structure of a raw-starch-degrading bacterial α-amylase belonging to subfamily 37 of the glycoside hydrolase family GH13. Sci Rep. 2017;7:44067. doi: 10.1038/srep44067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Singh R, Kumar M, Mittal A, Mehta PK. Microbial enzymes: industrial progress in 21st century. 3. Biotech. 2016;6:174. doi: 10.1007/s13205-016-0485-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gonçalves ACS, dos Santos ACF, dos Santos TF, et al. High yield of functional metagenomic library from mangroves constructed in fosmid vector. Genet Mol Res. 2015;14:11841–11847. doi: 10.4238/2015.October.2.17. [DOI] [PubMed] [Google Scholar]
- 20.Miqueletto PB, Andreote FD, Dias AC, et al. Cultivation-independent methods applied to the microbial prospection of oil and gas in soil from a sedimentary basin in Brazil. AMB Express. 2011;1:35. doi: 10.1186/2191-0855-1-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dias ACF, Andreote FD, Dini-Andreote F, et al. Diversity and biotechnological potential of culturable bacteria from Brazilian mangrove sediment. World J Microbiol Biotechnol. 2009;25:1305–1311. doi: 10.1007/s11274-009-0013-7. [DOI] [Google Scholar]
- 22.Andreote FD, Jiménez DJ, Chaves D, et al. The microbiome of Brazilian mangrove sediments as revealed by metagenomics. PLoS One. 2012;7:e38600. doi: 10.1371/journal.pone.0038600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tsai YL, Olson BH. Rapid method for direct extraction of DNA from soil and sediments. Appl Environ Microbiol. 1991;57:1070–1074. doi: 10.1128/AEM.57.4.1070-1074.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.de Vasconcellos SP, Angolini CFF, García INS, et al. Screening for hydrocarbon biodegraders in a metagenomic clone library derived from Brazilian petroleum reservoirs. Org Geochem. 2010;41:675–681. doi: 10.1016/j.orggeochem.2010.03.014. [DOI] [Google Scholar]
- 25.Kasana RC, Salwan R, Dhar H, et al. A rapid and easy method for the detection of microbial cellulases on agar plates using Gram’s iodine. Curr Microbiol. 2008;57:503–507. doi: 10.1007/s00284-008-9276-8. [DOI] [PubMed] [Google Scholar]
- 26.Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Amore A, Parameswaran B, Kumar R, et al. Application of a new xylanase activity from Bacillus amyloliquefaciens XR44A in brewer’s spent grain saccharification. J Chem Technol Biotechnol. 2015;90:573–581. doi: 10.1002/jctb.4589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Meddeb-Mouelhi F, Moisan JK, Beauregard M. A comparison of plate assay methods for detecting extracellular cellulase and xylanase activity. Enzym Microb Technol. 2014;66:16–19. doi: 10.1016/j.enzmictec.2014.07.004. [DOI] [PubMed] [Google Scholar]
- 29.Otero DM, Cadaval CL, Teixeira LM, et al. Screening of yeasts capable of producing cellulase-free xylanase. Afr J Biotechnol. 2015;14:1961–1969. doi: 10.5897/AJB2015.14476. [DOI] [Google Scholar]
- 30.Miller GL. Use of dinitrosaiicyiic acid reagent for determination of reducing sugar. Anal Chem. 1959;31:426–428. doi: 10.1021/ac60147a030. [DOI] [Google Scholar]
- 31.Samanta AK, Kolte AP, Senani S, et al. A simple and efficient diffusion technique for assay of endo β-1,4-xylanase activity. Braz J Microbiol. 2011;42:1349–1353. doi: 10.1590/S1517-83822011000400016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Teixeira RSS, da Silva AS, Ferreira-Leitão VS, Bon EPDS. Amino acids interference on the quantification of reducing sugars by the 3,5-dinitrosalicylic acid assay mislead carbohydrase activity measurements. Carbohydr Res. 2012;363:33–37. doi: 10.1016/j.carres.2012.09.024. [DOI] [PubMed] [Google Scholar]
- 33.Ko KC, Lee JH, Han Y, et al. A novel multifunctional cellulolytic enzyme screened from metagenomic resources representing ruminal bacteria. Biochem Biophys Res Commun. 2013;441:567–572. doi: 10.1016/j.bbrc.2013.10.120. [DOI] [PubMed] [Google Scholar]
- 34.Wejse PL, Ingvorsen K, Mortensen KK. Salinity and temperature effects on accessibility of soluble and cross-linked insoluble xylans to endo-xylanases. IUBMB Life. 2005;57:761–763. doi: 10.1080/15216540500364271. [DOI] [PubMed] [Google Scholar]
- 35.Giridhar PV, Chandra TS. Production of novel halo-alkali-thermo-stable xylanase by a newly isolated moderately halophilic and alkali-tolerant Gracilibacillus sp. TSCPVG. Process Biochem. 2010;45:1730–1737. doi: 10.1016/j.procbio.2010.07.012. [DOI] [Google Scholar]
- 36.Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a laboratory manual, 2nd edn. Cold Spring Harb Lab Press. 10.1039/b813545b
- 37.Tamura K, Peterson D, Peterson N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Anantharaman K, Brown CT, Hug LA, et al. Thousands of microbial genomes shed light on interconnected biogeochemical processes in an aquifer system. Nat Commun. 2016;7:13219. doi: 10.1038/ncomms13219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ibrahim M, Yusof N, Mohd Yusoff MZ, Hassan MA. Enrichment of anaerobic ammonium oxidation (anammox) bacteria for short start-up of the anammox process: a review. Desalin Water Treat. 2016;57:13958–13978. doi: 10.1080/19443994.2015.1063009. [DOI] [Google Scholar]
- 40.Fuchsman CA, Staley JT, Oakley BB, et al. Free-living and aggregate-associated Planctomycetes in the Black Sea. FEMS Microbiol Ecol. 2012;80:402–416. doi: 10.1111/j.1574-6941.2012.01306.x. [DOI] [PubMed] [Google Scholar]
- 41.Bondoso J, Albuquerque L, Nobre MF, et al. Aquisphaera giovannonii gen. nov., sp. nov., a planctomycete isolated from a freshwater aquarium. Int J Syst Evol Microbiol. 2011;61:2844–2850. doi: 10.1099/ijs.0.027474-0. [DOI] [PubMed] [Google Scholar]
- 42.Sheng L, Aijia Z, Rongyu W, et al. Effects of sugarcane ratooning cultivation on the alteration of bacterial communities in the rhizosphere soil. Sugar Tech. 2012;14:275–283. doi: 10.1007/s12355-012-0147-z. [DOI] [Google Scholar]
- 43.Zhou D, Jing T, Chen Y, et al. Deciphering microbial diversity associated with Fusarium wilt-diseased and disease-free banana rhizosphere soil. BMC Microbiol. 2019;19:161. doi: 10.1186/s12866-019-1531-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sindhu R, Binod P, Madhavan A, et al. Molecular improvements in microbial α-amylases for enhanced stability and catalytic efficiency. Bioresour Technol. 2017;245:1740–1748. doi: 10.1016/j.biortech.2017.04.098. [DOI] [PubMed] [Google Scholar]
- 45.Thompson CE, Beys-da-Silva WO, Santi L, et al. A potential source for cellulolytic enzyme discovery and environmental aspects revealed through metagenomics of Brazilian mangroves. AMB Express. 2013;3:1–35. doi: 10.1186/2191-0855-3-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Naumoff DG, Ivanova AA, Dedysh SN. Phylogeny of β-xylanases from Planctomycetes. Mol Biol. 2014;48:439–447. doi: 10.1134/S0026893314030145. [DOI] [PubMed] [Google Scholar]
- 47.Vasudevan G, Siddarthan V, Solai Ramatchandirane P. Predominance of Bacillus sp. in soil samples of the southern regions of Western Ghats, India. Ann Microbiol. 2015;65:431–441. doi: 10.1007/s13213-014-0876-1. [DOI] [Google Scholar]
- 48.Gao YC, Guo SH, Wang JN, et al. Effects of different remediation treatments on crude oil contaminated saline soil. Chemosphere. 2014;117:486–493. doi: 10.1016/j.chemosphere.2014.08.070. [DOI] [PubMed] [Google Scholar]
- 49.Goh KM, Kahar UM, Chai YY, et al. Recent discoveries and applications of Anoxybacillus. Appl Microbiol Biotechnol. 2013;97:1475–1488. doi: 10.1007/s00253-012-4663-2. [DOI] [PubMed] [Google Scholar]
- 50.El Nagar A, Huys R, Bishop JDD. Widespread occurrence of the Southern Hemisphere ascidian Corella eumyota Traustedt, 1882 on the Atlantic coast of Iberia. Aquat Invasions. 2010;5:169–173. doi: 10.3391/ai.2010.5.2.06. [DOI] [Google Scholar]
- 51.Romero-Garcia S, Hernandez-Bustos C, Merino E, et al. Homolactic fermentation from glucose and cellobiose using Bacillus subtilis. Microb Cell Factories. 2009;8:23. doi: 10.1186/1475-2859-8-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Asoodeh A, Chamani J, Lagzian M. A novel thermostable, acidophilic α-amylase from a new thermophilic “Bacillus sp. Ferdowsicous” isolated from Ferdows hot mineral spring in Iran: purification and biochemical characterization. Int J Biol Macromol. 2010;46:289–297. doi: 10.1016/j.ijbiomac.2010.01.013. [DOI] [PubMed] [Google Scholar]
- 53.Maki M, Leung KT, Qin W. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int J Biol Sci. 2009;5:500–516. doi: 10.7150/ijbs.5.500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Rivas R, Mateos PF, Martínez-Molina E, Velázquez E. Paenibacillus xylanilyticus sp. nov., an airborne xylanolytic bacterium. Int J Syst Evol Microbiol. 2005;55:405–408. doi: 10.1099/ijs.0.63173-0. [DOI] [PubMed] [Google Scholar]
- 55.Rhee Mun Su, Sawhney Neha, Kim Young Sik, Rhee Hyun Jee, Hurlbert Jason C., St. John Franz J., Nong Guang, Rice John D., Preston James F. GH115 α-glucuronidase and GH11 xylanase from Paenibacillus sp. JDR-2: potential roles in processing glucuronoxylans. Applied Microbiology and Biotechnology. 2016;101(4):1465–1476. doi: 10.1007/s00253-016-7899-4. [DOI] [PubMed] [Google Scholar]
- 56.Rashamuse KJ, Visser DF, Hennessy F, et al. Characterisation of two bifunctional cellulase-xylanase enzymes isolated from a bovine rumen metagenome library. Curr Microbiol. 2013;66:145–151. doi: 10.1007/s00284-012-0251-z. [DOI] [PubMed] [Google Scholar]
- 57.Farhat-Khemakhem A, Blibech M, Boukhris I, et al. Assessment of the potential of the multi-enzyme producer Bacillus amyloliquefaciens US573 as alternative feed additive. J Sci Food Agric. 2018;98:1208–1215. doi: 10.1002/jsfa.8574. [DOI] [PubMed] [Google Scholar]
- 58.Zhao S, Wang J, Bu D, et al. Novel glycoside hydrolases identified by screening a chinese holstein dairy cow rumen-derived metagenome library. Appl Environ Microbiol. 2010;76:6701–6705. doi: 10.1128/AEM.00361-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Carmo FL, Santos HF, Peixoto RS, et al. Tank bromeliad water: similar or distinct environments for research of bacterial bioactives? Braz J Microbiol. 2014;45:185–192. doi: 10.1590/S1517-83822014000100024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fathallh Eida M, Nagaoka T, Wasaki J, Kouno K. Isolation and characterization of cellulose-decomposing bacteria inhabiting sawdust and coffee residue composts. Microbes Environ. 2012;27:226–233. doi: 10.1264/jsme2.ME11299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Koroiva R, Souza CWO, Toyama D, et al. Lignocellulolytic enzymes and bacteria associated with the digestive tracts of Stenochironomus (Diptera: Chironomidae) larvae. Genet Mol Res. 2013;12:3421–3434. doi: 10.4238/2013.April.2.2. [DOI] [PubMed] [Google Scholar]
- 62.Singh S, Thavamani P, Megharaj M, Naidu R. Multifarious activities of cellulose degrading bacteria from koala (Phascolarctos cinereus) faeces. J Anim Sci Technol. 2015;57:23. doi: 10.1186/s40781-015-0056-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sena Letícia M. F., Morais Camila G., Lopes Mariana R., Santos Renata O., Uetanabaro Ana P. T., Morais Paula B., Vital Marcos J. S., de Morais Marcos A., Lachance Marc-André, Rosa Carlos A. d-Xylose fermentation, xylitol production and xylanase activities by seven new species of Sugiyamaella. Antonie van Leeuwenhoek. 2016;110(1):53–67. doi: 10.1007/s10482-016-0775-5. [DOI] [PubMed] [Google Scholar]
- 64.Seo JK, Park TS, Kwon IH, et al. Characterization of cellulolytic and xylanolytic enzymes of Bacillus licheniformis JK7 isolated from the rumen of a native Korean goat. Asian Australas J Anim Sci. 2013;26:50–58. doi: 10.5713/ajas.2012.12506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Techapun C, Sinsuwongwat S, Poosaran N, et al. Production of a cellulase-free xylanase from agricultural waste materials by a thermotolerant Streptomyces sp. Biotechnol Lett. 2001;23:1685–1689. doi: 10.1023/A:1012448229555. [DOI] [Google Scholar]
- 66.Sánchez MM, Fritze D, Blanco A, et al. Paenibacillus barcinonensis sp. nov., a xylanase-producing bacterium isolated from a rice field in the Ebro River delta. Int J Syst Evol Microbiol. 2005;55:935–939. doi: 10.1099/ijs.0.63383-0. [DOI] [PubMed] [Google Scholar]
- 67.Sá ALB, Dias ACF, Quecine MC, et al. Screening of endoglucanase-producing bacteria in the saline rhizosphere of Rhizophora mangle. Braz J Microbiol. 2014;45:193–197. doi: 10.1590/S1517-83822014000100025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Soares F, Marcon J, Pereira e Silva M, et al. A novel multifunctional β-N-acetylhexosaminidase revealed through metagenomics of an oil-spilled mangrove. Bioengineering. 2017;4:62. doi: 10.3390/bioengineering4030062. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The quality parameter determined by Phred value to the sequences belonging to MgrBr18, MgrBr61, and MgrBr135 clones. (PNG 1228 kb)
Identification of the α-amylase belonging to the family glycosyl hydrolase 13 (GH 13) with the carbohydrate-binding domain 48 (CBM 48) through dbCAN server. (BMP 481 kb)
The electrophoresis of the amplification products of the 24 transformants to check the subcloning result. Colonies 19 and 20 showed a pronounced amplification. A - GeneRuler DNA Ladder 1 kb. B - MassRuler Low Range DNA Ladder. (PNG 518 kb)
The of E. coli transformants exhibiting a hydrolysis halo in culture medium with starch. The expression test was performed in triplicate (Figures a, b, and c) with a one negative control (Figure d). (PNG 1434 kb)